
Human cerebellar responses to brush and heat stimuli in healthy and neuropathic pain subjects
21
ORIGINAL ARTICLE Human cerebellar responses to brush and heat stimuli in healthy and neuropathic pain subjects D. BORSOOK 1 , E. A. MOULTON 1 , S. TULLY 1 , J. D. SCHMAHMANN 2 & L. BECERRA 1 1 PAIN Group, Brain Imaging Center, McLean Hospital, Harvard Medical School, and 2 Cerebellar Research Group, Ataxia Unit, Department of Neurology, Massachusetts General Hospital and Harvard Medical School, Massachusetts, USA Abstract Though human pain imaging studies almost always demonstrate activation in the cerebellum, the role of the cerebellum in pain function is not well understood. Here we present results from two studies on the effects of noxious thermal heat and brush applied to the right side of the face in a group of healthy subjects (Group I) and a group of patients with neuropathic pain (Group II) who are more sensitive to both thermal and mechanical stimuli. Statistically significant activations and volumes of activations were defined in the cerebellum. Activated cerebellar structures were identified by colocalization of fMRI activation with the ‘MRI Atlas of the Human Cerebellum’. Functional data (obtained using a 3T magnet) were defined in terms of maximum voxels and volume of activation in the cerebellum. Volume maps were then mapped onto two millimeter serial slices taken through the cerebellum in order to identify activation within regions defined by the activation volume. The data indicate that different regions of the cerebellum are involved in acute and chronic pain processing. Heat produces greater contralateral activation compared with brush, while brush resulted in more ipsilateral/bilateral cerebellar activation. Further, innocuous brush stimuli in healthy subjects produced decreased cerebellar activation in lobules concerned with somatosensory processing. The data also suggest a dichotomy of innocuous stimuli/sensorimotor cerebellum activation versus noxious experience/cognitive/limbic cerebellum activation. These results lead us to propose that the cerebellum may modulate the emotional and cognitive experience that distinguishes the perception of pain from the appreciation of innocuous sensory stimulation. Key words: trigemino-cerebellar pathways, chronic pain, allodynia, cerebellar muclei, pain, fMRI, BOLD Introduction The cerebellum may have a role in a number of integrative functions including memory, associative learning, motor control (1–3), and more recently in sensory processing including nociception (4). The structure is traditionally divided into a medial zone involved in somatic and autonomic reflexes as well as complex movements; an intermediate zone that is involved in voluntary movements; and a lateral zone involved in higher order functions such as memory and cognitive functions in association with the cortex including language (5–7). In addition, connections between the cerebellum and the hypothalamus suggest a possible role in autonomic function, as well as a link to limbic structures involved with emotion (8–10). Thus, the cerebellum appears to be involved in integrating motor, sensory, autonomic, and cognitive responses to environmental stimuli including acute and chronic pain. Most fMRI studies of pain show activation in the cerebellum (See Table I) mostly described as either midline (vermis) or in the cerebellar hemispheres (11,12). One fMRI report specifically evaluated the effects of pain in the cerebellum (13). In the latter, a parametric analysis of the responses to four subject- applied temperatures, ranging in intensity from innocuous to painful, suggests that stimulus-inten- sity could be encoded in the vermis and ipsilateral hemispheric lobule VI. In addition, a number of cerebellar areas activated non-discriminately to different temperatures, including the anterior vermis (lobules III-V), contralateral lobule VIII, and bilat- erally in hemispheric lobule III-VI. Here we have taken separate studies of mechanical and thermal heat stimuli applied to the region of the right maxillary division of the trigeminal nerve in a group of healthy subjects (Moulton et al., submitted) and in a group of patients with chronic neuropathic pain (14), and mapped cerebellar activations evoked by thermal (heat) and mechanical (brush) stimuli. Cerebellar structures were identified using the atlas of the cerebellum (15). Although the studies were separate (14, Moulton et al., submitted), the Correspondence: David Borsook, MD, PhD, PAIN Group, Brain Imaging Center, McLean Hospital, 115 Mill Street, Belmont, MA, USA. E-mail: [email protected] The Cerebellum 2008, –2 ISSN 1473-4222 print/ISSN 1473-4230 online # 200 DOI: 10.10 8 Springer Science + Business Media, LLC 07/s12311-008-0011-6 Online first: 2008 3 April 252 72
-
Upload
onlineadvocate -
Category
Documents
-
view
0 -
download
0
Transcript of Human cerebellar responses to brush and heat stimuli in healthy and neuropathic pain subjects

ORIGINAL ARTICLE
Human cerebellar responses to brush and heat stimuli in healthy andneuropathic pain subjects
D. BORSOOK1, E. A. MOULTON1, S. TULLY1, J. D. SCHMAHMANN2 & L. BECERRA1
1PAIN Group, Brain Imaging Center, McLean Hospital, Harvard Medical School, and 2Cerebellar Research Group, Ataxia
Unit, Department of Neurology, Massachusetts General Hospital and Harvard Medical School, Massachusetts, USA
AbstractThough human pain imaging studies almost always demonstrate activation in the cerebellum, the role of the cerebellum inpain function is not well understood. Here we present results from two studies on the effects of noxious thermal heat andbrush applied to the right side of the face in a group of healthy subjects (Group I) and a group of patients with neuropathicpain (Group II) who are more sensitive to both thermal and mechanical stimuli. Statistically significant activations andvolumes of activations were defined in the cerebellum. Activated cerebellar structures were identified by colocalization offMRI activation with the ‘MRI Atlas of the Human Cerebellum’. Functional data (obtained using a 3T magnet) weredefined in terms of maximum voxels and volume of activation in the cerebellum. Volume maps were then mapped onto twomillimeter serial slices taken through the cerebellum in order to identify activation within regions defined by the activationvolume. The data indicate that different regions of the cerebellum are involved in acute and chronic pain processing. Heatproduces greater contralateral activation compared with brush, while brush resulted in more ipsilateral/bilateral cerebellaractivation. Further, innocuous brush stimuli in healthy subjects produced decreased cerebellar activation in lobulesconcerned with somatosensory processing. The data also suggest a dichotomy of innocuous stimuli/sensorimotorcerebellum activation versus noxious experience/cognitive/limbic cerebellum activation. These results lead us to proposethat the cerebellum may modulate the emotional and cognitive experience that distinguishes the perception of pain from theappreciation of innocuous sensory stimulation.
Key words: trigemino-cerebellar pathways, chronic pain, allodynia, cerebellar muclei, pain, fMRI, BOLD
Introduction
The cerebellum may have a role in a number of
integrative functions including memory, associative
learning, motor control (1–3), and more recently in
sensory processing including nociception (4). The
structure is traditionally divided into a medial zone
involved in somatic and autonomic reflexes as well as
complex movements; an intermediate zone that is
involved in voluntary movements; and a lateral zone
involved in higher order functions such as memory
and cognitive functions in association with the cortex
including language (5–7). In addition, connections
between the cerebellum and the hypothalamus
suggest a possible role in autonomic function, as
well as a link to limbic structures involved with
emotion (8–10). Thus, the cerebellum appears to be
involved in integrating motor, sensory, autonomic,
and cognitive responses to environmental stimuli
including acute and chronic pain.
Most fMRI studies of pain show activation in the
cerebellum (See Table I) mostly described as either
midline (vermis) or in the cerebellar hemispheres
(11,12). One fMRI report specifically evaluated the
effects of pain in the cerebellum (13). In the latter, a
parametric analysis of the responses to four subject-
applied temperatures, ranging in intensity from
innocuous to painful, suggests that stimulus-inten-
sity could be encoded in the vermis and ipsilateral
hemispheric lobule VI. In addition, a number of
cerebellar areas activated non-discriminately to
different temperatures, including the anterior vermis
(lobules III-V), contralateral lobule VIII, and bilat-
erally in hemispheric lobule III-VI. Here we have
taken separate studies of mechanical and thermal
heat stimuli applied to the region of the right
maxillary division of the trigeminal nerve in a group
of healthy subjects (Moulton et al., submitted) and
in a group of patients with chronic neuropathic pain
(14), and mapped cerebellar activations evoked by
thermal (heat) and mechanical (brush) stimuli.
Cerebellar structures were identified using the atlas
of the cerebellum (15). Although the studies were
separate (14, Moulton et al., submitted), the
Correspondence: David Borsook, MD, PhD, PAIN Group, Brain Imaging Center, McLean Hospital, 115 Mill Street, Belmont, MA, USA. E-mail:
The Cerebellum
2008, –2
ISSN 1473-4222 print/ISSN 1473-4230 online # 200
DOI: 10.10
8 Springer Science + Business Media, LLC
07/s12311-008-0011-6
Online first: 20083 April
252 72

