
Humoral immune response against tumoral mucin 1 (MUC1) in breast cancer patients
7
© 2013 Wichtig Editore - eISSN 1724-6008 Int J Biol Markers ( 2013; 00): 000-000 00 - 1 SHORT COMMUNICATION 20 amino acids and contains 5 potential glycosylation sites on Ser and Thr. In normal epithelia, carbohydrate chains are longer than in cancerous glycoforms, due to the ac- tion of distinct enzymes involved in elongation reactions (4, 5). Glycan changes may be important for the induc- tion of a humoral immune response. Mainly, carbohydrate antigens expressed at the cell surface have unexpectedly proved to be potent targets for immune recognition and attack against human cancers (6, 7). The search for the specific isotypes among circulating antibodies is an in- teresting approach to elucidate the nature of the humoral immune response. A study on the IgG response would be important not only for the potential protective role against breast cancer but also for highlighting their reactiv- ity directed to a widely studied tumor-associated antigen expressed in almost all breast tumors, as MUC1. INTRODUCTION Mucin-1 (MUC1) is a high-molecular-weight trans- membrane glycoprotein whose expression in normal epi- thelia is restricted to the apical border of normal cells. In cancer, this mucin is overexpressed in a hypoglycosylated form and is present not only on the entire cell surface but also at cytoplasmic level. MUC1 is involved in immune regulation, cell signaling, inhibition of cell-cell and cell- matrix adhesion (1, 2, 3). This glycoprotein is synthesized as a large peptide and is cleaved into 2 subunits. MUC1 contains 3 domains: the extracellular domain, the transmembrane domain and a cytoplasmic tail. The extracellular region has a protein core mainly composed by a variable number of tandem repeats (VNTR). Each tandem repeat consists of a sequence of Humoral immune response against tumoral mucin 1 (MUC1) in breast cancer patients Marina T Isla Larrain 1 , Andrea G Colussi 1,2 , Sandra O Demichelis 3 , Alberto Barbera 4 , Aldo Cretón 4 , Amada Segal-Eiras 1 , María V Croce 1 1 Centre of Basic and Applied Immunological Research, Faculty of Medical Sciences, National University of La Plata, CIC/PBA La Plata - Argentina 2 Faculty of Natural Sciences and Museum, National University of La Plata - Argentina 3 Department of Biology, Kennedy University of Argentina, Buenos Aires - Argentina 4 Breast Clinic, La Plata - Argentina ABSTRACT The aim of this study was to elucidate whether the IgG humoral immune response to breast cancer cells is directed to the ab- errant mucin-1 (MUC1) associated to this type of cancer. To this aim, an adaptation of immunohistochemistry (IHC) was per- formed on samples of 45 breast cancer tissues, 12 benign disease tissues, and 31 normal tissues, incubated with matched serum samples from the same patients. Each serum sample was also incubated, with a modified immunocytochemistry (ICC), with MCF7 cells. In both techniques, serum was employed instead of the primary antibody. In the case of IHC, the reactivity with sera diminished when added after previous incubation of the tumor/tissue with an anti-MUC1 mAb; the reduction in reactiv- ity was: from 93% to 44% in breast cancer tissues, and from 100% to 67% in benign disease tissues. The reactivity of normal samples (36%) remained unchanged. In the case of ICC, the reactivity with sera decreased after incubation with anti-MUC1 mAb from 71% to 16% in breast cancer tissues, from 83% to 0% in benign disease tissues, and from 52% to 10% in normal serum samples. These results were confirmed employing siRNA MUC1 transient gene knockdown. By Western blot analysis – after immunoprecipitation (IP) of the circulating MUC1– and ELISA, the TF antigen was detected in circulating MUC1 in all breast cancer and benign samples while Tn was detected in 38% of the samples. The existence of IgG autoantibodies against aberrantly glycosylated MUC1 may have a protective role and may contribute to a better prognosis in some patients. Enhancement of this natural immune response may constitute an alternative thera- peutic strategy. Key words: Breast cancer, Humoral immune response, MUC1, Tn, TF Received: September 7, 2012; Accepted: May 30, 2013 DOI: 10.5301/JBM.5000036
Transcript of Humoral immune response against tumoral mucin 1 (MUC1) in breast cancer patients

© 2013 Wichtig Editore - eISSN 1724-6008
Int J Biol Markers (2013; 00): 000-00000 -
1
SHORT COMMUNICATION
20 amino acids and contains 5 potential glycosylation sites on Ser and Thr. In normal epithelia, carbohydrate chains are longer than in cancerous glycoforms, due to the ac-tion of distinct enzymes involved in elongation reactions (4, 5). Glycan changes may be important for the induc-tion of a humoral immune response. Mainly, carbohydrate antigens expressed at the cell surface have unexpectedly proved to be potent targets for immune recognition and attack against human cancers (6, 7). The search for the specific isotypes among circulating antibodies is an in-teresting approach to elucidate the nature of the humoral immune response. A study on the IgG response would be important not only for the potential protective role against breast cancer but also for highlighting their reactiv-ity directed to a widely studied tumor-associated antigen expressed in almost all breast tumors, as MUC1.
IntroductIon
Mucin-1 (MUC1) is a high-molecular-weight trans-membrane glycoprotein whose expression in normal epi-thelia is restricted to the apical border of normal cells. In cancer, this mucin is overexpressed in a hypoglycosylated form and is present not only on the entire cell surface but also at cytoplasmic level. MUC1 is involved in immune regulation, cell signaling, inhibition of cell-cell and cell-matrix adhesion (1, 2, 3).
