
GEG Participates in the Regulation of Cell and Organ Shape during Corolla and Carpel Development in...
13
The Plant Cell, Vol. 11, 1093–1104, June 1999, www.plantcell.org © 1999 American Society of Plant Physiologists GEG Participates in the Regulation of Cell and Organ Shape during Corolla and Carpel Development in Gerbera hybrida Mika Kotilainen, a,1 Yrjö Helariutta, a Merja Mehto, a Eija Pöllänen, a Victor A. Albert, b Paula Elomaa, a and Teemu H. Teeri a a Institute of Biotechnology, Viikki Biocenter, P.O. Box 56, 00014–University of Helsinki, Finland b Lewis B. and Dorothy Cullman Program for Molecular Systematics Studies, New York Botanical Garden, Bronx, New York 10458-5126 The molecular mechanisms that control organ shape during flower development are largely unknown. By using differ- ential hybridization techniques, a cDNA designated GEG (for Ge rbera hybrida homolog of the g ibberellin [GA]–stimu- lated transcript 1 [GAST1] from tomato) was isolated from a library representing late stages of corolla development in Gerbera. GEG expression was detected in corollas and carpels, with expression spatiotemporally coinciding with flower opening. In corollas and styles, GEG expression is temporally correlated with the cessation of longitudinal cell expansion. In plants constitutively expressing GEG, reduced corolla lengths and carpels with shortened and radially ex- panded stylar parts were found, with concomitant reduction of longitudinal cell expansion in these organs. In addition, in styles, an increase in radial cell expansion was detected. Taken together, these observations indicate a regulatory role for the GEG gene product in determining the shape of the corolla and carpel. The deduced amino acid sequence of the GEG gene product shares high similarity with previously characterized putative cell wall proteins encoded by GA- inducible genes, namely, GAST1, GIP (for GA-induced gene of petunia), and the GASA (for GA-stimulated in Arabidopsis) gene family. Our studies suggest that GEG, the expression of which can also be induced by application of GA 3 , plays a role in phytohormone-mediated cell expansion. INTRODUCTION Plant cells do not migrate during development as do animal cells, and organ shape is deteminated by the organized and regulated control of cell expansion together with cell division (Meyerowitz, 1997). Despite its importance, the molecular and genetic regulation of cell expansion is not well under- stood. Emerging data support a view that the direction and magnitude of the enlargement of the primary cell wall largely determine the expansion pattern and thereby the final shapes of the cells. Water uptake causes constant turgor pressure in plant cells, which is the driving force of cell ex- pansion. Turgor pressure is counterbalanced by cell wall properties, such as the orientation of microfibrils. This in turn is thought to be regulated by cortical microtubules, one of the major components of cytoskeletal elements. It is be- lieved that phytohormones, for example, gibberellin (GA) and auxin, in turn regulate the orientation of the cortical mi- crotubules (Giddings and Staehelin, 1991; Shibaoka, 1991). Compared with those of vegetative organs, the shape and size of floral organs are highly invariable. This suggests that their shape is under strict developmental control. During flower development, floral organs typically assume their final shape after mitotic activity has basically ceased, indicating that cell expansion plays an important role in determining organ shape (Pyke et al., 1991; Tsuge et al., 1996). For ex- ample, during corolla development in petunia and Antirrhi- num, the induction of anthocyanin synthesis correlates with the end of cell division (Bianchi et al., 1978; Doodeman et al., 1985; Coen et al., 1986). In petunia, the unfolded flower is only 40% of its final length when cells cease to divide (Martin and Gerats, 1993). From an experimental point of view, the high degree of regularity of floral organ shape is advantageous for studying the basis of organogenesis in plants by using molecular and genetic approaches. In Gerbera ( Gerbera hybrida: Aster- aceae), the most prominent part of the corolla is the bladelike ligule, which results from a fusion of three petal lobes (Helariutta et al., 1993; Bremer, 1994). The styles of carpels are fine and elongated nonphotosynthetic struc- tures. By using differential hybridization of a corolla cDNA li- brary, we isolated a cDNA, GEG (for Ge rbera homolog of G AST1 gene). We propose that GEG plays a role in the reg- ulation of cell shape during corolla and carpel development in Gerbera. GEG expression both spatially and temporally correlates with the opening of the corolla and with cessation of corolla elongation. In the carpel, induction of GEG ex- pression coincides with the cessation of style elongation. In 1 To whom correspondence should be addressed. E-mail mika. [email protected]; fax 358-9-70859366.
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of GEG Participates in the Regulation of Cell and Organ Shape during Corolla and Carpel Development in...

The Plant Cell, Vol. 11, 1093–1104, June 1999, www.plantcell.org © 1999 American Society of Plant Physiologists
GEG
Participates in the Regulation of Cell and Organ Shape during Corolla and Carpel Development in
Gerbera hybrida
Mika Kotilainen,
a,1
Yrjö Helariutta,
a
Merja Mehto,
a
Eija Pöllänen,
a
Victor A. Albert,
b
Paula Elomaa,
a
and Teemu H. Teeri
a
a
Institute of Biotechnology, Viikki Biocenter, P.O. Box 56, 00014–University of Helsinki, Finland
b
Lewis B. and Dorothy Cullman Program for Molecular Systematics Studies, New York Botanical Garden, Bronx, New York 10458-5126
The molecular mechanisms that control organ shape during flower development are largely unknown. By using differ-ential hybridization techniques, a cDNA designated
GEG
(for
Gerbera hybrida
homolog of the gibberellin [GA]–stimu-lated transcript 1 [
GAST1
] from tomato) was isolated from a library representing late stages of corolla development inGerbera.
GEG
expression was detected in corollas and carpels, with expression spatiotemporally coinciding withflower opening. In corollas and styles,
GEG
expression is temporally correlated with the cessation of longitudinal cellexpansion. In plants constitutively expressing
GEG
, reduced corolla lengths and carpels with shortened and radially ex-panded stylar parts were found, with concomitant reduction of longitudinal cell expansion in these organs. In addition,in styles, an increase in radial cell expansion was detected. Taken together, these observations indicate a regulatoryrole for the
GEG
gene product in determining the shape of the corolla and carpel. The deduced amino acid sequence ofthe
GEG
gene product shares high similarity with previously characterized putative cell wall proteins encoded by GA-inducible genes, namely, GAST1, GIP (for GA-induced gene of petunia), and the GASA (for GA-stimulated in Arabidopsis)gene family. Our studies suggest that
GEG
, the expression of which can also be induced by application of GA
3
, plays arole in phytohormone-mediated cell expansion.
INTRODUCTION
Plant cells do not migrate during development as do animalcells, and organ shape is deteminated by the organized andregulated control of cell expansion together with cell division(Meyerowitz, 1997). Despite its importance, the molecularand genetic regulation of cell expansion is not well under-stood. Emerging data support a view that the direction andmagnitude of the enlargement of the primary cell wall largelydetermine the expansion pattern and thereby the finalshapes of the cells. Water uptake causes constant turgorpressure in plant cells, which is the driving force of cell ex-pansion. Turgor pressure is counterbalanced by cell wallproperties, such as the orientation of microfibrils. This inturn is thought to be regulated by cortical microtubules, oneof the major components of cytoskeletal elements. It is be-lieved that phytohormones, for example, gibberellin (GA)and auxin, in turn regulate the orientation of the cortical mi-crotubules (Giddings and Staehelin, 1991; Shibaoka, 1991).
