
Human MCRS2, a cell-cycle-dependent protein, associates with LPTS/PinX1 and reduces the telomere...
8
Human MCRS2, a cell-cycle-dependent protein, associates with LPTS/PinX1 and reduces the telomere length Hai Song, a Yiliang Li, a Guoyuan Chen, a Zhen Xing, a Jing Zhao, a Kazunari K. Yokoyama, b Tsaiping Li, a and Mujun Zhao a, * a State Key Laboratory of Molecular Biology, Institute of Biochemistry and Cell Biology, Shanghai Institutes for Biological Sciences, Chinese Academy of Sciences, 320 Yue Yang Road, Shanghai 200031, China b RIKEN Tsukuba Institute, 3-1-1 Koyadai, Tsukuba, Ibaraki 305-0074, Japan Received 25 February 2004 Abstract Human LPTS/PinX1 is a telomerase-inhibitory protein, which binds to the telomere protein Pin2/TRF1 and the catalytic subunit hTERT of telomerase. To explore the proteins that might be involved in the telomerase pathway, we performed a yeast two-hybrid screening with LPTS/PinX1 as the bait. A novel gene, MCRS2, encoding for an isoform of MCRS1/p78 and MSP58 was isolated. The expression of MCRS2 protein is cell-cycle dependent, accumulating in the very early S phase. MCRS2 interacts with LPTS/ PinX1 in vitro, in vivo and colocalizes with LPTS/PinX1 in cells. MCRS2 and its amino terminus inhibit telomerase activity in vitro and long-term overexpression of MCRS2 in SMMC-7721 cells results in a gradual and progressive shortening of telomeres. Our findings suggest that MCRS2 might be a linker between telomere maintenance and cell-cycle regulation. Ó 2004 Elsevier Inc. All rights reserved. Keywords: Nucleolar protein; Telomerase inhibitor; Cell-cycle regulation; Yeast two-hybrid screening; cDNA Telomeres, the ends of linear chromosomes, have multiple functions within the cell including maintaining genomic stability, preventing end-to-end fusions, and allowing cells escaping from crisis [1,2]. In mammals, the length, structure, and function of telomeres have been proposed to contribute to cellular and organism phe- notypes associated with cancer and aging [3,4]. Telo- meres consist of simple DNA repeats and associated proteins. In human, telomere maintenance is mainly performed by a specific reverse transcriptase, telome- rase, which is a ribonucleoprotein composed of a cata- lytic subunit, hTERT, and a 451 nt long RNA, which acts as a template for the addition of a short repetitive motif (TTAGGG) n on the 3 0 end of telomere [5]. Com- pelling evidences indicate that the ability of telomerase to elongate telomeres is regulated by other factors. In mammals, the telomeres are regulated by double-stran- ded telomere DNA binding protein TRF1 and TRF2 [6,7] and their interacting proteins, tankyrase, Tin2, PinX1, and hRap1 [8–11]. The Pot1, a single-stranded telomere DNA binding protein, is also involved in the telomere regulation [12,13]. LPTS, identified by allelic-loss mapping and posi- tional candidate cloning, is a putative human tumor- suppressor gene located on chromosome 8p23 [14]. PinX1 gene encodes a Pin2/TRF1-interacting protein and its coding sequence is almost same as LPTS gene [15,16]. LPTS and PinX1 are likely generated from the same gene. Both of LPTS and PinX1 have been dem- onstrated to be a potent telomerase inhibitor. Overex- pression of LPTS/PinX1 suppresses telomerase activity, results in shortened telomeres, and induces crisis in cells [10,15]. Depletion of LPTS/PinX1 increases telomerase activity and increases tumorigenicity [10]. In an attempt to characterize other proteins that might be involved in the regulation of telomeres, we performed a yeast two-hybrid screening to identify LPTS/PinX1-associated proteins. We isolated a cDNA, MCRS2, which was a novel isoform of human micro- spherule protein 1 (MCRS1)/p78 [17] and 58-kDa mi- crospherule protein MSP58 [18]. Previous reports have * Corresponding author. Fax: +86-21-5492-1011. E-mail address: [email protected] (M. Zhao). 0006-291X/$ - see front matter Ó 2004 Elsevier Inc. All rights reserved. doi:10.1016/j.bbrc.2004.02.166 Biochemical and Biophysical Research Communications 316 (2004) 1116–1123 BBRC www.elsevier.com/locate/ybbrc
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Human MCRS2, a cell-cycle-dependent protein, associates with LPTS/PinX1 and reduces the telomere...

Biochemical and Biophysical Research Communications 316 (2004) 1116–1123
BBRCwww.elsevier.com/locate/ybbrc
Human MCRS2, a cell-cycle-dependent protein, associateswith LPTS/PinX1 and reduces the telomere length
Hai Song,a Yiliang Li,a Guoyuan Chen,a Zhen Xing,a Jing Zhao,a
Kazunari K. Yokoyama,b Tsaiping Li,a and Mujun Zhaoa,*
a State Key Laboratory of Molecular Biology, Institute of Biochemistry and Cell Biology, Shanghai Institutes for Biological Sciences,
Chinese Academy of Sciences, 320 Yue Yang Road, Shanghai 200031, Chinab RIKEN Tsukuba Institute, 3-1-1 Koyadai, Tsukuba, Ibaraki 305-0074, Japan
Received 25 February 2004
Abstract
Human LPTS/PinX1 is a telomerase-inhibitory protein, which binds to the telomere protein Pin2/TRF1 and the catalytic subunit
hTERT of telomerase. To explore the proteins that might be involved in the telomerase pathway, we performed a yeast two-hybrid
screening with LPTS/PinX1 as the bait. A novel gene, MCRS2, encoding for an isoform of MCRS1/p78 and MSP58 was isolated.
The expression of MCRS2 protein is cell-cycle dependent, accumulating in the very early S phase. MCRS2 interacts with LPTS/
PinX1 in vitro, in vivo and colocalizes with LPTS/PinX1 in cells. MCRS2 and its amino terminus inhibit telomerase activity in vitro
and long-term overexpression of MCRS2 in SMMC-7721 cells results in a gradual and progressive shortening of telomeres. Our
findings suggest that MCRS2 might be a linker between telomere maintenance and cell-cycle regulation.
� 2004 Elsevier Inc. All rights reserved.