experimental imaging paradigm was similar across
the groups of healthy subjects and patients with a
sensitized pain state (see Methods).
Both data from human and animal studies strongly
implicate the cerebellum in the modulation of pain. In
human studies, for example, transcranial magnetic
stimulation of the cerebellum produces alteration of
sensory thresholds and attenuation of cold pain
sensation (16); functional imaging studies show
cerebellar activation to pain stimuli (see above);
Table I. Functional imaging of cerebellar activation in acute and neuropathic pain studies.
(A) Experimental pain (evoked stimuli).
Paper Method Stimulus Site Dominant activation Ref
Hsieh 1996 PET Ethanol injection R arm I (R) (105)
Casey 1996 PET Thermode L arm B (106)
Cold water bath L hand B
Svensson 1997 PET Laser L arm I (L) (107)
Electric L arm I (L)
Xu 1997 PET Laser L hand I (L) (108)
Derbyshire 1998 PET Hot water bath R hand I (R) (109)
Iadarola 1998 PET Capsaicin injection L arm B (110)
May 1998 PET Capsaicin injection R forehead B (111)
Paulson 1998 PET Thermode L arm I (L – males) (112)
B (females)
Becerra 1999 fMRI Thermode L hand B (113)
Coghill 1999 PET Thermode R arm B (114)
Peyron 1999 PET Thermode R/L hand B (115)
Becerra 2001 fMRI Thermode L hand B (116)
Casey 2001 PET Thermode L arm B (117)
Coghill 2001 PET Thermode R/L arm I (118)
Bingel 2002 fMRI Laser R/L hand B (119)
Derbyshire 2002 PET Thermode R hand B (120)
Helmchen 2003 fMRI Thermode R hand B (121)
Koyama 2003 fMRI Thermode R calf B (122)
Strigo 2003 fMRI Thermode Esophagus B (123)
Upper chest B
Helmchen 2004 fMRI Thermode R hand I (R) (13)
Ibinson 2004 fMRI Electric R arm I (R) (124)
Wager 2004 fMRI Electric R wrist I (R) (125)
Thermode L arm I (L)
Wiech 2005 fMRI Thermode L arm C (R) (126)
Albuquerque 2006 fMRI Thermode R masseter I (R) (127)
Choi 2006 fMRI Hot water bath L finger C (R) (33)
Kong 2006 fMRI Thermode R arm B (128)
Seminowicz 2006 fMRI Electric L arm C (R) (129)
Staud 2006 fMRI Thermode R foot I (R) (62)
B, bilateral; I, ipsilateral; C, contralateral; (L), left; (R), right.
(B) Studies on cognitive processes affecting pain-related activation.
Paper Method Cognitive process Stimulus Site Effect Ref
Ploghaus 1999 fMRI Anticipation Before heat pain L hand I (L) + (96)
Bantick 2002 fMRI Distraction During heat pain L hand B 2 (130)
Brooks 2002 fMRI Attention During heat pain R hand B + (131)
Distraction During heat pain R hand I (R) +Attention During heat pain L hand I (L) +Distraction During heat pain L hand none
Smith 2002 fMRI Anticipation Before heat pain L hand B + (132)
Gracely 2004 fMRI Catastrophizing During pressure pain L finger I (L) + (83)
Singer 2004 fMRI Empathy Partner in pain (Visual) B + (18)
Wager 2004 fMRI Placebo During heat pain L arm I 2 (125)
Jackson 2005 fMRI Empathy Pain-related pictures (Visual) B + (17)
Wiech 2005 fMRI Distraction During heat pain L arm B 2 (126)
Keltner 2006 fMRI Expectation During heat pain L hand I (L) + (85)
Moriguchi 2006 fMRI Empathy Pain-related pictures (Visual) R + (133)
Ogino 2006 fMRI Imagination Pain-related pictures (Visual) L + (84)
Seminowicz 2006 fMRI Distraction During electric pain L arm C (R) 2 (129)
B, bilateral; I, ipsilateral; (L), left; (R), right; +, increased activation; 2, decreased activation.
253Pain and the cerebellum

empathetic pain also produces similar cerebellar
activation (17,18); and studies of patients with
cerebellar damage impairs detection of somatosen-
sory input changes (19). Animal studies indicate that
cerebellar activation may be modulated by peripheral
afferent sensory inputs (20,21) including pain (22).
The trigeminal system has specific direct or collateral
inputs into the cerebellum (23–26). For example, in
rabbits, the paramedian lobule and the uvula receive
independent trigeminal sensory information from
neurons located in separate regions of the trigeminal
sensory nucleus (23; see Discussion – Trigemino-
cerebellar connections). Deafferentation of the infra-
obital branch of the trigeminal nerve has been
reported to result in reorganization of regions of the
cerebellum including Crus IIa (27). One report
suggests that C-fibers (that are activated by a number
of noxious stimuli including heat), may reach the
cerebellar Purkinje cells through climbing and mossy
fibers (28). Taken together, such clinical and
preclinical data suggest that there are direct and
indirect pathways from sensory systems, including the
trigeminal system, to the cerebellum. The salience of
trigeminal sensory input from the face is not known
but may be interpreted as being important in
orienting the face to protective motor planning (for
example, the blink reflex, nocifensive response to
pain, facial movements). Such processing of sensory
signals may be important in motor planning and
execution (see 29) in which the cerebellum may play
an integrative role. Given that the cerebellum may be
important in modulating sensory stimuli and a motor
response to such stimuli (30), the evaluation of
specific cerebellar functional activity in different
anatomical regions in response to noxious and non-
noxious stimuli in humans may provide insights into
how this structure may integrate or modulate this type
of information.
Methods
Data was obtained from two experiments using a
trigeminal model of pain (for a review, see 31). The
overall approach is shown in Figure 1. In our model,
pain was applied to the right side of the face in the
territory of the second division of the trigeminal
nerve (V2). The two experiments examined: (Group
I) acute noxious thermal pain in healthy subjects
(32), and (Group II) stimuli applied to the same
region of the face in patients with neuropathic pain
(14) affecting the second division of the trigeminal
nerve. All studies were thus similar or identical in the
following aspects – stimulation sites (second division
of the trigeminal nerve, V2); use of the same 3T
scanner; identical equipment for stimulation
(1.661.6 mm thermal contact probe/Peltier;
brush); and all experiments were performed by the
same research team at the same location. The
research protocols were approved by the institutional
IRB and all pain paradigms were conducted in
accordance with the Declaration of Helsinki.
Subjects
All subjects underwent a clinical review of systems
and urine screens for recreational drugs including
phencyclidine, barbiturates, tetra-hydrocannabinol,
opiates, benzodiazepines, amphetamines, and
cocaine. (Cortez Diagnostics 7 Drug RapidDip
INsta-scan). Subjects who were healthy and tested
negative for the drug screens were recruited to the
study. Two groups of subjects were recruited to two
separate studies.
Group I: 12 healthy volunteers (9 men and 3
women), with an average age of 27¡10 years, were
recruited for a study on facial cortical somatotopy
through advertisements circulated in the Boston
metropolitan area. Subjects were imaged once
(Moulton et al., in preparation).
Group II: For details see (14). Only brush and heat
stimuli to the affected side are reported here. Briefly,
6 subjects with right-sided neuropathic pain (1 man
and 5 women), with an average age of 49¡8 years,
were recruited to the study (see Table A). Subjects
were recruited through advertisements placed in the
local newspaper or at physicians’ offices (details are
(C) Neuropathic pain (Evoked stimuli).
Paper Method Pathology Stimulation
Dominant
activation Ref
Derbyshire 2002 PET Back pain Heat pain/R hand B (120)
Albuquerque 2006 fMRI Burning mouth Heat hyperalgesia/R face B (127)
Becerra 2006 fMRI Neuropathy Cold allodynia/R face B (14)
Brush allodynia/R face B
Heat hyperalgesia/R face B
Ducreux 2006 fMRI Syringomyelia Cold allodynia/affected hand I (134)
Schweinhardt 2006 fMRI Neuropathy Brush allodynia/Varied B (135)
Witting 2006 PET Neuropathy Brush allodynia/Varied I (136)
Geha 2007 fMRI Post-herpetic Spontaneous pain/Varied
Neuralgia
(L) (137)
B, bilateral; I, ipsilateral, (L), left.
254 D. Borsook et al.

provided in a published report, 14). Pain was rated
on a visual analogue scale (VAS; see methods). The
average spontaneous pain rating was 7.7¡0.6
(mean¡SEM), and the reported prior history of
pain evoked by stimuli (e.g., touching the area,
clothing etc) was 7.2¡0.42 years. Subjects were
imaged twice, with the second session coming 2–6
months after their first imaging session. Subjects
attended a screening session where a full medical
examination, medical history, and compliance with
inclusion/exclusion criteria were performed before
enrollment into the study. Medications were dis-
continued for one dosing interval. Note that most of
the subjects were women, and all premenopausal
women were scanned during their mid-follicular
phase (days 5–7) of their menstrual cycle, since
studies suggest that the variation in pain sensitivity
across the menstrual phase is lowest at this time (33)
and more consistent with responses in men.
Experimental paradigm
The experimental paradigm for data acquisition was
similar in each group. In addition, the tools used to
apply stimuli to the face were identical, and the
timing of the application of the brush and thermal
stimuli in both groups was similar. Temperatures
applied were different for Group I (46˚C) and Group
II (threshold-based; see below).
Group I: The experimental paradigm consisted of
one MR scanning visit. Anatomical scans were
collected, followed by separate functional scans for
heat (46˚C) and brush applied to the right V2 area of
the face.
Figure 1. Top: Summary table of the two groups. Bottom: Summary of data acquisition and anatomical mapping.
Table A. Clinical data on patients with trigeminal neuropathic pain.*
Subject Age Gender Affected side Origin of pain Diagnosis Pain medications
1 54 F Right V2 Following Antibiotic Rx Idiopathic facial
Neuropathy
None
2 39 F Right V2 Car accident Post traumatic Tylenol
Facial neuropathy
3 57 F Right Vi/V2/V3 Herpes Zoster Post-herpetic Neurontin
Neuralgia
4 48 F Right V1/V2 Ski accident Post traumatic Buproprion
Facial neuropathy Setraline
Tylenol #3
5 54 M Right V2 Herpes Zoster Post-herpetic None
6 41 F Right V2/V3 Car accident Post traumatic Vicodin
Paroxetine
Clonazepam
*Adapted from our previous study (14).
255Pain and the cerebellum

