
Identification and characterization of a prawn white spot syndrome virus gene that encodes an...
8
Identification and characterization of a prawn white spot syndrome virus gene that encodes an envelope protein VP31 Li Li, Xixian Xie, Feng Yang * Key Laboratory of Marine Biogenetic Resources, Third Institute of Oceanography, Xiamen, P.R. China Received 13 April 2005; returned to author for revision 9 May 2005; accepted 3 June 2005 Available online 14 July 2005 Abstract Based on a combination of SDS-PAGE and mass spectrometry, a protein with an apparent molecular mass of 31 kDa (termed as VP31) was identified from purified shrimp white spot syndrome virus (WSSV) envelope fraction. The resulting amino acid (aa) sequence matched an open reading frame (WSV340) of the WSSV genome. This ORF contained 783 nucleotides (nt), encoding 261 aa. A fragment of WSV340 was expressed in Escherichia coli as a glutathione S -transferase (GST) fusion protein with a 6His-tag, and then specific antibody was raised. Western blot analysis and the immunoelectron microscope method (IEM) confirmed that VP31 was present exclusively in the viral envelope fraction. The neutralization experiment suggested that VP31 might play an important role in WSSV infectivity. D 2005 Elsevier Inc. All rights reserved. Keywords: White spot syndrome virus; VP31; Envelope protein; The neutralization experiment Introduction White spot syndrome virus (WSSV) is a major shrimp pathogen that is highly virulent in penaeid shrimp and can also infect most species of crustacean (Lo et al., 1996a, 1996b; Chen et al., 1997; Flegel, 1997). The complete sequence of WSSV genome has now been published for three different isolates (Chen et al., 2002; van Hulten et al., 2001a; Yang et al., 2001; GenBank Accession Nos. AF332093, AF369029, AF440570 for viruses isolated from China, Thailand, and Taiwan, respectively). Recently, the International Committee on Taxonomy of Viruses approved a proposal to erect WSSV as the type species of the genus Whispovirus , family Nimaviridae (www.ncbi.nlm.nih.gov/ ICTVdb/Ictv/index.htm). To date, some major structural proteins have been identified (Huang et al., 2002b; Tsai et al., 2004). Among those proteins, VP28, VP26/P22, VP281, VP466, VP68, VP292, and VP124 were identified as viral envelope proteins by immunoelectron microscopy (Huang et al., 2002a, 2002b; Zhang et al., 2002a, 2002b, 2004). As far as functional research is concerned, VP28, VP281, VP466, and VP68 were suggested to play an important role in the systemic WSSV infection in shrimp by using an in vivo neutralization assay (van Hulten et al., 2001b; Wu et al., 2005), and VP28 was used for detection of WSSV in crustaceans (Yoganandhan et al., 2004); VP281, with a cell attachment RGD motif, was supposed to play an important role in mediating WSSV infectivity (Huang et al., 2002a); VP26 was reported to be capable of binding to actin or actin-associated proteins (Xie and Yang, 2005). In this paper, the purified WSSV virions, viral envelope fractions were separated by SDS-PAGE. Among these proteins, a 31 kDa protein (termed VP31) was analyzed using Nano-ESI MS/MS and further identified by Western blot and TEM (transmission electron microscopy) immuno- gold-labeling. In vivo neutralization assay was performed in this investigation to examine if VP31 can play a role in WSSV infection. 0042-6822/$ - see front matter D 2005 Elsevier Inc. All rights reserved. doi:10.1016/j.virol.2005.06.007 * Corresponding author. E-mail address: [email protected] (F. Yang). Virology 340 (2005) 125 – 132 www.elsevier.com/locate/yviro
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Identification and characterization of a prawn white spot syndrome virus gene that encodes an...

wwwelseviercomlocateyviro
Virology 340 (20
Identification and characterization of a prawn white spot syndrome virus
gene that encodes an envelope protein VP31
Li Li Xixian Xie Feng Yang
Key Laboratory of Marine Biogenetic Resources Third Institute of Oceanography Xiamen PR China
Received 13 April 2005 returned to author for revision 9 May 2005 accepted 3 June 2005
Available online 14 July 2005
Abstract
Based on a combination of SDS-PAGE and mass spectrometry a protein with an apparent molecular mass of 31 kDa (termed as VP31)
was identified from purified shrimp white spot syndrome virus (WSSV) envelope fraction The resulting amino acid (aa) sequence matched
an open reading frame (WSV340) of the WSSV genome This ORF contained 783 nucleotides (nt) encoding 261 aa A fragment of WSV340
was expressed in Escherichia coli as a glutathione S-transferase (GST) fusion protein with a 6His-tag and then specific antibody was raised
Western blot analysis and the immunoelectron microscope method (IEM) confirmed that VP31 was present exclusively in the viral envelope
fraction The neutralization experiment suggested that VP31 might play an important role in WSSV infectivity
D 2005 Elsevier Inc All rights reserved
Keywords White spot syndrome virus VP31 Envelope protein The neutralization experiment
Introduction
White spot syndrome virus (WSSV) is a major shrimp
pathogen that is highly virulent in penaeid shrimp and can
also infect most species of crustacean (Lo et al 1996a
1996b Chen et al 1997 Flegel 1997) The complete
sequence of WSSV genome has now been published for
three different isolates (Chen et al 2002 van Hulten et al
2001a Yang et al 2001 GenBank Accession Nos
AF332093 AF369029 AF440570 for viruses isolated from
China Thailand and Taiwan respectively) Recently the
International Committee on Taxonomy of Viruses approved
a proposal to erect WSSV as the type species of the genus
Whispovirus family Nimaviridae (wwwncbinlmnihgov
ICTVdbIctvindexhtm)
To date some major structural proteins have been
identified (Huang et al 2002b Tsai et al 2004) Among
those proteins VP28 VP26P22 VP281 VP466 VP68
0042-6822$ - see front matter D 2005 Elsevier Inc All rights reserved
doi101016jvirol200506007
Corresponding author
E-mail address mbiotechpublicxmfjcn (F Yang)
VP292 and VP124 were identified as viral envelope
proteins by immunoelectron microscopy (Huang et al
2002a 2002b Zhang et al 2002a 2002b 2004) As far
as functional research is concerned VP28 VP281
VP466 and VP68 were suggested to play an important
role in the systemic WSSV infection in shrimp by using
an in vivo neutralization assay (van Hulten et al 2001b
Wu et al 2005) and VP28 was used for detection of
WSSV in crustaceans (Yoganandhan et al 2004) VP281
with a cell attachment RGD motif was supposed to play
an important role in mediating WSSV infectivity (Huang
et al 2002a) VP26 was reported to be capable of
binding to actin or actin-associated proteins (Xie and
Yang 2005)
In this paper the purified WSSV virions viral envelope
fractions were separated by SDS-PAGE Among these
proteins a 31 kDa protein (termed VP31) was analyzed
using Nano-ESI MSMS and further identified by Western
blot and TEM (transmission electron microscopy) immuno-
gold-labeling In vivo neutralization assay was performed in
this investigation to examine if VP31 can play a role in
WSSV infection
05) 125 ndash 132
L Li et al Virology 340 (2005) 125ndash132126
Results
Identification of WSSV-VP31 by MS
The proteins of the WSSV virion the envelope and
nucleocapsid fraction were separated on 12 SDS-PAGE
gel respectively As shown in Fig 1A the 31 kDa protein
band (arrow indicated) was sequenced using Nano-ESI mass
spectrometry Ten experimentally derived peptide masses
were found to match the predicted peptide masses of the
VP31 protein covering 46 the amino acid sequence
(sequence coverage 120 aa261 aa = 46 date not shown)
Sequence alignment identified that the resulting amino acid
sequence matched WSV340 of the WSSV genome ORF
database WSV340 is located from nucleotide position
196252ndash195510 on the genome and encodes a protein of
261 amino acids with a theoretical molecular mass of
296 kDa
Characterization of WSSV vp31 gene
Rapid amplification of cDNA ends (RACE)
The 5V and 3V regions of the vp31 transcript were
obtained by rapid amplification of the cDNA end (RACE)
The RNA samples used in this study were prepared from
the crayfish at 24 h pi In 5V RACE after two rounds of
PCR amplifications the PCR products formed a single
Fig 1 Coomassie brilliant blue stained 12 SDS-PAGE gel of purified
WSSV virions WSSV envelope and WSSV nucleocapsids and Western
blot analysis (A) SDS-PAGE of purified WSSV virions WSSV envelope
and WSSV nucleocapsids Intact viral particles were obtained with
differential centrifugations The virus envelope was removed from the
virus particles by treatment with 02 Triton X-100 Then the purified
WSSV virions WSSV envelope and WSSV nucleocapsids were assayed
by 12 SDS-PAGE with Coomassie staining Lanes M low-molecular-
weight protein marker 1 purified WSSV virions 2 purified WSSV
envelope 3 purified WSSV nucleocapsids (B) Western blot analysis The
viral envelope and nucleocapsid proteins after separation by SDS-PAGE
were transferred onto a PVDF membrane followed by incubation with anti-
rVP31c serum (diluted at 15000) and alkaline phosphatase-conjugated goat
anti-mouse IgG respectively Lane 1 the viral envelope Lane 2 the viral
nucleocapsids
band in agarose gel at about 280 bp (Fig 2A) Analysis of
the products (Fig 2B) revealed that the transcription
initiation site was located at 94 nt upstream of the
predicted ATG initiation codon a putative TATA box
was found at 37 nt upstream of the transcriptional
initiation sites Amplification of the 3V region of the first-
strand cDNA in 3V RACE yielded PCR products of about
320 bp (Fig 2A) Although there was no putative
polyadenylation signal (AATAAA) a possible polyadeny-
lation signal (AATAAT) (Caron et al 2001 Halder et al
1998) was found at 22 nt upstream of a poly (A) tail by
sequencing 3V RACE fragments (Fig 2B)
Temporal expression of vp31 transcription in WSSV-infected
shrimp
RT-PCR was performed to detect vp31-specific tran-
scripts at different infection stages (0 to 48 h pi) The result
showed that this gene transcript was first detected at 12 h
pi and continued high transcription until 48 h pi (Fig
3A) This result suggested the vp31 gene was a late gene
just as most of WSSV late genes (Huang et al 2002b
Zhang et al 2002a 2002b) For the positive control the
results of RT-PCR with actin-specific primers were shown
in Fig 3B When RNA was treated with RNase and then
subjected to RT-PCR with vp31-specific primers no RT-
PCR amplicon was seen indicating that no virus genomic
DNA was left in the prepared RNA (negative control date
not shown)
Expression of rVP31c in E coli BL21 (DE3)
The C-terminal fragment of vp31 (vp31c) was cloned
into the pET-GST vector and overexpressed as a GST
fusion protein in BL21 (DE3) strain A band corre-
sponding to the GST-VP31c fusion protein was observed
in the induced GST-VP31c as the expected size (date
not shown) No protein band was found at the same
position in uninduced GST-VP31c The expressed fusion
protein was purified by Ni-NTA resin under denaturing
conditions Purified rVP31c was used for antibody
preparation
Western blot analysis
Purified WSSV virions envelope proteins and nucle-
ocapsid proteins were separated by SDS-PAGE respec-
tively (Fig 1A) Western blot results showed that the
rVP31c polyclonal antiserum was reacted specifically with
VP31 in the purified viral envelope proteins while no
binding could be found in the nucleocapsid proteins (Fig
1B) It confirmed that this protein should belong to viral
envelope
Localization of VP31 protein by IEM
Purified WSSV virions were tested to be intact under
transmission electron microscopy (Fig 4A) When both
WSSV virions and nucleocapsids were incubated with
rVP31c antiserum followed with gold-labeled secondary
Fig 2 Nucleotide sequence of WSSV vp31 containing the 5V and 3V terminal region and the deduced VP31 protein sequence (one-letter code) (A) Agarose gel
analysis of the 5V RACE and 3V RACE products Lane M 100 bp ladder DNA marker (Promega) After two rounds of PCR amplifications the PCR products
formed a single 280-bp band in 5V RACE and 320-bp band in 3V RACE (B) The nucleotide sequence of the presumptive WSSV vp31 gene and the deduced
VP31 amino acid sequence The predicted TATA box the possible polyadenylation signal (AATAAT) the start codons two N-glycosylation sites (NLSE
NRTG) and FArgndashGlyndashAsp_ motif were boxed Transcription start point and poly (A) were indicated with black dots Primers used for 5V3VRACE (VP31sp1
VP31sp2 VP31sp3) were underlined
Fig 3 Temporal transcription analysis of WSSV-vp31 at different post-
infection times (hpi) RT-PCR with the gene-specific primers was
performed using the total RNAs extracted from the infected crayfish (A)
RT-PCR with WSSV-vp31-specific primers The transcript was first
detected at 12 h pi and continued to be highly transcribed until 48 h
pi (B) RT-PCR with h-actin-specific primers No RT-PCR amplicon was
observed Lane M 100 bp DNA ladder marker Lane headings showed h
pi (0 2 4 6 8 12 18 24 and 48 h pi)
L Li et al Virology 340 (2005) 125ndash132 127
antibody the gold particles were found to be located on
the surface of the WSSV virions (Fig 4D) while no
particles could be found on the nucleocapsids (Fig 4B) In
the control experiments rVP31c antiserum was replaced
with a pre-immune mouse serum as the primary antibody
no gold particles could be seen for the WSSV virion too
(Fig 4C)
Neutralization assay
The results showed that the crayfish mortalities were
very low for the negative control (09 NaCl) The crayfish
from the positive control (WSSV only) displayed 100
mortality at 8 days post-infection and the crayfish from the
anti-GST IgG control displayed 100 mortality at 9 days
post-infection Whereas after the mixture of WSSV with
anti-rVP31c IgG was injected into crayfish the crayfish
mortality was significantly delayed (Fig 5) indicating that
the infection of WSSV could be delayed or neutralized by
antibodies against rVP31c Deceased shrimps were con-
Fig 4 Localization of VP31 in the WSSV virions by immunoelectron
microscopy (IEM) (Scale bar 90 nm) (A) Electron micrograph of
negatively stained intact WSSV virions (2 PTA) (B) Nucleocapsids of
WSSV incubated with anti-rVP31c antiserum as primary antibody followed
by gold-labeled secondary antibodies (C) Intact WSSV virions incubated
with pre-immune mouse serum as primary antibody followed by gold-
labeled secondary antibodies (D) Intact WSSV virions incubated with anti-
rVP31c antiserum as primary antibody followed by gold-labeled secondary
antibodies Gold particles were indicated by arrows
L Li et al Virology 340 (2005) 125ndash132128
firmed for the presence of WSSV by PCR detection (date
not shown)
Discussion
White spot syndrome virus is a major pathogen of
cultured penaeid shrimp worldwide After the completion of
the WSSV genomic DNA sequencing research has now
Fig 5 Neutralization of WSSV infection in crayfish by specific antibodies The so
crayfish were used After injections the crayfish mortality was monitored daily
mortality on the ordinate
focused on the functional analysis of the gene products
especially on the functions of the viral envelope proteins
Recently a simple and efficient method for purifying the
virions and separating envelope proteins from virions has
been developed in our laboratory (Xie et al 2005) which
facilitated the identification of the viral structural proteins
with low abundance By combining SDS-PAGE with mass
spectrometry vp31 was predicted to encode an envelope
protein This protein was also discovered by another study
in which the VP31 was shown to be a structural protein
without further analysis on protein location and function
(Tsai et al 2004) In this study Western blot showed that
VP31 was in the envelope fraction IEM further revealed
that gold particles were present along the intact WSSV
virions This provided visual evidence that the vp31 gene
encodes one of the envelope proteins of WSSV
Computer analysis showed that the predicted vp31 enco-
ding protein lacks a predominant transmembrane domain
which suggested that VP31 might be a fusion protein that
contained a domain involved in binding to other viral
envelope protein for membrane anchoring The same instance
has been found at the vaccinia virus membrane protein the
VV 14-kDa protein that is located on the surface of the
intracellular mature virus form and is essential for both the
release of extracellular enveloped virus from the cells and
virus spread (Marıa-Isabel et al 1998) Such form had also
been widely reported in some viral membrane fusion pro-
teins pathogen receptors and some cell adhesion molecules
(Boyle and Compton 1998 Jordens et al 1999) These pro-
teins could function to anchor the polypeptides to the
membrane hydrophobic phase by means of locating aqueous
activities or possibly interact with some membrane-spanning
components (Stevens and Arkin 2000) Therefore we pre-
sume that VP31 protein plays an important role in the biology
of the virus acting in virus-to-cell and cell-to-cell fusions
In addition the cell attachment motif RGD (Ruoslahti
and Pierschbacher 1986) occurred in vp31 gene sequence
(Fig 3) suggested that the tripeptide recognition motif might
lutions used for injection were shown at the bottom For each treatment 20
at 25 -C Days post-injection were shown on the abscissa and percentage
L Li et al Virology 340 (2005) 125ndash132 129
have an effect on VP31ndashhost cells interactions In previous
studies RGD structure has been reported widely to mediate
the virusndashhost interaction (Senn and Klaus 1993 Krezel et
al 1994 Verdaguer et al 1995 Yingmanee et al 2001
Mandl et al 1989 Ruoslahti 1996) Among WSSV
proteins six structure proteins including VP31 have been
reported to contain RGD motif (Tsai et al 2004) Moreover
VP31 possesses a threonine at the fourth position of the
RGD motif (RGDT) which was considered to be critical in
interacting with integrin (Plow et al 2000)
Antibodies raised against individual viral envelope
protein have been used successfully in neutralization assay
to identify envelope proteins involved in virus entry during
infection (Martignoni et al 1980 Schofield et al 2000
van Hulten et al 2001a 2001b Volkman and Goldsmith
1985 Wu et al 2005) In this study a specific antiserum
against VP31 protein was prepared to test for neutralization
in vivo The result suggested that VP31 should be involved
in the systemic infection of WSSV in crayfish by partici-
pating in virus attachment and membrane fusion Future
experiments would be performed to demonstrate which part
of VP31 was involved in the neutralization process and what
the role of VP31 in WSSV attachment and entry in the
systemic infection was
In summary we identified a structure protein VP31 as an
envelope protein VP31 might be a key protein in viral
invasion and could be a good candidate for screening
receptors on shrimp cell surface Study on VP31 will be
helpful to realize the mechanism of the virus infection and
to control the infection of WSSV in shrimp industry
Materials and methods
Virus purification and detergent extraction of intact virus
The WSSV isolate in this study originated from Penaeus
japonicus (Xiamen China) and was proliferated in an