Keywords: Nucleolar protein; Telomerase inhibitor; Cell-cycle regulation; Yeast two-hybrid screening; cDNA
Telomeres, the ends of linear chromosomes, have
multiple functions within the cell including maintaininggenomic stability, preventing end-to-end fusions, and
allowing cells escaping from crisis [1,2]. In mammals, the
length, structure, and function of telomeres have been
proposed to contribute to cellular and organism phe-
notypes associated with cancer and aging [3,4]. Telo-
meres consist of simple DNA repeats and associated
proteins. In human, telomere maintenance is mainly
performed by a specific reverse transcriptase, telome-rase, which is a ribonucleoprotein composed of a cata-
lytic subunit, hTERT, and a 451 nt long RNA, which
acts as a template for the addition of a short repetitive
motif (TTAGGG)n on the 30 end of telomere [5]. Com-
pelling evidences indicate that the ability of telomerase
to elongate telomeres is regulated by other factors. In
mammals, the telomeres are regulated by double-stran-
ded telomere DNA binding protein TRF1 and TRF2[6,7] and their interacting proteins, tankyrase, Tin2,
* Corresponding author. Fax: +86-21-5492-1011.
E-mail address: [email protected] (M. Zhao).
0006-291X/$ - see front matter � 2004 Elsevier Inc. All rights reserved.
doi:10.1016/j.bbrc.2004.02.166
PinX1, and hRap1 [8–11]. The Pot1, a single-stranded
telomere DNA binding protein, is also involved in thetelomere regulation [12,13].
LPTS, identified by allelic-loss mapping and posi-
tional candidate cloning, is a putative human tumor-
suppressor gene located on chromosome 8p23 [14].
PinX1 gene encodes a Pin2/TRF1-interacting protein
and its coding sequence is almost same as LPTS gene
[15,16]. LPTS and PinX1 are likely generated from the
same gene. Both of LPTS and PinX1 have been dem-onstrated to be a potent telomerase inhibitor. Overex-
pression of LPTS/PinX1 suppresses telomerase activity,
results in shortened telomeres, and induces crisis in cells
[10,15]. Depletion of LPTS/PinX1 increases telomerase
activity and increases tumorigenicity [10].
In an attempt to characterize other proteins that
might be involved in the regulation of telomeres, we
performed a yeast two-hybrid screening to identifyLPTS/PinX1-associated proteins. We isolated a cDNA,
MCRS2, which was a novel isoform of human micro-
spherule protein 1 (MCRS1)/p78 [17] and 58-kDa mi-
crospherule protein MSP58 [18]. Previous reports have

H. Song et al. / Biochemical and Biophysical Research Communications 316 (2004) 1116–1123 1117
demonstrated that MCRS1/p78 and MSP58 arecell-cycle-regulated nucleolar proteins and interact
with a nucleolar protein p120 and transcriptional
repressor Daxx [17–19]. In this study, we character-
ized both of biochemical and functional interactions
between MCRS2 and LPTS/PinX1. We describe here
that MCRS2 binds to LPTS/PinX1, the expression of
MCRS2 protein is cell-cycle dependent, accumulating in
the early S phase, and MCRS2 localizes to the nucleolusand telomeres and shortens telomere length.
Materials and methods
Yeast two-hybrid screen and plasmids. The cDNA of human LPTS/
PinX1 was inserted into the pGilda vector (Clontech) to produce the
bait for yeast two-hybrid screening. The LexA-LPTS/PinX1 construct
was used for screening with a human brain cDNA library (Clontech).
The yeast two-hybrid screen was performed as described in the manual
from Clontech. The positive clones were then subjected to DNA se-
quence analysis. MCRS2 was cloned into the vectors of pCMV-Tag
2B, pEGFP-C2, and pET-30a. LPTS/PinX1 was cloned into the vec-
tors of pCS2-RFP and pGEX-4T-1.
Expression and purification of recombinant proteins. To generate
GST-LPTS/PinX1 and GST- or 6His-MCRS2 fusion proteins, the
fragments encoding LPTS or MCRS2 and their different portions were
cloned into the pGEX-4T-1 or pET-30a vector, respectively. The
produced fusion proteins were purified by glutathione (Clontech) or
Ni–NTA (Shenergy Biocolor) agarose column as described in manual.
Production and affinity purification of antibodies. Rabbit polyclonal
antibodies were raised against full-length MCRS2 protein fused to
N-terminal 6His tag. Antibodies were affinity-purified by adsorption to
6His-MCRS2 proteins, conjugated to CNBr-activated Sepharose
(Amersham). The specificity of the antibodies was confirmed by
Western blotting.
Synchronization of SMMC-7721 cells. SMMC-7721 (hepatocellular
carcinoma cell line) cells (1� 105 cells/ 35mm dish) were incubated in
the medium supplemented with 2mM thymidine for 24 h at 37 �C,washed twice with PBS (20mM potassium phosphate, pH 7.4, 150mM
NaCl), and incubated for 12 h in regular medium. Cells were then in-
cubated again in the medium containing 2mM thymidine for another
24 h and released from the block by rinsing the cells twice with PBS.
Identification of MCRS2 and MCRS1 by RT-PCR. Total RNA was
extracted with Trizol reagent (Invitrogen) and the RT reaction was
performed in a 20 ll reaction mixture that contained 1 lg total RNA
and 1 ll AMV reverse transcriptase (TaKaRa). PCR was performed
using primers M1 (50-ATGACACGTGGCACCGGGGG-30) and M2
(50-AAGAATTTGAACGGCTGCTC-30) by reference to the sequence
of MCRS2 nt 1–246. The PCR was carried out in a volume of 25 ll for4min at 94 �C for initial denaturing, followed by 35 cycles of 94 �C for
30 s, 56 �C for 30 s, and 72 �C for 30 s. Final extension was continued
for 10min. Amplified products were visualized on 1.5% agarose gel
with ethidium bromide.
Colocalization of MCRS2 and LPTS/PinX1. For colocalization
experiments, SMMC-7721 cells were transiently cotransfected with
GFP-MCRS2, or GFP-MCRS2-N (1–261) and RFP-LPTS/PinX1.
SMMC-7721 cells were plated in 6-well plates that contained coverslips
and transfections were carried with Lipofectamine Plus (Invitrogen).
After 36–48 h, the cells were washed three times with PBS and then
fixed for 20min in 4% paraformaldehyde. The nuclei were stained by
40,60-diamidino-2-phenylindole (DAPI) for 10min. After several wa-
shes, the cells were viewed under a fluorescence microscope (Olympus
BX51, Japan).
