
I resti faunistici dall'insediamento romano di Pantani-Le Gore (Torrita di Siena-SI)
16
Rassegna di Archeologia 23B/2007-2008 Paolo Boscato* – Cynthia Mascione** I RESTI FAUNISTICI DALL’INSEDIAMENTO ROMANO DI PANTANI-LE GORE (TORRITA DI SIENA-SI) PREMESSA STORICO-ARCHEOLOGICA Lo scavo del sito di Pantani-Le Gore, situato sul margine occidentale della Valdichiana nei pressi di Torrita di Siena (SI), è stato condotto da chi scrive negli anni 1994-1999 su incarico del Comune, as- segnatario della concessione e principale sostenitore finanziario della ricerca, in accordo con la SBAT e in collaborazione con il gruppo archeologico locale Circolo Fra’ Iacopo da Torrita, che aveva segnalato già dal 1978 alle autorità competenti l’emergere di notevoli quantità di reperti archeologici, ascrivibili ad un arco cronologico piuttosto ampio, tra la fine del I secolo a.C. e il V d.C. 1 . Lo scavo stratigrafico realizzato su tre aree estese e contigue ha interes- sato una parte limitata dell’insediamento (Fig. 1), mentre un ulteriore settore è stato sottoposto ad indagini geofisiche (magnetometria e resistività), utili a tracciare, insieme alla realizzazione di trincee di verifica della consistenza delle anomalie registrate, la topografia generale del sito. Il primo nucleo insediativo è formato da un edificio caratterizzato da un’ampia corte interna e da un porticato sul lato meridionale, dove con tutta probabilità era l’ingresso principale. Dai pochi mate- riali restituiti dalle attività correlabili alla prima fase di vita dell’edificio (Periodo Ia), la sua fondazione sembra ascrivile ai decenni finali del I secolo a.C., mentre permangono incertezze rispetto alla funzione originaria della struttura. La planimetria, per quan- to restituito dall’indagine, presenta caratteristiche generiche che si adattano a più tipi di edifici e in particolare, data la sua ubicazione in un’area rurale, ad una villa rustica o ad una mansio 2 . La seconda ipotesi deriva dalla possibile identificazione del sito con Manliana, stazione di posta collocata dalla Tabula Peutingeriana tra Ad Novas-Acquaviva di Montepulciano 3 e Ad Mensulas-Sinalunga 4 , lungo un diverticolo della via Cassia che, distaccandosi dal tracciato principale immediatamente a nord di Ad Novas, conduceva a Saena Iulia 5 . L’indagine condotta non ha evidenziato elementi dirimenti ad una definitiva identificazione con la statio, indiziata dunque per il momento dalla corrispondenza della distanza di 8 miglia da Ad Novas e dall’apparente permanere dell’antico nome nella toponomastica attuale (via e poggio di Magliano). Infine, l’impronta commerciale che caratterizza l’insediamento fin dal I secolo d.C. e che, con vicende alterne, perdura fino alla metà del VI secolo, denuncia con evidenza la sua contiguità con la viabilità. Nel I secolo d.C. (Periodo Ib) l’insediamento si estende nel settore orientale con case, forse botteghe e probabilmente officine, dedite almeno in parte alla lavorazione del metallo 6 , mentre nel settore meridionale è allestito un vasto piazzale (Attività 86 e 110), formato da una massicciata piuttosto fine e compatta, che nel corso dei secoli vedrà nu- merose fasi di assestamento e rialzamento, realizzate progressivamente in modo sempre più grossolano, con rifiuti e macerie prodotte dalla manutenzione e ristrutturazione degli edifici circostanti o dalla loro demolizione. La maggior parte dei reperti proviene da queste attività, con un’alta percentuale di resi- dualità, mentre nelle aree edificate sono scarsissime le Unità Stratigrafiche diagnostiche in correlazione * Università degli Studi di Siena, Dipartimento di Scienze Ambientali “G. Sarfatti”, UR Ecologia Preistorica. ** Università degli Studi di Siena, Dipartimento di Archeologia e Storia delle Arti. 1 CASINI, 1992a. 2 MEZZOLANI, 1992; CHEVALLIER, 1998, pp. 284-291; CORSI, 2000; BASSO, 2007, pp. 66-68, dove si sottolinea la generale difficoltà di identificazione architettonica delle mansiones. 3 MINETTI, 1997. 4 PAOLUCCI, 1996. 5 Per la ricostruzione dei possibili tracciati: CASINI, 1992b. 6 Ferro e bronzo: PAPI, 1995; GLIOZZO-MASCIONE-MEMMI, 1999.
Transcript of I resti faunistici dall'insediamento romano di Pantani-Le Gore (Torrita di Siena-SI)

Rassegna di Archeologia 23B/2007-2008
Paolo Boscato* – Cynthia Mascione**
I RESTI FAUNISTICI DALL’INSEDIAMENTO ROMANO DI PANTANI-LE GORE (TORRITA DI SIENA-SI)
PREMESSA STORICO-ARCHEOLOGICA
Lo scavo del sito di Pantani-Le Gore, situato sul margine occidentale della Valdichiana nei pressi di Torrita di Siena (SI), è stato condotto da chi scrive negli anni 1994-1999 su incarico del Comune, as-segnatario della concessione e principale sostenitore finanziario della ricerca, in accordo con la SBAT e in collaborazione con il gruppo archeologico locale Circolo Fra’ Iacopo da Torrita, che aveva segnalato già dal 1978 alle autorità competenti l’emergere di notevoli quantità di reperti archeologici, ascrivibili ad un arco cronologico piuttosto ampio, tra la fine del I secolo a.C. e il V d.C.1. Lo scavo stratigrafico realizzato su tre aree estese e contigue ha interes-sato una parte limitata dell’insediamento (Fig. 1), mentre un ulteriore settore è stato sottoposto ad indagini geofisiche (magnetometria e resistività), utili a tracciare, insieme alla realizzazione di trincee di verifica della consistenza delle anomalie registrate, la topografia generale del sito.
Il primo nucleo insediativo è formato da un edificio caratterizzato da un’ampia corte interna e da un porticato sul lato meridionale, dove con tutta probabilità era l’ingresso principale. Dai pochi mate-riali restituiti dalle attività correlabili alla prima fase di vita dell’edificio (Periodo Ia), la sua fondazione sembra ascrivile ai decenni finali del I secolo a.C., mentre permangono incertezze rispetto alla funzione originaria della struttura. La planimetria, per quan-to restituito dall’indagine, presenta caratteristiche generiche che si adattano a più tipi di edifici e in particolare, data la sua ubicazione in un’area rurale,
ad una villa rustica o ad una mansio2. La seconda ipotesi deriva dalla possibile identificazione del sito con Manliana, stazione di posta collocata dalla Tabula Peutingeriana tra Ad Novas-Acquaviva di Montepulciano3 e Ad Mensulas-Sinalunga4, lungo un diverticolo della via Cassia che, distaccandosi dal tracciato principale immediatamente a nord di Ad Novas, conduceva a Saena Iulia5. L’indagine condotta non ha evidenziato elementi dirimenti ad una definitiva identificazione con la statio, indiziata dunque per il momento dalla corrispondenza della distanza di 8 miglia da Ad Novas e dall’apparente permanere dell’antico nome nella toponomastica attuale (via e poggio di Magliano). Infine, l’impronta commerciale che caratterizza l’insediamento fin dal I secolo d.C. e che, con vicende alterne, perdura fino alla metà del VI secolo, denuncia con evidenza la sua contiguità con la viabilità.
Nel I secolo d.C. (Periodo Ib) l’insediamento si estende nel settore orientale con case, forse botteghe e probabilmente officine, dedite almeno in parte alla lavorazione del metallo6, mentre nel settore meridionale è allestito un vasto piazzale (Attività 86 e 110), formato da una massicciata piuttosto fine e compatta, che nel corso dei secoli vedrà nu-merose fasi di assestamento e rialzamento, realizzate progressivamente in modo sempre più grossolano, con rifiuti e macerie prodotte dalla manutenzione e ristrutturazione degli edifici circostanti o dalla loro demolizione. La maggior parte dei reperti proviene da queste attività, con un’alta percentuale di resi-dualità, mentre nelle aree edificate sono scarsissime le Unità Stratigrafiche diagnostiche in correlazione
* Università degli Studi di Siena, Dipartimento di Scienze Ambientali “G. Sarfatti”, UR Ecologia Preistorica.** Università degli Studi di Siena, Dipartimento di Archeologia e Storia delle Arti.1 CASINI, 1992a.2 MEZZOLANI, 1992; CHEVALLIER, 1998, pp. 284-291; CORSI, 2000; BASSO, 2007, pp. 66-68, dove si sottolinea la generale
difficoltà di identificazione architettonica delle mansiones.3 MINETTI, 1997.4 PAOLUCCI, 1996.5 Per la ricostruzione dei possibili tracciati: CASINI, 1992b.6 Ferro e bronzo: PAPI, 1995; GLIOZZO-MASCIONE-MEMMI, 1999.

PAOLO BOSCATO – CYNTHIA MASCIONE96
Fig. 1 – Planimetria generale del sito di Pantani-Le Gore.
con le varie fasi d’uso dell’insediamento. L’identifica-zione del vasto piazzale come area di mercato deriva dall’alto numero di monete qui concentrate, insieme a strumenti per la pesatura, e dal rinvenimento di piccole buche di palo associate a concentrazioni di chiodi da carpenteria e riferibili all’allestimento, probabilmente temporaneo, di strutture leggere, come banchi per la vendita, tettoie o recinti per gli animali.
Con l’avanzare del II secolo (Periodo IIa) il cd. edificio principale è oggetto di modifiche profonde, che portano alla riconversione di parte della corte interna in ambienti, segno di un cambiamento d’uso
della struttura, forse associabile – soprattutto se nell’insediamento va riconosciuta la statio Manlia-na – con la ristrutturazione adrianea della Cassia e dunque con la costruzione di un percorso diretto verso Florentia, operazione che deve aver reso ancora più marginali i tracciati di età precedente. Forse non casualmente, è possibile associare a questo periodo una sola massicciata di ripavimentazione del piazzale (Attività 35), apparentemente poco estesa. Nell’area si accumulano per lo più terre, in parte identificabili come superfici d’uso, povere di materiali ceramici e prive di reperti numismatici, che sembrano indicare un momento di crisi e di probabile riconversione

I RESTI FAUNISTICI DALL’INSEDIAMENTO ROMANO DI PANTANI-LE GORE (TORRITA DI SIENA-SI) 97
dell’economia dell’insediamento, forse in favore di uno sviluppo dell’agricoltura e dell’allevamento, percepibile in un primo e sostanziale incremento nelle attestazioni di reperti ossei appartenenti a Bos taurus.