Group II: The experimental paradigm comprised
of two MR scanning visits, with the second visit
occurring 2–3 months after the first. For each visit,
anatomical scans were collected, followed by func-
tional scans of sensory stimulation to the face. The
sensory stimulation included heat and brush stimuli,
presented in separate blocks. During the functional
scans, sensory stimulation was applied to either the
maxillary division of the trigeminal nerve of the pain-
affected facial location (V2A), or the mirror location
on the unaffected contralateral side (V2U).
Stimuli.
Mechanical stimulation. Brush stimuli, applied by a
Velcro-topped (soft side) stick, were administered at
1–2 Hz (1–2 strokes per second) within the V2 area.
Brush stimuli were applied to the face using a lever
system attached to a wooden frame overlying the MRI
headcoil. Plastic/nylon materials allowed us to brush
the face while standing at the feet of the subject. Thus,
there was no manual reaching into the head coil, and
all movement occurred inferior to the location of the
brain. In Group I, stimuli were applied 3 times, each
for a period of 16 sec separated by 30 sec of no
stimulus. In Group II, stimuli were also applied 3
times, each for 25 sec and 30 sec of rest.
Thermal stimulation. An FDA approved Thermal
Sensory Analyzer (TSA, MEDOC, Haifa, Israel)
was used to deliver heat stimuli through a probe that
has been adapted to rest on the face. The peltier
probe is 1.661.6 cm, or about half the size of the
thumb. Temperatures applied to the V2 area were
different for Group I (46˚C) and II (V2A pain
threshold +1˚C). For both groups, the thermode was
heated at a rate of 4˚C/sec to the target temperature,
which was maintained for 15 sec. The temperature
returned to the baseline at a rate of 4˚C/sec to end
the stimulus event. The inter-stimulus interval was
30 sec, with 3 total stimulus events. For each group,
thermal stimuli were applied using the same timing
as described for the brush. The approach for thermal
stimulation of the face in the magnet has been used
by our group (14,31,32,34) and also recently by
others (35).
Stimulation of the face in the scanner. Prior to stimuli
administration, the right V2 areas of the face to be
stimulated (Group I applied to normal skin; Group
II applied to painful/neuropathic skin) were marked
with a water-soluble pen. A specially designed
module that allowed for placement of thermal
probes and the ability to apply brush stimuli to the
specific regions was used. While in the scanner, the
subjects were told to rate the intensity of stimulus-
evoked pain using an online visual analog scale
(VAS) from ‘No pain’ to ‘Max pain’, a method we
have previously described (36).
Ratings in the scanner. For online ratings of the VAS,
subjects used a dial that could be easily rotated
between index finger and thumb. An onscreen bar was
presented which had ‘no pain’ at one end and ‘max
pain’ at the other. By rotating the dial, a cursor could
be moved along the bar. During a stimulus, subjects
used their left hand to rate their pain. Thus, subjects
rated pain using their hand opposite to the side of the
face that was stimulated, so as to potentially segregate
activity due to sensory vs. motor events.
Imaging parameters
Imaging was carried out in a 3.0 T Siemens Trio
scanner. For anatomical localization, an MPRAGE
was used (16161.6 mm resolution). For Group I,
each functional scan consisted of 33 slices (3.5 mm
isotropic resolution) coronally oriented to match the
brainstem axis and covering the middle region of the
cerebrum. Functional images were acquired in Group
I using Gradient Echo (GE) echo planar imaging
(EPI) sequence with TR/TE52.5s/30 ms, for a total
of 74 volumes per functional scan. For Group II,
functional scans were acquired using a GE EPI
sequence with isotropic resolution of 3.5 mm, 41
slices (no-gaps) were prescribed obliquely perpendi-
cular to the AC-PC axis, a TR/TE52.5 s/30 ms was
used and 128 volumes were acquired per functional
scan. For both groups field map images were acquired
with the same prescription as for the functional
images. These images were then used to correct for
susceptibility-induced distortions in functional scans.
Note that in the case of healthy subjects the whole
cerebellum was scanned, but for the patients approxi-
mately 2/3 of the cerebellum was scanned, with the
posterior component not included (Figure 2). Due to
a limitation of the phase array coil to acquire more
slices (i.e., 41 instead of 33) within the same TR,
approximately 2/3 of the patients (Group II) cerebel-
lum was scanned wit the posterior component not
included (14). Group I had enough coverage with 41
slices to scan the whole cerebellum.
Data analysis
The image analysis package fsl 3.2 (FMRIB,
University of Oxford, UK; www.fmrib.ox.ac.uk/fsl)
and in-house programs implemented via MATLAB
(Release 7.2, Mathworks Inc., Natick, MA, USA)
were used for data processing. For both groups, head
motion correction, spatial smoothing (Group I:
6 mm Gaussian kernal; Group II: 5 mm), and pre-
whitening was performed using fsl tools. For Group
I, a high-pass filter of 75 sec was also used. The
general linear model (GLM) was utilized (fsl) for
individual subject statistical analysis. For Groups I
and II a contrast explanatory variable was con-
structed comparing periods of stimulation with
resting periods based on the temperature or brush
256 D. Borsook et al.

stroking temporal profiles (14, Moulton et al., in
preparation). For Group I, a mixed-effect analysis
was performed using fsl after transformation of
individual results to a standard brain (MNI
Standard Brain). For Group II, we combined all
subjects and both visits to report aggregate results
using a fixed effects approach. For both groups,
thresholds were determined from a partition of the
statistical map distribution utilizing a gaussian
mixture model approach (37). This approach
determines statistical thresholds from a partition of
the statistical map distribution by deconstructing the
overall statistical map into several optimized
Gaussian distributions. The group activation map
(zstat image) obtained from standard statistical
analysis is further classified using a generalized
mixture model approach (37). The classification
scheme produces a series of classes (active, deactive,
belonging to the null distribution). Each voxel has a
probability to be in a particular class. Each class (or
the others) has a corresponding map in which each
voxel depicts the probability that it belongs to its
class. Thresholding the activation class map with
pw0.5 (standard in classification schemes) allow us
Figure 2. Masks defined from functional scans used for cerebellar activation for Group I – Healthy Controls (Top) and Group II – Patients
(Bottom). MNI coordinates were adjusted to match the cerebellar atlas (15) – see text.
257Pain and the cerebellum

to determine a mask for which voxels can be
considered active. The mask is then applied to the
original zstat image.
In order to align fMRI images with the atlas, fMRI
images were transformed. The transformation con-
sisted of flipping the x-coordinate (atlas is in
neurological convention while fMRI images are in
radiological convention). Furthermore, fMRI data
was rendered over a 26262 mm3 template while
the atlas was on a 16161 mm3 atlas. The most
superior and inferior axial MRI images of the
cerebellum were matched with the atlas and labeled
accordingly in Figures 2–6 with the actual coordi-
nates in MNI space.
Anatomical evaluation of cerebellar activations
To measure regions activated in the cerebellum, we
first evaluated zmax and the volume of activations.
These measures were mapped using an automated
approach to identify regions activated (38). Once
zmax and volumes of activation were recorded, co-
ordinates of zmax were transformed onto the cere-
bellum atlas (15). This was necessary because the
MNI standard does not correspond precisely to the
Talairach-based cerebellum atlas. Cerebellar activa-
tions were then determined using the following
approach: activations were grouped into the following
domains: Vermis, Hemisphere, and Cerebellar nuclei
(dentate, eboliform, globose, and fastigial) as defined
in the Atlas (15). Vermal and hemisphere activations
were defined in the horizontal plane by examination of
the identical slice with the Atlas, which displays
specific coordinates in mm (Figure 3). To identify
activations of the nuclei, significant activations were
evaluated relative to the Atlas in the horizontal,
coronal, and sagittal planes (Figure A).
In this report, we use ‘cluster’ to refer to a group
of activated voxels that by statistical and spatial
extension can be considered as a single focus of
activation. A cluster is recorded in terms of peak
activation coordinates and its volume. If a cluster’s
Figure 3. Cerebellar vermal and hemisphere activations. Top: Serial sections with number indicating z axis as taken from the Atlas of
Schmahmann et al. Bottom: Mapping activations using MRI Atlas of Human Cerebellum (15). In this case the figure shows horizontal slice
251 from the atlas and the funtional activations mapped onto the same horizontal anatomical section acquired in the study. Note that the
numbers for the functional maps represent n-1 to match the cerebellar atlas cordinates. See text.
258 D. Borsook et al.

volume of activation encompassed any of the regions
within the cerebellum (i.e., lobule, vermis, or
nucleus), that region was considered active within
that cluster. We also include cerebellar structures
encompassed within the extension of each cluster in
a specific region (lobule, vermis or nucleus).
Common activations for healthy vs. neuropathic
The main focus of this report is to localize specific
activation in the cerebellum to thermal and mechanical
stimuli in these two groups, performed in separate
studies. Although not an optimal comparison due to
confounding issues (viz., un-matched controls for age,
gender, pain level, disease state, and medications) we
compared thermal and mechanical stimuli for the
‘neuropathic state’ vs. the ‘healthy state’. To identify
common activations for brush and for heat for Group I
and Group II, overlapping maps were created.
Since subjects rated pain intensity using the hand
opposite to the side of the face that was stimulated,
activity in the cerebellum may also be observed due
Figure 4. Activations for heat and brush – Group I. Circles mark activation within nuclei. MNI coordinates were adjusted to match the
cerebellar atlas (15) – see text.
259Pain and the cerebellum

to the motor input. The left hand was used to rate
both brush and heat. In order to evaluate the
potential contribution of the motor task, we eval-
uated overlapping areas of activation in the cerebel-
lum in each group separately.
Results
The results are presented as activations for heat and
brush in healthy subjects and in patients with
neuropathic pain (sensitized state; see 14).
Group I: Healthy subjects
Psychophysical ratings. During the functional scan,
heat to V2 produced significantly greater pain
intensity ratings than the near zero values
recorded for brush (paired-t-test t(11)53.72,
pv0.01). The mean (¡SE) pain intensity
ratings for brush were 0.04¡0.04, while the
ratings for heat were 2.59¡0.69. Only one subject
reported pain with brushing, with his average rating
being 0.5.
Figure 5. Activations for heat – Group II. MNI coordinates were adjusted to match the cerebellar atlas (15) – see text.
260 D. Borsook et al.

