
Fill Ups of Straight Lines and Pair of Straight Lines - SelfStudys
Upload
independentCategory
view
0download
0
ROMANIAN ARCHIVESOF
MICROBIOLOGY AND
IMMUNOLOGY
TOTAL PUBLISHING HOUSE
Founded byPROFESSOR ION CANTACUZINO
VOLUME 64 - Nos. 1-4January - December 2005
Published quarterly
by
CANTACUZINO INSTITUTE BUCHAREST

2
ROMANIAN ARCHIVES OF MICROBIOLOGY AND IMMUNOLOGY
Chief Editor: Dorel Lucian RADU
Editorial Assistant: Maria ªTEFÃNESCU
Editorial Board: Cornelia CEIANU, Maria DAMIAN, Emilia LUPULESCU,Cristiana MATACHE, Adrian ONU, Gabriela OPRIªAN, Mircea Ioan POPA,Aurora SÃLÃGEANU, Dan STERIU, Codruþa Romaniþa USEIN
Editorial Staff: Felicia RAPILAT, Monica POEANÃ
TOTAL PUBLISHING HOUSE
ROMANIAN ARCHIVESOF
MICROBIOLOGY AND
IMMUNOLOGY
Subscription orders:Orders can be placed directly with the publisher:
„Cantacuzino“ National Institute of Research-Development for Microbiology and Immunology
C.P. 1-525, Splaiul Independentei 103, 050096, Bucureºti, Romania, Fax: (40.21)318.44.14
E-mail: [email protected]
ISSN 1222-3891
INDEXED IN MEDLINE

ROMANIAN ARCHIVES OF MICROBIOLOGY AND IMMUNOLOGY
3
IMMUNOLOGY
Experimental studies on bacterial product CANTASTIMderived from Pseudomonas aeruginosa. VII. Activationof immune cells of healthy controls and cancer patientsIULIANA CARAª, IULIANA FRANCISCA ªERBÃNESCU, A.GRIGORESCU and AURORA SÃLÃGEANU
Microbiological and Immunologial study of Staphylococ-cus Vaccine Effects in Periodontitis CRINA STAVARU, VIORICA DUMITRESCU, DANIELA LEMENI,I.B.T. GEORGESCU, IRINA CODIÞÃ and D.L RADU
Patterns of peripheral cellular immune disorders in se-vere rheumatoid arthritis GINA MANDA, MONICA NEAGU, CAROLINA CONSTANTIN,ALINA RÃDULESCU and C. CODREANU
Neurotrophin-3, TNF-αα and IL-6 relations in serum andcerebrospinal fluid of ischemic stroke patients DANIELA PÃSÃRICÃ, MIHAELA GHEORGHIU, FLORICATOPÂRCEANU, CORALIA BLEOTU, LORETTA ICHIM and T.TRANDAFIR
BACTERIOLOGY
Incidence of Virulence-Encoding Genes among EntericEscherichia coli Strains Isolated from Healthy SubjectsMARIA DAMIAN, CODRUÞA-ROMANIÞA USEIN, DORINATATU-CHIÞOIU, A.M. PALADE, NATALIA POPOVICI, SIMONACIONTEA, MARIA NICA and LAURA GRIGORE
Application of Pulsed-Field Gel Electrophoresis to labo-ratory surveillance of small community outbreaks of Sal-monella enterica serovar Typhimurium CODRUTA-ROMANIÞA USEIN, A.M. PALADE, NATALIAPOPOVICI, DORINA TATU-CHIÞOIU, SIMONA CIONTEA,ªTEFANIA CECIU, MARIA COªMAN and MARIA DAMIAN
Rapid immunochromatographic serum assay of non-tuberculous mycobacterial infectionsHENRIETTE STAVRI, MANUELA BRÂNARU-GHEORGHIU,OLGA MOLDOVAN, MARINELA RÃILEANU, M.I. POPA,LOREDANA POPA and LUMINIÞA ENE
Intracellular degradation of poly(3-hydroxybutyric acid)accumulated by Azotobacter chroococcum MAL-201SOMA PAL SAHA and A.K. PAUL
CELL BIOLOGY
The effect of the plasma needle on tumoral cell linesapoptosisANA CÃLUGÃRU, LIDIA CREMER, AURORA HEROLD,ANDREEA LUPU, G. SZEGLI, C. LUNGU, ANA LUNGU and N.GEORGESCU
BIOCHEMISTRY
Preliminary identification and classification of five newYeast strains isolated from oil-polluted environmentORTANSA CSUTAK, RALUCA GHINDEA, ILEANA STOICA,SIMONA SOARE, ROBERTINA IONESCU, OANA CREANGÃ and
TATIANA VASSU
MYCOLOGY
Case-control study to evaluate the link betweenimmunosuppression and Candida spp. infectionLOREDANA GABRIELA POPA, M.I. POPA and IOANARUXANDRA MIHAI
AUTHOR INDEX
CONTENTS
VOLUME 64 NOS.1-4 JANUARY-DECEMBER 2005
5
11
17
27
34
42
49
57
65
72
77
39

4
ROMANIAN ARCHIVES OF MICROBIOLOGY AND IMMUNOLOGY
Aims and ScopeRomanian Archives of Microbiology and Immunoloy, an interna-tional journal dedicated to original research works, publishes papersfocusing on various aspects of microbiology and immunology.Romanian Archives of Microbiology and Immunology is indexedin MEDLINE. The frequency of the Journal is currently four issuesper year.
Categories of manuscriptsFull-length articles are full-length descriptions of original research(up to 10 printed pages).Reviews are comprehensive appraisals of research in a field of cur-rent interest. All reviews are subject to the normal review process(up to 15 printed pages)Rapid Communications are brief, definitive reports of highly sig-nificant and timely findings in the field (up to 5 printed pages).
Submission of manuscriptsManuscripts and illustration should be submitted in electronic formusing the e-mail address of the Editorial Office:[email protected] preferred software is: MS Word or Adobe PDF for text andAdobe Photoshop or Corel for images.
Editorial review and acceptanceAll manuscripts are subject to editorial review by professional peerreviewers (at least two). The Editor will decide if a paper is pub-lished or not. The acceptance criteria for all manuscripts are basedon the quality and originality.
Ethical considerationsA paper describing any experimental work with humans shouldinclude a statement that the Ethics Committee of the institution inwhich the work was done has approved it, and that the subjectsgave informed consent to the work. Experiments with animals should be done in accordance with thelegal requirements of the relevant local or national authority.Procedures should be such that animals used in experiments donot suffer unnecessarily. Papers should include details of the pro-cedures and anaesthetics used. The Editors will not accept paperswhere the ethical aspects are, in their opinion, open to doubt.
Preparation of manuscriptsManuscripts should be submitted in English. American or Britishspelling can be used provided that only one spelling style is con-sistently used throughout. Manuscripts must be typewritten on A4format (210x297 mm), with double spacing, margins of 25 mm, onone side only, consecutively numbered. Times New Roman font,12-point size, is required.Manuscripts should be divided into the following sections andorder: Title page, Abstract and key words, Introduction, Materialsand Methods, Results, Discussion, Acknowledgements, References,Tables, Figure Legends and Figures. Title page contain: title of the paper not longer than 80-100 cha-racters, including spaces and punctuation; full names (includingforenames) of the authors and the name of their institute(s); theauthor responsible for correspondence will be marked by an asterisk,and his full address, telephone/fax numbers, and e-mail address willbe indicated.Abstract must not exceed 250 words and must reflect the contentof the study. Following the abstract, a list of 3-10 keywords isessential for indexing purposes.Introduction contains a description of the problem under investi-gation and a brief survey of the existing literature on the subject. Material and Methods provide sufficient detail to allow the workto be reproduced.Results. Results should be clear and concise.
Discussion that enriches but does not repeat Section 3 or 5.Acknowledgements (if applicable) containing acknowledgement oftechnical help and of financial material support.References should be numbered consecutively in the order inwhich they are first mentioned in the text. Identify references in text,tables, and legends by Arabic numerals in square brackets (e.g. [1],[2-6], etc.). Please note the following examples:Journals:Allain F, Vanpouille C, Carpentier M, Slomianny MC, Durieux S,Spik G. Interaction with glycosaminoglycans is required forcyclophilin B to trigger integrin-mediated adhesion of peripheralblood T lymphocytes to extracellular matrix. Proc Natl Acad SciUSA 2002. 99: 2714-2719.Books:Theofilopoulos AN. Immune complexes in autoimmunity. In: BonaCA, Siminovitch KA, Zanetti M, Theofilopoulos AN (Eds.) TheMolecular Pathology of Autoimmune Diseases. HarwoodAcademic Publishers, Switzerland 1993, pp 229-244.Tables with suitable captions at the top and numbered with Arabicnumerals should be collected at the end of the text on separatesheets (one page per Table). Footnotes to tables should be markedwith a) b) c) etc and *, **, *** should be reserved for p values.Each table must be understood independently of the text. All tablesmust be cited in the text. Figures (illustrations) Figures should be submitted on separatepages at the end of the article (new page for each complete figure).They should be numbered in the order of their appearance with Arabicnumerals. Figures should be submitted as TIFF files at a proper reso-lution as follows: Graphs at 800-1200 dpi; Photos at 400-800 dpi;Color 300-400 dpi. Text in figures should be 8-10 point in size.Each figure must have a separate legend. The legends should notappear under the figures, but be gathered in a separate section(Figure legends). Color figures can only be printed if the author isprepared to pay the cost incurred.Figure legends should be supplied at the end of the manuscript,double spaced, with relevant figure numbers, labeling symbol andexplanation.
Units of measurement, Symbols and abbreviationsSymbols for physical units should be those of the Système Inter-nationale (SI) Units.Alternative or non-SI units may be used, but these must be definedat their first occurrence in the text.
Nomenclature of MicroorganismsBinary names, consisting of a generic name and a specific epithet(e.g., Escherichia coli), must be used for all microorganisms.
Genetic NomenclatureTo facilitate accurate communication, it is important that standardgenetic nomenclature be used whenever possible and that devia-tions or proposals for new naming systems be endorsed by anappropriate authoritative body.
Proofs and reprintsTen reprints of each article (free of charge) will be sent to the cor-responding author.
Cover letterEach manuscript submitted to the Romanian Archives of Micro-biology and Immunoloy must be accompanied by a Cover letterincluding statement that:- the material represents an original work, has not been previously
published, and that it has not been submitted simultaneously forpublication elsewhere.
- all authors of a manuscript concure with the submission and areresponsible for its content.
INSTRUCTIONS TO AUTHORS

5
Our previous reports demonstrated the ability of CAN-TASTIM (CS) a purified extract of Pseudomonas aerugi-nosa to protect against experimental bacterial infec-tions in mice and to modulate cytokine production inmurine macrophages /1, 2/.
Clinical uses of CANTASTIM include prevention andtreatment of viral and bacterial infections and immu-norestorative treatment of cancer patients. Nevertheless,its mechanism of action is not fully understood. In thispaper, we examined some in vitro effects of CS on hu-man leukocytes activated in whole blood. As CS is amixture of bacterial components, it might be presumedthat at least some of these components could be recog-nized by recently identified Toll-like receptors (TLR).
These are pathogen recognition receptors (PRRs)which recognize pathogen associated molecular pat-terns (PAMPs) expressed by microorganisms /3/. Theyare not only involved in innate immune response butinitiate and orientate adaptive immune response /4/.Signaling through TLRs leads to the activation of NF-kB
ABSTRACTCANTASTIM (CS) is a purified extract of Pseudomonas aeruginosa with beneficial effects relatedto enhancing the immune responses in conditions such as chronic viral and bacterial infections,immunodeficiencies and cancer immunosuppression. The aim of this study was to determinethe capacity of this biological product to stimulate in vitro human leukocytes in whole blood.Blood samples from healthy donors and cancer patients were incubated with CS for 24 h andleukocytes were assessed for induction of inflammatory cytokines (TNF-α and IL-1β) by ELISAand expression of early activation marker CD69 by flow-cytometry. For both groups of investi-gated subjects, the levels of TNF-α and IL-1β in the supernatants of whole blood culture stim-ulated with CS were significantly higher than in unstimulated cultures, although lower than inLPS-stimulated samples. Stimulation of whole blood cultures with CS increased both the fre-quency and the expression of CD69 on the surface of T lymphocytes and NK cells. Importantly,this was noticed not only for healthy controls, but also for cancer patients. These data demon-strate the capacity of bacterial immunomodulator CS to activate human leukocytes of healthysubjects and cancer patients.
transcriptional factor and synthesis of pro-inflammatorycytokines /5/. We were interested to find if CS inducesthe production of inflammatory cytokines in humanleukocytes. We chose to use blood cells from healthysubjects and also from cancer patients.
It is known that immunosuppression is associatedwith cancer and contributes not only to the increase ofthe risk of infections but also to the progression of can-cer itself /3/.
T lymphocytes and NK cells represent the main cyto-toxic effectors of immune response and therefore theiractivation is crucial for eliciting an effective anti-tumorimmunity.
In a previous paper we showed that peripheral Tlymphocytes and NK cells of cancer patients could beinduced to express early activation marker CD69 uponex vivo mitogen stimulation /6/. In this study we inves-tigated if cells from healthy subjects and cancer pa-tients could be induced to express CD69 in vitro by CSstimulation.
EXPERIMENTAL STUDIES ON BACTERIAL PRODUCT CANTASTIM DERIVED FROM PSEUDOMONAS AERUGINOSA. VII. ACTIVATION
OF IMMUNE CELLS OF HEALTHY CONTROLS AND CANCER PATIENTS
Iuliana Caras1, Iuliana Francisca ªerbãnescu1, A. Grigorescu2, Aurora Sãlãgeanu1
(1Infection and Immunity Laboratory, "Cantacuzino" National Institute of Research - Development for Microbiology and Immunology, 2Oncology Institute "Prof. Dr. Alex. Trestioreanu", Bucharest, Romania)

6
CARAª et al.
MATERIALS AND METHODS
PatientsEleven patients with lung cancer and four patients withbreast cancer were included in this study after infor-med consent. Eight healthy volunteers were includedas the control group. The main characteristics of the in-vestigated subjects are summarized in Table 1. The agedistribution between patients and controls was statisti-cally similar (Student's t-test).
Whole-blood cultures Peripheral blood samples were mixed with equal volu-mes of RPMI 1640 (Sigma, St. Louis, MO, USA) culturemedium containing 5 U of heparin per ml, dispensedon 96-well tissue culture plates (250 µl/well) and incu-bated at 37oC in 5% CO2 for 24 h. Cultures were stimu-lated 18-24 h with CS (10 µg/ml) and lipopolysaccha-ride (LPS E. coli, Sigma, St. Louis, MO, USA, 10 µg/ml)as a positive control.
Cytokines measurementAt the end of incubation, whole-blood samples collec-ted from 2 wells were centrifuged (quick run, 10 s) andthe supernatants were stored at -70oC until tested forcytokine secretion. The level of cytokines (IL-1β andTNF-α) in the samples was determined by enzyme-lin-ked immunosorbent assays, using DuoSet ELISA Deve-lopment Systems (R&D Systems).
Flow cytometry analysisThe expression of CD69 upon activation was determi-ned on the surface of T lymphocytes and NK cells afterCS or LPS stimulation. After 24 h of incubation, whole-blood samples (100 µl) were mixed with 10 µl of CD3FITC (fluorescein isothiocyanate) / CD16+ CD56 PE(phycoerythrin) and 10 µl of anti-CD69 antibody taggedwith peridinin chlorophyll protein (PerCP) and incu-bated for 30 min at room temperature in the dark. Afterincubation, 1 ml of Lysing Solution diluted 1/10 wasadded to samples to lyse erythrocytes. After centri-fugation and washing, the samples were fixed using0.5 ml of 1% paraformaldehyde. Samples were analy-
zed within 24 h using a BD FACS Calibur flow-cyto-meter with Cell Quest software for data acquisitionand analysis. Acquisition gates were set on a FL1/SSCdot-plot for T lymphocytes (CD3+) and a FL2/SSC dot-plot for NK cells (CD16+CD56+). Data for a mini-mum of 5,000 events were acquired within these ga-tes. The percentages of CD69 positive cells were determi-ned in histograms of CD69-PerCP fluorescence esta-blished for the gated populations of T lymphocytes andNK cells, respectively.
Statistical analysisData were statistically analyzed using paired Student'st-test for two tailed values. The p values below 0.05were considered significant. Results were presented asthe mean ± standard deviation.
RESULTS
Effect of CANTASTIM on the production of inflamma-tory cytokines by human blood leukocytes Previously we have shown that CS could induce theproduction of TNF-α in murine macrophages and thispartially explained its protective effect in experimentalbacterial infections /1/. In this study we stimulatedhuman whole blood cultures of healthy subjects andcancer patients with CS and LPS (as a positive control)and measured the level of TNF-α (Fig. 1) and IL-1β(Fig. 2) in culture supernatants after 18 h incubation.As can be seen, both bacterial extracts, LPS and CSinduced a marked secretion of inflammatory cytokinesin whole blood culture. The levels of both cytokineswere comparable between cancer patients (both breastand lung) and healthy subjects groups. It could be noti-ced that the concentration of both cytokines wasapproximately two times lower in the cultures stimu-lated with CS as compared to LPS, at the same con-centration (10 µg/ml).
Effect of CANTASTIM on expression of CD69 on Tlymphocytes and NK cellsCD69 is an activation marker expressed very early onlymphocytes surface. In these experiments, we assessed
Table 1 - Characteristics of cancer patients and healthy controls

Cantastim activates immune cells of healthy controls and cancer patients
7
the capability of CS to activate different blood leuko-cytes in whole blood by measuring the expression ofCD69 on cell surface by flow cytometry. Whole bloodsamples were incubated for 24 hours with saline, LPS(10 mg/ml) or CS (10 mg/ml). Stimulation with bothbacterial extracts significantly enhanced the percenta-ge of cells expressing CD69 molecule among CD16 +CD56+ NK cells (Fig. 3a) and CD3+ T lymphocytes(Fig. 3b). This was noticed not only for healthy donors,
but also for cancer patients. The differences betweenLPS-stimulated samples and unstimulated controls werestatistically significant for all the groups of patients. Animportant increase in the percentage of both T lympho-cytes and NK cells expressing CD69 was noticed alsofor CS-stimulated cultures, with differences reachingthe statistical significance in the case of healthy con-trols and lung cancer patients. To better evaluate theeffect on CD69 expression we compared the mean
Fig. 1 - Production of TNF- after 24h stimulation with 10 µg/ml LPS (positive control) or CS (10 µg/ml) in whole blood cul-tures from healthy donors and cancer patients as measured by ELISA in cell supernatants. Values are mean ± SD (n=5 forhealthy controls and n=10 cancer patients, 2 with breast cancer and 8 with lung cancer).
*p<0.001; **p<0.05 (Student's t-test) as compared to unstimulated cells
Fig. 2 - Production of IL-1 after 24h stimulation with 10 µg/ml LPS (positive control) or CS (10 µg/ml) in whole blood culturesfrom healthy donors and cancer patients as measured by ELISA in cell supernatants. Values are mean ± SD (n=5 for healthycontrols and n=10 cancer patients, 2 with breast cancer and 8 with lung cancer).
*p<0.001; **p<0.05 (Student's t-test) as compared to unstimulated cells

8
CARAª et al.
stimulation indices (SI) between control and cancergroups. SI was defined as:
Mean intensity of fluorescence (MIF) of CD69 mar-ker in stimulated cultures / MFI of CD69 marker in un-stimulated cultures. The results are presented in Fig. 4.As can bee seen, the expression of CD69 on NK cellsand T lymphocytes upon stimulation was comparableamong healthy controls and cancer patients.
DISCUSSION
In this study we have investigated the effects of CS ondifferent blood leukocytes using whole blood culture.Whole blood was used instead of purified peripheralblood leukocytes because this system represents a
more natural and physiological system that most close-ly mimics conditions in vivo /7/.
We have shown that bacterial extract CS inducedthe production of comparable levels of pro-inflamma-tory cytokines IL-1β and TNF-α in whole blood cultureof healthy controls and cancer patients. This suggestedthat CS was able to activate monocytes to produce pro-inflammatory cytokines. In other words, the capacityof immunocompetent cells to respond to activation bybacterial components by producing TNF-α and IL-1βwas not impaired in cancer patients. By contrast, inprevious study we showed that the production of IFN-α in peripheral blood leukocytes culture stimulatedwith LPS was impaired in breast and lung cancer
Fig. 3 - CD69 surface expression on NK cells (a) and T lymphocytes (b). Whole blood samples were cultured in the absenceor presence of LPS (10 µg/ml) or CS (10 µg/ml) for 24 h. Stimulated samples were labeled with CD3 FITC, CD16CD56 PE,and CD69 PerCP. Surface expression of CD69 was determined by three-color flow cytometric analysis. Analysis was per-formed by gating on CD3+ cells (T lymphocytes) or CD16+CD56+ cells (NK cells), and dead cells were excluded by for-ward/side scatter gating. Figures indicate percentages of CD69+ cells and are representative of samples from four patientswith breast cancer, eleven patients with lung cancer, and five healthy control subjects. Values are mean ± SD
*p<0.05; **p<0.01; ***p<0.001 (Student's t-test) as compared to unstimulated cells
a. NK cells
b. T lymphocytes

Cantastim activates immune cells of healthy controls and cancer patients
9
patients /8/. This is in agreement with other studies inliterature showing that suppression of cytokine secre-tion in patients with lung cancer was selective. Com-pared to normal controls, immunocompetent cells frompatients with small-cell lung cancer (SCLC) secretedsignificantly lower amounts of IL-2, IFN-α and IFN-γ,upon mitogen stimulation. TNF-α secretion was signi-ficantly reduced in SCLC extensive disease but not inSCLC limited disease. In contrast, secretion of IL-1αand IL-1β was not reduced /9/. In preliminary experi-ments, using whole blood culture of normal healthycontrols we did not notice an increase in the produc-tion of IFN-γ after a 24 hours stimulation with LPS or
CS (unpublished observations). However, a longer sti-mulation might be necessary to induce the productionof IFN-γ in cell supernatants, as suggested /10/.
CD69 is an early activation marker which is rapidlyacquired following in vitro activation of T and B lym-phocytes, NK cells, dendritic cells, neutrophils, andeosinophils /11/. Several immunomodulators havebeen shown to up-regulate the expression of CD69, es-pecially on the surface of NK cells /7, 12/. Bacterialimmunostimulant RU41740 from Klebsiella pneumo-niae highly activated NK cells as observed by the stronginduction of CD69 and early production of IFN-γ. Itincreased CD69 expression also on B lymphocytes
Fig. 4 - CD69 surface expression on NK cells (a) and T lymphocytes (b). Figures indicate stimulation indices and are repre-sentative of samples from four patients with breast cancer, eleven patients with lung cancer, and five healthy control subjects.Values are mean ± SD
a. NK cells
b. T lymphocytes

5. TAKEUCHI O. and AKIRA S. - "Toll-like receptors; theirphysiological role and signal transduction system", IntImmunopharmacol 1, 2001, 625-635
6. HADEN J.W. - "Immunodeficiency and cancer: prospectsfor correction", Int Immunopharmacol 3, 2003, 1061-1065
7. PICHYANGKUL S., YONGVANITCHIT K., KUM-ARB U.,KRIEG A.M., HEPPNER D.G. and WALSH D.S. - "Wholeblood cultures to assess the immunostimulatory activities ofCpG". J Immunol Meth 247, 2001, 83-94
8. CARAS I., GRIGORESCU A., STAVARU C., RADU D.L.,MOGOS I., SZEGLI G. and SALAGEANU A. - "Evidence forimmune defects in breast and lung cancer patients". CancerImmunol Immunother 53(12), 2004, 1146-1152
9. FISCHER J.R., SCHINDEL M., STEIN N., LAHM H., GALLATIH., KRAMMER P.H. and DRINGS P. - "Selective suppressionof cytokine secretion in patients with small-cell lung can-cer". Ann Oncol. 6, 1995, 921-926
10. POPA C., NETEA M.G., BARRERA P., RADSTAKE TRDS,VAN RIEL PL., KULLBERG B-J and VAN DER MEER JWM -"Cytokine production of stimulated whole blood culturesin rheumatoid arthritis patients receiving short-term infli-ximab therapy", Cytokine 30, 2005, 72-77
11. H-H, ZHANG L., HEBER D. and BONAVIDA B. - "Mecha-nism of activation of human peripheral blood NK cells atthe single cell level by Echinacea water soluble extracts:recruitment of lymphocyte-target conjugates and killercells and activation of programming for lysis". Int Immuno-pharmacol. 3, 2003, 811-24
12. PEDRAZA-SANCHEZ S., GONZALES-HERNANDEZ Y.,ESCOBAR-GUTIERREZ A. and RAMACHANDRA L. - "Theimmunostimulent RU41740 from Klebsiella pneumoniaeactivates human cells in whole blood to potentially stimu-late innate and adaptative immune response". Int Immuno-pharmacol 6, 2006, 635-646
13. HEIZELMANN M., POLK H.C. J.R., CHERNOBELSKY A.,STITES T.P. and GORDON L.E. - "Endotoxin and muramyldipeptide modulate surface receptor expression on humanmononuclear cells". Immunopharmacol, 14, 2000, 117-128
14. CLAUSEN J., VERGEINER B., ENK M., PETZER AL., GASTLG., GUNSILIUS E. - "Functional significance of the activation-associated receptors CD25 and CD69 on human NK-cellsand NK-like T-cells", Immunobiology, 207, 2003, 85-93
15. SIVORI S., CARLOMAGNO S., MORETTA L. andMORETTA A. - "Comparison of different CpG oligodeoxy-nucleotide classes for their capability to stimulate humanNK cells". Eur J Immunol. 36, 2006, 961-967
CARAª et al.
and, at higher concentration, on T lymphocytes /12/. Whencomparatively studying two bacterial components, LPSand muramyldipeptide (MDP) HEINZELMANN et al.have noticed that LPS induced the expression of CD69marker by a CD14-dependent mechanism, whereasMDP did not induce CD69 expression /13/.
In our study, stimulation of whole blood cultureswith bacterial product CS increased both the frequen-cy and the expression of CD69 on the surface of T lym-phocytes and NK cells. Importantly, this was noticed notonly for healthy controls, but also for cancer patients.
Recent studies showed that increased expression ofCD69 on the surface correlated with augmented cyto-toxic activity of NK cells in lymphocyte cultures fromperipheral blood of breast cancer patients /14, 15/. Theapplication of autologous ex vivo expanded cytotoxiclymphocytes to cancer patients may help to control mi-nimal residual disease and increased expression ofCD69 might be used as an activation marker to opti-mize this treatment.
Collectively, our results suggest that bacterial ex-tract CS may be used to activate immunocompetent cells,including cells from cancer patients. Administration ofCS to cancer patients may result in an improvement oftheir immunity, due to the ability of CS to stimulate cy-tokine production and activation of different cells.
Acknowledgements. This work was supported by agrant from the Romanian Ministry of Education, Re-search and Youth (Program Biotech, grant 097). Theauthors gratefully acknowledge the skillful technicalassistance of Camelia Tabarta, Rodica Tudorache, andFlorica Chioseolu.
REFERENCES
1. SALAGEANU A., CEACAREANU B., POPESCU D.,ISTRATE N. and SZEGLI G. - Experimental studies on bac-terial product CANTASTIM derived from Pseudomonas ae-ruginosa. II. Protective effect in Salmonella typhimurium in-fection, Rom. Arch. Microbiol. Immunol. T56, 1-2,17, 1997
2. SALAGEANU A., CEACAREANU B., ISTRATE N. andSZEGLI G. - "Experimental studies on bacterial productCANTASTIM derived from Pseudomonas aeruginosa. III. Su-ppression of lipopolysaccharide-induced tumor necrosisfactor alpha: are the lipid components involved?", Rom.Arch. Microbiol. Immunol. T56, 1-2, 27-35, 1997
3. AKIRA S. and HEMMI H. - "Recognition of pathogen-asso-ciated molecular patterns by TLR family", Immunol Letters85, 2003, 85-95
4. PASARE C. and MEDZHITOV R. - "Toll-like receptors: link-ing innate and adaptive immunity", Microb Infect 6, 2004,1382-1387
10