Compared with those of vegetative organs, the shape andsize of floral organs are highly invariable. This suggests thattheir shape is under strict developmental control. Duringflower development, floral organs typically assume their final
shape after mitotic activity has basically ceased, indicatingthat cell expansion plays an important role in determiningorgan shape (Pyke et al., 1991; Tsuge et al., 1996). For ex-ample, during corolla development in petunia and Antirrhi-num, the induction of anthocyanin synthesis correlates withthe end of cell division (Bianchi et al., 1978; Doodeman etal., 1985; Coen et al., 1986). In petunia, the unfolded floweris only 40% of its final length when cells cease to divide(Martin and Gerats, 1993).
From an experimental point of view, the high degree ofregularity of floral organ shape is advantageous for studyingthe basis of organogenesis in plants by using molecular andgenetic approaches. In Gerbera (
Gerbera hybrida
: Aster-aceae), the most prominent part of the corolla is thebladelike ligule, which results from a fusion of three petallobes (Helariutta et al., 1993; Bremer, 1994). The styles ofcarpels are fine and elongated nonphotosynthetic struc-tures. By using differential hybridization of a corolla cDNA li-brary, we isolated a cDNA,
GEG
(for Gerbera homolog of
GAST1
gene). We propose that
GEG
plays a role in the reg-ulation of cell shape during corolla and carpel developmentin Gerbera.
GEG
expression both spatially and temporallycorrelates with the opening of the corolla and with cessationof corolla elongation. In the carpel, induction of
GEG
ex-pression coincides with the cessation of style elongation. In
1
To whom correspondence should be addressed. E-mail [email protected]; fax 358-9-70859366.

1094 The Plant Cell
transgenic plants constitutively expressing
GEG
, corollasare shorter compared with those of nontransformed lines.Similar to that in the corolla, constitutive
GEG
expressioncauses shortening of the carpel but a concomitant radial ex-pansion of the style. We found that epidermal cells of boththe ligular part of the corolla and the style are reduced inlength along organ axes. Radial expansion of the epidermalcells in styles was also observed.
A database search showed that
GEG
belongs to a genefamily encoding putative small cell wall proteins with a cys-teine-rich domain and a putative signal peptide sequence.This family includes GAST1 (for GA-stimulated transcript 1)of tomato, GASA-1 (for GA stimulated in Arabidopsis), GIP(for GA-induced gene of petunia), and RSI-1 (for root systeminducible 1) of tomato (Shi et al., 1992; Taylor and Scheuring,1994; Herzog et al., 1995; Ben-Nissan and Weiss, 1996).
GEG
expression was experimentally induced by treatment with GA
3
,which is similar to previous reports indicating that thesegenes are GA or auxin inducible. We hypothesize that
GEG
is part of a phytohormone-mediated cell expansion mecha-nism that functions during corolla and carpel development.
RESULTS
Isolation of a
GEG
cDNA by Differential Screening
Both spatially restricted anthocyanin pigmentation patterns(Helariutta et al., 1993) and gene product accumulation pat-terns (Y. Helariutta, unpublished data) in various regions ofthe Gerbera ray floret corolla indicate region-specific controlof gene expression along the longitudinal axis of the corolla.We performed several differential screening schemes alongthis axis during the late stages of corolla morphogenesis toisolate genes with differential expression within the corolla.In this context, we isolated a cDNA as a clone whose ex-pression is stronger in the proximal part of the corolla thanin the distal part. Based on high similarity to
GAST1
-likegenes, we have named this cDNA
GEG
.Sequence comparisons show that
GEG
belongs to a fam-ily of genes that are transcriptionally regulated by phytohor-mones in different plants. The predicted GEG protein hashigh sequence similarity with proteins encoded by geneswhose expression is induced by GA (
GAST1
of tomato,
GASA1-4
of Arabidopsis, and
GIP
of petunia) or by auxin(
RSI-1
of tomato) (Shi et al., 1992; Taylor and Scheuring,1994; Herzog et al., 1995; Ben-Nissan and Weiss, 1996). Allof the derived polypeptides have a putative signal sequenceat their N termini, with cleavage sites predicted according tovon Heijne (1986). Because other targeting signals have notbeen identified, it has been proposed that these gene prod-ucts are targeted into the extracellular space or into the cellwall. Among 60 C-terminal amino acids, there are 22 identi-cal residues of which 12 are cysteines.
During DNA gel blot analysis, the probe (a 259-bp 3
9
frag-ment of the
GEG
cDNA, 90% noncoding) recognized one ortwo bands at the stringency used for RNA gel blotting. Thismost probably indicates that the expression analysis resultspresented below correspond to transcription of a single lo-cus and that the two bands found in some digests were dueto restriction length polymorphism in the heterozygous culti-var. During low-stringency DNA gel blot analysis, the full-length
GEG
cDNA probe recognized more bands, suggest-ing that there is a small gene family of
GEG
-like genes in theGerbera genome (data not shown).
GEG
mRNA Is Abundant in Corollas and Carpels
The developmental expression pattern of
GEG
was studiedby using RNA gel blot analyses. The expression of
GEG
ishighest in floral organs; in addition, a faint signal was de-tected in RNA from leaf blades. Strong
GEG
expression wasobserved in corolla tissue (both tube and ligule regions) andcarpels, with more moderate signals in the scape (floralstem) and the receptacle (terminal enlargement of floralstem) (Figure 1). To understand the role of GEG in plant de-velopment, we focused on
GEG
expression in corollas andcarpels.
GEG
expression was studied temporally at differentstages of ray floret corolla development (Figures 2A to 2F;Helariutta et al., 1993). The expression correlates temporallywith opening of both individual florets and the whole inflo-rescence, being induced at stage 7 (Figure 2G). Because of
Figure 1. RNA Gel Blot Hybridization Analysis Showing the TissueSpecificity of GEG Expression.
Autoradiography of an RNA gel blot probed with a 259-bp 39 frag-ment of the GEG cDNA (90% noncoding). Fifteen micrograms of to-tal RNA was loaded per lane, and equal loading was confirmed byethidium bromide staining. Organs covering several developmentalstages were examined. Receptacle, terminal enlargement of floralstem; scape, floral stem.

GAST1-like Gene Regulates Cell Shape 1095
the large size of ray floret corollas in Gerbera, we were ableto isolate RNA from various parts of the corolla over timeand study the developmental induction pattern of
GEG
expression by using RNA gel blot analysis. The pattern is in-triguing. Just before the opening of the flower—and unfold-ing of the corolla—the onset of
GEG
expression occursalmost simultaneously from both ends of the corolla. Thevery first signal can be seen in the proximal part of the co-rolla, more precisely, in the joint region of tube and ligule(Figure 2H, stage 7). During opening, the proximal expres-sion proceeds in both directions: basipetally into the tubeand acropetally into the ligule. Almost simultaneous to theonset of proximal expression,
GEG
expression also startsfrom the distal end of the corolla. This expression proceedsbasipetally toward the middle of the ligule, where both prox-imal and distal expression domains meet just as the corollahas opened (Figure 2H, stage 8).