alternate host the crayfish (Procambarus clarkii) Prepara-
tion of WSSV virions and nucleocapsids was carried out as
described previously (Xie et al 2005) Brief abundant viral
particles were obtained with only a few steps of conven-
tional differential centrifugations the upper loose pellet was
rinsed out and purified virus in white pellet were
resuspended and kept in TM buffer (50 mM TrisndashHCl 10
mM MgCl2 pH 75)
The virus envelope was removed from the virus
particles by treatment with Triton X-100 Generally the
purified WSSV was mixed with an equal volume of 02
Triton X-100 and incubated for 1 h at room temperature
The nucleocapsids were purified by centrifugation at
20000 g for 20 min at 4 -C The envelope fraction
was collected in the supernatant The nucleocapsid fraction
was subjected to a second round of Triton X-100
extraction to ensure the envelopes were removed com-
pletely The degree of purity of the intact virions isolated
and nucleocapsid fractions were evaluated by negative-
staining transmission electron microscopy
Mass spectrometric analysis of VP31
The intact virions envelope fractions and nucleocapsid
fractions were obtained as above Purified WSSV virions
and the viral envelope fractions were separated using 12
SDS-PAGE (Laemmli 1970) and stained with Coomassie
brilliant blue R-250 A 31 kDa target protein band was
excised from the gel and dehydrated several times with
acetonitrile After vacuum drying the gel band was
incubated with 10 mM dithiothreitol in 100 mM ammonium
bicarbonate (ABC buffer) at 57 -C for 60 min and
subsequently with 55 mM iodoacetamide (Sigma) in 100
mM ABC buffer at room temperature for 60 min Then the
gel was washed with 100 mM ABC buffer and dried In-gel
protein digestion was performed using sequencing grade
modified porcine trypsin (Promega Madison WI) in 50
mM ABC buffer at 37 -C for 15 h Digests were centrifuged
at 6000 g The supernatants were separated and the gel
pieces were extracted further first with 50 acetonitrile 5
formic acid and then with acetonitrile The extracts were
combined with the original digesting supernatants vacuum-
dried and redissolved in 05 trifluoroacetic acid and 50
acetonitrile (Shevchenko et al 1996) Subsequently in-gel-
digested sample was sequenced using Nano-ESI MSMS
mass spectrometry as described previously (Huang et al
2002b) The resulting mass spectrometric data were
searched against the theoretical ORF database of WSSV
Computer analysis of the vp31
The potential sites for posttranslational modifications
were analyzed by the PROSITE database (Hofmann et al
1999) The prediction of transmembrane domain and hydro-
phobic region of WSSV-VP31 was performed with TM-pred
program (Hofman and Stoffel 1993) and DNAMAN
software (Lynnon BioSoft Vaudreuil Canada) respectively
Characterization of WSSV vp31 gene
Rapid amplification of vp31 cDNA ends (5V and 3V RACE)Based on the nucleotide sequence of ORF340 the 5V and
3V ends of the cDNA encoding WSSV-VP31 were obtained
by 5V and 3V RACE using a commercial 5V3V RACE kit
(Roche) according to the manufacturerrsquos recommendations
The RNA samples used in this study were extracted from
WSSV-infected crayfish 24 h pi and then treated with
RNase-free DNase For 5VRACE the first-strand cDNAwas
synthesized using the specific primer sp1 (5V-GGCCGA-TACCAATGTACAAG-3V) and then a poly (A) tail was
added to the cDNA products using terminal transferase in
the presence of dATP The primer sp2 (5V-CTTA-
CAGTGTTCTTCAAC-3V) and an oligo (dT) anchor primer
supplied with the kit were used for PCR For 3VRACE first-
L Li et al Virology 340 (2005) 125ndash132130
strand cDNA was synthesized using an oligo (dT) anchor
primer The primer sp3 (5V-GAGGATGTAGATGCAGTG-3V) and an anchor primer supplied with the kit were used for
PCR The PCR products from 5V and 3V RACE were each
purified on a 2 agarose gel and subcloned into the
pMD18-T vector (TaKaRa) Arbitrarily selected clones were
sequenced and compared with the genomic DNA sequence
of WSSV
Transcriptional analyses of genes
The WSSV inoculum collected from virus-infected
shrimp P japonicus was inoculated into the crayfish
according to the protocol described before (Li et al
2004) Total RNAs at different times (ie 0 2 4 6 8 12
18 24 and 48 h post infection h pi) were extracted from
the hepatopancreas of WSSV-infected crayfish according to
the single-step RNA isolation method (Chomczynski and
Sacchi 1987) The obtained RNAwas further purified using
the SV Total RNA Isolation System (Promega) and
quantified by spectrophotometry at 260 nm The total
RNA (2 Ag) was reverse transcribed with M-MLV RT [H-]
(Promega) using gene-specific reverse primer (GGCCGA-
TACCAATGTACA) The cDNAs were subjected to PCR
using specific forward primer (5V-ATGAAGGATGCA-GAACTG-3V) and reverse primer (GGCCGATACCAATG-
TACA) The PCR cycles were as follows 94 -C for 2 min
30 cycles of 94 -C for 30 s 55 -C for 30 s 72 -C for 1 min
followed by an elongation at 72 -C for 10 min The
crayfish h-actin gene was used as the internal control for
RT-PCR with a gene specific primer set (5V-TCAT-CAGGGTGTGATGGT-3V and 5V-TCTGAGTCATCTTCT-CAC-3V) Total RNA from healthy crayfish was used as the
negative control
Partial expression of GST-VP31 fusion protein in
Escherichia coli
The WSSV-vp31 gene C-terminal fragment (termed as
vp31c) was amplified from WSSV genomic DNA using the
forward primer (5V-CGGGATCC CAAGAAACTGGT-
CAAGT-3V) and the reverse primer (5V-CGGAATTC
CTCCTCCTTAAAAGCAG-3V) that contained recognition
sequences for BamHI and EcoRI restriction enzymes
(underlined) The amplicon was cloned into the pET-GST
vector (Gene Power Lab Ltd Shenzhen office) The
recombinant plasmid was transformed into E coli BL21
(DE3) cells Liquid cultures were grown in a shaking
incubator (200 rpm) at 37 -C until the OD600 reached 07
and these were then induced with 02 mM IPTG for 5 h at
37 -C The cells were harvested by centrifugation at
4000 g for 5 min The recombinant VP31c (termed
rVP31c) was purified by Ni-NTA affinity chromatography
under denatured conditions following methodology in the
QIAexpressionist handbook (Qiagen) The E coli cells
containing pET-GST vector were also induced with IPTG
and total protein extracts were applied to the Ni-NTA
column as described above Final eluates were collected and
used as the negative control
Preparation of antibody
Purified rVP31c protein was used as antigen to immunize
mice by intradermal injection once every 10 days Antigen
(30 Ag) was mixed with an equal volume of Freundrsquos
complete adjuvant (Sigma) for the first injection Subse-
quent three injections were conducted using 30 Ag antigen
mixed with an equal volume of Freundrsquos incomplete
adjuvant (Sigma) Four days after the last injection mice
were exsanguinated and antisera were collected The titers
of the antisera were 115000 as determined by ELISA The
IgG fraction was purified with rProtein AndashSepharose Fast
Flow (Amersham) The optimal dilution of purified IgG
after serial dilutions was 15000 as determined by ELISA
Horseradish-peroxidase (HRP)-conjugated goat anti-mouse
IgG was obtained from Promega For a negative control
antigen was replaced with 1 PBS
Western blot
Virus envelope fraction and nucleocapsid fraction dis-
solved in loading buffer were separated by SDS-PAGE
(Laemmli 1970) respectively These proteins were trans-
ferred onto a PVDF membrane (Amersham Pharmacia) The
membrane was then immersed in blocking buffer (2 BSA
20 mM Tris 150 mM NaCl 01 Tween 20 pH 75) at
room temperature for 30 min followed by incubation with
anti-rVP31c serum (diluted 15000) for 1 h Following this
alkaline-phosphatase-conjugated goat anti-mouse IgG
(Promega) was used as the secondary antibody Detection
was performed using Western Blue Stabilized Substrate for
Alkaline Phosphatase (Promega)
Localization of VP31 protein by immunoelectron
microscopy
WSSV virion and nucleocapsid suspension were mounted
on carbon-coated nickel grids (300 meshes) for 1 h at room
temperature Respectively after washing with PBS the grids
were blocked in 3 BSA for 1 h Then the grids were rinsed
with PBS followed by incubation with anti-rVP31c IgG as
the primary antibody (1200 diluted in 3 BSA) for 2 h
After washing four times with 1 PBS 10-nm-gold-labeled
anti-mouse goat IgG as the secondary antibody (Sigma) was
added to the grids and incubated for 1 h at room temperature
Grids were washed further four times with 1 PBS and
stained with 2 phosphotungstic acid (PTA pH 70) for 25
min Specimens were examined under a TEM For control
experiment anti-rVP31c IgG was replaced with a pre-
immune mouse serum as the primary antibodies indicated
above
Neutralization assay
Prior to injection into crayfish WSSV virions were
incubated with anti-rVP31c IgG or the anti-GST IgG as a
L Li et al Virology 340 (2005) 125ndash132 131
control (final concentration 4 Ag IgGml 107 virionsml) for
1 h at room temperature Then the 100 Al mixture of
antibody and WSSV was intramuscularly injected into
crayfish using a syringe with 29-gauge needle At the same
time a negative (09 NaCl) and a positive control (virus
only 107 virionsml) were included in the injections For
each treatment 20 crayfish were used After injections the
crayfish mortality was monitored daily at 25 -C Deceasedshrimps were examined for the presence of WSSV by PCR
detection
Acknowledgments
We thank Ping Chen for her technical assistance in the
transmission electron microscopy This work was sup-
ported by the lsquolsquo863rsquorsquo Program of China (2003AA626020)
National Natural Science Foundation of China (30330470
40276038) and Fujian Science Fund (2003F001)
References
Boyle KA Compton T 1998 Receptor-binding properties of a
soluble form of human cytomegalovirus glycoprotein B J Virol 72
1826ndash1833
Caron H van Schaik B van der Mee M Baas F Riggins G van
Sluis P Hermus M-C van Asperen R Boon K Voute PA
Heisterkamp S van Kamper A Versteeg R 2001 The human
transcriptome map clustering of highly expressed genes in chromoso-
mal domainsmdashSupplementary material Science 291 1289ndash1292
Chen XF Chen C Wu DH Huai H Chi XC 1997 A new
baculovirus of cultured shrimp Sci China Ser C 40 630ndash635
Chen LL Leu JH 1 Huang CJ Chou CM Chen SM Wang CH
Lo CF Kou GH 2002 Identification of a nucleocapsid protein
(VP35) gene of shrimp white spot syndrome virus and characterization
of the motif important for targeting VP35 to the nuclei of transfected
insect cells Virology 293 44ndash53
Chomczynski P Sacchi N 1987 Single-step method of RNA isolation
by acid guanidinium thiocyanatendashphenolndashchloroform extraction Anal
Biochem 262 156ndash159
Flegel TW 1997 Special topic review major viral diseases of the black
tiger prawn (Penaeus monodon) in Thailand World J Microbiol
Biotechnol 13 433ndash442
Halder SK Takemori H Hatano O Nonaka Y Wada A Okamoto
M 1998 Cloning of a membrane-spanning protein with epidermal
growth factor-like repeat motifs from adrenal glomerulosa cells
Endocrinology 139 3316ndash3328
Hofman K Stoffel W 1993 TMbasemdashA database of membrane
spanning protein segments Biol Chem Hoppe-Seyler 374 166
Hofmann K Bucher P Falquet L Bairoch A 1999 The PROSITE
database its status in 1999 Nucleic Acids Res 27 215ndash219
Huang CH Zhang XB Lin QS Xu X Hew CL 2002a Character-
ization of a novel envelope protein (VP281) of shrimp white spot
syndrome virus by mass spectrometry J Gen Virol 83 2385ndash2392
Huang CH Zhang XB Lin QS Xu X Hu ZH Hew CL 2002b
Proteomic analysis of shrimp white spot syndrome viral proteins and
characterization of a novel envelope protein VP466 Mol Cell
Proteomics 1 223ndash231
Jordens R Thompson A Amons R Koning F 1999 Human dendritic
cells shed a functional soluble form of the mannose receptor Int
Immunol 11 1775ndash1780
Krezel AM Wagner G Seymour-Ulmer J Lazarus RA 1994
Structure of the RGD protein decorsin conserved motif and distinct
function in leech proteins that affect blood clotting Science 264
1944ndash1947
Laemmli UK 1970 Cleavage of structural proteins during the assembly
of the head of bacteriophage T4 Nature 227 680ndash685
Li Q Pan D Zhang J Yang F 2004 Identification of the thymidylate
synthase within the genome of white spot syndrome virus J Gen Virol
85 2035ndash2044
Lo CF Ho CH Peng SE Chen CH Hsu HC Chiu YL Chang
CF Liu KF Su MS Wang CH Kou GH 1996a White spot
syndrome baculovirus (WSBV) detected in cultured and captured
shrimp crabs and other arthropods Dis Aquat Org 27 215ndash226
Lo CF Leu JH Ho CH Chen CH Peng SE Chen YT Chou
CM Yeh PY Huang CJ Chou HY Wang CH Kou GH
1996b Detection of baculovirus associated with white spot syndrome
(WSBV) in penaeid shrimps using polymerase chain reaction Dis
Aquat Org 25 133ndash141
Mandl CW Guirakhoo F Holzmann H Heinz FX Kunz C 1989
Antigenic structure of the flavivirus envelope E protein at the molecular
level using tick-borne encephalitis virus as a model J Virol 63
564ndash571
Marıa-Isabel V German R David C Luis S Mariano E 1998
The vaccinia virus 14-kilodalton (A27L) fusion protein forms a triple
coiled-coil structure and interacts with the 21-kilodalton (A17L)
virus membrane protein through a C-terminal a-helix J Virol 72
10126ndash10137
Martignoni ME Iwai PJ Rohrmann GF 1980 Serum neutralization
of nucleopolyhedrosis viruses (baculovirus subgroup A) pathogenic for
Orgyia pseudotsugata J Invert Pathol 36 12ndash20
Plow EF Haas TA Zhang L Loftus J Smith JW 2000 Ligand
binding to integrins J Biol Chem 275 21785ndash21788
Ruoslahti E 1996 RGD and other recognition sequences for integrins
Annu Rev Cell Biol 12 697ndash715
Ruoslahti E Pierschbacher MD 1986 ArgndashGlyndashAsp a versatile cell
recognition signal Cell 44 517ndash518
Schofield DJ Glamann J Emerson U Purchell RH 2000 Identi-
fication by phage display and characterization of two neutralizing
chimpanzee monoclonal antibodies to the hepatitis E virus capsid
protein J Virol 74 5548ndash5555
Senn H Klaus W 1993 The nuclear magnetic resonance solution
structure of flavoridin an antagonist of the platelet GPIIbndash IIIa receptor
J Mol Biol 232 907ndash925
Shevchenko A Wilm M Vorm O Mann M 1996 Mass spectrometric
sequencing of protein silver-stained polyacrylamide gels Anal Chem
68 850ndash858
Stevens TJ Arkin IT 2000 Turning an opinion inside-out Rees and
Eisenbergrsquos commentary (Proteins 2000 38121ndash122) on FAre mem-
brane proteins lsquolsquoinside-outrsquorsquo proteins_ (Proteins 1999 36 135ndash143)
Proteins 40 463ndash464
Tsai JM Wang HC Leu JH Hsiao HH Wang AH Kou GH
Lo CF 2004 Genomic and proteomic analysis of thirty-nine
structural proteins of shrimp white spot syndrome virus J Virol 78
11360ndash11370
van Hulten MC Witteveldt J Peters S Kloosterboer N Tarchini
R Fiers M Sandbrink H Lankhorst RK Vlak JM 2001a
The white spot syndrome virus DNA genome sequence Virology
286 7ndash22
van Hulten MC Witteveldt J Snippe M Vlak JM 2001b White spot
syndrome virus envelope protein VP28 is involved in the systemic
infection of shrimp Virology 285 228ndash233
Verdaguer N Mateu MG Andreu D Giralt E Domingo E Fita
I 1995 Structure of the major antigenic loop of foot-and-mouth
disease virus complexed with a neutralizing antibody direct involve-
ment of the ArgndashGlyndashAsp motif in the interaction EMBO J 14
1690ndash1696
Volkman LE Goldsmith PA 1985 Mechanism of neutralization of
L Li et al Virology 340 (2005) 125ndash132132
budded Autographa californica nuclear polyhedrosis virus by a
monoclonal antibody inhibition of entry by adsorptive endocytosis
Virology 143 185ndash195
Wu WL Wang L Zhang XB 2005 Identification of white spot
syndrome virus (WSSV) envelope proteins involved in shrimp
infection Virology 332 578ndash583
Xie XX Yang F 2005 Interaction of white spot syndrome virus VP26
protein with actin Virology 336 93ndash99
Xie XX Li HY Xu LM Yang F 2005 A simple and efficient
method for purification of intact White Spot Syndrome Virus (WSSV)
viral particles Virus Res 108 63ndash67
Yang F He J Lin X Li Q Pan D Zhang X Xu X 2001 Complete
genome sequence of the shrimp white spot bacilliform virus J Virol 75
11811ndash11820
Yingmanee B Pamela JH Farideh G Glyn S 2001 Argininendash
glycinendashaspartic acid motif is critical for human parechovirus 1 entry
J Virol 75 10000ndash10004
Yoganandhan K Syed MS Narayanan RB Sahul HAS 2004
Production of polyclonal antiserum against recombinant VP28 protein
and its application for the detection of white spot syndrome virus in
crustaceans J Fish Dis 27 517ndash522
Zhang X Huang CH Xu X Hew CL 2002a Transcription and
identification of an envelope protein gene (p22) from shrimp white spot
syndrome virus J Gen Virol 83 471ndash477
Zhang X Huang CH Xu X Hew CL 2002b Identification and
localization of a prawn white spot syndrome virus gene that encodes an
envelope protein J Gen Virol 83 1069ndash1074
Zhang X Huang C Tang X Zhuang Y Choy LH 2004 Identification
of structural proteins from shrimp white spot syndrome virus (WSSV)
by 2-DE-MS Proteins Struct Funct Bioinform 55 229ndash235

L Li et al Virology 340 (2005) 125ndash132126
Results
Identification of WSSV-VP31 by MS
The proteins of the WSSV virion the envelope and
nucleocapsid fraction were separated on 12 SDS-PAGE
gel respectively As shown in Fig 1A the 31 kDa protein
band (arrow indicated) was sequenced using Nano-ESI mass
spectrometry Ten experimentally derived peptide masses
were found to match the predicted peptide masses of the
VP31 protein covering 46 the amino acid sequence
(sequence coverage 120 aa261 aa = 46 date not shown)
Sequence alignment identified that the resulting amino acid
sequence matched WSV340 of the WSSV genome ORF
database WSV340 is located from nucleotide position
196252ndash195510 on the genome and encodes a protein of
261 amino acids with a theoretical molecular mass of
296 kDa
Characterization of WSSV vp31 gene
Rapid amplification of cDNA ends (RACE)
The 5V and 3V regions of the vp31 transcript were
obtained by rapid amplification of the cDNA end (RACE)
The RNA samples used in this study were prepared from
the crayfish at 24 h pi In 5V RACE after two rounds of
PCR amplifications the PCR products formed a single
Fig 1 Coomassie brilliant blue stained 12 SDS-PAGE gel of purified
WSSV virions WSSV envelope and WSSV nucleocapsids and Western
blot analysis (A) SDS-PAGE of purified WSSV virions WSSV envelope
and WSSV nucleocapsids Intact viral particles were obtained with
differential centrifugations The virus envelope was removed from the
virus particles by treatment with 02 Triton X-100 Then the purified
WSSV virions WSSV envelope and WSSV nucleocapsids were assayed
by 12 SDS-PAGE with Coomassie staining Lanes M low-molecular-
weight protein marker 1 purified WSSV virions 2 purified WSSV
envelope 3 purified WSSV nucleocapsids (B) Western blot analysis The
viral envelope and nucleocapsid proteins after separation by SDS-PAGE
were transferred onto a PVDF membrane followed by incubation with anti-
rVP31c serum (diluted at 15000) and alkaline phosphatase-conjugated goat
anti-mouse IgG respectively Lane 1 the viral envelope Lane 2 the viral
nucleocapsids
band in agarose gel at about 280 bp (Fig 2A) Analysis of
the products (Fig 2B) revealed that the transcription
initiation site was located at 94 nt upstream of the
predicted ATG initiation codon a putative TATA box