Activation by heat and brush. Figure 4 shows serial
sections (2 mm thick) through the cerebellum
matching to the horizontal (z) axis of the cerebellar
atlas (15) showing activation following heat
(Figure 4 – top) or brush (Figure 4 – bottom)
stimulation. Based on the data, tables of activation
clusters (for max voxel, volume, coordinates for each
max volume) were created. Table IIA shows
activations following noxious heat. A total of 6
clusters met the criteria for significance (see
Methods). For increased signal, 2 clusters
contralateral to the stimulus and 2 clusters
ipsilateral were significant. For the larger volumes
activated (cerebellum_6_R545.92 cm3 or
cerebellum_6_L51.30 cm3) more than one
cerebellar region was observed within each cluster.
Two clusters showed significantly decreased
activation (Table IIB). For both increased and
decreased signal, activation was found in the
cerebellar hemispheres (n58) or dentate nucleus
(n52). A total of 5 right-sided (5 increased)
activations and 5 left-sided activations (3
Figure 6. Commonly activated areas for heat and brush for healthy subjects and neuropathic patients. MNI coordinates were adjusted to
match the cerebellar atlas (15) – see text.
261Pain and the cerebellum

Figure A. Method for evaluating cerebellar nuclei activations. Top: The location of the cerebellar nuclei within the x, y and z planes
(coordinates with atlas limits) defined by the Atlas (Schmahmann et al., 2004). D, dentate nucleus, E, emboliform nucleus; F, fastigial
nucleus; and G, globose nucleus. Bottom: Two examples of evaluating activation in nuclei by examination of activation in the x,y and z
planes. In both examples the activation maps are mapped onto the MRI and anatomical sections from the Atlas of Schmahmann et al. (15).
262 D. Borsook et al.
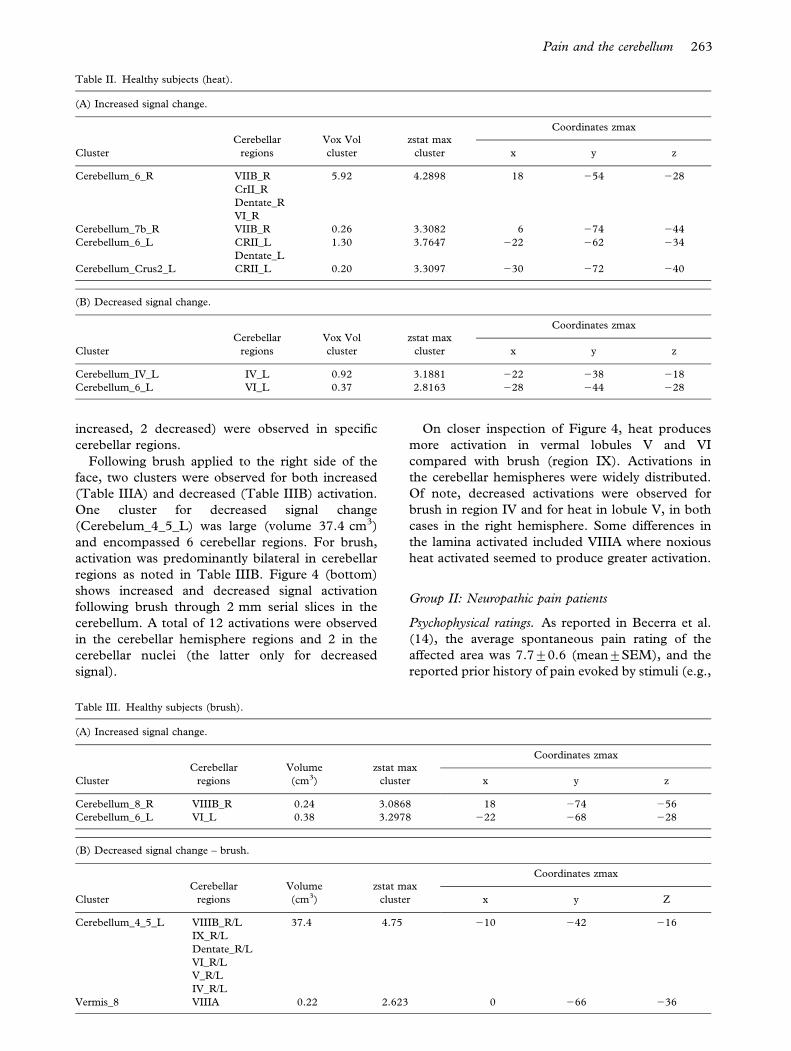
increased, 2 decreased) were observed in specific
cerebellar regions.
Following brush applied to the right side of the
face, two clusters were observed for both increased
(Table IIIA) and decreased (Table IIIB) activation.
One cluster for decreased signal change
(Cerebelum_4_5_L) was large (volume 37.4 cm3)
and encompassed 6 cerebellar regions. For brush,
activation was predominantly bilateral in cerebellar
regions as noted in Table IIIB. Figure 4 (bottom)
shows increased and decreased signal activation
following brush through 2 mm serial slices in the
cerebellum. A total of 12 activations were observed
in the cerebellar hemisphere regions and 2 in the
cerebellar nuclei (the latter only for decreased
signal).
On closer inspection of Figure 4, heat produces
more activation in vermal lobules V and VI
compared with brush (region IX). Activations in
the cerebellar hemispheres were widely distributed.
Of note, decreased activations were observed for
brush in region IV and for heat in lobule V, in both
cases in the right hemisphere. Some differences in
the lamina activated included VIIIA where noxious
heat activated seemed to produce greater activation.
Group II: Neuropathic pain patients
Psychophysical ratings. As reported in Becerra et al.
(14), the average spontaneous pain rating of the
affected area was 7.7¡0.6 (mean¡SEM), and the
reported prior history of pain evoked by stimuli (e.g.,
Table II. Healthy subjects (heat).
(A) Increased signal change.
Cluster
Cerebellar
regions
Vox Vol
cluster
zstat max
cluster
Coordinates zmax
x y z
Cerebellum_6_R VIIB_R 5.92 4.2898 18 254 228
CrII_R
Dentate_R
VI_R
Cerebellum_7b_R VIIB_R 0.26 3.3082 6 274 244
Cerebellum_6_L CRII_L 1.30 3.7647 222 262 234
Dentate_L
Cerebellum_Crus2_L CRII_L 0.20 3.3097 230 272 240
(B) Decreased signal change.
Cluster
Cerebellar
regions
Vox Vol
cluster
zstat max
cluster
Coordinates zmax
x y z
Cerebellum_IV_L IV_L 0.92 3.1881 222 238 218
Cerebellum_6_L VI_L 0.37 2.8163 228 244 228
Table III. Healthy subjects (brush).
(A) Increased signal change.
Cluster
Cerebellar
regions
Volume
(cm3)
zstat max
cluster
Coordinates zmax
x y z
Cerebellum_8_R VIIIB_R 0.24 3.0868 18 274 256
Cerebellum_6_L VI_L 0.38 3.2978 222 268 228
(B) Decreased signal change – brush.
Cluster
Cerebellar
regions
Volume
(cm3)
zstat max
cluster
Coordinates zmax
x y Z
Cerebellum_4_5_L VIIIB_R/L 37.4 4.75 210 242 216
IX_R/L
Dentate_R/L
VI_R/L
V_R/L
IV_R/L
Vermis_8 VIIIA 0.22 2.623 0 266 236
263Pain and the cerebellum
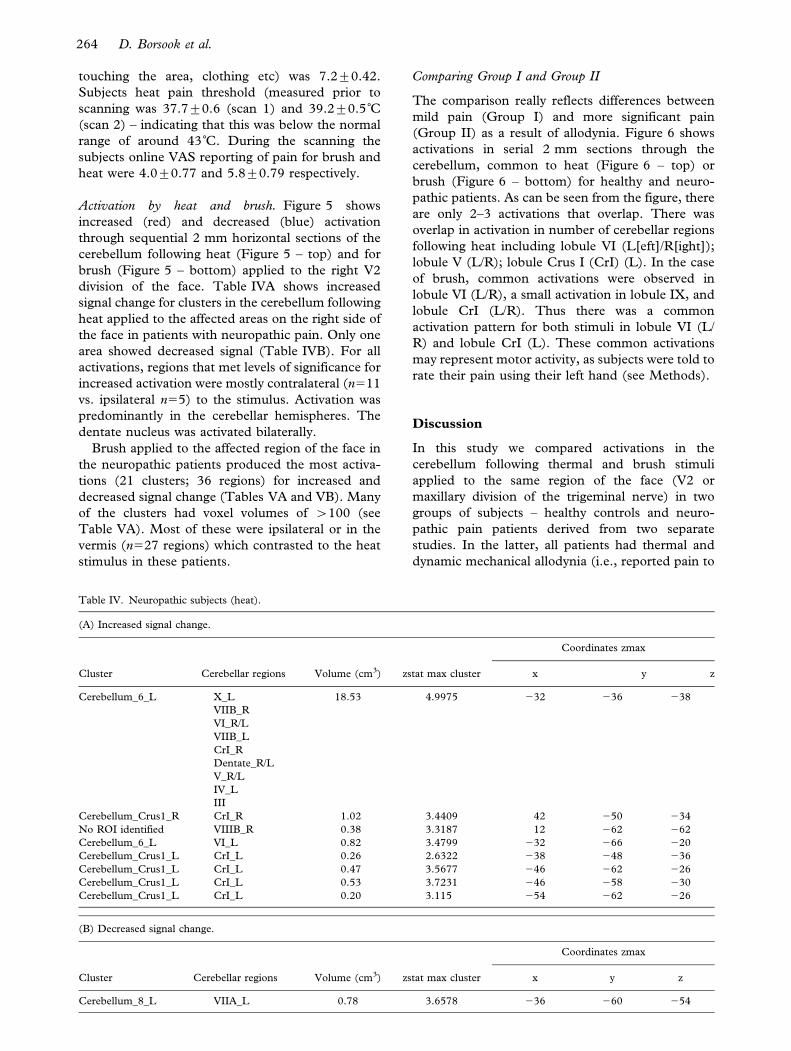
touching the area, clothing etc) was 7.2¡0.42.
Subjects heat pain threshold (measured prior to
scanning was 37.7¡0.6 (scan 1) and 39.2¡0.5˚C(scan 2) – indicating that this was below the normal
range of around 43˚C. During the scanning the
subjects online VAS reporting of pain for brush and
heat were 4.0¡0.77 and 5.8¡0.79 respectively.
Activation by heat and brush. Figure 5 shows
increased (red) and decreased (blue) activation
through sequential 2 mm horizontal sections of the
cerebellum following heat (Figure 5 – top) and for
brush (Figure 5 – bottom) applied to the right V2
division of the face. Table IVA shows increased
signal change for clusters in the cerebellum following
heat applied to the affected areas on the right side of
the face in patients with neuropathic pain. Only one
area showed decreased signal (Table IVB). For all
activations, regions that met levels of significance for
increased activation were mostly contralateral (n511
vs. ipsilateral n55) to the stimulus. Activation was
predominantly in the cerebellar hemispheres. The
dentate nucleus was activated bilaterally.
Brush applied to the affected region of the face in
the neuropathic patients produced the most activa-
tions (21 clusters; 36 regions) for increased and
decreased signal change (Tables VA and VB). Many
of the clusters had voxel volumes of w100 (see
Table VA). Most of these were ipsilateral or in the
vermis (n527 regions) which contrasted to the heat
stimulus in these patients.
Comparing Group I and Group II
The comparison really reflects differences between
mild pain (Group I) and more significant pain
(Group II) as a result of allodynia. Figure 6 shows
activations in serial 2 mm sections through the
cerebellum, common to heat (Figure 6 – top) or
brush (Figure 6 – bottom) for healthy and neuro-
pathic patients. As can be seen from the figure, there
are only 2–3 activations that overlap. There was
overlap in activation in number of cerebellar regions
following heat including lobule VI (L[eft]/R[ight]);
lobule V (L/R); lobule Crus I (CrI) (L). In the case
of brush, common activations were observed in
lobule VI (L/R), a small activation in lobule IX, and
lobule CrI (L/R). Thus there was a common
activation pattern for both stimuli in lobule VI (L/
R) and lobule CrI (L). These common activations
may represent motor activity, as subjects were told to
rate their pain using their left hand (see Methods).
Discussion
In this study we compared activations in the
cerebellum following thermal and brush stimuli
applied to the same region of the face (V2 or
maxillary division of the trigeminal nerve) in two
groups of subjects – healthy controls and neuro-
pathic pain patients derived from two separate
studies. In the latter, all patients had thermal and
dynamic mechanical allodynia (i.e., reported pain to
Table IV. Neuropathic subjects (heat).
(A) Increased signal change.
Cluster Cerebellar regions Volume (cm3) zstat max cluster
Coordinates zmax
x y z
Cerebellum_6_L X_L 18.53 4.9975 232 236 238
VIIB_R
VI_R/L
VIIB_L
CrI_R
Dentate_R/L
V_R/L
IV_L
III
Cerebellum_Crus1_R CrI_R 1.02 3.4409 42 250 234
No ROI identified VIIIB_R 0.38 3.3187 12 262 262
Cerebellum_6_L VI_L 0.82 3.4799 232 266 220
Cerebellum_Crus1_L CrI_L 0.26 2.6322 238 248 236
Cerebellum_Crus1_L CrI_L 0.47 3.5677 246 262 226
Cerebellum_Crus1_L CrI_L 0.53 3.7231 246 258 230
Cerebellum_Crus1_L CrI_L 0.20 3.115 254 262 226
(B) Decreased signal change.
Cluster Cerebellar regions Volume (cm3) zstat max cluster
Coordinates zmax
x y z
Cerebellum_8_L VIIA_L 0.78 3.6578 236 260 254
264 D. Borsook et al.

