
Regulation of Murine Telomere Length by Rtel
14
Cell, Vol. 117, 873–886, June 25, 2004, Copyright 2004 by Cell Press Regulation of Murine Telomere Length by Rtel : An Essential Gene Encoding a Helicase-like Protein Blackburn, 2001). In most eukaryotic species, telomeric DNA consists of short G-rich repeat sequences synthe- sized by telomerase (Greider and Blackburn, 1985; Hao Ding, 1,7 Mike Schertzer, 2,7 Xiaoli Wu, 1 Marina Gertsenstein, 1 Sara Selig, 2,4 Makoto Kammori, 2 Reza Pourvali, 2 Steven Poon, 2 Irma Vulto, 2 Elizabeth Chavez, 2 Patrick P.L. Tam, 3 Singer and Gottschling, 1994). In vertebrates, telomeric DNA consists of a double-strand region composed of Andras Nagy, 1,5 and Peter M. Lansdorp 2,6, * 1 Mount Sinai Hospital TTAGGG repeats (Moyzis et al., 1988). The very end of chromosomes consists of a 3 single-strand G-rich Samuel Lunenfeld Research Institute Toronto, Ontario, M5G 1X5 overhang (Makarov et al., 1997; McElligott and Wel- linger, 1997) that invades into the duplex telomeric re- 2 Terry Fox Laboratory British Columbia Cancer Agency peat array to form a T loop structure (Griffith et al., 1999). An important function of telomeres is to distinguish nor- Vancouver, British Columbia, V5Z 1L3 Canada mal chromosome ends from double-strand breaks, yet it is not clear exactly how the structure of telomere ends 3 Embryology Unit Children’s Medical Research Institute differs from sites of DNA damage (de Lange, 2002). Most likely activities that are specific for telomere mainte- University of Sydney Westmead, New South Wales, 2145 nance (such as telomerase) are recruited to telomeres via proteins that bind directly or indirectly to telomere Australia 4 Rambam Medical Center and repeats (Blackburn, 2001; Vega et al., 2003). In the ab- sence of such proteins, DNA repair-specific activities Technion Faculty of Medicine Haifa 31096 are recruited to chromosome ends that can trigger chro- mosome fusions (Mieczkowski et al., 2003) and end re- Israel 5 Department of Molecular and Medical Genetics section (Hackett and Greider, 2003) or induce other forms of potentially lethal DNA damage responses (Ta- University of Toronto Toronto, Ontario, M5S 1A8 kai et al., 2003; Zhu et al., 2003). Telomeric DNA is lost in cells because of incomplete 6 Department of Medicine University of British Columbia replication of lagging strand DNA (Greider, 1996; McEachern et al., 2000), C strand degradation (Makarov Vancouver, British Columbia, V5Z 4E3 Canada et al., 1997), oxidative stress (von Zglinicki, 2002), and possibly other mechanisms. Telomerase is dispensable in cells with sufficiently long telomeres (Blasco et al., 1997), but cells with short telomeres that lack telomerase Summary activity typically lose the ability to proliferate after a variable number of cell divisions (Blackburn, 2001). Little is known about the genes that regulate telomere length diversity between mammalian species. A candi- While the precise role of telomerase and telomeres in the proliferation of various cell types is subject to extensive date gene locus was previously mapped to a region on distal mouse Chr 2q. Within this region, we identified a investigations, it is clear that a minimum number of telo- mere repeats is required at each chromosome end for gene similar to the dog-1 DNA helicase-like gene in C. elegans. We cloned this Regulator of telomere proper telomere function and to distinguish telomeres from double-strand breaks. length (Rtel) gene and inactivated its expression in mice. Rtel / mice died between days 10 and 11.5 of The average length of telomere repeats is set by an equilibrium between the mechanisms that lengthen and gestation with defects in the nervous system, heart, vasculature, and extraembryonic tissues. Rtel / em- shorten telomere tracts. It is not known which factors are most important in establishing the average telomere bryonic stem cells showed telomere loss and displayed many chromosome breaks and fusions upon differentia- length in cells or the mechanisms that generate telomere tion in vitro. Crosses of Rtel / mice with Mus spretus length diversity between species. In a previous study, showed that Rtel from the Mus musculus parent is the genetic regulation of telomere length was investi- required for telomere elongation of M. spretus chro- gated by using two interfertile species of mice, which mosomes in F1 cells. We conclude that Rtel is an es- differ in their telomere length (Zhu et al., 1998). Mus sential gene that regulates telomere length and pre- musculus (telomere length 25 kb) and Mus spretus vents genetic instability. (telomere length 5–15 kb) were used to generate F1 crosses and reciprocal backcrosses, which were then Introduction analyzed for regulation of telomere length. This analysis indicated the presence of a dominant and transacting Telomeres are DNA-protein complexes at the ends of mechanism capable of extensive elongation of telo- linear eukaryotic chromosomes that play a critical role meres in somatic cells after fusion of parental germline in maintaining chromosome stability (Zakian, 1995; cells with discrepant telomere lengths. A genome-wide screen of F1 backcrosses using M. spretus as the recur- rent parent identified a 5 cM region on distal chromo- *Correspondence: [email protected] 7 These authors contributed equally to this work. some 2 that predominantly controls the observed spe-
-
Upload
independent -
Category
Documents
-
view
5 -
download
0
Transcript of Regulation of Murine Telomere Length by Rtel

Cell, Vol. 117, 873–886, June 25, 2004, Copyright 2004 by Cell Press
Regulation of Murine Telomere Length by Rtel:An Essential Gene Encoding a Helicase-like Protein
Blackburn, 2001). In most eukaryotic species, telomericDNA consists of short G-rich repeat sequences synthe-sized by telomerase (Greider and Blackburn, 1985;
Hao Ding,1,7 Mike Schertzer,2,7 Xiaoli Wu,1
Marina Gertsenstein,1 Sara Selig,2,4
Makoto Kammori,2 Reza Pourvali,2 Steven Poon,2
Irma Vulto,2 Elizabeth Chavez,2 Patrick P.L. Tam,3 Singer and Gottschling, 1994). In vertebrates, telomericDNA consists of a double-strand region composed ofAndras Nagy,1,5 and Peter M. Lansdorp2,6,*
1Mount Sinai Hospital TTAGGG repeats (Moyzis et al., 1988). The very endof chromosomes consists of a 3� single-strand G-richSamuel Lunenfeld Research Institute
Toronto, Ontario, M5G 1X5 overhang (Makarov et al., 1997; McElligott and Wel-linger, 1997) that invades into the duplex telomeric re-2 Terry Fox Laboratory
British Columbia Cancer Agency peat array to form a T loop structure (Griffith et al., 1999).An important function of telomeres is to distinguish nor-Vancouver, British Columbia, V5Z 1L3
Canada mal chromosome ends from double-strand breaks, yetit is not clear exactly how the structure of telomere ends3 Embryology Unit
Children’s Medical Research Institute differs from sites of DNA damage (de Lange, 2002). Mostlikely activities that are specific for telomere mainte-University of Sydney