was found at 37 nt upstream of the transcriptional
initiation sites Amplification of the 3V region of the first-
strand cDNA in 3V RACE yielded PCR products of about
320 bp (Fig 2A) Although there was no putative
polyadenylation signal (AATAAA) a possible polyadeny-
lation signal (AATAAT) (Caron et al 2001 Halder et al
1998) was found at 22 nt upstream of a poly (A) tail by
sequencing 3V RACE fragments (Fig 2B)
Temporal expression of vp31 transcription in WSSV-infected
shrimp
RT-PCR was performed to detect vp31-specific tran-
scripts at different infection stages (0 to 48 h pi) The result
showed that this gene transcript was first detected at 12 h
pi and continued high transcription until 48 h pi (Fig
3A) This result suggested the vp31 gene was a late gene
just as most of WSSV late genes (Huang et al 2002b
Zhang et al 2002a 2002b) For the positive control the
results of RT-PCR with actin-specific primers were shown
in Fig 3B When RNA was treated with RNase and then
subjected to RT-PCR with vp31-specific primers no RT-
PCR amplicon was seen indicating that no virus genomic
DNA was left in the prepared RNA (negative control date
not shown)
Expression of rVP31c in E coli BL21 (DE3)
The C-terminal fragment of vp31 (vp31c) was cloned
into the pET-GST vector and overexpressed as a GST
fusion protein in BL21 (DE3) strain A band corre-
sponding to the GST-VP31c fusion protein was observed
in the induced GST-VP31c as the expected size (date
not shown) No protein band was found at the same
position in uninduced GST-VP31c The expressed fusion
protein was purified by Ni-NTA resin under denaturing
conditions Purified rVP31c was used for antibody
preparation
Western blot analysis
Purified WSSV virions envelope proteins and nucle-
ocapsid proteins were separated by SDS-PAGE respec-
tively (Fig 1A) Western blot results showed that the
rVP31c polyclonal antiserum was reacted specifically with
VP31 in the purified viral envelope proteins while no
binding could be found in the nucleocapsid proteins (Fig
1B) It confirmed that this protein should belong to viral
envelope
Localization of VP31 protein by IEM
Purified WSSV virions were tested to be intact under
transmission electron microscopy (Fig 4A) When both
WSSV virions and nucleocapsids were incubated with
rVP31c antiserum followed with gold-labeled secondary
Fig 2 Nucleotide sequence of WSSV vp31 containing the 5V and 3V terminal region and the deduced VP31 protein sequence (one-letter code) (A) Agarose gel
analysis of the 5V RACE and 3V RACE products Lane M 100 bp ladder DNA marker (Promega) After two rounds of PCR amplifications the PCR products
formed a single 280-bp band in 5V RACE and 320-bp band in 3V RACE (B) The nucleotide sequence of the presumptive WSSV vp31 gene and the deduced
VP31 amino acid sequence The predicted TATA box the possible polyadenylation signal (AATAAT) the start codons two N-glycosylation sites (NLSE
NRTG) and FArgndashGlyndashAsp_ motif were boxed Transcription start point and poly (A) were indicated with black dots Primers used for 5V3VRACE (VP31sp1
VP31sp2 VP31sp3) were underlined
Fig 3 Temporal transcription analysis of WSSV-vp31 at different post-
infection times (hpi) RT-PCR with the gene-specific primers was
performed using the total RNAs extracted from the infected crayfish (A)
RT-PCR with WSSV-vp31-specific primers The transcript was first
detected at 12 h pi and continued to be highly transcribed until 48 h
pi (B) RT-PCR with h-actin-specific primers No RT-PCR amplicon was
observed Lane M 100 bp DNA ladder marker Lane headings showed h
pi (0 2 4 6 8 12 18 24 and 48 h pi)
L Li et al Virology 340 (2005) 125ndash132 127
antibody the gold particles were found to be located on
the surface of the WSSV virions (Fig 4D) while no
particles could be found on the nucleocapsids (Fig 4B) In
the control experiments rVP31c antiserum was replaced
with a pre-immune mouse serum as the primary antibody
no gold particles could be seen for the WSSV virion too
(Fig 4C)
Neutralization assay
The results showed that the crayfish mortalities were
very low for the negative control (09 NaCl) The crayfish
from the positive control (WSSV only) displayed 100
mortality at 8 days post-infection and the crayfish from the
anti-GST IgG control displayed 100 mortality at 9 days
post-infection Whereas after the mixture of WSSV with
anti-rVP31c IgG was injected into crayfish the crayfish
mortality was significantly delayed (Fig 5) indicating that
the infection of WSSV could be delayed or neutralized by
antibodies against rVP31c Deceased shrimps were con-
Fig 4 Localization of VP31 in the WSSV virions by immunoelectron
microscopy (IEM) (Scale bar 90 nm) (A) Electron micrograph of
negatively stained intact WSSV virions (2 PTA) (B) Nucleocapsids of
WSSV incubated with anti-rVP31c antiserum as primary antibody followed
by gold-labeled secondary antibodies (C) Intact WSSV virions incubated
with pre-immune mouse serum as primary antibody followed by gold-
labeled secondary antibodies (D) Intact WSSV virions incubated with anti-
rVP31c antiserum as primary antibody followed by gold-labeled secondary
antibodies Gold particles were indicated by arrows
L Li et al Virology 340 (2005) 125ndash132128
firmed for the presence of WSSV by PCR detection (date
not shown)
Discussion
White spot syndrome virus is a major pathogen of
cultured penaeid shrimp worldwide After the completion of
the WSSV genomic DNA sequencing research has now
Fig 5 Neutralization of WSSV infection in crayfish by specific antibodies The so
crayfish were used After injections the crayfish mortality was monitored daily
mortality on the ordinate
focused on the functional analysis of the gene products
especially on the functions of the viral envelope proteins
Recently a simple and efficient method for purifying the
virions and separating envelope proteins from virions has
been developed in our laboratory (Xie et al 2005) which
facilitated the identification of the viral structural proteins
with low abundance By combining SDS-PAGE with mass
spectrometry vp31 was predicted to encode an envelope
protein This protein was also discovered by another study
in which the VP31 was shown to be a structural protein
without further analysis on protein location and function
(Tsai et al 2004) In this study Western blot showed that
VP31 was in the envelope fraction IEM further revealed
that gold particles were present along the intact WSSV
virions This provided visual evidence that the vp31 gene
encodes one of the envelope proteins of WSSV
Computer analysis showed that the predicted vp31 enco-
ding protein lacks a predominant transmembrane domain
which suggested that VP31 might be a fusion protein that
contained a domain involved in binding to other viral
envelope protein for membrane anchoring The same instance
has been found at the vaccinia virus membrane protein the
VV 14-kDa protein that is located on the surface of the
intracellular mature virus form and is essential for both the
release of extracellular enveloped virus from the cells and
virus spread (Marıa-Isabel et al 1998) Such form had also
been widely reported in some viral membrane fusion pro-
teins pathogen receptors and some cell adhesion molecules
(Boyle and Compton 1998 Jordens et al 1999) These pro-
teins could function to anchor the polypeptides to the
membrane hydrophobic phase by means of locating aqueous
activities or possibly interact with some membrane-spanning
components (Stevens and Arkin 2000) Therefore we pre-
sume that VP31 protein plays an important role in the biology
of the virus acting in virus-to-cell and cell-to-cell fusions
In addition the cell attachment motif RGD (Ruoslahti
and Pierschbacher 1986) occurred in vp31 gene sequence
(Fig 3) suggested that the tripeptide recognition motif might
lutions used for injection were shown at the bottom For each treatment 20
at 25 -C Days post-injection were shown on the abscissa and percentage
L Li et al Virology 340 (2005) 125ndash132 129
have an effect on VP31ndashhost cells interactions In previous
studies RGD structure has been reported widely to mediate
the virusndashhost interaction (Senn and Klaus 1993 Krezel et
al 1994 Verdaguer et al 1995 Yingmanee et al 2001
Mandl et al 1989 Ruoslahti 1996) Among WSSV
proteins six structure proteins including VP31 have been
reported to contain RGD motif (Tsai et al 2004) Moreover
VP31 possesses a threonine at the fourth position of the
RGD motif (RGDT) which was considered to be critical in
interacting with integrin (Plow et al 2000)
Antibodies raised against individual viral envelope
protein have been used successfully in neutralization assay
to identify envelope proteins involved in virus entry during
infection (Martignoni et al 1980 Schofield et al 2000
van Hulten et al 2001a 2001b Volkman and Goldsmith
1985 Wu et al 2005) In this study a specific antiserum
against VP31 protein was prepared to test for neutralization
in vivo The result suggested that VP31 should be involved
in the systemic infection of WSSV in crayfish by partici-
pating in virus attachment and membrane fusion Future
experiments would be performed to demonstrate which part
of VP31 was involved in the neutralization process and what
the role of VP31 in WSSV attachment and entry in the
systemic infection was
In summary we identified a structure protein VP31 as an
envelope protein VP31 might be a key protein in viral
invasion and could be a good candidate for screening
receptors on shrimp cell surface Study on VP31 will be
helpful to realize the mechanism of the virus infection and
to control the infection of WSSV in shrimp industry
Materials and methods
Virus purification and detergent extraction of intact virus
The WSSV isolate in this study originated from Penaeus
japonicus (Xiamen China) and was proliferated in an
alternate host the crayfish (Procambarus clarkii) Prepara-
tion of WSSV virions and nucleocapsids was carried out as
described previously (Xie et al 2005) Brief abundant viral
particles were obtained with only a few steps of conven-
tional differential centrifugations the upper loose pellet was
rinsed out and purified virus in white pellet were
resuspended and kept in TM buffer (50 mM TrisndashHCl 10
mM MgCl2 pH 75)
The virus envelope was removed from the virus
particles by treatment with Triton X-100 Generally the
purified WSSV was mixed with an equal volume of 02
Triton X-100 and incubated for 1 h at room temperature
The nucleocapsids were purified by centrifugation at
20000 g for 20 min at 4 -C The envelope fraction
was collected in the supernatant The nucleocapsid fraction
was subjected to a second round of Triton X-100
extraction to ensure the envelopes were removed com-
pletely The degree of purity of the intact virions isolated
and nucleocapsid fractions were evaluated by negative-
staining transmission electron microscopy
Mass spectrometric analysis of VP31
The intact virions envelope fractions and nucleocapsid
fractions were obtained as above Purified WSSV virions
and the viral envelope fractions were separated using 12
SDS-PAGE (Laemmli 1970) and stained with Coomassie
brilliant blue R-250 A 31 kDa target protein band was
excised from the gel and dehydrated several times with
acetonitrile After vacuum drying the gel band was
incubated with 10 mM dithiothreitol in 100 mM ammonium
bicarbonate (ABC buffer) at 57 -C for 60 min and
subsequently with 55 mM iodoacetamide (Sigma) in 100
mM ABC buffer at room temperature for 60 min Then the
gel was washed with 100 mM ABC buffer and dried In-gel
protein digestion was performed using sequencing grade
modified porcine trypsin (Promega Madison WI) in 50
mM ABC buffer at 37 -C for 15 h Digests were centrifuged
at 6000 g The supernatants were separated and the gel
pieces were extracted further first with 50 acetonitrile 5
formic acid and then with acetonitrile The extracts were
combined with the original digesting supernatants vacuum-
dried and redissolved in 05 trifluoroacetic acid and 50
acetonitrile (Shevchenko et al 1996) Subsequently in-gel-
digested sample was sequenced using Nano-ESI MSMS
mass spectrometry as described previously (Huang et al
2002b) The resulting mass spectrometric data were
searched against the theoretical ORF database of WSSV
Computer analysis of the vp31
The potential sites for posttranslational modifications
were analyzed by the PROSITE database (Hofmann et al
1999) The prediction of transmembrane domain and hydro-
phobic region of WSSV-VP31 was performed with TM-pred
program (Hofman and Stoffel 1993) and DNAMAN
software (Lynnon BioSoft Vaudreuil Canada) respectively
Characterization of WSSV vp31 gene
Rapid amplification of vp31 cDNA ends (5V and 3V RACE)Based on the nucleotide sequence of ORF340 the 5V and
3V ends of the cDNA encoding WSSV-VP31 were obtained
by 5V and 3V RACE using a commercial 5V3V RACE kit
(Roche) according to the manufacturerrsquos recommendations
The RNA samples used in this study were extracted from
WSSV-infected crayfish 24 h pi and then treated with
RNase-free DNase For 5VRACE the first-strand cDNAwas
synthesized using the specific primer sp1 (5V-GGCCGA-TACCAATGTACAAG-3V) and then a poly (A) tail was
added to the cDNA products using terminal transferase in
the presence of dATP The primer sp2 (5V-CTTA-
CAGTGTTCTTCAAC-3V) and an oligo (dT) anchor primer
supplied with the kit were used for PCR For 3VRACE first-
L Li et al Virology 340 (2005) 125ndash132130
strand cDNA was synthesized using an oligo (dT) anchor
primer The primer sp3 (5V-GAGGATGTAGATGCAGTG-3V) and an anchor primer supplied with the kit were used for
PCR The PCR products from 5V and 3V RACE were each
purified on a 2 agarose gel and subcloned into the
pMD18-T vector (TaKaRa) Arbitrarily selected clones were
sequenced and compared with the genomic DNA sequence
of WSSV
Transcriptional analyses of genes
The WSSV inoculum collected from virus-infected
shrimp P japonicus was inoculated into the crayfish
according to the protocol described before (Li et al
2004) Total RNAs at different times (ie 0 2 4 6 8 12
18 24 and 48 h post infection h pi) were extracted from
the hepatopancreas of WSSV-infected crayfish according to
the single-step RNA isolation method (Chomczynski and
Sacchi 1987) The obtained RNAwas further purified using
the SV Total RNA Isolation System (Promega) and
quantified by spectrophotometry at 260 nm The total
RNA (2 Ag) was reverse transcribed with M-MLV RT [H-]
(Promega) using gene-specific reverse primer (GGCCGA-
TACCAATGTACA) The cDNAs were subjected to PCR
using specific forward primer (5V-ATGAAGGATGCA-GAACTG-3V) and reverse primer (GGCCGATACCAATG-
TACA) The PCR cycles were as follows 94 -C for 2 min
30 cycles of 94 -C for 30 s 55 -C for 30 s 72 -C for 1 min
followed by an elongation at 72 -C for 10 min The
crayfish h-actin gene was used as the internal control for
RT-PCR with a gene specific primer set (5V-TCAT-CAGGGTGTGATGGT-3V and 5V-TCTGAGTCATCTTCT-CAC-3V) Total RNA from healthy crayfish was used as the
negative control
Partial expression of GST-VP31 fusion protein in
Escherichia coli
The WSSV-vp31 gene C-terminal fragment (termed as
vp31c) was amplified from WSSV genomic DNA using the
forward primer (5V-CGGGATCC CAAGAAACTGGT-
CAAGT-3V) and the reverse primer (5V-CGGAATTC
CTCCTCCTTAAAAGCAG-3V) that contained recognition
sequences for BamHI and EcoRI restriction enzymes
(underlined) The amplicon was cloned into the pET-GST
vector (Gene Power Lab Ltd Shenzhen office) The
recombinant plasmid was transformed into E coli BL21
(DE3) cells Liquid cultures were grown in a shaking
incubator (200 rpm) at 37 -C until the OD600 reached 07
and these were then induced with 02 mM IPTG for 5 h at
37 -C The cells were harvested by centrifugation at
4000 g for 5 min The recombinant VP31c (termed
rVP31c) was purified by Ni-NTA affinity chromatography
under denatured conditions following methodology in the
QIAexpressionist handbook (Qiagen) The E coli cells
containing pET-GST vector were also induced with IPTG
and total protein extracts were applied to the Ni-NTA
column as described above Final eluates were collected and
used as the negative control
Preparation of antibody
Purified rVP31c protein was used as antigen to immunize
mice by intradermal injection once every 10 days Antigen
(30 Ag) was mixed with an equal volume of Freundrsquos
complete adjuvant (Sigma) for the first injection Subse-
quent three injections were conducted using 30 Ag antigen
mixed with an equal volume of Freundrsquos incomplete
adjuvant (Sigma) Four days after the last injection mice
were exsanguinated and antisera were collected The titers
of the antisera were 115000 as determined by ELISA The
IgG fraction was purified with rProtein AndashSepharose Fast
Flow (Amersham) The optimal dilution of purified IgG
after serial dilutions was 15000 as determined by ELISA
Horseradish-peroxidase (HRP)-conjugated goat anti-mouse
IgG was obtained from Promega For a negative control
antigen was replaced with 1 PBS
Western blot
Virus envelope fraction and nucleocapsid fraction dis-
solved in loading buffer were separated by SDS-PAGE
(Laemmli 1970) respectively These proteins were trans-
ferred onto a PVDF membrane (Amersham Pharmacia) The
membrane was then immersed in blocking buffer (2 BSA
20 mM Tris 150 mM NaCl 01 Tween 20 pH 75) at
room temperature for 30 min followed by incubation with
anti-rVP31c serum (diluted 15000) for 1 h Following this
alkaline-phosphatase-conjugated goat anti-mouse IgG
(Promega) was used as the secondary antibody Detection
was performed using Western Blue Stabilized Substrate for
Alkaline Phosphatase (Promega)
Localization of VP31 protein by immunoelectron
microscopy
WSSV virion and nucleocapsid suspension were mounted
on carbon-coated nickel grids (300 meshes) for 1 h at room
temperature Respectively after washing with PBS the grids
were blocked in 3 BSA for 1 h Then the grids were rinsed
with PBS followed by incubation with anti-rVP31c IgG as
the primary antibody (1200 diluted in 3 BSA) for 2 h
After washing four times with 1 PBS 10-nm-gold-labeled
anti-mouse goat IgG as the secondary antibody (Sigma) was
added to the grids and incubated for 1 h at room temperature
Grids were washed further four times with 1 PBS and
stained with 2 phosphotungstic acid (PTA pH 70) for 25
min Specimens were examined under a TEM For control
experiment anti-rVP31c IgG was replaced with a pre-
immune mouse serum as the primary antibodies indicated
above
Neutralization assay
Prior to injection into crayfish WSSV virions were
incubated with anti-rVP31c IgG or the anti-GST IgG as a
L Li et al Virology 340 (2005) 125ndash132 131
control (final concentration 4 Ag IgGml 107 virionsml) for
1 h at room temperature Then the 100 Al mixture of
antibody and WSSV was intramuscularly injected into
crayfish using a syringe with 29-gauge needle At the same
time a negative (09 NaCl) and a positive control (virus
only 107 virionsml) were included in the injections For
each treatment 20 crayfish were used After injections the
crayfish mortality was monitored daily at 25 -C Deceasedshrimps were examined for the presence of WSSV by PCR
detection
Acknowledgments
We thank Ping Chen for her technical assistance in the
transmission electron microscopy This work was sup-