GST pulldown, immunoprecipitation, and Western blotting. The
relevant proteins used for GST pulldown, immunoprecipitation, and
Western blotting were transiently expressed in the 293T cells or
translated in vitro using the TNT kit (Promega) in the presence of
[35S]Met. To prepare cell lysate, the cells were washed twice in PBS and
incubated at 2� 106 cells/ml in isotonic lysis buffer (20mM Tris–HCl,
pH 7.5, 150mM NaCl, 1mM EDTA, 0.5% Nonidet P-40, 1mM
phenylmethylsulfonyl fluoride (PMSF), 1mM aprotinin, and 1mM
leupeptin) for 0.5 h at 4 �C, after centrifugation the cellular superna-
tants were used for the assays. For immunoprecipitation or GST
pulldown experiment, cellular supernatants or proteins were incubated
with antibodies, or with 10 lg GST fusion proteins for 2 h at 4 �Cbefore addition of 15ll of protein A or glutathione–agarose beads
(Pharmacia), followed by incubation at 4 �C for 2 h. Beads were then
washed three times with lysis buffer and resuspended in 30 ll SDS
sample buffer (50mM Tris–HCl, pH 6.8, 100mM DTT, 2% sodium
dodecyl sulfate (SDS), 20% glycerol, and 0.2mg/ml bromophenol
blue). Ten microliters of each sample was subjected to SDS–PAGE
followed by Western blotting using appropriate antibodies. Proteins
were visualized by ECL.
Telomere length assay and measurement of telomerase activity. The
telomere length was measured by TRF length assay as described [20].
Briefly, genomic DNA was digested with HinfI and RsaI and sepa-
rated with 0.7% agarose. The gel was denatured in 0.5M NaOH/
1.5M NaCl for 30min, neutralized in 0.5M Tris–HCl/3M NaCl (pH
7.5) for 30min, and analyzed by Southern blotting using a 32P-labeled
(TTAGGG)6 DNA probe. The average TRF length versus PD
number was calculated by quantifying the hybridization signals using
Gen-Pro Analyzer software. For assaying telomerase activity,
SMMC-7721 cells were lysed in a lysis buffer and telomerase-con-
taining fraction was prepared, followed by assaying the telomerase
activity using the TRAP assay [21]. To examine the effect of MCRS2
on telomerase activity in vitro, 6His-MCRS2 or GST-MCRS2-N
(1–261) fusion proteins were incubated with telomerase for 15min at
4 �C before subjecting to TRAP assay. The telomerase products were
separated on 10% polyacrylamide gel, which was stained by silver
staining.
Results
Identification of MCRS2
To identify LPTS/PinX1-interacting proteins, we
screened a human brain yeast two-hybrid cDNA library
using LPTS as the bait. One of positive clones was
matched to the C-terminus of MCRS1/p78 or 58-kDamicrospherule protein (MSP58) [17,18]. In order to ob-
tain the full-length cDNA of MCRS1, we performed
PCR with primers that were based on the sequence in-
formation of MCRS1. After PCR amplification from a
liver cDNA library, a full-length cDNA encoding a
novel 475-amino acid protein was yielded. After se-
quence analysis, we found that this novel protein was a
short isoform of MCRS1. This protein was then namedas MCRS2. An overview of the cDNA sequences of
MCRS2, MCRS1, and MSP58 is shown in Fig. 1A. The
alignment of MCRS2 with MCRS1, MSP58, and mouse
MSP58 proteins (Fig. 1B) shows that the MCRS2 pro-
tein shares sequence identity in the C-terminal 448 res-
idues with all compared mammalian sequences, but
lacks 59 amino acids in N-terminus comparing with
MCRS1. Therefore, MCRS2 presumably represents ashorter isoform of MCRS1.
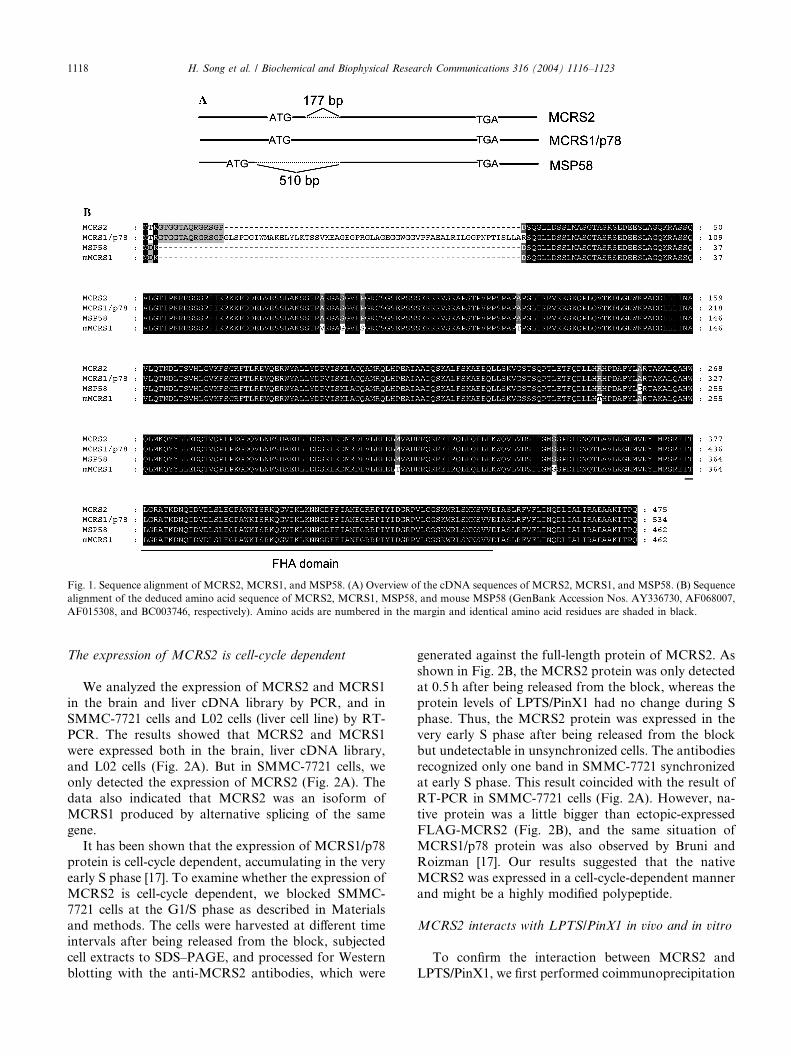
Fig. 1. Sequence alignment of MCRS2, MCRS1, and MSP58. (A) Overview of the cDNA sequences of MCRS2, MCRS1, and MSP58. (B) Sequence
alignment of the deduced amino acid sequence of MCRS2, MCRS1, MSP58, and mouse MSP58 (GenBank Accession Nos. AY336730, AF068007,
AF015308, and BC003746, respectively). Amino acids are numbered in the margin and identical amino acid residues are shaded in black.