This glycoprotein is synthesized as a large peptide and is cleaved into 2 subunits. MUC1 contains 3 domains: the extracellular domain, the transmembrane domain and a cytoplasmic tail. The extracellular region has a protein core mainly composed by a variable number of tandem repeats (VNTR). Each tandem repeat consists of a sequence of
Humoral immune response against tumoral mucin 1 (Muc1) in breast cancer patients
Marina t Isla Larrain1, Andrea G colussi1,2, Sandra o demichelis3, Alberto Barbera4, Aldo cretón4, Amada Segal-Eiras1, María V croce1
1 Centre of Basic and Applied Immunological Research, Faculty of Medical Sciences, National University of La Plata, CIC/PBA La Plata - Argentina
2Faculty of Natural Sciences and Museum, National University of La Plata - Argentina3Department of Biology, Kennedy University of Argentina, Buenos Aires - Argentina4Breast Clinic, La Plata - ArgentinaCentre of Basic and Applied Immunological Research, Faculty of Medical Sciences, National University of La Plata, CIC/PBA La Plata - ArgentinaCentre of Basic and Applied Immunological Research, Faculty of Medical Sciences, National University of La Plata, CIC/PBA La Plata - Argentina and Faculty of Natural Sciences and Museum, National University of La Plata - ArgentinaDepartment of Biology, Kennedy University of Argentina, Buenos Aires - ArgentinaBreast Clinic, La Plata - ArgentinaBreast Clinic, La Plata - ArgentinaCentre of Basic and Applied Immunological Research, Faculty of Medical Sciences, National University of La Plata, CIC/PBA La Plata - ArgentinaCentre of Basic and Applied Immunological Research, Faculty of Medical Sciences, National University of La Plata, CIC/PBA La Plata - Argentina
ABStrAct the aim of this study was to elucidate whether the IgG humoral immune response to breast cancer cells is directed to the ab-errant mucin-1 (Muc1) associated to this type of cancer. to this aim, an adaptation of immunohistochemistry (IHc) was per-formed on samples of 45 breast cancer tissues, 12 benign disease tissues, and 31 normal tissues, incubated with matched serum samples from the same patients. Each serum sample was also incubated, with a modified immunocytochemistry (Icc), with McF7 cells. In both techniques, serum was employed instead of the primary antibody. In the case of IHc, the reactivity with sera diminished when added after previous incubation of the tumor/tissue with an anti-Muc1 mAb; the reduction in reactiv-ity was: from 93% to 44% in breast cancer tissues, and from 100% to 67% in benign disease tissues. the reactivity of normal samples (36%) remained unchanged. In the case of Icc, the reactivity with sera decreased after incubation with anti-Muc1 mAb from 71% to 16% in breast cancer tissues, from 83% to 0% in benign disease tissues, and from 52% to 10% in normal serum samples. these results were confirmed employing sirnA Muc1 transient gene knockdown. By Western blot analysis – after immunoprecipitation (IP) of the circulating Muc1– and ELISA, the tF antigen was detected in circulating Muc1 in all breast cancer and benign samples while tn was detected in 38% of the samples.the existence of IgG autoantibodies against aberrantly glycosylated Muc1 may have a protective role and may contribute to a better prognosis in some patients. Enhancement of this natural immune response may constitute an alternative thera-peutic strategy.
Key words: Breast cancer, Humoral immune response, Muc1, tn, tF
Received: September 7, 2012; Accepted: May 30, 2013
DOI: 10.5301/JBM.5000036

© 2013 Wichtig Editore - eISSN 1724-60082
MUC1 antibodies in breast cancer
In a variety of different clinical settings, the correlation of antibodies –naturally acquired or vaccine-induced– with prognosis improvement is one of the basis for cancer vaccines primarily designed for antibody induction. It has been suggested that autoantibodies are not so effective as biomarkers for diagnosis but, rather, are more useful for prognosis (8).
The objectives of this study are: (i) to elucidate wheth-er the IgG response in serum samples of breast cancer patients, patients with a benign disease of the breast and healthy women is directed to aberrantly glycosylated tu-moral MUC1; (ii) to study the presence of tumor-associated short carbohydrate epitopes in circulating MUC1 through the detection of Tn and (Thomsen-Friedenreich) TF antigens.
MAtErIALS And MEtHodS
Patients
Fourty-five breast cancer patients (before surgery) were selected for this study, and 12 patients with breast benign disease (BBD), as well as 31 healthy women were selected as controls. BBD cases included 7 fibroadenomas (FA) and 5 usual epithelial hyperplasias (UEH). Median age of breast cancer patients was 53 years (range 29–83), while it was 41 years (range 26–68) for BBD patients, and 37 years (range 19–57) for healthy women. Breast cancer types were divided as follows: 89% not otherwise speci-fied ductal type (NOS ductal), 2% lobular, and 9% other types. Further classifications of the different breast cancers were: 8% carcinomas in situ, and 92% invasive breast car-cinomas; 42% stage I, 28% stage II, and 22% stage III. Samples from healthy women were obtained from breast reductive mammoplasty.
Specimen characteristics
Serum samples were frozen at -80°C, tumor and tissue samples were freshly collected and delivered to the labo-ratory with minimal delay. For histopathological diagno-sis and immunohistochemical analysis, a sample fraction was immediately fixed in Methacarn (methanol: chloro-form: acetic acid 60:30:10) for 2 hours and transferred into 70% ethanol until processing in paraffin.