ported by the lsquolsquo863rsquorsquo Program of China (2003AA626020)
National Natural Science Foundation of China (30330470
40276038) and Fujian Science Fund (2003F001)
References
Boyle KA Compton T 1998 Receptor-binding properties of a
soluble form of human cytomegalovirus glycoprotein B J Virol 72
1826ndash1833
Caron H van Schaik B van der Mee M Baas F Riggins G van
Sluis P Hermus M-C van Asperen R Boon K Voute PA
Heisterkamp S van Kamper A Versteeg R 2001 The human
transcriptome map clustering of highly expressed genes in chromoso-
mal domainsmdashSupplementary material Science 291 1289ndash1292
Chen XF Chen C Wu DH Huai H Chi XC 1997 A new
baculovirus of cultured shrimp Sci China Ser C 40 630ndash635
Chen LL Leu JH 1 Huang CJ Chou CM Chen SM Wang CH
Lo CF Kou GH 2002 Identification of a nucleocapsid protein
(VP35) gene of shrimp white spot syndrome virus and characterization
of the motif important for targeting VP35 to the nuclei of transfected
insect cells Virology 293 44ndash53
Chomczynski P Sacchi N 1987 Single-step method of RNA isolation
by acid guanidinium thiocyanatendashphenolndashchloroform extraction Anal
Biochem 262 156ndash159
Flegel TW 1997 Special topic review major viral diseases of the black
tiger prawn (Penaeus monodon) in Thailand World J Microbiol
Biotechnol 13 433ndash442
Halder SK Takemori H Hatano O Nonaka Y Wada A Okamoto
M 1998 Cloning of a membrane-spanning protein with epidermal
growth factor-like repeat motifs from adrenal glomerulosa cells
Endocrinology 139 3316ndash3328
Hofman K Stoffel W 1993 TMbasemdashA database of membrane
spanning protein segments Biol Chem Hoppe-Seyler 374 166
Hofmann K Bucher P Falquet L Bairoch A 1999 The PROSITE
database its status in 1999 Nucleic Acids Res 27 215ndash219
Huang CH Zhang XB Lin QS Xu X Hew CL 2002a Character-
ization of a novel envelope protein (VP281) of shrimp white spot
syndrome virus by mass spectrometry J Gen Virol 83 2385ndash2392
Huang CH Zhang XB Lin QS Xu X Hu ZH Hew CL 2002b
Proteomic analysis of shrimp white spot syndrome viral proteins and
characterization of a novel envelope protein VP466 Mol Cell
Proteomics 1 223ndash231
Jordens R Thompson A Amons R Koning F 1999 Human dendritic
cells shed a functional soluble form of the mannose receptor Int
Immunol 11 1775ndash1780
Krezel AM Wagner G Seymour-Ulmer J Lazarus RA 1994
Structure of the RGD protein decorsin conserved motif and distinct
function in leech proteins that affect blood clotting Science 264
1944ndash1947
Laemmli UK 1970 Cleavage of structural proteins during the assembly
of the head of bacteriophage T4 Nature 227 680ndash685
Li Q Pan D Zhang J Yang F 2004 Identification of the thymidylate
synthase within the genome of white spot syndrome virus J Gen Virol
85 2035ndash2044
Lo CF Ho CH Peng SE Chen CH Hsu HC Chiu YL Chang
CF Liu KF Su MS Wang CH Kou GH 1996a White spot
syndrome baculovirus (WSBV) detected in cultured and captured
shrimp crabs and other arthropods Dis Aquat Org 27 215ndash226
Lo CF Leu JH Ho CH Chen CH Peng SE Chen YT Chou
CM Yeh PY Huang CJ Chou HY Wang CH Kou GH
1996b Detection of baculovirus associated with white spot syndrome
(WSBV) in penaeid shrimps using polymerase chain reaction Dis
Aquat Org 25 133ndash141
Mandl CW Guirakhoo F Holzmann H Heinz FX Kunz C 1989
Antigenic structure of the flavivirus envelope E protein at the molecular
level using tick-borne encephalitis virus as a model J Virol 63
564ndash571
Marıa-Isabel V German R David C Luis S Mariano E 1998
The vaccinia virus 14-kilodalton (A27L) fusion protein forms a triple
coiled-coil structure and interacts with the 21-kilodalton (A17L)
virus membrane protein through a C-terminal a-helix J Virol 72
10126ndash10137
Martignoni ME Iwai PJ Rohrmann GF 1980 Serum neutralization
of nucleopolyhedrosis viruses (baculovirus subgroup A) pathogenic for
Orgyia pseudotsugata J Invert Pathol 36 12ndash20
Plow EF Haas TA Zhang L Loftus J Smith JW 2000 Ligand
binding to integrins J Biol Chem 275 21785ndash21788
Ruoslahti E 1996 RGD and other recognition sequences for integrins
Annu Rev Cell Biol 12 697ndash715
Ruoslahti E Pierschbacher MD 1986 ArgndashGlyndashAsp a versatile cell
recognition signal Cell 44 517ndash518
Schofield DJ Glamann J Emerson U Purchell RH 2000 Identi-
fication by phage display and characterization of two neutralizing
chimpanzee monoclonal antibodies to the hepatitis E virus capsid
protein J Virol 74 5548ndash5555
Senn H Klaus W 1993 The nuclear magnetic resonance solution
structure of flavoridin an antagonist of the platelet GPIIbndash IIIa receptor
J Mol Biol 232 907ndash925
Shevchenko A Wilm M Vorm O Mann M 1996 Mass spectrometric
sequencing of protein silver-stained polyacrylamide gels Anal Chem
68 850ndash858
Stevens TJ Arkin IT 2000 Turning an opinion inside-out Rees and
Eisenbergrsquos commentary (Proteins 2000 38121ndash122) on FAre mem-
brane proteins lsquolsquoinside-outrsquorsquo proteins_ (Proteins 1999 36 135ndash143)
Proteins 40 463ndash464
Tsai JM Wang HC Leu JH Hsiao HH Wang AH Kou GH
Lo CF 2004 Genomic and proteomic analysis of thirty-nine
structural proteins of shrimp white spot syndrome virus J Virol 78
11360ndash11370
van Hulten MC Witteveldt J Peters S Kloosterboer N Tarchini
R Fiers M Sandbrink H Lankhorst RK Vlak JM 2001a
The white spot syndrome virus DNA genome sequence Virology
286 7ndash22
van Hulten MC Witteveldt J Snippe M Vlak JM 2001b White spot
syndrome virus envelope protein VP28 is involved in the systemic
infection of shrimp Virology 285 228ndash233
Verdaguer N Mateu MG Andreu D Giralt E Domingo E Fita
I 1995 Structure of the major antigenic loop of foot-and-mouth
disease virus complexed with a neutralizing antibody direct involve-
ment of the ArgndashGlyndashAsp motif in the interaction EMBO J 14
1690ndash1696
Volkman LE Goldsmith PA 1985 Mechanism of neutralization of
L Li et al Virology 340 (2005) 125ndash132132
budded Autographa californica nuclear polyhedrosis virus by a
monoclonal antibody inhibition of entry by adsorptive endocytosis
Virology 143 185ndash195
Wu WL Wang L Zhang XB 2005 Identification of white spot
syndrome virus (WSSV) envelope proteins involved in shrimp
infection Virology 332 578ndash583
Xie XX Yang F 2005 Interaction of white spot syndrome virus VP26
protein with actin Virology 336 93ndash99
Xie XX Li HY Xu LM Yang F 2005 A simple and efficient
method for purification of intact White Spot Syndrome Virus (WSSV)
viral particles Virus Res 108 63ndash67
Yang F He J Lin X Li Q Pan D Zhang X Xu X 2001 Complete
genome sequence of the shrimp white spot bacilliform virus J Virol 75
11811ndash11820
Yingmanee B Pamela JH Farideh G Glyn S 2001 Argininendash
glycinendashaspartic acid motif is critical for human parechovirus 1 entry
J Virol 75 10000ndash10004
Yoganandhan K Syed MS Narayanan RB Sahul HAS 2004
Production of polyclonal antiserum against recombinant VP28 protein
and its application for the detection of white spot syndrome virus in
crustaceans J Fish Dis 27 517ndash522
Zhang X Huang CH Xu X Hew CL 2002a Transcription and
identification of an envelope protein gene (p22) from shrimp white spot
syndrome virus J Gen Virol 83 471ndash477
Zhang X Huang CH Xu X Hew CL 2002b Identification and
localization of a prawn white spot syndrome virus gene that encodes an
envelope protein J Gen Virol 83 1069ndash1074
Zhang X Huang C Tang X Zhuang Y Choy LH 2004 Identification
of structural proteins from shrimp white spot syndrome virus (WSSV)
by 2-DE-MS Proteins Struct Funct Bioinform 55 229ndash235

Fig 2 Nucleotide sequence of WSSV vp31 containing the 5V and 3V terminal region and the deduced VP31 protein sequence (one-letter code) (A) Agarose gel
analysis of the 5V RACE and 3V RACE products Lane M 100 bp ladder DNA marker (Promega) After two rounds of PCR amplifications the PCR products
formed a single 280-bp band in 5V RACE and 320-bp band in 3V RACE (B) The nucleotide sequence of the presumptive WSSV vp31 gene and the deduced
VP31 amino acid sequence The predicted TATA box the possible polyadenylation signal (AATAAT) the start codons two N-glycosylation sites (NLSE
NRTG) and FArgndashGlyndashAsp_ motif were boxed Transcription start point and poly (A) were indicated with black dots Primers used for 5V3VRACE (VP31sp1
VP31sp2 VP31sp3) were underlined
Fig 3 Temporal transcription analysis of WSSV-vp31 at different post-
infection times (hpi) RT-PCR with the gene-specific primers was
performed using the total RNAs extracted from the infected crayfish (A)
RT-PCR with WSSV-vp31-specific primers The transcript was first
detected at 12 h pi and continued to be highly transcribed until 48 h
pi (B) RT-PCR with h-actin-specific primers No RT-PCR amplicon was
observed Lane M 100 bp DNA ladder marker Lane headings showed h
pi (0 2 4 6 8 12 18 24 and 48 h pi)
L Li et al Virology 340 (2005) 125ndash132 127
antibody the gold particles were found to be located on
the surface of the WSSV virions (Fig 4D) while no
particles could be found on the nucleocapsids (Fig 4B) In
the control experiments rVP31c antiserum was replaced
with a pre-immune mouse serum as the primary antibody
no gold particles could be seen for the WSSV virion too
(Fig 4C)
Neutralization assay
The results showed that the crayfish mortalities were
very low for the negative control (09 NaCl) The crayfish
from the positive control (WSSV only) displayed 100
mortality at 8 days post-infection and the crayfish from the
anti-GST IgG control displayed 100 mortality at 9 days
post-infection Whereas after the mixture of WSSV with
anti-rVP31c IgG was injected into crayfish the crayfish
mortality was significantly delayed (Fig 5) indicating that
the infection of WSSV could be delayed or neutralized by
antibodies against rVP31c Deceased shrimps were con-
Fig 4 Localization of VP31 in the WSSV virions by immunoelectron
microscopy (IEM) (Scale bar 90 nm) (A) Electron micrograph of
negatively stained intact WSSV virions (2 PTA) (B) Nucleocapsids of
WSSV incubated with anti-rVP31c antiserum as primary antibody followed
by gold-labeled secondary antibodies (C) Intact WSSV virions incubated
with pre-immune mouse serum as primary antibody followed by gold-
labeled secondary antibodies (D) Intact WSSV virions incubated with anti-
rVP31c antiserum as primary antibody followed by gold-labeled secondary
antibodies Gold particles were indicated by arrows
L Li et al Virology 340 (2005) 125ndash132128
firmed for the presence of WSSV by PCR detection (date
not shown)
Discussion
White spot syndrome virus is a major pathogen of
cultured penaeid shrimp worldwide After the completion of
the WSSV genomic DNA sequencing research has now
Fig 5 Neutralization of WSSV infection in crayfish by specific antibodies The so
crayfish were used After injections the crayfish mortality was monitored daily
mortality on the ordinate
focused on the functional analysis of the gene products
especially on the functions of the viral envelope proteins
Recently a simple and efficient method for purifying the
virions and separating envelope proteins from virions has
been developed in our laboratory (Xie et al 2005) which
facilitated the identification of the viral structural proteins
with low abundance By combining SDS-PAGE with mass
spectrometry vp31 was predicted to encode an envelope
protein This protein was also discovered by another study
in which the VP31 was shown to be a structural protein
without further analysis on protein location and function
(Tsai et al 2004) In this study Western blot showed that
VP31 was in the envelope fraction IEM further revealed
that gold particles were present along the intact WSSV
virions This provided visual evidence that the vp31 gene
encodes one of the envelope proteins of WSSV
Computer analysis showed that the predicted vp31 enco-
ding protein lacks a predominant transmembrane domain
which suggested that VP31 might be a fusion protein that
contained a domain involved in binding to other viral
envelope protein for membrane anchoring The same instance
has been found at the vaccinia virus membrane protein the
VV 14-kDa protein that is located on the surface of the
intracellular mature virus form and is essential for both the
release of extracellular enveloped virus from the cells and
virus spread (Marıa-Isabel et al 1998) Such form had also
been widely reported in some viral membrane fusion pro-
teins pathogen receptors and some cell adhesion molecules
(Boyle and Compton 1998 Jordens et al 1999) These pro-
teins could function to anchor the polypeptides to the
membrane hydrophobic phase by means of locating aqueous
activities or possibly interact with some membrane-spanning
components (Stevens and Arkin 2000) Therefore we pre-
sume that VP31 protein plays an important role in the biology
of the virus acting in virus-to-cell and cell-to-cell fusions
In addition the cell attachment motif RGD (Ruoslahti
and Pierschbacher 1986) occurred in vp31 gene sequence
(Fig 3) suggested that the tripeptide recognition motif might
lutions used for injection were shown at the bottom For each treatment 20
at 25 -C Days post-injection were shown on the abscissa and percentage
L Li et al Virology 340 (2005) 125ndash132 129
have an effect on VP31ndashhost cells interactions In previous
studies RGD structure has been reported widely to mediate
the virusndashhost interaction (Senn and Klaus 1993 Krezel et
al 1994 Verdaguer et al 1995 Yingmanee et al 2001
Mandl et al 1989 Ruoslahti 1996) Among WSSV
proteins six structure proteins including VP31 have been
reported to contain RGD motif (Tsai et al 2004) Moreover
VP31 possesses a threonine at the fourth position of the
RGD motif (RGDT) which was considered to be critical in
interacting with integrin (Plow et al 2000)
Antibodies raised against individual viral envelope
protein have been used successfully in neutralization assay
to identify envelope proteins involved in virus entry during
infection (Martignoni et al 1980 Schofield et al 2000
van Hulten et al 2001a 2001b Volkman and Goldsmith
1985 Wu et al 2005) In this study a specific antiserum
against VP31 protein was prepared to test for neutralization
in vivo The result suggested that VP31 should be involved
in the systemic infection of WSSV in crayfish by partici-
pating in virus attachment and membrane fusion Future
experiments would be performed to demonstrate which part
of VP31 was involved in the neutralization process and what
the role of VP31 in WSSV attachment and entry in the
systemic infection was
In summary we identified a structure protein VP31 as an
envelope protein VP31 might be a key protein in viral
invasion and could be a good candidate for screening
receptors on shrimp cell surface Study on VP31 will be
helpful to realize the mechanism of the virus infection and
to control the infection of WSSV in shrimp industry
Materials and methods
Virus purification and detergent extraction of intact virus
The WSSV isolate in this study originated from Penaeus
japonicus (Xiamen China) and was proliferated in an
alternate host the crayfish (Procambarus clarkii) Prepara-
tion of WSSV virions and nucleocapsids was carried out as
described previously (Xie et al 2005) Brief abundant viral
particles were obtained with only a few steps of conven-
tional differential centrifugations the upper loose pellet was
rinsed out and purified virus in white pellet were
resuspended and kept in TM buffer (50 mM TrisndashHCl 10
mM MgCl2 pH 75)
The virus envelope was removed from the virus
particles by treatment with Triton X-100 Generally the
purified WSSV was mixed with an equal volume of 02
Triton X-100 and incubated for 1 h at room temperature
The nucleocapsids were purified by centrifugation at
20000 g for 20 min at 4 -C The envelope fraction
was collected in the supernatant The nucleocapsid fraction
was subjected to a second round of Triton X-100
extraction to ensure the envelopes were removed com-
pletely The degree of purity of the intact virions isolated
and nucleocapsid fractions were evaluated by negative-
staining transmission electron microscopy
Mass spectrometric analysis of VP31
The intact virions envelope fractions and nucleocapsid
fractions were obtained as above Purified WSSV virions
and the viral envelope fractions were separated using 12
SDS-PAGE (Laemmli 1970) and stained with Coomassie
brilliant blue R-250 A 31 kDa target protein band was
excised from the gel and dehydrated several times with
acetonitrile After vacuum drying the gel band was
incubated with 10 mM dithiothreitol in 100 mM ammonium
bicarbonate (ABC buffer) at 57 -C for 60 min and
subsequently with 55 mM iodoacetamide (Sigma) in 100
mM ABC buffer at room temperature for 60 min Then the
gel was washed with 100 mM ABC buffer and dried In-gel
protein digestion was performed using sequencing grade
modified porcine trypsin (Promega Madison WI) in 50
mM ABC buffer at 37 -C for 15 h Digests were centrifuged
at 6000 g The supernatants were separated and the gel
pieces were extracted further first with 50 acetonitrile 5
formic acid and then with acetonitrile The extracts were
combined with the original digesting supernatants vacuum-
dried and redissolved in 05 trifluoroacetic acid and 50
acetonitrile (Shevchenko et al 1996) Subsequently in-gel-
digested sample was sequenced using Nano-ESI MSMS
mass spectrometry as described previously (Huang et al
2002b) The resulting mass spectrometric data were
searched against the theoretical ORF database of WSSV
Computer analysis of the vp31
The potential sites for posttranslational modifications
were analyzed by the PROSITE database (Hofmann et al
1999) The prediction of transmembrane domain and hydro-
phobic region of WSSV-VP31 was performed with TM-pred
program (Hofman and Stoffel 1993) and DNAMAN
software (Lynnon BioSoft Vaudreuil Canada) respectively
Characterization of WSSV vp31 gene
Rapid amplification of vp31 cDNA ends (5V and 3V RACE)Based on the nucleotide sequence of ORF340 the 5V and
3V ends of the cDNA encoding WSSV-VP31 were obtained
by 5V and 3V RACE using a commercial 5V3V RACE kit
(Roche) according to the manufacturerrsquos recommendations
The RNA samples used in this study were extracted from
WSSV-infected crayfish 24 h pi and then treated with
RNase-free DNase For 5VRACE the first-strand cDNAwas
synthesized using the specific primer sp1 (5V-GGCCGA-TACCAATGTACAAG-3V) and then a poly (A) tail was
added to the cDNA products using terminal transferase in
the presence of dATP The primer sp2 (5V-CTTA-
CAGTGTTCTTCAAC-3V) and an oligo (dT) anchor primer
supplied with the kit were used for PCR For 3VRACE first-
L Li et al Virology 340 (2005) 125ndash132130
strand cDNA was synthesized using an oligo (dT) anchor
primer The primer sp3 (5V-GAGGATGTAGATGCAGTG-3V) and an anchor primer supplied with the kit were used for
PCR The PCR products from 5V and 3V RACE were each
purified on a 2 agarose gel and subcloned into the
pMD18-T vector (TaKaRa) Arbitrarily selected clones were
sequenced and compared with the genomic DNA sequence
of WSSV
Transcriptional analyses of genes
The WSSV inoculum collected from virus-infected
shrimp P japonicus was inoculated into the crayfish
according to the protocol described before (Li et al
2004) Total RNAs at different times (ie 0 2 4 6 8 12
18 24 and 48 h post infection h pi) were extracted from