Westmead, New South Wales, 2145 nance (such as telomerase) are recruited to telomeresvia proteins that bind directly or indirectly to telomereAustralia
4 Rambam Medical Center and repeats (Blackburn, 2001; Vega et al., 2003). In the ab-sence of such proteins, DNA repair-specific activitiesTechnion Faculty of Medicine
Haifa 31096 are recruited to chromosome ends that can trigger chro-mosome fusions (Mieczkowski et al., 2003) and end re-Israel
5 Department of Molecular and Medical Genetics section (Hackett and Greider, 2003) or induce otherforms of potentially lethal DNA damage responses (Ta-University of Toronto
Toronto, Ontario, M5S 1A8 kai et al., 2003; Zhu et al., 2003).Telomeric DNA is lost in cells because of incomplete6 Department of Medicine
University of British Columbia replication of lagging strand DNA (Greider, 1996;McEachern et al., 2000), C strand degradation (MakarovVancouver, British Columbia, V5Z 4E3
Canada et al., 1997), oxidative stress (von Zglinicki, 2002), andpossibly other mechanisms. Telomerase is dispensablein cells with sufficiently long telomeres (Blasco et al.,1997), but cells with short telomeres that lack telomeraseSummaryactivity typically lose the ability to proliferate after avariable number of cell divisions (Blackburn, 2001).Little is known about the genes that regulate telomere
length diversity between mammalian species. A candi- While the precise role of telomerase and telomeres in theproliferation of various cell types is subject to extensivedate gene locus was previously mapped to a region on
distal mouse Chr 2q. Within this region, we identified a investigations, it is clear that a minimum number of telo-mere repeats is required at each chromosome end forgene similar to the dog-1 DNA helicase-like gene in
C. elegans. We cloned this Regulator of telomere proper telomere function and to distinguish telomeresfrom double-strand breaks.length (Rtel) gene and inactivated its expression in
mice. Rtel�/� mice died between days 10 and 11.5 of The average length of telomere repeats is set by anequilibrium between the mechanisms that lengthen andgestation with defects in the nervous system, heart,
vasculature, and extraembryonic tissues. Rtel�/� em- shorten telomere tracts. It is not known which factorsare most important in establishing the average telomerebryonic stem cells showed telomere loss and displayed
many chromosome breaks and fusions upon differentia- length in cells or the mechanisms that generate telomeretion in vitro. Crosses of Rtel�/� mice with Mus spretus length diversity between species. In a previous study,showed that Rtel from the Mus musculus parent is the genetic regulation of telomere length was investi-required for telomere elongation of M. spretus chro- gated by using two interfertile species of mice, whichmosomes in F1 cells. We conclude that Rtel is an es- differ in their telomere length (Zhu et al., 1998). Mussential gene that regulates telomere length and pre- musculus (telomere length �25 kb) and Mus spretusvents genetic instability. (telomere length 5–15 kb) were used to generate F1
crosses and reciprocal backcrosses, which were thenIntroduction analyzed for regulation of telomere length. This analysis
indicated the presence of a dominant and transactingTelomeres are DNA-protein complexes at the ends of mechanism capable of extensive elongation of telo-linear eukaryotic chromosomes that play a critical role meres in somatic cells after fusion of parental germlinein maintaining chromosome stability (Zakian, 1995; cells with discrepant telomere lengths. A genome-wide
screen of F1 backcrosses using M. spretus as the recur-rent parent identified a 5 cM region on distal chromo-*Correspondence: [email protected]
7 These authors contributed equally to this work. some 2 that predominantly controls the observed spe-

Cell874
cies-specific telomere length regulation (Zhu et al., ure 1D). Of note, both RTEL and NHL contain a con-served eight-amino acid PIP-box motif near the carboxy1998).terminus that is commonly found in proteins capable ofAmong the candidate genes in this region, we identi-interacting with proliferating cell nuclear antigen (PCNA)fied the mouse homolog of the human novel helicase-(Warbrick, 2000). The protein encoded by the most com-like (NHL) gene (Bai et al., 2000). In order to study amon splice variant in M. spretus (AY481613) is lackingpossible role of this gene in telomere length regulation,six amino acids (AHFSKP) that are also absent in thewe decided to first study one of the two most similarhuman NHL protein. As a result, this splice variant con-genes in C. elegans, the dog-1 gene, for which a nulltains a predicted CK1 phosphorylation site (Kreegipuumutant was readily available. While dog-1 mutants dis-et al., 1999) that is not present in the full-length protein.played no clear telomere phenotype, they showed a
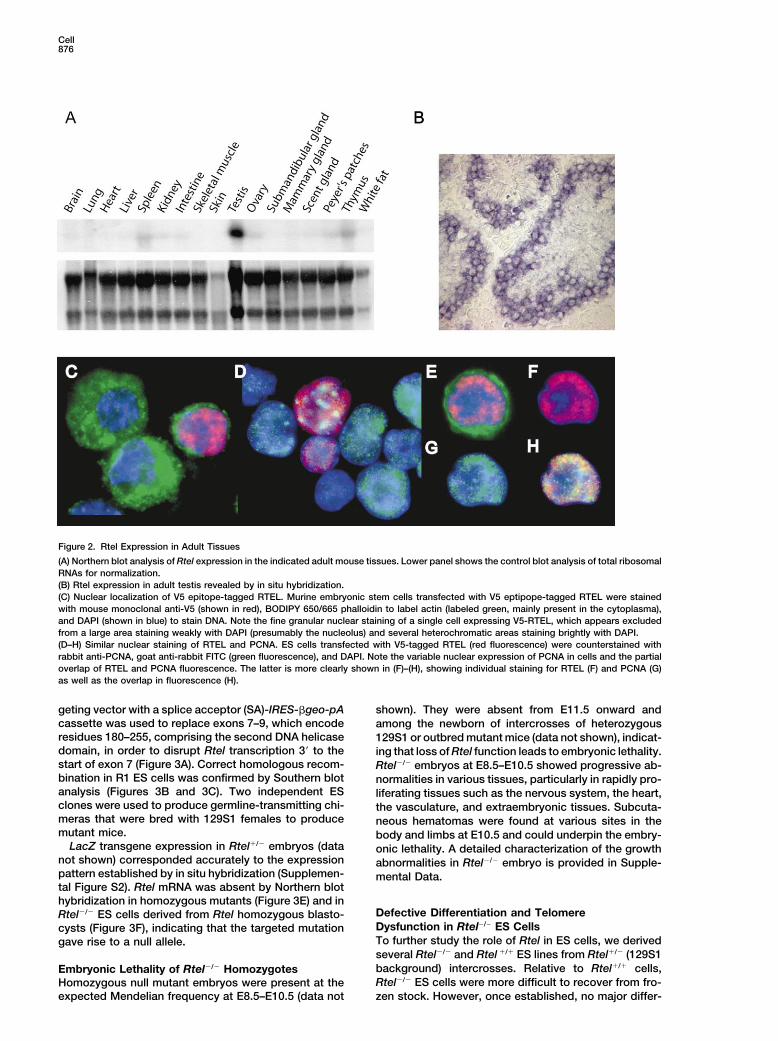
highly unusual mutator phenotype characterized by de-Expression of Rtel in Tissues and Cellsletions of G-rich DNA throughout genomic DNA (CheungRtel expression was analyzed in mouse embryos andet al., 2002). Deletions in dog-1 mutants invariably origi-adult tissues to reveal its potential function. Details ofnated around the 3� end of the G tracts and typicallythis analysis are shown in Figure 2 and Supplementalwere between 100 and 200 nucleotides in length. SuchFigure S2 (on the Cell web site). Rtel was widely ex-deletions were observed in �50% of genomic polygua-pressed in E8.5 and E9.5 embryos and showed a morenine tracts above a threshold length of more than 18restricted expression pattern at E13.5–E15.5. In general,consecutive Gs with a frequency that increased with theRtel expression in embryos appeared to coincide withlength of the tract. These results are consistent with aareas of actively proliferating cells (see Supplementalrole for the DOG-1 protein in the resolution of higherFigure S2). In adult mice, Rtel expression was detectableorder structures of G-rich DNA, such as guanine quadru-by Northern hybridization in spleen, thymus, Peyer’splex DNA (Sen and Gilbert, 1988; Williamson et al., 1989),patches, kidney, and intestine but not in brain, heart,that could arise stochastically during lagging strand rep-lung, skeletal muscles, skin, and white fat tissue (Figurelication of G-rich template sequences (Cheung et al.,2A). In the adult gonad, Rtel was expressed highly in2002). In view of these findings in C. elegans and thethe testis, mainly in the spermatogonia and meioticwell-known ability of telomeric G strands to assume G4spermatocytes (Figure 2B). Rtel expression was studiedDNA structures in vitro (Sen and Gilbert, 1988; William-in relation to cell proliferation in adult tissues in threeson et al., 1989; Parkinson et al., 2002), we decided toexperimental models. Significant induction of Rtel ex-study a possible role of the murine Rtel gene in telomerepression was observed in actively regenerating musclelength regulation by disrupting its expression in vitroand liver as well as in mitogen-stimulated spleen cellsand in vivo.(see Supplemental Figure S2).
