
Chitosanase displayed on liposome can increase its activity and stability
9
Please cite this article in press as: Ngo, K.X., et al., Chitosanase displayed on liposome can increase its activity and stability. J. Biotechnol. (2010), doi:10.1016/j.jbiotec.2010.01.014 ARTICLE IN PRESS G Model BIOTEC-5404; No. of Pages 9 Journal of Biotechnology xxx (2010) xxx–xxx Contents lists available at ScienceDirect Journal of Biotechnology journal homepage: www.elsevier.com/locate/jbiotec Chitosanase displayed on liposome can increase its activity and stability Kien Xuan Ngo a , Hiroshi Umakoshi a , Toshinori Shimanouchi a , Hiroyuki Sugaya a,b , Ryoichi Kuboi a,∗ a Division of Chemical Engineering, Graduate School of Engineering Science, Osaka University, 1-3 Machikaneyama, Toyonaka, Osaka 560-8531, Japan b Medical Devices Research Lab., Specialty Material Research Labs, TORAY Industries, 1-2 Sonoyama 3chome, Otsu, Shiga 520-0842, Japan article info Article history: Received 1 October 2009 Received in revised form 19 January 2010 Accepted 19 January 2010 Available online xxx Keywords: Immobilized chitosanase onto liposome Chitosanase Liposome Streptomyces griseus Signal peptide abstract The strategy to prepare a novel biocatalyst by the immobilization of chitosanase onto liposome (ICL) was carried out based on the direct interaction of liposomes with cell membrane of Streptomyces griseus cell. The ICL was characterized in relation to the molecular weight of protein, the chitosanase activity, the effect of the surface hydration of various liposomes on hydrolysis activity of immobilized chitosanase and the stability of ICL under various extreme conditions. The SDS-PAGE analysis of the purified ICL sample shows the existence of a protein with approximately 39 kDa that corresponded to the sum of weight of the mature chitosanase and its signal peptide (38.8 kDa). The above protein of ICL also expresses the chi- tosanase activity that is significantly higher than that of the conventional chitosanase. Furthermore, the surface hydration of liposomes used to prepare ICL that affected the activity of immobilized chitosanase verified the importance of liposome surfaces. Indeed, the stability of ICL assayed by measuring the chi- tosanase activity is significantly higher than that of conventional chitosanase under various temperatures and pH conditions. These characteristics of ICL show the possible preparation of the biocatalysts that can be prepared by immobilizing enzymes onto liposome vesicles properly. © 2010 Elsevier B.V. All rights reserved. 1. Introduction The production of the functional biomaterial oligochitosan has been attractive widely because of its valuable applications. The oligochitosan possesses many good functions for food (Shahidi et al., 1999), medicine (Majeti and Kumar, 2000) and cosmetic (Dodane and Vilivalam, 1998) usages. The pentameric and hex- americ oligochitosans have antibacterial activity (Uchida and Ohtakara, 1988), antitumor activity (Suzuki et al., 1986) and immune-enhancing effects (Hirano et al., 1991). Conventionally, oligochitosans were produced from chitosan by the acid hydrol- ysis method (Horowitz et al., 1957). However, the hydrolysis by enzymes has more advantages to produce oligochitosans because of milder and easier operation to control the enzymatic reaction to produce oligochitosans and to restrict the by-products such as glu- cosamine (Izume and Ohtakara, 1987). Indeed, several approaches have been studied to improve the hydrolysis potential of enzymes for the production of oligochitosans such as the direct immobiliza- tion of chitosanase onto solid surface (Ming et al., 2006; Kuroiwa et al., 2003), or circulation of chitosanase inside the ultrafiltration membrane reactor (Jeon and Kim, 2000a,b) to produce oligochi- tosans continuously. Although these approaches have been proved as prospective methods some disadvantages for commercial uti- ∗ Corresponding author. Tel.: +81 6 6850 6285; fax: +81 6 6850 6285. E-mail address: [email protected] (R. Kuboi). lization still need to improve due to the low production yield, low enzymatic activity and stability, high cost and limited availability. Liposome, a closely phospholipid bilayer membrane vesicle, has inner aqueous phase and a nano-order interface (∼5 nm) harbor- ing hydration layer on its surface. The enzymatic reaction inside small and giant liposome vesicles have been dedicated by many researchers (Fischer et al., 2002; Yoshimoto et al., 2004; Oberholzer et al., 1995, 1999; Treyer et al., 2002) because of its merits for appli- cations in medical and biomedical treatment (Walde and Ichikawa, 2001). Furthermore, the enzymes entrapped inside liposome have been proved to be more stable under high temperature (Wang et al., 2003; Han et al., 1998) and higher selectivity (Yoshimoto et al., 2004). However, the permeability of outside substrates across the lipid membrane of liposome is certainly main problem for enzy- matic reactions inside liposome reactors. In the efforts to enhance the permeability, most of these methods required organic solvents which could give a rise to considerable solvent residues that are unacceptable in food and medical products, and the low yield of entrapped enzyme inside liposome as well as the large-scale appli- cation should be taken into the difficulty. In other approaches utilizing the merits of liposome surface, the enzyme immobilized onto liposome has shown to be more effective and stable in enzymatic reaction (Durour et al., 1996). Liposome itself could recognize the biomolecules through the combined interactions based on the hydrogen bond stability (Yoshimoto et al., 2000; Kuboi et al., 2004); recruit the specific peptide of the oxi- dized and fragmented superoxide dismutase (SOD) onto its lipid 0168-1656/$ – see front matter © 2010 Elsevier B.V. All rights reserved. doi:10.1016/j.jbiotec.2010.01.014
Transcript of Chitosanase displayed on liposome can increase its activity and stability

G
B
C
Ka
b
a
ARRAA
KICLSS
1
boe(aOioyeopchftemta
0d
ARTICLE IN PRESSModel
IOTEC-5404; No. of Pages 9
Journal of Biotechnology xxx (2010) xxx–xxx
Contents lists available at ScienceDirect
Journal of Biotechnology
journa l homepage: www.e lsev ier .com/ locate / jb io tec
hitosanase displayed on liposome can increase its activity and stability
ien Xuan Ngoa, Hiroshi Umakoshia, Toshinori Shimanouchia, Hiroyuki Sugayaa,b, Ryoichi Kuboia,∗
Division of Chemical Engineering, Graduate School of Engineering Science, Osaka University, 1-3 Machikaneyama, Toyonaka, Osaka 560-8531, JapanMedical Devices Research Lab., Specialty Material Research Labs, TORAY Industries, 1-2 Sonoyama 3chome, Otsu, Shiga 520-0842, Japan
r t i c l e i n f o
rticle history:eceived 1 October 2009eceived in revised form 19 January 2010ccepted 19 January 2010vailable online xxx
eywords:
a b s t r a c t
The strategy to prepare a novel biocatalyst by the immobilization of chitosanase onto liposome (ICL)was carried out based on the direct interaction of liposomes with cell membrane of Streptomyces griseuscell. The ICL was characterized in relation to the molecular weight of protein, the chitosanase activity, theeffect of the surface hydration of various liposomes on hydrolysis activity of immobilized chitosanase andthe stability of ICL under various extreme conditions. The SDS-PAGE analysis of the purified ICL sampleshows the existence of a protein with approximately 39 kDa that corresponded to the sum of weight of
mmobilized chitosanase onto liposomehitosanaseiposometreptomyces griseusignal peptide
the mature chitosanase and its signal peptide (38.8 kDa). The above protein of ICL also expresses the chi-tosanase activity that is significantly higher than that of the conventional chitosanase. Furthermore, thesurface hydration of liposomes used to prepare ICL that affected the activity of immobilized chitosanaseverified the importance of liposome surfaces. Indeed, the stability of ICL assayed by measuring the chi-tosanase activity is significantly higher than that of conventional chitosanase under various temperaturesand pH conditions. These characteristics of ICL show the possible preparation of the biocatalysts that can
zing e
be prepared by immobili. Introduction
The production of the functional biomaterial oligochitosan haseen attractive widely because of its valuable applications. Theligochitosan possesses many good functions for food (Shahidit al., 1999), medicine (Majeti and Kumar, 2000) and cosmeticDodane and Vilivalam, 1998) usages. The pentameric and hex-meric oligochitosans have antibacterial activity (Uchida andhtakara, 1988), antitumor activity (Suzuki et al., 1986) and
mmune-enhancing effects (Hirano et al., 1991). Conventionally,ligochitosans were produced from chitosan by the acid hydrol-sis method (Horowitz et al., 1957). However, the hydrolysis bynzymes has more advantages to produce oligochitosans becausef milder and easier operation to control the enzymatic reaction toroduce oligochitosans and to restrict the by-products such as glu-osamine (Izume and Ohtakara, 1987). Indeed, several approachesave been studied to improve the hydrolysis potential of enzymes
or the production of oligochitosans such as the direct immobiliza-ion of chitosanase onto solid surface (Ming et al., 2006; Kuroiwa
Please cite this article in press as: Ngo, K.X., et al., Chitosanase displayed ondoi:10.1016/j.jbiotec.2010.01.014
t al., 2003), or circulation of chitosanase inside the ultrafiltrationembrane reactor (Jeon and Kim, 2000a,b) to produce oligochi-
osans continuously. Although these approaches have been proveds prospective methods some disadvantages for commercial uti-
∗ Corresponding author. Tel.: +81 6 6850 6285; fax: +81 6 6850 6285.E-mail address: [email protected] (R. Kuboi).