In most of the oral cavity surfaces there are plenty ofbacteria, which form a complex structure known asbiofilm. While the majority of these organisms arecommensals, a subset of them are likely to be oppor-tunistic pathogens that are able to cause oral disorderssuch as caries and periodontitis but also systemic dis-ease (23, 8). Epidemiological studies indicate thatapproximately 15% of the population is at high risk formoderate to severe periodontitis (22,16). Chronicinflammatory periodontal disease is characterized bymononuclear infiltration into gingival tissues leadingto connective tissue destruction and alveolar boneresorption. The causative agents of periodontitis arebacteria but the progression and disease severity arethought to be determined by host immune response (4,12, 14, 17, 18, 25).
Modern periodontal therapy aims to suppress oreradicate periodontal pathogens and to maintain a
posttreatment flora that is compatible with health.Orally administered antimicrobial agents often lead toecological disturbances in the normal oral and intes-tinal microflora. The use of antimicrobial agents alsopromotes the emergence of drug resistance so vacci-nation could represent a therapeutical approach (9, 22,6). Previous clinical studies demonstrated the benefitsof corpuscular staphylococcal vaccine in periodontitis(10,11,13).
The aim of the present study was to detect by clas-sical microbiology procedures the presence ofStaphlylococcus spp. and of anaerobic microorgan-isms in periodontal pockets and to investigate humoraland cellular immunity of the patients before and aftervaccine administration. The following procedures wereperformed: quantitation of C reactive protein (CRP),C3 complement fraction, immunoglobulin IgA, IgG, IgMby immunodifussion assay, PMN reactive oxygen
11
ABSTRACTThe aim of this study was to evaluate the immunomodulatory effect of the staphylococcal vac-cine inoculated subcutaneously in 15 patients with chronic periodontitis. Bacteriological inves-tigation of samples collected from the periodontal pocket for aerobic and anaerobic microor-ganisms was performed by classic bacteriological procedures before and after vaccination. Thefollowing immune system parameters were evaluated: C reactive protein (CRP), serum level ofC3 complement fraction, IgG, IgA, and IgM by immunodiffusion, PMN granulocytes ROSrelease after in vitro stimulation with opsonized zymosan (OZ) and Concanavalin A (ConA) bychemiluminescence assay and lymphocytes sets and subsets by flow-cytometry immunophe-notyping. The microbiological investigations revealed high frequency of Staphylococcus sppisolation and the presence of the most common anaerobe agents incriminated in human peri-odontitis like Fusobacterium, Porphyromonas, Peptostreptococcus, Veillonella spp and thereduction of this flora in the periodontal pocket after therapy. The immunological parametersquantification showed the absence of CRP, normal values of C3, IgG, IgA, IgM in the majorityof cases. All patients presented normal values of lymphocytes sets and subsets. Significantincrease of PMN respiratory burst after ConA stimulation was observed before vaccinationwhich turned to normal values after therapy and a low ROS level both before and after thera-py suggesting PMN Fc receptors dysfunction in this group of patients.The data presented in our study suggest an immunomodulatory effect of staphylococcal vac-cine therapy in periodontitis and high frequency of Staphylococcus spp recovering from theperiodontal pocket of investigated subjects.
MICROBIOLOGICAL AND IMMUNOLOGICAL STUDY OF STAPHYLOCOCCUS VACCINE EFFECTS IN PERIODONTITIS
Crina Stavaru1, Viorica Dumitrescu1, Daniela Lemeni1, Ion Bogdan Teodor Georgescu2, Irina Codita1, Dorel Lucian Radu1
(1“Cantacuzino” NIRDMI; 2“Titu Maiorescu” University, Bucharest - Romania)

species (ROS) generation by chemiluminescence assayand detection of lymphocytes sets and subsets by flow-cytometry.
MATERIALS AND METHODS
Subjects: Fifteen subjects with clinically documentedperiodontitis representing both genders, ranging in agefrom 35 to 70 and eight healthy volunteers with goodoral hygiene as controls for chemiluminescence assaywere included in the study. The patients did not use an-tibiotics and they were treated only by subcutaneousadministration of corpuscular staphylococcal vaccinewhich contains 15 Staphylococcus aureus strains (“Can-tacuzino” National Institute of Research - Developmentfor Microbiology and Immunology – commercial pro-duct VACCIN STAFILOCOCIC D). The microbiologicaland immunological analysis were performed before andone week after vaccination.
Specimen collectionSamples from periodontal pockets were collected induplicate from each patient using the bacteriologicalloop and, respectively, cotton pieces, specially prepa-red for being introduced into the pocket using a stoma-tological needle. Peripheral blood was aseptically har-vested on empty vacutainers to obtain serum and in va-cutainers with anticoagulant (heparin and NA-EDTA) forcell isolation.
Bacterial identificationSmears from collected samples were Gram stained.Identification and isolation of Staphylococcus spp. wasperformed by standard microbiology procedures con-sisting in cultivation on blood agar plates and on selec-tive media (7.5 % NaCl Chapmann type agar and broth).Biochemical and enzymatic identification was doneusing catalase test for differentiation of Micrococacceaefrom Streptococaceae (positive for Micrococacceae).S. aureus has been differentiated from other Staphylo-coccus spp. based on the coagulase test. API STAPHgallery and Analytic API STAPH Catalogue helped forfurther differentiation of Staphylococus species (2, 3,5, 6, 7).
Anaerobic bacteria were cultivated in anaerobicconditions (regenerated thioglycolic VF broth, cultiva-
tion in appropriate atmosphere on Nagler agar, bloodagar with 3% bile and antibiotics: Kanamycin 1000mg/mL, Neomycin 25 mg/mL, Polymixin 100 U/mL).The strictly anaerobic bacteria were biochemicallyidentified using the API 20 A gallery (3, 15, 24).
Mancini radial immunodiffusion was done for thedetection of CRP, IgA, IgG, IgM from patients serumusing Cantacuzino Institute commercial immunodiffu-sion microplates. Incubation time and samples dilutionusing the manufacturer protocol are presented in Table1. The diameters of the precipitation rings were meas-ured in mm and the conversion in mg/dL or IU/mL wasperformed as in the kit instructions.
PMN granulocytes isolationThe cells were isolated from heparinized venous PB ofthe donors by one step centrifugation on a mixture ofFicoll-Hipaque for 20 min at 400 x g.
The cell pellet was recovered, the red cells wereeliminated by NH4Cl lysis and the PMNs were washedtwice in Hank’s balanced salt solution (HBSS), pH 7.2and resuspended to a final number of 2 x 106 cells/mL.Cell viability was always greater than 97%, as deter-mined by tripan blue exclusion test.
Chemiluminescence assayTo detect reactive oxygen species (ROS) , a luminol-en-hanced assay was performed and ROS released levelswere automatically measured for 30 minutes with LKBBio Orbit 1251 luminometer, using Multiuse-Phago-cytosis test. The instrument was set to measure the in-tensity of light emission from each sample at 2 minu-tes intervals during an entry assay period of 30 minu-tes. The standard reaction mixture in the cuvettes con-sisted of 50 mL of 10-4 M luminol, 500 mL of cell sus-pension (2 x 106 PMN/mL), addition of either 100 mLopsonized zymosan (OZ) (10 mg/mL) or 100 mL Con-canavalin A (ConA) (0,1 mg/mL) and completed to 1 mLwith HBSS. The results are expressed as percent stimu-lation index (SI %) calculated by the following formu-la: maximum light intensity registered to the stimulatedsamples per maximum light intensity registered to un-stimulated samples x 100. The following protocol wasused: 1) unstimulated PMN granulocytes, 2) PMN stimu-lated with OZ, 3) PMN stimulated with ConA (19, 21).
12
STAVARU et al.
Table 1 - The incubation time and samples dilution
Protein Reference serum Samples Incubation time Dilution Dilution at 37oC (hours)
Ig G 1/4 SUR ¼ 24 Ig A 1/4 SUR ¼ 24 Ig M 1/2 SUR ½ 48 Complement C3 1/2 SUR ½ 24CRP CRP undiluted Undiluted 24

Flow cytometryEDTA peripheral blood cells were stained with the mo-noclonal antibodies from Simultest IMK-Plus kit (Bec-ton-Dickinson): CD45-FITC/CD14-PE, IgG1-FITC /IgG2a-PE, CD3-FITC/CD19-PE, CD 4-FITC/CD8-PE,CD3-FITC/ anti-HLA-DR-PE, CD3-FITC/ CD16-56-PE.Analyses were done on FACSCalibur (Becton-Dickin-son, San-Jose USA) using automatic analysis SimulSETsoftware. Cells were first gated according to their scattercharacteristics and then analyzed for the presence ofcell surface antigen (FITC on FL1, PE on FL2) (20).
RESULTS
Bacteriological investigation of the specimens collec-ted from the periodontal pockets before and after sta-phylococcal vaccine administration. The results showedno differences between the two methods used for col-lecting pocket samples. Bacterial strains isolated fromthe periodontal pocket before and after vaccine thera-py are presented in Table 2.
Periodontal pocket sample resulted in a positiveculture but in a single case. Most of patients (57%) pre-
sented two bacterial agents either strictly anaerobic orin association with aerobic gram positive cocci. In ninepatients (64%) Staphylococcus spp was present aloneor in association with anaerobic spp. Staphylococcusaureus was identified in two subjects and one subjectwas positive for Streptococcus viridans. In 9 patients(64%) anaerobic spp only were isolated before vaccineadministration or anaerobic species in association withfacultative anaerobic spp. After staphylococcal vaccineadministration the reduction of microbial flora fromthe periodontal pockets with total absence of bacterialgrowth in five patients were noticed (33%). Staphylo-coccus aureus was not isolated after corpuscular sta-phylococcal vaccine therapy. In six patients (40%) themicrobial isolated species spectrum was not modifiedafter therapy.
Serum levels of CRP, C3 complement subunit, IgM,IgG and IgA before and after staphylococcal vaccinetherapy are shown in Table 3. Serum concentration ofCRP protein was inferior to 1mg/dL in all determina-tions. In one patient, Complement C3 subunit was overthe normal values and persisted so after vaccination.
Microbiological and immunological study of staphylococcus vaccine effects in periodontitis
13
Table 2 - The bacterial strains isolated from periodontal pocket before and after treatment

Serum concentration of Ig M and Ig G immunoglobu-lins ranged within normal values before and after the-rapy in all investigated patients. In one patient, IgA se-rum level was high before vaccination and normalizedafter. In this patient, the anaerobic flora detected be-fore vaccination was no more isolated after but this se-cond time the culture was positive for Staphylococcusepidermidis.
PMN granulocytes ROS released in vitro after OZand ConA stimulation evaluated before and after ad-ministration of staphylococcal vaccine D as percent ofstimulation index (SI %). Before vaccination, the resultsshowed a decreased average level of ROS released bypatients PMN granulocytes after OZ stimulation to Fc-receptors compared with the control, with no statisticsignificance (p<0.08). Instead, before vaccination theROS generated by PMN granulocytes after lectine sti-
mulation with ConA showed a statistical significativeincreased value of SI % (p<0.015) compared with thecontrol. After staphylococcal vaccine therapy, the PMNrespiratory burst in patients was significantly decrea-sed (OZ p<0.03, ConA p<0.002) compared to theprevious values. These data are shown in Fig. 1.
Lymphocytes sets and subsets from patient’s pe-ripheral blood detected by flow cytometry before andafter staphylococcal vaccine therapy. The results eva-luated as percent average or absolute counts did notshow any significant difference related to the momentof vaccination and they were within the normal rangeof values. The presence of NKT cells in both determi-nation was demonstrated in 6 patients. The ratio ofCD4+/CD8+ T lymphocytes was over 1. These resultsare shown in Table 4.
14
STAVARU et al.
Tabelul 3 - The humoral immune parameters investigated before and after therapy at the patients with periodontitis

DISCUSSION
Periodontitis is a chronic inflammation caused bybacterial infection and its therapy is frequently associa-ted with resistance to antibiotics. An alternative thera-peutic method is vaccination with corpuscular staphy-lococcal vaccine D, which contains 15 selected strainsof Staphylococcus aureus. Previous clinical studies su-gested the efficacy of this therapeutic approach underan original therapeutic method (10,11,13).
In most studied patients, the microbiological inves-tigation performed from the periodontal pocket beforevaccination detected two microbial agents, both strictlyanaerobs or one anaerobe in association with aerobs. Asa peculiarity, a high frequency of coagulase-negativeStaphylococcus spp was noticed and in two patientsthe cultures were positive for S aureus. The anaerobeflora was also identified in most of the patients andbelongged to Fusobacterium, Bacteroides, Bifidobacte-rium, Porphyromonas, Peptostreptococcus, Veillonellaspp., which have been described in the literature asmicrobial agents most frequenly involved in periodon-tal diseases (23). From Gram positive cocci, we foundStreptococcus viridans. Our data revealed an increa-sed frequency of Staphylococcus spp. in the investiga-ted patients, though previous study in the Romanianpopulation described as more frequent Porphyromo-nas gingivalis, Prevotella intermedia and Fusobacte-rium nucleatum (1).
After subcutaneous administration of staphylococ-cal vaccine, the bacteriological investigation resultsshowed the reduction of microorganisms isolated fromperiodontal pocket. S. aureus was no more isolated andin 33% of the patients no previous flora was detected.Nevertheless, in 40% patients the initial detected florawas identified again after vaccination (Table 2).
CRP acute inflammation marker was inferior to 1mg/dL in all subjects. The C3 complement fraction wasin the normal range of values but in one subject, withno difference before or after the vaccination. At thesame time, the serum levels of IgM, IgG and IgA (ex-cept in one patient before vaccination) were in the nor-mal range of values without significant modificationafter the vaccination. The patient presenting increasedvalue of IgA before vaccination was detected with twoanaerobic bacteria before vaccination, which were nomore isolated after vaccination. This patient was posi-tive for Staphylococcus epidermidis after vaccination(Table 3).
The respiratory burst of PMN granulocytes stimu-lated with OZ showed a statistically significant decreaseafter vaccination compared with values obtainedbefore (p<0.003). The average of ROS released afterOZ stimulation of the PMN granulocytes before vac-cine therapy was less in the patients group than in thecontrol group (p<0.008) as shown in Fig. 1. The ROS
Microbiological and immunological study of staphylococcus vaccine effects in periodontitis
15
Table 4 - Immunophenotype of lymphocytes sets and subsets from peripheral blood before and after vaccine therapy
Fig. 1 - The PMN respiratory burst after in vitro stimulated with OZ or ConA before and after vaccination

released by patients PMN after in vitro stimulation withConA before therapy was statistical significant increa-sed compared with the control group (p<0.0015) andsignificantly decreased after vaccination (p<0,002) a-pproaching the values obtained in the control group(Fig. 1). These results suggest the existence of PMN Fcreceptors dysfunction in patients with periodontitis withan increased susceptibility to ConA lectine stimulation.
The immunophenotype of lymphocytes sets andsubsets from peripheral blood performed before andafter staphylococcal vaccine D therapy showed no sig-nificant differencies (Tabel 4). The CD4+/CD8+ cellsratio was under 1, leading us to concluding that there is nosupressed immune response in these patients. We noticedthe presence of NKT cells in 6 investigated patients.
Our findings suggest the immunomodulatory effectof staphylococcal vaccine D therapy in periodontitis.and Staphylococcus spp. to be more frequently impli-cated in Romania as microbial agents of periodontitis.
ACKNOWLEDGEMENT
This work was supported by the Ministry of Educa-tion and Research grant VIASAN 333.
REFERENCES
ALI R.W., VELCESCU C., JIVANESCU M.C., LOFTHUS B., SKAUG N. -Prevalence of 6 putative periodontal pathogens in sub-gingival plaque samples from Romanian adult periodon-titis patients, J. Clin. Periodontol., 23 (2), 133-139, 1996
Baird–Parker A.C. - The Staphylococci: In introduction. J.Appl. Bacteriol. Symposium Supplement, pp. 1s – 8s, 1990
BUIUC D., NEGUT M. - Tratat de microbiologie clinica (Cli-nical Microbiology), Ed. Medicala 1999
CHUNG WHASUN O., DALE BEVERLY A. - Innate Immune Res-ponse of Oral and Foreskin Keratinocytes: Utilization ofDifferent Signaling Pathways by Various BacterialSpecies. Infect Immun., 72(1), 352–358, 2004
CODITA I. - The genus Staphylococcus. Bacteriol. Virusol.Parazitol. Epidemiol 38, 5-32, 1993
CODITA I. DOROBAT O. - An update on the antibiotic resis-tance of staphylococci, Bacteriol. Virusol. Parazitol. Epi-demiol. 43 (1-2), 69-79, 1998.
CODITA I., DUMITRESCU V. - Possible taxonomic descriptionof coagulase-negative staphylococci isolated in infec-tious skin processes. Bacteriol. Virusol. Parazitol. Epide-miol. 33 (2), 147-153, 1988
DAHLEN G. - Role of suspected periodontopathogens inmicrobiological monitoring periodontitis. Adv. Dent.Res, 2, 163-174, 1993
EDLUND C., HEDBERG M., NORD C.E. - Antimicrobial treat-ment of periodontal diseases disturbs the human ecolo-gy: a review., J. Chemoter. 8 (5), 331-341, 1996
GAFAR M., GEORGESCU T., DUMITRIU H., CODITA I., NEGUT M. -The role of the staphylococcal infectious-allergic pro-cess in chronic marginal periodontal diseases, and spe-cific therapeutic possibilities with staphylococcal vaccine.Preliminary note., Rev.Chir. Oncol. ORL Oftalmol. Sto-matol. Ser. Stomatol., 34,(1), 1-10, 1987
GEORGESCU T - Modularea rãspunsului imun cu vaccin sta-filococic în boala parodontal?, (Immunomodulation ofperiodontitis by staphylococcic vaccine) Ed. Univ.Piteºti, 2004
GRENIER D., MICHAUD J. - Demonstration of human immu-noglobulin G Fc-binding activity in oral bacteria. ClinDiagn Lab Immunol., 1(2), 247–249, 1994
LAKI D., GEORGESCU T., CODITA I. - Anatomoclinical and bac-teriological study on chronic marginal periodontal disease.Therapy with staphylococcic vaccine., Rom J. Morphol.Embryol., 42 (3-4), 169-178, 1996
MARCOTTE HAROLD, LAVOIE MARC C. - Oral Microbial Ecolo-gy and the Role of Salivary Immunoglobulin A. Micro-biol Mol Biol Rev., 62(1): 71–109, 1998
Methode de laboratoire Bacteries anaerobies et leur iden-tification, 1997
MOORE W. E., HOLDEMAN L. V., CATO E. P., SMIBERT R. M.,BURMEISTER J. A., RANNEY R. R. - Bacteriology of moderate(chronic) periodontitis in mature adult humans. InfectImmun., 42(2): 510–515, 1983
MOORE W. E., HOLDEMAN L. V., SMIBERT R. M., GOOD I. J.,BURMEISTER J. A., PALCANIS K. G., RANNEY R. R. - Bacte-riology of experimental gingivitis in young adult hu-mans. Infect Immun., 38(2): 651–667, 1982
NAIR S. P., MEGHJI S., WILSON M., REDDI K., WHITE P., HEN-DERSON B. - Bacterially induced bone destruction: mecha-nisms and misconceptions. Infect Immun., 64(7):2371–2380, 1996
OLINESCU A., DENISA MICHELE, ANGELA DOLGANIUC, RADU
D.L.- Variation of oxygen free radicals release in vitro byhuman polymorphonuclear granulocytes (PMN).Physiology, 21, 10-19, 1999
OLINESCU A., RADU D.L., ANGELA DOLGANIUC - Immuno-logical studies by phenotyping tests and functional ana-lysis at the cellular level in populations of children from2 different communities. Bacteriol. Virusol. Parazitol.Epidemiol., 42, 225-228, 1997
OLINESCU R., RADU D.L., PINTEA G., MARIA MILITARU - Evalua-tion of the therapeutic efficiency of Voltarene by usingthe in vitro chemiluminescence emitted by polymorpho-nuclear leukocytes. Rom. J. Intern. Med., 29, 199-204,1991
PAGE R.C. - Vaccination and periodontitis: myth or reality.J. Int. Acad. Periodontol., 2, 31-43, 2000
PASTER B.J., BOCHESS.K., GALVIN J.L., ERICSON R.E., LAU C.N.,LEVANOS V.L., SAHASRABUDHE A. DEWHIRST F.L. - Bacterialdiversity in human subgingival plaque J. of Bacteriol,183 (12), 3770-3782, 2001
WADWORTH ANAEROBIC BACTERIOLOGY MANUAL - fifth edition,1993
YAMAZAKI K., OHSAWA Y., TABETA K., ITO H., UEKI K., ODA T.,YOSHIE H., SEYMOUR J.G. - Accumulation of human heatshock protein 60-reactive T cells in the gingival tissues ofperiodontitis patients. Inf. Immun., 70, 2492-2501, 2002
16
STAVARU et al.

Rheumatoid arthritis (RA) is an invalidating chronicinflammatory disease with still elusive pathogenic me-chanisms, characterized by progressive destruction ofarticular cartilage and surrounding tissues in multiplejoints. Recruitment of immune cells into the synovialmembrane, hyperplasia of synovial lining cells, a shiftin the phenotype and function of synovial fibroblastsare accepted as pathological coordinated events in RA(FIRESTEIN, 1997).
Lymphocyte accumulation in the synovial sublin-ing tissue may be the result of increased cellular migra-tion from blood, proliferation induced by an unknownantigen or inhibition of cell death (DUDLER et al, 1998;VANDERBORGHT et al, 2000; SCHIRMER et al, 1998; REPA-RON-SCHUJIT et al., X). Considerable evidence suggeststhat CD4+ T cells play an important role in the patho-genesis of RA that is not restrained to the recognitionof an arthritogenic antigen in the early phases of thedisease (FALTA, 1988).
Transendothelial migration of T-cells seems to bean intrinsic ability of certain subpopulations, such as
the CD4+ memory T cells biased towards a Th1 phe-notype (D’AMBROSIO et al, 2000). Virus-specific acti-vated CD8+ T cells were evidenced in synovium (TAN
et al, 2000), but conflicting evidence exists regardingvirus infection in the joint. Evidence exists about therole of CD8+ T lymphocytes in aggravating the patho-logical responses in RA synovitis via the cytokine net-work (SKAPENKO et al., 2005). Though a T cell-drivendisease, RA is critically dependent on B lymphocytes(EDWARDS et al, 2001); besides their role of arthrogenicimmunoglobulins secretors, B cells could provide helpfor T-cell activation or harbor the relevant antigen(TAKEMURA et al., 2001).
RA is a systemic disease with important life-threat-ening extra-articular symptoms (BAYRAKTAR et al., 2000)with a background of disease-associated peripheralimmune disorders that contribute to RA spreading. Ac-tivated T and B lymphocytes and memory cells weredetected in periphery, generated either as a conse-quence of cell traffic between blood and synovium orpersistent antigen challenge (NANKI et al., 2000).
17
ABSTRACTThe study is focused on the correlated peripheral cellular immune disorders registered in agroup of 23 patients with severe, progressive rheumatoid arthritis, on methotrexate therapy.We investigated a panel of peripheral immune parameters: leukocyte counts, the proportionsof lymphocyte populations (T, Thelper, Tcytotoxic/suppressor, B lymphocytes and NK cells)and the polyclonal activation of lymphocytes. Results show that leukocytosis is due to simul-taneously elevated values of monocytes, granulocytes and, to a lesser extent, lymphocytes. Theregistered high values of the Th to Tc/s ratio are mainly attributed to the abnormal low pro-portions of the Tc/s subpopulation. Inverse correlations were emphasized between B, Tc/s lym-phocytes and NK cells or granulocytes. The unbalance of the lymphocyte to monocyte ratio orof the Th to Tc/s ratio does not impair the polyclonal activation of lymphocytes. In conclusion,we have characterized different patterns of correlated cellular peripheral immune disorders inrheumatoid arthritis, associated to pathological processes in conjunction with the immunsup-pressive and anti-inflammatory action of methotrexate that might be relevant for further inves-tigation of disease and further therapy outcome. We emphasize the special relation betweenthe adaptive and innate immune system at the level of cell counts and proportions. The corre-lations between the peripheral abnormalities in the rheumatoid arthritis group are better high-lighted by analyzing subgroups of patients characterized by particular values of the investigat-ed parameters.
PATTERNS OF PERIPHERAL CELLULAR IMMUNE DISORDERS IN SEVERE RHEUMATOID ARTHRITIS
Gina Manda1*, Monica Neagu1, Carolina Constantin1, Alina Rãdulescu2, Cãtãlin Codreanu2
(1Department of Immunology, “Victor Babeº” National Institute, Bucharest, Romania, 2Rheumatic Diseases Center “Dr. Ion Stoia”, Bucharest, Romania)
*Address for correspondence: Gina Manda, Immunology Department, "Victor Babes" National Institute, Splaiul Independentei 99-101, 050096 Bucharest, Romania. Tel/fax: 40-21-4115105, e-mail: [email protected]

Our study is focused on patients with severe, pro-gressive RA treated with methotrexate, candidates forleflunomide therapy due to severe side-effects of me-thotrexate. In order to characterize the peripheral immunedisorders of these patients prior to novel therapy onset,several cellular immune parameters were investigatedin a correlating manner. Identification and characteri-zation of the patterns of peripheral immune distur-bances is a prerequisite step for the follow-up studiesregarding therapy efficiency.
MATERIALS AND METHODS
Reagents. Histopaque, phytohemagglutinin M (PHA),RPMI 1640 medium, fetal bovine serum, antibiotic-an-timycotic solution, POPOP, PPO were purchased fromSigma-Aldrich. SimultestTM IMK-Lymphocyte Kit waspurchased from Becton Dickinson. Tritium-labeled uri-dine [3H-Urd] was obtained from the Institute of Phy-sics and Nuclear Engineering “Horia Hulubei”, Ma-gurele, Romania.
Patients. We investigated a group of 23 outpatients withRA (RA patients), 21 females and 2 males, with a meanage of (55±3) years. Patients fulfilled the revised ARAcriteria (ARNETT et al., 1988), were positive for rheuma-toid factor and were placed in the functional class IIIdefined by STEINBROKER. They had a longstanding, se-vere, active disease (Table 1) whose progression couldnot be controlled by conventional therapy with gluco-corticoids, gold salts, anti-malarial drugs, sulfasalazineand finally methotrexate. The therapy with methotre-xate showed a poor benefit to side-effects ratio (mainlygastric disorders were registered). Patients did not re-ceive glucocorticoid therapy at least two months be-fore entering the study.
Normal values for the investigated parameters wereestablished in a group of age-matched healthy volun-teers that presented no clinical records or signs of in-flammation, infection or autoimmune disorders.
Isolation of peripheral leukocytes. Blood was collec-ted by venipuncture in sterile heparin-coated vials. Mo-nonuclear cells and granulocytes were isolated fromperipheral blood by density gradient centrifugation(BOYUM, 1968) using Histopaque (specific gravity 1.077).Red blood cells were lysed with a solution containing0.83% NH4Cl, 0.084% NaHCO3 (LIVESCU et al., 2003).Cellular viability, estimated by the trypan blue test,exceeded 98%. Differential counts of lymphocytes, mo-nocytes and granulocytes were performed accordingto their morphology (cellular size, nucleus shape) in1% acetic acid.
Immunophenotypyng of peripheral lymhocytes. Bloodwas collected by venipuncture in EDTA-coated vials.
Phenotyping was performed by flow cytometry(FACScan, Becton Dickinson) using the SimultestTM
IMK-Lymphocyte kit. Data acquisition and processingwere performed with the SimulSet software. We evalua-ted the percentage of the following cell populations inperipheral blood: T lymphocytes (CD3+), T CD4+ (Th)and T CD8+ (Tc/s) lymphocytes, B lymphocytes (CD19+)and NK cells (CD16+ CD56+).
Lymphocyte activation. Lymphocyte activation of iso-lated cells, nonstimulated or polyclonally activated invitro with PHA, was measured by the [3H-Urd] incorpo-ration test (SAXON, 1992). Briefly, test samples (200 mLfinal volume) containing 0.2 x 106 lymphocytes wereincubated in absence or presence of 10 µg/mL PHA for72 hrs at 370C in 5% CO2 atmosphere; 18 hrs prior toharvesting, cultures were labeled with 1µCi [3H-Urd] /sample. Cultures were harvested and were further mea-sured for radioactivity with a b-counter (Packard) inscintillation liquid [4 g PPO, 50 mg POPOP in 1 L of amixture of 60% toluene and 40% ethanol (v/v)]. Resultswere expressed as pulses/min (ppm). PHA stimulationindex was calculated as the response of PHA-stimulat-ed cells versus the response of unstimulated cells.
Statistics. Results were expressed as mean value ± stan-dard error of the mean (SEM). Statistical comparisonbetween groups/subgroups of patients and controlswere performed by the t-test considering unequal vari-ances. Statistical comparison between parameters withina group/subgroup was performed by the t-test for pairedsamples. Statistical significant differences were consi-dered when p<0.05. Correlation was established bythe Pearson correlation test and expressed as correla-tion coefficient (r).
RESULTS
The study was focused on a group of RA patientswith longstanding, severe, active disease (Table 1) thatcould not be controlled by conventional anti-rheumatictherapy with methotrexate, mainly due to severe side-effects. These patients were selected for further thera-py with leflunomide, a unique disease modifying anti-rheumatic drug with immunomodulatory properties(BREEDVELD et al., 2000).
We investigated in a correlative manner a panel ofperipheral cellular immune parameters of RA patients,described in Table 2. This investigation is a prerequi-site step for further follow-up studies regarding the po-tential immunomodulatory action and/or the immuno-toxicity of leflunomide and for identifying the potentialcellular targets of the drug (FRANSE et al., 2002).
RA patients presented moderate lymphocytosis par-alleled by markedly elevated counts of monocytes
18
MANDA et al.