To examine the subcellular localization of RTEL,Resultsmouse embryonic stem cells were transiently trans-fected with a plasmid encoding V5 epitope-taggedGenomic Organization of the Rtel Locus andRTEL. Cells expressing the transgene (�1% of total)Alternative Splicing of Rtel Transcriptswere easily recognized by bright nuclear staining, which,The murine Rtel gene has 34 exons, encompasses 36.6in all cells, had a fine granular appearance that did notkb of genomic DNA, and is located within 1 Mb of theinclude nucleoli or areas of heterochromatin (Figures 2Ctelomere on chromosome 2q (Figures 1A and 1B). North-and 2D). In view of the expression of RTEL in proliferating
ern blot analysis of transcripts yielded a species of �4cells and tissues and the presence of a predicted PCNA
kb in both M. musculus and M. spretus. No major differ-binding site in RTEL (Figure 1D), the expression of RTEL
ences between the two species were observed in �1.5 relative to PCNA was also examined (Figures 2D–2H).kb of putative Rtel promoter sequences upstream of PCNA was expressed at variable levels in most ES cellsthe first Rtel exon and Rtel transcript levels in different (Figure 2D). While, in cells that expressed both PCNAtissues varied in parallel (data not shown). Analysis of and the transgene, the nuclear staining pattern ap-cDNA clones from M. musculus, M. spretus, and F1 cells peared very similar, only a minority of the small nuclearshowed several splice variants involving the last four foci appeared to directly overlap (Figure 2H). Stainingexons of the Rtel gene (Figure 1C and dotted box in of replication foci using anti-BrdU yielded similar results,Figure 1A). Whereas full-length M. musculus Rtel tran- and epitope-tagged RTEL did not significantly colocal-scripts were readily obtained, most (�90%) M. spretus ize with either telomeric DNA or antibodies specific forRtel transcripts were missing 18 or more bp at the telomere binding proteins (data not shown).C terminus (Figure 1C). Taken together, these data indicate that Rtel expres-
sion is widespread in dividing cells during embryonicRtel Encodes a Highly Conserved Protein with development, in adult cells that are induced to prolifer-Characteristic Helicase Motifs ate, and, constitutively, in some adult tissues, most no-Full-length Rtel transcripts are predicted to encode a tably the testis. RTEL expression in ES cells appears1209 amino acids (aa) protein in M. musculus and M. restricted to the nucleus in numerous foci that appearspretus (see Supplemental Figure S1 at http://www.cell. similar but overlap only partially with replication focicom/cgi/content/full/117/7/873/DC1). Both proteins are containing PCNA.highly homologous (�98% aa identity) to each other and74% identical to the human NHL protein (Bai et al., 2000). Generation of a Null Mutation of RtelThe sequence identity is even higher for the first 750 aa To further study Rtel function, the Rtel locus was mu-
tated in ES cells by homologous recombination. A tar-containing the seven domains of known helicases (Fig-

Regulation of Telomere Length by Rtel875
Figure 1. Gene Structure, Splice Junction Sites, and Splice Variants of the Rtel Gene and Conserved Functional Domains in the RTEL Protein
(A) Location of the Rtel within 1 Mb of the telomere at chromosome 2q. The Rtel gene encompasses 36.6 kb of genomic DNA and is flankedby the Stathmin3 and Arfrp1 genes. Alternative splicing involves the boxed area at the 3� end of the gene.(B) Splice junction sites and exon size of the M. musculus Rtel gene. Exon sequences are in capitalized letters. Intron sequences are in lowercase.(C) Rtel splice variants obtained from cDNAs derived from M. musculus and M. spretus testes and (M. musculus � M. spretus) F1 spleencells. Regions homologous to the coding sequence of cDNA are indicated by black boxes, and the 3� UTR is indicated as a gray box. Skippingof a full or partial exon is indicated by brackets below the exons. The predicted protein products are shown as black bars under each splicevariant. The number of times a given splice variant sequence was observed is shown.(D) Conserved functional domains in the predicted RTEL amino acid sequence include the seven characteristic helicase motifs indicated byroman numbers. RTEL is classified in the NCBI conserved domain database as a member of the K0G1132, helicase of the DEAD superfamily(replication, recombination, and repair), and as a member of the DinG, Rad3-related DNA helicases containing both a DEXDc2 and a HELICc2domain (Marchler-Bauer et al., 2003). A PIP box (encoded by exon 34) is present near the C terminus of the protein.
Cell876
Figure 2. Rtel Expression in Adult Tissues
(A) Northern blot analysis of Rtel expression in the indicated adult mouse tissues. Lower panel shows the control blot analysis of total ribosomalRNAs for normalization.(B) Rtel expression in adult testis revealed by in situ hybridization.(C) Nuclear localization of V5 epitope-tagged RTEL. Murine embryonic stem cells transfected with V5 eptipope-tagged RTEL were stainedwith mouse monoclonal anti-V5 (shown in red), BODIPY 650/665 phalloidin to label actin (labeled green, mainly present in the cytoplasma),and DAPI (shown in blue) to stain DNA. Note the fine granular nuclear staining of a single cell expressing V5-RTEL, which appears excludedfrom a large area staining weakly with DAPI (presumably the nucleolus) and several heterochromatic areas staining brightly with DAPI.(D–H) Similar nuclear staining of RTEL and PCNA. ES cells transfected with V5-tagged RTEL (red fluorescence) were counterstained withrabbit anti-PCNA, goat anti-rabbit FITC (green fluorescence), and DAPI. Note the variable nuclear expression of PCNA in cells and the partialoverlap of RTEL and PCNA fluorescence. The latter is more clearly shown in (F)–(H), showing individual staining for RTEL (F) and PCNA (G)as well as the overlap in fluorescence (H).
geting vector with a splice acceptor (SA)-IRES-�geo-pA shown). They were absent from E11.5 onward andcassette was used to replace exons 7–9, which encode among the newborn of intercrosses of heterozygousresidues 180–255, comprising the second DNA helicase 129S1 or outbred mutant mice (data not shown), indicat-domain, in order to disrupt Rtel transcription 3� to the ing that loss of Rtel function leads to embryonic lethality.start of exon 7 (Figure 3A). Correct homologous recom- Rtel�/� embryos at E8.5–E10.5 showed progressive ab-bination in R1 ES cells was confirmed by Southern blot normalities in various tissues, particularly in rapidly pro-analysis (Figures 3B and 3C). Two independent ES liferating tissues such as the nervous system, the heart,clones were used to produce germline-transmitting chi- the vasculature, and extraembryonic tissues. Subcuta-meras that were bred with 129S1 females to produce neous hematomas were found at various sites in themutant mice. body and limbs at E10.5 and could underpin the embry-
LacZ transgene expression in Rtel�/� embryos (data onic lethality. A detailed characterization of the growthnot shown) corresponded accurately to the expression abnormalities in Rtel�/� embryo is provided in Supple-pattern established by in situ hybridization (Supplemen- mental Data.tal Figure S2). Rtel mRNA was absent by Northern blothybridization in homozygous mutants (Figure 3E) and in
Defective Differentiation and TelomereRtel�/� ES cells derived from Rtel homozygous blasto-Dysfunction in Rtel�/� ES Cellscysts (Figure 3F), indicating that the targeted mutationTo further study the role of Rtel in ES cells, we derivedgave rise to a null allele.several Rtel�/� and Rtel �/� ES lines from Rtel�/� (129S1background) intercrosses. Relative to Rtel�/� cells,Embryonic Lethality of Rtel�/� HomozygotesRtel�/� ES cells were more difficult to recover from fro-Homozygous null mutant embryos were present at the
expected Mendelian frequency at E8.5–E10.5 (data not zen stock. However, once established, no major differ-

Regulation of Telomere Length by Rtel877
Figure 3. Targeted Disruption of Rtel by Homologous Recombination
(A) Schematic representation of exons 1–13 of the mouse Rtel locus, the targeting vector, and the mutant allele. Black boxes represent exonsaccording to sequence comparison of Rtel cDNA and corresponding genomic sequences. Exons 7–9 were deleted and replaced with a splicingacceptor (SA)-linked IRES�geo cassette to generate a null allele. The locations of the hybridization probes (5� probe and 3� probe) for Southernblot analysis are shown.(B) Southern blot analysis of ES clones with insertion of targeting vector. The genomic DNA was digested with BamHI and probed with 5�
probe. The wild-type allele produced a 15.0 kb fragment, and the targeted allele produced a 9.0 kb fragment. The genomic DNA of wild-typeR1 ES cells was used as a control.(C) Southern blot analysis of targeted ES clones using the 3� probe. The genomic DNA was digested with NsiI and hybridized with the 3�
probe, giving a wild-type allele of 10.0 kb and a mutant allele of 16.0 kb.(D) Southern blot analysis of ES cells derived from blastocysts of Rtel�/� (129Sv background) intercrosses. Genomic DNA was digested withBamHI and probed with the 5� probe. Two homozygous ES clones produced only a mutant allele of 9.0 kb.(E) Northern blot analysis of Rtel mRNA in the E9.5 embryos of the indicated genotypes. A 750 bp cDNA fragment covering the 3� end of Rtelcoding sequence was used as a probe. GAPDH cDNA hybridization was used as a normalization control.(F) Northern blot analysis of total RNA isolated from the ES cells shown in (D). The 28S and 18S ribosomal RNAs are shown as normaliza-tion controls.
ences in the growth rate of Rtel�/� and Rtel �/� ES cells S phase decreased upon differentiation (Figure 4B), con-sistent with previous observations (Aladjem et al., 1998).were recorded. In contrast, when cells were cultured in
medium without leukemia inhibitory factor (LIF) to in- In contrast, a large number of cell fragments (reflectingcell death) and an almost complete loss of BrdU-labeledduce their differentiation, striking differences in the fre-
quency and size of day 5 embryoid bodies between cells was observed in Rtel�/� cells cultured without LIFfor 3 days (Figure 4B, bottom panels).Rtel�/� and Rtel �/� ES cells were observed (Figure 4A).