168-1656/$ – see front matter © 2010 Elsevier B.V. All rights reserved.oi:10.1016/j.jbiotec.2010.01.014
nzymes onto liposome vesicles properly.© 2010 Elsevier B.V. All rights reserved.
lization still need to improve due to the low production yield, lowenzymatic activity and stability, high cost and limited availability.
Liposome, a closely phospholipid bilayer membrane vesicle, hasinner aqueous phase and a nano-order interface (∼5 nm) harbor-ing hydration layer on its surface. The enzymatic reaction insidesmall and giant liposome vesicles have been dedicated by manyresearchers (Fischer et al., 2002; Yoshimoto et al., 2004; Oberholzeret al., 1995, 1999; Treyer et al., 2002) because of its merits for appli-cations in medical and biomedical treatment (Walde and Ichikawa,2001). Furthermore, the enzymes entrapped inside liposome havebeen proved to be more stable under high temperature (Wang etal., 2003; Han et al., 1998) and higher selectivity (Yoshimoto et al.,2004). However, the permeability of outside substrates across thelipid membrane of liposome is certainly main problem for enzy-matic reactions inside liposome reactors. In the efforts to enhancethe permeability, most of these methods required organic solventswhich could give a rise to considerable solvent residues that areunacceptable in food and medical products, and the low yield ofentrapped enzyme inside liposome as well as the large-scale appli-cation should be taken into the difficulty.
In other approaches utilizing the merits of liposome surface, theenzyme immobilized onto liposome has shown to be more effective
liposome can increase its activity and stability. J. Biotechnol. (2010),
and stable in enzymatic reaction (Durour et al., 1996). Liposomeitself could recognize the biomolecules through the combinedinteractions based on the hydrogen bond stability (Yoshimoto etal., 2000; Kuboi et al., 2004); recruit the specific peptide of the oxi-dized and fragmented superoxide dismutase (SOD) onto its lipid

ING
B
2 iotech
m(scb
msvci2oCommabtf
Ipsma
2
2
M3poc�tSch(
2
c(aiwaaioacttlogm
ARTICLEModel
IOTEC-5404; No. of Pages 9
K.X. Ngo et al. / Journal of B
embrane to recover the active SOD from non-active fragmentsTuan et al., 2008); regulate gene expression in cell free translationystem (Bui et al., 2008). Liposome modified with Mn–porphyrinomplex could simultaneously induce antioxidative like activity ofoth superoxide dismutase and peroxidase (Umakoshi et al., 2008).
In our previous studies, the interaction of liposome with cellembrane of Streptomyces griseus could effectively occur under the
pecific heat condition (Ngo et al., 2009). Such interaction couldary the surface properties of cell membrane such as surface netharge and surface net hydrophobicity (Ngo et al., 2008a), resultingn the enhanced production of chitosanase by these cells (Ngo et al.,005) and the enhanced secretion of this enzyme probably becausef the protein bound-lipid membrane mode (Ngo et al., 2008b).onventionally, the chitosanase of S. griseus cell is secreted basedn the regulation of SP bound to the lipid membrane and secretoryachinery and during or shortly after its secretion through cellembrane the SP is cleaved by signal peptidase type I (Tanabe et
l., 2003). The inhibition of signal peptidase to not cleave the SP thatinds chitosanase during its secretion across cell membrane andhe interaction of liposomes with cell membranes are importantactors to prepare ICL.
The purpose of this study is to prepare and characterize theCL from S. griseus cells. The ICL was prepared by S. griseus cellsretreated with the inhibitor of signal peptidase and various lipo-omes. The prepared ICL was characterized focusing on the proteinolecular weight of chitosanase and the variation of its enzymatic
ctivity and stability onto various liposome surfaces.
. Materials and methods
.1. Materials
Chitosanase from S. griseus was purchased from Sigma (St. Louis,O, USA). The lipids including 1-palmitoyl-2-oleoyl-sn-glycero-
-phosphatidylcholine (POPC), 1-palmitoyl-2-oleoyl-sn-glycero-3-hosphatidic acid (POPA), cholesterol (Ch) and 1-palmitoyl-2-leoyl-sn-glycero-3-[phospho-rac-(1-glycerol)] (POPG) were pur-hased from Avanti Polar Lipid (Alabaster City, AL, USA). The-lactam inhibitor (amoxicillin) was purchased from LKT Labora-
ories Inc. (St. Paul, USA). PhastGelTM Homogeneous 12.5, Silvertaining kit, Protein PlusOneTM were purchased from GE Health-are Bioscience AB (Uppsala, Sweden). Other chemical reagents ofighly analytical grade were purchased from Wako Pure ChemicalsOsaka, Japan).
.2. Preparation and characterization of ICL
The S. griseus cells (106/ml) were grown in 300 ml round flaskontaining 100 ml chitosanase production medium including KCl0.05%), KH2PO4 (0.1%), MgSO4·7H2O (0.05%), FeSO4·7H2O (0.001%)nd water-soluble chitosan (0.2%). The cultivation was carried outn water bath shaker at 140 rpm and 37 ◦C for 24 h. The cultivation
as stopped and amoxicillin that is �-lactam class of antibiotict minimum inhibition concentration was added to inhibit thectivity of signal peptidase (Andes and Craig, 1998). Parallelly, var-ous liposomes (100 nm in diameter) such as POPA, POPC, POPG,r POPC/Ch liposomes prepared as our previous paper (Ngo etl., 2005) was asceptically added to the cell broth at final con-entration of 0.4 mM lipid. The cell broth was cultivated by thereatment at 37 ◦C (20 min) and 41 ◦C (20 min) repeatedly with
Please cite this article in press as: Ngo, K.X., et al., Chitosanase displayed ondoi:10.1016/j.jbiotec.2010.01.014
otal 15 to 20 times. The above treatment was stopped and cells-iposomes suspension was cooled down to 4 ◦C and kept for 10 minn ice before separating the supernatant from cells by centrifu-ation at 7000 rpm at 4 ◦C for 10 min. The supernatant containsature chitosanase, chitosanase with SP bound-liposome (as ICL)
PRESSnology xxx (2010) xxx–xxx
and other impurities. The sample was further purified to removemature chitosanase and other impurities, which could not sta-bly bind to liposome surfaces. For this purpose, the module ofhollow membrane column (TORAY, Japan) was chosen to purifyICL. The supernatant was loaded and circulated inside this mem-brane module at low temperature for 30 min to wash out themature chitosanase and other impurities. The ICL could be purifiedeffectively and remained inside membrane column. The characteri-zation of ICL was based on protein molecular weight of chitosanasewith and without SP, chitosanse activity onto various liposomesurfaces, and stability of ICL at different temperatures and pHconditions.