(r=0.93) leading to low values of the lymphocyte tomonocyte ratio (Table 2). Lymphocytosis was alsoaccompanied by granulocytosis, but parameters werenot correlated.
Statistical analysis of the T lymphocytes subsets indi-cated that RA patients presented normal T cells percen-tage, but elevated proportions of the Th subset and ashift towards lower values of the Tc/s percentage wereregistered (Table 2). The obviously high values of theTh to Tc/s ratio were mainly determined by the redu-ced proportion of Tc/s lymphocytes (r = - 0.86).
The activation of the peripheral lymphocytes iso-lated from RA patients was in the normal range forboth unstimulated and PHA-activated cells (Table 2).Results show that the mentioned disorders at the levelof the lymphocyte to monocyte ratio and the Th to Tc/s
ratio did not affect the polyclonal activation of T lym-phocytes.
Individual data indicated a large heterogeneity ofthe cellular parameters range among the RA patients,that was not connected to disease severity (high num-ber of swollen and tender joints, prolonged morningstiffness or elevated values of CRP and ESR), but wasprobably determined by an inter-individual variabilityof responses to previous anti-rheumatic therapy (DANESI
et al., 2000; WOOD, 1999). We must also take into accountthat RA is not a single disease, but includes various dis-tinct variants (HOLMDAHI, 1999). Consequently, the cor-relations between the peripheral abnormalities in theRA group were better highlighted by analyzing sub-groups of patients characterized by particular values ofthe investigated parameters.
Patterns of peripheral cellular immune disorders in severe rheumatoid arthritis
19
Table 1 - Clinical and biological parameters of RA patients (n=23)
Table 2 - Peripheral cellular immune parameters of RA patients (n=23) with severe, progressive disease, treated with methotrexate.Statistical significant differences between patients and normal groups was considered when p<0,05; NS – not significant

When sorting RA patients according to the periph-eral lymphocyte counts (Fig. 1), data show that theleukocytosis registered in 14 out of 23 patients wasdue to simultaneously elevated lymphocyte, monocyteand granulocyte counts (p<0.05). The marked lym-phocytosis (p=0,017) registered in 6 out of 23 RApatients (Fig. 2) was associated with slightly higher Blymphocytes percentages (p=0.06) and moderatelylower NK values (p=0.09) leading to the disruption ofthe B-NK balance (B:NK=2,5±0,6 in the RA subgroup
versus the normal value 1.2±0.2, p<0.09). Lympho-cytosis or lymphopenia were not associated with animportant disturbance at the level of total T lymphocytesor T lymphocytes subpopulations (data not shown). Thelymphocytes isolated from RA patients presenting lym-phopenia [(0.51 ± 0.05)x106 lymphocytes/mL blood]were weakly responding in vitro to PHA stimulation(low stimulation index, p=0,01), while those from pa-tients with lymphocytosis developed normal cellularresponses (Fig. 3, Table 2).
20
MANDA et al.
Fig. 1 - Correlations between peripheral leukocytes populations in subgroups of RA patients (n=23), sorted according to theconcentration of peripheral lymphocytes. L=lymphocytes, Mo=monocytes, L:Mo=lymphocyte to monocyte ratio,PMN=granulocytes. Results are presented as mean ± SEM
Fig. 2 - The distribution of T, B lymphocytes and NK cells percentages in subgroups of RA patients (n=23) grouped accordingto the peripheral concentration of lymphocytes (see Fig. 1). Results are presented as mean ± SEM

We identified a subgroup of RA patients (15 out of23) with abnormal low values of the lymphocyte tomonocyte ratio (p<0.0003) (Fig. 4, Table 2) associatedwith elevated monocyte (p<0.0007) and moderatelyraised lymphocyte counts (p<0.03). These patients werecharacterized by significantly low Tc/s percentages(p<0.022) (Fig. 5, Table 2).
Most of the investigated RA patients (17 out of 23)were characterized by abnormally high values of theTh to Tc/s ratio (p=0.0003), determined by the down-re-gulation of the Tc/s subpopulation (p=0.005; r = - 0.80)(Fig. 6A). These patients showed peripheral disorders,
namely increased counts of lymphocytes (p=0.03),monocytes (p=0.006) and granulocytes (p=0.006)(Fig. 6A, Table 2), whereas T, B and NK proportionswere almost normal (Fig. 6B, Table 2).
The basal and PHA-induced activation of lympho-cytes isolated from RA patients was not dependent onthe Th to Tc/s ratio (data not shown), indicating thatthe significant high values of the Th to Tc/s ratio wereprobably due to the down-regulation of the cytotoxicsubpopulation, not the suppressor one.
We identified (Fig. 7) a small subgroup of RA pa-tients (8 out of 23) with extremely low values of the B
Patterns of peripheral cellular immune disorders in severe rheumatoid arthritis
21
Fig. 3 - The activation of peripheral lymphocytes isolated from 23 RA patients sorted according to the peripheral concentra-tion of lymphocytes (see Fig. 1). Lymphocyte activation was measured by the tritium-labeled uridine incorporation test.Results, expressed in radioactivity units (ppm) or as PHA stimulation index, are presented as mean ± SEM
Fig. 4 - Lymphocytes (L), monocytes (Mo) and granulocytes (PMN) concentrations in subgroups of RA patients (n=23), sortedaccording to the values of the lymphocyte to monocyte ratio (L:Mo). Results are presented as mean ± SEM

lymphocytes percentage (p=0.0004), accompanied byslightly higher values of NK cells, leading to an unba-lance of the B to NK ratio relative to normal values(p=0.003). Results indicate that a regulatory mecha-nism in the B-NK circuit might be active. Higher Blymphocytes proportions (15 out of 23 RA patients,p=0.06) were correlated (r=0.86) with low Tc/s per-centages (Fig. 7, Table 2). We also point out the nega-tive correlation between the percentage of B lympho-cytes and the granulocyte counts (r = - 0.95).
The marked granulocytosis (p=0.0002) registeredin a subgroup of RA patients (6 out of 23), was accom-panied by lympocytosis (p=0.03) and monocytosis(p=0.014) (Fig. 8A, Table 2). Reduced Tc/s proportions(p=0,03) and a related raise of the Th to Tc/s ratio(p=0.04) paralleled the moderate granulocytosis (Fig.8B, Table 2). A particular relation between granulocytecounts and the NK percentages was noticed (figure 8C);patients with normal or extremely high granulocytecounts had almost normal NK proportions, but the RAsubgroup with moderately high concentrations ofgranulocytes presented slightly elevated NK values(p=0.09).
CONCLUSION
We studied some aspects regarding the peripheralimmune status of RA patients with severe, refractory dis-ease, candidates for immunosuppressive therapy withleflunomide due to serious side-effects of methrotrexate.
The investigated RA patients presented heterogene-ity at the level of peripheral cellular parameters, thatwas not connected to disease severity. Consequently,
the analysis of subgroups of RA patients, sorted accor-ding to the magnitude of each peripheral cellular para-meter, rather than the statistical analysis of the total RAgroup, proved to be a valuable tool for identifying andcharacterizing the patterns of immune disorders in RApatients.
The study revealed various correlated immune dis-orders at the level of peripheral cell counts and pro-portions, but precaution must be taken when analysingthese parameters: the observed peripheral abnormali-ties might be the consequence either of impaired egressof cells from bone marrow or of transendothelial mi-gration of particular blood cells populations into theinflamed synovium (OPDENAKKER et al., 1998).
The elevated peripheral monocyte counts might havepathological consequences related to their migrationinto the synovium (ATHANSOU, 1995) in response tochemotactic signals delivered by synovial stromal cells(HAYASHIDA et al., 2001; GRISAR et al., 2004)). It is knownthat interaction with endothelial cells induces monocytedifferentiation in the inflamed tissue (SZAKANECZ et al.,2000) where they contribute to the pathogenesis of RAby functioning as antigen presenting cells (SZAKANECZ
et al., 2000) and by releasing inflammatory cytokines(EDWARDS, 1998).
Only a moderate lymphocytosis was statisticallyregistered in the investigated RA patients, showing anormal distribution of T and B lymphocytes. The mar-ked lymphocytosis registered in some RA patients wasassociated with a disruption of the B-NK balance infavour of B lymphocytes that might sustain pathologi-cal autoimmune processes. Previous results obtainedby us also showed a role of the B-NK circuits in RA
22
MANDA et al.
Fig. 5 - The correlations between the lymphocytes to monocytes ratio (L:Mo) and the percentage of Tc/s or the Th to Tc/s ratio(Th:Tc/s) in two subgroups of RA patients with distinct lymphocytes to monocytes ratio (see Fig. 4). Results are presented asmean ± SEM

(MANDA et al., 2003). Lymphocytes isolated from lym-phopenic patients were functionally depressed, possi-bly consequently to previous immunosuppressive the-rapy. Reduced proportion of peripheral T CD8+ lym-phocytes might be connected to their transendothelialmigration into the inflamed synovium, leading topathological processes.
The disturbance of the lymphocyte to monocyteratio due to high monocyte concentrations or the redu-ced peripheral Tc/s proportion have no major effect onthe polyclonal activation of lymphocytes, indicating thatthe cellular immune network is correctly functioning.
Although RA is a T cell-driven disease (SKAPENKO etal., 2005), no major abnormalities were recorded at
the level of the total T lymphocytes population, but animportant subgroup of patients was characterized byelevated values of the Th to Tc/s ratio determined mainlyby reduced Tc/s percentages. These data are in agree-ment with other reported observations (KURYLISZYN-MOSKAL, 1995), but, unlike these studies, the cellularfunctional experiments performed by us do not indica-te an activated state of peripheral lymphocytes. Proba-bly, the cytotoxic and not the suppressor subpopula-tion is down-regulated, leading to impaired antiviraldefence mechanisms and to disease progression (TAKEI
et al., 2001). Low Tc/s proportions are associated withhigh monocytes concentrations and, in some cases, withelevated NK percentages, that probably tend to com-
Patterns of peripheral cellular immune disorders in severe rheumatoid arthritis
23
Fig. 6 - Peripheral leukocytes distribution in subgroups of RA patients (n=23) sorted according the Th to Tc/s ratio (Th: Tc/s).A. Lymphocytes (L), monocytes (Mo) and granulocytes (PMN) counts. B. Percentages of T, Th, Tc/s, B lymphocytes and NKcells. Results are presented as mean ± SEM

24
MANDA et al.
Fig. 7 - The distribution of Tc/s lymphocytes and NK cells percentages in RA patients (n=23), sorted in ascending order of theB lymphocytes percentage (see Fig. 6). Results are presented as mean ± SEM
A. Correlations between peripheral leukocytes populations in subgroups of RA patients (n=23), grouped according to the concentration of peripheral granulocytes (PMN)
pensate the abnormalities of the Tc/s-mediated cyto-toxicity, at least at the level of cell proportion.
Particular inverse correlations between B lympho-cytes and NK cells were highlighted, with potential con-sequences on the production of rheumatoid factor (RF)by B lymphocytes, knowing that NK-like cells inducein vitro high levels of IgM RF synthesis in autologous Bcells (SANTIAGO-SCHWARTZ et al., 1992). Negative corre-lation between B lymphocytes percentage and PMN
counts was emphasized. We also showed that moder-ate granulocytosis is paralleled by reduced values ofthe T, Tc/s lymphocytes percentages and raised NKproportions.
These results point out an interrelation between theadaptive and innate immune system (MEDZITOV et al.,1997), at least at the level of cell counts and propor-tions, but further functional studies have to be per-formed in order to clarify these aspects. We highlight

especially the involvement of NK cells that react to thephysical and psychological stress to which the RApatients are submitted (MUKAI et al., 2000).
It is worth noticing that an unusual subset ofCD4+CD28- T lymphocytes, expressing NK receptors,exhibit autoreactivity and cytolytic functions(WARRINGTON et al., 2001), bridging specific and non-specific immune responses.
In conclusion, in a group of RA patients with se-vere, progressive, refractory disease we have identifieddifferent patterns of correlated peripheral cellularimmune disorders, associated to RA pathological pro-cesses in conjunction with the immunosuppressiveanti-inflamatory action of methotrexate. Results pointout disturbances at the level of cell counts and propor-tions and also particular correlation between theinnate and adaptive immune response.
Patterns of peripheral cellular immune disorders in severe rheumatoid arthritis
25
B. The distribution of T, Th, Tc/s lymphocytes percentages and the Th to Tc/s ratio in subgroups of RA patients, sorted according to the concentration of peripheral granulocytes (PMN)
C. The distribution of B lymphocytes and NK cells in subgroups of RA patients, sorted according to the concentration of peripheral granulocytes (PMN)
Fig. 8 - The distribution of leukocyte populations in subgroups of RA patients (n=23) sorted according to the concentrationof peripheral granulocytes (PMN). Results are presented as mean ± SEM

REFERENCES
ARNETT F.C., EDWORTHY S.M., BLOCH D.A, MCSHANED.J., FRIES J.F., COOPER N.S. et al. - The American Rheu-matism Association 1987 revised criteria for the classifica-tion of rheumatoid arthritis. Arthritis. Rheum., 31, 315, 1988
ATHANSOU N.A. - Synovial macrophages. Ann. Rheum. Dis.,54, 392, 1995.
BAYRAKTAR A., HUDSON S., WATSON A., FRASER S. - Phar-maceutical care (7) Arthritis, Pharmacol. J., 264, 57, 2000
BOYUM A. - Isolation of mononuclear cells and granulocytesfrom human blood. Scand. J. Clin. Lab. Invest., 21, 77, 1968
BREEDVELD F.C., DAYER J-M. - Leflunomide: mode of action inthe treatment of rheumatoid arthritis. Ann. Rheum. Dis., 59,841, 2000
D’AMBROSIO D., IELLEM A., COLANTONIO L., CLISSI B.,PARDI R. SINIGAGLIA F. - Localization of Th-cell subsets ininflammation: differential thresholds for extravasation ofTh1 and Th2 cells. Immunol. Today, 21: 183, 2000
DANESI R,. MOSCA M., BOGGI U., MOSCA F., DEL TACCAM. - Genetic of drug response to immunosuppressive treat-ment and prospects for personalized therapy. Mol. Med.Today, 6, 475, 2000
DUDLER J., KAI-LIK SO A. T cells and related cytokines. Curr.Op. Rheumatol., 10, 207, 1998
EDWARDS J.C.W. - The synovium. In Rheumathology. KlippelJ.H., Dieppe P.A., eds. London, Mosby, 6.1-6.8, 1998
EDWARDS J.C.W., CAMBRIDGE G. - Sustained improvementin rheumatoid arthritis following a protocol designed todeplete B lymphocytes. Rheumatol., 40, 205, 2001
FALTA M.T., KOTZIN B.L. – In T cells in Arthritis, Miossec P.,Vanden Berg W.B., Firestein G.S., Birkhauser eds., Basel,pp. 201-231, 1998
FIRESTEIN G.S. - Etiology and pathogenesis of rheumatoid ar-thritis. In Textbook of Rheumatology, eds. Kelley W.N., HarrisE.D. Jr, Rudy S., Sledge C.B., Philadelphia: WB Saunders,pp. 851-897, 1997
FRANSEN J., STUCKI, G., VAN RIEL P. - The merits of monito-ring: should we follow all our rheumatoid arthritis patientsin daily practice? Rheumatology, 41, 601, 2002
GRISAR J., ARINGER M., KOLLER M.D., STUMMVOLL G.H.,ESELBOCK D., ZWOLFER B., STEINER C.W., ZIERHUT B.WAGNER L., PIETSCHMANN P., SMOLEN J.S. - Lefluno-mide inhibits transendothelial migration of peripheral bloodmononuclear cells. Ann. Rheum. Dis., 63,1632, 2004
HAYASHIDA K., NANKI T., GIRSCHICK H., YAVUZ S., OCHIT., LIPDKY P.E. - Synovial stromal cells from rheumatoid ar-thritis patients attract monocytes by producing MCP-1 andIL-8. Arthritis Res., 3, 118, 2001
HOLMDAHI R. - Another pathway towards arthritis. Curr. Biol.,9: R528-R530, 1999.
KURYLISZYN-MOSKAL A. - Comparison of blood and synovialfluid lymphocyte subsets in rheumatoid arthritis and osteo-arthritis. Clin. Rheumatol., 14, 43, 1995
LIVESCU A., MANDA G., CONSTANTIN C., NEAGU M., IOR-DACHESCU D. - Plasma membrane potential interfers withthe respiratory burst of peripheral granulocytes, J. Cell. Mol.Med., 7(1), 73, 2003
MANDA G., NEAGU M., LIVESCU A., CONSTANTIN C., CO-DREANU C., RADULESCU A. - Immune consequences ofthe imbalance between peripheral B lymphocytes and NKcells in rheumatoid arthritis. J. Cell. Mol. Med., 7(1), 79, 2003
MEDZITOV R., JANEWAY JR C.A.- Innate immunity: impacton the adaptative immune response. Curr. Op. Immunol.,9, 4, 1997
MUKAI E., NAGASHIMA M., HIRANO, YOSHINO S. - Com-parative study of symptoms and neuroendocrine-immunenetwork mediator levels between rheumatoid arthritispatients and healthy subjects. Clin. Exp. Rheumathol., 18,585, 2000
NANKI T., LIPSKY P.E. - Cytokine, activation marker and che-mokine receptor expression by individual CD4+ memoryT cells in rheumatoid arthritis synovium. Arthritis Res., 2,415, 2000
NEAGU M., MANDA G., CONSTANTIN C., TANASEANU C.and MIKHAILIDIS D.P. The in vitro effect of a low molecularweight heparin, nadroparin, (Fraxiparine) on leukocytesobtained from patients with vascular disorders. Int. Angiol.,20: 164-173, 2001
OPDENAKKER G., FIBBE W.E., VAN DAMME J. - The molecu-lar basis of leukocytosis. Immunol. Today, 19, 182, 1998
REPARON-SCHUIJT C.C., VAN ESCH W. J.E., VAN KOOTEN C.,ROZIER B.C.D., LEVARHT E. W. N., BREEDWELD F.C.,VERWEIJ C.L. - Regulation of synovial B cell survival inrheumatoid arthritis by vascular cell adhesion molecule 1(CD106) expressed on fibroblast-like synoviocytes. ArthritisRheum., 43 (5):, 1115, 2000
SANTIAGO-SCHWARTZ F., KAY C., PANAGIOTOPOULOSC., CARSONS S.E. - Rheumatoid arthritis serum or synovialfluid and interleukin 2 abnormally expand natural killer-likecells that are potent stimulators of IgM rheumatoid factor. J.Rheumatol., 19, 223, 1992
SAXON A. - Functional B-cell studies. Chapter 63 In Manual ofClinical Laboratory Immunology, 4th ed. N.R. Rose, E.C. deMacario, J.L. Fahey, H. Freedman, G.M. Penn eds., Ameri-can Society for Microbiology, Washington DC, p.403, 1992
SCHIRMER M., VALLEJO A.N., WEYAND C.M., GORONZYJ.J. - Resistance to apoptosis and elevated expression of bcl-2 in clonally expanded CD4+CD28- T cells from rheuma-toid arthritis patients. J. Immunol., 161, 1018, 1998
SKAPENKO A., LEIPE J., LIPSKI E.P., SCHULZE-KOOPS H. – therole of T cells in autoimmune inflammation. Arthritis Res.Ther., 7 (Suppl. 2), S4, 2005
SZAKANECZ Z., KOCH A.E. - Cell-cell interactions in synovi-tis. Endothelial cells and immune cell migration. ArthritisRes., 2, 368, 2000
TAKEI M., ISHIWATA T., MITAMURA K., FUJIWARA S., SA-SAKI K., NISHI T., KUGA T., OOKUBO T., HORIE T., RYUJ., OHI H., SAWADA S. - Decreased expression of signalinglymphocytic-activation molecule-associated protein (SAP)transcripts in T cells from patients with rheumatoid arthritis.Int. Immunol., 13, 559, 2001
TAKEMURA S., KLIMIUK P.A., BRAUN A., GORONZY J.J.,WEYAND C.M. - T cell activation in rheumatoid synoviumis B cell dependent. J. Immunol., 167, 4710, 2001
TAN C.L., MORVAT A.G., FAZOU C., ROSTRIN T.,ROSKWELL H., DUNBAR P.R. et al. -Specificity of T cellsin synovial fluid: high frequencies of CD8+ T cells that arespecific for certain viral epitopes. Arthritis Res., 2, 154, 2000
VANDERBORGHT A., GENSENS P., VANDEVYVER C., RAUS G.,STINISSEN P. - Skewed T-cell receptor variable gene usagein the synovium of early and cronic rheumatoid arthritispatients and persistence of clonally expanded T cells in achronic patient. Rheumatol., 39, 1189, 2000
WARRINGTON K.J., TAKEMURA S., GORONZY J.J.,WEYAND C.M. - CD4+, CD28- T cells in rheumatoid arthritispatients combine features of the innate and adaptiveimmune systems. Arthritis. Rheum., 44, 13, 2001
WOOD J. - Rheumatoid arthritis: management with DMARDs.The Pharm. J., 263, 162, 1999
26
MANDA et al.

Experimental evidence indicates cytokine and neu-rotrophins production in brain tissue after stroke. Infla-mmatory mechanisms and immune activation havebeen hypothesized to play a role in the pathogenesis ofcerebral ischemia. Stroke could modulate peripheralneurotrophic factors levels.
Neurotrophic factors are endogenous moleculeswhich have a crucial role in maintenance, survival anddifferentiation of various neuronal population withinthe developing and adult brain. Also it could play a rolein repair-regeneration process such as axonal regene-ration, neuronal plasticity and neurogenesis, in fact pro-tection against degeneration and lessions, that could becritical for recovery after ischemic stroke.
Neurotrophins are peptides that act as growth orsurvival factors for specific neuronal population. Thefamily of neurotrophins includes Nerve growth factor(NGF), Brain-derived neurotrophic factor (BDNF),Neurotrophin-3 (NT-3) and Neurotrophin-4/5. They de-rive from a common ancestral gene and are similar in
sequence and structure. Six region of amino acid non-identity confer specificity to each neurotrophin. All fourmammalian neurotrophins are produced like precur-sors that are cleaved by convertases in order to gene-rate mature processed neurotrophins of 118-120 aminoacids that associate as dimmers.
NT-3 is a 119 amino acids protein with 57% ho-mology to NGF and 58% homology to BDNF. The bio-logical activity and its expression characteristics of NT-3 are quite different from NGF and BDNF. In contrastto these neurotrophins, the level of NT-3 mRNA in theCNS is high during fetal development and reduced inthe adult brain. It is actively transported retrograde,along axons, being predominantly expressed in thehippocampus and basal forebrain, mid-brain, cerebel-lum and spinal cord, in both neurons and glia. Theabsence of the NT-3 gene leads to a defect in the sen-sory nervous system, but it may be complemented byother neurotrophins in the motor nervous system duringdevelopment.
27
ABSTRACTPro-inflammatory cytokines and neurotrophins in the central nervous system (CNS) have beenrecognized as mediators of both neurodegenerative and neuroprotective mechanisms in a num-ber of CNS pathologies. A rapid, sustained elevation of these molecules was recently reportedafter traumatic and ischemic brain injury. Inflammatory mechanisms and immune activationhave been hypothesized to play a role in the pathogenesis of cerebral ischemia. Stroke is thethird largest cause of death next to heart disease and cancer in the world, and it is an impor-tant cause of death and disability in developed countries. Role of excitatory amino acids recep-tors activation, calcium overload, nitric oxide and oxidative stress in the pathogenesis ofischemic brain damage is well established. Stroke may modulate peripheral neurotrophic factorslevels. In experimental animal models, neurotrophin-3 (NT-3) has been shown to be producedby glial cells as an adaptability response to hypoxia. In spite of substantial research and significant number of neuroprotective drugs that have beendeveloped to limit ischemic brain damage and to improve the outcome for stroke patients, nospecific therapy for stroke is available. The neurotrophins have been proposed as therapeuticagents for the treatment of neurodegenerative disorders and ischemic injury.In the present work, we investigated the possible correlation of NT-3 with tumor necrosis fac-tor-alpha (TNF-α) and interleukin-6 (IL-6) in the serum and cerebrospinal fluid (CSF) frompatients with ischemic stroke (IS).
NEUROTROPHIN-3, TNF-α AND IL-6 RELATIONS IN SERUM AND CEREBROSPINAL FLUID OF ISCHEMIC STROKE PATIENTS
Daniela Pãsãricã1, Mihaela Gheorghiu1, Florica Topârceanu2, Coralia Bleotu2, Loretta Ichim2 and T. Trandafir1
(1"Carol Davila" University of Medicine and Pharmacy, Pathophysiology and Immunology Department; 2"St. S.Nicolau" Institute of Virology, Bucharest, Romania)

The action of NT-3 is dictated by two classes of cellsurface receptors: Trk C receptor tyrosine kinase andp75 neurotrophin receptor (p75NTR) a member of theTNF receptors superfamily. p75 and Trk receptors havebeen described as low and high affinity receptors. NT-3and NT-4/5 can also binds weakly to TrkA. The do-mains in the Trk receptors that determine specificity ofbinding are immunoglobulin-C2 domains. There areno sequence similarities between Trk and p75 recep-tors in either ligand binding or cytoplasmic domainsThe majority of neurotrophin-3 responsive cells ex-presses both p75 and TrkC. Expression of p75 increas-es the binding affinity of TrkC for NT-3, provides morespecificity and promotes survival in low concentrationof neurotrophins. In the absence of TrkC, or under stressconditions, p75 is frequently up regulated. Althoughp75 and TrkC receptors are coexpressed in many cells,independent expression of p75 and of individual Trkfamily members are also observed. p75 receptor is ca-pable to initiate a cell death program in selected cellsand to activate the ceramide production, NFkB or JNKpathways. The decision between survival and deathamong NT-3 responsive neurons is determined by theratio of p75 to Trk receptors and their individual sig-naling properties. Hence, neurotrophin signaling throughp75 is critical in regulating both survival and apoptoticevents
Binding of NT-3 to TrkC receptors causes receptordimerization and autophosphorylation on tyrosineresidues within the intracellular domain that activatesmultiple signal transduction pathways. The biologicalresponses are produced rapidly. It regulate cell growthand survival through Ras – MAP kinase (MAPK), phos-phatidylinositol 3-kinase (PI3-K) and phospholipase C(PLC). These intracellular second messengers trans-duce the signal, through cytoplasmatic and intranu-clear proteins in order to the gene expression apoptoticproteins are inhibited and downregulated, while anti-apoptotic proteins are activated or over expressed. Neu-rotrophins have been shown to attenuate both neu-ronal and glial apoptosis.
PATIENTS AND METHODS
Twenty patients aged between 38-93 years (72.65 ±12.81 years) were investigated after computed tomo-graphy confirmed IS. All the studied patients were hos-pitalized in the Cerebrovascular Disease Department“V. Voiculescu”, Bucharest, Romania. No patient wasstudied without prior consent signed by appropriaterelative.The patient outcomes were assessed 1 monthafter IS using the Glasgow outcome score (GOS). GOS5 = good recovery, 4 = moderate disability, 3 = severedisability, 2 = vegetative state, and 1 = death.
NT-3, TNF-alpha and IL-6 were determined in se-rum and CSF samples using venous puncture and lum-bar puncture, respectively.The CSF and serum weresubmitted for biochemical analysis (serum C-reactiveprotein concentration, cellularity and glucose-proteinconcentration in the CSF). All CSF and serum sampleswere centrifuged for 10 min at 1000 rpm, and super-natant was immediately stored at -70 oC until analysis.Serum and CSF samples were analyzed for NT-3, TNF-alpha and IL-6 using Coulter and EuroClone ELISA kits.
All the results were expressed as mean and stan-dard deviation. Statistical difference between meanswere determined using Student’s t test where a p valueunder 0.05 was significant. The Pearson correlationcoefficient was calculated too considering significant avalue of r > 0.5.
RESULTS
TNF-αα was detected in CSF and serum in only threepatients with GOS 1 and one patient with GOS 4(7.75±0.5 days after IS).
The patients were divided into 3 groups based ontheir IL-6 and NT-3 serum and CSF level.
- GROUP I (6.4 ± 3.4 days after IS) with low/mod-erate serum concentration of IL-6 (<40 pg/ml)and high/exacerbated CSF level of IL-6 (>76pg/ml) or inversely [GOS were 1 (n=5), 2 (n=2),3 (n=2)];
- GROUP II (10 ± 5.8 days after IS with high con-centration of IL-6 (>72 pg/ml) or NT-3 (>17pg/ml) in serum and high level of IL-6 (>50pg/ml) in CSF [GOS were 1 (n=2), 2 (n=1), 5(n=2)];
- GROUP III (14.2 ± 6.8 days after IS) with low /moderate concentration of IL-6 (<45 pg/ml) andNT-3 (<15 pg/ml) in serum and CSF [GOS were3 (n=1), 4 (n=3), 5 (n=2)].
There was a negative correlation between NT-3 andIL-6 in serum from survivors with GOS 2-5 (r = - 0.916,p = 0.05) in group I, (r = - 0.98, p = 0.003) in group II,(r = -0.814, p = 0.05) in group III and in CSF from sur-vivors with GOS 3-5 (r = - 0.775, p = 0.05) in group III.
On the contrary, there was a positive correlationbetween NT-3 and IL-6 in serum from non-survivors ingroup I (r = 0.88, p = 0.05). Non-survivors of group IIexhibited exacerbated serum levels of TNF-α (141-158pg/ml).
28
PÃSÃRICÃ et al.