1118 H. Song et al. / Biochemical and Biophysical Research Communications 316 (2004) 1116–1123
The expression of MCRS2 is cell-cycle dependent
We analyzed the expression of MCRS2 and MCRS1
in the brain and liver cDNA library by PCR, and in
SMMC-7721 cells and L02 cells (liver cell line) by RT-
PCR. The results showed that MCRS2 and MCRS1
were expressed both in the brain, liver cDNA library,
and L02 cells (Fig. 2A). But in SMMC-7721 cells, weonly detected the expression of MCRS2 (Fig. 2A). The
data also indicated that MCRS2 was an isoform of
MCRS1 produced by alternative splicing of the same
gene.
It has been shown that the expression of MCRS1/p78
protein is cell-cycle dependent, accumulating in the very
early S phase [17]. To examine whether the expression of
MCRS2 is cell-cycle dependent, we blocked SMMC-7721 cells at the G1/S phase as described in Materials
and methods. The cells were harvested at different time
intervals after being released from the block, subjected
cell extracts to SDS–PAGE, and processed for Western
blotting with the anti-MCRS2 antibodies, which were
generated against the full-length protein of MCRS2. As
shown in Fig. 2B, the MCRS2 protein was only detected
at 0.5 h after being released from the block, whereas the
protein levels of LPTS/PinX1 had no change during S
phase. Thus, the MCRS2 protein was expressed in the
very early S phase after being released from the block
but undetectable in unsynchronized cells. The antibodies
recognized only one band in SMMC-7721 synchronizedat early S phase. This result coincided with the result of
RT-PCR in SMMC-7721 cells (Fig. 2A). However, na-
tive protein was a little bigger than ectopic-expressed
FLAG-MCRS2 (Fig. 2B), and the same situation of
MCRS1/p78 protein was also observed by Bruni and
Roizman [17]. Our results suggested that the native
MCRS2 was expressed in a cell-cycle-dependent manner
and might be a highly modified polypeptide.
MCRS2 interacts with LPTS/PinX1 in vivo and in vitro
To confirm the interaction between MCRS2 and
LPTS/PinX1, we first performed coimmunoprecipitation

Fig. 2. The expression of MCRS2 is cell-cycle dependent. (A) The expression of MCRS2 and MCRS1 was analyzed by PCR in human liver and brain
cDNA library, and by RT-PCR in L02 and SMMC-7721cells. The presence of the 246 and 423bp fragments of amplified DNA indicated MCRS2
and MCRS1, respectively. (B) SMMC-7721 cell extracts were prepared from synchronized cells at 0, 0.5, 1, 3, 5, and 7 h after released from block at
the G1/S phase. MCRS2 was detected by Western blotting with anti-MCRS2 antibodies. The FLAG-MCRS2 was loaded as a positive control and
the empty vector as a negative control. The protein levels of LPTS/PinX1 were maintained through the S phage.
H. Song et al. / Biochemical and Biophysical Research Communications 316 (2004) 1116–1123 1119
assay. We transfected 293T cells with FLAG-MCRS2
expression construct and subjected cell extracts to im-
munoprecipitation with anti-LPTS antibodies. We de-tected FLAG-MCRS2 protein in immunoprecipitates,
but not in the preimmune serum control (Fig. 3A). Next
we examined the interaction betweenMCRS2 and LPTS/
PinX1 in vitro with GST pulldown experiment. GST-
LPTS, but not GST, precipitated 35S-MCRS2 synthe-
sized in a rabbit reticulocyte lysate (RRL) (Fig. 3B). This
result indicated that MCRS2 interacted with LPTS/
PinX1 in vitro. To identify the region in LPTS/PinX1that interacted with MCRS2, we expressed FLAG-
MCRS2 protein in 293T cells and subjected cell extracts
to GST pulldown experiment. The fragments of both the
C-terminal 133–328 amino acids as GST-LPTS-C and
the N-terminal 132 amino acids as GST-LPTS-N bound
to the protein of FLAG-MCRS2 (Fig. 3C).
Previous report demonstrated that LTPS/PinX1 lo-
calized at telomeres and in the nucleolus [10]. We nextexamined whether MCRS2 and LPTS/PinX1 were co-
localized at telomeres and in the nucleolus in cells. We
cotransfected SMMC-7721 cells with GFP-MCRS2 and
RFP-LPTS expression constructs and their subcellular
localizations were observed directly. MCRS2 and LPTS/
PinX1 colocalized in the nucleus, especially in nucleoli
and weakly at telomere speckles (Fig. 3E). Next, we
found that the N-terminal 1–261 amino acids of MCRS2were sufficient for telomere localization (Fig. 3E). Thus,
MCRS2 interacted with LPTS/PinX1 in vitro, in vivo
and colocalized with LPTS/PinX1 in cells.
Overexpression of MCRS2 shortens telomere length
Next we examined whether exogenous expression of
MCRS2 has an effect on telomere dynamics. We estab-
lished SMMC-7721 cell lines stably expressing FLAG-
MCRS2 or FLAG-MCRS2-N containing N-terminal
1–261 amino acids of MCRS2. The expression levels of
MCRS2 and MCRS2-N in stable cell lines were verifiedwith Western blotting using anti-FLAG antibody (Figs.
4A and B) and the SMMC-7721 cells transfected with
empty vector were loaded as a control. We found that
in FLAG-MCRS2 (Fig. 4C) and FLAG-MCRS2-N
(Fig. 4D) expressing cells, telomeres were slowly and
progressively shortened during passages, while control
cells maintained with the same TRF length, about 10 kb
on an average. The loss of telomere sequences was evi-dent from the data of the shortening of TRF length and
reduction of the TTAGGG hybridization signals. The
average TRF length versus PD number is presented in
Figs. 4E and F. TRF length in MCRS2 stable cells was
significantly shortened at the early population doublings
(PD). The 0 PD cells given in this experiment in
fact have been passaged for about 30 PD since the sin-
gle cell had to expand to the sufficient number forcharacterization.