GEG
expression continuesat a high level until senescence takes place (data notshown). In situ hybridization analysis of the
GEG
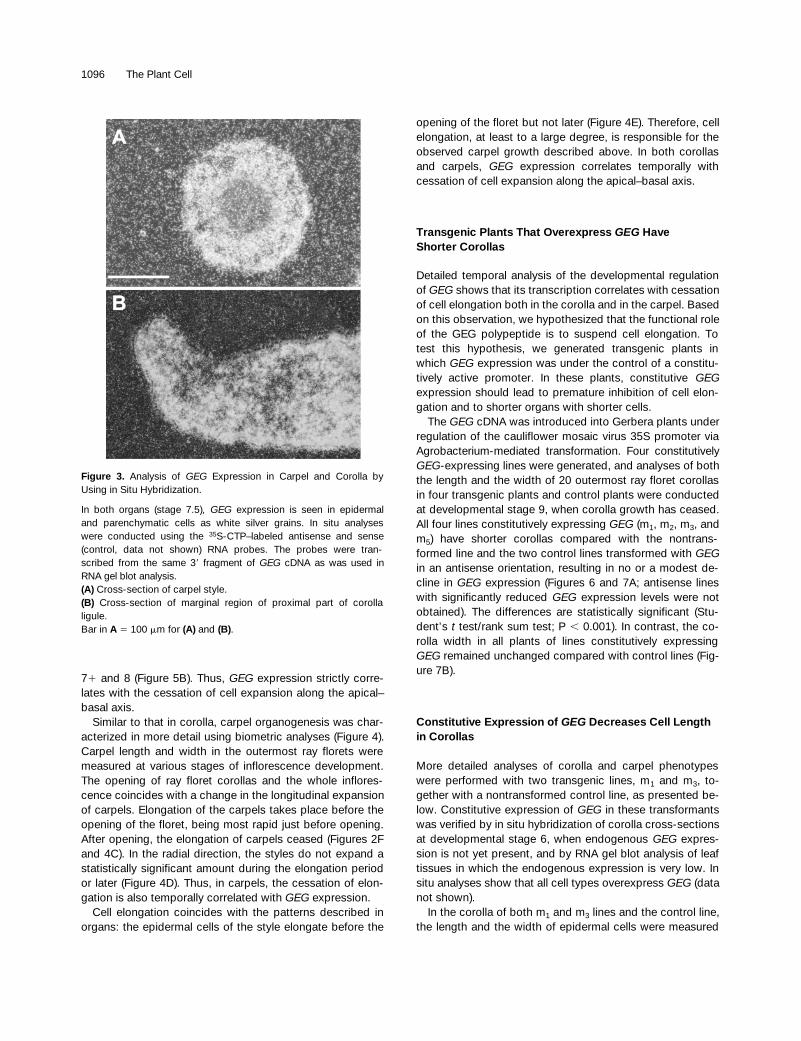
expressionpattern revealed that the transcript can be detected both inthe mesophyll and in the epidermis during opening of thecorolla (Figure 3B).
In carpels,
GEG
expression was studied by gel blottingusing RNA from samples taken before and after the openingof ray florets at developmental stages 6 and 8, respectively(Figures 2C to 2F; Helariutta et al. 1993). As shown in Figure2G, the onset of
GEG
expression coincides with the openingof the florets. In the style,
GEG
mRNA was detected in theouter epidermis and in the parenchyma (cortex) but not intransmitting tissues (Figure 3A).
Temporal Correlation of
GEG
Expression with Cessation of Organ and Cell Elongation in Corollas and Carpels
Biometric analyses (Figure 4) of corolla growth show thatbefore its opening, the corolla expands both longitudinallyand laterally. Soon after opening, growth ceases in both di-rections (Figures 4A and 4B). Temporally,
GEG
expressionfollows tightly the cessation of corolla growth, and it can bedetected everywhere in corolla tissues just after opening(Figures 2D to 2F and 2H).
The temporal pattern of
GEG
expression along the apical–basal axis of the corolla made it important to analyze whether
GEG
expression correlates with cessation of cell elongation.Cell length was measured in the distal and central regions ofcorolla (Figure 2H, regions 7 and 5, respectively) just afterstage 7 (7
1
) and at stage 8. At these stages,
GEG
mRNA ispresent in the distal region but reaches the central regionjust before stage 8 (Figure 2H). Cell length measurements re-vealed that cells in the distal region do not elongate,whereas in the central region, axial cell elongation takesplace (Figure 5A). The cell length differences between stage8 middle cells and other groups are statistically significant(rank sum tests; P
,
0.001). Cell width growth was detectedboth in distal and middle parts of the corolla between stages
Figure 2. Analysis of GEG Expression during Corolla and CarpelDevelopment.
(A) to (F) Different developmental stages of Gerbera inflorescence,according to Helariutta et al. (1993). (A) shows developmental stage1; (B), stage 3; (C), stage 5; (D), stage 7; (E), stage 7.5; and (F),stage 8.(G) The expression of GEG in carpel and corolla correlates withopening of both individual ray florets and the whole inflorescence.Numbers refer to developmental stage.(H) Spatial partition of ray floret corolla. Regions are indicated abovethe gel. The onset of expression occurs from both ends of the co-rolla (stage 7, regions 2 and 7) and just as the corolla has opened.Both expression domains meet at the middle of the ligule (stage 8,region 5). Region 1 is the tubular part of ray floret corolla (tube); re-gions 3 (proximal region) to 7 (distal region) represent the ligular partof the corolla. GDFR1 is used as a loading control. The developmen-tal stages (st.) are the same as presented in (A) to (F).
1096 The Plant Cell
7
1
and 8 (Figure 5B). Thus,
GEG
expression strictly corre-lates with the cessation of cell expansion along the apical–basal axis.
Similar to that in corolla, carpel organogenesis was char-acterized in more detail using biometric analyses (Figure 4).Carpel length and width in the outermost ray florets weremeasured at various stages of inflorescence development.The opening of ray floret corollas and the whole inflores-cence coincides with a change in the longitudinal expansionof carpels. Elongation of the carpels takes place before theopening of the floret, being most rapid just before opening.After opening, the elongation of carpels ceased (Figures 2Fand 4C). In the radial direction, the styles do not expand astatistically significant amount during the elongation periodor later (Figure 4D). Thus, in carpels, the cessation of elon-gation is also temporally correlated with
GEG
expression.Cell elongation coincides with the patterns described in
organs: the epidermal cells of the style elongate before the
opening of the floret but not later (Figure 4E). Therefore, cellelongation, at least to a large degree, is responsible for theobserved carpel growth described above. In both corollasand carpels,
GEG
expression correlates temporally withcessation of cell expansion along the apical–basal axis.
Transgenic Plants That Overexpress
GEG
HaveShorter Corollas
Detailed temporal analysis of the developmental regulationof
GEG
shows that its transcription correlates with cessationof cell elongation both in the corolla and in the carpel. Basedon this observation, we hypothesized that the functional roleof the GEG polypeptide is to suspend cell elongation. Totest this hypothesis, we generated transgenic plants inwhich
GEG
expression was under the control of a constitu-tively active promoter. In these plants, constitutive
GEG
expression should lead to premature inhibition of cell elon-gation and to shorter organs with shorter cells.
The
GEG
cDNA was introduced into Gerbera plants underregulation of the cauliflower mosaic virus 35S promoter viaAgrobacterium-mediated transformation. Four constitutively
GEG
-expressing lines were generated, and analyses of boththe length and the width of 20 outermost ray floret corollasin four transgenic plants and control plants were conductedat developmental stage 9, when corolla growth has ceased.All four lines constitutively expressing
GEG
(m
1
, m
2
, m
3
, andm
5
) have shorter corollas compared with the nontrans-formed line and the two control lines transformed with
GEG
in an antisense orientation, resulting in no or a modest de-cline in
GEG
expression (Figures 6 and 7A; antisense lineswith significantly reduced
GEG
expression levels were notobtained). The differences are statistically significant (Stu-dent’s
t
test/rank sum test; P
,
0.001). In contrast, the co-rolla width in all plants of lines constitutively expressing
GEG
remained unchanged compared with control lines (Fig-ure 7B).