2.3. Measurement of chitosanase activity of ICL
Chitosanase activity was measured with glycol chitosan as asubstrate (Ohtaka, 1988). The reaction mixture containing 0.5 mlof 2 wt% glycol chitosan in 0.1 M phosphate buffer (pH 5.6) and0.5 ml of ICL or standard chitosanase solution was incubated at45 ◦C for 10 min and then the enzymatic reaction was stoppedby boiling for 4 min. This solution was cooled down and addedto 1 ml of acetylacetone diluted in 0.5N Na2CO3 (1:50, v/v)and 1 ml of distilled water and boiled for 20 min. An amount5 ml of ethanol and 1 ml of enrich reagent (p-dimethylamino-benzaldehyde, DMAB) were then added and incubated at 65 ◦Cfor 10 min. Then, this sample was centrifuged at 16,000 rpm for30 min to remove the precipitated pellets. The absorbance of super-natant was measured at 530 nm using UV spectrophotometer(UV-1600A, Shimadzu, Japan). One unit of chitosanase is definedas the amount of enzyme that hydrolyzed glycol chitosan in orderto release 1 �M glucosamine per min at given assay conditions.In order to liberate the chitosanase from liposomes, triton X100was added at final concentration of 1% to disrupt the liposomemembranes.
2.4. Analysis
Protein concentration was measured by using BCA protein assaykit purchased from PIERCE (Rockland, IL, USA). The lipid concentra-tion of POPC, POPG, POPA and POPC/Ch liposomes was measuredby using lipid assay kit (Wako, Japan). For the analysis of molecu-lar weight of chitosanase with or without SP, SDS-PAGE was usedas the following: to liberate the chitosanase from liposomes, tritonX100 was added at final concentration of 1% to disrupt the lipo-some membranes of ICL solution. A 10 �l sample of disrupted ICLwas dissolved with 17 �l of SDS-PAGE sample buffer and 5 �l ofmercaptoethanol. This mixture was heated for 5 min at 95 ◦C fordenaturation of protein. The 4 �l samples were applied onto a com-mercial gel (Homologous 12.5, GE Healthcare, Chicago, IL) set forthe electrophoresis system (PhastSystem, GE Healthcare, Chicago,IL). The gel was finally stained with silver staining method and chi-tosanase with and without SP was detected as a major staining bandat 38.8 kDa and 33.5 kDa, respectively.
In another experiment, the surface hydration of POPA, POPC,POPC/Ch or POPG liposomes was characterized through the anti-symmetric infrared spectrum of PO2
− in their lipid headgroupsusing FTIR (JASCO, Japan). Liposomes with size (100 nm) and lipidconcentration (10 mM) were prepared in distilled water as theprevious study (Ngo et al., 2005). The sample of 30 �l liposomesuspensions at room temperature and neutral pH was applied to50 �m thick-cell with CaF2 window for the analysis of the anti-
liposome can increase its activity and stability. J. Biotechnol. (2010),
symmetric infrared spectrum of PO2−. The infrared (IR) spectra of
PO2− of liposomes were measured with a FT/IR 4100 spectrometer
(JASSCO, Japan) equipped with an Hg–Cd–Te detector. The reso-lution was 4 cm−1 and the frequency in range from 1700 cm−1 to950 cm−1 was collected for each sample. The subtraction of infrared

ARTICLE IN PRESSG Model
BIOTEC-5404; No. of Pages 9
K.X. Ngo et al. / Journal of Biotechnology xxx (2010) xxx–xxx 3
Table 1Relative estimation of average number of chitosanase molecules displayed onto one liposome.
Factor POPC (Huang andMason, 1978)
POPA (Huang andMason, 1978)
POPC/Ch (70:30 mol%) (Falck et al., 2004)
POPC Cholesterol
Surface area of outer head group (Å2) 74 60 44 382
st
cta
2m
wstwi3awWrdorsllslw
rgfti2olEtMss
�T
Surface area of inner head group (Å ) 61Thickness of bilayer (Å) 37Average number of chitosanasemolecules displayed onto one liposome
4.77
pectrum of PO2− to that of water as the blank was carried out with
he accuracy of the frequency reading better than ±0.1 cm−1.The influence of surface hydration of different liposomes on the
hitosanase activity of ICL was also figured out to verify the impor-ance of liposome surfaces that can be utilized to increase activitynd stability of immobilized chitosanase.
.5. Kinetic of enzymatic reaction using Michaelis–Mentonodels
The apparent activities of ICL and conventional chitosanasesere measured by using glycol chitosan (82.084 kDa) as the
ubstrate. In these enzymatic reactions, the concentration of chi-osanase enzymes of all ICL samples and conventional chitosanaseas adjusted to 1.76 × 10−10 M in order to utilize most enzymes
nto the reaction with the substrate that varied from 0.00 M to.65 × 10−8 M. The measured lipid concentrations of POPA, POPC,nd POPC/Ch liposomes used in these enzymatic reaction of ICLere 0.34 × 10−5 M, 0.32 × 10−5 M, and 0.39 × 10−5 M, respectively.e noted that the above chitosanase concentration of ICLs is
epresenting for the total concentration of chitosanases that areisplayed onto liposomes in the ICL solution. The average numbersf chitosanase molecules displayed onto one liposome were alsoelatively estimated and shown in Table 1. The numbers of lipo-omes in ICL solution was estimated by division of the total molaripid concentration of ICL solution to the molar concentration of oneiposome that were calculated based on parameters given in Table 1uch as surface area of outer and inner head groups and thickness ofipid bilayer of each liposome as well as the assumption of liposome
ith regularly spherical shape.For the steady-state approach to the analysis of an enzymatic
eaction, we assume the conversion of single substrate, S, to a sin-le product, P, by an enzyme, E. The breakdown of the ES complexollowing catalytic conversion into product and enzyme is so fasthat we may neglect its formation. Thus the slow step is the chem-cal step and we denote the rate constant as kcat (Gabi and Lee,007). Another assumption of equilibrium-state, the concentrationf enzyme and substrate complex, ES, is in thermodynamic equi-ibrium with the free forms of enzyme and substrate and that theS complex is held together by noncovalent force. In other words,he rate constants for the formation of ES complex (k1, units of
−1 s−1) and its breakdown to the free components (k−1, units of−1
Please cite this article in press as: Ngo, K.X., et al., Chitosanase displayed ondoi:10.1016/j.jbiotec.2010.01.014
) are considerably faster than the catalytic step (kcat, units of−1) (Scheme 1).
Assuming that [ES] complex is in a ‘steady state’, then kcat/Km
109 and the [ES] complex is in ‘equilibrium-state’, kcat/Km <109.hen steady-state assumption can also be performed (Scheme 2).
Scheme 1.
60 44 3837 34
5.03 4.47
The velocity of product formation versus the substrate con-centration is therefore performed by Michaelis–Menton model toestimate Michaelis constant (Km), maximum velocity (Vmax) andkcat.
2.6. Stability of ICL
The stability of ICL and conventional chitosanase with or with-out liposome under various heat and pH conditions was assayed bymeasuring chitosanase activity. The stability of ICLs and conven-tional chitosanase at different temperatures was assayed by theexposure of the enzyme samples at 25 ◦C, 30 ◦C, 37 ◦C, 41 ◦C, 45 ◦C,50 ◦C, 55 ◦C, and 65 ◦C for 10 min. The stability of the enzyme sam-ples under different pHs of 1.3, 2.3, 4.5, 5, 5.9, 7.6, and 10.2 wasassayed by incubation of these enzyme samples at given pH val-ues for 24 h. After exposure to above conditions of temperaturesand pHs, the chitosanase activity was measured at correspondedexperimental temperatures and pHs, similar to the measurementof chitosanase activity described above.