MCRS2 associates with hTERT and has a telomerase-
inhibitory activity in vitro
The observations that MCRS2 interacted with LPTS
(see above) and that LPTS/PinX1 binds directly to
hTERT [10] suggest that MCRS2 might also associate
with hTERT. To examine this possibility, we investi-
gated the interaction between MCRS2 and hTERT in
vivo. The 293T cells transiently cotransfected with
FLAG-MCRS2 and GFP-hTERT expression con-
structs, then subjected cell extracts to immunoprecipi-tation with anti-FLAG antibody. Anti-FLAG antibody,
but not normal mouse serum, immunoprecipitated
GFP-hTERT (Fig. 5A) indicated that MCRS2 had
formed a stable complex with hTERT in cells. Next we
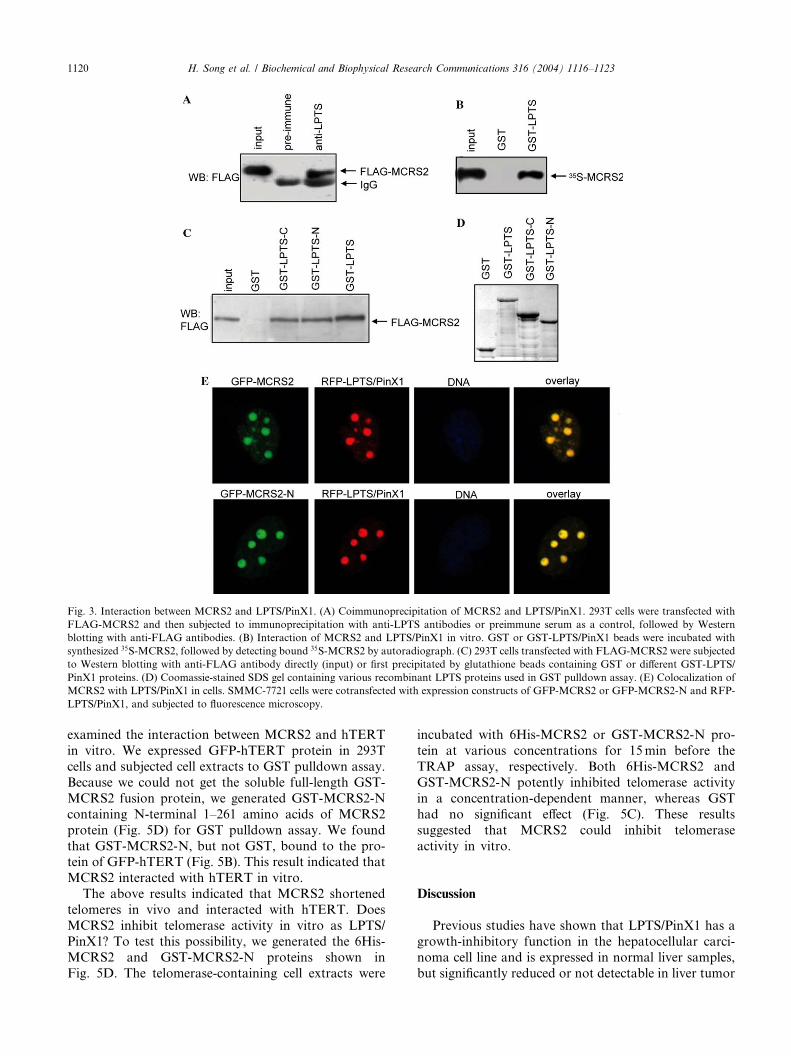
Fig. 3. Interaction between MCRS2 and LPTS/PinX1. (A) Coimmunoprecipitation of MCRS2 and LPTS/PinX1. 293T cells were transfected with
FLAG-MCRS2 and then subjected to immunoprecipitation with anti-LPTS antibodies or preimmune serum as a control, followed by Western
blotting with anti-FLAG antibodies. (B) Interaction of MCRS2 and LPTS/PinX1 in vitro. GST or GST-LPTS/PinX1 beads were incubated with
synthesized 35S-MCRS2, followed by detecting bound 35S-MCRS2 by autoradiograph. (C) 293T cells transfected with FLAG-MCRS2 were subjected
to Western blotting with anti-FLAG antibody directly (input) or first precipitated by glutathione beads containing GST or different GST-LPTS/
PinX1 proteins. (D) Coomassie-stained SDS gel containing various recombinant LPTS proteins used in GST pulldown assay. (E) Colocalization of
MCRS2 with LPTS/PinX1 in cells. SMMC-7721 cells were cotransfected with expression constructs of GFP-MCRS2 or GFP-MCRS2-N and RFP-
LPTS/PinX1, and subjected to fluorescence microscopy.
1120 H. Song et al. / Biochemical and Biophysical Research Communications 316 (2004) 1116–1123
examined the interaction between MCRS2 and hTERT
in vitro. We expressed GFP-hTERT protein in 293T
cells and subjected cell extracts to GST pulldown assay.
Because we could not get the soluble full-length GST-
MCRS2 fusion protein, we generated GST-MCRS2-N
containing N-terminal 1–261 amino acids of MCRS2
protein (Fig. 5D) for GST pulldown assay. We found
that GST-MCRS2-N, but not GST, bound to the pro-tein of GFP-hTERT (Fig. 5B). This result indicated that
MCRS2 interacted with hTERT in vitro.
The above results indicated that MCRS2 shortened
telomeres in vivo and interacted with hTERT. Does
MCRS2 inhibit telomerase activity in vitro as LPTS/
PinX1? To test this possibility, we generated the 6His-
MCRS2 and GST-MCRS2-N proteins shown in
Fig. 5D. The telomerase-containing cell extracts were
incubated with 6His-MCRS2 or GST-MCRS2-N pro-
tein at various concentrations for 15min before the
TRAP assay, respectively. Both 6His-MCRS2 and
GST-MCRS2-N potently inhibited telomerase activity
in a concentration-dependent manner, whereas GST
had no significant effect (Fig. 5C). These results
suggested that MCRS2 could inhibit telomerase
activity in vitro.