Constitutive Expression of
GEG
Decreases Cell Length in Corollas
More detailed analyses of corolla and carpel phenotypeswere performed with two transgenic lines, m
1
and m
3
, to-gether with a nontransformed control line, as presented be-low. Constitutive expression of
GEG
in these transformantswas verified by in situ hybridization of corolla cross-sectionsat developmental stage 6, when endogenous
GEG
expres-sion is not yet present, and by RNA gel blot analysis of leaftissues in which the endogenous expression is very low. Insitu analyses show that all cell types overexpress
GEG
(datanot shown).
In the corolla of both m
1
and m
3
lines and the control line,the length and the width of epidermal cells were measured
Figure 3. Analysis of GEG Expression in Carpel and Corolla byUsing in Situ Hybridization.
In both organs (stage 7.5), GEG expression is seen in epidermaland parenchymatic cells as white silver grains. In situ analyseswere conducted using the 35S-CTP–labeled antisense and sense(control, data not shown) RNA probes. The probes were tran-scribed from the same 39 fragment of GEG cDNA as was used inRNA gel blot analysis.(A) Cross-section of carpel style.(B) Cross-section of marginal region of proximal part of corollaligule.Bar in A 5 100 mm for (A) and (B).

GAST1-like Gene Regulates Cell Shape 1097
at the central part of the proximal end of the ray floret corollaligules on their adaxial sides at developmental stage 8 (Fig-ure 8A). In plants constitutively expressing GEG, cell lengthwas reduced in a statistically significant manner (Student’s ttest; P , 0.001), but no difference in cell width could bemeasured (Figures 8B, 8C, 9A, and 9B). In conclusion, themajor impact of constitutive GEG expression is the cessa-tion of the axial cell expansion of epidermal cells in theligule.
Epidermal Cells of the Style are Shorter and Wider in Lines Constitutively Expressing GEG
Compared with the control line, transgenic lines constitu-tively expressing GEG had a decrease in carpel length andan increase in carpel radius (Figures 10A, 10B, 11A, and11B). A comparison of cell length and the width of style epi-dermal cells between m1 and m3 lines constitutively ex-pressing GEG and the control line revealed a change inelongation pattern. Even before endogenous expression, atstage 6, statistically significant changes of cell length andwidth could be detected (Student’s t test; P , 0.001). In m1
and m3 lines constitutively expressing GEG, cell length wasreduced and the width was increased compared with thecontrol line (Figures 10A, 10B, 11C, and 11D). The constitu-tive expression phenotypes support the view that the GEGgene product regulates cell expansion in the axial dimensionduring carpel development as well as during corolla devel-opment. However, in the carpel, unlike in the corolla, weobserved a concomitant opposite effect in the radial dimen-sion.
Figure 4. Biometric Analysis of Corolla and Carpel Length andWidth Development.
(A) and (B) Corolla length and width, respectively. Temporal GEG
expression tightly follows the cessation of corolla expansion inboth longitudinal and lateral directions (see also Figure 2) .(C) Carpel length. In the carpel, cessation of elongation tempo-rally correlates with GEG expression.(D) Carpel width. Carpel styles do not expand in radial directionduring or after the elongation period.(E) Carpel cell length. In the carpel, the epidermal cells of the styleelongate before opening of the floret but not later; thus, the retar-dation of cell elongation correlates with GEG expression.Timing of different developmental stages (described by Helariutta etal., 1993) was measured by following the development of .50 inflo-rescences under our standard greenhouse conditions. The lengthsand widths of both corolla and carpel, shown in (A) to (D), weremeasured from 15 to 30 outermost ray florets at each developmen-tal stage. Samples were collected from at least two different inflo-rescences. Carpel cell length (E) of 72 epidermal cells at each timepoint was measured. Cell lengths of 18 epidermal cells 200 to 400mm below the stigma of each carpel were determined, and the aver-age cell length of four carpels was measured. Numbers below thecurves correspond to the development stage of the inflorescence(see Figures 2A to 2F). Error bars indicate the standard deviation.
1098 The Plant Cell
GA and Regulation of GEG Expression in the Corolla
Because all of the homologous genes (see Introduction) areinduced by phytohormones and because expression is spa-tially and temporally regulated, we studied whether GEGexpression reacts to GA3. Application of GA3 upregulated GEGexpression in ray floret corollas of detached inflorescences(Figure 12). A short pulse of GA3 was able to induce GEG in2 hr. However, because maximal levels of GEG mRNA werenot seen earlier than 24 hr after GA3 application, it is possi-ble that GA stimulation of GEG expression is indirect or thata decline (rather than an increase) in GA concentration in-duces GEG expression.
We have also isolated the genomic 59 flanking sequenceof GEG. It contains two sequence motifs that are found inthe flanking regions of rice and barley a-amylase geneswhose expression is regulated by GA (Huang et al., 1990;Skriver et al., 1991). This further supports the idea that GEGexpression is developmentally regulated by GA.
DISCUSSION
GEG Participates in the Regulation of Cell Shape
The expression pattern of GEG correlates with completionof organ and cell elongation in both corollas and carpels.Furthermore, in transgenic plants, constitutive GEG expres-sion demonstrates that excessive GEG production is able tocause alterations in organ and cell shape during corolla andcarpel development. This suggests that GEG plays a role in
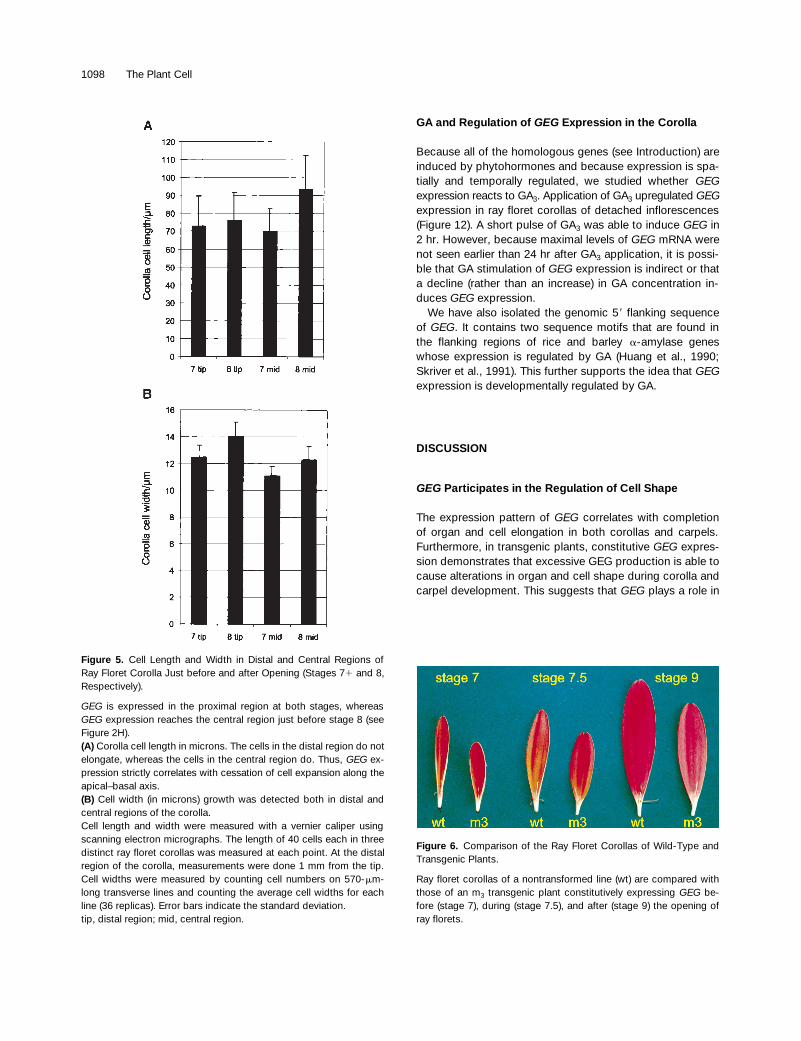
Figure 5. Cell Length and Width in Distal and Central Regions ofRay Floret Corolla Just before and after Opening (Stages 71 and 8,Respectively).