Whereas Rtel�/� and Rtel �/� ES cells yielded 367 � 101 To study a possible role of RTEL in telomere homeo-stasis, the average telomere length as well as the telo-and 274 � 83 embryoid bodies per 1500 cells plated,
this number was 66 � 19 for Rtel�/� ES cells. To study mere length of individual chromosome ends in Rtel�/�,Rtel �/�, and Rtel �/� ES cells was studied using flowthe nature of the defect in differentiating Rtel�/� ES cells,
the cell cycle distribution of ES cells grown without LIF FISH (Figure 4C), terminal restriction fragment length bySouthern blot analysis (Figure 4D), and Q-FISH (Figureswas analyzed using 5-bromo-2�-deoxyuridine (BrdU) in-
corporation. In Rtel�/� ES cells, the fraction of cells in 4E–4G), respectively. Flow FISH analysis showed that

Cell878
Figure 4. Defective Differentiation and Telomere Phenotype of Rtel�/� ES Cells
(A) Differentiation of Rtel�/� and Rtel�/� ES cells cultured for 20 population doublings in liquid culture prior to differentiation in the absenceof LIF in methylcellulose. Note that colonies observed at day 5 are notably smaller in Rtel�/� cells.(B) Cell cycle distribution of Rtel�/� and Rtel�/� ES cells before and after induction of differentiation by removal of LIF from the culture medium.Cells were pulsed with BrdU for 30 min prior to harvest to distinguish cells in the G1, S, and G2M phase of the cell cycle. Prior to differentiationthe majority of intact Rtel�/� and Rtel�/� cells (boxed areas, left quadrants) are in S phase (cells with high BrdU content that connect G1 cellswith low BrdU and low PI fluorescence to G2M cells with low BrdU and high DNA content). After 3 days of culture without LIF, very few (4.1%)intact cells are recovered from cultures of Rtel�/� ES cells (middle quadrants), and such cells contain very few cells in S phase (right quadrant,events within boxed area of middle quadrant only). In contrast, many cells in S phase (34.1%) were recovered from Rtel�/� control cultures.The percentage of events in the boxed area relative to the total number of acquired events is shown.(C) Telomere length (mean and standard deviation form duplicate measurements) in undifferentiated Rtel�/� (B4 and E3) and Rtel�/� (A4 andE1) ES cells measured by flow FISH. Telomere length in Rtel�/� cells is maintained at �37 kb or around 68% of the length in Rtel�/� cells(�54kb) over �25 population doublings.(D) Telomere length in Rtel�/�, Rtel �/�, and Rtel �/� ES cells measured by terminal restriction fragment (TRF) Southern blot analysis. Note thatfragments with an estimated length of �30 kb (boxed area) are observed in Rtel�/� cells but not Rtel�/� and Rtel �/� cells.(E–G) Telomere length analysis in Rtel�/� and Rtel�/� ES cells using Q-FISH. Ends with low or undetectable telomere signals (arrows) arepresent in both Rtel�/� (E) and Rtel�/� ES cells (F) but are much more abundant in Rtel�/� cells. (G) Q-FISH telomere length analysis in Rtel�/�
(E3) and Rtel�/� (A4) undifferentiated ES cells at PD20. Quantitative analysis of �30 metaphase cells using images similar to those shown in(E) and (F). The mean fluorescence intensity (in arbitrary units) is indicated in the fluorescence histogram. Note the skewed distribution oftelomeres and the large number of events with no or very little telomere signal in Rtel�/� relative to Rtel�/� cells .
Regulation of Telomere Length by Rtel879
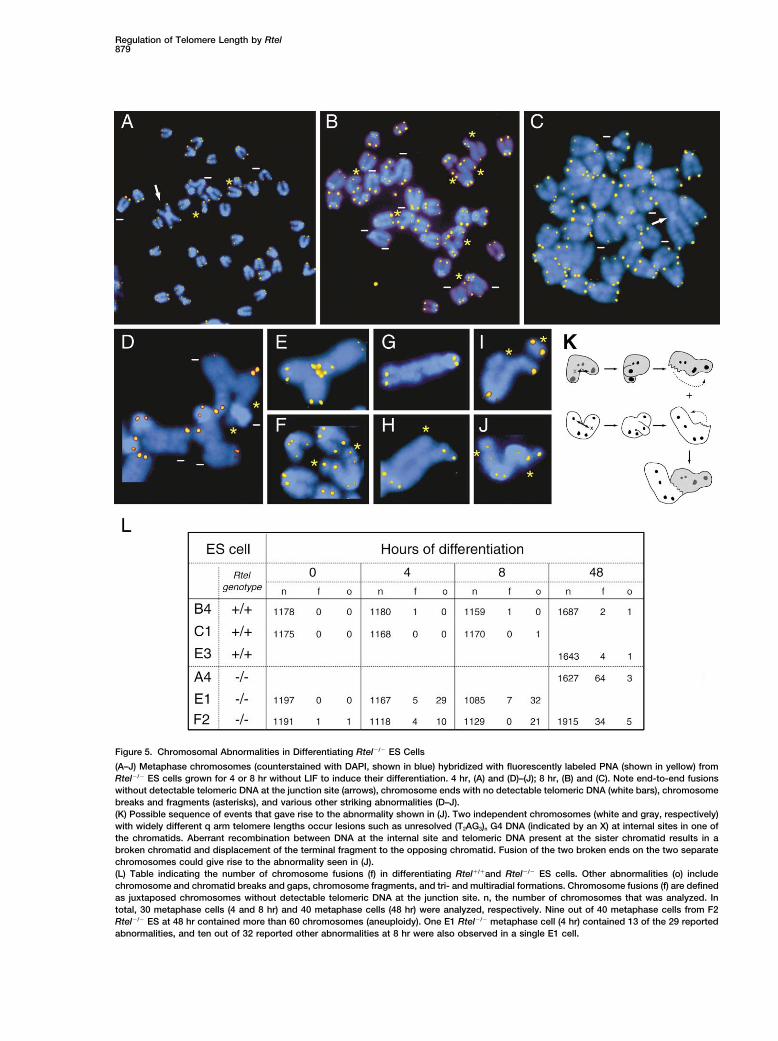
Figure 5. Chromosomal Abnormalities in Differentiating Rtel�/� ES Cells
(A–J) Metaphase chromosomes (counterstained with DAPI, shown in blue) hybridized with fluorescently labeled PNA (shown in yellow) fromRtel�/� ES cells grown for 4 or 8 hr without LIF to induce their differentiation. 4 hr, (A) and (D)–(J); 8 hr, (B) and (C). Note end-to-end fusionswithout detectable telomeric DNA at the junction site (arrows), chromosome ends with no detectable telomeric DNA (white bars), chromosomebreaks and fragments (asterisks), and various other striking abnormalities (D–J).(K) Possible sequence of events that gave rise to the abnormality shown in (J). Two independent chromosomes (white and gray, respectively)with widely different q arm telomere lengths occur lesions such as unresolved (T2AG3)n G4 DNA (indicated by an X) at internal sites in one ofthe chromatids. Aberrant recombination between DNA at the internal site and telomeric DNA present at the sister chromatid results in abroken chromatid and displacement of the terminal fragment to the opposing chromatid. Fusion of the two broken ends on the two separatechromosomes could give rise to the abnormality seen in (J).(L) Table indicating the number of chromosome fusions (f) in differentiating Rtel�/�and Rtel�/� ES cells. Other abnormalities (o) includechromosome and chromatid breaks and gaps, chromosome fragments, and tri- and multiradial formations. Chromosome fusions (f) are definedas juxtaposed chromosomes without detectable telomeric DNA at the junction site. n, the number of chromosomes that was analyzed. Intotal, 30 metaphase cells (4 and 8 hr) and 40 metaphase cells (48 hr) were analyzed, respectively. Nine out of 40 metaphase cells from F2Rtel�/� ES at 48 hr contained more than 60 chromosomes (aneuploidy). One E1 Rtel�/� metaphase cell (4 hr) contained 13 of the 29 reportedabnormalities, and ten out of 32 reported other abnormalities at 8 hr were also observed in a single E1 cell.

Cell880
the average telomere length in Rtel�/� and Rtel �/� ES cells from F1 littermates obtained from crosses betweena Rtel�/� 129Sl inbred female and a M. spretus malecells was kept relatively constant over 30 population
doublings (PD) at a similar length of 53.7 � 2.8 kb (Figure using flow FISH. The results (shown in Figure 6A) con-firmed that the average telomere length was long (43.34C) and 51.6 � 0.7 kb (data not shown). In contrast, the
telomere length in two separate Rtel�/� ES cells was kb) and short (9.3 kb) in the 129Sl M. musculus and M.spretus parental cells, respectively. In the offspring, themaintained at 36.7 � 1.9 kb or �68% of wild-type length
over 30 PD (Figure 4C). Southern blot analysis and average telomere length in cells from animals derivedfrom two litters that inherited the wild-type M. musculusQ-FISH analysis revealed that this difference in average
telomere length resulted from the presence of variably Rtel allele (n 5) was significantly longer than in lit-termates (n 7) that inherited the Rtel null allele fromshortened telomeres in Rtel�/� cells (boxed area in Fig-
ure 4D and skewed telomere length distribution in Figure the M. musculus parent (Figure 6B). These results wereconfirmed by performing Q-FISH telomere length analy-4G) next to telomeres of apparent wild-type length.