the hepatopancreas of WSSV-infected crayfish according to
the single-step RNA isolation method (Chomczynski and
Sacchi 1987) The obtained RNAwas further purified using
the SV Total RNA Isolation System (Promega) and
quantified by spectrophotometry at 260 nm The total
RNA (2 Ag) was reverse transcribed with M-MLV RT [H-]
(Promega) using gene-specific reverse primer (GGCCGA-
TACCAATGTACA) The cDNAs were subjected to PCR
using specific forward primer (5V-ATGAAGGATGCA-GAACTG-3V) and reverse primer (GGCCGATACCAATG-
TACA) The PCR cycles were as follows 94 -C for 2 min
30 cycles of 94 -C for 30 s 55 -C for 30 s 72 -C for 1 min
followed by an elongation at 72 -C for 10 min The
crayfish h-actin gene was used as the internal control for
RT-PCR with a gene specific primer set (5V-TCAT-CAGGGTGTGATGGT-3V and 5V-TCTGAGTCATCTTCT-CAC-3V) Total RNA from healthy crayfish was used as the
negative control
Partial expression of GST-VP31 fusion protein in
Escherichia coli
The WSSV-vp31 gene C-terminal fragment (termed as
vp31c) was amplified from WSSV genomic DNA using the
forward primer (5V-CGGGATCC CAAGAAACTGGT-
CAAGT-3V) and the reverse primer (5V-CGGAATTC
CTCCTCCTTAAAAGCAG-3V) that contained recognition
sequences for BamHI and EcoRI restriction enzymes
(underlined) The amplicon was cloned into the pET-GST
vector (Gene Power Lab Ltd Shenzhen office) The
recombinant plasmid was transformed into E coli BL21
(DE3) cells Liquid cultures were grown in a shaking
incubator (200 rpm) at 37 -C until the OD600 reached 07
and these were then induced with 02 mM IPTG for 5 h at
37 -C The cells were harvested by centrifugation at
4000 g for 5 min The recombinant VP31c (termed
rVP31c) was purified by Ni-NTA affinity chromatography
under denatured conditions following methodology in the
QIAexpressionist handbook (Qiagen) The E coli cells
containing pET-GST vector were also induced with IPTG
and total protein extracts were applied to the Ni-NTA
column as described above Final eluates were collected and
used as the negative control
Preparation of antibody
Purified rVP31c protein was used as antigen to immunize
mice by intradermal injection once every 10 days Antigen
(30 Ag) was mixed with an equal volume of Freundrsquos
complete adjuvant (Sigma) for the first injection Subse-
quent three injections were conducted using 30 Ag antigen
mixed with an equal volume of Freundrsquos incomplete
adjuvant (Sigma) Four days after the last injection mice
were exsanguinated and antisera were collected The titers
of the antisera were 115000 as determined by ELISA The
IgG fraction was purified with rProtein AndashSepharose Fast
Flow (Amersham) The optimal dilution of purified IgG
after serial dilutions was 15000 as determined by ELISA
Horseradish-peroxidase (HRP)-conjugated goat anti-mouse
IgG was obtained from Promega For a negative control
antigen was replaced with 1 PBS
Western blot
Virus envelope fraction and nucleocapsid fraction dis-
solved in loading buffer were separated by SDS-PAGE
(Laemmli 1970) respectively These proteins were trans-
ferred onto a PVDF membrane (Amersham Pharmacia) The
membrane was then immersed in blocking buffer (2 BSA
20 mM Tris 150 mM NaCl 01 Tween 20 pH 75) at
room temperature for 30 min followed by incubation with
anti-rVP31c serum (diluted 15000) for 1 h Following this
alkaline-phosphatase-conjugated goat anti-mouse IgG
(Promega) was used as the secondary antibody Detection
was performed using Western Blue Stabilized Substrate for
Alkaline Phosphatase (Promega)
Localization of VP31 protein by immunoelectron
microscopy
WSSV virion and nucleocapsid suspension were mounted
on carbon-coated nickel grids (300 meshes) for 1 h at room
temperature Respectively after washing with PBS the grids
were blocked in 3 BSA for 1 h Then the grids were rinsed
with PBS followed by incubation with anti-rVP31c IgG as
the primary antibody (1200 diluted in 3 BSA) for 2 h
After washing four times with 1 PBS 10-nm-gold-labeled
anti-mouse goat IgG as the secondary antibody (Sigma) was
added to the grids and incubated for 1 h at room temperature
Grids were washed further four times with 1 PBS and
stained with 2 phosphotungstic acid (PTA pH 70) for 25
min Specimens were examined under a TEM For control
experiment anti-rVP31c IgG was replaced with a pre-
immune mouse serum as the primary antibodies indicated
above
Neutralization assay
Prior to injection into crayfish WSSV virions were
incubated with anti-rVP31c IgG or the anti-GST IgG as a
L Li et al Virology 340 (2005) 125ndash132 131
control (final concentration 4 Ag IgGml 107 virionsml) for
1 h at room temperature Then the 100 Al mixture of
antibody and WSSV was intramuscularly injected into
crayfish using a syringe with 29-gauge needle At the same
time a negative (09 NaCl) and a positive control (virus
only 107 virionsml) were included in the injections For
each treatment 20 crayfish were used After injections the
crayfish mortality was monitored daily at 25 -C Deceasedshrimps were examined for the presence of WSSV by PCR
detection
Acknowledgments
We thank Ping Chen for her technical assistance in the
transmission electron microscopy This work was sup-
ported by the lsquolsquo863rsquorsquo Program of China (2003AA626020)
National Natural Science Foundation of China (30330470
40276038) and Fujian Science Fund (2003F001)
References
Boyle KA Compton T 1998 Receptor-binding properties of a
soluble form of human cytomegalovirus glycoprotein B J Virol 72
1826ndash1833
Caron H van Schaik B van der Mee M Baas F Riggins G van
Sluis P Hermus M-C van Asperen R Boon K Voute PA
Heisterkamp S van Kamper A Versteeg R 2001 The human
transcriptome map clustering of highly expressed genes in chromoso-
mal domainsmdashSupplementary material Science 291 1289ndash1292
Chen XF Chen C Wu DH Huai H Chi XC 1997 A new
baculovirus of cultured shrimp Sci China Ser C 40 630ndash635
Chen LL Leu JH 1 Huang CJ Chou CM Chen SM Wang CH
Lo CF Kou GH 2002 Identification of a nucleocapsid protein
(VP35) gene of shrimp white spot syndrome virus and characterization
of the motif important for targeting VP35 to the nuclei of transfected
insect cells Virology 293 44ndash53
Chomczynski P Sacchi N 1987 Single-step method of RNA isolation
by acid guanidinium thiocyanatendashphenolndashchloroform extraction Anal
Biochem 262 156ndash159
Flegel TW 1997 Special topic review major viral diseases of the black
tiger prawn (Penaeus monodon) in Thailand World J Microbiol
Biotechnol 13 433ndash442
Halder SK Takemori H Hatano O Nonaka Y Wada A Okamoto
M 1998 Cloning of a membrane-spanning protein with epidermal
growth factor-like repeat motifs from adrenal glomerulosa cells
Endocrinology 139 3316ndash3328
Hofman K Stoffel W 1993 TMbasemdashA database of membrane
spanning protein segments Biol Chem Hoppe-Seyler 374 166
Hofmann K Bucher P Falquet L Bairoch A 1999 The PROSITE
database its status in 1999 Nucleic Acids Res 27 215ndash219
Huang CH Zhang XB Lin QS Xu X Hew CL 2002a Character-
ization of a novel envelope protein (VP281) of shrimp white spot
syndrome virus by mass spectrometry J Gen Virol 83 2385ndash2392
Huang CH Zhang XB Lin QS Xu X Hu ZH Hew CL 2002b
Proteomic analysis of shrimp white spot syndrome viral proteins and
characterization of a novel envelope protein VP466 Mol Cell
Proteomics 1 223ndash231
Jordens R Thompson A Amons R Koning F 1999 Human dendritic
cells shed a functional soluble form of the mannose receptor Int
Immunol 11 1775ndash1780
Krezel AM Wagner G Seymour-Ulmer J Lazarus RA 1994
Structure of the RGD protein decorsin conserved motif and distinct
function in leech proteins that affect blood clotting Science 264
1944ndash1947
Laemmli UK 1970 Cleavage of structural proteins during the assembly
of the head of bacteriophage T4 Nature 227 680ndash685
Li Q Pan D Zhang J Yang F 2004 Identification of the thymidylate
synthase within the genome of white spot syndrome virus J Gen Virol
85 2035ndash2044
Lo CF Ho CH Peng SE Chen CH Hsu HC Chiu YL Chang
CF Liu KF Su MS Wang CH Kou GH 1996a White spot
syndrome baculovirus (WSBV) detected in cultured and captured
shrimp crabs and other arthropods Dis Aquat Org 27 215ndash226
Lo CF Leu JH Ho CH Chen CH Peng SE Chen YT Chou
CM Yeh PY Huang CJ Chou HY Wang CH Kou GH
1996b Detection of baculovirus associated with white spot syndrome
(WSBV) in penaeid shrimps using polymerase chain reaction Dis
Aquat Org 25 133ndash141
Mandl CW Guirakhoo F Holzmann H Heinz FX Kunz C 1989
Antigenic structure of the flavivirus envelope E protein at the molecular
level using tick-borne encephalitis virus as a model J Virol 63
564ndash571
Marıa-Isabel V German R David C Luis S Mariano E 1998
The vaccinia virus 14-kilodalton (A27L) fusion protein forms a triple
coiled-coil structure and interacts with the 21-kilodalton (A17L)
virus membrane protein through a C-terminal a-helix J Virol 72
10126ndash10137
Martignoni ME Iwai PJ Rohrmann GF 1980 Serum neutralization
of nucleopolyhedrosis viruses (baculovirus subgroup A) pathogenic for
Orgyia pseudotsugata J Invert Pathol 36 12ndash20
Plow EF Haas TA Zhang L Loftus J Smith JW 2000 Ligand
binding to integrins J Biol Chem 275 21785ndash21788
Ruoslahti E 1996 RGD and other recognition sequences for integrins
Annu Rev Cell Biol 12 697ndash715
Ruoslahti E Pierschbacher MD 1986 ArgndashGlyndashAsp a versatile cell
recognition signal Cell 44 517ndash518
Schofield DJ Glamann J Emerson U Purchell RH 2000 Identi-
fication by phage display and characterization of two neutralizing
chimpanzee monoclonal antibodies to the hepatitis E virus capsid
protein J Virol 74 5548ndash5555
Senn H Klaus W 1993 The nuclear magnetic resonance solution
structure of flavoridin an antagonist of the platelet GPIIbndash IIIa receptor
J Mol Biol 232 907ndash925
Shevchenko A Wilm M Vorm O Mann M 1996 Mass spectrometric
sequencing of protein silver-stained polyacrylamide gels Anal Chem
68 850ndash858
Stevens TJ Arkin IT 2000 Turning an opinion inside-out Rees and
Eisenbergrsquos commentary (Proteins 2000 38121ndash122) on FAre mem-
brane proteins lsquolsquoinside-outrsquorsquo proteins_ (Proteins 1999 36 135ndash143)
Proteins 40 463ndash464
Tsai JM Wang HC Leu JH Hsiao HH Wang AH Kou GH
Lo CF 2004 Genomic and proteomic analysis of thirty-nine
structural proteins of shrimp white spot syndrome virus J Virol 78
11360ndash11370
van Hulten MC Witteveldt J Peters S Kloosterboer N Tarchini
R Fiers M Sandbrink H Lankhorst RK Vlak JM 2001a
The white spot syndrome virus DNA genome sequence Virology
286 7ndash22
van Hulten MC Witteveldt J Snippe M Vlak JM 2001b White spot
syndrome virus envelope protein VP28 is involved in the systemic
infection of shrimp Virology 285 228ndash233
Verdaguer N Mateu MG Andreu D Giralt E Domingo E Fita
I 1995 Structure of the major antigenic loop of foot-and-mouth
disease virus complexed with a neutralizing antibody direct involve-
ment of the ArgndashGlyndashAsp motif in the interaction EMBO J 14
1690ndash1696
Volkman LE Goldsmith PA 1985 Mechanism of neutralization of
L Li et al Virology 340 (2005) 125ndash132132
budded Autographa californica nuclear polyhedrosis virus by a
monoclonal antibody inhibition of entry by adsorptive endocytosis
Virology 143 185ndash195
Wu WL Wang L Zhang XB 2005 Identification of white spot
syndrome virus (WSSV) envelope proteins involved in shrimp
infection Virology 332 578ndash583
Xie XX Yang F 2005 Interaction of white spot syndrome virus VP26
protein with actin Virology 336 93ndash99
Xie XX Li HY Xu LM Yang F 2005 A simple and efficient
method for purification of intact White Spot Syndrome Virus (WSSV)
viral particles Virus Res 108 63ndash67
Yang F He J Lin X Li Q Pan D Zhang X Xu X 2001 Complete
genome sequence of the shrimp white spot bacilliform virus J Virol 75
11811ndash11820
Yingmanee B Pamela JH Farideh G Glyn S 2001 Argininendash
glycinendashaspartic acid motif is critical for human parechovirus 1 entry
J Virol 75 10000ndash10004
Yoganandhan K Syed MS Narayanan RB Sahul HAS 2004
Production of polyclonal antiserum against recombinant VP28 protein
and its application for the detection of white spot syndrome virus in
crustaceans J Fish Dis 27 517ndash522
Zhang X Huang CH Xu X Hew CL 2002a Transcription and
identification of an envelope protein gene (p22) from shrimp white spot
syndrome virus J Gen Virol 83 471ndash477
Zhang X Huang CH Xu X Hew CL 2002b Identification and
localization of a prawn white spot syndrome virus gene that encodes an
envelope protein J Gen Virol 83 1069ndash1074
Zhang X Huang C Tang X Zhuang Y Choy LH 2004 Identification
of structural proteins from shrimp white spot syndrome virus (WSSV)
by 2-DE-MS Proteins Struct Funct Bioinform 55 229ndash235
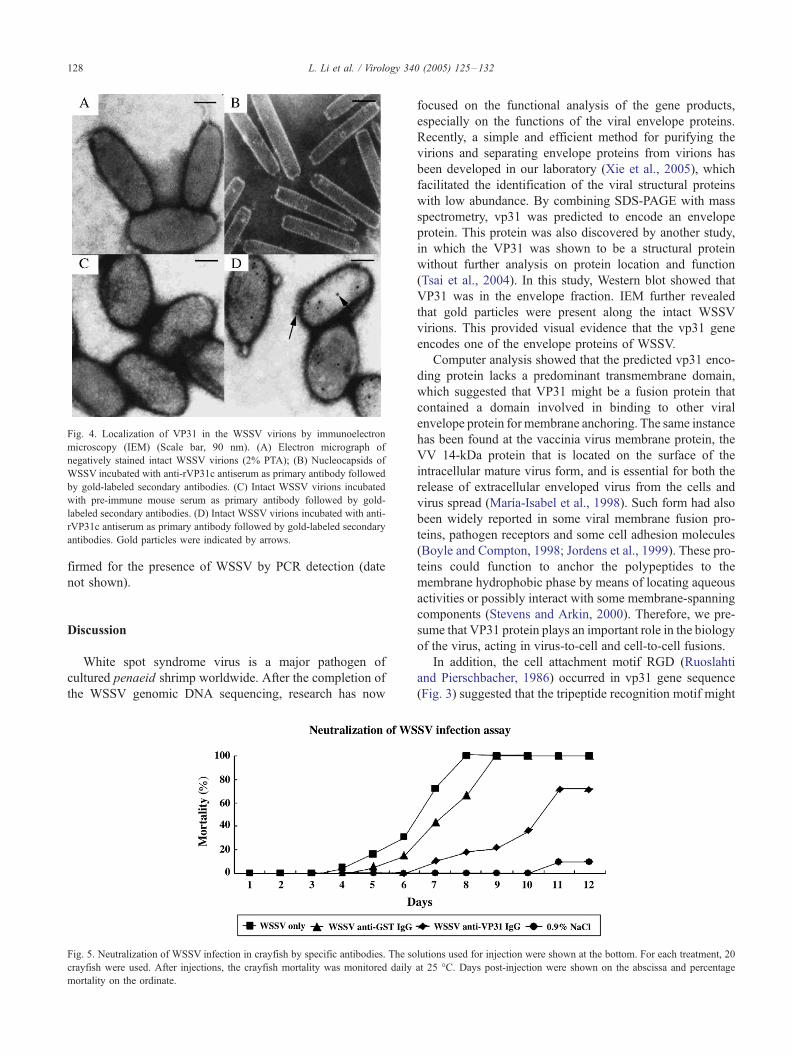
Fig 4 Localization of VP31 in the WSSV virions by immunoelectron
microscopy (IEM) (Scale bar 90 nm) (A) Electron micrograph of
negatively stained intact WSSV virions (2 PTA) (B) Nucleocapsids of
WSSV incubated with anti-rVP31c antiserum as primary antibody followed
by gold-labeled secondary antibodies (C) Intact WSSV virions incubated
with pre-immune mouse serum as primary antibody followed by gold-
labeled secondary antibodies (D) Intact WSSV virions incubated with anti-
rVP31c antiserum as primary antibody followed by gold-labeled secondary
antibodies Gold particles were indicated by arrows
L Li et al Virology 340 (2005) 125ndash132128
firmed for the presence of WSSV by PCR detection (date
not shown)
Discussion
White spot syndrome virus is a major pathogen of
cultured penaeid shrimp worldwide After the completion of
the WSSV genomic DNA sequencing research has now
Fig 5 Neutralization of WSSV infection in crayfish by specific antibodies The so
crayfish were used After injections the crayfish mortality was monitored daily
mortality on the ordinate
focused on the functional analysis of the gene products
especially on the functions of the viral envelope proteins
Recently a simple and efficient method for purifying the
virions and separating envelope proteins from virions has
been developed in our laboratory (Xie et al 2005) which
facilitated the identification of the viral structural proteins
with low abundance By combining SDS-PAGE with mass
spectrometry vp31 was predicted to encode an envelope
protein This protein was also discovered by another study
in which the VP31 was shown to be a structural protein
without further analysis on protein location and function
(Tsai et al 2004) In this study Western blot showed that
VP31 was in the envelope fraction IEM further revealed
that gold particles were present along the intact WSSV
virions This provided visual evidence that the vp31 gene
encodes one of the envelope proteins of WSSV
Computer analysis showed that the predicted vp31 enco-
ding protein lacks a predominant transmembrane domain
which suggested that VP31 might be a fusion protein that
contained a domain involved in binding to other viral
envelope protein for membrane anchoring The same instance
has been found at the vaccinia virus membrane protein the
VV 14-kDa protein that is located on the surface of the
intracellular mature virus form and is essential for both the
release of extracellular enveloped virus from the cells and
virus spread (Marıa-Isabel et al 1998) Such form had also
been widely reported in some viral membrane fusion pro-
teins pathogen receptors and some cell adhesion molecules
(Boyle and Compton 1998 Jordens et al 1999) These pro-
teins could function to anchor the polypeptides to the
membrane hydrophobic phase by means of locating aqueous
activities or possibly interact with some membrane-spanning
components (Stevens and Arkin 2000) Therefore we pre-
sume that VP31 protein plays an important role in the biology
of the virus acting in virus-to-cell and cell-to-cell fusions
In addition the cell attachment motif RGD (Ruoslahti
and Pierschbacher 1986) occurred in vp31 gene sequence
(Fig 3) suggested that the tripeptide recognition motif might
lutions used for injection were shown at the bottom For each treatment 20
at 25 -C Days post-injection were shown on the abscissa and percentage
L Li et al Virology 340 (2005) 125ndash132 129
have an effect on VP31ndashhost cells interactions In previous
studies RGD structure has been reported widely to mediate
the virusndashhost interaction (Senn and Klaus 1993 Krezel et
al 1994 Verdaguer et al 1995 Yingmanee et al 2001
Mandl et al 1989 Ruoslahti 1996) Among WSSV
proteins six structure proteins including VP31 have been
reported to contain RGD motif (Tsai et al 2004) Moreover
VP31 possesses a threonine at the fourth position of the
RGD motif (RGDT) which was considered to be critical in
interacting with integrin (Plow et al 2000)
Antibodies raised against individual viral envelope
protein have been used successfully in neutralization assay
to identify envelope proteins involved in virus entry during
infection (Martignoni et al 1980 Schofield et al 2000
van Hulten et al 2001a 2001b Volkman and Goldsmith
1985 Wu et al 2005) In this study a specific antiserum
against VP31 protein was prepared to test for neutralization
in vivo The result suggested that VP31 should be involved
in the systemic infection of WSSV in crayfish by partici-
pating in virus attachment and membrane fusion Future
experiments would be performed to demonstrate which part
of VP31 was involved in the neutralization process and what
the role of VP31 in WSSV attachment and entry in the
systemic infection was
In summary we identified a structure protein VP31 as an
envelope protein VP31 might be a key protein in viral
invasion and could be a good candidate for screening
receptors on shrimp cell surface Study on VP31 will be
helpful to realize the mechanism of the virus infection and
to control the infection of WSSV in shrimp industry
Materials and methods
Virus purification and detergent extraction of intact virus
The WSSV isolate in this study originated from Penaeus
japonicus (Xiamen China) and was proliferated in an
alternate host the crayfish (Procambarus clarkii) Prepara-
tion of WSSV virions and nucleocapsids was carried out as
described previously (Xie et al 2005) Brief abundant viral
particles were obtained with only a few steps of conven-
tional differential centrifugations the upper loose pellet was
rinsed out and purified virus in white pellet were
resuspended and kept in TM buffer (50 mM TrisndashHCl 10
mM MgCl2 pH 75)
The virus envelope was removed from the virus
particles by treatment with Triton X-100 Generally the
purified WSSV was mixed with an equal volume of 02
Triton X-100 and incubated for 1 h at room temperature
The nucleocapsids were purified by centrifugation at
20000 g for 20 min at 4 -C The envelope fraction
was collected in the supernatant The nucleocapsid fraction
was subjected to a second round of Triton X-100
extraction to ensure the envelopes were removed com-
pletely The degree of purity of the intact virions isolated
and nucleocapsid fractions were evaluated by negative-
staining transmission electron microscopy
Mass spectrometric analysis of VP31
The intact virions envelope fractions and nucleocapsid
fractions were obtained as above Purified WSSV virions
and the viral envelope fractions were separated using 12
SDS-PAGE (Laemmli 1970) and stained with Coomassie