normally non-noxious thermal and mechanical
(brush) stimuli). Both stimuli produced more pain
in the patients than heat and brush in healthy
subjects. Overall, heat and brush produced relatively
few activations in healthy subjects while a larger
number of activations were observed in patients. In
the latter group, with brush-induced pain (allodynia)
produced more activation than heat. In patients,
heat and brush had mirrored asymmetrical activa-
tion patterns: noxious heat produced a predomi-
nantly contralateral activation in the patient (and
healthy) group, while brush produced a greater
number of activations in the ipsilateral cerebellar
hemispheres (more so in patients than in healthy
subjects). A number of studies have reported
cerebellar activation following noxious stimuli and
these are summarized in Table I with a summary of
reports of cerebellar activation in acute experimental
pain (Table IA), cognitive influences on cerebellar
activation (Table IB) and in neuropathic pain
(Table IC). Note that most of the activations from
noxious stimuli were bilateral and or ipsilateral. Our
results show that cerebellar activation is observed in
distinct regions in response to brush and heat in two
different groups as discussed below.
Heat and brush activation in healthy subjects: Cognitive
vs. sensory processing
Our data show that heat produced activation in areas
thought to be involved in cognitive processing (39)
(i.e., lobules CrII and VIIB) and in sensory-cognitive
Table V. Neuropathic subjects (brush).
(A) Increased signal change.
Cluster Cerebellar Regions Volume (cm3) zstat max Cluster
Coordinates zmax
x y z
Cerebellum_6_R VIIB_R 9.67 5.8835 32 262 230
CrII_R
CrI_R
VI_R
Cerebellum_6_R VI_R 0.28 3.6028 38 242 232
Cerebellum_8_R VIIIA_R 0.59 3.801 16 266 256
Cerebellum_4_5_R V_R 0.22 3.3338 8 256 218
Vermis_7 VIIB 0.65 3.6009 6 268 226
Vermis_9 IX 0.37 3.6141 4 260 242
Cerebellum_9_R IX_R 0.30 3.6149 4 256 248
Vermis_6 VI 0.74 4.2462 2 268 212
Vermis_8 VIIIA 0.38 3.9221 2 262 236
Vermis_4_5 IV/V 0.53 4.0999 0 254 212
Vermis_4_5 III 0.42 4.1865 0 246 26
Vermis_8 VIIIA 1.01 3.9685 22 264 234
Cerebellum_4_5_L V_L 1.46 4.3947 24 260 216
No ROI identified Dentate_L 1.77 4.8849 216 256 228
No ROI identified White matter_L 0.61 4.4094 222 254 240
Dentate_L
No ROI identified Dentate_L 1.63 5.1305 222 254 232
Cerebellum_8_L VIIIA_L 0.89 4.2204 224 240 246
No ROI identified CrII_L 1.35 4.3673 230 252 240
CrI_L
Cerebellum_6_L VI_L 0.22 3.0393 234 246 232
(B) Decreased signal change.
Cluster Cerebellar Regions Volume (cm3) zstat max Cluster
Coordinates zmax
x y z
No ROI identified V_R 0.48 0.93465 26 232 234
No ROI identified IV_R 0.26 22.4171 12 238 232
No ROI identified VIIIB_R 41.30 0.97867 212 232 228
IX_R/L
VIIIA_R/L
Dentate_R/L
CrI_R/L
IX
VI_R/L
V_R/L
IV_R/L
III
265Pain and the cerebellum