Q-FISH analysis showed that, whereas occasional ends sis of cells from F1 animals (Figures 6C and 6D). Thetelomere length in M. spretus-derived chromosomes inwith a very faint or undetectable FISH signals were ob-
served in Rtel�/� ES cells (Figure 4E, arrows), such cells from F1 littermates with the Rtel null allele fromthe M. musculus parent (n 5) was similar to that ob-events were relatively abundant in Rtel�/� ES cells (Fig-
ure 4F, arrows, and Figure 4G). served in parental M. spretus cells and was markedlyshorter than in cells from littermates that inherited theTaken together, the data indicate that lack of RTEL
results in variable loss of telomere repeats in undifferen- wild-type Rtel allele from the M. musculus parent (n 5, Figure 6D). This was true for three different cell typestiated ES cells and maintenance of overall telomere
length at around 68% of that observed in wild-type from all animals of three different litters. These resultsdemonstrate that elongation of the shorter set of telo-ES cells.meres in chromosomes derived from the M. spretusparent in F1 cells requires Rtel expression from theCytogenetic Abnormalities in DifferentiatingM. musculus parent.Rtel�/� ES Cells
To study chromosome abnormalities observed in differ-entiating Rtel�/� ES cells in relation to telomeric DNA, DiscussionQ-FISH telomere analysis was performed on chromo-some spreads obtained from both Rtel�/� and Rtel �/� In this study, we describe the organization, expression
pattern, alternative splicing, and putative function ofES cells at various time intervals following inductionof differentiation. Strikingly, a variety of chromosomal Rtel, a murine gene encoding a DNA helicase-like pro-
tein. Rtel is an essential gene in that lack of Rtel resultsabnormalities was already observed in a minority of cells4 hr following removal of LIF (Figure 5). Apart from end- in genomic instability and embryonic lethality. Rtel�/�
ES cells were found to grow relatively normal, be it withto-end fusions (arrows) and ends without detectabletelomere signal (bars), the chromosomal abnormalities shortened telomeres, but showed genetic instability and
defective growth upon differentiation in vitro. Crossesthat were observed included clusters of chromosomesjoined at sites containing telomeric DNA (Figure 5E), between Rtel�/� mice and M. spretus revealed that Rtel
from the M. musculus parent is required for elongationvarious types of broken chromosomes and chromo-some fragments (asterisks in Figures 5A, 5B, 5D, 5F, of telomeres in M. spretus-derived chromosomes. We
conclude that Rtel is a dominant regulator of murineand 5H–5J), and chromatid gaps (Figure 5G). At latertime points, the most abundant cytogenetic abnormali- telomere length.ties were end-to-end fusions characterized by a lack ofdetectable telomere repeats at the junction site (Fig- Rtel Mode of Actionure 5L). The loss of telomere repeats and genetic instability in
The chromosomal abnormalities in differentiating Rtel�/� cells could result directly or indirectly from theRtel�/� ES cells indicate that RTEL is required to main- absence of RTEL. Further studies on the biochemicaltain the integrity of genomic as well as telomeric DNA. properties and molecules interacting with RTEL areIn order to explore possible similarities in genetic insta- needed to delineate its precise function. However,bility between Rtel�/� cells and dog-1 mutants (Cheung based on the observations described in this paper andet al., 2002), we studied the stability of several G-rich the sequence similarity between RTEL and the DOG-1sequences in genomic DNA from Rtel�/� and Rtel�/� ES protein in C. elegans (Cheung et al., 2002), we speculatecells (see Supplemental Data for details). No deletions that the functions of RTEL and DOG-1 are comparable.or insertions were observed in seven genomic poly- According to this model, RTEL is required to resolveguanine tracts (G � 18), 14 genomic (TTAGGG)n repeats higher order structures of G-rich DNA such as G4 DNA(n � 6) and 11 genomic (CCCTG)n repeats (n � 6). that could arise during replication, transcription, DNA
repair, and recombination of suitable G-rich DNA se-quences (Figure 7). In the absence of RTEL, higher orderRtel Is Required to Elongate M. spretus Telomeres
in Crosses with M. musculus structures of G-rich DNA at telomeres or elsewhere inthe genome could trigger repair by homologous recom-In a previous study, a trait involved in murine telomere
length regulation was mapped to the Tlq1 locus on distal bination. In general, repeat sequences pose a problemfor repair by homologous recombination (Petes and Hill,chromosome 2 (Zhu et al., 1998). In order to test the
hypothesis that Rtel is the gene responsible for this trait, 1988). For telomere repeats, the chances of a successfuloutcome could be further compromised by involvementwe first measured the average telomere length in spleen

Regulation of Telomere Length by Rtel881
Figure 6. M. musculus Rtel Is Required for Elongation of M. spretus Telomeres in F1 Cells
(A and B) Flow FISH analysis. (A) Dot plot histogram of telomere versus LDS-751 fluorescence of spleen cells obtained from parents (toppanels) and two representative F1 offspring (bottom panels) of a cross between a M. spretus male and a Rtel�/� M. musculus (129) female.The dotted line represents the analysis gate for the fixed bovine thymocytes that were added to every sample as an internal reference fortelomere length calculations (Baerlocher and Lansdorp, 2003).(B) Telomere length in spleen cells from littermates derived from crosses between a M. spretus male and a Rtel�/� M. musculus (129) female.Results of duplicate telomere length measurements by flow FISH of individual animals are shown. The difference in telomere length betweenF1 cells with (n 5) and without the M. musculus Rtel gene (n 7) is highly significant (p � 0.0001) using a hierarchical model (allowing forthe nesting within litters).(C and D) Analysis of telomere length at individual chromosome ends using Q-FISH. (C) Example of a multicolor FISH images used for Q-FISHanalysis. Metaphase chromosomes in an embryonic fibroblast from the F1 offspring of a cross between a M. spretus male and a Rtel�/� M.musculus (129) female. Cell were hybridized with a mixture of PNA probes specific for telomere repeats (probe labeled with Cy3, shown inyellow/red) and minor satellite repeats (labeled with FITC, shown in green). Note that the chromosomes derived from the M. spretus parentcan be easily recognized by the much brighter staining, with the probe for repetitive sequences present at mouse centromeres (except theY chromosome, arrow). The boxed area in the top panel is magnified in the lower panels which show individual (pseudo) colors and an exampleof the results of Q-FISH telomere length analysis generated using the TFL-Telo software program (Zijlmans et al., 1997).(D) Quantitative analysis of telomere length in chromosomes from different cell types derived from the offspring of crosses between Rtel�/�
M. musculus (129) and M. spretus parents. The mean fluorescence intensity (in arbitrary units) for each cell type is shown in the fluorescencehistogram. The telomere fluorescence in chromosomes derived only from the M. spretus parent (identified by bright green fluorescence asshown in Figure 6C) are shown in the panels on the right. Note that M. spretus telomeres are longer in F1 cells from animals that inheritedthe wild-type M. musculus Rtel allele than in F1 cells from animals that inherited the M. musculus Rtel null allele.

Cell882
Figure 7. Model of RTEL Action
In both wild-type and Rtel�/� cells, higher order structures of G-rich DNA such as guanine quadruplex (G4) DNA are postulated to arisesporadically and stochastically during lagging strand replication, repair, or recombination of suitable G-rich DNA sequences. (T2AG3)n DNA ispredicted to be particularly at risk for G4 DNA formation with the chance being proportional to the length of the repeat array. In wild-typecells, RTEL could be recruited to the G4 lesion either directly or via a DNA damage response and participate in its resolution. Effectiveinteractions between G4 lesions and RTEL may depend on RTEL levels and could vary between RTEL isoforms. In the absence of RTEL, theG4 lesion could trigger DNA “repair” by homologous recombination. Involvement of the 3� chromosome ends in the initial lesion or thesubsequent recombination reactions could result in deletions at telomeric sites (Figures 4C–4G) and more diverse abnormalities at intrachromo-somal sites (Figure 5).