brilliant blue R-250 A 31 kDa target protein band was
excised from the gel and dehydrated several times with
acetonitrile After vacuum drying the gel band was
incubated with 10 mM dithiothreitol in 100 mM ammonium
bicarbonate (ABC buffer) at 57 -C for 60 min and
subsequently with 55 mM iodoacetamide (Sigma) in 100
mM ABC buffer at room temperature for 60 min Then the
gel was washed with 100 mM ABC buffer and dried In-gel
protein digestion was performed using sequencing grade
modified porcine trypsin (Promega Madison WI) in 50
mM ABC buffer at 37 -C for 15 h Digests were centrifuged
at 6000 g The supernatants were separated and the gel
pieces were extracted further first with 50 acetonitrile 5
formic acid and then with acetonitrile The extracts were
combined with the original digesting supernatants vacuum-
dried and redissolved in 05 trifluoroacetic acid and 50
acetonitrile (Shevchenko et al 1996) Subsequently in-gel-
digested sample was sequenced using Nano-ESI MSMS
mass spectrometry as described previously (Huang et al
2002b) The resulting mass spectrometric data were
searched against the theoretical ORF database of WSSV
Computer analysis of the vp31
The potential sites for posttranslational modifications
were analyzed by the PROSITE database (Hofmann et al
1999) The prediction of transmembrane domain and hydro-
phobic region of WSSV-VP31 was performed with TM-pred
program (Hofman and Stoffel 1993) and DNAMAN
software (Lynnon BioSoft Vaudreuil Canada) respectively
Characterization of WSSV vp31 gene
Rapid amplification of vp31 cDNA ends (5V and 3V RACE)Based on the nucleotide sequence of ORF340 the 5V and
3V ends of the cDNA encoding WSSV-VP31 were obtained
by 5V and 3V RACE using a commercial 5V3V RACE kit
(Roche) according to the manufacturerrsquos recommendations
The RNA samples used in this study were extracted from
WSSV-infected crayfish 24 h pi and then treated with
RNase-free DNase For 5VRACE the first-strand cDNAwas
synthesized using the specific primer sp1 (5V-GGCCGA-TACCAATGTACAAG-3V) and then a poly (A) tail was
added to the cDNA products using terminal transferase in
the presence of dATP The primer sp2 (5V-CTTA-
CAGTGTTCTTCAAC-3V) and an oligo (dT) anchor primer
supplied with the kit were used for PCR For 3VRACE first-
L Li et al Virology 340 (2005) 125ndash132130
strand cDNA was synthesized using an oligo (dT) anchor
primer The primer sp3 (5V-GAGGATGTAGATGCAGTG-3V) and an anchor primer supplied with the kit were used for
PCR The PCR products from 5V and 3V RACE were each
purified on a 2 agarose gel and subcloned into the
pMD18-T vector (TaKaRa) Arbitrarily selected clones were
sequenced and compared with the genomic DNA sequence
of WSSV
Transcriptional analyses of genes
The WSSV inoculum collected from virus-infected
shrimp P japonicus was inoculated into the crayfish
according to the protocol described before (Li et al
2004) Total RNAs at different times (ie 0 2 4 6 8 12
18 24 and 48 h post infection h pi) were extracted from
the hepatopancreas of WSSV-infected crayfish according to
the single-step RNA isolation method (Chomczynski and
Sacchi 1987) The obtained RNAwas further purified using
the SV Total RNA Isolation System (Promega) and
quantified by spectrophotometry at 260 nm The total
RNA (2 Ag) was reverse transcribed with M-MLV RT [H-]
(Promega) using gene-specific reverse primer (GGCCGA-
TACCAATGTACA) The cDNAs were subjected to PCR
using specific forward primer (5V-ATGAAGGATGCA-GAACTG-3V) and reverse primer (GGCCGATACCAATG-
TACA) The PCR cycles were as follows 94 -C for 2 min
30 cycles of 94 -C for 30 s 55 -C for 30 s 72 -C for 1 min
followed by an elongation at 72 -C for 10 min The
crayfish h-actin gene was used as the internal control for
RT-PCR with a gene specific primer set (5V-TCAT-CAGGGTGTGATGGT-3V and 5V-TCTGAGTCATCTTCT-CAC-3V) Total RNA from healthy crayfish was used as the
negative control
Partial expression of GST-VP31 fusion protein in
Escherichia coli
The WSSV-vp31 gene C-terminal fragment (termed as
vp31c) was amplified from WSSV genomic DNA using the
forward primer (5V-CGGGATCC CAAGAAACTGGT-
CAAGT-3V) and the reverse primer (5V-CGGAATTC
CTCCTCCTTAAAAGCAG-3V) that contained recognition
sequences for BamHI and EcoRI restriction enzymes
(underlined) The amplicon was cloned into the pET-GST
vector (Gene Power Lab Ltd Shenzhen office) The
recombinant plasmid was transformed into E coli BL21
(DE3) cells Liquid cultures were grown in a shaking
incubator (200 rpm) at 37 -C until the OD600 reached 07
and these were then induced with 02 mM IPTG for 5 h at
37 -C The cells were harvested by centrifugation at
4000 g for 5 min The recombinant VP31c (termed
rVP31c) was purified by Ni-NTA affinity chromatography
under denatured conditions following methodology in the
QIAexpressionist handbook (Qiagen) The E coli cells
containing pET-GST vector were also induced with IPTG
and total protein extracts were applied to the Ni-NTA
column as described above Final eluates were collected and
used as the negative control
Preparation of antibody
Purified rVP31c protein was used as antigen to immunize
mice by intradermal injection once every 10 days Antigen
(30 Ag) was mixed with an equal volume of Freundrsquos
complete adjuvant (Sigma) for the first injection Subse-
quent three injections were conducted using 30 Ag antigen
mixed with an equal volume of Freundrsquos incomplete
adjuvant (Sigma) Four days after the last injection mice
were exsanguinated and antisera were collected The titers
of the antisera were 115000 as determined by ELISA The
IgG fraction was purified with rProtein AndashSepharose Fast
Flow (Amersham) The optimal dilution of purified IgG
after serial dilutions was 15000 as determined by ELISA
Horseradish-peroxidase (HRP)-conjugated goat anti-mouse
IgG was obtained from Promega For a negative control
antigen was replaced with 1 PBS
Western blot
Virus envelope fraction and nucleocapsid fraction dis-
solved in loading buffer were separated by SDS-PAGE
(Laemmli 1970) respectively These proteins were trans-
ferred onto a PVDF membrane (Amersham Pharmacia) The
membrane was then immersed in blocking buffer (2 BSA
20 mM Tris 150 mM NaCl 01 Tween 20 pH 75) at
room temperature for 30 min followed by incubation with
anti-rVP31c serum (diluted 15000) for 1 h Following this
alkaline-phosphatase-conjugated goat anti-mouse IgG
(Promega) was used as the secondary antibody Detection
was performed using Western Blue Stabilized Substrate for
Alkaline Phosphatase (Promega)
Localization of VP31 protein by immunoelectron
microscopy
WSSV virion and nucleocapsid suspension were mounted
on carbon-coated nickel grids (300 meshes) for 1 h at room
temperature Respectively after washing with PBS the grids
were blocked in 3 BSA for 1 h Then the grids were rinsed
with PBS followed by incubation with anti-rVP31c IgG as
the primary antibody (1200 diluted in 3 BSA) for 2 h
After washing four times with 1 PBS 10-nm-gold-labeled
anti-mouse goat IgG as the secondary antibody (Sigma) was
added to the grids and incubated for 1 h at room temperature
Grids were washed further four times with 1 PBS and
stained with 2 phosphotungstic acid (PTA pH 70) for 25
min Specimens were examined under a TEM For control
experiment anti-rVP31c IgG was replaced with a pre-
immune mouse serum as the primary antibodies indicated
above
Neutralization assay
Prior to injection into crayfish WSSV virions were
incubated with anti-rVP31c IgG or the anti-GST IgG as a
L Li et al Virology 340 (2005) 125ndash132 131
control (final concentration 4 Ag IgGml 107 virionsml) for
1 h at room temperature Then the 100 Al mixture of
antibody and WSSV was intramuscularly injected into
crayfish using a syringe with 29-gauge needle At the same
time a negative (09 NaCl) and a positive control (virus
only 107 virionsml) were included in the injections For
each treatment 20 crayfish were used After injections the
crayfish mortality was monitored daily at 25 -C Deceasedshrimps were examined for the presence of WSSV by PCR
detection
Acknowledgments
We thank Ping Chen for her technical assistance in the
transmission electron microscopy This work was sup-
ported by the lsquolsquo863rsquorsquo Program of China (2003AA626020)
National Natural Science Foundation of China (30330470
40276038) and Fujian Science Fund (2003F001)
References
Boyle KA Compton T 1998 Receptor-binding properties of a
soluble form of human cytomegalovirus glycoprotein B J Virol 72
1826ndash1833
Caron H van Schaik B van der Mee M Baas F Riggins G van
Sluis P Hermus M-C van Asperen R Boon K Voute PA
Heisterkamp S van Kamper A Versteeg R 2001 The human
transcriptome map clustering of highly expressed genes in chromoso-
mal domainsmdashSupplementary material Science 291 1289ndash1292
Chen XF Chen C Wu DH Huai H Chi XC 1997 A new
baculovirus of cultured shrimp Sci China Ser C 40 630ndash635
Chen LL Leu JH 1 Huang CJ Chou CM Chen SM Wang CH
Lo CF Kou GH 2002 Identification of a nucleocapsid protein
(VP35) gene of shrimp white spot syndrome virus and characterization
of the motif important for targeting VP35 to the nuclei of transfected
insect cells Virology 293 44ndash53
Chomczynski P Sacchi N 1987 Single-step method of RNA isolation
by acid guanidinium thiocyanatendashphenolndashchloroform extraction Anal
Biochem 262 156ndash159
Flegel TW 1997 Special topic review major viral diseases of the black
tiger prawn (Penaeus monodon) in Thailand World J Microbiol
Biotechnol 13 433ndash442
Halder SK Takemori H Hatano O Nonaka Y Wada A Okamoto
M 1998 Cloning of a membrane-spanning protein with epidermal
growth factor-like repeat motifs from adrenal glomerulosa cells
Endocrinology 139 3316ndash3328
Hofman K Stoffel W 1993 TMbasemdashA database of membrane
spanning protein segments Biol Chem Hoppe-Seyler 374 166
Hofmann K Bucher P Falquet L Bairoch A 1999 The PROSITE
database its status in 1999 Nucleic Acids Res 27 215ndash219
Huang CH Zhang XB Lin QS Xu X Hew CL 2002a Character-
ization of a novel envelope protein (VP281) of shrimp white spot
syndrome virus by mass spectrometry J Gen Virol 83 2385ndash2392
Huang CH Zhang XB Lin QS Xu X Hu ZH Hew CL 2002b
Proteomic analysis of shrimp white spot syndrome viral proteins and
characterization of a novel envelope protein VP466 Mol Cell
Proteomics 1 223ndash231
Jordens R Thompson A Amons R Koning F 1999 Human dendritic
cells shed a functional soluble form of the mannose receptor Int
Immunol 11 1775ndash1780
Krezel AM Wagner G Seymour-Ulmer J Lazarus RA 1994
Structure of the RGD protein decorsin conserved motif and distinct
function in leech proteins that affect blood clotting Science 264
1944ndash1947
Laemmli UK 1970 Cleavage of structural proteins during the assembly
of the head of bacteriophage T4 Nature 227 680ndash685
Li Q Pan D Zhang J Yang F 2004 Identification of the thymidylate
synthase within the genome of white spot syndrome virus J Gen Virol
85 2035ndash2044
Lo CF Ho CH Peng SE Chen CH Hsu HC Chiu YL Chang
CF Liu KF Su MS Wang CH Kou GH 1996a White spot
syndrome baculovirus (WSBV) detected in cultured and captured
shrimp crabs and other arthropods Dis Aquat Org 27 215ndash226
Lo CF Leu JH Ho CH Chen CH Peng SE Chen YT Chou
CM Yeh PY Huang CJ Chou HY Wang CH Kou GH
1996b Detection of baculovirus associated with white spot syndrome
(WSBV) in penaeid shrimps using polymerase chain reaction Dis
Aquat Org 25 133ndash141
Mandl CW Guirakhoo F Holzmann H Heinz FX Kunz C 1989
Antigenic structure of the flavivirus envelope E protein at the molecular
level using tick-borne encephalitis virus as a model J Virol 63
564ndash571
Marıa-Isabel V German R David C Luis S Mariano E 1998
The vaccinia virus 14-kilodalton (A27L) fusion protein forms a triple
coiled-coil structure and interacts with the 21-kilodalton (A17L)
virus membrane protein through a C-terminal a-helix J Virol 72
10126ndash10137
Martignoni ME Iwai PJ Rohrmann GF 1980 Serum neutralization
of nucleopolyhedrosis viruses (baculovirus subgroup A) pathogenic for
Orgyia pseudotsugata J Invert Pathol 36 12ndash20
Plow EF Haas TA Zhang L Loftus J Smith JW 2000 Ligand
binding to integrins J Biol Chem 275 21785ndash21788
Ruoslahti E 1996 RGD and other recognition sequences for integrins
Annu Rev Cell Biol 12 697ndash715
Ruoslahti E Pierschbacher MD 1986 ArgndashGlyndashAsp a versatile cell
recognition signal Cell 44 517ndash518
Schofield DJ Glamann J Emerson U Purchell RH 2000 Identi-
fication by phage display and characterization of two neutralizing
chimpanzee monoclonal antibodies to the hepatitis E virus capsid
protein J Virol 74 5548ndash5555
Senn H Klaus W 1993 The nuclear magnetic resonance solution
structure of flavoridin an antagonist of the platelet GPIIbndash IIIa receptor
J Mol Biol 232 907ndash925
Shevchenko A Wilm M Vorm O Mann M 1996 Mass spectrometric
sequencing of protein silver-stained polyacrylamide gels Anal Chem
68 850ndash858
Stevens TJ Arkin IT 2000 Turning an opinion inside-out Rees and
Eisenbergrsquos commentary (Proteins 2000 38121ndash122) on FAre mem-
brane proteins lsquolsquoinside-outrsquorsquo proteins_ (Proteins 1999 36 135ndash143)
Proteins 40 463ndash464
Tsai JM Wang HC Leu JH Hsiao HH Wang AH Kou GH
Lo CF 2004 Genomic and proteomic analysis of thirty-nine
structural proteins of shrimp white spot syndrome virus J Virol 78
11360ndash11370
van Hulten MC Witteveldt J Peters S Kloosterboer N Tarchini
R Fiers M Sandbrink H Lankhorst RK Vlak JM 2001a
The white spot syndrome virus DNA genome sequence Virology
286 7ndash22
van Hulten MC Witteveldt J Snippe M Vlak JM 2001b White spot
syndrome virus envelope protein VP28 is involved in the systemic
infection of shrimp Virology 285 228ndash233
Verdaguer N Mateu MG Andreu D Giralt E Domingo E Fita
I 1995 Structure of the major antigenic loop of foot-and-mouth
disease virus complexed with a neutralizing antibody direct involve-
ment of the ArgndashGlyndashAsp motif in the interaction EMBO J 14
1690ndash1696
Volkman LE Goldsmith PA 1985 Mechanism of neutralization of
L Li et al Virology 340 (2005) 125ndash132132
budded Autographa californica nuclear polyhedrosis virus by a
monoclonal antibody inhibition of entry by adsorptive endocytosis
Virology 143 185ndash195
Wu WL Wang L Zhang XB 2005 Identification of white spot
syndrome virus (WSSV) envelope proteins involved in shrimp
infection Virology 332 578ndash583
Xie XX Yang F 2005 Interaction of white spot syndrome virus VP26
protein with actin Virology 336 93ndash99
Xie XX Li HY Xu LM Yang F 2005 A simple and efficient
method for purification of intact White Spot Syndrome Virus (WSSV)
viral particles Virus Res 108 63ndash67
Yang F He J Lin X Li Q Pan D Zhang X Xu X 2001 Complete
genome sequence of the shrimp white spot bacilliform virus J Virol 75
11811ndash11820
Yingmanee B Pamela JH Farideh G Glyn S 2001 Argininendash
glycinendashaspartic acid motif is critical for human parechovirus 1 entry
J Virol 75 10000ndash10004
Yoganandhan K Syed MS Narayanan RB Sahul HAS 2004
Production of polyclonal antiserum against recombinant VP28 protein
and its application for the detection of white spot syndrome virus in
crustaceans J Fish Dis 27 517ndash522
Zhang X Huang CH Xu X Hew CL 2002a Transcription and
identification of an envelope protein gene (p22) from shrimp white spot
syndrome virus J Gen Virol 83 471ndash477
Zhang X Huang CH Xu X Hew CL 2002b Identification and
localization of a prawn white spot syndrome virus gene that encodes an
envelope protein J Gen Virol 83 1069ndash1074
Zhang X Huang C Tang X Zhuang Y Choy LH 2004 Identification
of structural proteins from shrimp white spot syndrome virus (WSSV)
by 2-DE-MS Proteins Struct Funct Bioinform 55 229ndash235

L Li et al Virology 340 (2005) 125ndash132 129
have an effect on VP31ndashhost cells interactions In previous
studies RGD structure has been reported widely to mediate
the virusndashhost interaction (Senn and Klaus 1993 Krezel et
al 1994 Verdaguer et al 1995 Yingmanee et al 2001
Mandl et al 1989 Ruoslahti 1996) Among WSSV
proteins six structure proteins including VP31 have been
reported to contain RGD motif (Tsai et al 2004) Moreover
VP31 possesses a threonine at the fourth position of the
RGD motif (RGDT) which was considered to be critical in
interacting with integrin (Plow et al 2000)
Antibodies raised against individual viral envelope
protein have been used successfully in neutralization assay
to identify envelope proteins involved in virus entry during
infection (Martignoni et al 1980 Schofield et al 2000
van Hulten et al 2001a 2001b Volkman and Goldsmith
1985 Wu et al 2005) In this study a specific antiserum
against VP31 protein was prepared to test for neutralization
in vivo The result suggested that VP31 should be involved
in the systemic infection of WSSV in crayfish by partici-
pating in virus attachment and membrane fusion Future
experiments would be performed to demonstrate which part
of VP31 was involved in the neutralization process and what
the role of VP31 in WSSV attachment and entry in the
systemic infection was
In summary we identified a structure protein VP31 as an
envelope protein VP31 might be a key protein in viral
invasion and could be a good candidate for screening
receptors on shrimp cell surface Study on VP31 will be
helpful to realize the mechanism of the virus infection and
to control the infection of WSSV in shrimp industry
Materials and methods
Virus purification and detergent extraction of intact virus
The WSSV isolate in this study originated from Penaeus
japonicus (Xiamen China) and was proliferated in an
alternate host the crayfish (Procambarus clarkii) Prepara-
tion of WSSV virions and nucleocapsids was carried out as
described previously (Xie et al 2005) Brief abundant viral
particles were obtained with only a few steps of conven-
tional differential centrifugations the upper loose pellet was
rinsed out and purified virus in white pellet were
resuspended and kept in TM buffer (50 mM TrisndashHCl 10
mM MgCl2 pH 75)
The virus envelope was removed from the virus
particles by treatment with Triton X-100 Generally the
purified WSSV was mixed with an equal volume of 02
Triton X-100 and incubated for 1 h at room temperature
The nucleocapsids were purified by centrifugation at
20000 g for 20 min at 4 -C The envelope fraction
was collected in the supernatant The nucleocapsid fraction
was subjected to a second round of Triton X-100
extraction to ensure the envelopes were removed com-
pletely The degree of purity of the intact virions isolated
and nucleocapsid fractions were evaluated by negative-
staining transmission electron microscopy
Mass spectrometric analysis of VP31
The intact virions envelope fractions and nucleocapsid
fractions were obtained as above Purified WSSV virions
and the viral envelope fractions were separated using 12
SDS-PAGE (Laemmli 1970) and stained with Coomassie
brilliant blue R-250 A 31 kDa target protein band was
excised from the gel and dehydrated several times with
acetonitrile After vacuum drying the gel band was
incubated with 10 mM dithiothreitol in 100 mM ammonium
bicarbonate (ABC buffer) at 57 -C for 60 min and
subsequently with 55 mM iodoacetamide (Sigma) in 100
mM ABC buffer at room temperature for 60 min Then the
gel was washed with 100 mM ABC buffer and dried In-gel