The dinamic of Il-6 and NT-3 level in serum and CSF in patients with IS
Neurotrophin-3, TNF-αα and IL-6 relations in serum and cerebrospinal fluid of ischemic stroke patients
29
Table 1 - The groups of patients divided on their serum and CSF levels of IL- 6 and NT-3
Table 3 - The dinamic of IL-6 serum level in patients with IS
Table 2 - The level of TNF-α at patients with IS

30
PÃSÃRICÃ et al.
Table 4 - The dinamic of NT-3 serum level in patients with IS
Table 5 -The dinamic of IL-6 CSF level in patients with IS
Table 6 - The dinamic of NT-3 CSF level in patients with IS
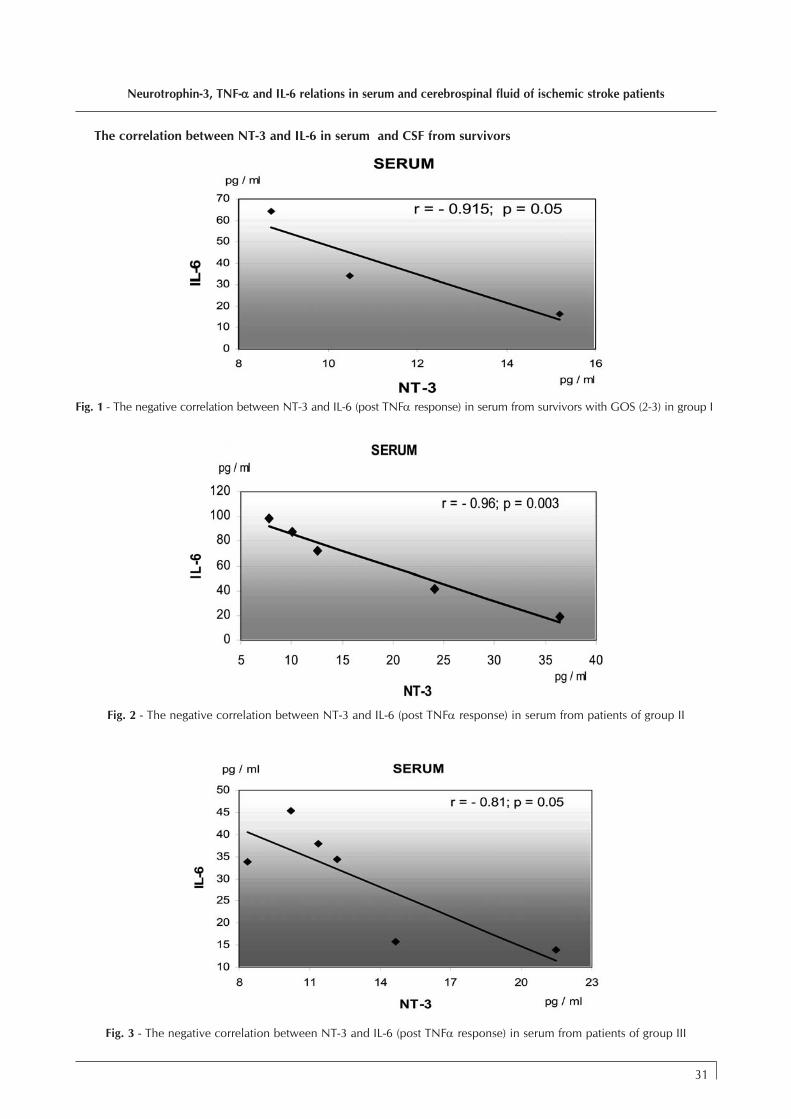
Neurotrophin-3, TNF-αα and IL-6 relations in serum and cerebrospinal fluid of ischemic stroke patients
31
The correlation between NT-3 and IL-6 in serum and CSF from survivors
Fig. 1 - The negative correlation between NT-3 and IL-6 (post TNFα response) in serum from survivors with GOS (2-3) in group I
Fig. 2 - The negative correlation between NT-3 and IL-6 (post TNFα response) in serum from patients of group II
Fig. 3 - The negative correlation between NT-3 and IL-6 (post TNFα response) in serum from patients of group III

CONCLUSION
The negative correlation between NT-3 and IL-6suggested that NT-3 could play a modulatory role onpost-TNF-alpha responses of IL-6 in patients with im-proved outcome after IS.
The absence of this correlation or presence of exa-cerbated serum levels of TNF-α corresponded to immi-nence of death.
DISCUSSION
This study demonstrates that increased the level of IL-6TNFα and NT-3 in CSF and serum of IS patients is notuniformly enhanced, suggesting that factors act diffe-
rent target cells and have different function in CNS.These elevation are result of enhanced central produc-tion of the cytokine since TNF are significantly higherin the first stage after IS. The exacerbated serum andCSF levels of this cytokine is present in principal to thenon-survivors. TNFα is synthesized de novo in neu-rons, activated microglia and macrophage after IS. Itexert a primary effect on microvascular inflammatoryresponse and induced neutrophin adhesion to brain ca-pillary endothelium. TNFα increases leukocyte adhe-rence to blood vessel and subsequent infiltration intothe brain. In addition TNFα induce the disruption ofthe blood-barrier, a phenomenon fundamental to IS.Several studies have shown that blocking TNFα im-
32
PÃSÃRICÃ et al.
Fig. 4 - The negative correlation between NT-3 and IL-6 (post TNFα response) in CSF from only 6 survivors patients of group II and III
Fig. 5 - The positive correlation between NT -3 and IL -6 in serum from non-survivors in group I

proved outcome in brain trauma and IS. Three patientswith high level of TNFα in serum and CSF did not sur-vivors (7.75±0.5 days after IS).
In our patients the CSF level of NT -3 and IL-6 andtheir serum level appear related in group II and III ofsurvivors.
The survivors present a significant negative correla-tion between IL-6 and NT-3. The serum level of IL-6 proinflammatory cytokine decrease but the level of NT-3increase that could involved in the protective effectagainst ischemic injury. IS are associated with the ex-pression of inflammatory mediators that disturb theblood - brain barrier and prepare endothelium for leu-cocytes adhesion. The presence of pro-inflammatorylymphocytes influences the occurrence and severity ofstroke, stroke recurrence and death. This inflammatoryreaction not only contributes to lipid-membrane per-oxidation but also exacerbates the degree of tissueinjury due to the release of cytotoxic products fromactivates leucocytes. The generation and release of oxy-gen radicals are cytodistructive to the already compro-mised tissue.
NT-3 up-regulation might represent an activeresponse of the injuries brain or a result to inflamma-tory stimuli. That can represent a protective mechanismfor neuronal survival. We believe that interaction bet-ween NT-3 and IL-6 may also play a central roll amongbiochemical and molecular changes triggered by IS.
In CSF of our patients, the moderate CSF level ofNT-3 was found only to 6 survivors patients.
Although in patients with GOS=1 an extensive dis-ruption in the blood brain barrier can by hypothesized,the transfer of neurotrophin factors and cytokines proin-flammatory across biological barrier is unpredictable.The patients non survivors present a positive serumcorrelation between IL-6 and NT-3, high serum level ofneurotrophin and pro-inflammatory cytokine (IL-6),corresponded to imminence of death.
The level of IL-6 in serum and CSF from survivorswere lower that non-survivors with IS. Studied of therole of IL-6 in brain injury have produces inconsistentresult. Singhal et all. (2002) reported that elevated IL-6were linked to good outcome, while Kalabilikis et al.(1999) found no correlation between raised Il-6 andoutcome. Other studies have suggested that elevatedIL-6 in serum or CSF is associated with poor outcome.
In this regard NT-3 have been shown to modulatethe production of inflammatory cytokine IL-6 by thevascular cerebral endothelium, supporting that neuro-trophic factor protect the brain with hypoxic-ischemicbrain injury. The observed changes in the CSF and plas-ma survivors patients with IS of NT-3 may reflect anendogenous attempt at neuroprotection against bio-chemical and molecular cascades triggered by IS. The ne-
urotophin expression in the CSF could represent animportant marker of improve recuperation of this pa-tients and it is possible to discover prognostic and thera-peutic application of neurotrophins in patients with IS.
REFERENCES
BARBACID, M. - The Trk family of neurotrophin receptors.J. Neurobiol. 1994. 25. 1386-1403.
BIBEL, M. et al. - Biochemocal and functional interactionsbetween the neurotrophin receptors trk and p75 NTREMBO J. 1999. 18. 616-622.
SHYAM S SHARMA - Emerging neuroprotective approachesin stroke treatment.CRIPS 2003.4. 8-12.
ZOPPO, G., GINIS, I. et al. - Inflammation and stroke, puta-tive role for cytokines, adhesion molecules and ions inthe brain response to ischemia, Brain Pathology 2000.10. 95-112.
CAPLAN, L.R. - Insights and ideas from the recent stroke li-terature. Stroke 2005. 2 (1) 45-49.
MIDORI, A. - Yenari Pathophysiology of acute ischemicstroke. Clinic J.of Medicine 2004. 71. 25-27.
WINTER, D.C. et al. - Raised parenchimal interleukin-6 levelscorrelate with improved outcome after traumatic braininjury, Brain, 2004 127, 315-320.
HOCHHAUS F. et al. - Elevated nerve growth factor andneurotrophin-3 levels in cerebrospinal fluid of childrenwith hydrocephalus, BMC Pediatrics 2001, 1(2), 1-8.
KAPLAN D.R and MILLER F.D - Neurotrophin signal trans-duction in the nervous system, Curr Opin Neurobiol,2002: 10, 381-391.
WOICIECHOWSKY C. et al. - Early IL-6 concentration cor-relate with severity of brain injury, Journal of Trauma-Injury 2002, 52(2), 339-345.
XINKANG WANG - Investigational anti-inflammatoryagents for the treatment of ischaemic brain injury. ExpertOpin. Investig. Drugs 2005, 14, (4) 393-408.
CONTE, V. ROYO, N.C. et al. - Neurotrophic factors. EurJ.Trauma 2003, 29. 335-355
KOKAIA, Z. and LINDVALL, O. - Neurogenesis afterischaemic brain insults. Curr. Opin Neurobiol 2003, 13.127-132.
LENZLINGER, P.M. LAURER, H,L. et al. - The duality of theinflammatory response to traumatic brain injury. MolNeurobiol 2001, 24. 169-181.
Neurotrophin-3, TNF-αα and IL-6 relations in serum and cerebrospinal fluid of ischemic stroke patients
33

Escherichia coli is a common facultative anaerobemember of human intestinal micro flora. Although mostE.coli strains are living in a perfect symbiosis in thegastrointestinal tract, many of them are causing a varietyof diseases in animals and humans.
Escherichia coli is a very heterogeneous specieswith a genome in a continuous dynamic. Most genes,even those encoding conserved functions are poly-morphic. E. coli is very permissive to lateral genetictransfer and is now recognized that more than 18% ofthe contemporary genes of its genome are obtainedfrom other species by horizontal transfer (1, 4). Most ofthe virulence factors that distinguish different patho-types were acquired from diverse sources includingplasmids, bacteriophages or mobilized genomic regionsfrom other bacteria.
Pathogenic E. coli strains associated with human andanimal diseases are expressing a set of virulence fac-
tors, such as adherence factors (adhesions and fimbriae),toxins, proteins involved in internalization and inva-sion of host cells, systems of iron uptake (sideropho-res), etc (16). Based on these properties pathogenicstrains cause enteric diseases ranging from cholera-likediarrhoea to severe dysentery. Beside these there arestrains able to colonize the urinary tract and to causecystitis and / or pyelonephritis. Such strains could alsodetermine other extra intestinal infections like septi-caemia and / or meningitis.
Microbiological analyses of foods involved in food-borne infections reveal multiple bacterial species con-tamination and among them, with high frequency, aredetected ETEC and VTEC (21). Diarrheagenic strains ofE.coli contribute to water related diseases in urban andrural environment, all over the world, in developingcountries as well as in developed ones. Animal farmscould also be an important reservoir of E. coli. Human
34
ABSTRACTEscherichia coli, heterogeneous species consisting of commensal and pathogenic strains,is causing a broad spectrum of intestinal and extra intestinal diseases, ranging from asymp-tomatic infections to septicaemia, according to its capacity to produce different virulencefactors. The incidence of different virulence - associated genes among the strains isolated from healthysubjects, taking into account that the human gastrointestinal tract is considered an importantsource for spreading E. coli strains, was evaluated. A total of 241 E. coli strains isolated from 41 healthy subjects, working in the food chain andcoming to the laboratory for periodical medical control, were investigated for harbouring pato-genicity factors - encoding genes. Extra intestinal virulence - associated genes, pap, sfa/foc, afa,hly, cnf and intestinal ones eaeA, bfp, agg, lt, st, vtx1 (stx1), vtx2 (stx2) and ipaH, were target-ed by PCR using cellular lysate for total DNA. Genes encoding for adherence were the most prevalent. A number of 67 strains (27.80%) werepositive for pap genes and 34 strains (14.11%) presented PCR positive results when afa geneswere targeted, but sfa/foc genes were identified in only 10 strains (4.15%). Genes encoding fortoxigenesis were less prevalent. A total of 9 strains amplified hly genes, 2.49% were positivefor cnf genes and only 2 strains presented vtx1(stx1) gene.The results are in concordance with those which demonstrate that healthy subjects carryingstrains possessing virulence - encoding genes could represent a reservoir for environmental cir-culation of such strains, considered life - threatening when a receptive host is encountered.
INCIDENCE OF VIRULENCE-ENCODING GENES AMONG ENTERICESCHERICHIA COLI STRAINS ISOLATED FROM HEALTHY SUBJECTS
Maria Damian1*, Codruþa-Romaniþa Usein1, Dorina Tatu-Chiþoiu1, A.M.Palade1, Natalia Popovici1, Simona Ciontea1, Maria Nica2, Laura Grigore1
(1“Cantacuzino” National Institute of Research-Development for Microbiology and Immunology, Bucharest, Romania;2Hospital for Infectious and Tropical Diseases “V. Babes”, Bucharest, Romania)
*Address for correspondence: Maria Damian, INCDMI “Cantacuzino”, Splaiul Independentei 103,Sector 5, Bucharest, [email protected]

infections with VTEC strains have been associated withbovine contact as well as with consumption of raw milk,and beef products. Use of manure and water resultingfrom farms for fertilization represent also a source ofsurface water as well as for vegetables contamination.Person-to-person contact contributes, in addition to thefood and water contaminated, to the spread of infec-tion (2, 5, 11, 21, 22).
Both pathogenic and non-pathogenic strains are ex-creted by humans, the presence of E. coli in surface wateror/and foods and feeds being considered as an indica-tor of faecal contamination.
The aim of the present study is to evaluate healthysubjects as a risk factor for spreading of non-pathogen-ic, virulence encoding genes bearing strains, for publichealth.
As classical cultural methods are not able to dis-criminate between pathogenic and non-pathogenicstrains, molecular diagnostic techniques have provento be very useful tools (15). Genetic structures involvedin synthesis of virulence factors in E. coli isolates arelocated on chromosome or mobile genetic elementsand PCR and/or nucleic acid probes are used for theiridentification (14).
MATERIALS AND METHODS
Bacterial strains and DNA preparationA total of 41 healthy subjects, male and female, agedbetween 19 and 55 years, working in food chain, addre-ssing to the laboratory for periodic microbiologic con-trol, were screened for the presence of E. coli strainscarrying virulence encoding genes. Stool samples wereplated on MacConkey agar, ADCL, and MacConkeywith sorbitol. A number of 5 to10 colonies both lac-tose positive and lactose negative as well as sorbitolpositive and sorbitol negative were analysed for E. coliidentification. A total of 241 strains were collected inorder to be submitted to further analysis.
E. coli K12 strain HB 101 and E. coli reference strainsfor different E. coli pathotypes (kindly provided by ChantalLe Bouguénec from Pasteur Institute, Paris) were used asnegative and positive controls respectively.
The DNA used as target molecule in PCR was ob-tained by thermic lyses. Briefly, bacteria were grownin Brain Hearth Infusion broth overnight, at 37 °C and250 rpm. An aliquot of 1,5 ml broth culture was cen-trifuged at 10000 rpm for 5 min. The pellet was resus-pended in 100 µl sterile ultrapure water (MilliQ -Millipore) and boiled for 10 min.
PCR analysesStrains were analysed for virulence factors distinc-
tive for both enteric and extra intestinal pathotypes.
Genes coding for fimbrial adhesins associated withpyelonefritis (pap), S and F1C fimbriae (sfa/foc), afim-brial adhesin (afa), haemolysin (hly), cytotoxic necro-tizing factor (cnf) production as well as genes respon-sible for synthesis of attachment-effacement factor(eaeA), bundling forming factor (bfpA), verotoxin/shigatoxin (vtx/stx1 and vtx/stx2), thermo-labile (lt) and ther-mo-stabile toxin (st) and invasion protein (ipaH) weredetected by PCR. Primers used for annealing were cho-sen from those cited previously (6, 10, 17, 18, 20). Thereactions were performed in a final volume of 50 µlcontaining 10 µl of bacterial lysate, 30 pmol of eachprimer, 200 µM of each deoxynucleoside triphosphateand 1.5 U of Taq DNA Polymerase (Roche). The con-centration of MgCl2 was 1.5 mM per reaction.
PCR amplification was done in a Mycyler thermo-cycler (BioRad) and consisted of one cycle at 94°C for5 min, 25 cycles of denaturation at 94°C for 30s,annealing at each pair of primers’ specific temperaturefor 30s, and extension at 72°C for 1 min, followed bya 2s time increase per cycle. Aliquots (10 µl) of thefinal reaction mixtures underwent standard gel electro-phoresis in 1.5% agarose. Amplified DNA fragmentswere detected by UV fluorescence after ethidium bro-mide staining.
RESULTS
All 241 strains isolated from healthy subjects wereanalysed for the presence of genes involved in intes-tinal and extra intestinal virulence of E.coli strains.
Genes coding for adherence to the epithelial cellswere the most prevalent. A number of 67 (27.80%)strains presented pap genes, 34 strains (14.11%) werepositive for afa genes and 10 (4.15%) strains gave apositive PCR when sfa/foc genes were targeted. Lessprevalent were genes involved in toxins production.Genes encoding for haemolysin were identified in 9(3.73%) strains, cnf genes in 6 (2.49%) strains andvtx/stx1 genes only in 2 (0.83%) strains (table 1).
Among the 241 strains analysed a number of 165(68.46%) were negative for all the genetic structurestargeted. The others presented one gene (32 strains),two genes (38 strains), three genes (4 strains) and fourgenes (2 strains). The most prevalent gene associationwas pap and afa (28 strains) and the less prevalent wasafa and hly. Generally, 31.54% among the strains iso-lated from healthy subjects presented at least one viru-lence - associated gene (Fig. 1).
DISCUSSION
Based on clinical and microbiological criteria, the po-tential pathogenic strains were classified in so-calledintestinal pathotypes and extra intestinal pathotype.
Incidence of virulence-encoding genes among enteric Escherichia coli strains isolated from healthy subjects
35

Six different enteric pathotypes are identified whichcause symptomatology ranging from mild auto limita-tive diarrhoea to severe haemorrhagic colitis, often com-plicated with haemolytico-uremic syndrome. The sixintestinal pathotypes recognised are: enterotoxigenicE. coli (ETEC), enteropathogenic E.coli (EPEC), entero-haemorragic E. coli (EHEC)/shiga-like toxin-producingE. coli (SLTEC)/verotoxin producing E. coli (VTEC),enteroinvasive E. coli (EIEC), enteroaggregative E. coli(EAEC) and diffusely adherent E. coli (DAEC). They areexpressing virulence factors encoded by genes locatedon chromosome or mobile genetic structures. Besidesthe virulence factors involved in adherence, coloniza-tion and internalization (bundle forming pilus, intimine,invazins) they are carrying genes encoding the toxinproduction (thermo-labile toxin, thermo-stable toxin,verotoxines/shiga like toxins). They are identified bydetecting the genes coding for such factors: eaeA andbfp for EPEC, lt and st for ETEC, ipaH for EIEC, vtx/stx1and/or vtx/stx2 and eaeA for EHEC, agg for EaggEC (6,17, 18).
The extra intestinal pathotype is represented bystrains involved in urinary tract infections, neonatal me-ningitis and sepsis. The so-called urinary pathogenicstrains are able to express adherence factors encodedby pap, sfa/foc and afa genes, siderophores encoded byaer genes as well as toxins encoded by cnf and hlygenes (8, 9, 10, 11, 13).
Human and animal gut is considered as the reser-voir of E. coli strains and identification of such strainsin aliments or surface water is an indicator for pollu-tion/faecal contamination. Other authors demonstratedthe presence of E. coli strains bearing virulence genesin environmental water and in stool of healthy volun-teers (13). In our previous studies, investigating stoolsof patients with urinary tract infections but withoutdigestive simptomathology, as source of urinary infec-tions, we found a percent of strains carrying virulencegenes (20). The results obtained on a restricted numberof patients encouraged us to extend the study on a largergroup of healthy individuals working in alimentarychain.
Among the strains isolated from normal intestinalflora we found 31.54% strains carrying from 1 to 4 viru-lence encoding genes grouped in 9 different geneticprofiles. Most prevalent association was pap and afafollowed by pap and sfa/foc. Other associations werepap, sfa/foc, hly and cnf, afa and hly, pap and sfa/focor hly and cnf. This is in accordance with other authorswho investigated the genes associated with urinaryinfection and postulated that the genes are located onpathogenicity island and they are transmitted in link-age (7).
We have to note that from 76 strains carrying viru-lence genes only in 2 of them we detected virulencegenes distinctive for enteric pathotypes. All the others
36
DAMIAN et al.
Table 1 - Distribution of virulence genes among the E. coli strains

presented virulence genes characteristic for extra intes-tinal pathotypes. That could explain why the subjectsdidn’t develop simptomatology of enteric infections.These strains could be considered non-pathogenic,based on their behaviour in the host of origin, but theycould be of risk for new immunocompromised ormore receptive host.
The fact that 2 strains isolated from the same sub-ject, carrying stx1/vtx1 gene didn’t develop diseasecould be explained on one hand by the relationshipbetween carriage of a gene and its expression and onthe other hand by the relationship between the patho-genicity power of strains and host receptivity. The vir-ulence factors are not considered independently of thehost’s defences. The clinical aspects of diseases dependof the interaction of virulence factors with the host’sresponse. This is in concordance with the results obtai-ned by the other authors who demonstrated that intro-duction of a single virulence-associated gene into thegenome of E. coli K12 doesn’t convert the strain topathogenicity (7).
The high percentage of strains carrying extra intes-tinal virulence genes could support the hypothesis thatintestinal micro flora represents a reservoir for UTI in-fection (19). In the proper environmental and host con-ditions the virulence genes could revert to expressionand we could assume that in a receptive host the strainscan develop extra intestinal infections.
Among the 41 healthy subjects screened in thisstudy for carrying virulent E. coli in their normal diges-tive flora, 16 were positive. The number of positive strainsamong the 5-10 analysed for each individual rangedfrom 1 to 10, but all positive ones presented the samevirulence pattern. This fact suggests that at least 10colonies have to be analysed when healthy subjects
are evaluated. On the contrary, in diarrhoea infections,the strains are equipped with all necessary tools tomultiply in the host and consecutively to disturb theequilibrium of cell physiology. In this case, more then80 % of bacteria in a pathological sample are identicaland a number of 3 colonies seem to be sufficient toidentify the pathogen.
Horizontal gene transfer is one of the mechanismscontributing to the evolution of prokaryotic genomes.Acquisition of plasmids and phages as well as genom-ic islands plays an important role in the variability ofbacterial species (3). Considering the high frequencyof genetic transfer among the E. coli strains, and thefact that most of virulence-encoding genes are locatedon mobile genetic structures, the human gut could beconsidered a perfect ecological site for development ofa rich flora carrying virulence factors by conversion tovirulence of a great population of non-virulent cells. Asstudies demonstrated (12) there is a widespread distri-bution of potentially virulent E. coli strains in the envi-ronment and healthy humans carrying virulent strainscould represent an important source. This is a matter ofconcern for individuals but also a real threat for publichealth when the individuals are working in the chainfood.
ACKNOWLEDGEMENTS
Partly, this work was supported by the Project 47 ofthe CEEX Programme of Romanian Ministry of Educa-tion and Research.
Incidence of virulence-encoding genes among enteric Escherichia coli strains isolated from healthy subjects
37
Fig. 1 - Distribution of E. coli strains carrying virulence encoding genes. Among 241 strains 68% are negative for all investigatedvirulence determinants. A proportion of 16% are positive for pap and sfa/foc, pap and afa or afa and hly. Strains presenting onevirulence gene (pap or afa or hly or vtx/stx1) represent 13% of the total strains. 1.66% of the investigated strains are carrying 3virulence determinants (pap, hly and cnf) and only 0.83% of them are positive in PCR for 4 genes (pap, sfa/foc, hly and cnf)

REFERENCES
1. BINGEN E., PICARD B., BRAHIMI N., MATHY S., DES-JARDINS P., ELION J. and DENAMUR E. - Phylogeneticanalysis of Escherichia coli strains causing neonatalmeningitis suggests horizontal gene transfer from a pre-dominant pool of highly virulent B2 group strains. J.Infect. Dis., 177:642-650, 1998
2. CLARKE S. C., HAIGH R.D., FREESTONE P.P.E. andWILLIAMS P.H. - Virulence of Enteropathogenic Esche-richia coli, a Global Pathogen. Clin. Microbiol. Ten. 16,3:365-378, 2003
3. DOBRINDT U., AGERER F., MICHAELIS K., JANKA A.,BUCHRIESER C., SAMUELSON M., SVANBORG C.,GOTTSCHALK G., KARCH H. and HACKER J. - Analysisof Genome Plasticity in Pathogenic and CommensalEscherichia coli Isolates by use of DNA Arrays. J. Bac-teriol., 185, 6:1831-1840, 2003.
4. DONNENBERG M.S. and WHITTAM T.S. - Pathogenesisand evolution of virulence in enteropathogenic andenterohemorragic Escherichia coli. J. Clin. Investig. 107,5:539-548, 2001.
5. FOXMAN B. and RILEY L. - Molecular epidemiology: focuson infection. Am. J. Epidemiol. 153: 1135–1141, 2001.
6. GERMANI Y., BÉGAUD E. and LE BOUGUÉNEC C. -Detection of the Escherichia coli attaching and effacinggene (eae A) in enteropathogenic strains by polymerasechain reaction. Res. Microbiol. 148:177-181, 1997.
7. HACKER J., BLUM-OEHLER G., MUHLDORFER I. andTSCHAPE H. - Pathogenicity islands of virulent bacteria:structure, function and impact on microbial evolution.Mol. Microbiol. 23: 1089-1097, 1997.
8. JOHNSON J.R. and RUSSO T.A.- Extraintestinal patho-genic Escherichia coli: „the other bad E.coli“. J. Lab.Clin. Med., 139: 155-162, 2002.
9. JOHNSON J.R. - Virulence factors in Escherichia coli uri-nary tract infections. Clin. Microbiol. Rev., 4: 80-128,1991.
10. LE BOUGUENEC C., ARCHAMBAUD M., andLABIGNE A. - Rapid and specific detection of the pap,afa, and sfa adhesin-encoding operons in uropathogenicEscherichia coli strains by polymerase chain reaction. J.Clin. Microbiol. 30, (5):1189-1193. 1992.
11. MAINIL J.G. and DAUBE G. - Verotoxigenic Escheri-chia coli from animals, humans and foods: who’s who?J. Appl. Microbiol. 98, 1332-1344, 2005.
12. MALDONATO Y., FISER J.C., NAKATSU C.H. andBHUNIA A.K. - Cytotoxicity Potential and Genotypic Cha-racterization of Escherichia coli Isolates from Envi-ronmental and Food sources. Appl. Environ. Microbiol.,71 4:1890-1898, 2005.
13. MUHLDORFER I., BLUM G., DONOHUE-ROLFE A.,HEIER H., OLSCHLAGER T., TSCHAPE H, WALLNER U.and HACKER J. - Characterization of Escherichia colistrains isolated from environmental water habitats and
from stool samples of healthy volunteers. Res. Micro-biol. 147, 625-635, 1996.
14. NATARO J.P. AND KAPER J.B. - Diarrheagenic Esche-richia coli. Clin. Microbiol. Rev. 11:142-201, 1998.
15. NATARO J.P. and MARTINEZ J. - Diagnosis and In-vestigation of Diarrheagenic Escherichia coli, in Mole-cular Bacteriology Protocols and Clinical Applications(Woodford N and Johnson A.P.) Humana Press Inc,Totowa, NJ, 387-405, 1999.
16. PETERSON J.W. - Bacterial Pathogenesis, in Medical Mi-crobiology (Samuel Baron) Univ of Texas Ed. USA, 1996.
17. POLLARD, D. R., W. M. JOHNSON, H. LIOR, S. D.TYLER, and K. R. ROZEE - Rapid and specific detectionof verotoxin genes in Escherichia coli by the polymerasechain reaction. J. Clin. Microbiol. 28:540-545, 1990.
18. TORNIEPORTH N. G., JOHN J., SALGADO K., JESUS P.,LATHAM E., MELO M. C., GUNZBURG S. T. and RILEYL. W. - Differentiation of pathogenic Escherichia colistrains in Brazilian children by PCR. J. Clin. Microbiol.33(5): 1371–1374, 1995.
19. USEIN C.R., DAMIAN M., TATU-CHITOIU D., CA-PUSA C., FAGARAS R. and MIRCESCU G. - Comparisonof Genomic Profiles of Escherichia coli Isolates from Uri-nary Tract infections. Rom. Arch. Microbiol. Immunol.62, 3-4:137-153, 2003.
20. USEIN C.R., DAMIAN M., TATU-CHITOIU D., CAPUSAC., FAGARAS R., TUDORACHE D., NICA M. and LEBOUGUENEC C. - Prevalence of virulence genes inEscherichia coli strains isolated from Romanian adulturinary tract infection cases. J. Cell. Mol. Med. 5:303-310, 2001.
21. WANG R.F., CAO W.W. and CERNIGLIA C.E. - A uni-versal protocol for PCR detection of 13 species of food-borne pathogens in foods. J. Appl. Microbiol., 83:727-736, 1997.
22. WANI S.A., PANDIT F., SAMANTA I., BHAT M.A. andBUCHH A.S. - Molecular epidemiology of Shiga toxin-production Escherichia coli in India. Curr. Sci. 87,10:1345-1353, 2004.
38
DAMIAN et al.