Discussion
Previous studies have shown that LPTS/PinX1 has a
growth-inhibitory function in the hepatocellular carci-
noma cell line and is expressed in normal liver samples,
but significantly reduced or not detectable in liver tumor
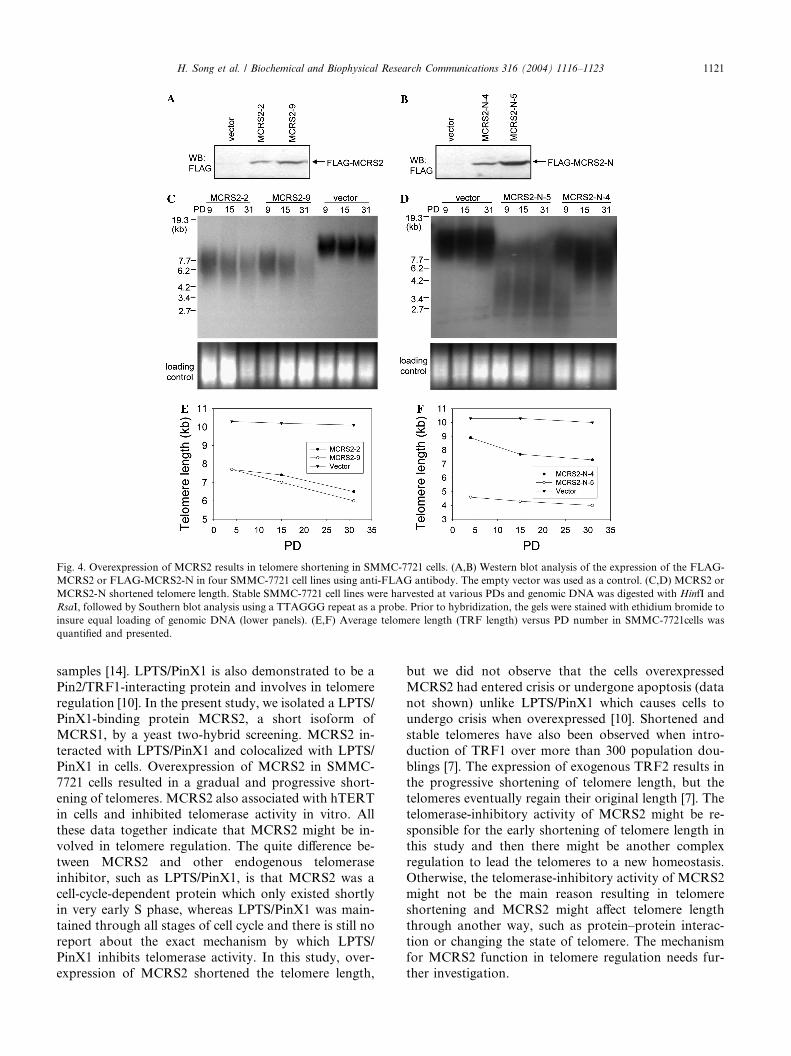
Fig. 4. Overexpression of MCRS2 results in telomere shortening in SMMC-7721 cells. (A,B) Western blot analysis of the expression of the FLAG-
MCRS2 or FLAG-MCRS2-N in four SMMC-7721 cell lines using anti-FLAG antibody. The empty vector was used as a control. (C,D) MCRS2 or
MCRS2-N shortened telomere length. Stable SMMC-7721 cell lines were harvested at various PDs and genomic DNA was digested with HinfI and
RsaI, followed by Southern blot analysis using a TTAGGG repeat as a probe. Prior to hybridization, the gels were stained with ethidium bromide to
insure equal loading of genomic DNA (lower panels). (E,F) Average telomere length (TRF length) versus PD number in SMMC-7721cells was
quantified and presented.
H. Song et al. / Biochemical and Biophysical Research Communications 316 (2004) 1116–1123 1121
samples [14]. LPTS/PinX1 is also demonstrated to be a
Pin2/TRF1-interacting protein and involves in telomere
regulation [10]. In the present study, we isolated a LPTS/
PinX1-binding protein MCRS2, a short isoform of
MCRS1, by a yeast two-hybrid screening. MCRS2 in-
teracted with LPTS/PinX1 and colocalized with LPTS/
PinX1 in cells. Overexpression of MCRS2 in SMMC-7721 cells resulted in a gradual and progressive short-
ening of telomeres. MCRS2 also associated with hTERT
in cells and inhibited telomerase activity in vitro. All
these data together indicate that MCRS2 might be in-
volved in telomere regulation. The quite difference be-
tween MCRS2 and other endogenous telomerase
inhibitor, such as LPTS/PinX1, is that MCRS2 was a
cell-cycle-dependent protein which only existed shortlyin very early S phase, whereas LPTS/PinX1 was main-
tained through all stages of cell cycle and there is still no
report about the exact mechanism by which LPTS/
PinX1 inhibits telomerase activity. In this study, over-
expression of MCRS2 shortened the telomere length,
but we did not observe that the cells overexpressed
MCRS2 had entered crisis or undergone apoptosis (data
not shown) unlike LPTS/PinX1 which causes cells to
undergo crisis when overexpressed [10]. Shortened and
stable telomeres have also been observed when intro-
duction of TRF1 over more than 300 population dou-
blings [7]. The expression of exogenous TRF2 results inthe progressive shortening of telomere length, but the
telomeres eventually regain their original length [7]. The
telomerase-inhibitory activity of MCRS2 might be re-
sponsible for the early shortening of telomere length in
this study and then there might be another complex
regulation to lead the telomeres to a new homeostasis.
Otherwise, the telomerase-inhibitory activity of MCRS2
might not be the main reason resulting in telomereshortening and MCRS2 might affect telomere length
through another way, such as protein–protein interac-
tion or changing the state of telomere. The mechanism
for MCRS2 function in telomere regulation needs fur-
ther investigation.