protein digestion was performed using sequencing grade
modified porcine trypsin (Promega Madison WI) in 50
mM ABC buffer at 37 -C for 15 h Digests were centrifuged
at 6000 g The supernatants were separated and the gel
pieces were extracted further first with 50 acetonitrile 5
formic acid and then with acetonitrile The extracts were
combined with the original digesting supernatants vacuum-
dried and redissolved in 05 trifluoroacetic acid and 50
acetonitrile (Shevchenko et al 1996) Subsequently in-gel-
digested sample was sequenced using Nano-ESI MSMS
mass spectrometry as described previously (Huang et al
2002b) The resulting mass spectrometric data were
searched against the theoretical ORF database of WSSV
Computer analysis of the vp31
The potential sites for posttranslational modifications
were analyzed by the PROSITE database (Hofmann et al
1999) The prediction of transmembrane domain and hydro-
phobic region of WSSV-VP31 was performed with TM-pred
program (Hofman and Stoffel 1993) and DNAMAN
software (Lynnon BioSoft Vaudreuil Canada) respectively
Characterization of WSSV vp31 gene
Rapid amplification of vp31 cDNA ends (5V and 3V RACE)Based on the nucleotide sequence of ORF340 the 5V and
3V ends of the cDNA encoding WSSV-VP31 were obtained
by 5V and 3V RACE using a commercial 5V3V RACE kit
(Roche) according to the manufacturerrsquos recommendations
The RNA samples used in this study were extracted from
WSSV-infected crayfish 24 h pi and then treated with
RNase-free DNase For 5VRACE the first-strand cDNAwas
synthesized using the specific primer sp1 (5V-GGCCGA-TACCAATGTACAAG-3V) and then a poly (A) tail was
added to the cDNA products using terminal transferase in
the presence of dATP The primer sp2 (5V-CTTA-
CAGTGTTCTTCAAC-3V) and an oligo (dT) anchor primer
supplied with the kit were used for PCR For 3VRACE first-
L Li et al Virology 340 (2005) 125ndash132130
strand cDNA was synthesized using an oligo (dT) anchor
primer The primer sp3 (5V-GAGGATGTAGATGCAGTG-3V) and an anchor primer supplied with the kit were used for
PCR The PCR products from 5V and 3V RACE were each
purified on a 2 agarose gel and subcloned into the
pMD18-T vector (TaKaRa) Arbitrarily selected clones were
sequenced and compared with the genomic DNA sequence
of WSSV
Transcriptional analyses of genes
The WSSV inoculum collected from virus-infected
shrimp P japonicus was inoculated into the crayfish
according to the protocol described before (Li et al
2004) Total RNAs at different times (ie 0 2 4 6 8 12
18 24 and 48 h post infection h pi) were extracted from
the hepatopancreas of WSSV-infected crayfish according to
the single-step RNA isolation method (Chomczynski and
Sacchi 1987) The obtained RNAwas further purified using
the SV Total RNA Isolation System (Promega) and
quantified by spectrophotometry at 260 nm The total
RNA (2 Ag) was reverse transcribed with M-MLV RT [H-]
(Promega) using gene-specific reverse primer (GGCCGA-
TACCAATGTACA) The cDNAs were subjected to PCR
using specific forward primer (5V-ATGAAGGATGCA-GAACTG-3V) and reverse primer (GGCCGATACCAATG-
TACA) The PCR cycles were as follows 94 -C for 2 min
30 cycles of 94 -C for 30 s 55 -C for 30 s 72 -C for 1 min
followed by an elongation at 72 -C for 10 min The
crayfish h-actin gene was used as the internal control for
RT-PCR with a gene specific primer set (5V-TCAT-CAGGGTGTGATGGT-3V and 5V-TCTGAGTCATCTTCT-CAC-3V) Total RNA from healthy crayfish was used as the
negative control
Partial expression of GST-VP31 fusion protein in
Escherichia coli
The WSSV-vp31 gene C-terminal fragment (termed as
vp31c) was amplified from WSSV genomic DNA using the
forward primer (5V-CGGGATCC CAAGAAACTGGT-
CAAGT-3V) and the reverse primer (5V-CGGAATTC
CTCCTCCTTAAAAGCAG-3V) that contained recognition
sequences for BamHI and EcoRI restriction enzymes
(underlined) The amplicon was cloned into the pET-GST
vector (Gene Power Lab Ltd Shenzhen office) The
recombinant plasmid was transformed into E coli BL21
(DE3) cells Liquid cultures were grown in a shaking
incubator (200 rpm) at 37 -C until the OD600 reached 07
and these were then induced with 02 mM IPTG for 5 h at
37 -C The cells were harvested by centrifugation at
4000 g for 5 min The recombinant VP31c (termed
rVP31c) was purified by Ni-NTA affinity chromatography
under denatured conditions following methodology in the
QIAexpressionist handbook (Qiagen) The E coli cells
containing pET-GST vector were also induced with IPTG
and total protein extracts were applied to the Ni-NTA
column as described above Final eluates were collected and
used as the negative control
Preparation of antibody
Purified rVP31c protein was used as antigen to immunize
mice by intradermal injection once every 10 days Antigen
(30 Ag) was mixed with an equal volume of Freundrsquos
complete adjuvant (Sigma) for the first injection Subse-
quent three injections were conducted using 30 Ag antigen
mixed with an equal volume of Freundrsquos incomplete
adjuvant (Sigma) Four days after the last injection mice
were exsanguinated and antisera were collected The titers
of the antisera were 115000 as determined by ELISA The
IgG fraction was purified with rProtein AndashSepharose Fast
Flow (Amersham) The optimal dilution of purified IgG
after serial dilutions was 15000 as determined by ELISA
Horseradish-peroxidase (HRP)-conjugated goat anti-mouse
IgG was obtained from Promega For a negative control
antigen was replaced with 1 PBS
Western blot
Virus envelope fraction and nucleocapsid fraction dis-
solved in loading buffer were separated by SDS-PAGE
(Laemmli 1970) respectively These proteins were trans-
ferred onto a PVDF membrane (Amersham Pharmacia) The
membrane was then immersed in blocking buffer (2 BSA
20 mM Tris 150 mM NaCl 01 Tween 20 pH 75) at
room temperature for 30 min followed by incubation with
anti-rVP31c serum (diluted 15000) for 1 h Following this
alkaline-phosphatase-conjugated goat anti-mouse IgG
(Promega) was used as the secondary antibody Detection
was performed using Western Blue Stabilized Substrate for
Alkaline Phosphatase (Promega)
Localization of VP31 protein by immunoelectron
microscopy
WSSV virion and nucleocapsid suspension were mounted
on carbon-coated nickel grids (300 meshes) for 1 h at room
temperature Respectively after washing with PBS the grids
were blocked in 3 BSA for 1 h Then the grids were rinsed
with PBS followed by incubation with anti-rVP31c IgG as
the primary antibody (1200 diluted in 3 BSA) for 2 h
After washing four times with 1 PBS 10-nm-gold-labeled
anti-mouse goat IgG as the secondary antibody (Sigma) was
added to the grids and incubated for 1 h at room temperature
Grids were washed further four times with 1 PBS and
stained with 2 phosphotungstic acid (PTA pH 70) for 25
min Specimens were examined under a TEM For control
experiment anti-rVP31c IgG was replaced with a pre-
immune mouse serum as the primary antibodies indicated
above
Neutralization assay
Prior to injection into crayfish WSSV virions were
incubated with anti-rVP31c IgG or the anti-GST IgG as a
L Li et al Virology 340 (2005) 125ndash132 131
control (final concentration 4 Ag IgGml 107 virionsml) for
1 h at room temperature Then the 100 Al mixture of
antibody and WSSV was intramuscularly injected into
crayfish using a syringe with 29-gauge needle At the same
time a negative (09 NaCl) and a positive control (virus
only 107 virionsml) were included in the injections For
each treatment 20 crayfish were used After injections the
crayfish mortality was monitored daily at 25 -C Deceasedshrimps were examined for the presence of WSSV by PCR
detection
Acknowledgments
We thank Ping Chen for her technical assistance in the
transmission electron microscopy This work was sup-
ported by the lsquolsquo863rsquorsquo Program of China (2003AA626020)
National Natural Science Foundation of China (30330470
40276038) and Fujian Science Fund (2003F001)
References
Boyle KA Compton T 1998 Receptor-binding properties of a
soluble form of human cytomegalovirus glycoprotein B J Virol 72
1826ndash1833
Caron H van Schaik B van der Mee M Baas F Riggins G van
Sluis P Hermus M-C van Asperen R Boon K Voute PA
Heisterkamp S van Kamper A Versteeg R 2001 The human
transcriptome map clustering of highly expressed genes in chromoso-
mal domainsmdashSupplementary material Science 291 1289ndash1292
Chen XF Chen C Wu DH Huai H Chi XC 1997 A new
baculovirus of cultured shrimp Sci China Ser C 40 630ndash635
Chen LL Leu JH 1 Huang CJ Chou CM Chen SM Wang CH
Lo CF Kou GH 2002 Identification of a nucleocapsid protein
(VP35) gene of shrimp white spot syndrome virus and characterization
of the motif important for targeting VP35 to the nuclei of transfected
insect cells Virology 293 44ndash53
Chomczynski P Sacchi N 1987 Single-step method of RNA isolation
by acid guanidinium thiocyanatendashphenolndashchloroform extraction Anal
Biochem 262 156ndash159
Flegel TW 1997 Special topic review major viral diseases of the black
tiger prawn (Penaeus monodon) in Thailand World J Microbiol
Biotechnol 13 433ndash442
Halder SK Takemori H Hatano O Nonaka Y Wada A Okamoto
M 1998 Cloning of a membrane-spanning protein with epidermal
growth factor-like repeat motifs from adrenal glomerulosa cells
Endocrinology 139 3316ndash3328
Hofman K Stoffel W 1993 TMbasemdashA database of membrane
spanning protein segments Biol Chem Hoppe-Seyler 374 166
Hofmann K Bucher P Falquet L Bairoch A 1999 The PROSITE
database its status in 1999 Nucleic Acids Res 27 215ndash219
Huang CH Zhang XB Lin QS Xu X Hew CL 2002a Character-
ization of a novel envelope protein (VP281) of shrimp white spot
syndrome virus by mass spectrometry J Gen Virol 83 2385ndash2392
Huang CH Zhang XB Lin QS Xu X Hu ZH Hew CL 2002b
Proteomic analysis of shrimp white spot syndrome viral proteins and
characterization of a novel envelope protein VP466 Mol Cell
Proteomics 1 223ndash231
Jordens R Thompson A Amons R Koning F 1999 Human dendritic
cells shed a functional soluble form of the mannose receptor Int
Immunol 11 1775ndash1780
Krezel AM Wagner G Seymour-Ulmer J Lazarus RA 1994
Structure of the RGD protein decorsin conserved motif and distinct
function in leech proteins that affect blood clotting Science 264
1944ndash1947
Laemmli UK 1970 Cleavage of structural proteins during the assembly
of the head of bacteriophage T4 Nature 227 680ndash685
Li Q Pan D Zhang J Yang F 2004 Identification of the thymidylate
synthase within the genome of white spot syndrome virus J Gen Virol
85 2035ndash2044
Lo CF Ho CH Peng SE Chen CH Hsu HC Chiu YL Chang
CF Liu KF Su MS Wang CH Kou GH 1996a White spot
syndrome baculovirus (WSBV) detected in cultured and captured
shrimp crabs and other arthropods Dis Aquat Org 27 215ndash226
Lo CF Leu JH Ho CH Chen CH Peng SE Chen YT Chou
CM Yeh PY Huang CJ Chou HY Wang CH Kou GH
1996b Detection of baculovirus associated with white spot syndrome
(WSBV) in penaeid shrimps using polymerase chain reaction Dis
Aquat Org 25 133ndash141
Mandl CW Guirakhoo F Holzmann H Heinz FX Kunz C 1989
Antigenic structure of the flavivirus envelope E protein at the molecular
level using tick-borne encephalitis virus as a model J Virol 63
564ndash571
Marıa-Isabel V German R David C Luis S Mariano E 1998
The vaccinia virus 14-kilodalton (A27L) fusion protein forms a triple
coiled-coil structure and interacts with the 21-kilodalton (A17L)
virus membrane protein through a C-terminal a-helix J Virol 72
10126ndash10137
Martignoni ME Iwai PJ Rohrmann GF 1980 Serum neutralization
of nucleopolyhedrosis viruses (baculovirus subgroup A) pathogenic for
Orgyia pseudotsugata J Invert Pathol 36 12ndash20
Plow EF Haas TA Zhang L Loftus J Smith JW 2000 Ligand
binding to integrins J Biol Chem 275 21785ndash21788
Ruoslahti E 1996 RGD and other recognition sequences for integrins
Annu Rev Cell Biol 12 697ndash715
Ruoslahti E Pierschbacher MD 1986 ArgndashGlyndashAsp a versatile cell
recognition signal Cell 44 517ndash518
Schofield DJ Glamann J Emerson U Purchell RH 2000 Identi-
fication by phage display and characterization of two neutralizing
chimpanzee monoclonal antibodies to the hepatitis E virus capsid
protein J Virol 74 5548ndash5555
Senn H Klaus W 1993 The nuclear magnetic resonance solution
structure of flavoridin an antagonist of the platelet GPIIbndash IIIa receptor
J Mol Biol 232 907ndash925
Shevchenko A Wilm M Vorm O Mann M 1996 Mass spectrometric
sequencing of protein silver-stained polyacrylamide gels Anal Chem
68 850ndash858
Stevens TJ Arkin IT 2000 Turning an opinion inside-out Rees and
Eisenbergrsquos commentary (Proteins 2000 38121ndash122) on FAre mem-
brane proteins lsquolsquoinside-outrsquorsquo proteins_ (Proteins 1999 36 135ndash143)
Proteins 40 463ndash464
Tsai JM Wang HC Leu JH Hsiao HH Wang AH Kou GH
Lo CF 2004 Genomic and proteomic analysis of thirty-nine
structural proteins of shrimp white spot syndrome virus J Virol 78
11360ndash11370
van Hulten MC Witteveldt J Peters S Kloosterboer N Tarchini
R Fiers M Sandbrink H Lankhorst RK Vlak JM 2001a
The white spot syndrome virus DNA genome sequence Virology
286 7ndash22
van Hulten MC Witteveldt J Snippe M Vlak JM 2001b White spot
syndrome virus envelope protein VP28 is involved in the systemic
infection of shrimp Virology 285 228ndash233
Verdaguer N Mateu MG Andreu D Giralt E Domingo E Fita
I 1995 Structure of the major antigenic loop of foot-and-mouth
disease virus complexed with a neutralizing antibody direct involve-
ment of the ArgndashGlyndashAsp motif in the interaction EMBO J 14
1690ndash1696
Volkman LE Goldsmith PA 1985 Mechanism of neutralization of
L Li et al Virology 340 (2005) 125ndash132132
budded Autographa californica nuclear polyhedrosis virus by a
monoclonal antibody inhibition of entry by adsorptive endocytosis
Virology 143 185ndash195
Wu WL Wang L Zhang XB 2005 Identification of white spot
syndrome virus (WSSV) envelope proteins involved in shrimp
infection Virology 332 578ndash583
Xie XX Yang F 2005 Interaction of white spot syndrome virus VP26
protein with actin Virology 336 93ndash99
Xie XX Li HY Xu LM Yang F 2005 A simple and efficient
method for purification of intact White Spot Syndrome Virus (WSSV)
viral particles Virus Res 108 63ndash67
Yang F He J Lin X Li Q Pan D Zhang X Xu X 2001 Complete
genome sequence of the shrimp white spot bacilliform virus J Virol 75
11811ndash11820
Yingmanee B Pamela JH Farideh G Glyn S 2001 Argininendash
glycinendashaspartic acid motif is critical for human parechovirus 1 entry
J Virol 75 10000ndash10004
Yoganandhan K Syed MS Narayanan RB Sahul HAS 2004
Production of polyclonal antiserum against recombinant VP28 protein
and its application for the detection of white spot syndrome virus in
crustaceans J Fish Dis 27 517ndash522
Zhang X Huang CH Xu X Hew CL 2002a Transcription and
identification of an envelope protein gene (p22) from shrimp white spot
syndrome virus J Gen Virol 83 471ndash477
Zhang X Huang CH Xu X Hew CL 2002b Identification and
localization of a prawn white spot syndrome virus gene that encodes an
envelope protein J Gen Virol 83 1069ndash1074
Zhang X Huang C Tang X Zhuang Y Choy LH 2004 Identification
of structural proteins from shrimp white spot syndrome virus (WSSV)
by 2-DE-MS Proteins Struct Funct Bioinform 55 229ndash235

L Li et al Virology 340 (2005) 125ndash132130
strand cDNA was synthesized using an oligo (dT) anchor
primer The primer sp3 (5V-GAGGATGTAGATGCAGTG-3V) and an anchor primer supplied with the kit were used for
PCR The PCR products from 5V and 3V RACE were each
purified on a 2 agarose gel and subcloned into the
pMD18-T vector (TaKaRa) Arbitrarily selected clones were
sequenced and compared with the genomic DNA sequence
of WSSV
Transcriptional analyses of genes
The WSSV inoculum collected from virus-infected
shrimp P japonicus was inoculated into the crayfish
according to the protocol described before (Li et al
2004) Total RNAs at different times (ie 0 2 4 6 8 12
18 24 and 48 h post infection h pi) were extracted from
the hepatopancreas of WSSV-infected crayfish according to
the single-step RNA isolation method (Chomczynski and
Sacchi 1987) The obtained RNAwas further purified using
the SV Total RNA Isolation System (Promega) and
quantified by spectrophotometry at 260 nm The total
RNA (2 Ag) was reverse transcribed with M-MLV RT [H-]
(Promega) using gene-specific reverse primer (GGCCGA-
TACCAATGTACA) The cDNAs were subjected to PCR
using specific forward primer (5V-ATGAAGGATGCA-GAACTG-3V) and reverse primer (GGCCGATACCAATG-
TACA) The PCR cycles were as follows 94 -C for 2 min
30 cycles of 94 -C for 30 s 55 -C for 30 s 72 -C for 1 min
followed by an elongation at 72 -C for 10 min The
crayfish h-actin gene was used as the internal control for
RT-PCR with a gene specific primer set (5V-TCAT-CAGGGTGTGATGGT-3V and 5V-TCTGAGTCATCTTCT-CAC-3V) Total RNA from healthy crayfish was used as the
negative control
Partial expression of GST-VP31 fusion protein in
Escherichia coli
The WSSV-vp31 gene C-terminal fragment (termed as
vp31c) was amplified from WSSV genomic DNA using the
forward primer (5V-CGGGATCC CAAGAAACTGGT-
CAAGT-3V) and the reverse primer (5V-CGGAATTC
CTCCTCCTTAAAAGCAG-3V) that contained recognition
sequences for BamHI and EcoRI restriction enzymes
(underlined) The amplicon was cloned into the pET-GST
vector (Gene Power Lab Ltd Shenzhen office) The
recombinant plasmid was transformed into E coli BL21
(DE3) cells Liquid cultures were grown in a shaking
incubator (200 rpm) at 37 -C until the OD600 reached 07
and these were then induced with 02 mM IPTG for 5 h at
37 -C The cells were harvested by centrifugation at
4000 g for 5 min The recombinant VP31c (termed
rVP31c) was purified by Ni-NTA affinity chromatography
under denatured conditions following methodology in the
QIAexpressionist handbook (Qiagen) The E coli cells
containing pET-GST vector were also induced with IPTG
and total protein extracts were applied to the Ni-NTA
column as described above Final eluates were collected and
used as the negative control
Preparation of antibody
Purified rVP31c protein was used as antigen to immunize
mice by intradermal injection once every 10 days Antigen
(30 Ag) was mixed with an equal volume of Freundrsquos
complete adjuvant (Sigma) for the first injection Subse-
quent three injections were conducted using 30 Ag antigen
mixed with an equal volume of Freundrsquos incomplete
adjuvant (Sigma) Four days after the last injection mice
were exsanguinated and antisera were collected The titers
of the antisera were 115000 as determined by ELISA The
IgG fraction was purified with rProtein AndashSepharose Fast
Flow (Amersham) The optimal dilution of purified IgG
after serial dilutions was 15000 as determined by ELISA
Horseradish-peroxidase (HRP)-conjugated goat anti-mouse
IgG was obtained from Promega For a negative control
antigen was replaced with 1 PBS
Western blot
Virus envelope fraction and nucleocapsid fraction dis-
solved in loading buffer were separated by SDS-PAGE
(Laemmli 1970) respectively These proteins were trans-
ferred onto a PVDF membrane (Amersham Pharmacia) The
membrane was then immersed in blocking buffer (2 BSA
20 mM Tris 150 mM NaCl 01 Tween 20 pH 75) at
room temperature for 30 min followed by incubation with
anti-rVP31c serum (diluted 15000) for 1 h Following this