Infectious diarrhea syndrome is an important cause ofhuman morbidity around the world, and Salmonellagenus remains one of the most prevalent etiology.Salmonellosis are zoonosis that may pass underrepor-ted when presenting as community sporadic cases, butwhen occuring as community foodborne outbreaks arelegally subjected to epidemiologic investigations andpublic health coordinated registration.
Our study focused on isolates of nontyphoidalSalmonella serotypes received by the Laboratory forEnteric Pathogens in N.I.R.D.M.I. “Cantacuzino” forconfirmation and subtyping. We investigated by pul-sed-field gel electrophoresis (PFGE) their XbaI macro-restriction profiles for providing evidence that epide-miologically related isolates collected during commu-nity outbreaks of disease were also genetically related.We reasoned that this information is helpful for thenational surveillance of foodborne pathogens, and maycontribute to building a harmonized public health in-frastructure for dealing with foodborne bacterial disea-ses at European level.
MATERIALS AND METHODS
Bacterial strainsThe 7 strains of Salmonella enterica serovar Typhi-murium included in this study were outbreak isolatessubmitted to Laboratory for Enteric Pathogens inN.I.R.D.M.I. “Cantacuzino”. We analyzed 2 small co-mmunity outbreaks that occurred in two Romanian vi-llages during this year in July. They were designated asO1 and O2. The clinical cases were associated withadults and children, and were reported as severe clini-cal cases of foodborne infections. O1 included 2 adultsand 1 child and was represented by 3 human isolatesplus 1 food isolate, while O2 included 2 children and1 adult, and was represented only by the human iso-lates.
Reference strainsSalmonella enterica serovar Braenderup H9812(obtained within the EQA system organized byPULSENET Europe) was used as PFGE control.
39
ABSTRACTInfectious diarrhea syndrome is an important cause of human morbidity around the world, andSalmonella genus remains one of the most prevalent etiology. Salmonella enterica serovar Typhimurium outbreak-associated isolates received by theLaboratory for Enteric Pathogens from N.I.R.D.M.I. “Cantacuzino” for confirmation and typingwere analyzed by genomic pulsed-field gel electrophoresis (PFGE) and phage susceptibilitytesting to establish their relatedness. Both typing methods proved to have similar discriminato-ry power. The isolates originating from the same outbreak belonged to the same phage typeand showed indistinguishable PFGE profiles. The molecular characterization of autochthonalSalmonella enterica Typhimurium outbreak human isolates provided laboratory evidence thatepidemiologically related isolates collected from community outbreaks of disease were alsogenetically related. In order to improve the national and international surveillance of major foodborne pathogensthe reference laboratory centers are required to establish and maintain the capacity to performa wide range of both phenotypic and genotypic methods to support outbreak investigations.
APPLICATION OF PULSED-FIELD GEL ELECTROPHORESIS TO LABORATORY SURVEILLANCE OF SMALL COMMUNITY OUTBREAKS
OF SALMONELLA ENTERICA SEROVAR TYPHIMURIUM
Codruþa-Romaniþa Usein1, A.M. Palade1, Natalia Popovici1, Dorina Tatu-Chiþoiu2, Simona Ciontea2, ªtefania Ceciu2, Maria Coºman2, Maria Damian1
(1Molecular Microbiology Laboratory, N.I.R.D.M.I. ”Cantacuzino”; 2Laboratory for Enteric Pathogens, N.I.R.D.M.I.”Cantacuzino”, Bucharest – Romania)

Phage typingA 2 hours broth culture plated on agar was used forphage susceptibility testing. Phage typing was perfor-med in accordance with the methods of the Laboratoryof Enteric Pathogens, Health Protection Agency (HPA),Colindale, London, United Kingdom. (ANDERSON et al.,1977) using a set of Salmonella Typhimurium phagesand the interpreting scheme provided by HPA throughENTERNET EQA system.
PFGE For our study we used the PFGE standard protocoldesigned by Centers for Disease Control and Preven-tion Atlanta (SWAMINATHAN et al. 2001), and recommen-ded within the MedVetNet/European PulseNet Projectfor non-typhoidal Salmonella strains. N.I.R.D.M.I.”Cantacuzino” as member of ENTERNET network isalso included in PULSENET Europe group.
The XbaI-restricted genomic DNA fragments wereresolved in a 1% Pulsed Field Certified Agarose (BioRad)using a CHEF-Mapper system (BioRad). For each gelthe well-characterized strain Salmonella BraenderupH9812 was included as both procedure control andmolecular size marker.
The electrophoretic conditions used were as fol-lows: initial switch time, 2.16 s, final switch time54.17 s, run time 19 h, angle 120°, gradient 6.0 V/cm,temperature 14°C, ramping factor linear. After elec-trophoresis the gel was stained for 20 min in 500 ml ofsterile distilled water containing 50ml ethidium bro-mide (10 mg/ml), destained for 1 h in distilled waterand photographed under UV light.
The Polaroid photos of PFGE patterns werescanned and subsequently subjected to computerizedanalysis using Dice similarity coefficient, UPGMA(unweighted pair group arithmetic average) algorithmand FingerPrinting II software (BioRad).
RESULTS AND DISCUSSION
The nature of foodborne disease investigations haschanged significantly, with more complex and wide-ranging investigations becoming the norm (O’BRIEN etal., 2003). Investigators rely on advanced microbiolo-gical evidence to supplement field epidemiology, andrequire strain typing data to identify outbreak-relatedstrains and to distinguish from sporadic isolates.
Although there is no current “golden standard” typingsystem for Salmonella fingerprinting, particularly inthe case of less commonly studied serotypes, PFGE iswidely used in local, national and international sur-veillance. The Centers for Disease Control and Preven-tion in Atlanta have developed the PulseNet system of
surveillance for typing human isolates of Salmonella,Listeria, and E. coli O157:H7 using PFGE (SWAMI-NATHAN et al., 2001). Europe also commonly usesPFGE as a complementary tool to traditional pheno-typic methods of analysis. This has developed into theSalm-gene Project and allowed the exchange of PFGEprofiles for common Salmonella serotypes across Eu-rope (PETERS et al., 2003). The system has proven valu-able for identifying the source of outbreaks and pro-vides a library of DNA patterns for future reference.
All the tested strains were phage typable and 2phage types (PT) were identified among the isolates.The O1 outbreak-associated isolates were identified asPT2, and O2 outbreak-associated isolates were PT85.
It has been previously reported that different PTscan give the same PFGE pattern and that the same PTcan give different PFGE patterns (CORBETT-FEENEY et al,.1998; DORAN et al., 2005), but within our study thePFGE molecular profiles of the isolates were in con-cordance with the phage typing results.
Macrorestriction patterns generated by XbaI, thefirst choice enzyme recommended for Salmonella,were resolved into 2 band patterns, which were desig-nated pulsotype XbaO1 and XbaO2 (Fig. 1). There wasno PFGE diversity among the same PT, the PFGE pat-terns for both PT2 and PT85 being indistinguishable.The food isolate was considered as the source of O1outbreak, and all O1 human isolates had identical bandpatterns when compared with it. According to TENOVER
et al. (1995) the epidemiologically related isolates withindistinguishable PFGE band patterns are consideredas outbreak isolates, having a common ancestor.
A tolerance of 1.5% was accepted for the genera-tion of the dendrogram (data not shown), and the clus-ter analysis revealed the different clonal origins of thetwo distinctive outbreak strains, with a similarity of84% among them. Further studies are needed to eval-uate the diversity of Salmonella serovar Typhimuriumisolates from human infections circulating in Romania.
In conclusion, the 7 Salmonella Typhimurium iso-lates were identified as outbreak strains by both phagetyping and PFGE typing. This is the first molecular PFGEanalysis of Salmonella serovars carried out in Roma-nia, so the existing knowledge on the subject is limited.Our national routine typing of Salmonella entericarelies on serotyping and phage typing. Previous studiesshowed that these conventional methods may lack thesensitivity to detect outbreaks involving commonserotypes (BENDER et al., 2001). Gaining our own expe-rience regarding the epidemiological and microbiologi-cal autochthonal characteristics of pathogens will per-mit a better evaluation of the utility of laboratory me-thods applied for the national infectious disease sur-veillance strategy. At the same time, taking into account
40
USEIN et al.

the enlargement of EU, integrating Romania also, theharmonization of our routine protocols and the rapidmolecular subtyping, at least for major foodborne dis-ease pathogens such as Salmonella enterica serovarsand E. coli O157:H7, will be mandatory.
REFERENCES
1. ANDERSON E.S., WARD L.R. and DE SAXE M.J. - Bac-teriophage-typing designations of Salmonella Typhimu-rium. J. Hyg 78, 297-300 (1977)
2. BENDER J.B., HEDBERG C.W., BOXRUD D.J., BESSERJ.M., WICKLUND J.H., SMITH K.E. and OSTERHOLMM.T. - Use of molecular subtyping in surveillance for Sal-monella enterica serotype Typhimurium. N Engl J Med433: 189-195 (2001)
3. CORBETT-FEENEY G. and RIAIN N.U. - The use ofpulsed-field gel electrophoresis for subdivision of Sal-monella typhimurium in an outbreak situation. J Infect36: 175-177 (1998)
4. DORAN G., MORRIS D., O’HARE C., DELAPPE N.,BRADSAW B., CORBETT-FEENEY G. and CORMICAN M.- Cost-effective application of pulsed-field electrophore-sis to typing of Salmonella enterica serovar Typhimu-rium. Appl Environ Microbiol 71: 8236-8240 (2005).
5. O’BRIEN S.J. and DE VALK H. - Salmonella - “Old” orga-nism, continued challenges. Euro Surveil, 8, 29-31 (2003)
6. PETERS T.M., MAGUIRE C., THREFALL E.J., FISHER I.S.,GILL N. and GATTO A.J. - The Salm-gene project – a Eu-ropean collaboration for DNA fingerprinting for food-related salmonellosis. Euro Surveil. 8: 46-50 (2003)
7. SWAMINATHAN B., BARRETT T.J., HUNTER S.B. andTAUXE R.V. - PulseNet: the molecular subtyping networkfor foodborne bacterial disease surveillance, UnitedStates. Emerg Infect Dis 7: 382-389 (2001)
8. TENOVER F.C., ARBEIT R.D., GOERING R.V., MIC-KELSEN P.A., MURRAY B.E., PERSING D.H. and SWA-MINATHAN B. - Interpreting chromosomal DNA restric-tion patterns produced by pulsed-field gel electrophore-sis: criteria for bacterial strain typing. J Clin Microbiol33: 2233- 39 (1995)
PFGE Typing of Salmonella Typhimurium outbreaks isolates
41
Fig. 1 - PFGE profiles of XbaI-digested genomic DNA from Salmonella enterica serovar Typhimurium outbreak isolates. Lanes1,6,10 Salmonella enterica serovar Braenderup; lanes 2-5 O1 outbreak-associated isolates; lanes 7-9 O2 outbreak-associatedisolates

Nontuberculous mycobacteria (NTM) includethose Mycobacterium species that are not members ofMycobacterium tuberculosis complex (M. tuberculo-sis, M. africanum, M. bovis, etc.), responsible fortuberculosis (TB) or for leprosis (M.leprae), hence theuse of the terms “nontuberculous mycobacteria” and“mycobacteria other than tuberculosis” (MOTT), orcarried the epithet “atypical” (originated from the mis-taken belief that they were unusual strains ofMycobacterium tuberculosis). Only few of thesemycobacteria, natural inhabitants of soil and waters, arepathogenic (but mostly less pathogenic), for the healthyimmunocompetent adult, and are responsible for thehuman infections called mycobacteriosis. Unlike tubercu-losis and leprosy, the environmental mycobacteria arenot transmitted from person to person (HEIFETS, 2004).
The infections produced by NTM have an in-creased incidence in the developed countries and re-latively lower incidence in developing countries butthe prevalence of mycobacterioses is continuouslygrowing (CORBETT et al., 1999, KATOCH, 2004).
The prevalence of TB in Romania is estimated for2005 as high as 126,4 0/0000. There are also an impor-tant number of NTM infections. These could be diag-nosed in immunocompetent people but, mostly, inimmunocompromised ones (HIV/AIDS patients, neo-plasies etc.). In North America and Western Europe,where the prevalence of TB is low, the incidence ofNTM infection is high, especially in AIDS patients (Am.Thoracic Soc., 1990). In Romania, there are cases when,concomitantly with Mycobacterium tuberculosis infec-tion, some of the patients are co-infected with one ortwo different NTM, the diagnosis and the treatmentbeing very difficult.
In the natural environment throughout the world,there are several NTM species, either slightly or notpathogenic for healthy persons. These could becomepathogenic when the local and general defense mecha-nisms are compromised (FALKINHAM, 1996; FRENCH et al,1997). Unlike tuberculosis and leprosy, the environ-mental mycobacteria are not transmitted from personto person.
42
ABSTRACTA rapid immunochromatographic serologic assay (Dot assay) is proposed to be applied onpatients infected with nontuberculous mycobacteria (NTM). This assay could evidentiatethe infecting species and allow the beginning of the treatment.The test is based on the principle of immunoblotting chromatography, a rapid membrane-based assay, capable of diagnosing NTM infections in serum, in less than 1 hour, with noneed of special equipment or skilled staff. The secreted extracellular antigens have beenisolated from the unheated culture filtrates of the clinically significant NTM (M. avium,MAI, M. kansasii, M. xenopi, M. chelonaei, M. scrofulaceum, M. marinum, M. fortuitum,M. abscesus, M. szulgai). The patients have been tested against these antigens, as well asfrom M.tuberculosis H37Rv, due to the possibility of co-infection with tuberculous bacilli.A number of 385 tests on patient sera have been performed (10, with NTM suspected infec-tion, with or without M.tuberculosis co-infection, 5 with confirmed diagnosis of NTMinfection, 10 with TB, 10 with other respiratory diseases). The preliminary results present-ed in this paper support the fact that the rapid immunochromatographic serum assay, com-bined with clinical and radiographic evidence, could evidentiate the infecting NTM speciesand allow the start of an earlier treatment, but must be confirmed on a higher number ofpatients.
RAPID IMMUNOCHROMATOGRAPHIC SERUM ASSAY OF NONTUBERCULOUS MYCOBACTERIAL INFECTIONS
Henriette Stavri1, Manuela Brânaru-Gheorghiu2, Olga Moldovan2, Marinela Rãileanu2, M.I. Popa3, Loredana Popa3, Luminiþa Ene4
(1National Institute of Research-Development for Microbiology and Immunology ”Cantacuzino”, Mycobacterial Antigens Department; 2Pneumophthisiology Institute “Marius Nasta”; 3University of Medicine and Pharmacy ”Carol Davila”; 4Infectious and Tropical Diseases Clinical Hospital „Dr. Victor Babes”, Bucharest - Romania)

Usually, the infections due to NTM are part of thegroup of opportunistic infections, meaning two-wayopportunities for causing infection in humans (HEIFETS,2004). One is the level/intensity of interaction with anatural or human-influenced environment that mayhave high contents of mycobacteria. The other is anenhanced sensitivity to infection with these organismsin some individuals. NTM infections had been descri-bed between 25 to 50 % of AIDS patients in the earlyepidemics. The mycobacteria most frequently respon-sible for the disease is M. avium (M. avium-intracellu-lare complex), but there are cases where a species con-sidered nonpathogenic, such as M. gordonae, couldtrigger the disease. Generally, in humans, 95 % of NTMinfections are due to M. avium, M. xenopi, M. kansasii,M. chelonei, M. scrofulaceum and M. marinum (NOLTE
et al, 1995).NTM infections as well as co-infection with TB and
non-TB strains are difficult and time consuming todiagnose. The diagnosis of NTM infections is difficultto establish in both cases, as well as in mixed infec-tions with tuberculous and nontuberculous bacilli.Despite the fact that some clinical manifestations andspecific mycobacteriosis treatment resemble classic TB,problems encountered are due to the high rate of re-sistance of most NTM to major usual TB drugs, stress-ing the need of individualized treatment, according toresistance testing (NAGAI et al, 2001)
This individualization is based on species of myco-bacteria recovered, site and severity of the infection,antimicrobial drug susceptibility test results, concurrentdiseases and the patient general condition. The golddiagnostic standard is the culture, NTM and Mycobac-terium tuberculosis possessing different characteristics(HEIFETS, 2005). But this takes approximately twomonths, a very long period of time. If the NTM diag-nosis is confirmed, it is mandatory to do the antibioticsusceptibility test in order to establish the treatment,taking into consideration the fact that the disease pro-duced by NTM is difficult to be treated, due to its mul-tiple resistance to TB drugs. It is very important to diffe-rentiate between M. tuberculosis and NTM infections,since the treatments for these two diseases are some-times different. Therefore, a rapid, sensitive and inex-pensive assay to confirm the NTM infections, for a my-cobacteriosis positive diagnostic and a differential diag-nosis in TB is needed.
The classical techniques of bacteriological diagno-sis: direct examination, culture and resistance testingutilized in NTM infections diagnosis, require an extre-mely long period of time (WALLACE, 1997). In order toshorten it, the blood culture (diagnostic method for di-sseminated infections, especially with M. avium, inAIDS patients) is used. Also, it uses the a medium cul-
ture (BACTEC/ MGIT), which allows the reduction byhalf of the time necessary for the growth of mycobac-teria: it could take 3 weeks for M.kansasii, to 3 monthsfor M. xenopi, to have a positive result ( WILSON et al.,1997). The use of nucleic acid probes represents arapid non-cultural diagnosis. Polymerase chain reac-tion technology may be an ideal method for identifi-cation of mycobacterial DNA in clinical samples (thereare commercial probes for M. avium, M. kansasii, M.gordonae complex, and Mycobacterium tuberculosiscomplex) but it requires sophisticated technology andit is too expensive for the developing world (TONJUM etal, 1998, YAJKO et al, 1995).The most popular approachesused now in top clinical mycobacteriololgy are cellwall lipid (mycolic acid ) analysis by HPLC (HEIFETS,2005) and the use of DNA probes.
A rapid identification of Mycobacterium species bylectin agglutination was recently presented (ATHAMNA
et al, 2006).An indirect diagnostic method is ELISA serology for
tuberculosis, which has been widely explored (COLE etal, 1996; STAVRI et al, 1988, 1993; NICULESCU et al,1995). Some of the assays, using a cocktail of five anti-gens as DETECT-TB (AMICOSANTE et al., 1999) providea potentially inexpensive alternative. But even so, per-forming an ELISA requires skilled staff and specialequipment.
Until now, there have been neither production norpublications about diagnostic assays for the most im-portant NTM, that are able to cause disease, whichcould analyze, in the same assay, the presence of thecirculating antibodies against all these clinically signif-icant species, by rapid immunochromatographic assayon immunocompetent and immunocompromisedpatients.
In this paper, we are proposing a rapid immuno-chromatographic serologic assay on patients infectedwith NTM, which could evidentiate the infecting spe-cies and allow the beginning of the treatment. The testis based on the principle of immunoblotting chroma-tography, a rapid membrane assay (STAVRI H. et al.,2003). Using the same assay principle, but with un-heated antigens isolated from the culture filtrates of NTMwith circulation in population, implied in pathogenic-ity, as well as from M. tuberculosis, it is possible to es-tablish a rapid serologic diagnostic, in less than onehour, when a mycobacterial infection is suspected.
MATERIAL AND METHODS
Study groups. Serum samples were obtained frompatients admitted to “M. Nasta” National Pneumoph-thisiology Institute, Bucharest, the reference clinic andhospital for patients with pulmonary diseases and „Dr.Victor Babes”, Infectious and Tropical Diseases
Rapid immunochromatographic serum assay of nontuberculous mycobacterial infections
43

Clinical Hospital Selection criteria included: (a)patients with suspicions of NTM infections, with orwithout M. tuberculosis co-infections; (b) patients withNTM infections, bacteriological, radiological and clin-ically confirmed (positive controls); (c) patients withTB; (d) patients with respiratory nontuberculous dis-eases or without any NTM infections (negative con-trols).
Bacteriological diagnosis (direct examination bymicroscopy, culture and resistance testing) was per-formed on all patients and the sera were collected. Thesera were kept at - 70°C until utilization.
All diagnoses of mycobacteriosis or/and tuberculo-sis were confirmed by identification of M. tuberculosisby culture according to standard methodology (RUNYON
et al., 1980), using Lowenstein-Jensen medium. Theacid fast smears were prepared from sputum, by themodified Petroff technique, described by KENT andKUBICA, 1985. Sputum specimens for Ziehl-Neelsenstaining were obtained from some patients in the firstthree days after admission to hospital, usually beforethe beginning of specific treatment (KUBICA, 1984), whileothers have been admitted to hospital for control, dur-ing and after their treatment. The sera were collectedin the same way, either at the start of the treatment, forsuspected infection with nontuberculous mycobacteriaor/and tuberculosis or other respiratory disease, orwhen the patients have been observed, during or afterdifferent periods of the therapy.
Sera of patients with other respiratory diseasesincluding: lung infection (acute respiratory tract infec-tions i.e. lower respiratory tract infections and pneu-monia, but not nosocomial forms), lung cancer, COPD,sleep apnea syndrome and Cor pulmonale (as compli-cations of COPD), acute or chronic bronchitis, asthma,pneumothorax, benign tumor, were included in thestudy. These patients were diagnosed by clinical, X-rayand microbial evaluation.
Informed consent was obtained from all patients.All sputa and sera samples were code labeled and keptat -70 °C until being tested. All samples were testedwithout knowledge of the clinical diagnosis of eachpatient, but the study groups were not randomly se-lected, as samples were intentionally collected for thedevelopment of the dot assay from patients stronglysuspected of having mycobacteriosis or tuberculosisand from those with other respiratory diseases.
All the subjects in this study were supposed to beBCG vaccinated at birth according to NationalVaccination programme.
Microorganisms and growth conditionsThe 11 mycobacteria species used were obtained fromATCC (M. abscessus, M. xenopi, M. chelonae), Pasteur
Institute Paris, France (M. tuberculosis H37Rv, M.avium, M. kansasii, M. fortuitum, M. scrofulaceum, M.marinum, M. szulgai), Dr. Joseph Lam, Dept. Micro-biology, College of Biological Sciences, University ofGuelph, Ontario, Canada (M.avium-intracellulare MAI).All species were cultivated on the surface of a synthe-tic liquid medium, Sauton [ L-asparagine monohy-drate, 4 g ; citric acid, 2 g; dipotassium phosphate, 0.5g; magnesium sulfate heptahydrate, 0,5 g; zinc sulfateheptahydrate, 0.04 g; ammonium iron (III) citrate (approx.7 % Fe), 0.05 g; glycerol, 65 ml, distilled water at 1000ml, (pH 7.3 – 7.4 )] at 37°C 3- 28 days ( depending onthe rapid or slow growth type of mycobacteria), withone exception: M.xenopi was grown at 42°C (GOOD
and SHINNICK, 1998). When each of the bacterium spe-cies had been grown as a compact pellicle on the liq-uid media surface, the respective culture filtrate wasfiltered twice, through Jena Schott G5 sintered glass fil-ters. In order to obtain the secreted extracellular pro-teins, the unheated culture filtrate was precipitatedwith 100 % trichlor acetic acid, to a 4 % final concen-tration. The precipitate was centrifuged, dissolved anddialyzed in PBS, 0.2 M, pH 7.4 and freeze-dried in 2ml ampoules, each containing 2 mg protein.
The method, i.e. SpA - colloidal gold conjugate pre-paration, the devices used for the assay as well as theprinciple of mycobacterial antibodies evidentiation,was similar with that previously described (STAVRI et al,2003) for pulmonary tuberculosis assay in patient sera.
The assay was performed on cassette devices, witha membrane disk on which the antigen could be fixedand utilizing a SpA-gold colloid conjugate which turnred when a positive result is reached.
The patient sera have been tested against these anti-gens, as well as from M. tuberculosis H37Rv, due to thepossibility of co-infection with tuberculous bacilli.When, in a patient serum there were antibodies to acertain mycobacteria, a red dot development wasobserved.
It is important to mention that in Romania, all thenewborns are BCG vaccinated.
RESULTS
There have been 10 patients with suspicion of NTMinfection, 5 patients with a bacteriological, clinical andradiographic diagnosis of NTM infection, 10 patientswith TB and 10 patients with other respiratory diseases.
It was extremely difficult to find patients infectedonly with NTM, since almost all who proved to beinfected with atypical mycobacteria were simultane-ously infected with M. tuberculosis.
The results of immunochromatographic assays onpatients with suspicion of NTM infection are presented
44
STAVRI et al.

in Table1, while the bacteriological, clinical and radio-graphic evidence on these patients is presented in Table 2.
Corroborating the results obtained using the rapidimmunochromatographic serum assay with bacterio-logical, clinical and radiographic data, 5 out of 10 pa-tients with suspicion of mycobacteriosis (with or with-out M. tuberculosis co-infection), were confirmed to beco-infected with M. tuberculosis and, respectively, M.avium, M. kansasii (2 patients), M. xenopi, and MAI +M. kansasii (1 patient).
The results obtained in immunochromatographicserologic assays in patients with mycobacterioses, NTMinfections, are presented in Tabel 3. Their diagnoseswere confirmed by bacteriological, clinical and radio-graphic data as shown in Table 4.
As shown in Table 3 and 4, 4 out of 5 patients withmycobacteriosis were confirmed to have NTM infec-tion, while one patient had a false negative diagnosis.
The assays were performed also on TB patients sera(Table 5). The TB patients as well as the patients with
Rapid immunochromatographic serum assay of nontuberculous mycobacterial infections
45
Table 2 - Bacteriological, clinical and radiographic evidence on patients with NTM suspicions
Table 1 - Immunochromatograhic assays for specific antibodies identification in the sera of patients with NTM suspected infection
Table 3 - Immunochromatographic serologic assays identification in NTM patients

pulmonary diseases other than TB were considered ne-gative controls for this assay.
2 out of 10 patients with TB, tested with each of 11mycobacterial antigens, had false positive results ( MAIand M.xenopi) in the dot serum assay.
In TB patients, the diagnosis was established basedon direct microscopy and culture in 9 out 10 patientsand by culture alone in 1 patient.
The assay results on the negative controls, i.e. pa-tients with pulmonary diseases, non TB and non NTMare presented in Table 7 and their bacteriological, cli-nical and X-ray results can be observed in Table 8.
8 out of 10 patients with other respiratory diseases,were confirmed by the rapid dot assay as negative,while 2 were false positives (one with M. tuberculosisand one with M. abscessus).
46
STAVRI et al.
Table 6 - Bacteriological, clinical and radiographic evidence on patients with TB
Table 4 - Bacteriological, clinical and radiographic evidence on patients with NTM infections
Table 5 - Immunochromatographic serologic assays for identification of NTM antibodies in TB patients

DISCUSSIONLack of reports on disease caused by NTM in many
countries is most likely related to insufficient efforts todetect such patients, mainly due to practices of thebacteriological testing, especially in countries withpoor resources. Another problem that may limit therecovery of NTM is that in many countries, culture iso-lation is performed only on egg-based medium, whichis not an optimal choice for isolation of NTM (HEIFETS,2004).
AIDS patients and also other severely immunosup-pressed patients (with corticosteroids treatments, can-cer, etc.), have higher susceptibility to mycobacterialinfections and develop disseminated mycobacterioses,especially with M.avium complex (KWARA et al, 2005).The infection with NTM could be asymptomatic orcould resemble Tb symptoms (e.g.: cough, sputumproduction, as those of TB) making the diagnosis diffi-cult (WILSON et al, 1997, PHILIPS et al, 2001). Clinicalmanifestations of NTM infections and also specifictreatment may overlap TB, but due to the high rate ofresistance of most NTM to major usual anti-tubercu-
lous drugs, stresses the need of individualized treat-ment, according to resistance testing (DAUZENBERG etal, 1994, MUELLER et al, 2001).
Among the physiologic characteristics of NTMwhich could contribute to their resistance to a widerange of antibiotics, the high hydrophobicity of thebacterial cell wall and its low permeability could betaken into consideration.
It is well documented that the reservoir for most ofthe infections with NTM, that we have diagnosed bythe rapid assay, and most likely for many other NTM,is soil, natural watersand aerosols created from thesesources (WALLACE, 1997; WOLINSKY 1968; MARRAS,2003). Finding so many infections with M .xenopi,could be explained by the fact that hospital heatingtanks and hospital hot water taps all around the worldhave been identified as a source of infection with M.xenopi, which may have entered the hospital tanksfrom general water supply and multiplied in tempera-tures (42°-45° C) favorable for this organism. Local tapwater and hospital water tanks have been reported asa source of infection with M.kansasii.
As analyzed in a recent review (MARRAS, 2003) themain risk factors for developing an infection with NTMare those associated with coexisting medical condi-tions, living and work environment, and demograph-ics.
The most important among the coexisting condi-tions are those that affect the general immune andlocal defenses. Thus, in the cases we had in this study,these were: previous and abandoned cases of tubercu-losis, HIV/AIDS, bronchiectasis, chronic obstructivepulmonary disease.
Samples obtained from individuals that have re-cently taken antibiotics or are currently undergoing an-tibiotic therapy may not yield appropriate results. An-tibody levels diminish rapidly after taking antibioticsand may decrease them below detectable levels in theserum even if an infection is still present.
Rapid immunochromatographic serum assay of nontuberculous mycobacterial infections
47
Table 7 - Immunochromatographic serologic assays for identification of NTM antibodies in patients with pulmonary diseases other than TB
Table 8 - Clinical and radiographic evidence onpatients with pulmonary diseases other than TB