Le scarsissime tracce d’uso degli edifici, databili nel III secolo (Periodo IIb), sono verosimilmente segno di una progressiva rarefazione dell’abitato, tendenza che pare confermata anche dalla poca consistenza quantitativa dei materiali residui prove-nienti dai successivi rifacimenti del piazzale (Periodi IV-V), fra cui vasellame da mensa a vernice rossa, databile in base alla tipologia fra fine II e III secolo d.C. e riconosciuto come di manifattura locale7. La produzione è indiziata per il momento dalle analisi archeometriche condotte sugli impasti, che ne hanno verificato l’appartenenza ad un gruppo omogeneo, omologo peraltro ad un secondo di età tardoantica (cfr. infra). Si tratta dunque di una produzione di lunga durata, che deve essere collocata nei dintorni o forse anche all’interno dell’insediamento stesso, in un’area non toccata dallo scavo. È importante segnalare che una delle attività attribuibili a questo periodo è la massicciata 74, a conferma di una conti-nuità della vocazione commerciale del sito, probabil-mente sempre più rivolta a scambi di prodotti locali e di ridotta entità. L’abbandono dell’insediamento appare definitivo tra la fine del III e la prima metà del IV secolo, mentre un nuovo periodo di occupazione (Periodo IV) si registra tra la seconda metà del IV e il V secolo, con la ristrutturazione di parte del cd. edificio principale e delle abitazioni orientali, forse in funzione di una rinnovata vivacità del commercio, probabilmente improntato sempre più su scambi di prodotti locali: dalla ripavimentazione del piazzale (Attività 18, 73, 101), e soprattutto dal consistente uso delle massicciate (Attività 17 e 48), provengono nuovamente monete, discrete quantità di vasellame da mensa di manifattura locale8 e ossa animali, per lo più di bovini, presenti in quantità elevata e indicatori di un forte sviluppo dell’allevamento e dell’agricoltura. All’aspetto commerciale del sito è probabilmente da associare anche l’apprestamento di un’area artigianale (attività 45, 72, 88), collocata immediatamente a sud del piazzale e connotata da più punti di fuoco, fra cui una piccola fornace rettangolare, con più fasi di riutilizzo difficilmente
databili per l’assenza di materiale diagnostico (Atti-vità 24, 31), e da accumuli di scorie ferrose, residuo di attività di forgiatura9.
Nell’avanzato V secolo il settore abitativo è definitivamente abbandonato, con dinamiche non percepibili dallo scavo; risulta infatti del tutto assente una stratigrafia relativa ad episodi violenti o ai crolli progressivi delle strutture, poiché le successive fasi di riuso del piazzale, databili nei primi decenni del VI secolo (Periodo V), hanno portato ad un consistente recupero di materiale edilizio e terre, da reimpiegare nella stesura di nuove massicciate. A quest’ultimo periodo di frequentazione sono attribuibili due fasi di rialzamento del piano di calpestio del piazzale (Fase a: Attività 5, 19, 40; Fase b: Attività 10-11), che invade anche il settore meridionale del cd. edificio principale (Attività 36), privato di buona parte dei muri perimetrali. All’interno di uno degli ambienti si insedia una capanna di forma ellittica, mentre altre buche di palo con allineamenti rettilinei sono probabilmente relative alla costruzione di recinti per gli animali o di ripari, come anche nell’area dell’an-tico piazzale. La poca consistenza delle strutture e la quasi totale assenza di materiale ceramico coevo sembrano indicare una frequentazione sporadica del sito, probabilmente come area di mercato stagionale, ancora collegato allo scambio e alla vendita di pro-dotti agricoli locali, aspetto che pare permanere, in considerazione della forte presenza nelle massicciate e nelle terre di uso del piazzale (Attività 23, 33, 47) di monete di piccolo taglio e in particolare di ossa di bovini, il cui allevamento rimane predominante, in continuità con la tendenza registrata soprattutto a partire dalla seconda metà del IV secolo e che sembra connotare ancora l’economia agricola del territorio.
C.M.
I RESTI FAUNISTICI. STUDIO PRELIMINARE10
I resti di macrofauna studiati in questo lavoro comprendono la totalità dei rinvenimenti della cam-pagna di scavo del 1994 (saggio I) e una parte del-l’abbondante lotto recuperato nel 1995 (saggio II). I reperti sono relativi ai periodi Ib (I-inizi II secolo d.C.), IIa (pieno II secolo d.C.), IV (metà IV-metà V
7 APROSIO et ALII, c.s.8 Ibidem.9 GLIOZZO-MASCIONE-MEMMI, 1999.10 Ringrazio la dott.ssa Silvia Pallecchi e Lara Marelli per il lavoro di preparazione delle ossa, il dott. Alfredo Riedel per la lettura critica
del manoscritto e il dott. Frank Salvadori per il suo contributo nei dati bibliografici.

PAOLO BOSCATO – CYNTHIA MASCIONE98
secolo d.C.), Va e Vb (fine V, prima metà VI secolo d.C.). La quantità maggiore di reperti ossei è stata rinvenuta nei periodi IV e Va. La consistenza e il tipo di campione (1060 reperti determinati distribuiti nei cinque periodi considerati) forniscono un valido, seppur preliminare, inquadramento della gestione faunistica di questo sito. Il futuro studio sull’intero materiale a disposizione sarà fondamentale per l’ap-profondimento delle conoscenze su alcuni ambiti di indagine e sui periodi meno rappresentati.
Il materiale: stato di conservazione e contesti di rinvenimento
Le ossa, in gran parte relative a bovini, sono state rinvenute in un buono stato di conservazione. Il materiale, per lo più frammentato, presenta sia segni di rotture volontarie dovute ad azione antropica, che fratturazioni post-deposizionali. Sono relativamente abbondanti le tracce di macellazione.
L’assenza di setacciatura del terreno di scavo ha probabilmente determinato il mancato recupero di alcune ossa di vertebrati di piccola e media taglia. Per questo motivo, pur rimanendo certa la forte dominanza di resti di bue, è da ritenere sottostimata la presenza di ovicaprini e di suini.
Gran parte del materiale osseo del campione esa-minato è stato rinvenuto all’interno di strati d’uso e di massicciate. Nella tabella 1 figura la distribuzione dei frammenti determinati a livello tassonomico per i contesti strutturali più comuni. I valori percentuali servono indicativamente a quantificare la distri-buzione della massa totale dei frammenti ossei, in buona parte non determinabili.
Ad una prima analisi sul materiale osseo degli strati d’uso (Attività 17, P. IV; 47 P. Va) non è evi-dente un particolare stato di frammentazione. Sono presenti ossa integre anche di grosse dimensioni: metapodiali di bovini e parti fragili come porzioni di scapole. Sembra, quindi, che le ossa abbandonate su questi piani non abbiano subito forti danni da calpestio. È ipotizzabile che un substrato morbido abbia inglobato anche parzialmente le ossa in tempi brevi successivamente al loro abbandono, proteggen-dole da stress meccanici.
Nelle massicciate il materiale osseo era concentrato sulle parti sommitali (Fig. 2) (Attività 35, Periodo IIa; 18 e 101, Periodo IV; 5 e 40, Periodo Va; 10, Periodo Vb). Anche in questi contesti, oltre alla consueta fram-mentazione, non sono evidenti sulle ossa forti abra-sioni. È probabile che il materiale osseo di scarto della macellazione sia stato utilizzato associato a pietrame e a frammenti di laterizi nella messa a punto di piani drenanti utili nella bonifica del substrato argilloso del
Tab. 1 – Distribuzione dei reperti determinati per contesti strutturali di rinvenimento.
ATTIVITÀ DI PROVENIENZA NR %Strati d’uso delle massicciate 374 35,3Massicciate 278 26,2Preparazione massicciate 70 6,6Crolli 170 16,0Riempimento fosse 46 4,3Altro 122 11,5NR totale 1060
Fig. 2 – Superficie di massicciata tardoantica con concen-trazione di ossa.
sito. L’interpretazione della funzione delle massicciate nel contesto generale e l’analisi microscopica dei segni sulle ossa potranno aggiungere maggiori informazioni su questa questione.
Frammenti ossei di dimensioni inferiori alle medie del materiale di questo sito provengono dal-l’attività 33 (Periodo Va), strato formato da concotto e laterizi sbriciolati, interpretato come possibile crollo dell’alzato del muro che divide l’area delle massicciate e quella artigianale nel Periodo IV (att. 76). In questo strato sono stati rinvenuti numerosi denti sciolti di bovini.
Le faune
Il campione faunistico oggetto di questo lavoro esprime una netta prevalenza di resti di bovini (Tabb. 2a, 2b, 3). Nei periodi Ib, IV, Va le percentuali sul numero totale dei reperti determinati vedono per questo grosso ungulato valori compresi tra l’88,1% e il 91,5%. Suini e ovicaprini sono segnalati in quanti-tà decisamente inferiori. I primi variano dal 5,1% del periodo IV al 6,6% del periodo Va, mentre i secondi, nella somma con i resti attribuiti specificamente alla capra e alla pecora, vanno dal 3,1% al 5,6%.
I RESTI FAUNISTICI DALL’INSEDIAMENTO ROMANO DI PANTANI-LE GORE (TORRITA DI SIENA-SI) 99
Tab. 2a – Numero dei reperti determinati divisi per periodo e per attività.
Tab. 2b – Numero dei reperti determinati divisi per periodo e per attività.
Periodi Ib IIa IVCronologia I d.C. II d.C. IV-V d.C.Attività 110 35 75 108 14 17 18 45 48 72 73 88 101Equus asinus 1Equidae sp. 1 1Bos taurus 7 29 31 23 9 24 14 4 77 8 6 91 35Ovis vel Capra 2 1 2 1 2 2Capra hircus 1 1 1Sus domesticus 1 4 1 8 6Tot. NR 7 32 32 28 12 25 15 5 77 10 6 103 42
Periodi IV-V Va VbCronologia IV-metà VI Prima metà VI d.C.Attività 24 31 49b 5 23 32 33 40 47 99 106 10Equus ferus 1Equidae sp. 1Bos taurus 50 80 29 84 36 109 31 99 33 7 31Ovis vel Capra 5 3 1 7 1 2 1 1 5 1Ovis aries 1 1Capra hircus 2 1Sus domesticus 3 4 2 18 2 5 2 3 2Canis familiaris 1Cervus elaphus 2Tot. NR 58 89 32 111 38 2 116 32 100 38 16 35
Periodi Ib IIa IV IV-V Va VbCronologia I d.C. II d.C. IV-V d.C. Prima metà VI d.C.