alkaline-phosphatase-conjugated goat anti-mouse IgG
(Promega) was used as the secondary antibody Detection
was performed using Western Blue Stabilized Substrate for
Alkaline Phosphatase (Promega)
Localization of VP31 protein by immunoelectron
microscopy
WSSV virion and nucleocapsid suspension were mounted
on carbon-coated nickel grids (300 meshes) for 1 h at room
temperature Respectively after washing with PBS the grids
were blocked in 3 BSA for 1 h Then the grids were rinsed
with PBS followed by incubation with anti-rVP31c IgG as
the primary antibody (1200 diluted in 3 BSA) for 2 h
After washing four times with 1 PBS 10-nm-gold-labeled
anti-mouse goat IgG as the secondary antibody (Sigma) was
added to the grids and incubated for 1 h at room temperature
Grids were washed further four times with 1 PBS and
stained with 2 phosphotungstic acid (PTA pH 70) for 25
min Specimens were examined under a TEM For control
experiment anti-rVP31c IgG was replaced with a pre-
immune mouse serum as the primary antibodies indicated
above
Neutralization assay
Prior to injection into crayfish WSSV virions were
incubated with anti-rVP31c IgG or the anti-GST IgG as a
L Li et al Virology 340 (2005) 125ndash132 131
control (final concentration 4 Ag IgGml 107 virionsml) for
1 h at room temperature Then the 100 Al mixture of
antibody and WSSV was intramuscularly injected into
crayfish using a syringe with 29-gauge needle At the same
time a negative (09 NaCl) and a positive control (virus
only 107 virionsml) were included in the injections For
each treatment 20 crayfish were used After injections the
crayfish mortality was monitored daily at 25 -C Deceasedshrimps were examined for the presence of WSSV by PCR
detection
Acknowledgments
We thank Ping Chen for her technical assistance in the
transmission electron microscopy This work was sup-
ported by the lsquolsquo863rsquorsquo Program of China (2003AA626020)
National Natural Science Foundation of China (30330470
40276038) and Fujian Science Fund (2003F001)
References
Boyle KA Compton T 1998 Receptor-binding properties of a
soluble form of human cytomegalovirus glycoprotein B J Virol 72
1826ndash1833
Caron H van Schaik B van der Mee M Baas F Riggins G van
Sluis P Hermus M-C van Asperen R Boon K Voute PA
Heisterkamp S van Kamper A Versteeg R 2001 The human
transcriptome map clustering of highly expressed genes in chromoso-
mal domainsmdashSupplementary material Science 291 1289ndash1292
Chen XF Chen C Wu DH Huai H Chi XC 1997 A new
baculovirus of cultured shrimp Sci China Ser C 40 630ndash635
Chen LL Leu JH 1 Huang CJ Chou CM Chen SM Wang CH
Lo CF Kou GH 2002 Identification of a nucleocapsid protein
(VP35) gene of shrimp white spot syndrome virus and characterization
of the motif important for targeting VP35 to the nuclei of transfected
insect cells Virology 293 44ndash53
Chomczynski P Sacchi N 1987 Single-step method of RNA isolation
by acid guanidinium thiocyanatendashphenolndashchloroform extraction Anal
Biochem 262 156ndash159
Flegel TW 1997 Special topic review major viral diseases of the black
tiger prawn (Penaeus monodon) in Thailand World J Microbiol
Biotechnol 13 433ndash442
Halder SK Takemori H Hatano O Nonaka Y Wada A Okamoto
M 1998 Cloning of a membrane-spanning protein with epidermal
growth factor-like repeat motifs from adrenal glomerulosa cells
Endocrinology 139 3316ndash3328
Hofman K Stoffel W 1993 TMbasemdashA database of membrane
spanning protein segments Biol Chem Hoppe-Seyler 374 166
Hofmann K Bucher P Falquet L Bairoch A 1999 The PROSITE
database its status in 1999 Nucleic Acids Res 27 215ndash219
Huang CH Zhang XB Lin QS Xu X Hew CL 2002a Character-
ization of a novel envelope protein (VP281) of shrimp white spot
syndrome virus by mass spectrometry J Gen Virol 83 2385ndash2392
Huang CH Zhang XB Lin QS Xu X Hu ZH Hew CL 2002b
Proteomic analysis of shrimp white spot syndrome viral proteins and
characterization of a novel envelope protein VP466 Mol Cell
Proteomics 1 223ndash231
Jordens R Thompson A Amons R Koning F 1999 Human dendritic
cells shed a functional soluble form of the mannose receptor Int
Immunol 11 1775ndash1780
Krezel AM Wagner G Seymour-Ulmer J Lazarus RA 1994
Structure of the RGD protein decorsin conserved motif and distinct
function in leech proteins that affect blood clotting Science 264
1944ndash1947
Laemmli UK 1970 Cleavage of structural proteins during the assembly
of the head of bacteriophage T4 Nature 227 680ndash685
Li Q Pan D Zhang J Yang F 2004 Identification of the thymidylate
synthase within the genome of white spot syndrome virus J Gen Virol
85 2035ndash2044
Lo CF Ho CH Peng SE Chen CH Hsu HC Chiu YL Chang
CF Liu KF Su MS Wang CH Kou GH 1996a White spot
syndrome baculovirus (WSBV) detected in cultured and captured
shrimp crabs and other arthropods Dis Aquat Org 27 215ndash226
Lo CF Leu JH Ho CH Chen CH Peng SE Chen YT Chou
CM Yeh PY Huang CJ Chou HY Wang CH Kou GH
1996b Detection of baculovirus associated with white spot syndrome
(WSBV) in penaeid shrimps using polymerase chain reaction Dis
Aquat Org 25 133ndash141
Mandl CW Guirakhoo F Holzmann H Heinz FX Kunz C 1989
Antigenic structure of the flavivirus envelope E protein at the molecular
level using tick-borne encephalitis virus as a model J Virol 63
564ndash571
Marıa-Isabel V German R David C Luis S Mariano E 1998
The vaccinia virus 14-kilodalton (A27L) fusion protein forms a triple
coiled-coil structure and interacts with the 21-kilodalton (A17L)
virus membrane protein through a C-terminal a-helix J Virol 72
10126ndash10137
Martignoni ME Iwai PJ Rohrmann GF 1980 Serum neutralization
of nucleopolyhedrosis viruses (baculovirus subgroup A) pathogenic for
Orgyia pseudotsugata J Invert Pathol 36 12ndash20
Plow EF Haas TA Zhang L Loftus J Smith JW 2000 Ligand
binding to integrins J Biol Chem 275 21785ndash21788
Ruoslahti E 1996 RGD and other recognition sequences for integrins
Annu Rev Cell Biol 12 697ndash715
Ruoslahti E Pierschbacher MD 1986 ArgndashGlyndashAsp a versatile cell
recognition signal Cell 44 517ndash518
Schofield DJ Glamann J Emerson U Purchell RH 2000 Identi-
fication by phage display and characterization of two neutralizing
chimpanzee monoclonal antibodies to the hepatitis E virus capsid
protein J Virol 74 5548ndash5555
Senn H Klaus W 1993 The nuclear magnetic resonance solution
structure of flavoridin an antagonist of the platelet GPIIbndash IIIa receptor
J Mol Biol 232 907ndash925
Shevchenko A Wilm M Vorm O Mann M 1996 Mass spectrometric
sequencing of protein silver-stained polyacrylamide gels Anal Chem
68 850ndash858
Stevens TJ Arkin IT 2000 Turning an opinion inside-out Rees and
Eisenbergrsquos commentary (Proteins 2000 38121ndash122) on FAre mem-
brane proteins lsquolsquoinside-outrsquorsquo proteins_ (Proteins 1999 36 135ndash143)
Proteins 40 463ndash464
Tsai JM Wang HC Leu JH Hsiao HH Wang AH Kou GH
Lo CF 2004 Genomic and proteomic analysis of thirty-nine
structural proteins of shrimp white spot syndrome virus J Virol 78
11360ndash11370
van Hulten MC Witteveldt J Peters S Kloosterboer N Tarchini
R Fiers M Sandbrink H Lankhorst RK Vlak JM 2001a
The white spot syndrome virus DNA genome sequence Virology
286 7ndash22
van Hulten MC Witteveldt J Snippe M Vlak JM 2001b White spot
syndrome virus envelope protein VP28 is involved in the systemic
infection of shrimp Virology 285 228ndash233
Verdaguer N Mateu MG Andreu D Giralt E Domingo E Fita
I 1995 Structure of the major antigenic loop of foot-and-mouth
disease virus complexed with a neutralizing antibody direct involve-
ment of the ArgndashGlyndashAsp motif in the interaction EMBO J 14
1690ndash1696
Volkman LE Goldsmith PA 1985 Mechanism of neutralization of
L Li et al Virology 340 (2005) 125ndash132132
budded Autographa californica nuclear polyhedrosis virus by a
monoclonal antibody inhibition of entry by adsorptive endocytosis
Virology 143 185ndash195
Wu WL Wang L Zhang XB 2005 Identification of white spot
syndrome virus (WSSV) envelope proteins involved in shrimp
infection Virology 332 578ndash583
Xie XX Yang F 2005 Interaction of white spot syndrome virus VP26
protein with actin Virology 336 93ndash99
Xie XX Li HY Xu LM Yang F 2005 A simple and efficient
method for purification of intact White Spot Syndrome Virus (WSSV)
viral particles Virus Res 108 63ndash67
Yang F He J Lin X Li Q Pan D Zhang X Xu X 2001 Complete
genome sequence of the shrimp white spot bacilliform virus J Virol 75
11811ndash11820
Yingmanee B Pamela JH Farideh G Glyn S 2001 Argininendash
glycinendashaspartic acid motif is critical for human parechovirus 1 entry
J Virol 75 10000ndash10004
Yoganandhan K Syed MS Narayanan RB Sahul HAS 2004
Production of polyclonal antiserum against recombinant VP28 protein
and its application for the detection of white spot syndrome virus in
crustaceans J Fish Dis 27 517ndash522
Zhang X Huang CH Xu X Hew CL 2002a Transcription and
identification of an envelope protein gene (p22) from shrimp white spot
syndrome virus J Gen Virol 83 471ndash477
Zhang X Huang CH Xu X Hew CL 2002b Identification and
localization of a prawn white spot syndrome virus gene that encodes an
envelope protein J Gen Virol 83 1069ndash1074
Zhang X Huang C Tang X Zhuang Y Choy LH 2004 Identification
of structural proteins from shrimp white spot syndrome virus (WSSV)
by 2-DE-MS Proteins Struct Funct Bioinform 55 229ndash235

L Li et al Virology 340 (2005) 125ndash132 131
control (final concentration 4 Ag IgGml 107 virionsml) for
1 h at room temperature Then the 100 Al mixture of
antibody and WSSV was intramuscularly injected into
crayfish using a syringe with 29-gauge needle At the same
time a negative (09 NaCl) and a positive control (virus
only 107 virionsml) were included in the injections For
each treatment 20 crayfish were used After injections the
crayfish mortality was monitored daily at 25 -C Deceasedshrimps were examined for the presence of WSSV by PCR
detection
Acknowledgments
We thank Ping Chen for her technical assistance in the
transmission electron microscopy This work was sup-
ported by the lsquolsquo863rsquorsquo Program of China (2003AA626020)
National Natural Science Foundation of China (30330470
40276038) and Fujian Science Fund (2003F001)
References
Boyle KA Compton T 1998 Receptor-binding properties of a
soluble form of human cytomegalovirus glycoprotein B J Virol 72
1826ndash1833
Caron H van Schaik B van der Mee M Baas F Riggins G van
Sluis P Hermus M-C van Asperen R Boon K Voute PA
Heisterkamp S van Kamper A Versteeg R 2001 The human
transcriptome map clustering of highly expressed genes in chromoso-
mal domainsmdashSupplementary material Science 291 1289ndash1292
Chen XF Chen C Wu DH Huai H Chi XC 1997 A new
baculovirus of cultured shrimp Sci China Ser C 40 630ndash635
Chen LL Leu JH 1 Huang CJ Chou CM Chen SM Wang CH
Lo CF Kou GH 2002 Identification of a nucleocapsid protein
(VP35) gene of shrimp white spot syndrome virus and characterization
of the motif important for targeting VP35 to the nuclei of transfected
insect cells Virology 293 44ndash53
Chomczynski P Sacchi N 1987 Single-step method of RNA isolation
by acid guanidinium thiocyanatendashphenolndashchloroform extraction Anal
Biochem 262 156ndash159
Flegel TW 1997 Special topic review major viral diseases of the black
tiger prawn (Penaeus monodon) in Thailand World J Microbiol
Biotechnol 13 433ndash442
Halder SK Takemori H Hatano O Nonaka Y Wada A Okamoto
M 1998 Cloning of a membrane-spanning protein with epidermal
growth factor-like repeat motifs from adrenal glomerulosa cells
Endocrinology 139 3316ndash3328
Hofman K Stoffel W 1993 TMbasemdashA database of membrane
spanning protein segments Biol Chem Hoppe-Seyler 374 166
Hofmann K Bucher P Falquet L Bairoch A 1999 The PROSITE
database its status in 1999 Nucleic Acids Res 27 215ndash219
Huang CH Zhang XB Lin QS Xu X Hew CL 2002a Character-
ization of a novel envelope protein (VP281) of shrimp white spot
syndrome virus by mass spectrometry J Gen Virol 83 2385ndash2392
Huang CH Zhang XB Lin QS Xu X Hu ZH Hew CL 2002b
Proteomic analysis of shrimp white spot syndrome viral proteins and
characterization of a novel envelope protein VP466 Mol Cell
Proteomics 1 223ndash231
Jordens R Thompson A Amons R Koning F 1999 Human dendritic
cells shed a functional soluble form of the mannose receptor Int
Immunol 11 1775ndash1780
Krezel AM Wagner G Seymour-Ulmer J Lazarus RA 1994
Structure of the RGD protein decorsin conserved motif and distinct
function in leech proteins that affect blood clotting Science 264
1944ndash1947
Laemmli UK 1970 Cleavage of structural proteins during the assembly
of the head of bacteriophage T4 Nature 227 680ndash685
Li Q Pan D Zhang J Yang F 2004 Identification of the thymidylate
synthase within the genome of white spot syndrome virus J Gen Virol
85 2035ndash2044
Lo CF Ho CH Peng SE Chen CH Hsu HC Chiu YL Chang
CF Liu KF Su MS Wang CH Kou GH 1996a White spot
syndrome baculovirus (WSBV) detected in cultured and captured
shrimp crabs and other arthropods Dis Aquat Org 27 215ndash226
Lo CF Leu JH Ho CH Chen CH Peng SE Chen YT Chou
CM Yeh PY Huang CJ Chou HY Wang CH Kou GH
1996b Detection of baculovirus associated with white spot syndrome
(WSBV) in penaeid shrimps using polymerase chain reaction Dis
Aquat Org 25 133ndash141
Mandl CW Guirakhoo F Holzmann H Heinz FX Kunz C 1989
Antigenic structure of the flavivirus envelope E protein at the molecular
level using tick-borne encephalitis virus as a model J Virol 63
564ndash571
Marıa-Isabel V German R David C Luis S Mariano E 1998
The vaccinia virus 14-kilodalton (A27L) fusion protein forms a triple
coiled-coil structure and interacts with the 21-kilodalton (A17L)
virus membrane protein through a C-terminal a-helix J Virol 72
10126ndash10137
Martignoni ME Iwai PJ Rohrmann GF 1980 Serum neutralization
of nucleopolyhedrosis viruses (baculovirus subgroup A) pathogenic for
Orgyia pseudotsugata J Invert Pathol 36 12ndash20
Plow EF Haas TA Zhang L Loftus J Smith JW 2000 Ligand
binding to integrins J Biol Chem 275 21785ndash21788
Ruoslahti E 1996 RGD and other recognition sequences for integrins
Annu Rev Cell Biol 12 697ndash715
Ruoslahti E Pierschbacher MD 1986 ArgndashGlyndashAsp a versatile cell
recognition signal Cell 44 517ndash518
Schofield DJ Glamann J Emerson U Purchell RH 2000 Identi-
fication by phage display and characterization of two neutralizing
chimpanzee monoclonal antibodies to the hepatitis E virus capsid
protein J Virol 74 5548ndash5555
Senn H Klaus W 1993 The nuclear magnetic resonance solution
structure of flavoridin an antagonist of the platelet GPIIbndash IIIa receptor
J Mol Biol 232 907ndash925
Shevchenko A Wilm M Vorm O Mann M 1996 Mass spectrometric
sequencing of protein silver-stained polyacrylamide gels Anal Chem
68 850ndash858
Stevens TJ Arkin IT 2000 Turning an opinion inside-out Rees and
Eisenbergrsquos commentary (Proteins 2000 38121ndash122) on FAre mem-
brane proteins lsquolsquoinside-outrsquorsquo proteins_ (Proteins 1999 36 135ndash143)
Proteins 40 463ndash464
Tsai JM Wang HC Leu JH Hsiao HH Wang AH Kou GH
Lo CF 2004 Genomic and proteomic analysis of thirty-nine
structural proteins of shrimp white spot syndrome virus J Virol 78
11360ndash11370
van Hulten MC Witteveldt J Peters S Kloosterboer N Tarchini
R Fiers M Sandbrink H Lankhorst RK Vlak JM 2001a
The white spot syndrome virus DNA genome sequence Virology
286 7ndash22
van Hulten MC Witteveldt J Snippe M Vlak JM 2001b White spot
syndrome virus envelope protein VP28 is involved in the systemic
infection of shrimp Virology 285 228ndash233
Verdaguer N Mateu MG Andreu D Giralt E Domingo E Fita
I 1995 Structure of the major antigenic loop of foot-and-mouth
disease virus complexed with a neutralizing antibody direct involve-
ment of the ArgndashGlyndashAsp motif in the interaction EMBO J 14
1690ndash1696
Volkman LE Goldsmith PA 1985 Mechanism of neutralization of
L Li et al Virology 340 (2005) 125ndash132132
budded Autographa californica nuclear polyhedrosis virus by a
monoclonal antibody inhibition of entry by adsorptive endocytosis
Virology 143 185ndash195
Wu WL Wang L Zhang XB 2005 Identification of white spot
syndrome virus (WSSV) envelope proteins involved in shrimp
infection Virology 332 578ndash583
Xie XX Yang F 2005 Interaction of white spot syndrome virus VP26
protein with actin Virology 336 93ndash99
Xie XX Li HY Xu LM Yang F 2005 A simple and efficient
method for purification of intact White Spot Syndrome Virus (WSSV)
viral particles Virus Res 108 63ndash67
Yang F He J Lin X Li Q Pan D Zhang X Xu X 2001 Complete
genome sequence of the shrimp white spot bacilliform virus J Virol 75
11811ndash11820
Yingmanee B Pamela JH Farideh G Glyn S 2001 Argininendash
glycinendashaspartic acid motif is critical for human parechovirus 1 entry
J Virol 75 10000ndash10004
Yoganandhan K Syed MS Narayanan RB Sahul HAS 2004
Production of polyclonal antiserum against recombinant VP28 protein
and its application for the detection of white spot syndrome virus in
crustaceans J Fish Dis 27 517ndash522
Zhang X Huang CH Xu X Hew CL 2002a Transcription and
identification of an envelope protein gene (p22) from shrimp white spot
syndrome virus J Gen Virol 83 471ndash477
Zhang X Huang CH Xu X Hew CL 2002b Identification and
localization of a prawn white spot syndrome virus gene that encodes an
envelope protein J Gen Virol 83 1069ndash1074
Zhang X Huang C Tang X Zhuang Y Choy LH 2004 Identification
of structural proteins from shrimp white spot syndrome virus (WSSV)
by 2-DE-MS Proteins Struct Funct Bioinform 55 229ndash235

L Li et al Virology 340 (2005) 125ndash132132
budded Autographa californica nuclear polyhedrosis virus by a
monoclonal antibody inhibition of entry by adsorptive endocytosis
Virology 143 185ndash195
Wu WL Wang L Zhang XB 2005 Identification of white spot
syndrome virus (WSSV) envelope proteins involved in shrimp
infection Virology 332 578ndash583
Xie XX Yang F 2005 Interaction of white spot syndrome virus VP26
protein with actin Virology 336 93ndash99
Xie XX Li HY Xu LM Yang F 2005 A simple and efficient
method for purification of intact White Spot Syndrome Virus (WSSV)
viral particles Virus Res 108 63ndash67
Yang F He J Lin X Li Q Pan D Zhang X Xu X 2001 Complete
genome sequence of the shrimp white spot bacilliform virus J Virol 75
11811ndash11820
Yingmanee B Pamela JH Farideh G Glyn S 2001 Argininendash
glycinendashaspartic acid motif is critical for human parechovirus 1 entry
J Virol 75 10000ndash10004
Yoganandhan K Syed MS Narayanan RB Sahul HAS 2004
Production of polyclonal antiserum against recombinant VP28 protein
and its application for the detection of white spot syndrome virus in
crustaceans J Fish Dis 27 517ndash522
Zhang X Huang CH Xu X Hew CL 2002a Transcription and
identification of an envelope protein gene (p22) from shrimp white spot
syndrome virus J Gen Virol 83 471ndash477
Zhang X Huang CH Xu X Hew CL 2002b Identification and
localization of a prawn white spot syndrome virus gene that encodes an
envelope protein J Gen Virol 83 1069ndash1074
Zhang X Huang C Tang X Zhuang Y Choy LH 2004 Identification
of structural proteins from shrimp white spot syndrome virus (WSSV)
by 2-DE-MS Proteins Struct Funct Bioinform 55 229ndash235




