3. Results and discussion
3.1. Preparation of ICL
It has been reported that the extracellular chitosanase isreleased through the secretory machinery of S. griseus cell by thehydrolysis of its SP catalyzed by the signal peptidase (Tanabe etal., 2003). Fig. 1 shows the schematic illustration for the prepa-ration of ICL by treatment of S. griseus cell with liposomes, heatand inhibitor of signal peptidase. The operation has been describedin Section 2. The nascent chitosanase synthesized inside cytosolof S. griseus cell is secreted across secretion machinery. Duringor shortly after the translocation across secretion machinery, theSP is cleaved from mature protein and consequently degradedby the specific signal peptidase (SSPase). After its translocation,the mature protein folds into its native conformation (Tjallsma etal., 2000). In the conventional case at 37 ◦C (arrow A), liposomedid not plays a role to enhance secretion and production of chi-tosanase from S. griseus cells as our previous papers (Ngo et al.,2005, 2008b). In the other cases, our strategy to prepare ICL is totreat S. griseus cell with inhibitor of signal peptidase to not cleaveSP bound mature chitosanase during or shortly after translocationacross secretion machinery. Then, the liposomes were applied tocoincubate with these cells and the heat-up at 41 ◦C and heat-down at 37 ◦C were continuously applied to control the adsorption
liposome can increase its activity and stability. J. Biotechnol. (2010),
and desorption of liposomes onto cell membrane. The adsorp-tion of intact liposomes onto cell membrane plays a key for theinteraction of liposomes with the secreted chitosanase harboringSP onto cell membrane interface to form ICL (shown as B arrow)although the mode of this interaction should be further elucidated.
Scheme 2.

ARTICLE IN PRESSG Model
BIOTEC-5404; No. of Pages 9
4 K.X. Ngo et al. / Journal of Biotechnology xxx (2010) xxx–xxx
F incub( nd he
Tb(acrbmhoa(obamet
adBrcbat
TS
n
ig. 1. Schematic illustration for the preparation of ICLs by S. griseus. (A) Cells wereB) cells treated with inhibitor of signal peptidase were incubated with liposomes a
he adsorption and desorption of liposomes onto cell membraney the heat treatment was well established in our previous reportNgo et al., 2009). As summarized and shown in Table 2, the vari-tion of surface properties of cell membrane was significant byhanging the temperature of 37 ◦C or 41 ◦C. It shows that theaising temperature caused the increased surface net hydropho-icity, the reduced surface hydration and membrane fluidity of cellembranes. The variation of surface net hydrophobicity, surface
ydration, and membrane fluidity of cell membrane at temperaturef 37 ◦C and 41 ◦C is important to establish the modes of its inter-ction with liposomes such as adsorption, fusion or internalizationNgo et al., 2009). In addition, the increasing of membrane fluidityf POPC, POPC/Ch, and POPA liposomes was obviously predictedy raising temperature. Hence, the controlling the adsorptionnd desorption of POPC, POPC/Ch, or POPA liposomes onto cellembrane of S. griseus could be simply based on hydrophobic
ffects, membrane fluidity and hydration effects by changing theemperature.
After heat treatment to solution of liposomes and S. griseus cellss described in Section 2, the cell suspension was finally cooledown at 4 ◦C for 10 min to separate ICL from the cell membrane.y centrifugation at 18,000 rpm at 4 ◦C for 30 min, ICL solution was
Please cite this article in press as: Ngo, K.X., et al., Chitosanase displayed ondoi:10.1016/j.jbiotec.2010.01.014
ecovered as the supernatant. This raw solution was purified byirculation of raw ICL solution inside the module of hollow mem-rane column (TORAY, Japan) to wash out the small-size impuritiesnd proteins through the membrane. The ICL with larger size (morehan 100 nm) remained inside membrane column. This operation
able 2ummary the heat effects on the variation surface properties of cell membrane of S. grise
Terms Cell membrane of S. griseus Lip
PO
37 ◦C 41 ◦C 37
Surface net hydrophobicity, HFS (kJ/mol) −110a −90a n.dSurface hydration, � (cm−1) 1232.3 1251.6 n.dSurface charge at pH 7.5, Z (−) −13.1b −13.1b 0Membrane fluidity, 1/P (−) 3.8c 2.9c 4.2
.d.: not determineda Ngo et al. (2009).b Ngo et al. (2008a).c Ngo et al. (2008b).
ated with liposomes at 37 ◦C without the treatment of inhibitor of signal peptidase;ated at 37 ◦C (20 min) and at 41 ◦C (20 min) repeatedly from 15 to 20 times.
will have two effects: (i) condensation of ICL sample and (ii) sep-aration and purification of ICL from impurities such as non/weakbinding proteins onto liposomes.
3.2. Characteristics of chitosanase and its signal peptide
The SP of chitosanase from S. griseus can herewith be classi-fied as the signal peptide type I (leader peptide) (Tanabe et al.,2003). The primary structure analysis of the SP associated withmature chitosanase is shown in Fig. 2. The SP contains 53 aminoacids in length with molecular weight approximately 5.3 kDa andisoelectric point (pI) at 13. The analysis shows that the positivelycharged and hydrophobic amino acids are abundant in the primarystructure of SP, as shown in signal peptide region in Fig. 2b andc. In another case, the primary structure analysis of the SP associ-ated mature chitosanase is also shown in Fig. 2a–c. This enzymecontains 359 amino acids in length with molecular weight approx-imate 38.8 kDa and pI at 9.3 while mature chitosanase (without SP)contains 306 amino acids in length with molecular weight approx-imately 33.5 kDa and pI at 7.9. The charged and hydrophobic aminoacids are poorly included though the hydrophilic and neutral aminoacids are abundant in the primary structure of mature chitosanase,
liposome can increase its activity and stability. J. Biotechnol. (2010),
as the analysis shown in Fig. 2b and c.This analysis implies that the SP could be a key to prepare
ICL because SP bound chitosanase is able to mimic its biologicalfunction to interact with liposome membrane by electrostatic andhydrophobic effects.
us bacterium and liposome membranes for preparation of ICLs.
osome
PC POPC/Ch POPA
◦C 41 ◦C 37 ◦C 41 ◦C 37 ◦C 41 ◦C
. −400a n.d. n.d. n.d. n.d.
. 1232.3 n.d. 1230.4 n.d. 1189.90 0 0 −1 −1
c 4.8c n.d. 2.5 n.d. 5.7

ARTICLE IN PRESSG Model
BIOTEC-5404; No. of Pages 9
K.X. Ngo et al. / Journal of Biotechnology xxx (2010) xxx–xxx 5
Fig. 2. Characteristics of chitosanase from S. griseus and its sig-nal peptide analyzed based on its amino acid sequence by using:http://us.expasy.org/tools/protscale.html. Amino acids sequence of SP andmature part of chitosanase are shown as italic and normal text, respectively(mhqrttpptg tapttgrrrm arkallaasa lgvvtalmap vragaapapa aptaatplaa ngqlsvcgrqlcnasgqava lngmsthgtq wyaqcvtdgs lnalaqdwra dvlrvstyvq ggyetdpag ftaraqkfidavfi
3
cltepStpiLctp3s(pl
3r
Oh
Fig. 3. SDS-PAGE analysis of chitosanase harboring signal peptide (SP) produced byStreptomyces griseus using Phastgel homologue 12.5 with silver staining for observa-tion of proteins. The cells treated with inhibitor and liposomes were heated at 41 ◦C.L1: Protein band shown by red arrow as chitosanase + SP produced by S. griseus cells;
ahargmyav idwhmlspgd pnanlarakt fftamakkyk nhpgvlyeia nepsgvswsa iksyaeqiipiraqdpdav vlvgtrawss fgvsegsnes evvnnpvras nimytfhfya ashreeylsa ldrasdklpvvtefgtqny agegandftm sqryldlmkr kkiswtnwny sddhrsgavf ktgtcngtnw tgtgvlkeagwirdrirq).
.3. Protein molecular weight of chitosanase of ICL
A possibility of the recovered SP associated-chitosanase washaracterized by measuring its activity and analysis of its molecu-ar weight by SDS-PAGE. The expression of chitosanase activity ofhe purified ICL sample confirmed the abundance of chitosanasenzyme. The chitosanase activity is well corresponding with ourrevious studies (Ngo et al., 2005, 2008a,b). The existence ofP associated-chitosanase in this sample was verified by usinghe SDS-PAGE, shown in Fig. 3. The molecular weight of therotein shown as red arrow mark in L1 was estimated approx-
mate 39 kDa that is higher than that of mature chitosanase in2 (33.5 kDa). The result also showed that there was no band ofhitosanase (without SP) existed in L1 although some impure pro-eins were remained with distinct molecular weights after theurification. The protein with molecular weight approximately9 kDa expressed chitosanase activity is well corresponding to theum of the molecular weight of mature chitosanase and its SP38.8 kDa). Indeed, the chitosanase–liposome complex was pro-osed to be formed via its SP associated-lipid membrane of the
iposome absorbing onto cell membrane of S. griseus.