processing (VI). However, in healthy subjects brush
stimuli produced predominantly decreased signal in
areas involved in sensory-motor integration (lobules
IV, V and VI), and in secondary sensory processing
(lobule VIIIB). Thus, there is a differentiation of
increased cognitive cerebellar regional activity for a
low heat stimulus vs. a predominant decrease in the
sensory processing component for brush. We infer
from these results that there is a distinct difference in
cerebellar mechanisms integrating information for
mild thermal heat vs. brush, the latter producing
activation almost completely restricted to cerebellar
sensory processing.
Heat and brush activation in patients: Cognitive vs.
sensory processing
In patients, both heat and brush produces pain (see
Figure 1) when applied to the area of the face
affected by nerve damage resulting in the clinical
condition of neuropathic pain. Heat pain produced
activation in regions involved in sensory-motor
processing (anterior lobe, lobules III–V, and lobules
VI and VIIIA), but also included areas involved in
cognitive processing (lobule CRI). The activation
pattern was not very different from heat in healthy
subjects. For brush-induced allodynia, activation in
the cerebellum was observed in sensorimotor regions
in lobules III to V, putative secondary somatosen-
sory regions (lobule VIII; (40)), vestibular regions
(lobule IX), cognitive regions (lobules VIIB, CRI,
CRII), and prominent dentate nucleus activation.
The distribution of sensory and cognitive activation
following brush may result from the nature of the
sensation: pain, dysesthesia, differential input by the
same fiber type (Aß fibers) that normally conveys the
sensation of mechanical stimuli such as brush. Thus,
there might be additive inputs from the trigeminal
nuclei that connect with various brain regions
including the cerebellum in response to mechanical
stimuli as well as painful stimuli. In the current
study, there is nothing disrupting cerebellar function
(i.e., no injury), rather, cognitive cerebellar regions
are active in the cognitive state. Thus, the cerebel-
lum appears to be active in its sensorimotor as well
as its cognitive and limbic regions for situations in
which a subject’s experience of a stimulus is painful.
Trigemino-cerebellar pathways
Possible pathways mediating noxious (thermal and
mechanical) and innocuous (mechanical) informa-
tion to the cerebellum have been described using
electorphysiological and anatomical tracing techni-
ques in mammals and birds (21,25,41–46).
Relatively few trigeminal afferents project to the
cerebellar nuclei (47) and most project to the
cerebellar cortex. Trigeminal afferents are reported
to arise in the the nucleus interpolaris,nucleus oralis
and principal nucleus, and project ipsilaterally to the
three cerebellar cortical regions – the lobulus
simplex and part of lobule V, rostral folia of the
paramedian lobule with surrounding parts of crus I
and II, and lobule IX (48). Some projections are
observed from the mesencepahlic nucleus and
nucleus caudalis directed to vermal regions. In a
study of trigeminocerebellar projections in sheep,
secondary trigeminocerebellar connections have
been described in some detail using tract tracing
techniques (49). The results indicate most of the
cerebellar cortex receives bilateral (but mostly
ipsilateral) fibers from the trigeminal nuclei (except
flocculus, ventral paraflocculus and lobules I–IV)
with some topographic organization (mesencephalic
nucleus to the anterior lobe, lobules VI, VIII and
dorsal paraflocculus; principal nucleus to all lobules
in vermis and hemispheres). These projections have
not been described in man. Our results may be
interpreted in the light of these studies. For noxious
heat, the primary inputs are to the trigeminal
nucleus caudalis and interpolaris (50–52), and based
on the rat data, projections would be to vermal
regions. Non-noxious brush stimuli are transmitted
via the main or principal sensory nucleus and have
collateral projections to the more caudal nuclei (53–
57). Inputs to the trigeminal nucleus are complex
(see (31) for a review) and given patterns we observe
(e.g., heat pain activating a disproportionate number
of Crus I and Crus II regions and painful brush
activating a large number of vermal regions), it is
difficult to extrapolate from the animal studies. In
addition, many indirect pathways, for example, the
trigemino-olivary-cerebellar projection (46) may also
contribute. MRI tract-tracing studies may help
delineate these pathways in humans (58,59).
Pain and the cerebellum
Although there are numerous pain studies that
report activation in the cerebellum, only one group
has reported activation in specific cerebellar regions
(13,60). In these two prior studies, the authors
employed a similar method for producing heat, but
applied heat to the hand in healthy subjects.
Activations were reported in the anterior vermis
(lobules II–V), and bilaterally in the hemispheres
(lobules III–VI). These activations were observed
predominantly ipsilateral to the stimulus. The
studies were performed in healthy volunteers.
Other studies have shown activation in the cerebel-
lum to temporal summation of C-fiber evoked pain,
and suggest cerebellar activation may correlate with
premotor activity (61,62). In our studies, we
observed more widespread activations within the
vermis and hemispheres for the patient group and in
the nuclei. Based on our current understanding of
input-output systems of the cerebellum the activa-
tions in the cerebellar cortex are a result of afferent
266 D. Borsook et al.

sensory inputs (63), including pain (64,65), while
activation in the nuclei relate to outputs (66).
Decreased activations were also observed in the
present study (Tables IB–IVB). If decreased activa-
tion represents inhibitory processing (67,68), we
interpret these changes as such and relate to
inhibitory systems in cerebellar cortical regions.
The cerebellum has been called ‘the great
modulator of neurologic function’ (69). It has
outputs to numerous limbic structures (including
the hippocampus, amygdala, intralaminar thalamic
nuclei, the hypothalamus, the periaqaueductal gray
(70–75) and thus may influence emotion, cognition,
and sensory function – all dimensions involved in the
response to pain or pain modulation. Having said
this, the role of the cerebellum in emotional
processing remains controversial (see 76).
The role of the cerebellum in nociception has been
reviewed by Saab and Willis (4). In preclinical
studies, cerebellar stimulation modulates thalamic
neural responses to painful stimuli (77) and also
modulates the intensity of a visceral nociceptive
reflex in rats (78). Conversely, nociceptive stimuli
modulate the activity of cerebellar Purkinje cells
(22,64,65). Pharmacological manipulation of the
cerebellum also produces analgesia. For example,
microinjection of morphine into the anterior cere-
bellum of rats produced analgesia that was reversible
by systemic naloxone (79). In addition, the same
authors reported that brief electrical stimulation of
the same area resulted in analgesia after the
stimulation ended. Lesions of the cerebellar vermis
in rats results in a larger increase in the threshold to
the reaction to electrical shock in rats compared with
lesions of the cerebellar cortex (80). Human studies
have not been able to produce controlled lesions or
stimulation for pain, although cerebellar stimulation
has been used for motor disorders (81). Pain
imaging studies across the spectrum from visceral
(82) to somatic, acute to chronic (83) all show
cerebellar activation. Few other structures are
activated in such a consistent manner. In addition
to responding to direct pain stimulation, activation is
reported across a number of pain experiments
including imagination of pain (84), modulation of
pain (85) and perception of pain in others (17),
empathy (18), and acupuncture stimulation (86).
Indeed many aspects of sensory discrimination
described above pertain to anticipation, error pre-
diction, cognitive responses, attention, and integra-
tion of the brain’s response to sensation (87).
Little is known about the specific role of the
cerebellum in nociceptive processing. Are these
systems generic to sensory processing, or is there
specific modulation relating to pain and integrating
the CNS response including autonomic, cognitive,
emotional and sensory dimensions? Although the
cerebellum has been considered to be involved in
cognitive behaviors, no clinical or preclinical studies
have yet shown a role in acute or chronic pain.
Recent work has considered many pain responses as
learned (88,89) and the role of the cerebellum is still
unclear. The possible role of the cerebellum in pain
processing may be considered in the light of several
theoretical formulations:
(i) The cerebellum may optimize performance by
modulating behavior automatically according
to context (39) such as the need to integrate
appropriate motor function. This modulation
may be applied to learned behaviors in pain
(sensory) processing, as the cerebellum has
been considered to be involved in monitoring
and adjusting sensory acquisition (90). The
cerebellum is considered to be involved in
controlling error signals, and may be a com-
parator for errors in somatosensory processing
(64,65,91,92). Such error signals may play a
role, for example, in wrongly executed move-
ment (4).
(ii) Cerebellar processing may play an important
role in response to expected vs. unexpected events.
The cerebellum may play a bigger role in
processing unexpected events than expected
events. In a recent imaging study of sponta-
neous trigeminal neuralgia (Borsook et al.,
under review), no activation was observed in
the cerebellum following evoked (expected) tics
but activation was observed following sponta-
neous (unexpected tics). This type of informa-
tion may infer a role in processing anticipatory
sensory input with a high level of temporal
accuracy, and optimize temporal responses in
the sensory and integrative systems (93,94).
For example, measures of cerebellar activity
using MEG in response to electric shocks of the
median nerve suggests that the signals are
probably elicited by the first afferent sensory
volley from peripheral nerve endings and
mediated by spinocerebellar (cuneocerebellar)
tracts (95). The results imply strong coherent
activation of cerebellar neuronal populations
after purely sensory stimulation based on
observation of changes notes measured in
millisecond scale temporal resolution.
Whether the cerebellum is directly involved in
anticipation of pain (96) or plays an integrative
role is unknown.
(iii) The cerebellum may play an integrative role
with respect to sensory information flow to and
from the somatosensory cortex. Alterations in
somatosensory cortex have been reported in
patients with cerebellar lesions (97), and
altered sensory processing has also been
reported in such patients (19). In the latter
study, the authors report that the cerebellum
may be involved in pre-attentive detection of
incoming somatosensory inputs.
267Pain and the cerebellum