NR % NR % NR % NR % NR % NR %Equus ferus 1 0,2Equus asinus 1 0,3Equidae sp. 2 0,6 1 0,2Bos taurus 7 83 90,2 267 91,5 159 88,8 399 88,1 31Ovis vel Capra 3 3,3 7 2,4 9 5,0 17 3,7 1Capra hircus 1 1,1 2 0,7 3 0,7Ovis aries 1 0,6 1Sus domesticus 5 5,4 15 5,1 9 5,0 30 6,6 2Canis familiaris 1 0,6Cervus elaphus 2 0,4Tot. NR 7 92 294 179 453 35
Tab. 3 – Numero dei reperti determinati (NR) con relative percentuali per periodo.
Completano questo quadro di specie domestiche sporadiche presenze di equidi (cavallo e asino) e un solo reperto di cane.
L’unica testimonianza di attività di caccia è data per il momento da due denti di cervo.
BoviniI 948 reperti attribuiti a bovini rappresentano,
seppure in quantità diverse, l’intero apparato schele-
trico (Tab. 4). Gli elementi più comuni sono i denti isolati che, sommati a parti mandibolari, mascellari e craniali arrivano al 52% del totale.
Complessivamente, in rapporto al numero dei frammenti di metapodiali, risultano scarse le ossa relative al carpo e alle parti distali degli arti (falangi e sesamoidi). La ragione di tale carenza potrebbe in parte dipendere dalla mancata setacciatura del sedimento e quindi, durante il recupero “a vista”,

PAOLO BOSCATO – CYNTHIA MASCIONE100
Periodi Ib IIa IV IV-V Va VbCavicchia 2 1Mascellare 2 2 4 1Mandibola 2 10 42 11 21 4Neurocranio 4 14 7Denti decidui 4Denti permanenti sup. 7 40 25 80 4Denti permanenti inf. 13 60 24 98 6Denti indeterminati 1 2Ioide 2 7 3Vertebre 2 5 8 8 9 2Costole 4 4 3Scapola 5 9 4 13 1Omero 2 7 3 12 2Radio 1 4 9 5 9 2Ulna 1 5 2Radio-ulna 2 3Carpali 1 4 8Metacarpo 2 10 9 13 28 1Coxale 4 3 14Femore 2 6 2 6Tibia 1 3 2 6 1Malleolare 1 1Tarsali 7 11 5 11 1Metatarso 1 12 17 4 15 1Metapodiali 3 2 12 2Falange 1 2 12 10 13Falange 2 7 4 9 1Falange 3 1 6Sesamoidi 1Tot. NR 8 83 267 159 399 31
Tab. 4 – Bue: numero dei reperti determinati divisi per elementi scheletrici.
Fig. 3 – Bue: localizzazione delle tracce di macellazione.
nella non individuazione di alcuni elementi di mi-nori dimensioni. Questo squilibrio numerico può pure dipendere dalle modalità di macellazione e dal diverso utilizzo delle parti scheletriche. Le falangi con lo zoccolo potrebbero essere state scartate du-rante una prima fase di macellazione e avere subito un percorso di abbandono diverso dalle altre parti scheletriche legate a grosse masse muscolari o con-tenenti maggiori quantità di midollo.
Nella frequenza degli elementi scheletrici, un altro dato da specificare riguarda le costole e le ver-tebre. Nella tabella 4 figurano solo i pochi elementi rinvenuti integri o con caratteri morfologici di sicura attribuzione specifica. In realtà, tra il materiale non determinato, esistono discrete quantità di fram-menti vertebrali e costali riferibili genericamente ad ungulati di grosse dimensioni. Come esempio, nel materiale della campagna di scavo 1994, nel pe-riodo Va, sono state determinate 143 ossa di bovini e, separatamente, 19 frammenti di vertebre e 64 di costole di grossi ungulati; nel periodo IV, sempre
nello stesso anno di scavo, a 77 ossa determinate di bovini vanno aggiunti 16 frammenti di vertebre e 46 di costole. Come è evidente, risulta non tra-scurabile la presenza di questi elementi scheletrici, sebbene sovrastimata vista la facilità di attribuzione generica anche di piccoli frammenti. Le dimensioni riportano questi reperti a costole e vertebre di bue o cavallo-asino e la sporadicità delle presenze di questi ultimi fa supporre trattarsi principalmente di frammenti di bovini.Tracce di macellazione e patologie
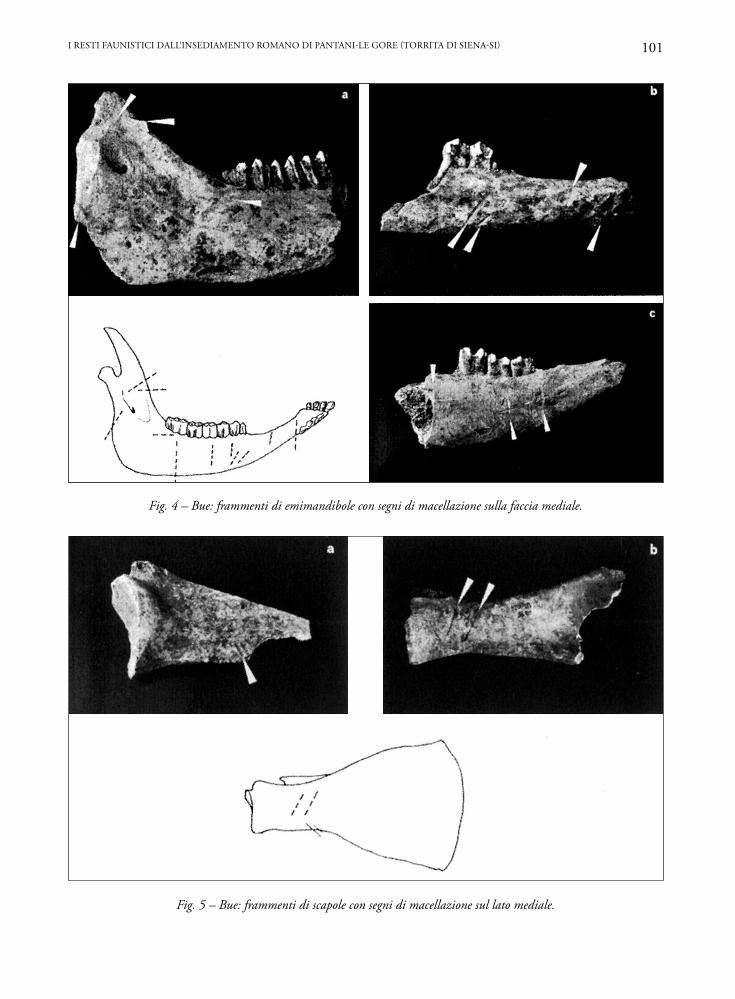
Nella Fig. 3 è schematizzata la localizzazione delle tracce di macellazione osservate negli elementi scheletrici dei bovini. Si tratta di strie o di segni più profondi da fendente lasciati da strumenti da taglio durante lo smembramento e spellamento degli animali e nella scarnificazione o rottura delle varie ossa.Cranio e mandibola
Tracce lasciate da strumenti da taglio sullo zigo-matico e sulla parte posteriore mandibolare, tra il condilo e il processo coronoideo, sono probabilmen-te legate alla separazione delle due emimandibole dal cranio. In un caso, l’emimandibola è stata isolata con fendenti di sbieco sulla branca mandibolare (Fig. 4a). Anteriormente, in alcuni casi le emimandibole sono state separate con fendenti trasversali nell’area com-presa tra il foro mentoniero e il terzo premolare (Fig. 4b) o spezzate all’altezza del terzo molare (Fig. 4c).
Sottili strie sono state notate su un osso ioide, probabilmente lasciate durante la disarticolazione della lingua. Sono evidenti anche segni di separa-zione della cavicchia dal frontale.Quarti anteriori
Vari frammenti di scapola portano tracce lasciate nell’isolamento delle masse muscolari lungo la fossa
I RESTI FAUNISTICI DALL’INSEDIAMENTO ROMANO DI PANTANI-LE GORE (TORRITA DI SIENA-SI) 101
Fig. 4 – Bue: frammenti di emimandibole con segni di macellazione sulla faccia mediale.
Fig. 5 – Bue: frammenti di scapole con segni di macellazione sul lato mediale.
PAOLO BOSCATO – CYNTHIA MASCIONE102
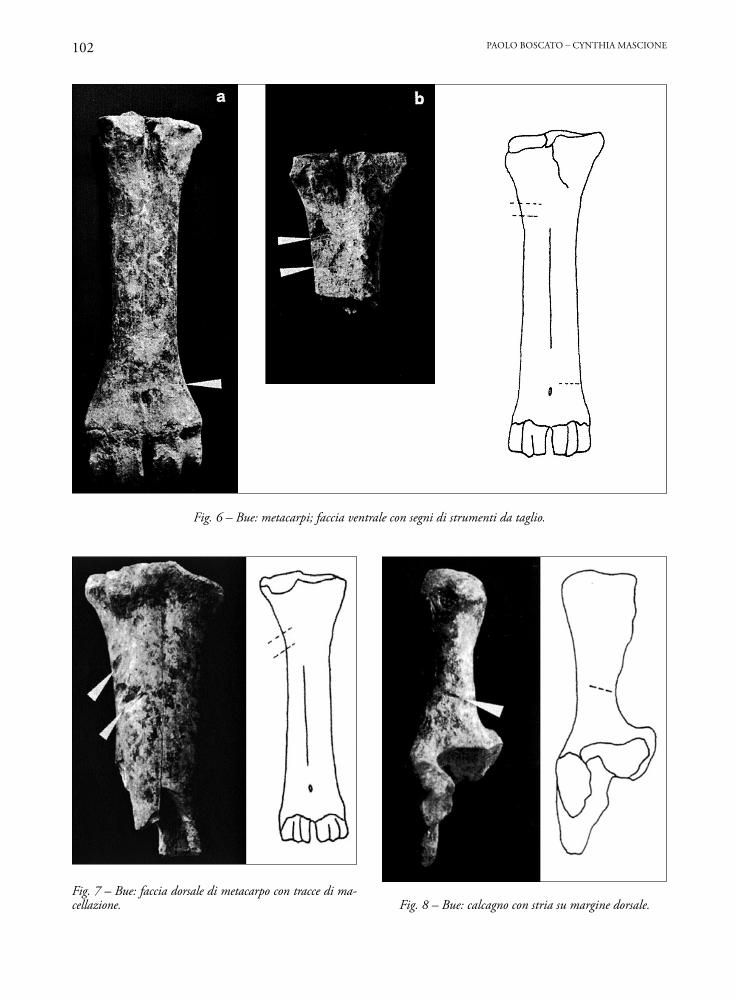
Fig. 6 – Bue: metacarpi; faccia ventrale con segni di strumenti da taglio.