GEG is expressed in the proximal region at both stages, whereasGEG expression reaches the central region just before stage 8 (seeFigure 2H).(A) Corolla cell length in microns. The cells in the distal region do notelongate, whereas the cells in the central region do. Thus, GEG ex-pression strictly correlates with cessation of cell expansion along theapical–basal axis. (B) Cell width (in microns) growth was detected both in distal andcentral regions of the corolla.Cell length and width were measured with a vernier caliper usingscanning electron micrographs. The length of 40 cells each in threedistinct ray floret corollas was measured at each point. At the distalregion of the corolla, measurements were done 1 mm from the tip.Cell widths were measured by counting cell numbers on 570-mm-long transverse lines and counting the average cell widths for eachline (36 replicas). Error bars indicate the standard deviation.tip, distal region; mid, central region.
Figure 6. Comparison of the Ray Floret Corollas of Wild-Type andTransgenic Plants.
Ray floret corollas of a nontransformed line (wt) are compared withthose of an m3 transgenic plant constitutively expressing GEG be-fore (stage 7), during (stage 7.5), and after (stage 9) the opening ofray florets.

GAST1-like Gene Regulates Cell Shape 1099
determining cell shape during carpel and corolla morpho-genesis, thus providing functional information for the role ofGEG-like genes in plants.
In carpels, constitutive GEG expression reveals a negativeinterrelationship between longitudinal and radial growth. Asdescribed above, this is also evident in epidermal cells.However, in corollas, no radial expansion of epidermal cellsdue to constitutive GEG expression was observed. Further-more, in carpels, no increase in style width was observed
Figure 7. Analysis of Corolla Length and Width for Four LinesConstitutively Expressing GEG and for the Control Lines.
(A) Corolla length (in millimeters). Length of the outermost ray flo-ret corollas of wild-type plants (wt), constitutively GEG-expressinglines (m1, m3, m2, and m5) and two GEG antisense lines with no (t4) ora modest (t9; 80% remaining) decline of GEG expression. All fourlines constitutively expressing GEG have statistically shorter corollascompared with the control lines.(B) The corolla width (in millimeters) of all lines constitutively ex-pressing GEG remained unchanged when compared with controllines. Forty outermost ray floret corollas of two inflorescences ofeach line at stage 9 were collected. Corolla width and length offour lines constitutively expressing GEG (m1, m3, m2, and m5), anontransformed line (wt), and two antisense lines (with no ormodest effect in GEG expression) were measured.Error bars indicate the standard deviations.
Figure 8. Analysis of the Effects of Constitutive GEG Expression onCorolla Epidermal Cells.
(A) The epidermal cells are organized into longitudinal files runningalong the apical–basal axis of the corolla. Measurements of cell di-mensions took place in the area marked with a white box.(B) Scanning electron microscopy of the adaxial (upper) side of theproximal part of a ray floret corolla of a nontransformed control line(stage 8). One of the epidermal cells is highlighted in yellow.(C) Scanning electron microscopy of the corresponding region of anm1 line constitutively expressing GEG.Bar beneath (C) 5 75 mm for (B) and (C).
1100 The Plant Cell
during the endogenous GEG expression stage (Figure 4D).This suggests that the primary role of GEG is to inhibit cellelongation. According to this hypothesis, constitutive GEGexpresssion prematurely inhibits cell expansion in the longi-tudinal direction. This could allow the growth potential of thecell to be directed passively in the radial direction, as seen inthe epidermal cells of the style. The alternative hypothesisthat GEG would promote radial and inhibit longitudinal ex-pansion simultaneously is less likely. To distinguish betweenthese hypotheses, we need to study GEG function at a cel-lular level.
The angustifolia and rotundifolia mutations for leaf devel-opment (Tsuge et al., 1996) and several cell expansion mu-tations for root development in Arabidopsis (Aeschbacher etal., 1995; Hauser et al., 1995) point to an interrelationship
between the longitudinal and radial dimensions of the cell.For example, the palisade cells of leaves in the angustifoliamutant exhibit restricted expansion in the leaf-width di-mension and enhanced expansion in leaf thickness. Inrotundifolia3, leaf morphology is affected in one directiononly: cell elongation is reduced in the leaf-length (axial) di-mension (Tsuge et al., 1996).
In our analyses, we have measured the dimensions of epi-dermal cells by using scanning electron microscopy. Be-cause GEG expression was also observed in the underlyingparenchymatic cells, it is probable that analogous changesin cell shape are occurring there. The parallels between cellexpansion and organ expansion are evident, although it ispossible that in addition to cell expansion, cell divisionevents could also contribute to the determination of the finalshape at the stages analyzed. However, during the stagesthat we have investigated, anthocyanin accumulation andflavonoid biosynthetic gene expression, which have beenconsidered as markers for postmitotic cell differentiation,occurred (Martin and Gerats, 1993).Figure 9. Cell Lengths and Widths in the Same Regions as De-
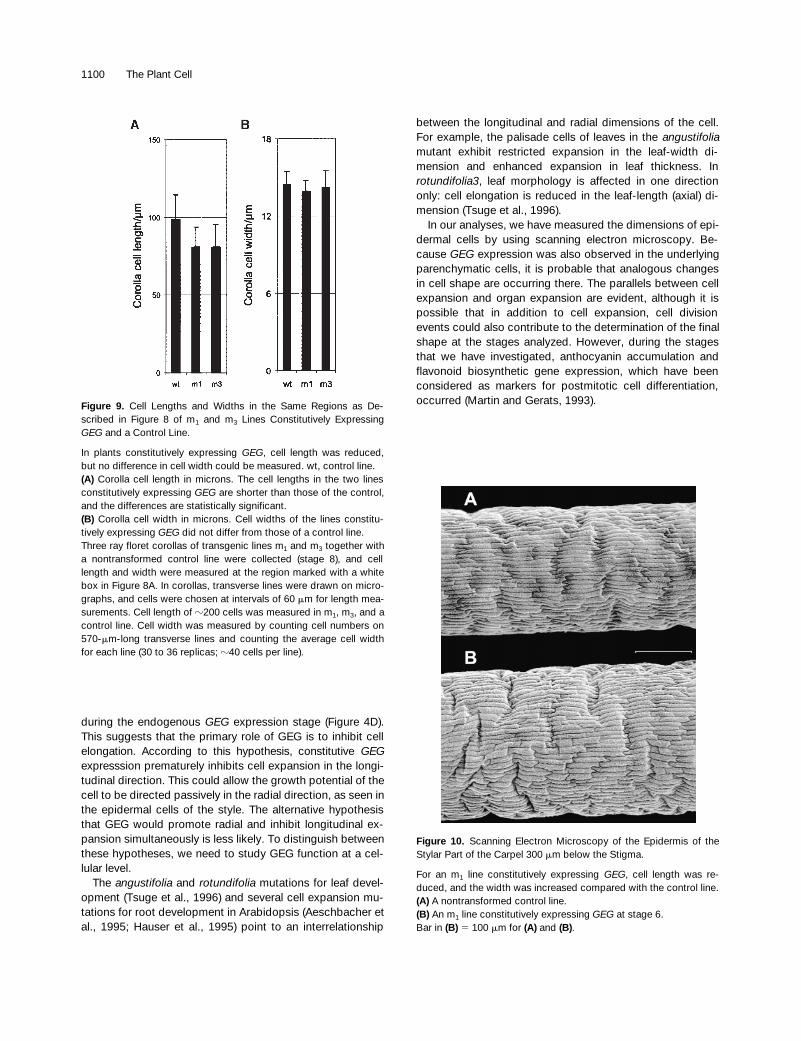
scribed in Figure 8 of m1 and m3 Lines Constitutively ExpressingGEG and a Control Line.