of the single-strand overhang (Makarov et al., 1997, Rtel Is Required to Prevent ChromosomalInstability in Differentiating ES CellsMcElligott and Wellinger, 1997) in the recombination
reaction (Lustig, 2003; Zhu et al., 2003). The very rapid onset of chromosomal instability in aminor subset of differentiating Rtel�/� ES cells (FigureThe above model is consistent with several of the
observations reported here. First, the telomere length 5) is surprising. While chromosome fusions were mostabundant after 48 hr, other types of abnormalities werein Rtel�/� ES shows an unusual distribution compatible
with the presence of wild-type as well as variably short- already observed in a minority of cells 4 hr followinginduction of differentiation (Figure 5). What could beened telomeres (Figures 4D and G). G4 DNA at telomeres
could presumably form anywhere along a telomere tract, the immediate change(s) that trigger the chromosomalinstability and demise of differentiating Rtel�/� ES cellsresulting in deletions of variable size. Second, we did
observe chromosomal abnormalities in differentiating and embryos? It has been reported that DNA damageresponses involving p53 are activated upon differentia-Rtel�/� ES cells (Figure 6J) that indicate that genetic
instability in those cells is not restricted to telomeres, tion (Aladjem et al., 1998). However, we could not rescuethe Rtel�/� mutant phenotype by breeding with p53�/�consistent with recombination between telomeric DNA
and intrachromosomal sequences (Figures 6J and 6K mice (data not shown). We assume that the geneticinstability in differentiating Rtel�/� ES is triggered byand legend). A role for RTEL in preventing illegitimate
recombination is also compatible with the high level of either posttranslational modifications of proteins orchanges in gene expression. For example, a decreaseRTEL expression in meiotic cells of the testis (Figure
2B). However, Rtel�/� ES cells did not show deletions in telomerase activity upon differentiation (Armstrong etal., 2000) could explain the increased number of fusionssimilar to those observed in dog-1 mutant animals (see
Supplemental Data for data and further discussion). in differentiating Rtel�/� ES cells (Figure 5L), as active

Regulation of Telomere Length by Rtel883
telomerase could be required to rescue critically short aspects of DNA replication and processing, and it hasbeen proposed that sequential and regulated bindingtelomeres (Figure 7). Alternatively, telomerase in undif-
ferentiated ES cells could “cap” the single-strand over- of PIP motif-containing proteins to PCNA may contributeto the ordering of events during DNA replication andhang present at telomeres and prevent their involvement
in recombination reactions (Blackburn, 2000). Clearly, repair (Warbrick, 2000). In Saccharomyces cerevisiae,the DNA helicase Rrm3p both interacts with PCNA viaRtel�/� ES cells provide an excellent model for further
studies of DNA damage responses and telomerase ac- a PIP-box motif (Schmidt et al., 2002) and promotesreplication fork progression through telomeric andtivity in relation to differentiation and genetic instability.subtelomeric DNA (Ivessa et al., 2002). Telomerase alsointeracts with PCNA and the lagging-strand DNA replica-Rtel and the Differences in Telomere Lengthtion machinery in S. pombe (Dahlen et al., 2003) and,between Murine Speciesduring macronuclear development but not in vegeta-The chromosomal location of Rtel and its role in thetively growing cells, in the ciliate Euplotes crassus (Rayelongation of telomeres in chromosomes derived fromet al., 2002). Interactions of RTEL and telomerase withthe M. spretus parent in F1 cells (Figure 6) are compati-PCNA and other components of the replication and DNAble with Rtel being the candidate gene(s) previouslyrepair machinery could be developmentally controlledimplicated in telomere length regulation (Zhu et al.,in mammalian cells as well, perhaps explaining the in-1998). Based on the observations reported here, wecreased genetic instability in Rtel�/� ES cells upon differ-propose that RTEL, by preventing recombination be-entiation. Presently, it is not clear if RTEL is part of thetween telomere repeats, is a primary determinant ofnormal telomere replication machinery or is recruitedtelomere length in the mouse. How is Rtel related to theto replication forks stalled at G4 DNA. While RTEL isdifferences in telomere length between M. spretus andlocalized in the nucleus (Figures 2C and 2E), the majorityM. musculus? The RTEL protein sequence of both spe-of RTEL foci did not show clear overlap with PCNAcies is very similar, and no differences in overall expres-(Figures 2D and 2F–2H), and further studies are neededsion pattern or promoter sequences have been identi-to distinguish between these possibilities.fied. However, we did note differences in Rtel transcripts
between the two species (Figure 1C). Whereas full-length M. musculus Rtel transcripts were readily ob-
Functional Relationship of Rtel to Other Telomeretained, full-length M. spretus Rtel transcripts were rareMaintenance Factors(Figure 1C). The predominant M. spretus transcript uti-Telomere length is regulated by highly dynamic andlizes a secondary 3� splice site in exon 32, which resultscomplex processes that are cell cycle-dependent andin the deletion of 18 nt. A possible explanation for thisdevelopmentally controlled (Zakian, 1996; McEacherndifference is a nucleotide divergence between the twoet al., 2000; Ray et al., 2002). A surprisingly large numberMus species at position –24 relative to exon 32. Thisof helicases has been implicated in telomere homeosta-nucleotide difference creates a new binding site forsis. In S. cerevisiae, at least four different helicases,U2AF65, the large subunit of U2 snRNP auxiliary factorincluding Pif1p, Rrm3p, Sgs1p, and Dna2p, have been(Maniatis and Tasic, 2002; Kent et al., 2003) and, as areported to be involved in telomere function (Schulz andconsequence, an alternative 3� splice site for exon 32Zakian, 1994; Ivessa et al., 2002; Choe et al., 2002). Inin M. spretus that does not exist in M. musculus. Differ-humans, members of the RecQ helicase family, includingences resulting from alternative splicing of Rtel tran-the Werner and Bloom syndrome helicases, are knownscripts affect the carboxy terminus of the RTEL proteinto interact with telomeres (Schulz et al., 1996; Opreskoand create a novel CK1 phosphorylation site potentiallyet al., 2002; Hickson, 2003). Here, we demonstrate thatdisturbing the predicted functional interaction withRTEL is yet another helicase-like protein with an impor-PCNA. Further studies are needed to clarify if alternativetant role in telomere maintenance. It is possible thatsplicing indeed provides an explanation for the apparentother helicases perform functions similar to RTEL. Forhypomorphic M. spretus Rtel allele and to delineate theexample, in cells with insufficient expression of RTELprecise difference in RTEL protein structure, expression,(or in cells that do not express the correct RTEL part-or posttranslational modification between the two spe-ners), the proposed function of RTEL could be executedcies in relation to their widely different telomere lengths.by BRCA1 binding helicase-like protein BACH1 (Cantoret al., 2001) or by members of the RecQ helicase familyRole of RTEL at Telomeres(Hickson, 2003). Stochastic loss of telomeres has beenIn addition to telomere length variation between differentreported in cells deficient in WRN protein (Schulz etorganisms, large variations in the telomere length atal., 1996; Bai and Murnane, 2003), and the defectiveindividual chromosome ends also exist in individual hu-helicases associated with Werner and Bloom syndromeman and murine cells (Lansdorp et al., 1996; Martensare known to be stimulated by interaction with the telo-et al., 2000; Zijlmans et al., 1997). For example, verymere binding protein TRF2 (Opresko et al., 2002) and areshort telomeres were also observed, be it at a low fre-capable of unwinding telomeric G4 DNA (Mohaghegh etquency, in Rtel�/� ES cells (Figure 4E). Perhaps RTELal., 2001). These proteins could also play a role in theactivity in wild-type cells increases the chance of (butresolution of higher order structures of telomeric DNAdoes not guarantee) successful resolution of higher or-and thereby assist in the maintenance of telomere lengthder structures of G-rich DNA. The limiting factor couldand function. Future studies will no doubt reveal the rolebe the chance of a functional interaction between RTELof RTEL relative to these other molecules in telomereand proliferating cell nuclear antigen (PCNA) via the PIP
box motif in RTEL (Figure 1D). PCNA is involved in many function and the growth of normal and malignant cells.