Fig. 7 – Bue: faccia dorsale di metacarpo con tracce di ma-cellazione. Fig. 8 – Bue: calcagno con stria su margine dorsale.
I RESTI FAUNISTICI DALL’INSEDIAMENTO ROMANO DI PANTANI-LE GORE (TORRITA DI SIENA-SI) 103
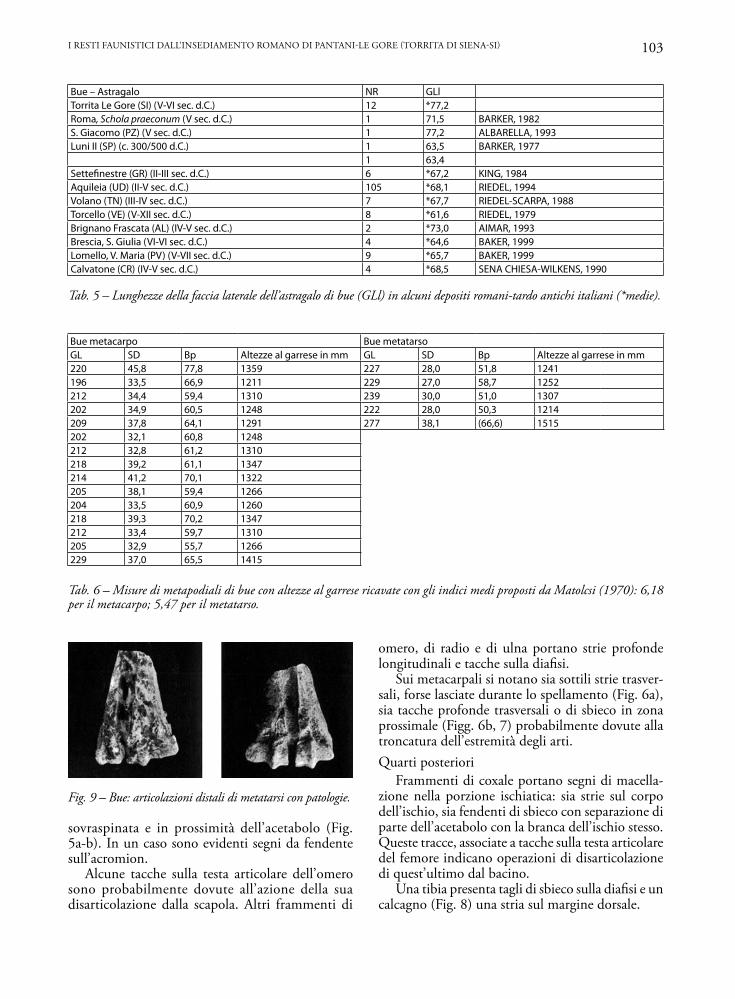
Bue – Astragalo NR GLlTorrita Le Gore (SI) (V-VI sec. d.C.) 12 *77,2Roma, Schola praeconum (V sec. d.C.) 1 71,5 BARKER, 1982S. Giacomo (PZ) (V sec. d.C.) 1 77,2 ALBARELLA, 1993Luni II (SP) (c. 300/500 d.C.) 1 63,5 BARKER, 1977
1 63,4Settefinestre (GR) (II-III sec. d.C.) 6 *67,2 KING, 1984Aquileia (UD) (II-V sec. d.C.) 105 *68,1 RIEDEL, 1994Volano (TN) (III-IV sec. d.C.) 7 *67,7 RIEDEL-SCARPA, 1988Torcello (VE) (V-XII sec. d.C.) 8 *61,6 RIEDEL, 1979Brignano Frascata (AL) (IV-V sec. d.C.) 2 *73,0 AIMAR, 1993Brescia, S. Giulia (VI-VI sec. d.C.) 4 *64,6 BAKER, 1999Lomello, V. Maria (PV) (V-VII sec. d.C.) 9 *65,7 BAKER, 1999Calvatone (CR) (IV-V sec. d.C.) 4 *68,5 SENA CHIESA-WILKENS, 1990
Bue metacarpo Bue metatarsoGL SD Bp Altezze al garrese in mm GL SD Bp Altezze al garrese in mm220 45,8 77,8 1359 227 28,0 51,8 1241196 33,5 66,9 1211 229 27,0 58,7 1252212 34,4 59,4 1310 239 30,0 51,0 1307202 34,9 60,5 1248 222 28,0 50,3 1214209 37,8 64,1 1291 277 38,1 (66,6) 1515202 32,1 60,8 1248212 32,8 61,2 1310218 39,2 61,1 1347214 41,2 70,1 1322205 38,1 59,4 1266204 33,5 60,9 1260218 39,3 70,2 1347212 33,4 59,7 1310205 32,9 55,7 1266229 37,0 65,5 1415
Tab. 6 – Misure di metapodiali di bue con altezze al garrese ricavate con gli indici medi proposti da Matolcsi (1970): 6,18 per il metacarpo; 5,47 per il metatarso.
Tab. 5 – Lunghezze della faccia laterale dell’astragalo di bue (GLl) in alcuni depositi romani-tardo antichi italiani (*medie).
Fig. 9 – Bue: articolazioni distali di metatarsi con patologie.
sovraspinata e in prossimità dell’acetabolo (Fig. 5a-b). In un caso sono evidenti segni da fendente sull’acromion.
Alcune tacche sulla testa articolare dell’omero sono probabilmente dovute all’azione della sua disarticolazione dalla scapola. Altri frammenti di
omero, di radio e di ulna portano strie profonde longitudinali e tacche sulla diafisi.
Sui metacarpali si notano sia sottili strie trasver-sali, forse lasciate durante lo spellamento (Fig. 6a), sia tacche profonde trasversali o di sbieco in zona prossimale (Figg. 6b, 7) probabilmente dovute alla troncatura dell’estremità degli arti.Quarti posteriori
Frammenti di coxale portano segni di macella-zione nella porzione ischiatica: sia strie sul corpo dell’ischio, sia fendenti di sbieco con separazione di parte dell’acetabolo con la branca dell’ischio stesso. Queste tracce, associate a tacche sulla testa articolare del femore indicano operazioni di disarticolazione di quest’ultimo dal bacino.
Una tibia presenta tagli di sbieco sulla diafisi e un calcagno (Fig. 8) una stria sul margine dorsale.
PAOLO BOSCATO – CYNTHIA MASCIONE104
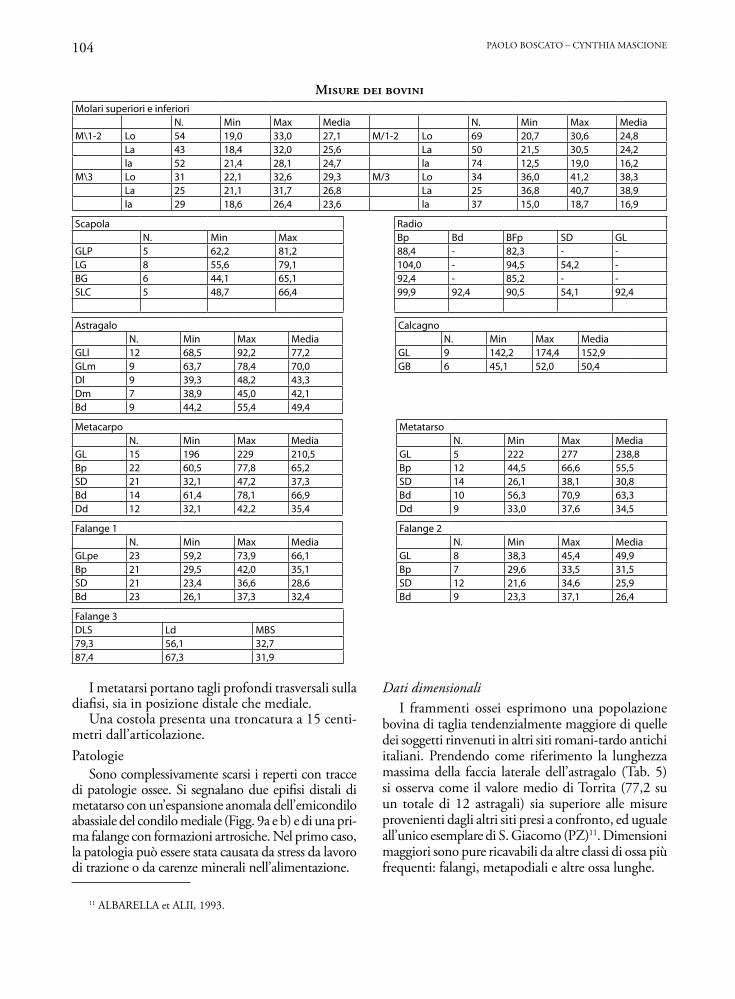
Molari superiori e inferioriN. Min Max Media N. Min Max Media
M\1-2 Lo 54 19,0 33,0 27,1 M/1-2 Lo 69 20,7 30,6 24,8La 43 18,4 32,0 25,6 La 50 21,5 30,5 24,2la 52 21,4 28,1 24,7 la 74 12,5 19,0 16,2
M\3 Lo 31 22,1 32,6 29,3 M/3 Lo 34 36,0 41,2 38,3La 25 21,1 31,7 26,8 La 25 36,8 40,7 38,9la 29 18,6 26,4 23,6 la 37 15,0 18,7 16,9
Scapola RadioN. Min Max Bp Bd BFp SD GL
GLP 5 62,2 81,2 88,4 - 82,3 - -LG 8 55,6 79,1 104,0 - 94,5 54,2 -BG 6 44,1 65,1 92,4 - 85,2 - -SLC 5 48,7 66,4 99,9 92,4 90,5 54,1 92,4
Astragalo CalcagnoN. Min Max Media N. Min Max Media
GLl 12 68,5 92,2 77,2 GL 9 142,2 174,4 152,9GLm 9 63,7 78,4 70,0 GB 6 45,1 52,0 50,4Dl 9 39,3 48,2 43,3Dm 7 38,9 45,0 42,1Bd 9 44,2 55,4 49,4
Metacarpo MetatarsoN. Min Max Media N. Min Max Media
GL 15 196 229 210,5 GL 5 222 277 238,8Bp 22 60,5 77,8 65,2 Bp 12 44,5 66,6 55,5SD 21 32,1 47,2 37,3 SD 14 26,1 38,1 30,8Bd 14 61,4 78,1 66,9 Bd 10 56,3 70,9 63,3Dd 12 32,1 42,2 35,4 Dd 9 33,0 37,6 34,5
Falange 1 Falange 2N. Min Max Media N. Min Max Media
GLpe 23 59,2 73,9 66,1 GL 8 38,3 45,4 49,9Bp 21 29,5 42,0 35,1 Bp 7 29,6 33,5 31,5SD 21 23,4 36,6 28,6 SD 12 21,6 34,6 25,9Bd 23 26,1 37,3 32,4 Bd 9 23,3 37,1 26,4
Falange 3DLS Ld MBS79,3 56,1 32,787,4 67,3 31,9
I metatarsi portano tagli profondi trasversali sulla diafisi, sia in posizione distale che mediale.