In plants constitutively expressing GEG, cell length was reduced,but no difference in cell width could be measured. wt, control line.(A) Corolla cell length in microns. The cell lengths in the two linesconstitutively expressing GEG are shorter than those of the control,and the differences are statistically significant.(B) Corolla cell width in microns. Cell widths of the lines constitu-tively expressing GEG did not differ from those of a control line.Three ray floret corollas of transgenic lines m1 and m3 together witha nontransformed control line were collected (stage 8), and celllength and width were measured at the region marked with a whitebox in Figure 8A. In corollas, transverse lines were drawn on micro-graphs, and cells were chosen at intervals of 60 mm for length mea-surements. Cell length of z200 cells was measured in m1, m3, and acontrol line. Cell width was measured by counting cell numbers on570-mm-long transverse lines and counting the average cell widthfor each line (30 to 36 replicas; z40 cells per line).
Figure 10. Scanning Electron Microscopy of the Epidermis of theStylar Part of the Carpel 300 mm below the Stigma.
For an m1 line constitutively expressing GEG, cell length was re-duced, and the width was increased compared with the control line.(A) A nontransformed control line.(B) An m1 line constitutively expressing GEG at stage 6.Bar in (B) 5 100 mm for (A) and (B).

GAST1-like Gene Regulates Cell Shape 1101
GEG Homologs in Plants
The genes orthologous to GEG have been described earlierin various plants, for example, GAST1 (tomato), GASA1-4(Arabidopsis), GIP (petunia), and RSI-1 (tomato) (Shi et al.,1992; Taylor and Scheuring, 1994; Herzog et al., 1995; Ben-Nissan and Weiss, 1996). The GEG-like gene/protein familyshares several features that may suggest a role for GEG inregulating cell expansion. Based on our studies of GEG ex-pression and the fact that several members (GEG, GAST1,and RSI-1) have been isolated by using differential screen-ing methods, we conclude that the mRNA is relatively abun-dant, characteristic of a structural role for the gene product.Furthermore, the putative signal sequence and the absenceof other targeting signals suggest that the gene products aresecreted, possibly to the cell wall (Shi et al., 1992). Anothercharacteristic feature is regulation of gene expression withphytohormones. The variability in the effective hormone in-dicates that the role of the genes may be downstream ofvarious signal transduction pathways after their conver-gence. Taken together with the data from transgenic plantssuggesting that the primary role of GEG may be inhibitingaxial cell expansion, it is possible that the GEG-like functionmay be generally related to establishing cell wall propertiesduring organogenesis in plants.
GEG-like gene products are characterized by a highlyconserved C-terminal domain, with 12 invariable cysteineresidues and an N-terminal domain variable in length andhydrophobicity. Because the genes are expressed in differ-ent developmental contexts, this might reflect involvementin similar but not necessarily identical processes. However,phylogenetic analysis using the nucleotide sequences corre-sponding to 60 amino acids of the conserved C terminusdoes not reveal subgroups that might reflect split functionalrelationships (data not shown).
No information about the function of these proteins hasbeen reported until this study, but we can compare the othermembers of the gene family to GEG concerning their geneexpression. Similar to GEG, two other members (GIP andGASA4) are expressed during corolla or carpel organogene-sis. GIP expression in petunia corollas is reported to be thehighest just before anthesis (stage 5). When the corollareached maximum size (stage 7), no transcript was detected(Ben-Nissan and Weiss, 1996). Thus, in contrast to GEG,GIP expression is transient and temporally correlates withthe cell elongation period of corolla growth instead of itscompletion. If the role of GEG as a negative regulator of cellelongation is representative for GIP, the latter could be in-volved in preventing overelongation during cell expansion.However, there is also a requirement for competence to re-spond to GEG, because constitutive expression of GEGdoes not lead to a phenotype in all organs. In Gerbera, thecompetence may rise before GEG expression, but in petu-nia, GIP expression may be present before the competencedevelops, leading to a different timing of gene expression.Alternatively, GEG and GIP may be involved in the same
Figure 11. Analysis of the Effects of Constitutive GEG Expressionon Epidermal Cells of the Carpel Style.
(A) Carpel length in millimeters. Carpel length of lines constitutivelyexpressing GEG is shorter compared with that of a control line.(B) Carpel width in microns. Compared with that of a control line, thecarpel radius has increased in lines constitutively expressing GEG.(C) Carpel cell length in microns. Comparison of cell length of m1
and m3 with a nontransformed control line reveals that epidermalcells of the style of lines constitutively expressing GEG are shorterthan those of a control line.(D) Carpel cell width in microns. The epidermal cells of constitutivelyGEG-expressing lines are wider than those of a control line.Twelve to 20 carpels of two lines constitutively expressing GEG (m1
and m3) and a nontransformed control line (wt) were measured. Car-pel length was measured at developmental stage 9; carpel width,epidermal cell length, and width were measured at stage 6. Celllength of 18 individual epidermal cells 200 to 400 mm below thestigma of each carpel and the average cell length of 12 to 20 carpelsof each line were measured. The differences presented in (A), (B),(C), and (D) are statistically significant. Error bars indicate the stan-dard deviations.

1102 The Plant Cell
process but act in an opposite manner. In this scheme, forexample, the N-terminal variable domain of putative proteinscould be responsible for allowing GEG to inhibit and GIP topromote cell elongation.
During style development, the expression pattern of Ara-bidopsis GASA4 has temporal and spatial similarities withGEG. GASA4 expression was also detected in the short sty-lar region of gynoecium at the stage during which thegrowth rate of overall gynoecium length ceases (Bowman,1994; Aubert et al., 1998). Therefore, GASA4 could have arole similar to GEG in regulating cell elongation in style de-velopment.
However, Aubert et al. (1998) reported that GASA4 in Ara-bidopsis is expressed in regions undergoing rapid cell divi-sion in various organs. Recently, Sablowski and Meyerowitz(1998) reported on a gene (NO APICAL MERISTEM, NAP)whose constitutive expression, like that of GEG, results inthe inhibition of cell elongation. However, it was concludedthat NAP probably functions in the transition between celldivision and cell expansion during stamen and petal devel-opment, because in the absence of NAP this transition doesnot occur, and constitutive NAP expression delays theswitch. The transient expression pattern of NAP betweenstages of frequent cell division and cell expansion furthersupports a transition function (Sablowski and Meyerowitz,1998). In contrast to NAP expression, GEG expression dur-ing corolla and carpel development is clearly induced afterthe active cell division phase and occurs simultaneously tothe cessation of the elongation phase (as evidenced by theanthocyanin accumulation and Gerbera dihydroflavonol-4-
reductase gene 1 [GDFR1] expression at this stage; Figures2 and 4), indicating that GEG is an inhibitor of cell elonga-tion, as discussed above. If the inhibitory role of GEG in cellexpansion is representative of GASA4 in this developmentalcontext, then it is possible that the elongation of dividingcells is controlled by similar molecules as it is after the elon-gation stage.