.4. Characterization of enzymatic activity of ICL based on
Please cite this article in press as: Ngo, K.X., et al., Chitosanase displayed ondoi:10.1016/j.jbiotec.2010.01.014
eaction rate
Units of chitosanase indicate the enzymatic activity of ICL.ne unit of chitosanase is defined as the amount of enzyme thatydrolyzed glycol chitosan in order to release 1 �M glucosamine
L2: band shown by arrow as standard chitosanase; M: Marker. To liberate the chi-tosanase from liposomes before SDS-PAGE running, triton X100 was added at finalconcentration of 1% to disrupt the liposomes membrane.
per min at given assay conditions. For these words, the enzymaticactivity of ICL prepared by POPC, POPC/Ch, or POPA liposomes wascharacterized through the kinetic analysis of the glucosamine pro-duction reaction in certainly fixed time. Fig. 4A shows the rate ofproduct (glucosamine) formation versus the substrate (glycol chi-tosan) concentration. The result shows that ICL prepared by POPA,POPC or POPC/Ch liposomes significantly accelerated the rate ofproduct formation about twice to four times higher than that of thecontrol. The maximum velocity (Vmax) of enzymatic reaction of ICLsprepared by different liposomes was measured through the rate ofglucosamine formation. Among three kinds of ICLs assayed here,the Vmax of ICL prepared by POPA liposome expresses the high-est rate of glucosamine formation at 12.4 × 10−10 M/s. The otherICLs prepared by POPC or POPC/Ch liposomes express slower rateof glucosamine formation at 7.1 × 10−10 M/s and 5.8 × 10−10 M/s,respectively. Indeed, the Vmax value is only 3.9 × 10−10 M/s for con-ventional chitosanase as the control. This comparison shows theeffect of different liposome surfaces on the rate of enzymatic reac-tion of immobilized chitosanase. The comparison between aboveexperiments is carried out by normalization of total concentrationof chitosanase fixed at 1.76 × 10−10 M.
In order to verify the influence of liposome surfaces on the effi-ciency of catalytic rate (kcat/Km) of ICL, the kcat/Km values of variouskinds of ICLs was calculated and shown in Fig. 4B. As the result,the kcat/Km value of ICL prepared by POPA liposome is 1.6 × 109
(M−1 s−1) that corresponds to the steady state of enzymatic reac-tion. The kcat/Km values of ICL prepared by POPC or POPC/Chliposomes are 6.9 × 108 and 6.2 × 108 (M−1 s−1), respectively, corre-sponding to the equilibrium state of enzymatic reaction. Indeed, thekcat/Km of conventional chitosanase is only 2.8 × 108 (M−1 s−1), sig-nificantly lower than those of ICLs. The above results confirm thatthe efficiency of catalytic rate of ICLs prepared by POPC, POPC/Ch, orPOPA liposomes are significantly higher than that of conventionalchitosanase.
There are several effects of liposome membranes on the cat-alytic potential of ICL such as charge, membrane fluidity and surfacehydration. As above hypothesis (Fig. 1), SP might play a role toimmobilize chitosanase onto liposomes properly. Although we esti-
liposome can increase its activity and stability. J. Biotechnol. (2010),
mated the average numbers of chitosanase molecules that aredisplayed onto one liposome, it is noted that this value was not trulyreflecting the real numbers of chitosanase molecules displayedonto one liposome in ICLs solution. The displayed chitosanase onto

ARTICLE IN PRESSG Model
BIOTEC-5404; No. of Pages 9
6 K.X. Ngo et al. / Journal of Biotechnology xxx (2010) xxx–xxx
Fig. 4. Rate of product (glucosamine) formation versus concentration of substrate (glycol chitosan) (A) and efficiency of catalytic rate kcat/Km (B) catalyzed by different kindsof ICLs and conventional chitosanase (control). The enzymatic reaction was carried out by the coincubation of 150 �l ICL prepared by POPC or POPA, or POPC/Ch liposomes(final total protein concentration that was normalized at fixed concentration of 1.76 × 10−10 M for all ICL samples) with 150 �l glycol chitosan (final concentration was variedf ried ow 5 ◦C fg
osoPrteswbehCrisbraotentPtttissIt(tbts
rom 0.00 M to 3.65 × 10−8 M) at 45 ◦C for 10 min. The control experiment was carith the same concentration of chitosanase and glycol chitosan of ICL samples at 4
lucosamine concentration was quantified as the described method in Section 2.
ne liposome was probably not same amount with every lipo-ome. The estimation is shown in Table 1. The average numbersf chitosanase molecules displayed onto one POPC, POPC/Ch, orOPA liposome are 4.77, 4.47 and 5.03, respectively. This resulteflects chitosanase with SP that is highly positively charged por-ion favorably interacts with negatively charged liposomes bylectrostatic effects. In addition, the membrane fluidity of lipo-omes also plays crucial role for the immobilization of chitosanaseith SP onto membrane of liposomes by insertion of hydropho-
ic portion of SP into lipid bilayer membrane by hydrophobicffects. The result reveals that the higher fluidity liposome theigher amount of immobilized chitosanases onto its surface is.oncerning about electrostatic effects on the enhanced reactionate of chitosanase of ICL, liposome surface probably played anmportant role to attract the binding of the substrate onto itsurface. In such mode, chitosanases displayed onto liposome canind to substrate more frequently to carry out the enzymaticeactions. Previously, it has been reported that neutral and neg-tively charged liposomes prepared by phosphatidylcholine (PC)r phosphatidic acid (PA) strongly attract the binding of chi-osan (positively charged polymer) on their surfaces (Henriksent al., 1994; Mertins et al., 2010). Indeed, the ICL prepared byegatively charged POPA liposomes expressed the highest reac-ion rate as compared to those prepared by neutral POPC andOPC/Ch liposomes. The cryo-electron microscope also showedhat the binding of chitosan on liposome surface caused the clus-er formation of liposome vesicles (Henriksen et al., 1994). Forhis model, we predicted that each ICL in solution did not workndependently. They gathered and worked together on liposomeurfaces in the cluster forms once chitosans were added to ICLsolution. Consequently, the amount of produced glucosamine byCLs was significantly increased at the same period of reactionime as compared to that obtained by conventional chitosanases
Please cite this article in press as: Ngo, K.X., et al., Chitosanase displayed ondoi:10.1016/j.jbiotec.2010.01.014
same normalized concentration of chitosanase). As the defini-ion of chitosanase activity, the amount of glucosamine producedy ICLs or conventional chitosanases at certain period of reactionime show the activity of chitosanase enzyme. Here, it is clearlyhown the effects of liposome surfaces on the enhanced activity
ut the enzymatic reaction between conventional chitosanase and glycol chitosanor 10 min. The enzymatic reactions were stopped by boiling at 95 ◦C for 4 min and
of displayed chitosanase. Hence, some characteristics of liposomesshould be clarified to give a deeper insight of its effects on thecatalytic potential of immobilized chitosanase such as electro-static effects, membrane fluidity, and surface hydration of liposomemembranes.
3.5. Relationship between surface hydration of liposome andchitosanase activity
A possible mechanism on the ICL formation was further char-acterized by the effect of the hydration of liposome surface on theactivity of immobilized chitosanase.