Assay methods
The following anti-MUC1 antibodies against the VNTR epitopes were used: SM3 (IgG, which recognizes PDTRP), Human Milk Fat Globule 1 (HMFG1) and 2 (HMFG2) mAbs (both IgG and both recognizing DTR). Anti-GalNAc (Tn) and anti-Gal β-1,4-GalNAc (TF) (IgM, DAKO, Copenhagen, Denmark) were employed to study these carbohydrates on MUC1.
The MCF7 breast cancer cell line was used for the experiments employing a cell line. Cells were seeded in flasks (Gibco-BRL, Gaithersburg, Maryland, USA) and maintained in 5% CO2 atmosphere, with RPMI 1640 sup-plemented with 10% fetal bovine serum (Gen Argentina, Buenos Aires, Argentina) and 1% antibiotics.
Immunohistochemistry (IHC)
IHC was performed following standard procedures, with tumoral, BBD, or normal tissue samples incubated with the different anti-MUC1 mAbs. Antigen retrieval was performed with sodium citrate buffer pH 6.0. After incubation with the mAb, a horseradish peroxidase (HRP)-conjugated anti-mouse IgG (DAKO, Copenhagen, Denmark) was added, incubated for 1 hour and rinsed; the chromogen employed was 3,3’-diaminodiazobenzi-dine (DAB) (SIGMA, St. Louis, MO, USA) in 1% bovine serum albumin (BSA)/PBS containing 30% H2O2.
Modified IHC
An adaptation of the IHC was performed employing serum samples instead of the primary antibody as to eval-uate the humoral immune response in patients by incubat-ing with tumoral, BBD, and normal tissue samples. After rinsing, a 1/600 HRP-conjugated anti-human IgG (DAKO, Copenhagen, Denmark) was added and incubated for 1 hour and then rinsed; the chromogen employed was DAB (SIGMA, St. Louis, MO, USA) in 1% BSA/PBS con-taining 30% H2O2.
Inhibition of reactivity with an anti-MUC1 mAb
Tumoral, BBD, and normal tissues were incubated with an anti-MUC1 mAb (dilutions: 1/1 SM3, 1/500 HMFG1, and 1/1 HMFG2) before the sera-addition step. Further steps were performed as described above.
The intensity of the staining was scored as negative (-), low (+), moderate (++), or strong (+++). The pattern of reac-tion was classified as linear (membrane reaction), cytoplas-mic or mixed (both cytoplasmic and membrane reactions) (9); the positive reaction of the lumen content was identi-fied as cellular debris or secretion; apical and non-apical reactions were also considered.
Immunocytochemistry (ICC)
ICC was performed on MCF7 cells grown on glass coverslips and fixed in methanol. Blockage for non- specific binding was performed with normal horse se-rum diluted 1/10 in 1% BSA/PBS for 15 minutes, rinsed with PBS and then incubated with SM3 for 1 hour. After incubation, an HRP-conjugated anti-mouse IgG (DAKO, Copenhagen, Denmark) was added and incubated for

© 2013 Wichtig Editore - eISSN 1724-6008 3
Isla Larrain et al
1 hour before rinsing; the chromogen employed was DAB (SIGMA, St. Louis, MO, USA) in 1% BSA/PBS containing 30% H2O2.
Modified ICC
An adaptation of the ICC was performed employing se-rum samples, instead of the primary antibody, as to evaluate the humoral immune response in patients. Thus, serum was incubated with the MCF7 cell line. After rinsing, a 1/600 HRP-conjugated anti-human IgG (DAKO, Copenhagen, Denmark) was added and incubated for 1 hour before rins-ing; the chromogen employed was DAB (SIGMA, St. Louis, MO, USA) in 1% BSA/PBS containing 30% H2O2.
Inhibition of reactivity with the SM3 anti-MUC1 mAb
MCF7 cells were incubated with the SM3 mAb before the sera-addition step. Further steps were performed as described above.
Transfection of siRNA (Invitrogen) with Lipofectamine 2000 (Invitrogen)
This method was performed following the manufac-turer’s instructions. Briefly, MCF7 cells were transfected with siRNA when cells were 30%-50% confluent. After 72 hours, the transient gene knockdown was evaluated with an anti-MUC1 mAb. Modified ICC was performed with transfected cells to evaluate a decrease in reactivity or intensity of the reaction when incubated with sera. The rest of the technique was performed as described above.
Immunoprecipitation (IP) of MUC1 from serum samples
A total of 500 mL of serum were added to 50 mL of protein A-Sepharose CL-4B (SIGMA, St. Louis, MO, USA), incubated on ice for 30-60 minutes and centrifuged at 10,000 g at 4ºC for 10 minutes. Pre-cleared serum was in-cubated at 4ºC for 1 hour with 10 mL of HMFG1 mAb. Fifty microliters of protein A-Sepharose CL-4B were added to the immune complexes and shook on a rotator at 4ºC for 1 hour. After spinning, the supernatant was removed and the pellet was washed with lysis buffer (1% NP40, 1 mM phenyl methyl sulphonyl fluoride, 150 mM NaCl, 50 mM Tris-HCl, pH 8.0) (SIGMA, St. Louis, MO, USA). Then, 50 ml of Laemmli buffer (2% SDS, 5% 2-mercapoethanol, and 10% glycerol) were added and heated to 90ºC-100ºC for 10 minutes. After centrifugation, the supernatant was loaded on the gel for SDS-PAGE analysis.