Una costola presenta una troncatura a 15 centi-metri dall’articolazione.Patologie
Sono complessivamente scarsi i reperti con tracce di patologie ossee. Si segnalano due epifisi distali di metatarso con un’espansione anomala dell’emicondilo abassiale del condilo mediale (Figg. 9a e b) e di una pri-ma falange con formazioni artrosiche. Nel primo caso, la patologia può essere stata causata da stress da lavoro di trazione o da carenze minerali nell’alimentazione.
Dati dimensionaliI frammenti ossei esprimono una popolazione
bovina di taglia tendenzialmente maggiore di quelle dei soggetti rinvenuti in altri siti romani-tardo antichi italiani. Prendendo come riferimento la lunghezza massima della faccia laterale dell’astragalo (Tab. 5) si osserva come il valore medio di Torrita (77,2 su un totale di 12 astragali) sia superiore alle misure provenienti dagli altri siti presi a confronto, ed uguale all’unico esemplare di S. Giacomo (PZ)11. Dimensioni maggiori sono pure ricavabili da altre classi di ossa più frequenti: falangi, metapodiali e altre ossa lunghe.
MISURE DEI BOVINI
11 ALBARELLA et ALII, 1993.
I RESTI FAUNISTICI DALL’INSEDIAMENTO ROMANO DI PANTANI-LE GORE (TORRITA DI SIENA-SI) 105
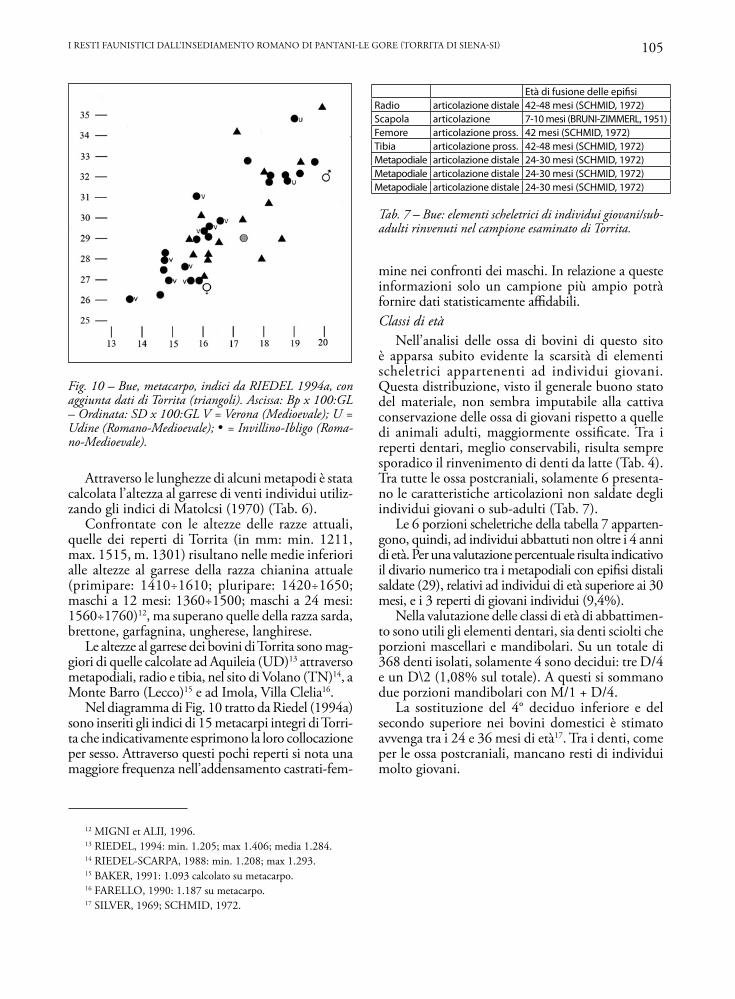
Fig. 10 – Bue, metacarpo, indici da RIEDEL 1994a, con aggiunta dati di Torrita (triangoli). Ascissa: Bp x 100:GL – Ordinata: SD x 100:GL V = Verona (Medioevale); U = Udine (Romano-Medioevale); = Invillino-Ibligo (Roma-no-Medioevale).
12 MIGNI et ALII, 1996.13 RIEDEL, 1994: min. 1.205; max 1.406; media 1.284.14 RIEDEL-SCARPA, 1988: min. 1.208; max 1.293.15 BAKER, 1991: 1.093 calcolato su metacarpo.16 FARELLO, 1990: 1.187 su metacarpo.17 SILVER, 1969; SCHMID, 1972.
Età di fusione delle epifisiRadio articolazione distale 42-48 mesi (SCHMID, 1972)Scapola articolazione 7-10 mesi (BRUNI-ZIMMERL, 1951)Femore articolazione pross. 42 mesi (SCHMID, 1972)Tibia articolazione pross. 42-48 mesi (SCHMID, 1972)Metapodiale articolazione distale 24-30 mesi (SCHMID, 1972)Metapodiale articolazione distale 24-30 mesi (SCHMID, 1972)Metapodiale articolazione distale 24-30 mesi (SCHMID, 1972)
Tab. 7 – Bue: elementi scheletrici di individui giovani/sub-adulti rinvenuti nel campione esaminato di Torrita.
Attraverso le lunghezze di alcuni metapodi è stata calcolata l’altezza al garrese di venti individui utiliz-zando gli indici di Matolcsi (1970) (Tab. 6).
Confrontate con le altezze delle razze attuali, quelle dei reperti di Torrita (in mm: min. 1211, max. 1515, m. 1301) risultano nelle medie inferiori alle altezze al garrese della razza chianina attuale (primipare: 1410÷1610; pluripare: 1420÷1650; maschi a 12 mesi: 1360÷1500; maschi a 24 mesi: 1560÷1760)12, ma superano quelle della razza sarda, brettone, garfagnina, ungherese, langhirese.
Le altezze al garrese dei bovini di Torrita sono mag-giori di quelle calcolate ad Aquileia (UD)13 attraverso metapodiali, radio e tibia, nel sito di Volano (TN)14, a Monte Barro (Lecco)15 e ad Imola, Villa Clelia16.
Nel diagramma di Fig. 10 tratto da Riedel (1994a) sono inseriti gli indici di 15 metacarpi integri di Torri-ta che indicativamente esprimono la loro collocazione per sesso. Attraverso questi pochi reperti si nota una maggiore frequenza nell’addensamento castrati-fem-
mine nei confronti dei maschi. In relazione a queste informazioni solo un campione più ampio potrà fornire dati statisticamente affidabili.Classi di età
Nell’analisi delle ossa di bovini di questo sito è apparsa subito evidente la scarsità di elementi scheletrici appartenenti ad individui giovani. Questa distribuzione, visto il generale buono stato del materiale, non sembra imputabile alla cattiva conservazione delle ossa di giovani rispetto a quelle di animali adulti, maggiormente ossificate. Tra i reperti dentari, meglio conservabili, risulta sempre sporadico il rinvenimento di denti da latte (Tab. 4). Tra tutte le ossa postcraniali, solamente 6 presenta-no le caratteristiche articolazioni non saldate degli individui giovani o sub-adulti (Tab. 7).
Le 6 porzioni scheletriche della tabella 7 apparten-gono, quindi, ad individui abbattuti non oltre i 4 anni di età. Per una valutazione percentuale risulta indicativo il divario numerico tra i metapodiali con epifisi distali saldate (29), relativi ad individui di età superiore ai 30 mesi, e i 3 reperti di giovani individui (9,4%).
Nella valutazione delle classi di età di abbattimen-to sono utili gli elementi dentari, sia denti sciolti che porzioni mascellari e mandibolari. Su un totale di 368 denti isolati, solamente 4 sono decidui: tre D/4 e un D\2 (1,08% sul totale). A questi si sommano due porzioni mandibolari con M/1 + D/4.
La sostituzione del 4° deciduo inferiore e del secondo superiore nei bovini domestici è stimato avvenga tra i 24 e 36 mesi di età17. Tra i denti, come per le ossa postcraniali, mancano resti di individui molto giovani.