Methodological limitations
A number of methodological caveats apply to this
report:
(i) Separate studies. The data for heat and brush
were from two different studies and although
many processes were the same (3T magnet,
number of stimuli, exactly the same method
used for applying the stimuli etc), there are a
number of differences. Differences in the
scanning were present for stimulation sites
and number of times subjects were scanned
(twice in Group II). However as reported in
our paper (14) we did not observe significant
differences to pain thresholds prior to scan-
ning, or differences in activation for the two
scans. Patients were scanned twice to enhance
the power or the study.
(ii) Un-matched studies. The main data reported
relate to separate activation patterns to heat
and brush in two groups – healthy and
neuropathic pain. As noted data reported here
were derived from two prior studies (Moulton
et al., in preparation; (14)) that were not
specifically designed to compare activation
between healthy and neuropathic pain sub-
jects. As a consequence, the groups were not
matched for age or gender, or ongoing back-
ground pain. While some issues can be
addressed in a proper comparison of such
data, there are a few that are always proble-
matic when using a patient group. These
include the disease state for which there are
no good surrogate models (except perhaps
capsaicin hyperalgesia, but this does not
reflect the underlying changes in neuropathic
pain including altered chemical, anatomical
and functional processes observed in neuro-
pathic pain (98–100). In addition, patients are
usually on or have been on medication for
long periods of time, and thus a true compar-
ison is very difficult. What we show in the
comparison are essentially areas of activation
that are common or different between these
groups. One way to interpret this may be the
effects of a low level input to heat and brush in
Group I and a high level input to heat and
brush in Group II. Such a group would still
have some inherent inequities e.g., analgesics.
(iii) Motor function. Another confound may relate
to the motor function by left hand movements
for rating pain that could have produced
activation within the cerebellum, since the
cerebellum is well known to be involved in
motor integration (101). The common and
similar activation sites for both heat and brush
stimuli suggest that motor activation is present
but does not encompass all of the regions
demonstrated in the two cohorts. Further, we
do not see common activation in the dentate
nuclei. In an fMRI study, activation in
cerebellar output nuclei (dentate) showed
increases only when subjects experienced
cutaneous stimulation alone but showed little
activation in nuclei with combined sensory
stimulation and coordinated movements of
the hand (see (102)). This and other imaging
studies (103,104) suggest that the cerebellum
has an important role in sensory discrimina-
tion.
(iv) Cerebellar acquisition. The cerebellum was not
fully imaged in the patient cohort, potentially
missing activation in the most posterior aspect
of the cerebellum. However, healthy subjects,
that had full coverage of the cerebellum, did
not display activation beyond y5274, the
posterior extent for the patient group, and
hence our common activation results are likely
valid.
(v) Differences in pain responses. In addition, heat
applied in the control subjects produced mild
pain whereas the pain produced by heat in the
neuropathic pain subjects was more severe. An
increase in the intensity of pain would
probably enhance cerebellar activation.
Conclusions
Cerebellar sensorimotor regions are activated by
touch, whereas cerebellar lobules thought to be
involved in cognitive and emotional processing are
activated during noxious stimulation both in healthy
subjects and in those with neuropathic pain. These
results led us to propose that the cerebellum
modulates the emotional and cognitive experience
that distinguishes the perception of pain from the
appreciation of innocuous sensory stimulation.
These results add to the weight of preclinical and
clinical imaging studies in healthy subjects and
patients that demonstrate cerebellar activation by
painful stimulation, but a clinical condition in which
there is altered pain processing as a result of
cerebellar lesions in humans has not yet been
described.
Acknowledgements
This work was supported by a grant from NINDS
(NS 042721).
References
1. Ito M. Cerebellar circuitry as a neuronal machine. Prog
Neurobiol. 2006;78:272–303.
2. Schmahmann JD. An emerging concept. The cerebellar
contribution to higher function. Arch Neurol. 1991;48:
1178–87.
268 D. Borsook et al.

3. Schmahmann JD. Rediscovery of an early concept. Int Rev
Neurobiol. 1997;41:3–27.
4. Saab CY, Willis WD. The cerebellum: organization, func-
tions and its role in nociception. Brain Res Brain Res Rev.
2003;42:85–95.
5. Chambers WW, Sprague JM. Functional localization in the
cerebellum. II. Somatotopic organization in cortex and
nuclei. AMA Arch Neurol Psychiatry. 1955;74:653–80.
6. Leiner HC, Leiner AL, Dow RS. Does the cerebellum
contribute to mental skills? Behav Neurosci. 1986;100:
443–54.
7. Schmahmann JD, Sherman JC. Cerebellar cognitive affec-
tive syndrome. Int Rev Neurobiol. 1997;41:433–40.
8. Haines DE, Dietrichs E. An HRP study of hypothalamo-
cerebellar and cerebello-hypothalamic connections in squir-
rel monkey (Saimiri sciureus). J Comp Neurol. 1984;229:
559–75.
9. Levisohn L, Cronin-Golomb A, Schmahmann JD.
Neuropsychological consequences of cerebellar tumour
resection in children: cerebellar cognitive affective syndrome
in a paediatric population. Brain. 2000;123(Pt 5):1041–50.
10. Schutter DJ, van Honk J. The cerebellum on the rise in
human emotion. Cerebellum. 2005;4:290–4.
11. Apkarian AV, Bushnell MC, Treede RD, Zubieta JK.
Human brain mechanisms of pain perception and regulation
in health and disease. Eur J Pain. 2005;9:463–84.
12. Peyron R, Laurent B, Garcia-Larrea L. Functional imaging
of brain responses to pain. A review and meta-analysis
(2000). Neurophysiol Clin. 2000;30:263–88.
13. Helmchen C, Mohr C, Erdmann C, Binkofski F. Cerebellar
neural responses related to actively and passively applied
noxious thermal stimulation in human subjects: a parametric
fMRI study. Neurosci Lett. 2004;361:237–40.
14. Becerra L, Morris S, Bazes S, Gostic R, Sherman S, Gostic J,
et al. Trigeminal neuropathic pain alters responses in CNS
circuits to mechanical (brush) and thermal (cold and heat)
stimuli. J Neurosci. 2006;26:10646–57.
15. Schmahmann JD, Doyon J, Toga AW, Petrides M,
Evans AC. MRI atlas of the human cerebellum. San
Diego: Academic Press, 2000.
16. Landgrebe M, Langguth B, Barta W, Hajak G,
Eichhammer P. [Modulation of cold-/warm-sensation by
cerebellar repetitive transcranial magnetic stimulation
(rTMS).]. Psychiatr Prax. 2007;34(1 Suppl.):10–12.
17. Jackson PL, Meltzoff AN, Decety J. How do we perceive the
pain of others? A window into the neural processes involved
in empathy. Neuroimage. 2005;24:771–9.
18. Singer T, Seymour B, O’Doherty J, Kaube H, Dolan RJ,
Frith CD. Empathy for pain involves the affective but not
sensory components of pain. Science. 2004;303(5661):
1157–62.
19. Restuccia D, Marca GD, Valeriani M, Leggio MG,
Molinari M. Cerebellar damage impairs detection of somato-
sensory input changes. A somatosensory mismatch-negativity
study. Brain. 2007;130(Pt 1):276–87.
20. Hartmann MJ, Bower JM. Tactile responses in the granule
cell layer of cerebellar folium crus IIa of freely behaving rats.
J Neurosci. 2001;21:3549–63.
21. Holtzman T, Rajapaksa T, Mostofi A, Edgley SA. Different
responses of rat cerebellar Purkinje cells and Golgi cells
evoked by widespread convergent sensory inputs. J Physiol.
2006;574(Pt 2):491–507.
22. Saab CY, Willis WD. Nociceptive visceral stimulation
modulates the activity of cerebellar Purkinje cells. Exp
Brain Res. 2001;140:122–6.
23. Bukowska D, Mierzejewska-Krzyzowska B, Zguczynski L.
Topography and axonal collaterals of trigeminocerebellar
projection to the paramedian lobule and uvula in the
rabbit cerebellum. Acta Neurobiol Exp (Wars). 2006;66:
145–51.
24. Patrick GW, Robinson MA. Collateral projections from
trigeminal sensory nuclei to ventrobasal thalamus and
cerebellar cortex in rats. J Morphol. 1987;192:229–36.
25. Somana R, Kotchabhakdi N, Walberg F. Cerebellar
afferents from the trigeminal sensory nuclei in the cat. Exp
Brain Res. 1980;38:57–64.
26. Steindler DA. Trigeminocerebellar, trigeminotectal, and
trigeminothalamic projections: a double retrograde axonal
tracing study in the mouse. J Comp Neurol. 1985;237:
155–75.
27. Shumway CA, Morissette J, Gruen P, Bower JM. Plasticity
in cerebellar tactile maps in the adult rat. J Comp Neurol.
1999;413:583–92.
28. Jie W, Pei-Xi C. Discharge response of cerebellar Purkinje
cells to stimulation of C-fiber in cat saphenous nerve. Brain
Res. 1992;581:269–72.
29. Heekeren HR, Marrett S, Ruff DA, Bandettini PA,
Ungerleider LG. Involvement of human left dorsolateral
prefrontal cortex in perceptual decision making is indepen-
dent of response modality. Proc Natl Acad Sci USA.
2006;103:10023–8.
30. Molinari M, Leggio MG, Thaut MH. The cerebellum and
neural networks for rhythmic sensorimotor synchronization
in the human brain. Cerebellum. 2007;6:18–23.
31. Borsook D, Burstein R, Becerra L. Functional imaging of
the human trigeminal system: opportunities for new insights
into pain processing in health and disease. J Neurobiol.
2004;61:107–25.
32. Moulton E, Pendse G, Morris S, Strassman A, Aiello-
Lammens M, Becerra L, et al. Capsaicin-induced thermal
hyperalgesia and sensitization in the human trigeminal
nociceptive pathway: an fMRI study. NeuroImage. 2007;
35(4):1586–600.
33. Choi JC, Park SK, Kim YH, Shin YW, Kwon JS, Kim JS,
et al. Different brain activation patterns to pain and pain-
related unpleasantness during the menstrual cycle.
Anesthesiology. 2006;105:120–7.
34. DaSilva AF, Becerra L, Makris N, Strassman AM,
Gonzalez RG, Geatrakis N, et al. Somatotopic activation
in the human trigeminal pain pathway. J Neurosci.
2002;22:8183–92.
35. de Leeuw R, Davis CE, Albuquerque R, Carlson CR,
Andersen AH. Brain activity during stimulation of the
trigeminal nerve with noxious heat. Oral Surg Oral Med
Oral Pathol Oral Radiol Endod. 2006;102:750–7.
36. Borras MC, Becerra L, Ploghaus A, Gostic JM, DaSilva A,
Gonzalez RG, et al. fMRI measurement of CNS responses
to naloxone infusion and subsequent mild noxious thermal
stimuli in healthy volunteers. J Neurophysiol. 2004;91:
2723–33.
37. Pendse G, Borsook D, Becerra L. A generalized mixture
modeling approach applied to the problem of thresholding
fMRI statistical maps. Society for Neuroscience. Atlanta,
GA; 2006.
38. Maldjian JA, Laurienti PJ, Kraft RA, Burdette JH. An
automated method for neuroanatomic and cytoarchitectonic
atlas-based interrogation of fMRI data sets. Neuroimage.
2003;19:1233–9.
39. Schmahmann JD. Disorders of the cerebellum: ataxia,
dysmetria of thought, and the cerebellar cognitive affective
syndrome. J Neuropsychiatry Clin Neurosci. 2004;16:
367–78.
40. Woolsey T. Summary of papers on the cerebellum. In:
Philip B, editor. Patterns of organization in the central
nervous system, 1952. pp 334–6.
41. Arends JJ, Zeigler HP. Cerebellar connections of the
trigeminal system in the pigeon (Columba livia). Brain Res.
1989;487:69–78.
42. Bukowska D. Trigeminocerebellar projection to the para-
median lobule with emphasis on the climbing fibre zones: a
269Pain and the cerebellum