Cell884
Experimental Procedures Derivation of Rtel�/� ES CellsRtel�/� mice (129S1 background) were intercrossed, and the daythat vaginal plugs were detected was designated embryonic dayIsolation of Rtel Genomic DNA
Murine genomic DNA containing the Rtel gene was obtained from 0.5 (E0.5). Blastocysts were flushed and then cultured in the ESculture medium to establish cell lines.BAC RPCI-23-60M5 (CIHR Genome Resource Facility, Toronto, Can-
ada), which was identified by hybridization with EST AA030845 (ex-ons 22–34 of Rtel) as well as with cDNAs from the flanking genes Culture and Cell Cycle Analysis of ES CellsStathim3 (AA058212) and Arfrp1 (all from ATCC, Manassas, VA). For Rtel�/�, Rtel �/�, and Rtel �/� ES cells were maintained under stan-determination of the exon/intron structure, EcoRI or EcoRI/NcoI dard culture conditions in the presence of LIF on gelatin coatedfragments, subcloned in pBluescript KS II� (Stratagene), were iso- dishes, as described previously (Gertsenstein et al., 2002). Forlated and hybridized with the full-length cDNA. Plasmids containing in vitro differentiation in semisolid medium, the cells were platedcoding sequences were sequenced at the NAPS facility (University at 1500 cells per dish in methylcellulose medium (ES-Cult M3120,of British Columbia, Vancouver, Canada). To identify the transcrip- StemCell Technologies Inc., Vancouver, Canada). For differentiationtion start site, 5� RACE was performed using Marathon-Ready cDNA in liquid culture, cells were plated in medium without LIF onto culturefrom Balb/c testis (BD Biosciences Clontech, Palo Alto, CA). Five dishes that were not treated with gelatin. Cell cycle analysis wasdifferent start sites within a 65 bp region were found. The 5� UTR performed as described (Aladjem et al., 1998) with minor modifica-genomic sequence was generated by PCR on DNA isolated from tions. ES cells were grown with or without LIF for 30 min at 37C inBalb/c liver. Genomic DNA was isolated from skin fibroblasts of the presence of 30 �M BrdU prior to fixation with ethanol and stain-M. spretus (Jackson Laboratory, Bar Harbor, ME), and sequencing ing with propidium iodide and anti-BrdU-FITC antibody (Bectonwas done on amplified genomic DNA. Dickinson, San Jose, CA) following the protocol supplied by Becton
Dickinson. Flow cytometry data files were acquired on a FACSCali-bur Flow Cytometer (Becton Dickinson), and data were analyzedCloning of Rtel cDNAusing CellQuest and WinMDI software.RNA was isolated from M. spretus and M. musculus (Balb/c) adult
testis using Trizol (Invitrogen). First strand synthesis was done usingoligo dT and Superscript II (Invitrogen). Full-length Balb/c and M. Telomere Length Analysisspretus cDNAs were assembled using the 5� primer with an EcoRI The average telomere length in various cells was measured by flowsite inserted A(AAGAATTCAGGCTGATATGCCCAGGGTA), a 3� FISH (Rufer et al., 1998) using modifications (including fixed bovineprimer also with an EcoRI site (GAGAATTCCCCTATTGCTTACTC thymocytes as internal controls) as described (Baerlocher and Lans-CGA), and internal forward (TGTGATTGTCACGGGCCTCCC) and re- dorp, 2003). Q-FISH analysis was performed as previously describedverse (CAATAGCCTGGTTCACAGCCC) primers. (Lansdorp et al., 1996; Zijlmans et al., 1997). Results of Q-FISH
Analysis of splice variants was done using the following primers: analysis are expressed in arbitrary units rather than telomere fluores-forward (exon 23) CGTGTTGTCCTCAAGATGCAG, reverse (exon 34) cence units (TFU), because calibration based on hybridizations withTTGCTTACTCCGATGAGGCTTC. plasmid DNA containing telomere inserts resulted in shorter average
telomere values than were actually measured by flow FISH. Chromo-somes from the M. spretus parent in cells derived from F1 animalsGeneration of Mice Deficient for Rtelobtained from crosses between a M. musculus (129Sl background)The mouse Rtel genomic DNA fragments were cloned by PCR basedfemale and a M. spretus male were identified by including fluores-on the mouse Rtel genomic sequence in the Celera database tocein labeled peptide nucleic acid (PNA) specific for repetitive DNAbuild a target vector, which was designed to replace a 5 kb genomicpresent at centromeres (Fl-OO-CTTCGTTGGAAACGGGGT) at 0.3fragment containing exons 7–9 with an SA-IRES-�geopA expression�g/ml together with Cy3-labeled (CCCTAA)3 PNA probe in the hy-cassette. The targeting construct was linearized with NotI and elec-bridization mixture. Analysis of terminal restriction fragments bytroporated into R1 ES cells and selected with G418 as describedSouthern blot was performed as previously described (Harley etpreviously (Gertsenstein et al., 2002). Southern blot analysis wasal., 1990).carried out to screen for the presence of a disrupted Rtel gene using
BamHI (for the 5� external probe) and NsiI digestion (for the 3�
Acknowledgmentsexternal probe). We used two independently targeted ES cell clonesto generate chimeric mice that subsequently were bred with 129S1
We thank Gloria Shaw for help with cytogenetic procedures; Camfemales to obtain germ-line transmission. The phenotypes of Rtel�/�
Smith for performing colony-forming assays; Susan McPhail andmutants derived from both targeted ES cell lines were indistin-Peggy Olive for help with immunofluorescence procedures; Jackyguishable.Schein for analysis of BACs; Michael Schulzer for help with statisticalanalysis; Dixie Mager and Keith Humphries for helpful discussions;Genotypingand Carolyn Price, Lea Harrington, and Peter Rowe for criticallyPCR and Southern blot analysis were applied for genotyping. PCRreading the manuscript. Peptide nucleic acid probes were gener-was performed on ear-punched DNA. Primers to amplify the targetedously provided by Applied Biosystems, Bedford, MA. This work isallele were sense primer, located in intron 6 (TGTGTTTCTAGCCTCTsupported by the Terry Fox Foundation (P.M.L.); a CIHR grant toGCAGCT) and antisense primer specific for the SA-IRES-�geopAA.N.; and a Neuromuscular Disease Research Development Grantcassette (GGGACAGGATAAGTATGACATCA). The primers to am-from the American Muscular Dystrophy Association (to H.D.). H.D.plify the wild-type Rtel allele were sense primer locating on intronis the holder of CRC Chair. P.P.L.T. is a Senior Principal Research7 (CCTTGAGGCTCTGTGTACAGA) and antisense primer locating onFellow of the NHMRC of Australia. A.N. is a senior scientist of theintron 8 (GACATCAGTAGCCATCAGCTT). Southern blot analysisCIHR.was undertaken using standard protocols.
Received: November 17, 2003Northern Blot AnalysisRevised: April 23, 2004Total RNA from flash-frozen mouse tissues was extracted usingAccepted: April 28, 2004TRIzol (Life Technologies, Inc.), while total RNA from the cells wasPublished: June 24, 2004purified using the RNeasy Mini Kit (Qiagen). RNA (20 �g of total)
was fractionated on a 1% agarose-formaldehyde gel and transferredReferencesto Hybond nylon membrane (Amersham). Hybridization was carried
out in ExpressHyb (Clontech) containing 2 � 106 cpm/ml probe.Aladjem, M.I., Spike, B.T., Rodewald, L.W., Hope, T.J., Klemm, M.,Jaenisch, R., and Wahl, G.M. (1998). ES cells do not activate p53-Subcellular Expression of RTELdependent stress responses and undergo p53-independent apopto-Details of the experimental procedures used to study the expressionsis in response to DNA damage. Curr. Biol. 8, 145–155.and subcellular localization of epitope-tagged RTEL are described
in Supplemental Data. Armstrong, L., Lako, M., Lincoln, J., Cairns, P.M., and Hole, N. (2000).

Regulation of Telomere Length by Rtel885
mTert expression correlates with telomerase activity during the dif- Heterogeneity in telomere length of human chromosomes. Hum.Mol. Genet. 5, 685–691.ferentiation of murine embryonic stem cells. Mech. Dev. 97, 109–116.
Lustig, A.J. (2003). Clues to catastrophic telomere loss in mammalsBaerlocher, G.M., and Lansdorp, P.M. (2003). Telomere length mea-from yeast telomere rapid deletion. Nat. Rev. Genet. 4, 916–923.surements in leukocyte subsets by automated multicolor flow-FISH.
Cytometry 55A, 1–6. Makarov, V.L., Hirose, Y., and Langmore, J.P. (1997). Long G tails atboth ends of human chromosomes suggest a C strand degradationBai, Y., and Murnane, J.P. (2003). Telomere instability in a humanmechanism for telomere shortening. Cell 88, 657–666.tumor cell line expressing a dominant-negative WRN protein. Hum.