PAOLO BOSCATO – CYNTHIA MASCIONE106
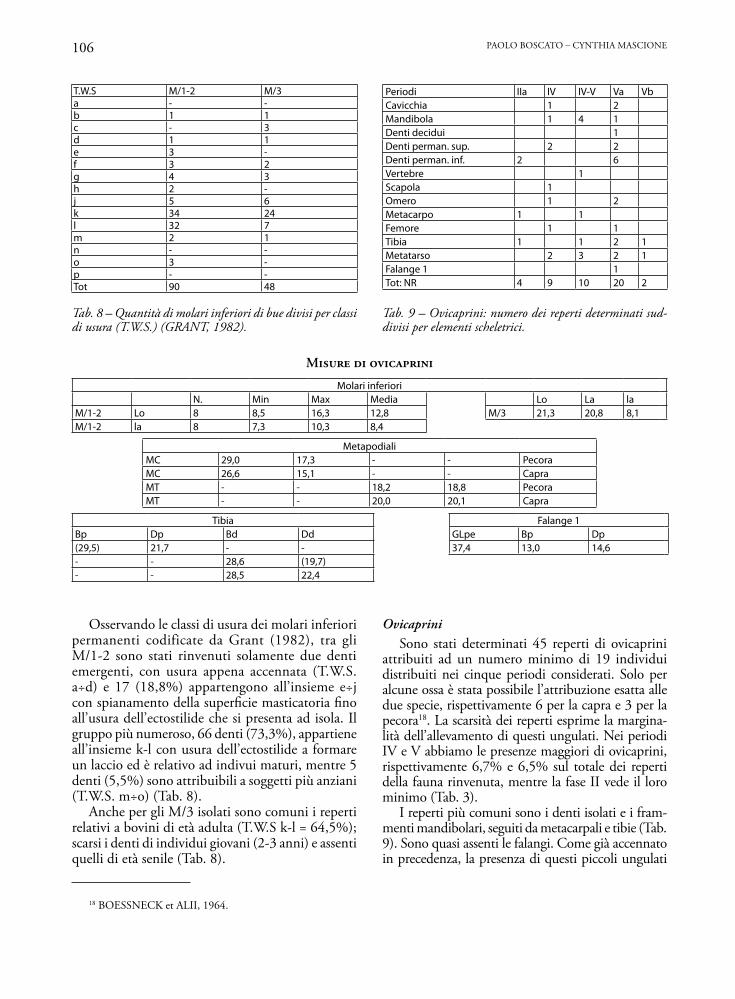
T.W.S M/1-2 M/3a - -b 1 1c - 3d 1 1e 3 -f 3 2g 4 3h 2 -j 5 6k 34 24l 32 7m 2 1n - -o 3 -p - -Tot 90 48
Tab. 8 – Quantità di molari inferiori di bue divisi per classi di usura (T.W.S.) (GRANT, 1982).
Periodi IIa IV IV-V Va VbCavicchia 1 2Mandibola 1 4 1Denti decidui 1Denti perman. sup. 2 2Denti perman. inf. 2 6Vertebre 1Scapola 1Omero 1 2Metacarpo 1 1Femore 1 1Tibia 1 1 2 1Metatarso 2 3 2 1Falange 1 1Tot: NR 4 9 10 20 2
Tab. 9 – Ovicaprini: numero dei reperti determinati sud-divisi per elementi scheletrici.
MISURE DI OVICAPRINI
Molari inferioriN. Min Max Media Lo La la
M/1-2 Lo 8 8,5 16,3 12,8 M/3 21,3 20,8 8,1M/1-2 la 8 7,3 10,3 8,4
MetapodialiMC 29,0 17,3 - - PecoraMC 26,6 15,1 - - CapraMT - - 18,2 18,8 PecoraMT - - 20,0 20,1 Capra
Tibia Falange 1Bp Dp Bd Dd GLpe Bp Dp(29,5) 21,7 - - 37,4 13,0 14,6- - 28,6 (19,7)- - 28,5 22,4
Osservando le classi di usura dei molari inferiori permanenti codificate da Grant (1982), tra gli M/1-2 sono stati rinvenuti solamente due denti emergenti, con usura appena accennata (T.W.S. a÷d) e 17 (18,8%) appartengono all’insieme e÷j con spianamento della superficie masticatoria fino all’usura dell’ectostilide che si presenta ad isola. Il gruppo più numeroso, 66 denti (73,3%), appartiene all’insieme k-l con usura dell’ectostilide a formare un laccio ed è relativo ad indivui maturi, mentre 5 denti (5,5%) sono attribuibili a soggetti più anziani (T.W.S. m÷o) (Tab. 8).
Anche per gli M/3 isolati sono comuni i reperti relativi a bovini di età adulta (T.W.S k-l = 64,5%); scarsi i denti di individui giovani (2-3 anni) e assenti quelli di età senile (Tab. 8).
OvicapriniSono stati determinati 45 reperti di ovicaprini
attribuiti ad un numero minimo di 19 individui distribuiti nei cinque periodi considerati. Solo per alcune ossa è stata possibile l’attribuzione esatta alle due specie, rispettivamente 6 per la capra e 3 per la pecora18. La scarsità dei reperti esprime la margina-lità dell’allevamento di questi ungulati. Nei periodi IV e V abbiamo le presenze maggiori di ovicaprini, rispettivamente 6,7% e 6,5% sul totale dei reperti della fauna rinvenuta, mentre la fase II vede il loro minimo (Tab. 3).
I reperti più comuni sono i denti isolati e i fram-menti mandibolari, seguiti da metacarpali e tibie (Tab. 9). Sono quasi assenti le falangi. Come già accennato in precedenza, la presenza di questi piccoli ungulati
18 BOESSNECK et ALII, 1964.
I RESTI FAUNISTICI DALL’INSEDIAMENTO ROMANO DI PANTANI-LE GORE (TORRITA DI SIENA-SI) 107
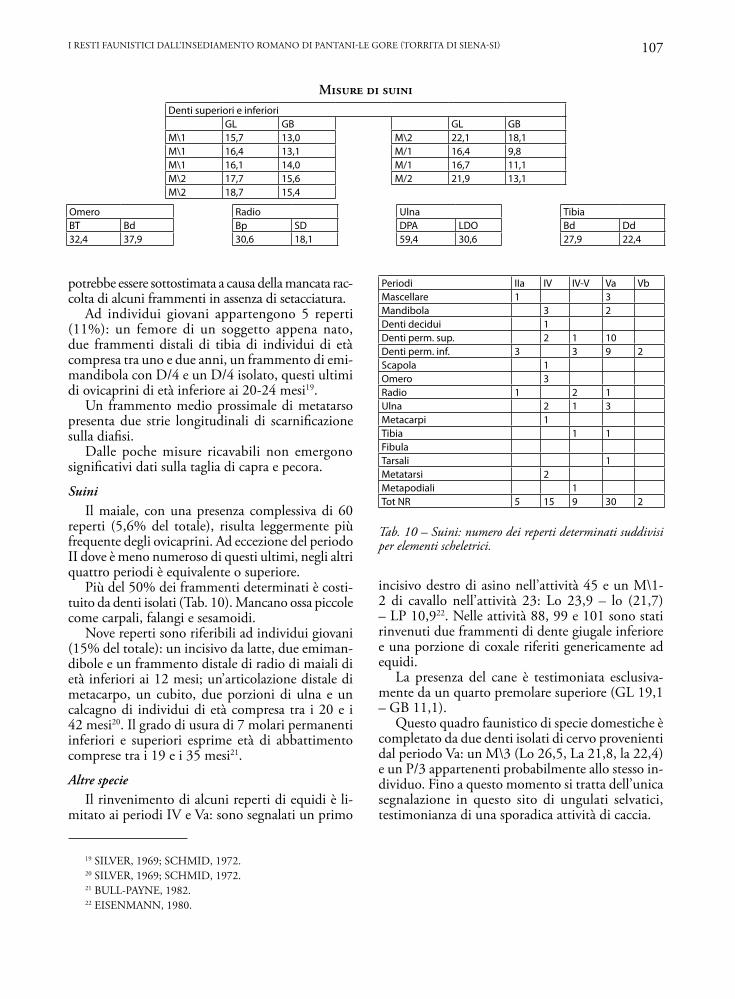
Periodi IIa IV IV-V Va VbMascellare 1 3Mandibola 3 2Denti decidui 1Denti perm. sup. 2 1 10Denti perm. inf. 3 3 9 2Scapola 1Omero 3Radio 1 2 1Ulna 2 1 3Metacarpi 1Tibia 1 1FibulaTarsali 1Metatarsi 2Metapodiali 1Tot NR 5 15 9 30 2
Tab. 10 – Suini: numero dei reperti determinati suddivisi per elementi scheletrici.
Denti superiori e inferioriGL GB GL GB
M\1 15,7 13,0 M\2 22,1 18,1M\1 16,4 13,1 M/1 16,4 9,8M\1 16,1 14,0 M/1 16,7 11,1M\2 17,7 15,6 M/2 21,9 13,1M\2 18,7 15,4
Omero Radio Ulna TibiaBT Bd Bp SD DPA LDO Bd Dd32,4 37,9 30,6 18,1 59,4 30,6 27,9 22,4
potrebbe essere sottostimata a causa della mancata rac-colta di alcuni frammenti in assenza di setacciatura.
Ad individui giovani appartengono 5 reperti (11%): un femore di un soggetto appena nato, due frammenti distali di tibia di individui di età compresa tra uno e due anni, un frammento di emi-mandibola con D/4 e un D/4 isolato, questi ultimi di ovicaprini di età inferiore ai 20-24 mesi19.
Un frammento medio prossimale di metatarso presenta due strie longitudinali di scarnificazione sulla diafisi.
Dalle poche misure ricavabili non emergono significativi dati sulla taglia di capra e pecora.
SuiniIl maiale, con una presenza complessiva di 60
reperti (5,6% del totale), risulta leggermente più frequente degli ovicaprini. Ad eccezione del periodo II dove è meno numeroso di questi ultimi, negli altri quattro periodi è equivalente o superiore.
Più del 50% dei frammenti determinati è costi-tuito da denti isolati (Tab. 10). Mancano ossa piccole come carpali, falangi e sesamoidi.
Nove reperti sono riferibili ad individui giovani (15% del totale): un incisivo da latte, due emiman-dibole e un frammento distale di radio di maiali di età inferiori ai 12 mesi; un’articolazione distale di metacarpo, un cubito, due porzioni di ulna e un calcagno di individui di età compresa tra i 20 e i 42 mesi20. Il grado di usura di 7 molari permanenti inferiori e superiori esprime età di abbattimento comprese tra i 19 e i 35 mesi21.
Altre specieIl rinvenimento di alcuni reperti di equidi è li-
mitato ai periodi IV e Va: sono segnalati un primo
incisivo destro di asino nell’attività 45 e un M\1-2 di cavallo nell’attività 23: Lo 23,9 – lo (21,7) – LP 10,922. Nelle attività 88, 99 e 101 sono stati rinvenuti due frammenti di dente giugale inferiore e una porzione di coxale riferiti genericamente ad equidi.
La presenza del cane è testimoniata esclusiva-mente da un quarto premolare superiore (GL 19,1 – GB 11,1).