First, the surface hydration of various liposomes was measuredthrough the hydration level of PO2
− in phospholipid head groupsof different liposomes. In fact, the infrared spectra of PO2
− showthe hydration of this functional group in the different head groupsof POPC, POPA, and POPC/Ch liposomes (Casal et al., 1987) and inthe principle, the lower frequency of PO2
− the higher hydrationof liposome is. Fig. 5A shows the infrared frequencies of the PO2
−
antisymmetric stretching modes of POPC, POPA and POPC/Ch smallunilamellar vesicles (SUVs). In POPA–SUVs, infrared frequency ofPO2
− group had the maximum frequency at 1189.9 cm−1 while thisvalue of POPC-SUVs is 1232.3 cm−1. Incorporation of 30 mol% ofcholesterol (Ch) into POPC-SUVs brings this value about 1.9 cm−1
to lower frequency, as shown for POPC/Ch liposome. These resultsmean that among three liposomes here the surface hydration ofPOPA liposome is highest and the incorporation of cholesterol(30 mol%) into POPC liposome increases surface hydration this lipo-some. The above measured infrared spectra of PO2
− are quite fittedwith previous reports. The infrared frequency of PO2
− group in eggyolk phosphatidic acid (EPA) vesicles in the gel phase had the maxi-mum frequency at 1187.2 cm−1 (Nabet et al., 1994). In the Egg-yorkphosphatidylethanolamine (EPC) vesicle in liquid-crytalline state,
liposome can increase its activity and stability. J. Biotechnol. (2010),
this value was 1231.7 cm−1 and the incorporation of more than26 mol% cholesterol into EPC–SUVs brought about 3.1 cm−1 shiftto lower frequency (Yamamoto et al., 1995) because cholesterolincreased the space around P O bond and enhanced the hydrationof PO2
−.
ARTICLE IN PRESSG Model
BIOTEC-5404; No. of Pages 9
K.X. Ngo et al. / Journal of Biotechnology xxx (2010) xxx–xxx 7
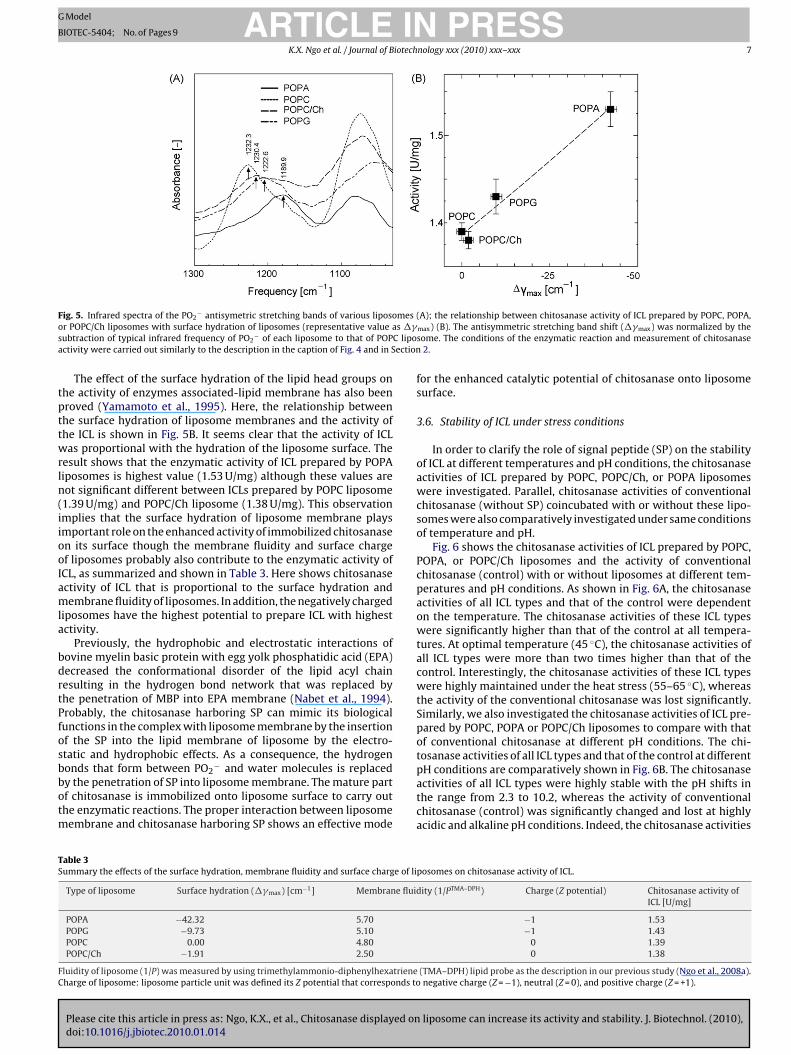
F meso s ��s C lipoa ection
tpttwrln(iiooIamla
bdrtPfosbbotm
TS
FC
ig. 5. Infrared spectra of the PO2− antisymetric stretching bands of various liposo
r POPC/Ch liposomes with surface hydration of liposomes (representative value aubtraction of typical infrared frequency of PO2
− of each liposome to that of POPctivity were carried out similarly to the description in the caption of Fig. 4 and in S
The effect of the surface hydration of the lipid head groups onhe activity of enzymes associated-lipid membrane has also beenroved (Yamamoto et al., 1995). Here, the relationship betweenhe surface hydration of liposome membranes and the activity ofhe ICL is shown in Fig. 5B. It seems clear that the activity of ICLas proportional with the hydration of the liposome surface. The
esult shows that the enzymatic activity of ICL prepared by POPAiposomes is highest value (1.53 U/mg) although these values areot significant different between ICLs prepared by POPC liposome1.39 U/mg) and POPC/Ch liposome (1.38 U/mg). This observationmplies that the surface hydration of liposome membrane playsmportant role on the enhanced activity of immobilized chitosanasen its surface though the membrane fluidity and surface chargef liposomes probably also contribute to the enzymatic activity ofCL, as summarized and shown in Table 3. Here shows chitosanasectivity of ICL that is proportional to the surface hydration andembrane fluidity of liposomes. In addition, the negatively charged
iposomes have the highest potential to prepare ICL with highestctivity.
Previously, the hydrophobic and electrostatic interactions ofovine myelin basic protein with egg yolk phosphatidic acid (EPA)ecreased the conformational disorder of the lipid acyl chainesulting in the hydrogen bond network that was replaced byhe penetration of MBP into EPA membrane (Nabet et al., 1994).robably, the chitosanase harboring SP can mimic its biologicalunctions in the complex with liposome membrane by the insertionf the SP into the lipid membrane of liposome by the electro-tatic and hydrophobic effects. As a consequence, the hydrogen
Please cite this article in press as: Ngo, K.X., et al., Chitosanase displayed ondoi:10.1016/j.jbiotec.2010.01.014
onds that form between PO2− and water molecules is replaced
y the penetration of SP into liposome membrane. The mature partf chitosanase is immobilized onto liposome surface to carry outhe enzymatic reactions. The proper interaction between liposome
embrane and chitosanase harboring SP shows an effective mode
able 3ummary the effects of the surface hydration, membrane fluidity and surface charge of li
Type of liposome Surface hydration (��max) [cm−1] Membrane fluid
POPA −42.32 5.70POPG −9.73 5.10POPC 0.00 4.80POPC/Ch −1.91 2.50
luidity of liposome (1/P) was measured by using trimethylammonio-diphenylhexatrieneharge of liposome: liposome particle unit was defined its Z potential that corresponds to
(A); the relationship between chitosanase activity of ICL prepared by POPC, POPA,max) (B). The antisymmetric stretching band shift (��max) was normalized by thesome. The conditions of the enzymatic reaction and measurement of chitosanase
2.
for the enhanced catalytic potential of chitosanase onto liposomesurface.
3.6. Stability of ICL under stress conditions
In order to clarify the role of signal peptide (SP) on the stabilityof ICL at different temperatures and pH conditions, the chitosanaseactivities of ICL prepared by POPC, POPC/Ch, or POPA liposomeswere investigated. Parallel, chitosanase activities of conventionalchitosanase (without SP) coincubated with or without these lipo-somes were also comparatively investigated under same conditionsof temperature and pH.