GEG Belongs to a Class of Genes Abundantly Expressed in Styles and Corollas of Gerbera
We have previously isolated several genes from Gerberathat are functional during corolla morphogenesis. Interest-ingly, two of them have the same organ specificity as doesGEG, involving epidermal and/or parenchymatic cells of car-pel and corolla. In addition to GEG, this group of genes con-sists of GDFR1 and the nonspecific lipid transfer proteingene 1 (GLTP1) (Helariutta et al., 1993; Kotilainen et al.,1994). The style in Gerbera is nonphotosynthetic, differingfrom other well-studied species, and in addition has antho-cyanin pigmentation. Although the spatial and temporal ex-pression patterns of these genes are different, the datasuggest that similar genetic programs are occurring duringthe differentiation of the epidermal and parenchymatic celltypes of the corolla and style in Gerbera. It is likely thatthese differentiation-related molecules are regulated by pro-grams/factors that are independent from or converge afterthe determination of floral organ identity, because the ABCmodel predicts that corolla and carpel development are trig-gered by a nonoverlapping set of regulatory factors in thefloral meristem (Coen and Meyerowitz, 1991). One suchcandidate is GA, which has been previously shown to regu-late flavonoid biosynthetic genes and GAST1-like genes, in-cluding GEG, as has been described here.
METHODS
Plant Material
Gerbera hybrida var Terra Regina used in this research was obtainedfrom Terra Nigra BV (De Kwakel, Holland). The control and transgenicplants were grown under identical conditions (side by side) at thesame time, and the age of plants was the same. Developmentalstages of the inflorescence are described by Helariutta et al. (1993).For all analyses, samples were collected from outermost ray florets(flowers) of the inflorescence, and each transgenic and control plantsample was harvested and treated at the same time.
Plant Transformation
Gerbera transformation was performed using Agrobacterium tumefa-ciens–mediated gene transfer, as described previously (Elomaa et
Figure 12. GA Application Upregulates GEG Expression in Ray Flo-ret Corollas.
RNA gel blot showing GEG expression after the addition of GA3 justbefore the opening of the inflorescence (stage 71). The scape (floralstem) was cut 5 cm below the inflorescence.(A) The control inflorescences were grown on 50 mM sucrose.(B) Induction of GEG expression was detected when inflorescenceswere grown on 50 mM sucrose with 5 mm GA3.(C) Induction of GEG expression was also detected when inflores-cences were first incubated in 50 mM sucrose with 50 mm GA3 for 5min and then transferred to 50 mM sucrose medium.Approximately 10 ray floret corollas were collected for RNA isolationat each time point (indicated above gels in hours) after GA3 addition.

GAST1-like Gene Regulates Cell Shape 1103
al., 1993, 1998). Transformation was verified by RNA gel blot analysisshowing GEG (for Gerbera homolog of the gibberellin [GA]–stimu-lated transcript 1 [GAST1]) expression in leaves and by DNA gel blotanalysis. The analyses were performed on clones of the originaltransgenic plants (T0).
Isolation of Plant DNA and RNA
Plant DNA was isolated using the method of Dellaporta et al. (1983).Total RNA was isolated as described by Jones et al. (1985) or by us-ing the RNeasy plant total RNA kit (Qiagen, Chatsworth, CA).Poly(A)1 RNA was isolated using oligo(dT) cellulose affinity chroma-tography (Sambrook et al., 1989).
Construction and Differential Screening of a CorollacDNA Library
Polyadenylated RNA (5 mg) extracted from proximal parts of ray floretcorollas at developmental stages 5 to 9 (Helariutta et al., 1993) wasused to construct a cDNA library in the l ZAPII vector (ZAP-cDNAsynthesis kit; Stratagene, La Jolla, CA). From the nonamplified cDNAlibrary, z50,000 plaques were plated and transferred onto replicanylon membranes and then screened differentially with radiolabeledfirst-strand cDNA pools from the ray floret tube region of the proxi-mal and distal parts of the ligule (first-strand cDNA synthesis kit; Am-ersham).
GEG cDNA was isolated as a clone that is expressed morestrongly in the proximal part than in the distal part of the ligule. Twoindependent but similar cDNA clones were isolated, subcloned into apUC18 derivative, and sequenced using the AutoRead kit (Pharma-cia, Uppsala, Sweden). The 813-bp genomic fragment containingpart of the GEG promoter was obtained by applying a 59 rapid ampli-fication of cDNA ends–like polymerase chain reaction amplificationon genomic DNA. The GEG cDNA sequence and sequence of the 59
flanking region of the GEG gene have been submitted to the EMBLdatabase; the accession numbers are AJ005206 and AJ006273, re-spectively.
RNA Gel Blot Analyses and in Situ Hybridization
Fifteen micrograms of total RNA was loaded per lane. The amount ofRNA to be loaded was measured spectrophotometrically, and equalloading was confirmed by ethidium bromide staining of rRNA bands.Electrophoresis and hybridizations were as described by Sambrooket al. (1989). The 259-bp 39 fragment (of which 234 bp is from thenoncoding region) served as the probe. Washing conditions of 0.2 3SSC (1 3 SSC is 0.15 M NaCl and 0.015 M sodium citrate) and 0.1%SDS at 588C were applied for all RNA blots. In situ hybridization wasconducted as described previously by Kotilainen et al. (1994) using35S-CTP–labeled antisense and sense (control) RNA probes. Theprobes were transcribed from the same fragment as was used in thegel blot studies under the T7 promoter in vector pSP72/73 (SP6/T7transcription kit; Roche Diagnostics, Mannheim, Germany).
Scanning Electron Microscopic Analysis
Corolla and carpel samples of control and transgenic plants werecollected and further treated side by side at the same time. They
were fixed in FAA buffer (50% ethanol, 5% acetic acid, and 2% form-aldehyde) overnight and then transferred through an ethanol series to100% ethanol, critical point dried (Balzers CPD 030 critical pointdryer; Bal-Tec, Balzers, Liechtenstein), and coated with platinum/pal-ladium (agar sputter coater; Agar Scientific Ltd., Stansel, UK). Spec-imens were mounted on aluminium stubs using graphite adhesive ortape and examined with a scanning electron microscope (digitalscanning microscope, model DSM 962; Karl Zeiss, Oberkochen,Germany) in the Electron Microscopy Laboratory of the Institute ofBiotechnology, University of Helsinki.
Organ and Cell Measurements and Statistical Analysis
Organ length and width measurements were done with a vernier cal-iper in vivo, except for carpel width, which was measured from scan-ning electron micrographs 200 mm below the stigma. Cell length andwidth were measured by using scanning electron micrographs.
To study whether the differences in organ and cell expansioncaused by constitutive GEG expression are statistically significant,we performed Student’s t tests and/or rank sum tests. A parametrict test was used if the normality and equal variances of samples wereconfirmed (P values to reject , 0.050). A nonparametric rank sumtest was used if either was not confirmed. The level of confidence isP , 0.001 in all statistically significant differences mentioned in thisstudy.
ACKNOWLEDGMENTS
We thank Philip Benfey, Xuemei Chen, Pekka Lappalainen, and threeanonymous reviewers for valuable comments on the manuscript. Wethank Eija Takala, Marja Huovila, and Anu Immonen for excellenttechnical assistance. We also thank Eija Saarikko and Anne Aaltonenfor greenhouse care of plant material, Jyrki Juhanoja for his help inelectron microscopy, Tapio Linkosalo for advice on statistical analy-ses, and Jari Penttinen for editing the figures. This work was partiallyfunded by the Academy of Finland.
Received January 4, 1999; accepted March 29, 1999.
REFERENCES
Aeschbacher, R.A., Hauser, M.-T., Feldmann, K.A., and Benfey,P.N. (1995). The SABRE gene is required for normal cell expan-sion in Arabidopsis. Genes Dev. 9, 330–340.