Questo quadro faunistico di specie domestiche è completato da due denti isolati di cervo provenienti dal periodo Va: un M\3 (Lo 26,5, La 21,8, la 22,4) e un P/3 appartenenti probabilmente allo stesso in-dividuo. Fino a questo momento si tratta dell’unica segnalazione in questo sito di ungulati selvatici, testimonianza di una sporadica attività di caccia.
MISURE DI SUINI
19 SILVER, 1969; SCHMID, 1972.20 SILVER, 1969; SCHMID, 1972.21 BULL-PAYNE, 1982.22 EISENMANN, 1980.
PAOLO BOSCATO – CYNTHIA MASCIONE108
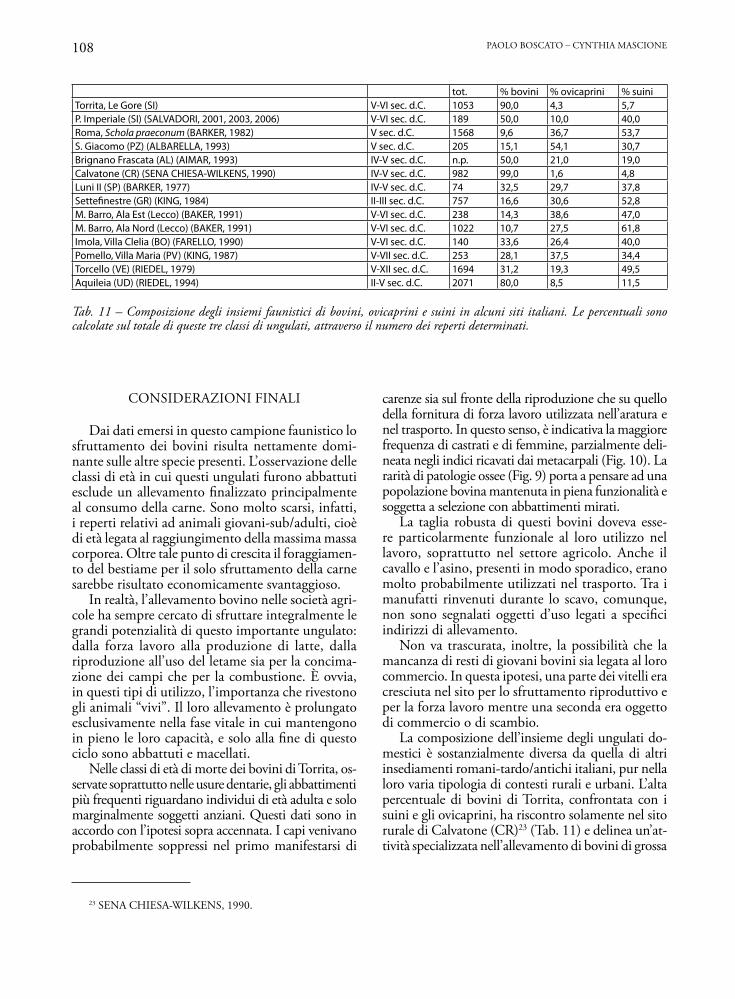
tot. % bovini % ovicaprini % suiniTorrita, Le Gore (SI) V-VI sec. d.C. 1053 90,0 4,3 5,7P. Imperiale (SI) (SALVADORI, 2001, 2003, 2006) V-VI sec. d.C. 189 50,0 10,0 40,0Roma, Schola praeconum (BARKER, 1982) V sec. d.C. 1568 9,6 36,7 53,7S. Giacomo (PZ) (ALBARELLA, 1993) V sec. d.C. 205 15,1 54,1 30,7Brignano Frascata (AL) (AIMAR, 1993) IV-V sec. d.C. n.p. 50,0 21,0 19,0Calvatone (CR) (SENA CHIESA-WILKENS, 1990) IV-V sec. d.C. 982 99,0 1,6 4,8Luni II (SP) (BARKER, 1977) IV-V sec. d.C. 74 32,5 29,7 37,8Settefinestre (GR) (KING, 1984) II-III sec. d.C. 757 16,6 30,6 52,8M. Barro, Ala Est (Lecco) (BAKER, 1991) V-VI sec. d.C. 238 14,3 38,6 47,0M. Barro, Ala Nord (Lecco) (BAKER, 1991) V-VI sec. d.C. 1022 10,7 27,5 61,8Imola, Villa Clelia (BO) (FARELLO, 1990) V-VI sec. d.C. 140 33,6 26,4 40,0Pomello, Villa Maria (PV) (KING, 1987) V-VII sec. d.C. 253 28,1 37,5 34,4Torcello (VE) (RIEDEL, 1979) V-XII sec. d.C. 1694 31,2 19,3 49,5Aquileia (UD) (RIEDEL, 1994) II-V sec. d.C. 2071 80,0 8,5 11,5
CONSIDERAZIONI FINALI
Dai dati emersi in questo campione faunistico lo sfruttamento dei bovini risulta nettamente domi-nante sulle altre specie presenti. L’osservazione delle classi di età in cui questi ungulati furono abbattuti esclude un allevamento finalizzato principalmente al consumo della carne. Sono molto scarsi, infatti, i reperti relativi ad animali giovani-sub/adulti, cioè di età legata al raggiungimento della massima massa corporea. Oltre tale punto di crescita il foraggiamen-to del bestiame per il solo sfruttamento della carne sarebbe risultato economicamente svantaggioso.
In realtà, l’allevamento bovino nelle società agri-cole ha sempre cercato di sfruttare integralmente le grandi potenzialità di questo importante ungulato: dalla forza lavoro alla produzione di latte, dalla riproduzione all’uso del letame sia per la concima-zione dei campi che per la combustione. È ovvia, in questi tipi di utilizzo, l’importanza che rivestono gli animali “vivi”. Il loro allevamento è prolungato esclusivamente nella fase vitale in cui mantengono in pieno le loro capacità, e solo alla fine di questo ciclo sono abbattuti e macellati.
Nelle classi di età di morte dei bovini di Torrita, os-servate soprattutto nelle usure dentarie, gli abbattimenti più frequenti riguardano individui di età adulta e solo marginalmente soggetti anziani. Questi dati sono in accordo con l’ipotesi sopra accennata. I capi venivano probabilmente soppressi nel primo manifestarsi di
carenze sia sul fronte della riproduzione che su quello della fornitura di forza lavoro utilizzata nell’aratura e nel trasporto. In questo senso, è indicativa la maggiore frequenza di castrati e di femmine, parzialmente deli-neata negli indici ricavati dai metacarpali (Fig. 10). La rarità di patologie ossee (Fig. 9) porta a pensare ad una popolazione bovina mantenuta in piena funzionalità e soggetta a selezione con abbattimenti mirati.
La taglia robusta di questi bovini doveva esse-re particolarmente funzionale al loro utilizzo nel lavoro, soprattutto nel settore agricolo. Anche il cavallo e l’asino, presenti in modo sporadico, erano molto probabilmente utilizzati nel trasporto. Tra i manufatti rinvenuti durante lo scavo, comunque, non sono segnalati oggetti d’uso legati a specifici indirizzi di allevamento.
Non va trascurata, inoltre, la possibilità che la mancanza di resti di giovani bovini sia legata al loro commercio. In questa ipotesi, una parte dei vitelli era cresciuta nel sito per lo sfruttamento riproduttivo e per la forza lavoro mentre una seconda era oggetto di commercio o di scambio.
La composizione dell’insieme degli ungulati do-mestici è sostanzialmente diversa da quella di altri insediamenti romani-tardo/antichi italiani, pur nella loro varia tipologia di contesti rurali e urbani. L’alta percentuale di bovini di Torrita, confrontata con i suini e gli ovicaprini, ha riscontro solamente nel sito rurale di Calvatone (CR)23 (Tab. 11) e delinea un’at-tività specializzata nell’allevamento di bovini di grossa
Tab. 11 – Composizione degli insiemi faunistici di bovini, ovicaprini e suini in alcuni siti italiani. Le percentuali sono calcolate sul totale di queste tre classi di ungulati, attraverso il numero dei reperti determinati.
23 SENA CHIESA-WILKENS, 1990.

I RESTI FAUNISTICI DALL’INSEDIAMENTO ROMANO DI PANTANI-LE GORE (TORRITA DI SIENA-SI) 109
taglia. Altri valori elevati sono registrati ad Aquileia24 con l’80% di bovini. Si tratta, però, di un insedia-mento urbano con una situazione di importazione di derrate alimentari, incluse le carni, difficilmente confrontabile con Torrita che con ogni probabilità è identificabile come un sito di produzione.
La macellazione doveva avvenire all’interno del-l’insediamento e parte delle ossa scartate, molte delle quali portano tracce di strumenti da taglio, anziché essere smaltite in rifiutaie erano probabilmente utilizzate assieme a pietrisco e laterizi nell’appron-tamento di piani di frequentazione, funzionali per bonificare il substrato argilloso del sito.
Minor importanza nell’economia di questo in-sediamento riveste l’allevamento dei suini (5,6%) e degli ovicaprini (4,2% sul totale dei reperti determi-nati). Mentre i primi erano allevati essenzialmente per la carne, la bassa percentuale di individui giovani tra gli ovicaprini (11%) fa ipotizzare un allevamento di tipo misto, indirizzato alla produzione di latte, lana e secondariamente carne. Tra gli ovicaprini le determinazioni specifiche vedono una prevalenza delle capre sulle pecore.
P.B.Abbreviazioni usate nel testo (da von den DRIESCH, 1976)Bd: massima larghezza distaleBfp: massima larghezza faccia articolare prossimaleBG: larghezza cavità glenoideaBp: massima larghezza prossimaleBT: massima larghezza trocleaDd: massima profondità distaleDl: massima profondità lateraleDLS: massima lunghezza diagonale della suolaDm: massima profondità medialeGB: massima larghezzaGL: massima lunghezzaGLl: massima lunghezza lateraleGLm: massima lunghezza medialeGLP: massima lunghezza processo articolareGlpe: massima lunghezza abassialela: larghezza alveolareLa: lunghezza alveolareLd: lunghezza faccia dorsaleLG: lunghezza cavità glenoideaLo: lunghezza occlusalelo: larghezza occlusaleLP: lunghezza occlusale protoconoM\: molare superioreM/: molare inferioreMBS: larghezza parte mediana della suolaMC: metacarpoMT: metatarsoP: premolareSLC: lunghezza minima del collo della scapolaSD: larghezza minima della diafisi
RIFERIMENTI BIBLIOGRAFICI
AIMAR A. (1993) – Brignano Frascata, Frazione San Gior-gio. in G. PANTÒ (a cura di) Archeologia nella Valle del Curone, Torino, pp. 181-186.