Fig. 6 shows the chitosanase activities of ICL prepared by POPC,POPA, or POPC/Ch liposomes and the activity of conventionalchitosanase (control) with or without liposomes at different tem-peratures and pH conditions. As shown in Fig. 6A, the chitosanaseactivities of all ICL types and that of the control were dependenton the temperature. The chitosanase activities of these ICL typeswere significantly higher than that of the control at all tempera-tures. At optimal temperature (45 ◦C), the chitosanase activities ofall ICL types were more than two times higher than that of thecontrol. Interestingly, the chitosanase activities of these ICL typeswere highly maintained under the heat stress (55–65 ◦C), whereasthe activity of the conventional chitosanase was lost significantly.Similarly, we also investigated the chitosanase activities of ICL pre-pared by POPC, POPA or POPC/Ch liposomes to compare with thatof conventional chitosanase at different pH conditions. The chi-tosanase activities of all ICL types and that of the control at different
liposome can increase its activity and stability. J. Biotechnol. (2010),
pH conditions are comparatively shown in Fig. 6B. The chitosanaseactivities of all ICL types were highly stable with the pH shifts inthe range from 2.3 to 10.2, whereas the activity of conventionalchitosanase (control) was significantly changed and lost at highlyacidic and alkaline pH conditions. Indeed, the chitosanase activities
posomes on chitosanase activity of ICL.
ity (1/PTMA–DPH) Charge (Z potential) Chitosanase activity ofICL [U/mg]
−1 1.53−1 1.43
0 1.390 1.38
(TMA–DPH) lipid probe as the description in our previous study (Ngo et al., 2008a).negative charge (Z = −1), neutral (Z = 0), and positive charge (Z = +1).

ARTICLE IN PRESSG Model
BIOTEC-5404; No. of Pages 9
8 K.X. Ngo et al. / Journal of Biotechnology xxx (2010) xxx–xxx
F f ICLo h lipoc ig. 4 a
optPvs
oScastwaepbjsll
h
ig. 6. Effect of different temperatures and pH conditions on chitosanase activity of conventional chitosanase in the presence or absence of POPC, POPA, or POPC/Chitosanase activity were carried out similarly to the description in the caption of F
f all ICL types were significantly higher than that of the control atH conditions from 2.3 to 10.2. This observation implies that chi-osanase harboring SP could be immobilized properly onto POPA,OPC or POPC/Ch liposomes. Here, the surface of liposomes playsery important role to enhance the chitosanase activity ICL and itstability under stress condition.
The above results might show the roles of SP in the complexf chitosanase with liposome membrane. To verify the role of theP, the chitosanase activities of the conventional chitosanase coin-ubated with or without POPC, POPA or POPC/Ch liposomes werelso investigated at different temperatures and pH conditions. Ashown in Fig. 6C and D, there was no significant difference in chi-osanase activities of the conventional chitosanase coincubatedith or without POPA, POPC or POPC/Ch liposomes at given temper-
tures and pH conditions although slight effects of liposomes on thenhanced activity of chitosanase could be observed at higher tem-eratures (Fig. 6C). These observations confirm that the complexetween conventional chitosanase and liposomes was not stable
ust based on the conventional absorption of chitosanase onto lipo-
Please cite this article in press as: Ngo, K.X., et al., Chitosanase displayed ondoi:10.1016/j.jbiotec.2010.01.014
ome surface. Thus, the association of SP with lipid membranes ofiposomes was playing crucial roles to immobilize chitosanase ontoiposome properly.
The key factor to stabilize the conformation of protein isydrophobic interaction between hydrophobic residues inside pro-
prepared by POPC, POPA, or POPC/Ch liposomes (A and B) and chitosanase activitysomes (C and D). The conditions of the enzymatic reaction and measurement ofnd in Section 2.
tein. The hydration is essential feature in the native conformationof a protein in aqueous solution. The hydrophilic residues located atthe molecular surface and the hydrophobic residues located insideto stabilize the conformation of protein. The loss in hydration freeenergy; and the gain in conformational free energy were producedby non-bonded interaction between dehydrated atomic groups. Inone model, a hydration shell is formed based on the interactionof protein with water molecules in the first hydration layer sur-rounding the protein (Ooi et al., 1987). The interactions of proteinwith water molecules in the first hydrated layer interfere with thewater structure of the second and further remote layers, result-ing in the changes in free energy (Ooi et al., 1987). It has also beenfound that charge–charge interactions are attractive at all pH valueswhere the protein unfolds. The temperature and pH dependencesin these interactions contribute to acid denaturation of protein butthe other effects such as hydrogen bonding and solvation of proteinare important as well (Yang and Honig, 1993). Based on their sug-gestions, it is possible that the stability of chitosanase on the surfaceof liposomes at high temperature and acidic and alkali conditions is
liposome can increase its activity and stability. J. Biotechnol. (2010),
dependent on the stability of the hydration shell surrounding chi-tosanase mediated by hydrogen bond interaction with the surfacehydration of PO2
− head group of liposomes. In addition, the associ-ation of the SP into lipid membrane of liposome by the electrostaticand hydrophobic interactions probably strengthened the hydrogen

ING
B
iotech
besmiltohp
A
Rt1MoCTCta
R
A
B
C
D
D
F
F
G
H
H
H
H
H
I
J
J
K
ARTICLEModel
IOTEC-5404; No. of Pages 9
K.X. Ngo et al. / Journal of B
onds network of chitosanase onto liposome membranes. How-ver, the conventional adsorption of chitosanase onto liposomeurface was not proper to immobilize chitosanase onto liposomeembrane. For this mode, Durour et al. have reported that the
mmobilization efficiency of chymotripsin on phosphatidylcholineiposomes was thermally dependent on the hydrophobic interac-ions between hydrophobic parts of enzymes and hydrophobic tailsf phospholipids in liposomes. These may show the importance ofydrophobic interactions on the stability of the protein/lipid com-lex.
cknowledgements
The fundamental concept of this study was supported by theesearch Group of “Membrane Stress Biotechnology”. It was par-ially supported by a Grant-in-Aid for Scientific Research (Nos.9656203, 19656220, 20360350, 20760539, 21246121) from theinistry of Education, Culture, Sports, Science and Technology
f Japan, a grant from the Global COE program Bio-Environmenthemistry of Japan Society for the Promotion of Science (JSPS).he authors are grateful to the Research Center for Solar Energyhemistry of Osaka University and the Gas Hydrate Analyzing Sys-em of Osaka University. One of the authors (K.X. Ngo) also deeplycknowledges the financial support of JSPS.
eferences
ndes, D., Craig, W.A., 1998. In vivo activities of amoxicillin and amoxicillin-clavulanate against Streptococcus pneumoniae: application to breakpointdeterminations. Antimicrob. Agents Chemother. 42 (9), 2375–2379.
ui, H.T., Umakoshi, H., Ngo, K.X., Nishida, M., Shimanouchi, T., Kuboi, R., 2008. Lipo-some membrane itself can affect gene expression in the Escherichia coli cell-freetranslation system. Langmuir 24 (19), 10537–10542.
asal, H.L., Mantsch, H.H., Hauser, H., 1987. Infrared studies of fully hydrated satu-rated phosphatidylserine bilayers. Effect of lithium and calcium. Biochemistry26, 4408–4416.
odane, V., Vilivalam, V.D., 1998. Pharmaceutical applications of chitosan. Pharm.Sci. Tech. 1 (6), 246–253.
urour, P., Vuillemard, J.C., Laloy, E., Simard, R.E., 1996. Characterization of enzymeimmobilization in liposomes prepared from proliposomes. J. Microencaps. 13(2), 185–194.
alck, E., Patra, M., Karttunen, M., Hyvönen, M.T., Vattulainen, I., 2004. Lessonsof slicing membranes: interplay of packing, free area, and lateral diffusion inphospholipid/cholesterol bilayers. Biophys. J. 87 (2), 1076–1091.
ischer, A., Franco, A., Oberholzer, T., 2002. Giant vesicles as microreactor for enzy-matic mRNA synthesis. ChemBiochem 3, 409–417.
abi, N.W., Lee, R.W., 2007. Applied Cell and Molecular Biology for Engineers.McGraw-Hill Professional.
an, X., Li, G., Lin, K., 1998. FTIR study of the thermal denaturation of �-actinin inits lipid-free and dioleoylphosphatidylglycerol-bound state and the central andN-terminal domain of �-actinin in D2O. Biochemistry 37, 10730–10737.
enriksen, I., Smistad, G., Karlsen, J., 1994. Interaction between liposomes and chi-tosan. Int. J. Pharm. 101, 227–236.
irano, S., Iwata, M., Yamanaka, K., Tanaka, H., Toda, T., Inui, H., 1991. Enhancementof serum lysozyme activity by induction a mixture of chitosan oligosaccharidesintravenously in rabbits. A. Biol. Chem. 55, 2623–2625.
orowitz, S.T., Roseman, S., Blumenthal, H.J., 1957. The preparation of glucosamineoligosaccharides. 1. Separation. J. A. Chem. Soc. 79, 5046–5049.
uang, C., Mason, J.T., 1978. Geometric packing constraints in egg phosphatidyl-choline vesicles. Proc. Natl. Acad. Sci. U.S.A. 75 (1), 308–310.
zume, M., Ohtakara, A., 1987. Preparation of d-glucosamine oligosaccharides by theenzymatic hydrolysis of chitosan. A. Biol. Chem. 51, 1189–1191.
eon, Y.J., Kim, S.K., 2000a. Continuous production of chitooligosaccharides using adual reactor system. Proc. Biochem. 35, 623–632.