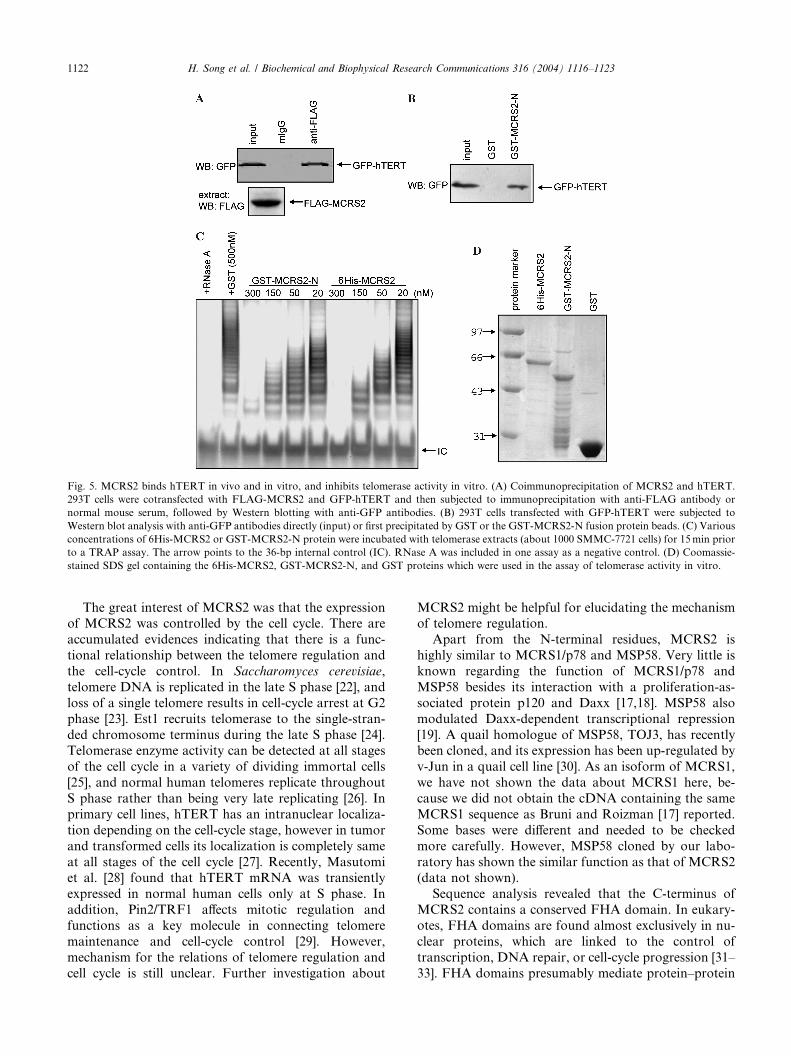
Fig. 5. MCRS2 binds hTERT in vivo and in vitro, and inhibits telomerase activity in vitro. (A) Coimmunoprecipitation of MCRS2 and hTERT.
293T cells were cotransfected with FLAG-MCRS2 and GFP-hTERT and then subjected to immunoprecipitation with anti-FLAG antibody or
normal mouse serum, followed by Western blotting with anti-GFP antibodies. (B) 293T cells transfected with GFP-hTERT were subjected to
Western blot analysis with anti-GFP antibodies directly (input) or first precipitated by GST or the GST-MCRS2-N fusion protein beads. (C) Various
concentrations of 6His-MCRS2 or GST-MCRS2-N protein were incubated with telomerase extracts (about 1000 SMMC-7721 cells) for 15min prior
to a TRAP assay. The arrow points to the 36-bp internal control (IC). RNase A was included in one assay as a negative control. (D) Coomassie-
stained SDS gel containing the 6His-MCRS2, GST-MCRS2-N, and GST proteins which were used in the assay of telomerase activity in vitro.
1122 H. Song et al. / Biochemical and Biophysical Research Communications 316 (2004) 1116–1123
The great interest of MCRS2 was that the expressionof MCRS2 was controlled by the cell cycle. There are
accumulated evidences indicating that there is a func-
tional relationship between the telomere regulation and
the cell-cycle control. In Saccharomyces cerevisiae,
telomere DNA is replicated in the late S phase [22], and
loss of a single telomere results in cell-cycle arrest at G2
phase [23]. Est1 recruits telomerase to the single-stran-
ded chromosome terminus during the late S phase [24].Telomerase enzyme activity can be detected at all stages
of the cell cycle in a variety of dividing immortal cells
[25], and normal human telomeres replicate throughout
S phase rather than being very late replicating [26]. In
primary cell lines, hTERT has an intranuclear localiza-
tion depending on the cell-cycle stage, however in tumor
and transformed cells its localization is completely same
at all stages of the cell cycle [27]. Recently, Masutomiet al. [28] found that hTERT mRNA was transiently
expressed in normal human cells only at S phase. In
addition, Pin2/TRF1 affects mitotic regulation and
functions as a key molecule in connecting telomere
maintenance and cell-cycle control [29]. However,
mechanism for the relations of telomere regulation and
cell cycle is still unclear. Further investigation about
MCRS2 might be helpful for elucidating the mechanismof telomere regulation.
Apart from the N-terminal residues, MCRS2 is
highly similar to MCRS1/p78 and MSP58. Very little is
known regarding the function of MCRS1/p78 and
MSP58 besides its interaction with a proliferation-as-
sociated protein p120 and Daxx [17,18]. MSP58 also
modulated Daxx-dependent transcriptional repression
[19]. A quail homologue of MSP58, TOJ3, has recentlybeen cloned, and its expression has been up-regulated by
v-Jun in a quail cell line [30]. As an isoform of MCRS1,
we have not shown the data about MCRS1 here, be-
cause we did not obtain the cDNA containing the same
MCRS1 sequence as Bruni and Roizman [17] reported.
Some bases were different and needed to be checked
more carefully. However, MSP58 cloned by our labo-
ratory has shown the similar function as that of MCRS2(data not shown).
Sequence analysis revealed that the C-terminus of
MCRS2 contains a conserved FHA domain. In eukary-
otes, FHA domains are found almost exclusively in nu-
clear proteins, which are linked to the control of
transcription, DNA repair, or cell-cycle progression [31–
33]. FHA domains presumably mediate protein–protein

H. Song et al. / Biochemical and Biophysical Research Communications 316 (2004) 1116–1123 1123
interactions, and it has recently been shown that distinctFHA domains are competent to bind to phosphopeptides
or to peptides in a phosphorylation-dependent manner
[32,33]. Further studies on the function ofMCRS2 will be
crucial to the understanding of the relationship between
cell-cycle progression and telomere regulation.
Acknowledgments
The authors thank Dr. Yanhua Xu for critical comments on the
manuscript. This work was supported by the grants from the National
High Technology Research and Development of China (863 Program)
No. 2001AA221021 and No. 2002BA711A02 and special funds
for Major State Basic Research of China (973 Program), No.
2001CB510205. The nucleotide sequence for MCRS2 has been de-
posited in the GenBank Sequence Database under GenBank Accession
No. AY336730.
References
[1] E.H. Blackburn, Switching and signaling at the telomere, Cell 106
(2001) 661–673.
[2] T. de Lange, Protection of mammalian telomeres, Oncogene 21
(2002) 532–540.
[3] S.H. Kim, P. Kaminker, J. Campisi, Telomeres, aging and cancer:
in search of a happy ending, Oncogene 21 (2002) 503–511.