Genet. 113, 337–347. Maniatis, T., and Tasic, B. (2002). Alternative pre-mRNA splicingand proteome expansion in metazoans. Nature 418, 236–243.Bai, C., Connolly, B., Metzker, M.L., Hilliard, C.A., Liu, X., Sandig,
V., Soderman, A., Galloway, S.M., Liu, Q., Austin, C.P., and Caskey, Marchler-Bauer, A., Anderson, J.B., DeWeese-Scott, C., Fedorova,C.T. (2000). Overexpression of M68/DcR3 in human gastrointestinal N.D., Geer, L.Y., He, S., Hurwitz, D.I., Jackson, J.D., Jacobs, A.R.,tract tumors independent of gene amplification and its location in Lanczycki, C.J., et al. (2003). CDD: a curated Entrez database ofa four-gene cluster. Proc. Natl. Acad. Sci. USA 97, 1230–1235. conserved domain alignments. Nucleic Acids Res. 31, 383–387.Blackburn, E.H. (2000). Telomere states and cell fates. Nature Martens, U.M., Chavez, E.A., Poon, S.S., Schmoor, C., and Lansdorp,408, 53–56. P.M. (2000). Accumulation of short telomeres in human fibroblasts
prior to replicative senescence. Exp. Cell Res. 256, 291–299.Blackburn, E.H. (2001). Switching and signaling at the telomere. Cell106, 661–673. McEachern, M.J., Krauskopf, A., and Blackburn, E.H. (2000). Telo-
meres and their control. Annu. Rev. Genet. 34, 331–358.Blasco, M.A., Lee, H.-W., Hande, M.P., Samper, E., Lansdorp, P.M.,DePinho, R.A., and Greider, C.W. (1997). Telomere shortening and McElligott, R., and Wellinger, R.J. (1997). The terminal DNA structuretumor formation by mouse cells lacking telomerase RNA. Cell 91, of mammalian chromosomes. EMBO J. 16, 3705–3714.25–34. Mieczkowski, P.A., Mieczkowska, J.O., Dominska, M., and Petes,Cantor, S.B., Bell, D.W., Ganesan, S., Kass, E.M., Drapkin, R., Gross- T.D. (2003). Genetic regulation of telomere-telomere fusions in theman, S., Wahrer, D.C., Sgroi, D.C., Lane, W.S., Haber, D.A., and yeast Saccharomyces cerevisae. Proc. Natl. Acad. Sci. USA 100,Livingston, D.M. (2001). BACH1, a novel helicase-like protein, inter- 10854–10859.acts directly with BRCA1 and contributes to its DNA repair function. Mohaghegh, P., Karow, J.K., Brosh, J.R., Jr., Bohr, V.A., and Hick-Cell 105, 149–160. son, I.D. (2001). The Bloom’s and Werner’s syndrome proteins are
DNA structure-specific helicases. Nucleic Acids Res. 29, 2843–2849.Cheung, I., Schertzer, M., Rose, A., and Lansdorp, P.M. (2002). Dis-ruption of dog-1 in Caenorhabditis elegans triggers deletions up- Moyzis, R.K., Buckingham, J.M., Cram, L.S., Dani, M., Deaven, L.L.,stream of guanine-rich DNA. Nat. Genet. 31, 405–409. Jones, M.D., Meyne, J., Ratliff, R.L., and Wu, J.-R. (1988). A highly
conserved repetitive DNA sequence, (TTAGGG)n, present at the telo-Choe, W., Budd, M., Imamura, O., Hoopes, L., and Campbell, J.L.meres of human chromosomes. Proc. Natl. Acad. Sci. USA 85, 6622–(2002). Dynamic localization of an Okazaki fragment processing pro-6626.tein suggests a novel role in telomere replication. Mol. Cell. Biol.
22, 4202–4217. Opresko, P.L., von Kobbe, C., Laine, J.P., Harrigan, J., Hickson, I.D.,and Bohr, V.A. (2002). Telomere-binding protein TRF2 binds to andDahlen, M., Sunnerhagen, P., and Wang, T.S. (2003). Replicationstimulates the Werner and Bloom syndrome helicases. J. Biol.proteins influence the maintenance of telomere length and telom-Chem. 277, 41110–41119.erase protein stability. Mol. Cell. Biol. 23, 3031–3042.
Parkinson, G.N., Lee, M.P., and Neidle, S. (2002). Crystal structurede Lange, T. (2002). Protection of mammalian telomeres. Oncogeneof parallel quadruplexes from human telomeric DNA. Nature 417,21, 532–540.876–880.Gertsenstein, M., Lobe, C., and Nagy, A. (2002). ES cell-mediatedPetes, T.D., and Hill, C.W. (1988). Recombination between repeatedconditional transgenesis. Methods Mol. Biol. 185, 285–307.genes in microorganisms. Annu. Rev. Genet. 22, 147–168.Greider, C.W. (1996). Telomere length regulation. Annu. Rev. Bio-Ray, S., Karamysheva, Z., Wang, L., Shippen, D.E., and Price, C.M.chem. 65, 337–365.(2002). Interactions between telomerase and primase physically link
Greider, C.W., and Blackburn, E.H. (1985). Identification of a specificthe telomere and chromosome replication machinery. Mol. Cell. Biol.
telomere terminal transferase activity in Tetrhymena extracts. Cell22, 5859–5868.
43, 405–413.Rufer, N., Dragowska, W., Thornbury, G., Roosnek, E., and Lansdorp,
Griffith, J.D., Comeau, L., Rosenfield, S., Stansel, R.M., Bianchi, A.,P.M. (1998). Telomere length dynamics in human lymphocyte sub-
Moss, H., and de Lange, T. (1999). Mammalian telomeres end in apopulations measured by flow cytometry. Nat. Biotechnol. 16,
large duplex loop. Cell 97, 503–514.743–747.
Hackett, J.A., and Greider, C.W. (2003). End resection initiates geno-Schmidt, K.H., Derry, K.L., and Kolodner, R.D. (2002). Saccharo-
mic instability in the absence of telomerase. Mol. Cell. Biol. 23, 8450–myces cerevisiae RRM3, a 5� to 3� DNA helicase, physically interacts
8461.with proliferating cell nuclear antigen. J. Biol. Chem. 277, 45331–
Harley, C.B., Futcher, A.B., and Greider, C.W. (1990). Telomeres 45337.shorten during ageing of human fibroblasts. Nature 345, 458–460. Schulz, V.P., and Zakian, V.A. (1994). The saccharomyces PIF1 DNAHickson, I.D. (2003). RecQ helicases: caretakers of the genome. Nat. helicase inhibits telomere elongation and de novo telomere forma-Rev. Cancer 3, 169–178. tion. Cell 76, 145–155.
Ivessa, A.S., Zhou, J.Q., Schulz, V.P., Monson, E.K., and Zakian, Schulz, V.P., Zakian, V.A., Ogburn, C.E., McKay, J., Jarzebowicz,V.A. (2002). Saccharomyces Rrm3p, a 5� to 3� DNA helicase that A.A., Edland, S.D., and Martin, G.M. (1996). Accelerated loss ofpromotes replication fork progression through telomeric and telomeric repeats may not explain accelerated replicative declinesubtelomeric DNA. Genes Dev. 16, 1383–1396. of Werner syndrome cells. Hum. Genet. 97, 750–754.
Kent, O.A., Reayi, A., Foong, L., Chilibeck, K.A., and MacMillan, A.M. Sen, D., and Gilbert, W. (1988). Formation of parallel four-stranded(2003). Structuring of the 3� splice site by U2AF65. J. Biol. Chem. complexes by guanine-rich motifs in DNA and its implications for278, 50572–50577. meiosis. Nature 334, 364–366.
Kreegipuu, A., Blom, N., and Brunak, S. (1999). PhosphoBase, a Singer, M.S., and Gottschling, D.E. (1994). TLC1: template RNA com-database of phosphorylation sites: release 2.0. Nucleic Acids Res. ponent of Saccharomyces cerevisiae telomerase. Science 266,27, 237–239. 404–409.
Takai, H., Smogorzewska, A., and de Lange, T. (2003). DNA damageLansdorp, P.M., Verwoerd, N.P., van de Rijke, F.M., Dragowska,V., Little, M.-T., Dirks, R.W., Raap, A.K., and Tanke, H.J. (1996). foci at dysfunctional telomeres. Curr. Biol. 13, 1549–1556.

Cell886
Vega, L.R., Mateyak, M.K., and Zakian, V.A. (2003). Getting to theend: telomerase access in yeast and humans. Nat. Rev. Mol. CellBiol. 4, 948–959.
von Zglinicki, T. (2002). Oxidative stress shortens telomeres. TrendsBiochem. Sci. 27, 339–344.
Warbrick, E. (2000). The puzzle of PCNA’s many partners. Bioessays22, 997–1006.
Williamson, J.R., Raghuraman, M.K., and Cech, T.R. (1989). Monova-lent cation-induced structure of telomeric DNA: the G-quartet model.Cell 59, 871–880.
Zakian, V.A. (1995). Telomeres: beginning to understand the end.Science 270, 1601–1607.
Zakian, V.A. (1996). Structure, function, and replication of Saccharo-myces cerevisiae telomeres. Annu. Rev. Genet. 30, 141–172.
Zhu, L., Hathcock, K.S., Hande, P., Lansdorp, P.M., Seldin, M.F.,and Hodes, R.J. (1998). Telomere length regulation in mice is linkedto a novel chromosome locus. Proc. Natl. Acad. Sci. USA 95, 8648–8653.
Zhu, X.D., Niedernhofer, L., Kuster, B., Mann, M., Hoeijmakers, J.H.,and de Lange, T. (2003). ERCC1/XPF removes the 3� overhang fromuncapped telomeres and represses formation of telomeric DNA-containing double minute chromosomes. Mol. Cell 12, 1489–1498.
Zijlmans, J.M., Martens, U.M., Poon, S.S., Raap, A.K., Tanke, H.J.,Ward, R.K., and Lansdorp, P.M. (1997). Telomeres in the mousehave large inter-chromosomal variations in the number of T2AG3
repeats. Proc. Natl. Acad. Sci. USA 94, 7423–7428.
Accession Numbers
The GenBank accession numbers for Rtel full-length, promoter, andsplice variant sequences (see Figure 1C) are the following: M. mus-culus, AY481619, AY48618, AY481620, AY481621, AY481622,AY481623, and AY481624; and M. spretus, AY530632, AY48612,AY481613, AY481614, AY481615, AY481616, and AY481617.




