CONCLUSIONSFor serum samples that test positive by this assay,
more specific confirmatory testing should be done. Aclinical evaluation of the patient situation and historyshould also be made before a final diagnosis is estab-lished. The use of a rapid test alone is not sufficient todiagnose an infection, with nontuberculous or/andtuberculous mycobacteria, even if antibodies are pres-ent. Also, a negative result does not preclude the pos-sibility of infection with a nontuberculous or/andtuberculous mycobacteria.
Therefore, the use of an inexpensive and rapidtechnique, which requires no equipment, little or noskill to perform, could be of value for the diagnosis ofthe NTM disease, regardless of the site of infection:pulmonary, cutaneous or other extrapulmonary loca-tion. Such assays that increased the indication of pos-sible mycobacteriosis as well as tuberculosis (BOTHAMLEY
et al.,1995). are of great value in confirming the clini-cal decision to begin specific treatment.
However, in the event of a negative test and thepresence of clinical signs, this is not enough to excludethe possibility that the patient has mycobacteriosis.Therefore, the patients should be tested by the otheraccepted technical and clinical methods to determinethe real status. The diagnostic value of an immunocro-matographic test depends on the context of its use(POTTUMARTHY et al., 2000). In a population with highprevalence of tuberculosis, a positive result could aidin clinical decision-making.
The possibility of identifying the NTM species,responsible for the patient`s infection, would be veryuseful, especially for those who have negative culturesor with extrapulmonary localizations.
The preliminary results presented in this paper sup-port the fact that the rapid immunochromatographicserum assay, combined with clinical and radiographicdata, could evidentiate the infecting NTM species andallow the start of an earlier treatment, but must be con-firmed on a higher number of patients.
REFERENCES
ATHAMNA A, D. COHEN, M. ATHAMNA, I. OFEKand HENRIETTE STAVRI - Rapid identification of Mycobac-terium species by lectin agglutination. J. Microbiol. Meth.,65, 209-215, 2006
BOTHAMLEY, G.H. -. Serological diagnosis of tubercu-losis. Eur.Respir.J., 8, 676-688.
COLE, R.A., LU, H.M., SHI, T.Z., WANG J, DE-HUA, T.and ZHOU A.T. - Clinical evaluation of a rapid immu-nochromatographic assay based on the 38 kDa antigen ofMycobacterium tuberculosis on patients with pulmonarytuberculosis in China. Tuber. Lung Dis. 77:363-368, 1996
CORBETT E.L., BLUMBERG L., CHURCHYARD G.J.,MOLOI N., MALLORY K. et al.- Nontuberculous my-cobacteria. Am. J. Respir. Crit. Care Med., 160 (1):15-21,1999
DAUTZENBERG, B. and A. MERCAT - Mycobacteriosesatypiques, Presse Med., 23:1483-8, 1994
FALKINHAM J. O. - Epidemiology of infection by non-tuberculous mycobacteria. Clin. Microbiol.Rev, 9(2):177-215, 1996
FRENCH A.L.,BENATOR D.A., GORDIN F.M. - Nontu-berculous mycobacterial infections, Med. Clin. North Ame-rica, 81(2), 361-379, 1997
GOOD, R.C., T.M. SHINNICK - Mycobacterium. InTopley and Wilson`s Microbiology and MicrobialInfections, Ninth Ed., chapt.26: 549-576, 1998
HEIFETS L., E. DESMOND - Clinical Mycobacteriology(Tuberculosis) Laboratory: Services and Methods. In S.T.Cole et al (ed.) “Tuberculosis and the Tubercle Bacillus”,2005, ASM Press, Washington, D.C.
HEIFETS L. - Mycobacterial infections caused by nontu-berculous mycobacteria. Seminars in Respiratory and Cri-tical Care Medicine, 25 (3): 283-295, 2004
KATOCH,V.M. - Infections due to non-tuberculous my-cobacteria (NTM). Indian J. Med. Res., 120: 290-304, 2004
KENT P.T., KUBICA, G.P. - Public Health mycobacteri-ology, a guide for level III laboratory. US Department ofHealth and Human Services, Public Health Services CenterClinical Microbiology, 3rd ed., American Society forMicrobiology, Washington DC, pp.1-257, 1985
KONDO A., OKETANI N., MARUYAMA M., SAITO Y.,MIYAO H., OHNO M., WADA K., TSUCHIYA T., NAGAIT., TSUCHIYA K., WATANABE Y. - Serological diagnosis ofpulmonary tuberculosis and non-tuberculous pulmonarymycobacteriosis. Kekkaku, 76(9):603-14, 2001
KUBICA, G.P. - Clinical microbiology. In The mycobac-teria. A Source book, Part A, 133-175, G.P. Kubica andL.G. Wayne ed., Marcel Dekker, New York., 1984
KWARA A., ROAHEN-HARRISON S., PRYSTOWSKYE., KISSINGER R. et al. - Manifestations of extrapulmonarytuberculosis: impact of human immunodeficiency virus co-infection. Int.J.Tuberc Lung Dis., 9 (5): 485-93, 2005
48
STAVRI et al.

MARRAS TK, DALEY CL. - Epidemiology of humanpulmonary infection with nontuberculous mycobacteria.In: Catanzaro A., Daley CL, eds. Lung disease due to non-tuberculous mycobacterial infections. Philadelphia: WBSaunders; 2003 : 553-568, Clinics in chest Medicine,Vol.23
MUELLER P.S., EDSON R.S. - Disseminated Mycobac-terium abscessus infection manifesting as fever of unknownorigin and intra-abdominal lymphadenitis:case report andliterature review. Diagn. Microbiol. Infect. Dis., 39:33-37 ,2001
NICULESCU D., STAVRI H., TEODOR I., POPA M.,STAVRI D., CIOMU E., HOMOS M. and POPESCU L. -Clinical usefulness of rapid serodiagnosis in pulmonarytuberculosis by ELISA with glycolipidic antigens extractedfrom Mycobacterium tuberculosis and with whole BCGsuspension. Rom.Arch.Microbiol.Immunol., T.54, 277-284, 1995
NOLTE F.S. and METCHOCK B. - “Mycobacterium” inP.R. Murray Manual of clinical microbiology, p.400-426,1995
PHILLIPS M.S. and C. FORDHAM VON REYN - Noso-comial Infections due to nontuberculous mycobacteria.Clin. Infec. Dis., 33: 1363-74, 2001
POTTUMARTHY S., WELLS V.C. and MORRIS A.J. - Acomparison of seven tests for serological diagnosis of tuber-culosis.J.Clin.Microbiol., 38(6):2227-2231, 2000
RUNYON, E.H., KARLSON A.G., KUBICA G.P. andWAYNE L.G. - Mycobacterium. In: Lennette, E.H., BallowsA., Hausler, Jr., W.J., Truant J.P. (Eds). Manual of ClinicalMicrobiology, 3rd ed., American Society for Microbiology,Washington, DC, pp.50-179, 1980
STAVRI H., O. MOLDOVAN, F. MIHALTAN, D.BANICA AND R.J.DOYLE - Rapid dot sputum and serumassay in pulmonary tuberculosis. J. Microbiol. Meth.,52:285-296, 2003
STAVRI, D., NICULESCU D., STAVRI H., BOREA F,STANCIU, G. and BASACOPOL, A. - Humoral immuneresponse in human pulmonary tuberculosis: antibodiesagainst Mycobacterium tuberculosis- polyssacharide, pro-tein and glycolipid antigens. Rom. Arch. Microbiol.Immunol., 52, 109-119, 1993
STAVRI, D., NICULESCU D., STAVRI H. and FUIOREAI. - Efficacy of two mycobacterial antigen preparations inthe serology of tuberculosis by enzyme-linked immuno-sorbent assay. Arch. Roum. Pathol. Exp. Microbiol., 47, 11-15, 1988
TONJUM T., WELTY D.B., E. JANTZEN, SMALL P.L. -Differentiation of Mycobacterium ulcerans, M. marinumand M. haemophilum: mapping of their relationships toM.tuberculosis by fatty acid profile analysis, DNA-DNAhybridization, and 16S rRNA gene sequence analysis. J.Clin. Microbiol, 36(4): 918-925, 1998
WALLACE R.J., GLASSROTH J., GRIFFITH, D.E.,OLIVIER K.N., COOK J.L. and GORDIN F. - Diagnosis andtreatment of disease caused by nontuberculous mycobacte-ria. Am. J. Respir. Crit. Care Med., 156:1-25, 1997
WILSON R., S. and ABDALLAH - Pulmonary diseasecaused by nontuberculous mycobacteria in immunocom-petent patients. Eur.Respir.Mon., 4:247-272, 1997
WILSON, M.L., STONE, B.L., HILDRED, M.V. andREVES, R.R. - Comparison of recovery rates for mycobacte-ria from BACTEC 12B vials, Middlebrook 7H11 biplates,and Lowenstein-Jensen slants in a public helath my-cobacteriology laboratory. J. Clin. Microbiol., 33, 2516-2518, 1995
WOLINSKY E. and RYNEARSON T.K. - Mycobacteriain soil and their relation to disease associated strains. Amer.Rev. Resp. Dis., 97:1031-1037, 1968
YAJKO, D.M., WAGNER, C., TEVERE, U.J.,KOCAGOZ, T., HADLEY, W.K. and CHAMBERS, H.F. -Quantitative culture of Mycobacterium tuberculosis fromclinical sputum specimens and dilution end-point of itsdetection by the Amplicor PCR assay. J.Clin.Microbiol., 33,1944-1947, 1995
xxx - American Thoracic Society. Diagnosis and treat-ment if disease caused by nontuberculous mycobacteria.Amer. Rev. Resp. Dis. 142:940-953, 1990
Rapid immunochromatographic serum assay of nontuberculous mycobacterial infections
49

Poly(3-hydroxybutyric acid) [P(3HB)], the unique intra-cellular carbon and energy reserve of many bacteriahas been identified as a biodegradable natural poly-ester. It undergoes both extracellular as well as intrace-llular degradations. Extracellular degradation involvesthe utilization of exogenous crystalline polymer bynon-accumulating microbes through the secretion ofextracellular P(3HB) depolymerase, (e-depolymerase)[16] whereas intracellular degradation of endogenousamorphous P(3HB) usually follows the exponentialaccumulation phase of accumulating organism pro-ducing intracellular P(3HB) depolymerase, (i-depoly-merase) [10]. The extracellular P(3HB) degradation isimportant from environmental view points which wasfirst studied by A. A. CHOWDHURY in 1963 [5]. The mi-crobial degradation of both endogenous and exoge-nous P(3HB) proceeds by enzymatic erosion of poly-mer to water soluble monomer and/or oligomers [8,25]. Unlike e-depolymerases, most of the i-depolyme-rases share the characteristics of less stability at widerange of temperature, pH, and ionic strength. Both of
the enzymes are relatively low molecular weight pro-teins and contain only one peptide chain in each [16].
In vivo degradation by the producer organism itselfis also important in relation to its physiology and opti-mization of production process. MACRAE and WILKINSON
[20] described the reduction in P(3HB) content ofBacillus megaterium cells during aerobic incubation ofpolymer rich cells in phosphate buffer and the lesserstability of P(3HB)-poor cells has also been recordedwith Ralstonia eutropha [12]. Later on the cyclic na-ture of P(3HB) metabolism (i.e. simultaneous synthesisand degradation) was confirmed in R. eutropha byfeeding 14C glucose as carbon source [8]. The rapid uti-lization of polymer in absence of exogenous carbonhas been studied in R. eutropha [24], Legionella pneu-mophila [15] and Hydrogenophaga pseudoflava [4,29] in details and in each case the key enzyme formobilization of native amorphous granule hydrolysiswas intracellularly located P(3HB) depolymerase.STEINBUCHEL et al [26] considered the enzyme as gran-ule associated protein, responsible for reutilization ofpolyesters under changed environmental conditions
50
ABSTRACTAzotobacter chroococcum MAL-201 accumulates poly(3-hydroxybutyric acid) [P(3HB)]accounting 69% of cell dry weight (CDW) from glucose during growth in nitrogen-freeStockdale medium. Degradation of the accumulated polymer by the organism was stud-ied under carbon-free medium following two-step cultivation method. P(3HB) content ofcells decreased rapidly from 69% to 4.8% of CDW after 35h under carbon-deprived con-dition. Autodigestion of P(3HB) was evident from the estimation of intracellular P(3HB)depolymerase (i-depolymerase) activity in cell-free extract using artificial P(3HB) granulesas substrate. Polymer content decreased rapidly along with the increase in i-depolymeraseactivity and rate of polymer degradation when medium was supplemented with(NH4)2SO4 at 0.1% (w/v) level. However, the effects were reverse when organic nitroge-nous substrate, beef extract at similar concentration was present in the medium. The opti-mum temperature and pH for i-depolymerase activity were 35°C and 7.7 respectively.The oxygen-limiting condition (culture volume per flask volume, 50%) decreased 10.7%activity of i-depolymerase over control resulting a slow P(3HB) degradation. The presenceof NaCl (6 X 103 µg/ml) showed a positive effect on i-depolymerase where as EDTA (40µg/ml) resulted in 20% less activity. Furthermore, the intracellular degradation of P(3HB)decreased the intrinsic viscosity, molecular weight and tensile strength of the accumula-ted polymer.
INTRACELLULAR DEGRADATION OF POLY(3-HYDROXYBUTYRIC ACID)ACCUMULATED BY AZOTOBACTER CHROOCOCCUM MAL-201
Soma Pal Saha and A.K. Paul*
(University of Calcutta, Dept. of Botany, India)
*Author for correspondence: E-mail: [email protected], Phone: 91(033)2475 3681; Fax: (033)2476 4419

and this enzyme is completely different from those ofe-depolymerases in relation to their origin and proper-ties [13]. The first biochemical studies on i-depolyme-rase were performed in Bacillus megaterium and Rhodos-pirillum rubrum by MERRICK and DOUDOROFF [21], thoughthe unstable nature of purified i-depolymerase wasdescribed later [9, 28].
Azotobacter chroococcum MAL-201 accumulatesP(3HB) accounting 69% of CDW during late exponen-tial growth and it utilizes the polymer during subse-quent phases of growth [22]. The present communica-tion describes the influence of cultural conditions andeffect of some chemicals on intracellular degradationof P(3HB) by the strain.
MATERIALS AND METHODS
Bacterial strain and mediaAzotobacter chroococcum MAL-201 (MTCC-3853), apotent P(3HB) producing, nonsymbiotic nitrogen-fixingsoil bacterium was used throughout the study. The nitro-gen-free Stockdale medium [27] containing 2% (w/v) glu-cose was principally used for growth and polymer pro-duction. The 24h old liquid culture in same mediumwas used as inoculum at 4%, (v/v) level. The Stockdalemedium without carbon source was used for degrada-tion studies. Organism was maintained on agar slopeof Stockdale medium at 4 oC. Salts and chemicals wereadded to the carbon-free medium as and when required.
Intracellular P(3HB) degradationDuring intracellular P(3HB) degradation a two stepcultivation method was used. Cells from early stationaryphase of growth containing P(3HB) about 69% of CDWwere harvested by centrifugation (X 10,000g) using aHitachi SCR 20B centrifuge and a rotor RPR-20-2-2548.Biomass was washed with carbon-free medium, trans-ferred aseptically to the same medium and incubated fora period under shake flask condition (120 rpm) at 32°C.
Analytical proceduresDetermination of growthThe growth was measured by viable cell count follow-ing direct dilution and plating on nitrogen-free Stock-dale agar medium and expressed as c.f.u./ml.
Determination of soluble proteinTotal soluble protein present in biomass was estimatedfollowing the method of LOWRY et al. [19] using Folinphenol reagent and bovine serum albumin (Sigma,USA) as a standard.
Assay of polymerThe accumulated P(3HB) in biomass was determinedfollowing the method of LAW and SLEPECKY [18] using
Jenway 6505 UV-VIS-spectrophotometer. P(3HB) ob-tained from Sigma (USA) was used as standard.
Assay of i-depolymeraseThe activity of P(3HB) i-depolymerase was assayed fol-lowing the method of SAITO et al. [24]. The cells weredisrupted by two passages through a French PressureCell at 140 X 106 Pa. The resulting cell free extract (CFE)was used as crude enzyme. To assay i-depolymerase ac-tivity, 1 ml of CFE as crude enzyme was added to theassay mixture (4ml) containing artificial amorphous gra-nules (1 mg as polymer) in Tris-HCI (50 µmol) buffer.One unit of activity is defined as the decrease of 0.01absorbency (A650) unit per minute.
Preparation of artificial granulesArtificial amorphous P(3HB) granules were preparedthrough emulsification of chloroform solution of poly-mer and sodium dodicyl sulphate (SDS, 100 mM) solu-tion using a sonicator following the method ofHOROWITZ and SANDERS [14]. The granules were washedwith SDS (1 mM) solution before assay.
Isolation and characterization of polymerThe intracellular polymer was extracted from oven-dried biomass with chloroform at 60°C following themethod of RAMSAY et al. [23]. The polymer was purifiedthrough diethyl ether precipitation. The viscometricmeasurement was performed in chloroform solution at30°C using Ubbelohde’s dilution viscosimeter [2] andmol. wt. of polymer was determined through intrinsicviscosity [η](dlg-1) measurement following the MARK-HOWINK equation: [η] = 7.7 x 10-5 x M0.82 [3]. The ten-sile strength of polymer sheet (0.75 mm thick and 4.0mm wide) was tested using Instron model 4301 with apulling speed 20 mm/min.
RESULTS
Time course of intracellular P(3HB) degradationAzotobacter chroococcum MAL-201 cells, pre-grown inStockdale medium which contained P(3HB) account-ing 69% of CDW were harvested after 27 h of incuba-tion. Cells were resuspended aseptically in equal volumeof carbon-free Stockdale medium and incubated at32°C under continuous shaking. Compared to normalgrowth conditions the number of viable cells (c.f.u./ml)in carbon-free medium declined sharply along with asteady fall of polymer content from 69.0% to 4.8%after 35 h (Fig. 1). The rates of polymer degradation ascalculated from the data obtained were 12.42 mg/hand 18.34 mg/h in normal growth condition and incarbon-free medium respectively.
Intracellular degradation of poly(3-hydroxybutyric acid) accumulated by Azotobacter chroococcum MAL-201
51

During the course of degradation of P(3HB), the i-depolymerase activity was evaluated. Cell-free extractof MAL-201 was obtained by disrupting the cellsthrough a French Pressure cell and its intracellular de-polymerase activity was assayed using artificial P(3HB)granules as substrates. The extent of intracellular de-gradation of P(3HB) was further increased along withan increment of i-depolymerase activity when the car-bon-free Stockdale medium was supplemented withammonium sulphate at a concentration of 0.1% (w/v).The situations were reverse with beef extract at similarconcentration for both parameters (Fig. 2, A and B).The rates of polymer degradation after 35 h (as calcula-
ted from Fig. 2A) were 18.34, 19.7 and 16.7 mg/h innitrogen-free medium and in media with (NH4)2SO4 andbeef extract respectively. Almost 99.9%, polymer wasconsumed by the cells in medium with (NH4)2SO4
where as more than 15% of cellular P(3HB) was left inpresence of beef extract and 32-40% inhibition of i-de-polymerase activity was noticed. The maximum de-polymerase activity in cells was observed after 15-20 hof incubation under nitrogen-limiting condition andwith (NH4)2SO4 in presence of organic nitrogen itrequired 25h to reach its maxima.
Influence of temperature and pHThe temperature profile of intracellular degradation ofP(3HB) after 15 h incubation in nitrogen and carbon-free medium indicated maximum polymer degradationrate (32.2 mg/h), i-depolymerase activity (5.15 unit) andspecific activity (7.2 unit) at a temperature of 35°C(Table 1). Similarly, optimum pH for polymer degra-dation was recorded at 7.5 although optimum depoly-merase activity (5.2 unit) and specific activity (6.45)were noticed at pH 7.7 (Table 2).
Influence of oxygen-limitationOxygen-limitation was imposed with a variation in cul-ture volume per flask volume (CVF). The extent of via-bility and polymer content in cell showed a raisingtrend with increase in CVF % (Table 3). Simultaneousdecrease in i-depolymerase activity from 5.1 to 4.55unit (i.e. 10.7 % decrease than control) and rate ofP(3HB) degradation from 26.8 to 22.26 mg/h wererecorded when CVF increased from 20-50%.
Influence of chemicalsThe influence of a number of salts (NaCl, KC1, etc.),alcohols (propanol, ethanol), reducing agent (dithio-
52
SOMA PAL SAHA and A.K. PAUL
Fig. 1 – Time course of growth (n) and P(3HB) content (�) ofA. chroococcum MAL-201 cells grown in glucose (2%, w/v ath) containing medium as represented by smooth lines (¾). The dotted lines (- -) represent the growth (o) and P(3HB)content of cells (∆) in carbon-free medium under shakeflask condition (140 rpm) at 32 oC
Fig. 2 – Effect of nitrogen on P(3HB) content of cell (A) and i-depolymerase activity (B) of A. chroococcum MAL-201 duringthe course of cultivation in carbon-free medium. Cultural conditions are same as Fig. 1. Symbols: (n) – control, without nitro-gen; (l) – with (NH4)SO4 0.1% w/v and (�) – with beef extract, 0/1% w/v

Intracellular degradation of poly(3-hydroxybutyric acid) accumulated by Azotobacter chroococcum MAL-201
53
Table 1 – Effect of incubation temperature on viability of cell, P(3HB) degradation and i-polymerase activity of A. chrococcum MAL-201 during cultivation in carbon-free Stockdale medium
a – Cells with P(3HB) accounting 69.0% of CDW were suspended in carbon-free medium and incubated at different temperatures under-shake flask condition (120 rpm).Biomass was harvested after 15 h of incubation.
b – P(3HB) was estimated following the method of LAW and SLEPECKY (1961)c – The rate of polymer degradation was determined by calculating the total P(3HB) consumption by a fixed volume of culture per hour.d – Intracellular depolymerase activity and its specific activity were determined following the method of SAITO et al. (1995). Cell protein
was estimated following the method of LOWRY et al. (1951).
Table 2 – Effect of initial pH of culture medium on viability of cell, P(3HB) degradation and i-depolymerase activity of A. chroococcum MAL-201 during cultivation in carbon-free Stockdale medium
a – Cells with P(3HB) accounting 69.0% of CDW were suspended in carbon-free medium and incubated at different temperatures under-shake flask condition (120 rpm).Biomass was harvested after 15 h of incubation.
b c d – Same as Table 1
Table 3 – Effect of oxygen-limitation on viability of cell, P(3HB) degradation and i-depolymerase activity of A. chroococcum MAL/201 during cultivation in carbon/free Stockdale medium
a – Increase in culture volume per flask volume (CVF) decreases the aeration in medium.b c d – Same as Table 1

threitol, DTT), alkylating agent (sodium azide), non-io-nic detergents (tritonX-100, tween 80), ionic detergent(sodium dodicyl sulphate, SDS), esterase inhibitors(phenyl methyl sulphonyl fluoride, PMSF), chelatingagent (ethylene diamine tetraacetate, EDTA) and othertoxic chemicals (marcaptoethanol, MCE, etc.) onP(3HB) degradation and i-depolymerase activity weretested (Table 4). Among the tested chemicals, the pre-
sence of NaCl resulted in maximum rate of degrada-tion (40.66 mg/h) with 10% more enzyme activity thancontrol. The chelating agent EDTA inhibited the de-polymerase activity maximum and consequently therate of polymer degradation was found as low as 15.43mg/h when after 15 h intracellular P(3HB) was still45.85% of CDW and it was more than 58% higherpolymer content compared to control.
Changes in polymer propertiesDuring incubation in carbon-free medium A chroococ-cum degraded and utilized its intracellular P(3HB) in-clusions when simultaneous changes in physico-me-chanical properties of polymer were noticed (Fig. 3).Decrease in intrinsic viscosity (15.5 to 5.0 dl/g), accom-panied with a slow reduction in molecular weight (2830to 834 kda) of the accumulated polymer and its tensilestrength (35 to 10 Mpa) were recorded.
DISCUSSION
Poly(3-hydroxybutyric acid) is synthesized by polyme-rization of 3-hydroxybutyryl-coA catalyzed by P(3HB)polymerase [11] and in most of the organism P(3HB) isutilized as carbon and energy source under carbon-deprived conditions [l]. Results as obtained with A.chroococcum MAL-201 clearly indicated triggering ofrapid degradation of accumulated polymer by sudden
54
SOMA PAL SAHA and A.K. PAUL
Table 4 – Effect of chemicals on P(3HB) degradation in A. chroococcum MAL- 201 during cultivation in carbon-free Stockdale medium
a Chemicals at sublethal dose were added individually to carbon-free media at h.b c d Same as Table 1
Fig. 3 – Changes in properties during intracellular degrada-tion of P(3HB) of A. chroococcum MAL-201 in carbon-freemedium. Cultural conditions are same as Fig.1. Symbols:(*) – intrinsic viscosity, (�) – molecular weight and (l) –tensile strength

carbon deprived condition and more than 93% poly-mer in cells were degraded after 35 h (Fig. 1). Thehydrolyzing activity of i-depolymerase in cell-free ex-tract was established [24, 26]. The results of Fig. 2Bshowed that i-depolymerase was present even at Oh,which was actually the maximum polymer accumula-tion phase for those cells during cultivation in carboncontaining medium (Fig. 1). More than 16 fold increaseof enzyme activity after 15 h in carbon-free mediumand then a decrease in activity with time suggests thati-depolymerase activity was proportional to polymercontent of cells (Fig. 2 A and B). DOI et al. [8] describeda simultaneous operation of P(3HB) polymerase anddepolymerase enzyme in Ralstonia eutropha under nongrowth condition; later on it was proved that in pre-sence of carbon, P(3HB) chain propagation with poly-merase was 10 times faster than chain degradation withdepolymerase and vice versa in absence of carbon [17].
Stimulatory effect of (NH4)2SO4 on native P(3HB)digestion was recorded where the rate of degradationand enzyme activity increased by 7.47% and 6% respec-tively compared to those of control. The decrease in theprofile of polymer content and i-depolymerase activitywere consistent with each other. Such an influence of(NH4)2SO4 was also recorded in Hydrogenophaga pseu-doflava [4, 7]. The decrease in enzyme activity as re-corded with beef extract (Fig. 2B) suggested the invol-vement of complex nitrogen in the formation of someprimary metabolites other than P(3HB) which did notallow the cell to digest its storage material efficiently.
The i-depolymerase is heat sensitive [16] but theendogenous enzyme showed a minimum activity at50°C (Table 1). MERRICK and DOUDOROFF [21] des-cribed the decrease in polymer degradation rate afteran exposure of Rhodospirillum rubrum cells to highertemperature as the less susceptibility of substrate frac-tion for subsequent hydrolysis with depolymerase. Ta-ble 2 showed that there were two distinct optimum pHfor endogenous i-depolymerase, active on native gran-ules (pH 7.5) and the i-depolymerase in cell-free extractactive on artificial SDS coated P(3HB) granules (pH 7.7).These findings suggested the dependence of results onextensive substrate specificity and assay systems.
As Azotobacters are free-living nitrogen-fixers, about20% air saturation may also inhibit their growth [6].Under carbon and nitrogen deprived condition, limita-tion of oxygen imposed a further stress condition anddid not allow the organism to utilize accumulated po-lymer where more than 10.7% decrease in depoly-merase activity was noted. MACRAE and WILKINSON [20]proved that during active phase of degradation a sig-nificant amount of P(3HB) were broken down to CO2
and water when a good volume of oxygen was availableto cell, required for the complete oxidation of polymer.
Addition of higher concentration of inorganic salts result-ed in an osmotic stress condition, which promoted theenzyme activity up to 10%. Inhibition of i-depoly-merase in presence of EDTA suggested that the en-zyme was either dependent on Ca++ or other divalentcations. The reduction of enzyme activity by other che-micals was not so effective, though further experimentson i-depolymerase with EDTA might be hopeful.
During intracellular degradation the time depend-ent changes in molecular weight (Fig.3) and so tensilestrength were due the hydrolysis of polymer by i-depolymerase. KWAGUCHI and DOI [17] described i-depolymerase as an exotype hydrolase which hydroly-ses an ester bond of P(3HB) at carbonyl terminus ofchain and released 3-hydroxy butyric acid monomerultimately. Therefore, studies on intracellular P(3HB)degradation in A. chroococcum MAL-201 thus allowone step ahead to large scale production of this indus-trially promising polymer with controlled quantity andquality.
AcknowledgementThe authors are indebted to Prof. S. N. Gupta, Depart-ment of Applied Chemistry, University of Calcutta andDr. Shaon Roy Chowdhury, Department of Biophysics,University of Calcutta for their invaluable assistance ininstrumental facilities. This research was conductedfrom the financial support of the Council of Scientificand Industrial Research (CSIR), New Delhi.
REFERENCES
ANDERSON A.J. and DAWES E.A.- Occurrence, metabo-lism, metabolic role and industrial uses of bacterial poly-hydroxyalkanoates. Microbiol. Rev., 54, 450 (1990)BIBERS I and KALNINS M. - Control of biopolymerpoly-p-hydroxybutyrate characteristics by γ-irradiation.Mech. Comp. Mat,. 35, 169 (1999)
2. BRUCKNER S, MEILLE S.V. and MALPEZZI L.-The struc-ture of poly -β-hydroxybutyrate). A refinement based onthe Reietveld method. Macromol, 21, 967 (1988)
3. CHOI M.H., YOON S.C. and LENZ R.W. - Production ofpoly(3-hydroxybutyric acid-co-4-hydroxybutyric acid)and poly (4-hydroxybutyric acid) without subsequentdegradation by Hydrogenophaga pseudoflava. Appl.Environ. Microbiol., 65, 1570 (1999)
Intracellular degradation of poly(3-hydroxybutyric acid) accumulated by Azotobacter chroococcum MAL-201
55