ALBARELLA U. (1993) – The fauna, in U. ALBARELLA, V. CEGLIA, P. ROBERTS, S. Giacomo degli Schiavoni (Molise): an early fifth century AD deposit of pottery and animal bones from Central Adriatic Italy, P.B.S.R., 61, pp. 203-230.
APROSIO M., MASCIONE C., PECCHIONI E., AZ-ZARO E. (c.s.) – Le ceramiche da mensa tardoantiche dall’insediamento di Pantani-Le Gore (Torrita di Siena-SI), in MENCHELLI S., PASQUINUCCI M. (a cura di), Late Roman Coarse Wares, cooking wares and amphorae in the mediterranean: archaeology and archaeometry, 3rd International Conference (Parma-Pisa 2008).
BARKER G. (1977) – L’economia del bestiame a Luni, in A. FROVA (a cura di) Scavi di Luni, II, Roma, pp. 725-735.
BARKER G. (1982) – The animal bones, in D. WHIT-EOUSE, G. BARKER, R. REECE, D. REESE (eds.) The Schola Praeconum I: the coins, pottery, lamps and fauna, P.B.S.R., 50, pp. 53-101.
BAKER P. (1999) – The vertebrate remains from the longobard and 9th-10th C. occupation at S. Giulia, Brescia, in G.P. BROGIOLO (a cura di), S. Giulia di Brescia, gli scavi dal 1980 al 1992. Reperti preromani, romani e altomedievali, Firenze, pp. 425-449.
BAKER P. (1991) – Fauna, in G.P. BROGIOLO, L. CA-STELLETTI (a cura di), Archeologia a Monte Barro, I, Il grande edificio e le torri, Lecco, pp. 153-167.
BASSO P. (2007) – Strade romane: storia e archeologia, Roma.
BRUNI A.C., ZIMMERL U. (1951) – Anatomia degli animali domestici, vol. 1, Milano.
BOESSNECK J., MÜLLER H., TEICHERT M. (1964) – Osteologische Unterscheidungsmerkale zwischen Schaf (Ovis aries LINNE’) und Ziege (Capra hircus LINNE’), Kühn-Archiv, 78 Band, heft 1/2, Berlin.
BULL G., PAYNE S. (1982) – Tooth eruption and epiphy-sial fusion in pigs and wild boar, in B. WILSON, C. GRIGSON, S. PAYNE (eds.), Ageing and Sexing Animal Bones from Archaeological Sites, BAR, British Series 109, Oxford, pp. 55-72.
CASINI S. (1992a) – Pantani (Le Gore), in Pucci G. (a cura di), La fornace di Umbricio Cordo. L’officina di un cerami-sta romano e il territorio di Torrita di Siena nell’antichità, Firenze, pp. 30-32.
CASINI S. (1992b) – Torrita di Siena e la viabilità romana, in G. PUCCI (a cura di), La fornace di Umbricio Cordo. L’officina di un ceramista romano e il territorio di Torrita di Siena nell’antichità, Firenze, pp. 23-28.
CHEVALLIER R. (1998) – Voies romaines, Paris.CORSI C. (2000) – Le strutture di servizio del Cursus Publicus
in Italia: ricerche topografiche ed evidenze archeologiche, BAR, Int. Series, 875, Oxford.
DRIESCH von den A. (1976) – A guide to the measurement of animal bones from archaeological sites, Peabody Museum, bull. 1, Harvard Univ., Cambridge, Massachusetts.
24 RIEDEL, 1994.

PAOLO BOSCATO – CYNTHIA MASCIONE110
EISENMANN V. (1980) – Les Chevaux (Equus sensu lato) fossiles et actuels: crânes et jugales supérieures, Cahiers Paléontologie, p. 186.
FARELLO P. (1990) – La fauna, in AA.VV., Contesti tar-do-antichi e altomedievali dal sito di Villa Clelia (Imola, Bologna), Arch. Med., 17, pp. 208-215.
GLIOZZO E., MASCIONE C., MEMMI I. (1999) – Stu-dio archeometallurgico delle scorie di Le Gore, Abstract del 2° Forum Italiano di Scienze della Terra (Belluria 1999), Plinius. Supplemento Italiano all’European Journal of Mineralogy, 22, pp. 206-208.
GRANT A. (1982) – The use of tooth wear as a guide to the age of domestic ungulates, in B. WILSON, C. GRIGSON, S. PAYNE (eds.), Ageing and sexing animal bones from archaeological sites, BAR, Brithis Series 109, Oxford, pp. 91-108.
KING A. (1984) – I resti animali, in A. RICCI (ed.), Set-tefinestre: una villa schiavistica nell’Etruria Romana, II, Bologna, pp. 278-299.
KING A. (1987) – Ossa animali, in H. BLAKE, C. MAC-CABRUNI (a cura di), Dallo scavo a Villa Maria di Lomello (Pavia), 1984: la buca tardo-antica 203, Arch. Med., 14, pp. 175-177.
MATOLCSI J. (1970) – Historische Erforschung der Korper-grobe des Rindes auf Grund von ungarischen Knochenmate-rial, Zeitschrift Fur Tierzuchtung und Zuchtungbiologie, 87 (2), pp. 89-137.
MEZZOLANI A. (1992) – Appunti sulle mansiones in base ai dati archeologici, in L. QUILICI, S. QUILICI GIGLI, Tecnica stradale romana, Roma, pp. 105-113.
MIGNI L., RIDOLFI M., FILIPPINI F., SENSI B. (1996) – La razza chianina, Perugia.
MINETTI A. (1997) (a cura di) – Etruschi e romani ad Acquaviva di Montepulciano, Montepulciano.
PAOLUCCI G. (1996) – Sinalunga e Bettolle. Due centri etruschi della Valdichiana, Sinalunga.
PAPI E. (1995) – Bollo su lamina di piombo dalla mansio manliana, Zeitschrift für Papirologie und Epigraphik, 106, pp. 255-258.
RIEDEL A. (1979) – La fauna degli scavi di Torcello (1961-1962) – The fauna of the Torcello excavations (1961-1962), Atti Mus. Civ. St. Nat., Trieste, XXXI, f. 2, pp. 75-154.
RIEDEL A. (1985) – Die fauna von Altino (Venetien) im Verhältnis zu den Faunen Nordostitaliens und der Alpen-länder, Razprave IV, Zbornik Ivana Rakovca sazu XXVI, Ljubljana, pp. 131-146.
RIEDEL A. (1986) – Risultati di ricerche archeozoologiche eseguite nella regione fra la costa adriatica e il crinale alpino (dal Neolitico al recente Medio Evo), Padusa, 22, pp. 1-220.
RIEDEL A. (1994) – Roman animal bones from the forum of Aquileia, in M. VERZAR-BASS (a cura di), Scavi ad Aquileia I. L’area ad est del foro. Rapporto degli scavi 1989-91, Roma, pp. 583-591.
RIEDEL A. (1994a) – The animal remains of medioeval Verona: an Archaeozoological and palaeoeconomical study, Mem. Mus. Civ. St. Nat.Verona, 3, p. 142.
RIEDEL A., SCARPA G. (1988) – Resti animali di un com-plesso produttivo di età Tardoromana a Volano, Annali dei Musei Civici di Rovereto, pp. 37-54.
SALVADORI F. (2001) – I reperti osteologici animali di Poggio Imperiale (SI) e della Rocca di Campiglia Marittima (LI): un contributo per la storia economica ed alimentare me-dievale e la sperimentazione di un sistema di archiviazione ed elaborazione dei dati faunistici. Tesi di laurea inedita, Università degli Studi di Siena, Facoltà di Lettere e Fi-losofia, a.a. 2000-2001.
SALVADORI F. (2003) – Archeozoologia e Medioevo: lo stato degli studi, in R. FIORILLO, P. PEDUTO (a cura di), III Congresso Nazionale di Archeologia Medievale (Salerno 2003), Firenze, pp. 176-181.
SALVADORI F. (2006) – Resti osteologici animali: elementi di continuità e discontinuità tra Tardoantico e Altome-dioevo, in R. FRANCOVICH, M. VALENTI (a cura di), IV Congresso Nazionale di Archeologia Medieoevale, Abbazia San Galgano (Chiusdino, Siena 2006), Firenze, pp. 520-524.
SENA CHIESA G., WILKENS B. (1990) – I resti fauni-stici di Calvatone (CR), Archeologia Medievale, 17, pp. 307-322.
SCHMID E.F. (1972) – Atlas of Animal Bones, New York.SILVER I.A. (1969) – The aging of Domestic Animals, in
BROTHWELL D., HIGGS E. (eds.), Science in Archae-ology, New York, pp. 250-268.
RIASSUNTO
L’insediamento romano di Pantani-Le Gore di Torrita di Siena ha restituito insiemi faunistici relativi al I-VI secolo d.C. e caratterizzati dalla netta prevalenza di resti di bovini. I reperti analizzati di questa specie sono relativi, seppur in quantità diverse, all’intero apparato scheletrico. Su varie ossa sono state individuate evidenti tracce di macellazione. Le misure degli elementi scheletrici esprimono una popolazione bovina di grossa taglia. L’allevamento dei bovini sembra indirizzato alla riproduzione e allo sfruttamento della forza lavoro: dall’analisi dell’età di abbattimento emergono quasi esclusivamente indi-vidui adulti. La mancanza di giovani può essere legata al loro commercio. L’allevamento di suini e di ovicaprini ha un ruolo marginale nell’economia di questo insediamento. Sono segnalate sporadiche presenze di cavallo, asino e cane.
ABSTRACT
The roman settlement of Pantani-Le Gore (Torrita di Siena, Siena-Italy) provides I-VI century a.C. faunal remains with high predominance of cattle. Analyzed bones of this taxa represent the whole skeletal system and several of them bear clear butchering marks. Bone measurements indicate a big-sized bovine population. The aim of their breeding seems to be only reproduction and working use: in fact the killing age analysis show exclusively adult animals. Perhaps the lack of young su-bjects can be correlated to their trade. The breeding of pigs, sheep and goats have a lower role in the economy of this settlement. Finally horses, donkeys and dogs are rarely testified.




