SDS-PAGE and Western blot of IP products
Supernatants were analyzed under reducing condi-tions in SDS-PAGE in a discontinuous buffer system ac-
cording to standard procedures. After electrophoresis, gels were electrophoretically transferred to nitrocellulose membranes (Whatman GmbH, Protran BA83 nitrocellu-lose, Dassel, Germany), which were blocked with PBS/5% skimmed milk (blocking buffer). After washing with 0,05% Tween20-PBS (PBST), sheets were incubated with either the HMFG1 mAb (undiluted) or the anti-Tn and anti-TF mAbs (diluted 1/100 in blocking buffer). Sheets were in-cubated overnight at 4ºC and rinsed with PBST buffer. A final incubation with 1/400 peroxidase-conjugated anti-human immunoglobulins was performed according to the manufacturer’s instructions (SIGMA, St. Louis, MO, USA). Nitrocellulose sheets were developed with developed with Enhanced chemiluminiscence (ECL).
MUC1 detection by the CASA test
MUC1 serum levels were measured by a commer-cial CASA test using a dual determinate ELISA (Medical Innovations Limited, Artarmon, Australia). All steps of the CASA test were performed according to the manufactur-ers’ instructions. The working range was between 2 and 64 units/mL; samples that exceeded the 64 units/mL were diluted 1/5 in negative control and re-assayed. This test utilizes the mAbs BC2 (IgG) and BC3 (IgM), both detect-ing the peptide epitope APDTR on the VNTR region of the protein core of the MUC1 mucin; 1/1000 HRP-conjugated anti-mouse IgG were added and detected with ABTS (2,2’-azinobis(3-ethylbenzothiazoline)-6-sulphonic acid). The cutoff level was set at 2 units/mL (0,400 OD units).
ELISA for MUC1-Tn and MUC1-TF detection
Ninety-six-well microplates covered with BC2 anti-MUC1 mAb were incubated with human serum in du-plicates for 1 hour at room temperature; this step was followed by incubation with 1/500 anti-Tn and 1/500 anti-TF mAbs; 1/1000 HRP-conjugated anti-mouse IgM were added and then detected with ABTS.
Study design
The study design was prospective. Samples were tak-en from 2005 to 2011 at institutions related to the Faculty of Medical Sciences of the National University of La Plata, La Plata, Argentina. According to our inclusion criteria, we selected women with breast cancer at different stages, women with BBD and healthy women (controls), from whom it was possible to obtain matched serum and nor-mal/tumor tissue samples before surgery.
Procedures followed the World Medical Association Declaration of Helsinki (Finland, 1964) and its further modifications. Informed consent was obtained from all patients included in this project. This research was approved by the local human investigation committee of
Marina
Resaltado
Marina
Nota adhesiva
Repace by "IgM"
© 2013 Wichtig Editore - eISSN 1724-60084
MUC1 antibodies in breast cancer
the Faculty of Medical Sciences at the National University of La Plata, Argentina.
Statistical analysis methods
Repeated measures ANOVA was employed.
rESuLtS
In a first step, MUC1 expression in normal/tumor tis-sue samples was studied by IHC. Most samples reacted to the HMFG1 mAb: 98% of breast cancer tissues, 100% BBDs and 100% of the normal tissues. By contrast, re-activity to the SM3 mAb was limited to 45% of breast cancer samples, 8% of BBDs and 5% of normal tissue samples.
In order to study the serum reactivity directed to MUC1 expressed by breast cancer tissues, a modified IHC was employed by incubating the normal/tumor breast tissue samples with matched sera from the same patient (autologous). Reactivity and intensity of the reaction were then evaluated.
Next, we verified the anti-MUC1 specificity of the humoral immune response by inhibiting serum reactivity to the anti-MUC1 mAb. Tumoral or normal tissues were incubated with anti-MUC1 mAbs (dilutions: 1/1 SM3, 1/500 HMFG1, and 1/1 HMFG2) before the sera-addition step. At this point, any decrease in reactivity and/or inten-sity of the reaction was analyzed. Both these observations would be suggestive of the presence of circulating anti-bodies against an aberrant glycoform of MUC1 associated to breast cancer.
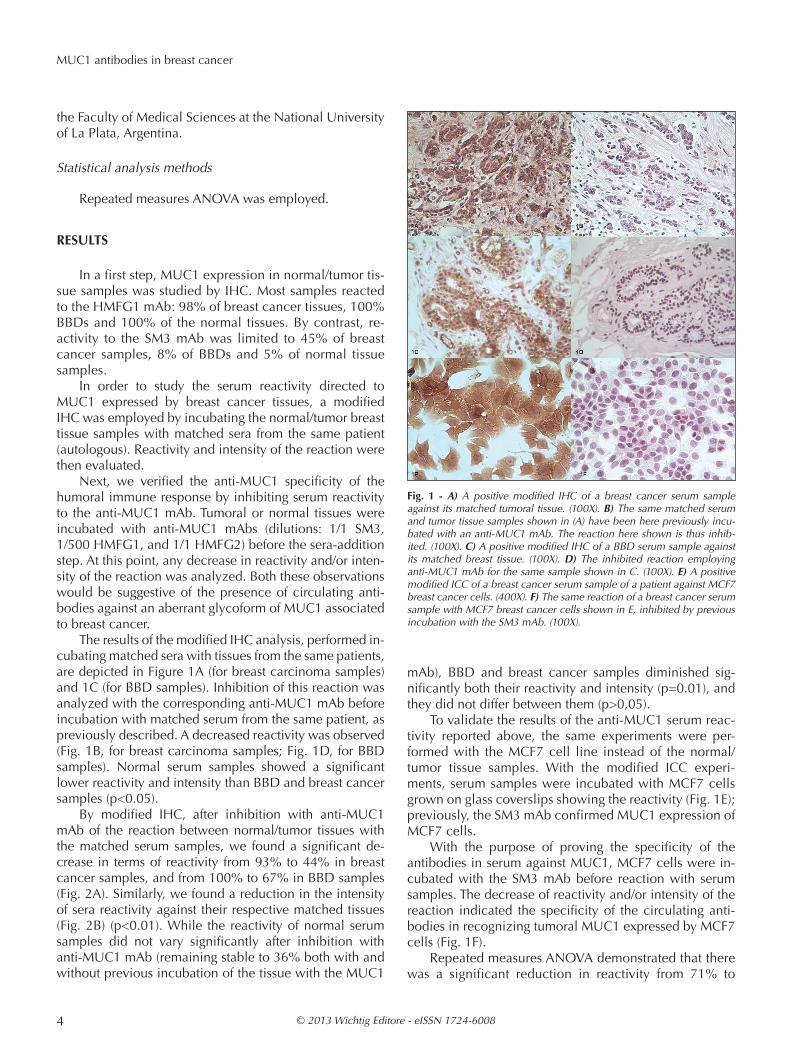
The results of the modified IHC analysis, performed in-cubating matched sera with tissues from the same patients, are depicted in Figure 1A (for breast carcinoma samples) and 1C (for BBD samples). Inhibition of this reaction was analyzed with the corresponding anti-MUC1 mAb before incubation with matched serum from the same patient, as previously described. A decreased reactivity was observed (Fig. 1B, for breast carcinoma samples; Fig. 1D, for BBD samples). Normal serum samples showed a significant lower reactivity and intensity than BBD and breast cancer samples (p<0.05).