4. CHOWDHURY A. A. - Poly-β-hydroxybuttersaureabbauende bakterien und exoenzym. Arch. Microbiol,47, 167(1963)
5. DALTON H. and PSTGATE J.R. - Growth and physiol-ogy of Azotobacter chroococcum in continuous culture.J Gen Microbiol, 56, 307 (1969)
6. DOI Y., KUNIOKA M, NAKAMURA V. and SOGA K. -Nuclear magnetic resonance studies on unusual bacter-ial copolyesters of 3-hydroxybutyrate and 4-hydroxybu-tyrate. Macromol, 21, 2722 (1988).
7. DOI Y., SASAESUGA A., KAWAGUCHI Y. and KUNIO-KA M. - Cyclic nature of poly(3-hydroxyalkanoate) me-tabolism in Alcaligenes eutrophus. FEMS Micribiol Lett,67, 165 (1990)
8. HANDRICK R. and JENDROSSEK D. - Extracellular andintracellular polyhydroxyalkanoate depolymerase: homo-logies and differences. In: Steinbutchel A (ed) Bioche-mical principles and mechanisms of biodegradation anddegradation of polymers. Wiley VCH, Weinheim, pp 57(1998)
9. HANDRICK R, REINHARD S. and JENDROSSEK D. -Mobilization of poly(3-hydroxybutyrate) in Ralstoniaeutropha. J. Bacteriol, 182, 5916 (2000)
10. HAYWOOD G.W., ANDERSON A.J. and DA WES E.A.- The importance of PHB-synthase substrate specificityin polyhydroxyalkanoate synthesis by Alcaligenes eu-trophous. FEMS Microbiol Lett.,51, 1 (1989).
11. HIPPE H. and SCHEGEL H.G. - Hydrolyse Von PHBsduerch intracellulare depolymerase von Hydrogeno-monus HI6. Arch Mikrobiol, 56, 278 (1967)
12.HOCKING P.J and MARCHESSAULT R.H - Biopo-lyesters. In: Griffin GJL (ed) Chemistry and technology ofbiodegradable polymers. Chapman and Hall, London,pp 48 (1994)
13. HOROWITZ D.M. and SANDRERS J.K.M. - Amor-phous, biomimetic granules of polyhydroxybutyrate:preparation, characterization and biological implica-tions. J.Am.Chem. Soc, 116, 2695 (1993)
14. JAMES B.W., MAUCHLINE W.S, DENNIS P.J., KEEVILC.W. and WAIT R. - Poly-3-hydroxybutyrate in Legio-nella pneumophila an energy source: for survival in lownutrient environments. Appl. Environ. Microbiol., 65,822 (1999)
15. JENDROSSEK D. - Extracellular PHA depolymerases: thekey enzymes for PHA degradation Ann. Rev. Microbiol.56, 41 (2001)
16. KAWAGUCHI X, DOI Y - Kinetic and mechanism ofsynthesis and degradation of poly(3-hydroxybutyrate) inAlcaligenes eutrophous. Macromol., 25, 2324 (1992)
17. LAW J.H. and SLEPECKY R.A. - Assay of Poly-p-hydroxy-butyric acid./ Bacteriol,82,33(1961).
18. LOWRY O.H., ROSEBROUGH N.J., FARR A.L. andRANDALL R.J.- Protein measurement with Folin phenolreagent. J Biol Chem 193, 265 (1951)
19. MACRAE R.M. and WILKINSON J.E.- Poly-P-hydroxy-butyrate metabolism in washed suspension of Bacilluscereus and Bacillus megaterium. J. Gen. Microbiol. 19,210(1958)
20. MERRICK J. M. and DOUDOROFF M. - Depolyme-rization of poly-p-hydroxybutyrate by an intracellularenzyme system. J Bacteriol 88, 60 (1964)
21. PAL S, MANNA A., PAUL A.K. Nutritional and culturalconditions for production of poly-P-hydroxybutyric acidby Azotobacter chroococcum. Folia Microbiol, 43, 117(1998).
22. RAMSAY B.A., LOMALIZA K., CHAVERIE C, DUBE B.,BATAILLE P., RAMSAY J.A. - Production of poly-β-hydro-xybutyrate-co-β-hydroxyvalerate). Appl. Environ Microbiol.,55, 584 (1990)
23. SAITO T., TAKIZAWA K. and SAESUGA H. - Intracellularpoly(3-hydroxybutyrate) depolymerase in Alcaligeneseutrophous. Can. J. Microbiol.41, 187(1995)
24. SCANDOIA M. and FOCARETE M.L. - Frisoni G Simplekinetic model of heterogenous enzymatic hydrolysis ofnatural poly(3-hydroxybutyrate). Macromol., 31 3846(1998)
25. STEINBUTCHEL A., AERTS K., BABEL W, FOLLNER, C,LIEBERGESELL M,MADKOUR MH, MAYER F, PIPER-FRUST U, PRIES A, VALENTIN HE, WIECZOREK R -Considerations on the structure of biochemistry of bac-terial polyhydroxyalkanoic acid inclusions. Can J Mi-crobiol 41: 94-105 (1995)
26. STCKDALE H., RIBBONS D.W. and DAWES E.A. - Occu-rrence of poly-β-hydroxybutyrate in the Azotobacteria-ceae. J Bacteriol, 95, 1798 (1968).
27. TECHNOW U. - Isolierung und substratbindedomaneder extrazellularen PHB depolymerase a von Pseudomo-nas lemoignei aus recombinanten Escherichia coli undAnreicherung und character isierung des aktivitors inin-trazellularen PHB abbau von rhodospirillum rubrum.Diplomrbeit Univ Gottingen, pp.108 (1999)
28. YOON S.Y., CHOI M.H.H - Local sequence dependenceof polyhydroxyalkanoic acid degradation in Hydroge-nophagapseudoflava. J. Biol. Chem. 274, 37800 (1999)
56
SOMA PAL SAHA and A.K. PAUL

Plasma application in life sciences is a recent trend,naturally born from the well-established and thrivingsolid state processing technology. Most non-thermaldischarges are generated under reduced pressures(<1mbar), but not all organic materials can withstandsuch conditions (OHL et al., 1999). Nevertheless, low-pressure plasmas are being successfully used in bio-medical engineering, e.g. for surface pattering to con-trol cell adhesion or spraying of bio-compatible mate-rials to improve the performance of artificial implants(HEIMANN and VU, 1997)
Many plasma based devices have been constructedto sterilize plastic medical equipment; air filters havebeen equipped with corona –like discharges to destroybacteria and spores in public areas (BIRMINGHAM andHAMMERSTROM, 2000). Atmospheric plasmas aremore flexible and less expensive in operation, becausethey do not require costly vacuum systems. The appli-cations of the plasma needle in vivo deactivation ofdental bacteria: plaque destruction and treatment ofcaries (SLADEK and STOFFELS, 2005). Deactivation ofharmful bacteria can be performed by applying chem-ical or physical factors. Conventional methods of ster-ilization involve heat (steam and dry autoclaves),chemical agents (active gases like chlorine and ozone,or solution) and irradiation (UV, gamma). An alterna-tive method of sterilization is treatment with non-ther-mal plasmas (LAROUSSI. M., 2002; MOISAN M. et al.,2001; LAROUSSI and LEIPOLD, 2004). These plasmasoperate under moderate temperature and use non to-xic gases. Bacterial deactivation has been reported in
low- pressure as well as atmospheric plasmas (STOF-FELS et al., 2002).
Numerous techniques of surface modification havebeen investigated and successfully implemented onthe industrial scale: reactive ion etching of semicon-ductor/photoresist, deposition of amorphous siliconfor solar cells, production of hard/ protective/hydrophobic/ decorative layers, etc. In all these appli-cations, refined treatment is performed using non-ther-mal gas discharge plasmas. All processes occur atroom or slightly elevated temperature, so that treatedsurfaces do not suffer any thermal damage (RADETICet al., 2001; HOCKNEY et.al , 2004; WHITTAKERA.G. et al., 2004).
We have developed an atmospheric plasma (theplasma needle), which operate at room temperature.
For the first time in our country, plasma needle wasdesigned by Plasma Physics Department, NationalInstitute for Laser, Plasma and Radiation Physics,Bucharest, Romania. The plasma is generated in areactor chamber at the tip of a metal needle by using aradio-frequency generator (13.56 MHz, 100’s V ampli-tude). The feed gases are helium, argon, nitrogen, air,with flow rates of hundreds of ml/min. Ranges of sta-ble plasma operation are established for various plas-ma chemistries. Electron excitation, vibrational androtational temperatures are determined using opticalemission spectroscopy. Regarding plasma needlegeometries it is known that the first two geometries arecoaxial ones. The metal needle is placed coaxially inthe last geometries and removed the outer cylinder.
57
ABSTRACTIn this paper, we present the effect of the plasma needle on tumor cell surface. The plasma isgenerated at the tip of a metal needle by using a radio-frequency generator of 13.56 MHz,100’s V amplitude. In our study we investigated the interaction of non – thermal plasma (plas-ma needle) with living monolayer tumour cells in culture medium. We applied short needleto sample distance (1 mm) at temperature of 25 oC, 30 oC and 37 oC, respectively. Our datasugest that the plasma needle reduces the viability and induces apoptosis of tumour cells.These activities may be very useful in dermatology, where a part of the tissue must be removedwith high-precision, without damage to the adjacent cells and without inflammatory reaction.
THE EFFECT OF THE PLASMA NEEDLE ON TUMORAL CELL LINES APOPTOSIS
Ana Cãlugãru1, Lidia Cremer1, Aurora Herold1, Andreea Lupu1, G. Szegli1, C. Lungu2, Ana Lungu2 and N. Georgescu2
(N.I.R.D.M.I. “Cantacuzino”, Immunomodulation Department; N.I.L.P.R.PH, Plasma Pysics Department, Bucharest- Romania)

The cylinder is filled with the working gas, dimensionsare much smaller. The working gas is blown towardsthe needle tip only, flowing along the needle. The heliumflow rate was of 3 l/min. The RF voltage for a stableplasma needle was 150- 400Vrms. The typical emissionspectrum was in the range of 273 – 432 nm.
The plasma needle is a small-sized (1mm) atmos-pheric glow sustained by radio-frequency excitation.
In this paper we studied the interactions of a small-size, non thermal plasma (plasma needle) with diffe-rent tumour cells: murine melanoma (B16 line); humanbreast carcinoma ( BrSk line). Treatment of the abovementioned cells was restricted to the membrane surface.
We measured the cell viability and studied theapoptosis/necrosis processes induced by plasma nee-dle on different tumour cell lines.
MATERIALS AND METHODS
Plasma needle Plasma needle was designed by Plasma Physics De-partment, National Institute for Laser, Plasma and Radi-ation Physics, Bucharest, Romania.
Cell preparationTumour cell lines (BrSk – human breast cancer; and B16– murine melanoma) were grown in flasks containing cul-tured at 37 oC and 5% CO2 humidified atmosphere inculture complete medium (DULBECCO’S MODIFIEDEAGLE’S MEDIUM NUTRIENT MIXTURE F-12 HAM)supplemented with 10% FCS, antibiotics and 2mMglutamine.
Flow cytometry analysis
For plasma needle treatment, the cells (2x105 cells /ml) were cultured at 37 oC and 5% CO2 humidified at-mosphere in culture complete medium (DULBECCO’SMODIFIED EAGLE’S MEDIUM NUTRIENT MIXTUREF-12 HAM) supplemented with 10% FCS, antibiotics and2 mM glutamine, for 24 hrs in flat-bottomed 24 well tis-sue culture plates (COSTAR). After 24 hrs an optimumconfluence – monolayer (the percentage of the cell-co-vered area on the well sample was about 90%) wasreached.
After 24 h, the adherent cells were treated withplasma needle for 30 seconds, 60 seconds and 180 se-conds, respectivelly. The treatment was performed at25 oC, 30 oC and 37 oC respectively. To prevent cellsdrying out, samples were covered with a film of mediumof 0.3 mm. Finally, the cells were trypsinized and resus-pended in 100 µl PBS buffer. To each sample, 10 µlPropidium Iodide (PI -10 µg stock solution) wereadded and incubated 30 minutes in darkness, at roomtemperature. The apoptotic cells were analysed on
FACSCalibur (Becton-Dickinson, San Jose –USA) usingCellQuest software. Cells were first gated according totheir scatter characteristics and then analyzed forapoptosis by WINDMI.2.7 programme.
Morphological modifiers assayComplementary methods Giemsa stain slides and pic-tures collected by digital camera from OM (OpticalMicroscop) to compare the results obtained by flowcytometry were used.
The results were represented on pictures.
RESULTS AND DISCUSSION
Literature reported that the plasma needle became arefined surgical tool to dispose pathological cells (can-cer) and unwanted tissues (peeling), to fight bacterialinfectious in vivo sterilization of skin and dental cavi-ties and to improve wound healing by controlling celladhesion (STOFFLES E. et al., 2003.
Our results were focused on the capacity of the plas-ma needle to affect the viability of tumoral cell lines.
I. FLOW CYTOMETRY RESULTS
Effect of plasma needle on B16 tumour cells lineB16 tumour cell line (murine melanoma – are basaltype fibroblasts) was used in our experiments as posi-tive control. After 24 h of cultivation, these cells weretreated with plasma needle different exposure peri-ods:30 seconds, 60 seconds and 180 seconds, respec-tively; the temperature of plasma needle developedwas of 25 oC, 32 oC or 37 oC and this temperature wascontrolled by thermocouple system. Plasma needleactioned at 1 mm distance to monolayer cells. Wehave observed that the viability of B16 cells decreaseddepending on the action timing of plasma needle atcellular surface. In case of plasma needle treatment at32 oC for 30 seconds, B16 cells viability was of 55%;when plasma needle actioned 60 seconds or 180 sec-onds, the viability was 28% and 20% respectively(Table 1, Fig.1). Flow cytometric measurements de-monstrated that plasma needle induced apoptosis ofB16 cells. Were registered 42% apoptotic cells at 30seconds; 67% apoptotic cells at 60 seconds and 74%at 180 seconds respectively. The necrosis was verylow. We have established that plasma needle treat-ment does not induce necrosis, but a temporary inter-ruption of cell adhesion could be possible.
In the case of plasma needle treatment at 37 oC, theeffect of plasma needle on B16 tumour cells for 30 sec-onds, 60 seconds, 180 seconds respectively is present-ed in Table 1, Fig.2. Our data revealed that the viabil-ity of B16 cells under plasma needle treatment wasdecreased to control sample (cells untreated by plasma
58
CÃLUGÃRU et al.

needle at time 0). Thus, was registered of 41%, 26%and 20% living cells for 30 seconds, 60 seconds and180 seconds respectively plasma needle action time.Apoptosis was increased at both parameters; tempera-ture and plasma needle action time.
Effect of plasma needle on BrSk tumour cells line Another tumour cell line used in our research wasBrSk (human breast cancer). Plasma needle conditionswere the same for B16 tumour cells.
Table 1, Fig. 3 and Fig. 4, revealed the results ob-tained after BrSk human breast cancer plasma needletreatment at 32 oC and 37 oC temperature for 30 sec-onds, 60 seconds and 180 seconds. We observed thatBrSk human breast cancer are more resistant againstplasma needle treatment at 32 oC comparatively withB16 tumour cells. For instance, the viability of BrSkcells was 81% and 75% for 30s and 60 s plasma nee-dle action time respectively. During 180 seconds treat-ment, these cells viability decreased to 40%.
A necrotic effect of plasma needle was not detec-ted. We noticed that at 37 oC BrSk cells treated withplasma needle become apoptotic cells, especially at60 seconds and 180 seconds timing action of plasmaneedle. The percent of apoptotic cells was of 93%.
Effect of plasma needle on BrSk and B16 tumour celllines at room temperatureAs can be seen in Fig. 5, the viability of BrSk and B16tumour cells was not affected by 30 seconds plasma nee-dle treatment at room temperature (25 oC). The apop-tosis of BrSk tumour cells induced by plasma needleaction was only 6%. In case of B16 tumour cells theapoptosis was of 16%. The necrosis was decreased, 2%for BrSk cells and 7% for B16 tumour cells.
II. MORPHOLOGICAL MODIFIERS OF TUMOURCELLS AFTER PLASMA NEEDLE TREATMENT
The pictures are representative for morphologicalmodifiers produced by plasma needle treatment on
The effect of the plasma needle on tumoral cell lines apoptosis
59
Plasma needle installation for experiment
Table 1 - Status of B16 and BrSk tumour cells (percent of cells) after plasma needle treatment. Analysis by flow cytometry

60
CÃLUGÃRU et al.
Fig. 1 - The viability of B16 tumour cells line after plasma needle at 32 oC
Fig. 2 - The viability of B16 tumour cells line after plasma needle at 37 oC
Fig. 3 - The viability of BrSk tumour cells line after plasma needle treatment at 32 oC

The effect of the plasma needle on tumoral cell lines apoptosis
61
Fig. 4 - The viability of BrSk tumour cells line after plasma needle treatment at 37 oC
Fig. 5 - The viability of B16 and BrSk tumour cell lines after plasma needle treatment at room temperature for 30 seconds
Pictures b -B16 cells after plasma needle treatment
at 32 oC for 30 seconds
Pictures a -B 16 cells control

62
CÃLUGÃRU et al.
Pictures d - B16 cells after plasma needle treatment at 37 oC for 180 seconds
Pictures c - B16 cells after plasma needle treatment at 32 oC for 60 seconds
Pictures f - BrSk cells after plasma needle treatment at 32oC for 30 seconds
Pictures e - BrSk cells control

The effect of the plasma needle on tumoral cell lines apoptosis
63
Pictures h - BrSk cells after plasma needle treatment at 37oC for 60 seconds
Pictures h - BrSk cells after plasma needle treatment at 37oC for 180 seconds
Pictures g - BrSk cells after plasma needle treatment at 32oC for 60 seconds
tumour cells (line B16 and line BrSk) at 30 oC and 37 oCfor 30 seconds, 60 seconds and 180 seconds, respec-tively. Plasma needle actioned at 1 mm distance tomonolayer cells.
These results are correlated with the resultsobtained by flow cytometry analysis. As can be seenfrom pictures , plasma needle treatment on tumourcells induced apoptosis phenomena of these cells.The morphological modifiers produced by plasmaneedle on tumour cells (B16 and BrSk lines) were thefollowing: loss of membrane asymmetry (b and d pic-tures), cell shrinkage (b and c pictures), chromatin con-densation and chromosomal DNA fragmentation (pic-tures b, c, f, and h pictures), superaggregation andclumping chromatin (i picture), different features ofchromatin clumps (b and h pictures), vacuolization (bpicture), surface blebbing and cell retraction (d, f and ipictures). All morphological modifiers after plasmaneedle treatment were analysed to control sample,cells untreated by plasma needle at time 0 (a - B16 ande BrSk tumour cells pictures ).
CONCLUSIONS
Our results sugest the capacity of plasma needle toreduce the viability and to induce apoptosis of tumour
cells. We observed that the necrotic effect of plasmaneedle was not detected by flow cytometry analysis orGiemsa stain slides.
We conclude that the plasma needle may be apotential instrument to modulate the apoptosis oftumour cells in future as applications for dermatologytreatment.

ACKNOWLEDGEMENTS
This work was supported by Ministry of Education andResearch programme CERES 4-141,2/2004-2006
REFERENCES
OHL A., SCHRODER K., KELLER D., MEYER-PLATH A.,BIENERT H., HUSEN B., and RUNE G.M. - J. Mater. Sci.Mater. Med. 10, 747, 1999
HEIMANN R.B., and VU T.A. - J. Thermanl Spray Technol,6, 145, 1997
BIRMINGHAM J.G. and HAMMERSTROM D.J. - IEEETrans. Plasma, Sci. 28, 51, 2000
SLADEK R.E.J and STOFFELS E. - Deactivation ofEscherichia coli by the plasma needle. J. Phys. D. Appl.Phys, 38, 1716-1721, 2005
LAROUSSI. M., IEEE Trans Plasma Sci., 30.1409. 2002;
MOISAN M., BARBEAU J., MOREAU S., PELLETIER J.,TABRIZIAN M., and YAHIA L.H., Int.J. Pharm., 226,1,2001
LAROUSSI M. and LEIPOLD F. - Int. J. Mass Spectrum.233, 81 2004
STOFFELS E., FLICKWEERT A.J. and KROESEN G.M.W -Plasmas Sources Sci. Technol. 11, 383 , 2002
STOFFLES E., KIEFT I.E., and SLADEK R.E.J. - Superficialtreatment of mammalian cells using plasma needle, J.Phys. D: Appl. Phys. 36, 2908-2913, 2003
RADETIC M., JOCIC D., JOVANCIC P., TRAJCOVIC R andPETROVIC Z. Lj. - Textile Chemist Colorist Am. DyestuffReporter, 32, 55, 2000
MOSELHY M., SHI W., STARK R.H and SCHOENBACHK.H, Appl. Phys. Lett, 79, 1240, 2001
HOCKNEY R.C. - Trends Biotechnol. 12, 456,1994WHITTAKER A.G., et.al - J. Hosp. Infect. 56, 37, 2004
64
CÃLUGÃRU et al.

During the last decades, due to massive pollution ofsoil and water environments with oil wastes, there hasbeen a growing international interest in the possibilityof bioremediating these ecosystems by using variousmicrobial communities.
Even though there is quite a lot of information onthe role of bacteria in biodegradation, there is ratherpoor knowledge on yeasts with biodegradative abili-ties. Until now, the yeast species able to use hydrocar-bons as sole carbon source have been identified asbelonging to several genera: Rhodotorula, Candida,Pichia, Hansenula and Yarrowia [3; 5; 8; 10; 11; 14].
Petroleum, or rock oil, contains mainly hydrocar-bons: linear alkanes or “n-paraffins”, branched par-rafins or “isoparaffins”, cyclic alkanes and aromatichydrocarbons (compounds with one or more benzenerings). The ability of microorganisms of degradingthese hydrocarbons depends on their chemical struc-ture and molecular weight. Thus, the alkanes withshort strands are toxic for most of the microbial speciesand rather difficult to metabolize. The C5 – C8 alkanes
are not metabolized due to their ability to dissolve thecellular membranes and cellular structures involved inthe oxidation and transport of hydrocarbons in thecell. Yeasts with biodegrading abilities use mostly C10
– C24 n-alkanes [15]. Recent studies showed that yeastsdegrade in small rates aromatic compounds, and espe-cially those with more benzene rings. Phenols are alsobiodegraded, usually in combination with other car-bon sources [4]. Cyclic alkanes can be metabolized ifthey present also a branched aliphatic linear strand [1].
The major international interest regarding the pos-sibility to use yeasts in the degradation of oil com-pounds led to intensive studies regarding the isolationand identification of new yeast species and strains withdegradative abilities, using conventional [13] correlat-ed with molecular taxonomy data [6; 12]. The aim ofour study was the preliminary taxonomical identifica-tion and classification as well as the analysis, at labo-ratory scale, of the biodegrading abilities of five newyeast strains, named D1, D2, D3, D4 and D6, isolatedfrom oil-polluted environment.
65
ABSTRACTBioremediation is a very interesting alternative for restoring the oil-polluted ecosystems. Manystudies concerning the possibility of using microorganisms (bacteria and yeasts) in the degra-dation of oil compounds have as starting point the isolation and taxonomical identification ofnew species and strains with degradative abilities. Our study focusses on the preliminary clas-sification of five yeast strains (D1, D2, D3, D4 and D6) isolated from oil-polluted environ-ments. The strains were characterized by conventional taxonomical techniques: microscopicaland macroscopical appearance, fermentation abilities, assimilation of various carbon or nitro-gen compounds, growth under stress conditions (non-permissive temperatures, high glucoseconcentration) and urea degradation. According to these tests, D1, D2 and D4 showed greatsimilarity to Rhodotorula glutinis, D3 to Candida parapsilosis and D6 to Candida tropicalis.Further supplementary tests were performed in order to establish their ability to degrade hydro-carbons, by observing growth in media with n-alkanes (n-decane, n-dodecane, n-tetradecane,n-hexadecane). Thus, D1, D2 and D4 were the best alkane-consuming strains, presenting pos-sible similar degrading abilities and pathways, which correlates well to our identification asRhodotorula strains. For D3 and D6 the growth was not so spectacular as for D1, D2 and D4,but continuous along the entire experiment. The resemblance between the curves profiles con-firms the idea that both belong to the same genus, Candida.
PRELIMINARY IDENTIFICATION AND CLASSIFICATION OF FIVE NEW YEAST STRAINS ISOLATED FROM OIL-POLLUTED ENVIRONMENT
Ortansa Csutak1, Raluca Ghindea2, Ileana Stoica1, Simona Soare2, Robertina Ionescu2, Oana Creangã2 and Tatiana Vassu1
(1University of Bucharest, Faculty of Biology, Department of Genetics; 2University of Bucharest, Faculty of Biology, MICROGEN – Centre for Research, Education and Consulting
in Genetics, Microbiology and Biotechnology, Bucharest - Romania)

MATERIALS AND METHODS
1. Yeast strains and mediaStudies were performed on five yeast strains named
D1, D2, D3, D4 and D6 isolated from oil-pollutedenvironment (soil and water) from Pitesti area (Roma-nia). Standard yeast strains were used as reference: S. ce-revisiae ATTC 26108, C. parapsilosis GM (MICROGEN,Faculty of Biology, University of Bucharest), C. tropi-calis GM, Rh. glutinis GM.
Yeasts were maintained at -70 oC on yeast peptoneglucose (YPG) medium (yeast extract 5 g/l, peptone10 g/l, glucose 20 g/l) supplemented with 20% glycerol.Prior to taxonomical tests, yeasts were cultured onYPG slants, and grown for 18 hours, at 30 oC.
2. Conventional taxonomy tests2.1. Morphological characterizationYeasts from fresh growing culture were cultivated inPetri dishes, on YPGA medium (YPG medium supple-mented with agar 20 g/l), for observing the macrosco-pical appearance of the colony type. From the samegrowing culture, yeasts were inoculated in flasks withYPG medium, and the appearance of the cells wasexamined microscopically.
2.2. Physiological characterization
Ability to use various carbon compounds for semi-anaerobical growthThe ability to use semi-anaerobically various carboncompounds (fermentation ability) was tested using theDurham technique. In order to test the fermentationability, the yeast strains were cultivated on yeast extractmedia (YE – yeast extract 0.5 g/l) supplemented with50 mM: D- glucose, D-galactose, maltose, lactose, raf-finose or sucrose. The results were recorded by obser-ving the amount of accumulated gas – CO2 – in theDurham tubes, using a positive control (standard strainS. cerevisiae ATTC 26108 with the ability of ferment-ing all the tested carbon sources) and a negative control(all strains cultivated on YE medium with no carbonsource). The fermentation ability of the yeast strainswas evaluated using a scale from (–) to (++++) cor-responding to the amount of CO2 that has been accu-
mulated.
Ability to assimilate carbon compoundsThe ability to aerobically use carbon compounds (assi-milation) was tested using the auxanogram technique, bycultivation of yeast strains on solid YNB medium (YeastNitrogen Base DIFCO) supplemented with: D-glucose,D(+)-galactose, sucrose, raffinose, D(+)-lactose, D-mannitol, ramnose, L(-)-sorbose, D-xylose, D-ribose,
maltose, starch, D-arabinose or L(+)-arabinose. Theresults were recorded by observing the growth com-paratively with a positive control (YNB + D-glucose)and a negative control (YNB with no carbon source).
Ability to assimilate nitrogen compoundsThe ability to assimilate various nitrogen compoundswas tested by cultivating yeast strains on YCB (SIGMA)medium (Yeast Carbon Base – 11.5 g/l) supplementedwith agar 20 g/l, and: sodium nitrite, potassium nitrate,L-lysine and creatinine. The results were recorded byobserving the growth comparatively with a negativecontrol (YCB with no nitrogen source).
Growth at non-permissive temperaturesThis test allows the identification and classification ofyeast species based on their ability to grow at non-per-missive temperatures (e.g. 37oC), comparatively with apermissive temperature (27oC).
Growth on media with high glucose concentrationFrom a fresh grown culture, yeast strains were culti-vated on slants of modified YPGA medium with 50%glucose, and incubated at 28 oC. Growth was recordedafter 4, 6, 12 and 27 days from cultivation, compara-tively with a marker (strains cultivated on regular YPGAmedium, under same growth conditions).
Ability to degrade ureaOnly some yeast species have the ability to degradeurea. The tested yeast strains were cultivated on me-dium supplemented with urea and phenol-red (0.012g/l) as pH indicator, and the ability to degrade ureawas determined after 5 days when the color of themedium changed from yellow to pink.
3. Biodegradation of hydrocarbonsDegradation experiments were performed at laborato-ry scale, using the method described by KANAYAMA etal., 1998 [5]. For the preculture, yeast cells were grownfor 18 hours at 30 oC on slants on YPG medium (yeastextract 1%, peptone 1%, glucose 2%) supplementedwith agar 1%, then on YPG. Cells were washed, resus-pended and dilluted in YNB w/o aminoacids media(DIFCO, 6.7 g/l; pH – 5.2) to obtain a medium opticaldensity (OD) at 640 nm of approximately 0.008. Theinoculum (1 %) was cultured in flasks with stirring, inYNB w/o aminoacids media supplemented with 1% n-decane, n-dodecane, n-tetradecane, n-hexadecane(SIGMA), and 0.05% Tween 80. Growth curves weredrawn for each strain, by measuring the OD at 640 nmafter 12, 24, 36 and 48 hours of incubation.
66
CSUTAK et al.