[4] R.S. Maser, R.A. DePinho, Connecting chromosomes, crisis, and
cancer, Science 297 (2002) 565–569.
[5] C.I. Nugent, V. Lundblad, The telomerase reverse transcriptase:
components and regulation, Genes Dev. 12 (1998) 1073–1085.
[6] B. van Steensel, T. de Lange, Control of telomere length by the
human telomeric protein TRF1, Nature 385 (1997) 740–743.
[7] A. Smogorzewska, B. van Steensel, A. Bianchi, S. Oelmann, M.R.
Schaefer, G. Schnapp, T. de Lange, Control of human telomere
length by TRF1 and TRF2, Mol. Cell. Biol. 20 (2000) 1659–1668.
[8] S. Smith, I. Giriat, A. Schmitt, T. de Lange, Tankyrase, a poly
(ADP-ribose) polymerase at human telomeres, Science 282 (1998)
1484–1487.
[9] S.H. Kim, P. Kaminker, J. Campisi, TIN2, a new regulator of
telomere length in human cells, Nat. Genet. 23 (1999) 405–412.
[10] X.Z. Zhou, K.P. Lu, The Pin2/TRF1-interacting protein PinX1 is
a potent telomerase inhibitor, Cell 107 (2001) 347–359.
[11] B. Li, S. Oestreich, T. de Lange, Identification of human Rap1:
implications for telomere evolution, Cell 101 (2000) 471–483.
[12] D. Loayza, T. de Lange, POT1 as a terminal transducer of TRF1
telomere length control, Nature 423 (2003) 1013–1018.
[13] L.M. Colgin, K. Baran, P. Baumann, T.R. Cech, R.R. Reddel,
Human POT1 facilitates telomere elongation by telomerase, Curr.
Biol. 13 (2003) 942–946.
[14] C. Liao, M.J. Zhao, H. Song, K. Uchida, K.K. Yokoyama, T.P.
Li, Identification of the gene for a novel liver-related putative
tumor suppressor at a high-frequency loss of heterozygosity
region of chromosome 8p23 in human hepatocellular carcinoma,
Hepatology 32 (2000) 721–727.
[15] C. Liao, M.J. Zhao, J. Zhao, D. Jia, H. Song, Z.P. Li, Over-
expression of LPTS-L in hepatocellular carcinoma cell line
SMMC-7721 induces crisis, World J. Gastroenterol. 8 (2002)
1050–1052.
[16] W.S. Park, J.H. Lee, J.Y. Park, S.W. Jeong, M.S. Shin, H.S. Kim,
S.K. Lee, S.N. Lee, S.H. Lee, C.G. Park, N.J. Yoo, J.Y. Lee,
Genetic analysis of the liver putative tumor suppressor (LPTS)
gene in hepatocellular carcinomas, Cancer Lett. 178 (2002) 199–
207.
[17] R. Bruni, B. Roizman, Herpes simplex virus 1 regulatory protein
ICP22 interacts with a new cell cycle-regulated factor and
accumulates in a cell cycle-dependent fashion in infected cells, J.
Virol. 72 (1998) 8525–8531.
[18] Y. Ren, R.K. Busch, L. Perlaky, H. Busch, The 58-kDa
microspherule protein (MSP58), a nucleolar protein, interacts
with nucleolar protein p120, Eur. J. Biochem. 253 (1998) 734–742.
[19] D.Y. Lin, H.M. Shih, Essential role of the 58-kDa microspherule
protein in the modulation of daxx-dependent transcriptional
repression as revealed by nucleolar sequestration, J. Biol. Chem.
277 (2002) 25446–25456.
[20] C.B. Harley, A.B. Futcher, C.W. Greider, Telomeres shorten
during aging of human fibroblasts, Nature 345 (1990) 458–
460.
[21] N.W. Kim, F. Wu, Advances in quantification and characteriza-
tion of telomerase activity by the telomeric repeat amplification
protocol (TRAP), Nucleic Acids Res. 25 (1997) 2595–2597.
[22] W.L. Fangman, B.J. Brewer, A question of time: replication
origins of eukaryotic chromosomes, Cell 71 (1992) 363–366.
[23] L.L. Sandell, V.A. Zakian, Loss of a yeast telomere: arrest,
recovery, and chromosome loss, Cell 75 (1993) 729–739.
[24] A.K. Taggart, S.C. Teng, V.A. Zakian, Est1p as a cell cycle-
regulated activator of telomere-bound telomerase, Science 297
(2002) 1023–1026.
[25] S.E. Holt, W.E. Wright, J.W. Shay, Regulation of telomerase
activity in immortal cell lines, Mol. Cell. Biol. 16 (1996) 2932–
2939.
[26] W.E. Wright, V.M. Tesmer, M.L. Liao, J.W. Shay, Normal
human telomeres are not late replicating, Exp. Cell Res. 251
(1999) 492–499.
[27] J.M. Wong, L. Kusdra, K. Collins, Subnuclear shuttling of human
telomerase induced by transformation and DNA damage, Nat.
Cell Biol. 4 (2002) 731–736.
[28] K. Masutomi, E.Y. Yu, S. Khurts, I. Ben-Porath, J.L. Currier,
G.B. Metz, M.W. Brooks, S. Kaneko, S. Murakami, J.A.
DeCaprio, R.A. Weinberg, S.A. Stewart, W.C. Hahn, Telomerase
maintains telomere structure in normal human cells, Cell 114
(2003) 241–253.
[29] X.Z. Zhou, K. Perrem, K.P. Lu, Role of Pin2/TRF1 in telomere
maintenance and cell cycle control, J. Cell Biochem. 89 (2003) 19–
37.
[30] A.G. Bader, M.L. Schneider, K. Bister, M. Hartl, TOJ3, a target
of the v-Jun transcription factor, encodes a protein with
transforming activity related to human microspherule protein 1
(MCRS1), Oncogene 20 (2001) 7524–7535.
[31] K. Hofmann, P. Bucher, The FHA domain: a putative nuclear
signalling domain found in protein kinases and transcription
factors, Trends Biochem. Sci. 20 (1995) 347–349.
[32] D. Durocher, J. Henckel, A.R. Fersht, S.P. Jackson, The FHA
domain is a modular phosphopeptide recognition motif, Mol. Cell
4 (1999) 387–394.
[33] J. Li, G.I. Lee, S.R. Van Doren, J.C. Walker, The FHA domain
mediates phosphoprotein interactions, J. Cell Sci. 113 (2000)
4143–4149.




