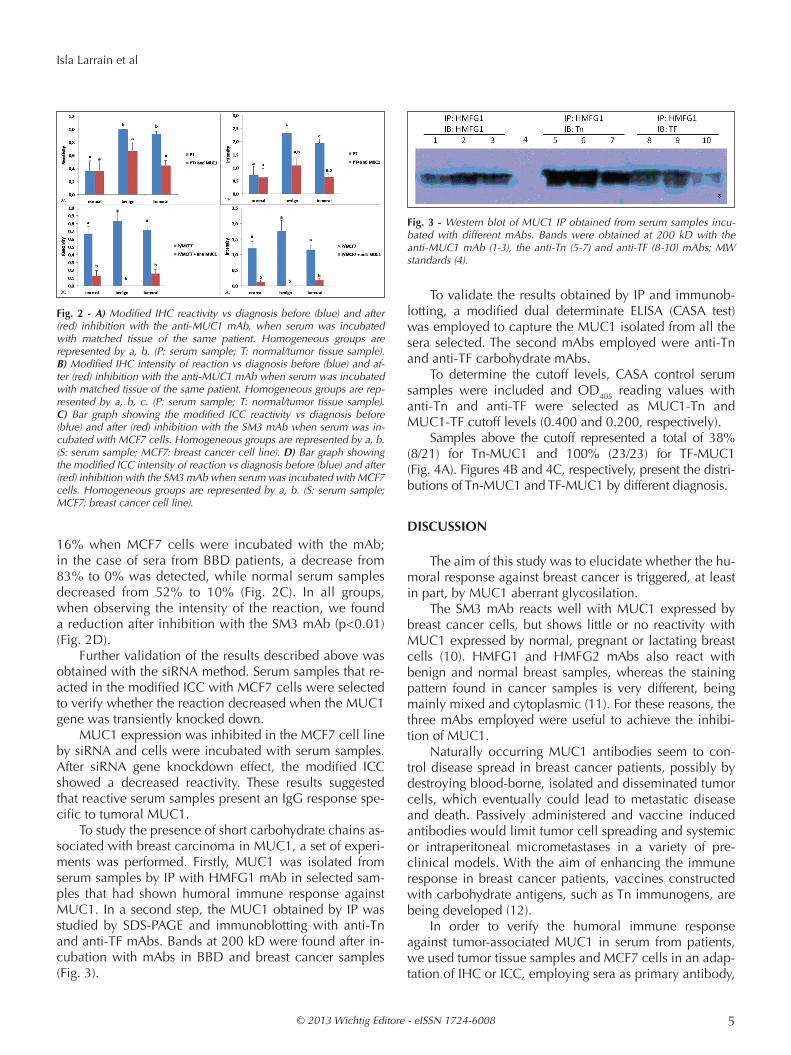
By modified IHC, after inhibition with anti-MUC1 mAb of the reaction between normal/tumor tissues with the matched serum samples, we found a significant de-crease in terms of reactivity from 93% to 44% in breast cancer samples, and from 100% to 67% in BBD samples (Fig. 2A). Similarly, we found a reduction in the intensity of sera reactivity against their respective matched tissues (Fig. 2B) (p<0.01). While the reactivity of normal serum samples did not vary significantly after inhibition with anti-MUC1 mAb (remaining stable to 36% both with and without previous incubation of the tissue with the MUC1
mAb), BBD and breast cancer samples diminished sig-nificantly both their reactivity and intensity (p=0.01), and they did not differ between them (p>0,05).
To validate the results of the anti-MUC1 serum reac-tivity reported above, the same experiments were per-formed with the MCF7 cell line instead of the normal/ tumor tissue samples. With the modified ICC experi-ments, serum samples were incubated with MCF7 cells grown on glass coverslips showing the reactivity (Fig. 1E); previously, the SM3 mAb confirmed MUC1 expression of MCF7 cells.