Aubert, D., Chevillard, M., Dorne, A.-M., Arlaud, G., and Herzog,M. (1998). Expression patterns of GASA genes in Arabidopsisthaliana: The GASA4 gene is up-regulated by gibberellins in mer-istematic regions. Plant Mol. Biol. 36, 871–883.
Ben-Nissan, G., and Weiss, D. (1996). The petunia homologue oftomato GAST1: Transcript accumulation coincides with gibberel-lin-induced corolla cell elongation. Plant Mol. Biol. 32, 1067–1074.
Bianchi, F., Cornelissen, P.T.J., Gerats, A.G.M., and Hogervorst,J.M.W. (1978). Regulation of gene action in Petunia hybrida:

1104 The Plant Cell
Unstable alleles for a gene for flower color. Theor. Appl. Genet.53, 157–167.
Bowman, J. (1994). Arabidopsis, An Atlas of Morphology andDevelopment. (New York: Springer-Verlag).
Bremer, K. (1994). Asteraceae, Cladistics and Classification. (Port-land, OR: Timber Press).
Coen, E.S., and Meyerowitz, E.M. (1991). The war of whorls:Genetic interaction controlling flower development. Nature 353,31–37.
Coen, E.S., Carpenter, R., and Martin, C. (1986). Transposableelements generate novel patterns of gene expression in Antirrhi-num majus. Cell 47, 285–296.
Dellaporta, S.L., Wood, J., and Hicks, J.B. (1983). A plant DNAminipreparation: Version II. Plant Mol. Biol. Rep. 1, 19–21.
Doodeman, M., Bino, R.J., Uytewaal, B., and Bianchi, F. (1985).Genetic analysis of instability in Petunia hybrida. The effect ofenvironmental factors on the reversion rate of unstable alleles.Theor. Appl. Genet. 69, 489–495.
Elomaa, P., Honkanen, J., Puska, R., Seppänen, P., Helariutta,Y., Mehto, M., Kotilainen, M., Nevalainen, L. and Teeri, T.H.(1993). Agrobacterium-mediated transfer of antisense chalconesynthase cDNA to Gerbera hybrida inhibits flower pigmentation.Bio/Technology 11, 508–511.
Elomaa, P., Mehto, M., Kotilainen, M., Helariutta, Y., Nevalainen,L., and Teeri, T.H. (1998). A bHLH transcription factor mediatesorgan, region and flower type specific signals on dihydroflavonol-4-reductase (dfr) gene expression in the inflorescence of Gerberahybrida (Asteraceae). Plant J. 16, 93–100.
Giddings, T.H., and Staehelin, L.A. (1991). Microtubule-mediatedmicrofibril deposition: A reexamination of the hypothesis. In TheCytoskeletal Basis of Plant Growth and Form, C.W. Lloyd, ed(London: Academic Press) pp. 85–100.
Hauser, M.-T., Morikami, A., and Benfey, P.N. (1995). Conditionalroot expansion mutants of Arabidopsis. Development 121, 1237–1252.
Helariutta, Y., Elomaa, P., Kotilainen, M., Seppänen, P., andTeeri, T.H. (1993). Cloning of cDNA coding for dihydroflavonol-4-reductase (DFR) and characterization of dfr expression in thecorollas of Gerbera hybrida var. Regina (Compositae). Plant Mol.Biol. 22, 183–193.
Herzog, M., Dorne, A.-M., and Grellet, F. (1995). GASA, a gibberel-lin-regulated gene family from Arabidopsis thaliana related to thetomato GAST1 gene. Plant Mol. Biol. 27, 743–752.
Huang, N., Sutliff, T.D., Litts, J.C., and Rodriguez, R.L. (1990).Classification and characterization of the rice a-amylase multi-gene family. Plant Mol. Biol. 14, 655–668.
Jones, J.D.G., Duismuir, P., and Bedbrook, J. (1985). High levelexpression of introduced chimeric genes in regenerated trans-formed plants. EMBO J. 4, 2411–2418.
Kotilainen, M., Helariutta, Y., Elomaa, P., Paulin, L., and Teeri,T.H. (1994). A corolla- and carpel-abundant, non-specific lipidtransfer protein gene is expressed in the epidermis and paren-chyma of Gerbera hybrida var. Regina (Compositae). Plant Mol.Biol. 26, 971–978.
Martin, C., and Gerats, T. (1993). Control of pigment biosynthesisgenes during petal development. Plant Cell 5, 1253–1264.
Meyerowitz, E.M. (1997). Genetic control of cell division patterns indeveloping plants. Cell 88, 299–308.
Pyke, K.A., Marrison, J.L., and Leech, R.M. (1991). Temporal andspatial development of the cells of the expanding first leaf of Ara-bidopsis thaliana (L.). J. Exp. Bot. 42, 1407–1416.
Sablowski, R.W.M., and Meyerowitz, E.M. (1998). A homolog ofNO APICAL MERISTEM is an immediate target of the floralhomeotic genes APETALA3/PISTILLATA. Cell 92, 93–103.
Sambrook, J., Fritsch, E.F., and Maniatis, T. (1989). MolecularCloning: A Laboratory Manual. (Cold Spring Harbor, NY: ColdSpring Harbor Laboratory Press).
Shi, L., Gast, R.T., Gopalraj, M., and Olszewski, N.E. (1992). Char-acterization of a shoot-specific, GA3- and ABA-regulated genefrom tomato. Plant J. 2, 153–159.
Shibaoka, H. (1991). Microtubules and the regulation of cell mor-phogenesis by plant hormones. In The Cytoskeletal Basis of PlantGrowth and Form, C.W. Lloyd, ed (London: Academic Press), pp.159–168.
Skriver, K., Olsen, F.L., Rogers, J.C., and Mundy J. (1991). cis-Acting DNA elements responsive to gibberellin and its antagonistabscisic acid. Proc. Natl. Acad. Sci. USA 88, 7266–7270.
Taylor, B.H., and Scheuring, C.F. (1994). A molecular marker forlateral root initiation: The Rsi-1 gene of tomato (Lycopersiconesculentum Mill.) is activated in early lateral root primordia. Mol.Gen. Genet. 243, 148–157.
Tsuge, T., Tsukaya, H., and Uchimiya, H. (1996). Two independentand polarized processes of cell elongation regulate leaf bladeexpansion in Arabidopsis thaliana (L.) Heynh. Development 122,1589–1600.
von Heijne, G. (1986). A new method for predicting signal sequencecleavage sites. Nucleic Acids Res. 14, 4683–4690.

DOI 10.1105/tpc.11.6.1093 1999;11;1093-1104Plant Cell
Teemu H. TeeriMika Kotilainen, Yrjö Helariutta, Merja Mehto, Eija Pöllänen, Victor A. Albert, Paula Elomaa and
Gerbera hybridaDevelopment in Participates in the Regulation of Cell and Organ Shape during Corolla and CarpelGEG
This information is current as of June 27, 2015
References http://www.plantcell.org/content/11/6/1093.full.html#ref-list-1
This article cites 24 articles, 7 of which can be accessed free at:
Permissions https://www.copyright.com/ccc/openurl.do?sid=pd_hw1532298X&issn=1532298X&WT.mc_id=pd_hw1532298X
eTOCs http://www.plantcell.org/cgi/alerts/ctmain
Sign up for eTOCs at:
CiteTrack Alerts http://www.plantcell.org/cgi/alerts/ctmain
Sign up for CiteTrack Alerts at:
Subscription Information http://www.aspb.org/publications/subscriptions.cfm
is available at:Plant Physiology and The Plant CellSubscription Information for
ADVANCING THE SCIENCE OF PLANT BIOLOGY © American Society of Plant Biologists




