Please cite this article in press as: Ngo, K.X., et al., Chitosanase displayed ondoi:10.1016/j.jbiotec.2010.01.014
eon, Y.J., Kim, S.K., 2000b. Production of chitooligosaccharides using an ultra-filtration membrane reactor and their antibacterial activity. Carbo. Pol. 41,133–141.
uboi, R., Shimanouchi, T., Yoshimoto, M., Umakoshi, H., 2004. Detection of proteinconformation under stress conditions using liposomes as sensor. Mat. Sens. Mat.16 (5), 241–254.
PRESSnology xxx (2010) xxx–xxx 9
Kuroiwa, T., Ichikawa, S., Sato, S., Mukataka, S., 2003. Improvement of the yieldof physiologically active oligosaccharides in continuous hydrolysis of chitosanusing immobilized chitosanases. Biotech. Bioeng. 84, 121–127.
Majeti, N.V., Kumar, R., 2000. A review of chitin and chitosan applications. React.Funct. Polym. 46 (1), 1–27.
Mertins, O., Schneider, P.H., Pohnmann, A.R., Silveira, N.P.D., 2010. Interactionbetween phospholipids bilayer and chitosan in liposomes investigated by 31PNMR spectroscopy. Colloid Surf. B: Bioint. 75, 294–299.
Ming, M., Kuroiwa, T., Ichikawa, S., Sato, S., Mukataka, S., 2006. Production of chitosanoligosaccharides at high concentration by immobilized chitosanase. Food Sci.Tech. Res. 12 (2), 85–90.
Nabet, A., Boggs, J.M., Pezolet, M., 1994. Study by FTIR spectroscopy of the interactionof bovine myelin basic protein with phosphatidic acid. Biochemistry 33 (49),14792–14799.
Ngo, K.X., Umakoshi, H., Shimanouchi, T., Jung, H.S., Morita, S., Kuboi, R., 2005. Heat-enhanced production of chitosanase from Streptomyces griseus in the presenceof liposome. J. Biosci. Bioeng. 100 (5), 495–501.
Ngo, K.X., Umakoshi, H., Shimanouchi, T., Kuboi, R., 2008a. Variation of surface prop-erties of Streptomyces griseus cells after heat treatment with liposome. Solv. Extr.Res. Dev. Jpn. 15, 133–137.
Ngo, K.X., Umakoshi, H., Shimanouchi, T., Kuboi, R., 2008b. Enhanced release of chi-tosanase from Streptomyces griseus through direct interaction of liposome withcell membrane under heat stress. J. Biosci. Bioeng. 106 (6), 602–605.
Ngo, K.X., Umakoshi, H., Shimanouchi, T., Kuboi, R., 2009. Characterization of heatenhanced-interaction of neutral liposome with lipid membrane of Streptomycesgriseus cell. Colloid Surf. B: Bioint. 73, 399–407.
Oberholzer, T., Albrizio, M., Luisi, P.L., 1995. Polymerase chain reaction in liposomes.Chem. Biol. 2 (10), 677–682.
Oberholzer, T., Meyer, E., Amato, I., Lustig, A., Monnard, P.A., 1999. Enzymaticreactions in liposomes using the detergent-induced liposome loading method.Biochim. Biophys. Acta 1416, 57–68.
Ohtaka, A., 1988. Meth. Enzymol. 161, 458–459.Ooi, T., Oobatake, M., Némethy, G., Scheraga, H.A., 1987. Accessible surface areas as
a measure of the thermodynamic parameters of hydration of peptides. PNAS 84(10), 3086–3090.
Shahidi, F., Kamil, J., Arachchi, V., Jeon, Y.J., 1999. Food applications of chitin andchitosans. Trends Food Sci. Tech. 10, 37–51.
Suzuki, K., Mikami, T., Okawa, Y., Tokoro, A., Suzuki, S., Suzuki, M., 1986. Antitu-mor effect of hexa-N-acetylchitohexaose and chitohexaose. Carbohydr. Res. 151,403–408.
Tanabe, T., Morinaga, K., Fukamizo, T., Mitsutomo, M., 2003. Novel chi-tosanase from Streptomyces griseus HUT 6037 with transglycosylation activity(biochemistry and molecular biology). Biosci. Biotech. Biochem. 67 (2),354–364.
Tjallsma, H., Bolhuis, A., Jongbloed, J.D.H., Bron, S., Dijl, J.M.V., 2000. Signal peptide-dependent protein transport in Bacillus subtilis: a genome-based survey of thesecretome. Microbiol. Mol. Biol. Rev. 64 (3), 515–547.
Treyer, M., Walde, P., Oberholzer, T., 2002. Permeability enhancement of lipid vesi-cles to nucleotides by use of sodium cholate: basic studies and applicationto an enzyme-catalyzed reaction occurring inside the vesicles. Langmuir 18,1043–1050.
Tuan, L.Q., Umakoshi, H., Shimanouchi, T., Kuboi, R., 2008. Liposome-recruitedactivity of oxidized and fragmented superoxide dismutase. Langmuir 24 (2),350–354.
Uchida, Y., Ohtakara, A., 1988. Chitosanase from Bacillus species. Met. Enz. B161,501–505.
Umakoshi, H., Morimoto, K., Ohama, Y., Nagami, H., Shimanouchi, T., Kuboi, R., 2008.Liposome modified with Mn–Porphyrin complex can simultaneously induceantioxidative enzyme-like activity of both superoxide dismutase and peroxi-dase. Langmuir 24 (9), 4451–4455.
Walde, P., Ichikawa, S., 2001. Enzymes inside lipid vesicles: preparation, reactivityand applications. Biomol. Bioeng. 18, 143–177.
Wang, S., Yoshimoto, M., Fukunaga, K., Nakao, K., 2003. Optimal covalent immobi-lization of glucose oxidase-containing liposomes for highly stable biocatalyst inbioreactor. Biotech. Bioeng. 83, 444–453.
Yamamoto, I., Konto, A., Handa, T., Miyajima, K., 1995. Regulation of phospholipaseD activity by neutral lipids in egg-york phophatidylcholine small unilamellarvesicles and by calcium ion in aqueous medium. Biochim. Biophys. Acta 233,21–121.
Yang, A.S., Honig, B., 1993. On the pH dependence of protein stability. J. Mol. Biol.231, 459–474.
Yoshimoto, M., Shimanouchi, T., Umakoshi, H., Kuboi, R., 2000. Immobilized lipo-
liposome can increase its activity and stability. J. Biotechnol. (2010),
some chromatography for refolding and purification of protein. J. Chromatogr.B Biomed. Sci. A 743 (1–2), 93–99.
Yoshimoto, M., Wang, S., Fukunaga, K., Treyer, M., Walde, P., Kuboi, R., Nakao, K.,2004. Enhancement of apparent substrate selectivity of proteinase K encap-sulated in liposomes through a cholate-induced alteration of the bilayerpermeability. Biotech. Bioeng. 85 (2), 222–233.




