With the purpose of proving the specificity of the antibodies in serum against MUC1, MCF7 cells were in-cubated with the SM3 mAb before reaction with serum samples. The decrease of reactivity and/or intensity of the reaction indicated the specificity of the circulating anti-bodies in recognizing tumoral MUC1 expressed by MCF7 cells (Fig. 1F).
Repeated measures ANOVA demonstrated that there was a significant reduction in reactivity from 71% to
Fig. 1 - A) A positive modified IHC of a breast cancer serum sample against its matched tumoral tissue. (100X). B) The same matched serum and tumor tissue samples shown in (A) have been here previously incu-bated with an anti-MUC1 mAb. The reaction here shown is thus inhib-ited. (100X). C) A positive modified IHC of a BBD serum sample against its matched breast tissue. (100X). D) The inhibited reaction employing anti-MUC1 mAb for the same sample shown in C. (100X). E) A positive modified ICC of a breast cancer serum sample of a patient against MCF7 breast cancer cells. (400X). F) The same reaction of a breast cancer serum sample with MCF7 breast cancer cells shown in E, inhibited by previous incubation with the SM3 mAb. (100X).
© 2013 Wichtig Editore - eISSN 1724-6008 5
Isla Larrain et al
16% when MCF7 cells were incubated with the mAb; in the case of sera from BBD patients, a decrease from 83% to 0% was detected, while normal serum samples decreased from 52% to 10% (Fig. 2C). In all groups, when observing the intensity of the reaction, we found a reduction after inhibition with the SM3 mAb (p<0.01) (Fig. 2D).
Further validation of the results described above was obtained with the siRNA method. Serum samples that re-acted in the modified ICC with MCF7 cells were selected to verify whether the reaction decreased when the MUC1 gene was transiently knocked down.
MUC1 expression was inhibited in the MCF7 cell line by siRNA and cells were incubated with serum samples. After siRNA gene knockdown effect, the modified ICC showed a decreased reactivity. These results suggested that reactive serum samples present an IgG response spe-cific to tumoral MUC1.
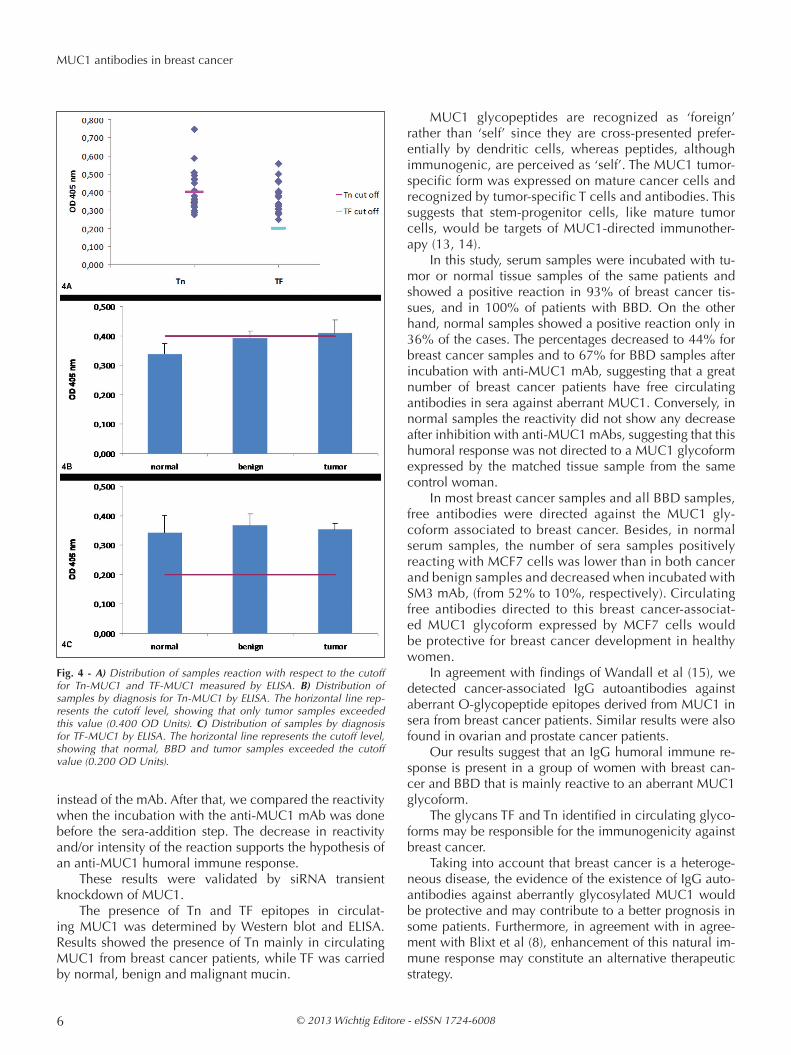
To study the presence of short carbohydrate chains as-sociated with breast carcinoma in MUC1, a set of experi-ments was performed. Firstly, MUC1 was isolated from serum samples by IP with HMFG1 mAb in selected sam-ples that had shown humoral immune response against MUC1. In a second step, the MUC1 obtained by IP was studied by SDS-PAGE and immunoblotting with anti-Tn and anti-TF mAbs. Bands at 200 kD were found after in-cubation with mAbs in BBD and breast cancer samples (Fig. 3).
To validate the results obtained by IP and immunob-lotting, a modified dual determinate ELISA (CASA test) was employed to capture the MUC1 isolated from all the sera selected. The second mAbs employed were anti-Tn and anti-TF carbohydrate mAbs.