RESULTS
Conventional taxonomy testsThe macroscopical observations revealed similaritiesbetween strains D1, D2 and D4, which formed redcolonies on YPGA medium, like those of Rhodotorulaspecies. When analysed by microscopy, the threestrains showed round or oval cells with polar budding.D3 and D6 formed creamy, rough colonies with fila-mentous cells with pseudohyphae, similar to Candidacells. These observations were used as starting pointfor further biochemical tests (Table 1).
The fermentation ability (semi-anaerobical growth)was evaluated after 12 days of incubation (Table 2).Strains D1, D2 and D4 did not use any of the testedcarbon compounds, like Rh. glutinis. While D3 resem-bled C. parapsilosis, D6 showed some differences fromC. tropicalis, such as delayed growth on D-glucose andabsence of growth on D-galactose and maltose.
The similarities between D1, D2 and D4 and Rh.glutinis were observed also from the tests regarding the
ability to aerobically assimilate various carbon com-pounds, exception being recorded only for raffinoseand D-ribose. Strain D3 assimilated no sugars, whileD6, unlike C. tropicalis, did not assimilate starch andmaltose (Table 3).
Variability was observed also during nitrogenouscompounds assimilation, for D1, D2 and D4 on nitriteand creatinine.
The strains behaved similarly to the reference strainswhen assessing the ability to grow under stress condi-tions (Table 4).
Urea degradation is a highly selective physiologicaltest. The positive results obtained for D1, D2 and D4strongly supported their classification in Rhodotorulagenus. It was also confirmed the possible belonging ofstrains D3 and D6 to Candida genus (Table 5).
HYDROCARBON DEGRADATION
The ability of yeast strains to degrade hydrocarbonswas studied by observing their growth in special mediacontaining n-alkanes as sole carbon sources.
Preliminary identification and classification of five new Yeast strains isolated from oil-polluted environment
67
Table 1 - Macroscopical and microscopical appearance of yeast strains
Table 2 - Ability of yeast strains D1, D2, D3, D4 and D6 to use various carbon compounds for semi-anaerobic growth

68
CSUTAK et al.
Table 3 - Ability of yeast strains D1, D2, D3, D4 and D6 to assimilate various carbon and nitrogen compounds for aerobic growth
Table 4 - Growth under stress condition
Table 5 - Ability to degrade urea

Preliminary identification and classification of five new Yeast strains isolated from oil-polluted environment
69
Fig. 1 - Hydrocarbon degradation by yeast strains D1 – D6: (a) strain D1; (b) strain D2; (c) strain D3; (d) strain D4; (e) strain D6.
(e)

In general, all the strains grew well during the first12 hours after inoculation. The best results were ob-tained for D1 and D2, on n-decane and n-hexadecane,this period being followed by another 12 hours plateau(Figure 1a and 1b).
Significant differences between the strains, wereobserved within 24 - 48 hours: strains D1 and D3 de-graded with good rates n-decane and n-tetradecane(Figure 1a and 1c); strain D2 grew on all media; thebest alkane uptake was recorded for strain D4 in me-dia with n-decane and n-dodecane (Figure 1d), and forstrain D6 in media with n-dodecane and n-hexadecane(Figure 1e).
The results obtained for the degrading abilities ofD1, D2 and D4 on n-decane, n-tetradecane and n-he-xadecane, matched with those of our previous studies,in which the growing curves of the three strains weremonitored over long periods of time (21 days) [O.Csutak, data not shown].
Analysing the data for D3, we observed a slow slope,especially within the first 24 hours. While for n-hexade-cane and n-dodecane, the degradation rates diminishedafter 35 hours, growth continued on n-decane and n-tetradecane media.
An interesting case was D6, which grew less, butalmost constantly, during the entire period of the ex-periment, especially on n-dodecane and n-hexadecane.Moreover, strain D6 showed good degrading rates of n-hexadecane, when analysed over long periods of time(21 days) [O. Csutak, data not shown].
DISCUSSION
The process of identification of new yeast strains al-ways begins with their morphological and biochemicalobservations comparatively to previously describedtaxonomically identified species. This provides the basisfor a preliminary taxonomical classification.
By comparing the data regarding the macroscopicaland microscopical appearances of our five yeast strainsisolated from polluted environment with the data fromreference scientific literature [2; 7], we found similari-ties with standard laboratory yeast strains belonging toRhodotorula (strains D1, D2 and D4) and Candida(strains D3 and D6) genera.
Further, biochemical analyses were performed fordetermination of physiological characteristics of thestrains: fermentation and assimilation abilities, growthunder stress conditions (temperature and high glucosecontent) and urea degradation. Due to the fact that atpresent, yeast taxonomical identification comprises alarge number of physiological tests (approximately 83),finally, we selected only few considered as the mostselective and representative for establishing the possi-ble belonging of our unknown strains to the three
described species. The obtained results supported stron-gly our identification, mainly for D1, D2, D4 and D6.Nevertheless, some variability was observed regardingthe assimilation abilities of the studied strains and,especially, in the case of D3. We think that one expla-nation could reside in the fact that our studies dealtwith newly isolated strains that might present somemetabolic differences compared with the laboratorystrains described until now. Another explanation wouldbe that, in fact, our taxonomical classification is notcomplete. In order to achieve this, we will perform fur-ther molecular biology analysis.
The ability of yeasts to degrade different hydrocar-bons resides in fact in their ability to use these com-pounds as sole carbon source for aerobic growth. Inorder to determine the degradative abilities of our strains,tests were performed on n-alkanes. Aromatic com-pounds were not used in the present study, knowingthat they are assimilated at rather low rates by yeasts.
We noticed that strains D1, D2 and D4, presentedthe best hydrocarbon-assimilation rates. On the otherhand, the similarities of their growth curves’ profilesmight be determined by some eventual similarities intheir metabolic pathways. This observation correlateswith our finding that both strains might belong to Rh.glutinis, well known for its alkane degrading abilities.
The results obtained for strain D3, which showedlower degrading abilities comparared to D1, D2 andD4, confirmed our preliminary taxonomic identifica-tion, according to which D3 would be a C. parapsilo-sis strain. It is known that this species has rather low-a-ffinity for hydrocarbons, but may be isolated frommicrobial communities from oil-polluted environments,along with other hydrocarbon-biodegrading yeastssuch as Rh. glutinis (strains D1, D2 and D4) and C.tropicalis (D6).
Finally, we showed that, although D6 presentedslow growth during our experiment, it degraded most-ly the n-hexadecane. Since this is mentioned as beingone of the alkanes assimilated preferentially by someCandida strains, we considered our result as a newproof for the possible belonging of D6 to this genus.
CONCLUSIONS
Five yeast strains named D1, D2, D3, D4 and D6,isolated from oil-polluted soil and water, were subjectto preliminary tests in order to establish their possibletaxonomic clasiffication.
The macroscopical and microscopical appearance,as well as the biochemical tests, revealed a great simi-larity between D1, D2 and D4 and Rhodotorula gluti-nis, D3 and Candida parapsilosis, D6 and Candidatropicalis.
70
CSUTAK et al.

Since the yeast strains were isolated from pollutedarea, we also performed tests for assessing their abilityto degrade n-alkanes (n-decane, n-dodecane, n-tetrade-cane, n-hexadecane).
D1 and D2 were the best hydrocarbons consumingstrains on n-decane, n-tetradecane and n-hexadecane,while D4 grew best on media with n-decane and n-dodecane. Their growth patterns pointed out to possi-ble similar degrading abilities and pathways, whichcorelates well to our identification as Rhodotorula glu-tinis strains.
For D3 and D6 the growth was not so spectacularas for D1, D2 and D4, but continuos along the entireexperiment. The resemblings between the curves pro-files, confirmed the idea that both belong to the samegenus, Candida.
REFERENCES
ATLAS R. M, UNTERMAN R. - Bioremediation. In: Manualof industrial microbiology and biotechnology, Sec.Edition, Ed: A. L. Demain, J. E. Davis, R. M. Atlas, G.Cohen, C. L. Hershberger, W-S Hu, D. H. Sherman, R.C. Wilson, J. H. D. Wu. ASM Press, Washington D.C,USA, pp. 666-682, 1999
BARNETT J. A., PAYNE R. W., YARROW D. - Yeasts - char-acteristics and identification, Cambridge UniversityPress, pp. 13 - 60, 1983
BARTH G., GAILLARDIN C. - Nonconventional yeasts inbiotechnology. A handbook, K. Wolf, ed., Springer, pp.313 - 383, 1996
DURHAM D. R., MCNAMEE C. G., STEWART D. B. –Dissimilation of aromatic compounds in Rhodotorulagraminis: biochemical characterization of pleiotropical-ly negative mutants, J. Bacteriol., 160 (2), 771 (1984)
KANAYAMA N., UEDA M., ATOMI H., TANAKA A. ––Genetic evaluation of physiological functions of thiolaseisozymes in the n-alkane-assimilating yeast Candidatropicalis. J. Bacteriol., 180 (3), 690 (1998)
KURTZMAN C. P. – Molecular taxonomy of the yeasts,Yeast, 10, 1727 (1994).
LODDER J. - The yeasts. A taxonomic study, North -Holland Publishing Company, pp. 1-54, 1970
MACGILLIVRAY A. R., SHIARIS M. P. – Biotransformationof polycyclic aromatic hydrocarbons by yeasts isolatedform coastal sediments, Appl. Env. Microbiol., 59 (5),1613 (1993)
MAUERSBERGER S., OHKUMA M., SCHUNK W-H.,TAKAGI M. – Chapter 12: Candida maltosa. In: Non-con-ventional yeasts in biotechnology. A handbook., Ed. K.Wolf, Springer-Verlag Berlin, Germany, pp.411-580, 1996
MIDDELHOVEN W. J., COENEN A., KRAAKMAN B.,SOLLEWIJN GELPKE M. D. – Degradation of some phe-nols and hydroxybenzoates by the imperfect ascomyce-tous yeasts Candida parapsilosis and Arxula adeninivo-rans: evidence for an operative gentisate pathway,Antonie van Leeuwenhoek, 62, 181(1992)
MIDDELHOVEN W. J., SPAAIJ F. – Rhodotorula cresolicasp. nov., a cresol-assimilating yeast species isolated fromsoil, Int. J. System. Bacteriol., 47 (2), 324 (1997)
QUEROL A., BARRIO E., RAMON D. – A comparativestudy of different methods of yeast strain characteriza-tion, System. Appl. Microbiol., 15, 439 (1992)
QUEROL A., RAMON D. – The application of moleculartechniques in wine microbiology, Trends in FoodScience and Technology, 7, 73 (1996)
UEDA M., KANAYAMA N., TANAKA A. – Genetic evalua-tion of peroxisomal and cytosolic acetoacetyl-CoA thio-lase isozymes in n-alkane-assimilating diploid yeastCandida tropicalis. Cell Biochem. Biophys., 32, 285(2000)
A. VAMANU, D. ARIZAN, R. ARIZAN, C. POPA – Biologyand technology of yeasts (In Romanian), vol.III, Ed. I.Anghel, Editura Tehnica, pp.218-227, 1993
Preliminary identification and classification of five new Yeast strains isolated from oil-polluted environment
71

BACKGROUND
In different international studies, candidiasis wasthe fungal infection with the highest incidence inimmunosuppressed hosts. Candida spp. cause a vari-ety of clinical syndromes ranging from relativelymucocutaneous infection to life-threatening can-didemia and deeply invasive candidiasis (1-2, 9-15,17-23, 25-26, 31-38). The reported frequency ofCandida infections has increased, with candidemiarepresenting now the fourth most common cause ofnosocomial bloodstream infection in the UnitedStates of America and Western Europe (3, 5, 8, 24,27-29, 30, 39). Among the species of Candida causingnosocomial infection, Candida albicans accounts foralmost 60% of all infections. The increased use ofbroad-spectrum antifungal agents and the recognitionof Candida albicans as a prominent nosocomialpathogen with the potential for developing resistanceto these agents emphasize the need for ongoing sur-veillance of antifungal susceptibility patterns of clini-cal isolates (4, 6-7, 10, 24).
MATERIAL AND METHODS
Starting from the actual data, we decided to evaluateand prove the existence or not (H0 hypothesis) of theconnection between the immunosuppressed host andCandida infection.
Therefore, we have carried out a case-controlstudy, which is an observational study, to prove theexistence or not of an epidemiological connection,and to check the hypothesis worded as a result of clin-ical observations that confirm that there is a linkbetween the immunosuppressed host and Candidainfection. The study was performed in a retrospectiveway, meaning that the immunosuppressed host wasobserved and recorded after the beginning of the dis-ease (16).
As an analytical investigation, we identified twogroups of patients: a. the lot of cases (case group); b.the comparison lot (control group).
For the group of cases we have selected 61 patientshospitalized in the Dermatology Clinic of “N. G.Lupu” Hospital Bucharest, during two years. Allpatients were identified as presenting various Candidainfections with different expressions as mucous, cuta-neous etc.
72
ABSTRACTCandidiasis is one of the fungal infections with the highest incidence in the immuno-suppressed host. The evolution of infection and the increase of antifungal medical drugsresistance could both contribute to the mortality attributable to Candida infection in theimmunosuppressed host. Even though the data from international studies are well known,few studies have been published in Romania on this subject. In the case-control study we demonstrated the link between the immunosuppression andthe presence of Candida infection. Further studies are to be carried out in order to identi-fy more accurately this link and to extend the study to other fungi. There is a need toincrease the microbiological diagnosis use at least at the hospital laboratory level in orderto better identify the real situation of fungal infections and the link between them and theconcrete status of different hosts.Continued surveillance for infections caused by C. albicans and other species of Candidaamong hospitalized patients is recommended. Control of antimicrobial resistance amongnosocomial infections caused by C. albicans and other species of Candida requires ration-al policies for use of both antifungal and antibacterial agents and appropriate surveillancefor the emergence of resistant strains and species.
CASE-CONTROL STUDY TO EVALUATE THE LINK BETWEEN IMMUNOSUPPRESSION AND CANDIDA SPP. INFECTION
Loredana Gabriela Popa1,2, M.I. Popa1, Ioana Ruxandra Mihai1
(1“Carol Davila” University of Medicine and Pharmacy, 2“N. Gh. Lupu” Hospital, Bucharest, Romania)

The control group included patients hospitalized inthe same period of time, being diagnosed with fungalinfections other than Candida spp. infection; the etio-logical agent was represented by a heterogeneous fun-gal group: Pitirosporum orbicularis, Dermatophitonspp. or Geotrichum candidum.
The criteria that supported the diagnosis ofCandida spp. infection were: a. clinical criteria linkedto immunosuppression and b. laboratory criteria.
Criteria linked to the immunosuppression In the group of clinical criteria were included thosediseases and conditions that induce the alteration ofthe immune response (cellular or humoral) allowingthe development of the opportunistic infection and theprogression to disease. We consider, based on the datapresented in literature, that the development of theinfections is favored by the existence of:
- hematological neoplasia (leukemia, Hodgkin lym-phoma etc);
- other cancers;- collagen diseases (rheumatoid polyarthritis, dis-
seminated lupus erythematosus etc);- immunosuppressive treatments (cytotoxic drugs,
cortisone drugs, chemotherapy drugs);- other drug treatments (for longer periods);- radiation therapies (or other causes);- traumas;- repeated hospitalizations;- some invasive diagnostic procedures, either me-
dical or surgical;- malnutrition;- diabetes mellitus;- congenital malformations that alter either totally
the immune response or only some cells involvedin the immune response;
- alcoholism and other situations
Laboratory criteriaWe took into consideration in the group of laboratorycriteria the following:
- neutropenia (<1500/mm3);- functional alterations of the granulocytes (alter-
ation of chemo attraction phagocytes, etc);- lymphopaenia (their relative and absolute value);- the decrease of the LT number (globally and also
lymphocyte subpopulation CD4+, CD8+);- LT functional alterations;- LB number decrease;- LB functional alterations;- quantitative and qualitative alterations of immuno-
globulin (IgG, IgA, IgM).
Other criteria used for patients’ inclusion in thegroups were represented by:
- concomitance of an immunosuppression status,proved by laboratory tests;
- Candida infection without immunosuppression atleast not proved by lab tests;
- lack of Candida infections with immuno-suppression and
- lack of Candida infections without immuno-suppression.
Data collectionIn order to collect the information, we had to accom-plish 2 conditions:
- Use the same methods also for case lots and forthe control lots;
- A high level of accuracy
The information was obtained and registered usingthe patients’ files, through direct examination, inter-viewing and examining the patients. For a few caseswe have done the laboratory examination.
RESULTS
Measuring the associationThe data obtained from the investigation were intro-duced in the 2/2 contingency table.
The frequency of the immunosuppressed patientsin the case lots is f1= 35 / (35+26).
The frequency of the immunosuppressed patientsin the control cases is f2= 17 / (17+25)
The power (strength) of association is measured bythe relative risk (RR) that indicates how much higher isthe risk of Candida infection in the immunosuppressedpatients, compared to immunocompetent ones.
RR = the risk for the Candida infection in theimmunosuppressed patients/ risk of the disease at theimmunocompetent patients; the relative risk representsthe likelihood of disease in exposed individuals rela-tive to those who are not exposed (16, 39).
The relative risk could have certain different values:1: no association>1: there is a link; as the link becomes stronger,
the rate is much higher than 1;<1: the risk factor is actually a protective one.In other words, to prove the association with a cer-
tain risk factor, the relative risk must be higher than 1(but other conditions could be taken into considera-tion in a more powerful study).
Case-control study to evaluate the link between immunosuppression and Candida spp. infection
73

In the case-control study, the relative risk cannot bedirectly calculated because the disease in immuno-suppressed and immunocompetent patients fails to bemeasured. The proof of the association strength is theodds ratio (OR), which represents the value betweentwo probabilities:
OR = the probability of the case exposure / theprobability of the control exposure.
Odds Ratio is similar to the relative risk, which canbe estimated by OR.
The attributable risk: AR% = [(OR-1)/OR] * 100,shows the difference of the risk value at immunosu-ppressed compared to immunocompetent patients. ARis a measure of association that provides informationabout the absolute effect of the exposure or the excessrisk of disease in those exposed compared with thosenonexposed (15-16, 39).
The following results were obtained:F1 = 35 / 61 = 0.57
F2 = 17 / 42 = 0.4
OR = 1.98AR = 49.4 %
DISCUSSION
These data show that the frequency of the immuno-suppressed patients with Candida infection was 0.57compared to 0.4, which is the frequency of the immuno-suppressed patients having no Candida infection.
The power of the association measured by OR,being higher than 1, respectively 1.98, shows that thereis a connection between the immunosuppressed hostand the Candida infection, and the attributable riskindicates how much higher is this risk in the immuno-suppressed (49.4 %) compared to immunocompetentpatients.
LimitsThis study has a certain number of limitations, startingwith the number of controls compared with the numberof cases. It is needed to better ensure the rate betweenthe two groups. One major issue is the interpretationof the potential different types of bias. A major concern
in conducting a case-control study was that cases andcontrols differed in characteristics of exposure other thanthe one that has been targeted for the study (matchingissue). It has not enough statistical power in order tohave statistical significance. Our study was a quite sim-ple one but we did not identify other studies in the na-tional literature for this subject and we considered thatthere is a need to prove this association in Romaniaand not to take into consideration only the data frominternational resources.
CONCLUSION
In conclusion, for these groups of patients, onegroup being diagnosed with Candida infections (61patients), and the group of patients hospitalized duringthe same period of time, in the same hospital, with noCandida infection (42 patients), the link between theimmunosuppression and the presence of Candidainfection was demonstrated. Further studies are to becarried out in order to identify more accurately thislink and to extend the study to other fungi. As identi-fied during our research, there is a need to increase themicrobiological diagnostic use at least at the hospitallaboratory level in order to better identify the real situ-ation of fungal infections and the link between themand the concrete status of different hosts.
Continued surveillance of infections caused by C.albicans and other species of Candida among hospi-talized patients is recommended. The treatment of andprophylaxis for Candida infection have led to theemergence of resistant species and the acquisition ofresistance in previously susceptible species.
Control of antimicrobial resistance among nosoco-mial infections caused by C. albicans and other spe-cies of Candida require rational policies for use of bothantifungal and antibacterial agents and appropriatesurveillance for the emergence of resistant strains andspecies.
REFERENCES
1. Abelson JA, Moore T, Bruckner D et al., 2005,Frequency of fungemia in hospitalized pediatricinpatients over 11 years at a tertiary care institution,Pediatrics, 116, 61-67
74
POPA et al.
Table 1 – Contingency table for cases and controls

2. Abi Said D, E Anaissie, O Lzun et al., 1997, The epi-demiology of hematogenous candidiasis caused bydifferent Candida species, Clin. Infect. Dis., 24,1122–1128
3. Almirante B, Rodriguez D, Park BJ et al., 2005,Epidemiology and predictors of mortality in cases ofCandida bloodstream infection: Results from popu-lation-based surveillance, Barcelona, Spain, from2002 to 2003, J. Clin. Microbiol., 43, 1829-1835
4. Anaissie EJ, SE Vartivarian, 1996, Fluconazole ver-sus amphotericin B in the treatment of hematoge-nous candidiasis: a matched cohort study, Am. J.Med., 101, 170–176
5. Beck-Sague CM, Jarvis WR, 1993, National Noso-comial Infections Surveillance System. Seculartrends in the epidemiology of nosocomial fungalinfections in the United States, 1980-1990., J. Infect.Dis., 167, 1247–1251
6. Bille J, MP Glauser, 1997, Evaluation of the suscep-tibility of pathogenic Candida species to flucona-zole. Eur. J. Clin. Microbiol. Infect. Dis., 16,924–928
7. Blumberg EA, AC Reboli, 1996, Failure of systemicempirical treatment with amphotericin B to preventcandidemia in neutropenic patients with cancer,Clin. Infect. Dis., 22, 462–466
8. Blumberg H, W Jarvis, J Soucie et al., 2001, Risk fac-tors for candidal bloodstream infections in surgicalintensive care unit patients: the NEMIS prospectivemulticenter study. The National Epidemiology ofMycosis Survey, Clin. Infect. Dis., 33, 177–186
9. Boo TW, O’Reilly B, O’Leary J et al., 2005, Candi-daemia in an Irish tertiary referral hospital: epidemi-ology and prognostic factors, Mycoses, 48, 251-259
10. Buchner T, Fegeler W, Bernhardt H et al., 2002,Treatment of severe Candida infections in high-riskpatients in Germany: consensus formed by a panelof interdisciplinary investigators, Eur. J. Clin.Microbiol. Infect. Dis., 21, 337–352
11. Diekema J, Messer S, Brueggemann A et al., 2002,Epidemiology of candidemia: 3-year results fromthe emerging infections and the epidemiology ofIowa organisms study, J. Clin. Microbiol., 40,1298–1302
12. DiNubile MJ, Hille D, Sable CA et al, 2005,Invasive candidiasis in cancer patients: observa-tions from a randomized clinical trial, J. Infect., 50,443-449
13. Edwards J, Bodey G, Bowden R et al., 1997, Inter-national Conference for the Development of a Con-
sensus on the Management and Prevention of SevereCandida Infections, Clin. Infect. Dis., 25, 43-59
14. Fraser VJ, Jones M, J Dunkel et al., 1992, Candi-demia in a tertiary care hospital: epidemiology, riskfactors and predictors of mortality, Clin. Infect.Dis., 15, 414–421
15. Gudlaugsson O, S Gillespie, K Lee et al., 2003,Attributable mortality of nosocomial candidemia,revisited, Clin. Infect. Dis., 37, 1172–1177
16. Hennekens CH, Buring JE, 1987, Epidemiology inmedicine, Lippincott Williams and Wilkins
17. Hung CC, Yang YL, Lauderdale TL et al., 2005, Co-lonization of human immunodeficiency virus-infected outpatients in Taiwan with Candidaspecies, J. Clin. Microbiol., 43, 1600-1603
18. Kaiserova E, V Krcmery, 1998, Fungemia in chil-dren with cancer, Pediatr. Infect. Dis. J., 19,844–845
19. Karlowsky J, G Zhanel, K Klym et al., 1997, Candi-demia in a Canadian tertiary care hospital from1976–1996, Diagn. Microbiol. Infect. Dis., 29,5–9
20. Krcmery V, J Barnes, 2002, Non-albicans Candidaspp. causing fungaemia: pathogenicity and antifun-gal resistance, J. Hosp. Infection, 4, 243-260
21. Krcmery V, M Mrazova, A Kunova, 1999,Nosocomial candidaemias due to species otherthan Candida albicans in cancer patients.Aetiology, risk factors, and outcome of 45 episodeswithin 10 years in a single cancer institution,Support Care Cancer, 7, 428–431
22. Martin D, Persat F, Piens MA et al., 2005, Candidaspecies distribution in bloodstream cultures inLyon, France, 1998-2001, Eur. J. Clin. Microbiol.Inf. Dis., 24, 329-333
23. McDonald L, C Baker, C Chenoweth, 1998, Riskfactors for candidemia in a children’s hospital,Clin. Infect. Dis., 26, 642–645
24. Miller L, R Hajjeh, J Edwards, 2001, Estimating thecost of nosocomial candidemia in the UnitedStates, Clin. Infect. Dis., 32, 1110
25. Nucci M, Marr KA, 2005, Emerging fungal dis-eases, Clin. Inf. Dis., 41, 521-526
26. Pasqualotto AC, Nedel WL, Machado TS et al.,2005, A comparative study of risk factors and out-come among outpatient-acquired and nosocomialcandidaemia, J. Hosp. Inf., 60, 129-134
Case-control study to evaluate the link between immunosuppression and Candida spp. infection
75

27. Pfaller MA, Jones RN, Messer SA et al., 1997,National surveillance of nosocomial blood streaminfection due to species of Candida other thanCandida albicans: frequency of occurrence andantifungal susceptibility in the SCOPE Program,Diagn. Microbiol. Infect. Dis., 29, 121–129
28. Pfaller MA, R Jones, G Doern et al., 1999, Interna-tional surveillance of blood stream infections dueto Candida species in the European SENTRYProgram: species distribution and antifungal sus-ceptibility including the investigational triazole andechinocandin agents, Diagn. Microbiol. Infect.Dis., 35, 19-25
29. Pfaller MA, R Jones, S Messer et al., 1999, NationalSurveillance of Nosocomial Blood Stream InfectionDue to Candida albicans: Frequency of Occur-rence and Antifungal Susceptibility in the SCOPEProgram, J. Inf. Dis., 169, 218-225
30. Rangel-Frausto M, Wiblin T, Blumberg H et al.,1999, National epidemiology of mycoses survey(NEMIS): variations in rates of bloodstream infec-tions due to Candida species in seven surgical inten-sive care units and six neonatal intensive care units,Clin. Infect. Dis., 29, 253–258
31. Richardson MD, 2005, Changing patterns andtrends in systemic fungal infections, J. Antimicrob.Chemotherapy, 56, Suppl. 1, 5-11
32. Sims CR, L Ostrosky-Zeichner, JH Rex, 2005,Invasive Candidiasis in ImmunocompromisedHospitalized Patients, Arch. Med. Res., 36, 660-71
33. Soub H, W Estinoso, 1997, Hospital acquired can-didaemia experience from a developing country, J.Hosp. Infect., 35, 141–147.
34. Trick W., Fridkin S, Edwards J et al., 2002, Seculartrend of hospital-acquired candidemia among inten-sive care unit patients in the United States during1989–1999, Clin. Infect. Dis., 35, 627–630
35. Tumbarello M, E Tacconelli, G Gaetano K et al.,1999, Candidemia in HIV-infected subjects, Eur. J.Clin. Microbiol. Infect. Dis., 18, 478–483
36. Voss A, JL le Noble, FM Lunel et al., 1997, Candi-demia in intensive care unit: patients risk factors formortality, Infection, 25, 8–15
37. Walsh T, Rex H, 2002, All catheter-related can-didemia is not the same: assessment of the balancebetween the risks and benefits of removal of vascu-lar catheters, Clin. Infect. Dis., 34, 600–602
38. Wingard JR, 1994, Infections due to resistantCandida species in patients with cancer who are
receiving chemotherapy, Clin. Infect. Dis., 19,49–53
39. Zaoutis TE, Argon J, Chu J et al., 2005, The epi-demiology and attributable outcomes of candidemiain adults and children hospitalized in the UnitedStates: A propensity analysis, Clin. Inf. Dis., 41,1232-1239
76
POPA et al.
Copyright © 2022 FDOKUMEN




