To determine the cutoff levels, CASA control serum samples were included and OD405 reading values with anti-Tn and anti-TF were selected as MUC1-Tn and MUC1-TF cutoff levels (0.400 and 0.200, respectively).
Samples above the cutoff represented a total of 38% (8/21) for Tn-MUC1 and 100% (23/23) for TF-MUC1 (Fig. 4A). Figures 4B and 4C, respectively, present the distri-butions of Tn-MUC1 and TF-MUC1 by different diagnosis.
dIScuSSIon
The aim of this study was to elucidate whether the hu-moral response against breast cancer is triggered, at least in part, by MUC1 aberrant glycosilation.
The SM3 mAb reacts well with MUC1 expressed by breast cancer cells, but shows little or no reactivity with MUC1 expressed by normal, pregnant or lactating breast cells (10). HMFG1 and HMFG2 mAbs also react with benign and normal breast samples, whereas the staining pattern found in cancer samples is very different, being mainly mixed and cytoplasmic (11). For these reasons, the three mAbs employed were useful to achieve the inhibi-tion of MUC1.
Naturally occurring MUC1 antibodies seem to con-trol disease spread in breast cancer patients, possibly by destroying blood-borne, isolated and disseminated tumor cells, which eventually could lead to metastatic disease and death. Passively administered and vaccine induced antibodies would limit tumor cell spreading and systemic or intraperitoneal micrometastases in a variety of pre-clinical models. With the aim of enhancing the immune response in breast cancer patients, vaccines constructed with carbohydrate antigens, such as Tn immunogens, are being developed (12).
In order to verify the humoral immune response against tumor-associated MUC1 in serum from patients, we used tumor tissue samples and MCF7 cells in an adap-tation of IHC or ICC, employing sera as primary antibody,
Fig. 2 - A) Modified IHC reactivity vs diagnosis before (blue) and after (red) inhibition with the anti-MUC1 mAb, when serum was incubated with matched tissue of the same patient. Homogeneous groups are represented by a, b. (P: serum sample; T: normal/tumor tissue sample). B) Modified IHC intensity of reaction vs diagnosis before (blue) and af-ter (red) inhibition with the anti-MUC1 mAb when serum was incubated with matched tissue of the same patient. Homogeneous groups are rep-resented by a, b, c. (P: serum sample; T: normal/tumor tissue sample). C) Bar graph showing the modified ICC reactivity vs diagnosis before (blue) and after (red) inhibition with the SM3 mAb when serum was in-cubated with MCF7 cells. Homogeneous groups are represented by a, b. (S: serum sample; MCF7: breast cancer cell line). D) Bar graph showing the modified ICC intensity of reaction vs diagnosis before (blue) and after (red) inhibition with the SM3 mAb when serum was incubated with MCF7 cells. Homogeneous groups are represented by a, b. (S: serum sample; MCF7: breast cancer cell line).
Fig. 3 - Western blot of MUC1 IP obtained from serum samples incu-bated with different mAbs. Bands were obtained at 200 kD with the anti-MUC1 mAb (1-3), the anti-Tn (5-7) and anti-TF (8-10) mAbs; MW standards (4).
© 2013 Wichtig Editore - eISSN 1724-60086
MUC1 antibodies in breast cancer
instead of the mAb. After that, we compared the reactivity when the incubation with the anti-MUC1 mAb was done before the sera-addition step. The decrease in reactivity and/or intensity of the reaction supports the hypothesis of an anti-MUC1 humoral immune response.
These results were validated by siRNA transient knockdown of MUC1.
The presence of Tn and TF epitopes in circulat-ing MUC1 was determined by Western blot and ELISA. Results showed the presence of Tn mainly in circulating MUC1 from breast cancer patients, while TF was carried by normal, benign and malignant mucin.
MUC1 glycopeptides are recognized as ‘foreign’ rather than ‘self’ since they are cross-presented prefer-entially by dendritic cells, whereas peptides, although immunogenic, are perceived as ‘self’. The MUC1 tumor-specific form was expressed on mature cancer cells and recognized by tumor-specific T cells and antibodies. This suggests that stem-progenitor cells, like mature tumor cells, would be targets of MUC1-directed immunother-apy (13, 14).
In this study, serum samples were incubated with tu-mor or normal tissue samples of the same patients and showed a positive reaction in 93% of breast cancer tis-sues, and in 100% of patients with BBD. On the other hand, normal samples showed a positive reaction only in 36% of the cases. The percentages decreased to 44% for breast cancer samples and to 67% for BBD samples after incubation with anti-MUC1 mAb, suggesting that a great number of breast cancer patients have free circulating antibodies in sera against aberrant MUC1. Conversely, in normal samples the reactivity did not show any decrease after inhibition with anti-MUC1 mAbs, suggesting that this humoral response was not directed to a MUC1 glycoform expressed by the matched tissue sample from the same control woman.
In most breast cancer samples and all BBD samples, free antibodies were directed against the MUC1 gly-coform associated to breast cancer. Besides, in normal serum samples, the number of sera samples positively reacting with MCF7 cells was lower than in both cancer and benign samples and decreased when incubated with SM3 mAb, (from 52% to 10%, respectively). Circulating free antibodies directed to this breast cancer-associat-ed MUC1 glycoform expressed by MCF7 cells would be protective for breast cancer development in healthy women.
In agreement with findings of Wandall et al (15), we detected cancer-associated IgG autoantibodies against aberrant O-glycopeptide epitopes derived from MUC1 in sera from breast cancer patients. Similar results were also found in ovarian and prostate cancer patients.
Our results suggest that an IgG humoral immune re-sponse is present in a group of women with breast can-cer and BBD that is mainly reactive to an aberrant MUC1 glycoform.
The glycans TF and Tn identified in circulating glyco-forms may be responsible for the immunogenicity against breast cancer.
Taking into account that breast cancer is a heteroge-neous disease, the evidence of the existence of IgG auto-antibodies against aberrantly glycosylated MUC1 would be protective and may contribute to a better prognosis in some patients. Furthermore, in agreement with in agree-ment with Blixt et al (8), enhancement of this natural im-mune response may constitute an alternative therapeutic strategy.
Fig. 4 - A) Distribution of samples reaction with respect to the cutoff for Tn-MUC1 and TF-MUC1 measured by ELISA. B) Distribution of samples by diagnosis for Tn-MUC1 by ELISA. The horizontal line rep-resents the cutoff level, showing that only tumor samples exceeded this value (0.400 OD Units). C) Distribution of samples by diagnosis for TF-MUC1 by ELISA. The horizontal line represents the cutoff level, showing that normal, BBD and tumor samples exceeded the cutoff value (0.200 OD Units).

© 2013 Wichtig Editore - eISSN 1724-6008 7
Isla Larrain et al
AcKnoWLEdGEMEntS
Grateful acknowledgement is expressed to Professor J. Taylor-Papadimitriou and to Professor J. Burchell (Research Oncology, King’s College London, Guy’s Hospital, UK) for the generous gift of the SM3 and HMFG2 mAbs. Many thanks are expressed to Mr. Juan Carlos Molina and Miss Yamila Blas for technical assistance. María Virginia Croce and Marina Isla Larrain are members of the Research Career of the Comisión de Investigaciones Científicas de la Provincia de Buenos Aires (CIC/PBA).
Informed consent: Signed informed consent was obtained from all patients included in this project.
Financial Support: Supported by the National University of La Plata (UNLP).
conflict of Interest Statements: The authors have declared that no conflict of interest exists.
Address for correspondence: Marina Isla LarrainCentre of Basic and Applied Immunological Research, Faculty of Medical Sciences National University of La PlataSt. 60 and 120, 1900, La Plata, Argentinae-mail: [email protected]
rEFErEncES
1. Gendler SJ. MUC1, the renaissance molecule. J Mammary Gland Biol Neoplasia 2001; 6: 339-53.
2. Finn OJ, Jerome KR, Henderson RA, et al. MUC-1 epithe-lial tumor mucin-based immunity and cancer vaccines. Immunol Rev 1995; 145: 61-89.
3. Croce MV, Isla Larrain MT, Rua CE, Rabassa ME, Gendler SJ, Segal-Eiras A. Patterns of MUC1 tissue expression defined by an anti-MUC1 cytoplasmic tail monoclonal antibody in breast cancer. J Histochem Cytochem 2003; 51: 781-8.
4. Hanisch FG, Müller S. MUC1: the polymorphic appear-ance of a human mucin. Glycobiology 2000; 10: 439-49.
5. Burchell JM, Mungul A, Taylor-Papadimitriou J. O-linked glycosylation in the mammary gland: changes that oc-cur during malignancy. J Mammary Gland Biol Neoplasia 2001; 6: 355-64.
6. von Mensdorff-Pouilly S, Verstraeten AA, Kenemans P, et al. Survival in early breast cancer patients is favorably influ-enced by a natural humoral immune response to polymor-phic epithelial mucin. J Clin Oncol 2000; 18: 574-83.
7. Ragupathi G, Livingston P. The case for polyvalent cancer vaccines that induce antibodies. Expert Rev Vaccines 2002; 1: 193-206.
8. Blixt O, Bueti D, Burford B, et al. Autoantibodies to aber-rantly glycosylated MUC1 in early stage breast cancer are associated with a better prognosis. Breast Cancer Res 2011; 13: R25.
9. Croce MV, Isla Larrain MT, Demichelis SO, Gori JR, Price MR, Segal-Eiras A. Tissue and serum MUC1 mucin de-tection in breast cancer patients. Breast Cancer Res Treat 2003; 81: 195-207.
10. Girling A, Bartkova J, Burchell J, Gendler S, Gillett C, Taylor-Papadimitriou J. A core protein epitope of the polymorphic epithelial mucin detected by the monoclonal antibody SM3 is selectively exposed in a range of primary carcinomas. Int J Cancer 1989; 43: 1072-6.
11. Burchell J, Gendler S, Taylor-Papadimitriou J, et al. Development and characterization of breast cancer reac-tive monoclonal antibodies directed to the core protein of the human milk mucin. Cancer Res 1987; 47: 5476-82.
12. Tarp MA, Sørensen AL, Mandel U, et al. Identification of a novel cancer-specific immunodominant glycopeptide epi-tope in the MUC1 tandem repeat. Glycobiology 2007; 17: 197-209.
13. Ryan SO, Vlad AM, Islam K, Gariépy J, Finn OJ. Tumor-associated MUC1 glycopeptide epitopes are not subject to self-tolerance and improve responses to MUC1 peptide epitopes in MUC1 transgenic mice. Biol Chem 2009; 390: 611-8.
14. Engelmann K, Shen H, Finn OJ. MCF7 side population cells with characteristics of cancer stem/progenitor cells express the tumor antigen MUC1. Cancer Res 2008; 68: 2419-26.
15. Wandall HH, Blixt O, Tarp MA, et al. Cancer biomark-ers defined by autoantibody signatures to aberrant O-glycopeptide epitopes. Cancer Res 2010; 70: 1306-13.




