retrograde tracing study in the rabbit. J Hirnforsch.
1996;37:159–72.
43. Elias SA, Taylor A, Somjen G. Direct and relayed projection
of periodontal receptor afferents to the cerebellum in the
ferret. Proc R Soc Lond B Biol Sci. 1987;231(1263):
199–216.
44. Falls WM. Direct connections of primary trigeminal afferent
axons with trigeminocerebellar projection neurons in the
border zone of rat trigeminal nucleus oralis. Neurosci Lett.
1987;83:247–52.
45. Hayashi H, Sumino R, Sessle BJ. Functional organization of
trigeminal subnucleus interpolaris: nociceptive and innoc-
uous afferent inputs, projections to thalamus, cerebellum,
and spinal cord, and descending modulation from periaque-
ductal gray. J Neurophysiol. 1984;51:890–905.
46. Yatim N, Billig I, Compoint C, Buisseret P, Buisseret-
Delmas C. Trigeminocerebellar and trigemino-olivary pro-
jections in rats. Neurosci Res. 1996;25:267–83.
47. Dietrichs E, Walberg F. Cerebellar nuclear afferents – where
do they originate? A re-evaluation of the projections from
some lower brain stem nuclei. Anat Embryol (Berl).
1987;177:165–72.
48. Jacquin MF, Semba K, Rhoades RW, Egger MD.
Trigeminal primary afferents project bilaterally to dorsal
horn and ipsilaterally to cerebellum, reticular formation, and
cuneate, solitary, supratrigeminal and vagal nuclei. Brain
Res. 1982;246:285–91.
49. Saigal RP, Karamanlidis AN, Voogd J, Mangana O,
Michaloudi H. Secondary trigeminocerebellar projections
in sheep studied with the horseradish peroxidase tracing
method. J Comp Neurol. 1980;189:537–53.
50. Amano N, Hu JW, Sessle BJ. Responses of neurons in feline
trigeminal subnucleus caudalis (medullary dorsal horn) to
cutaneous, intraoral, and muscle afferent stimuli. J
Neurophysiol. 1986;55:227–43.
51. Sessle BJ, Greenwood LF. Inputs to trigeminal brain stem
neurones from facial, oral, tooth pulp and pharyngolaryngeal
tissues: I. Responses to innocuous and noxious stimuli.
Brain Res. 1976;117:211–26.
52. Young RF, Perryman KM. Pathways for orofacial
pain sensation in the trigeminal brain-stem nuclear
complex of the Macaque monkey. J Neurosurg. 1984;61:
563–8.
53. Jacquin MF, Chiaia NL, Klein BG, Rhoades RW. Structure-
function relationships in the rat brainstem subnucleus
interpolaris: VI. Cervical convergence in cells deafferented
at birth and a potential primary afferent substrate. J Comp
Neurol. 1989;283:513–25.
54. Jacquin MF, Mooney RD, Rhoades RW. Morphology,
response properties, and collateral projections of trigemi-
nothalamic neurons in brainstem subnucleus interpolaris of
rat. Exp Brain Res. 1986;61:457–68.
55. Jacquin MF, Renehan WE, Mooney RD, Rhoades RW.
Structure-function relationships in rat medullary and cervi-
cal dorsal horns. I. Trigeminal primary afferents. J
Neurophysiol. 1986;55:1153–86.
56. Jacquin MF, Rhoades RW. Cell structure and response
properties in the trigeminal subnucleus oralis. Somatosens
Mot Res. 1990;7:265–88.
57. Renehan WE, Jacquin MF, Mooney RD, Rhoades RW.
Structure-function relationships in rat medullary and cervi-
cal dorsal horns. II. Medullary dorsal horn cells. J
Neurophysiol. 1986;55:1187–201.
58. Mori S, Zhang J. Principles of diffusion tensor imaging and
its applications to basic neuroscience research. Neuron.
2006;51:527–39.
59. Yamada K, Nagakane Y, Yoshikawa K, Kizu O, Ito H,
Kubota T, et al. Somatotopic organization of thalamocor-
tical projection fibers as assessed with MR tractography.
Radiology. 2007;242:840–5.
60. Helmchen C, Rambold H, Sprenger A, Erdmann C,
Binkofski F. Cerebellar activation in opsoclonus: an fMRI
study. Neurology. 2003;61:412–5.
61. Christmann C, Koeppe C, Braus DF, Ruf M, Flor H. A
simultaneous EEG-fMRI study of painful electric stimula-
tion. Neuroimage. 2007;34:1428–37.
62. Staud R, Craggs JG, Robinson ME, Perlstein WM,
Price DD. Brain activity related to temporal summation of
C-fiber evoked pain. Pain. 2007;129(1–2):130–42.
63. Jorntell H, Ekerot CF. Properties of somatosensory synaptic
integration in cerebellar granule cells in vivo. J Neurosci.
2006;26:11786–97.
64. Ekerot CF, Gustavsson P, Oscarsson O, Schouenborg J.
Climbing fibres projecting to cat cerebellar anterior lobe
activated by cutaneous A and C fibres. J Physiol. 1987;386:
529–38.
65. Ekerot CF, Oscarsson O, Schouenborg J. Stimulation of cat
cutaneous nociceptive C fibres causing tonic and synchro-
nous activity in climbing fibres. J Physiol. 1987;386:539–46.
66. Delgado-Garcia JM. [Structure and function of the cere-
bellum]. Rev Neurol. 2001;33:635–42.
67. Harel N, Lee SP, Nagaoka T, Kim DS, Kim SG. Origin of
negative blood oxygenation level-dependent fMRI signals. J
Cereb Blood Flow Metab. 2002;22:908–17.
68. Stefanovic B, Warnking JM, Pike GB. Hemodynamic and
metabolic responses to neuronal inhibition. Neuroimage.
2004;22:771–8.
69. Snider RS. Recent contributions to the anatomy and
physiology of the cerebellum. Arch Neurol Psych, 1950.
pp 196–219.
70. Allen G, McColl R, Barnard H, Ringe WK, Fleckenstein J,
Cullum CM. Magnetic resonance imaging of cerebellar-
prefrontal and cerebellar-parietal functional connectivity.
Neuroimage. 2005;28:39–48.
71. Braak H, Braak E, Yilmazer D, Bohl J. Functional anatomy
of human hippocampal formation and related structures. J
Child Neurol. 1996;11:265–75.
72. Middleton FA, Strick PL. Cerebellar output channels. Int
Rev Neurobiol. 1997;41:61–82.
73. Onat F, Cavdar S. Cerebellar connections: hypothalamus.
Cerebellum. 2003;2:263–9.
74. Schmahmann JD, Pandya DN. The cerebrocerebellar
system. Int Rev Neurobiol. 1997;41:31–60.
75. Snider RS, Maiti A. Cerebellar contributions to the Papez
circuit. J Neurosci Res. 1976;2:133–46.
76. Manto MU. On the cerebello-cerebral interactions.
Cerebellum. 2006;5:286–8.
77. Liu FY, Qiao JT, Dafny N. Cerebellar stimulation mod-
ulates thalamic noxious-evoked responses. Brain Res Bull.
1993;30:529–34.
78. Saab CY, Willis WD. Cerebellar stimulation modulates the
intensity of a visceral nociceptive reflex in the rat. Exp Brain
Res. 2002;146:117–21.
79. Dey PK, Ray AK. Anterior cerebellum as a site for morphine
analgesia and post-stimulation analgesia. Indian J Physiol
Pharmacol. 1982;26:3–12.
80. Peters M, Bleek C, Monjan AA. Reaction to electrical shock
after cerebellar lesions in the rat. Physiol Behav.
1973;10:429–33.
81. Iwata NK, Ugawa Y. The effects of cerebellar stimulation on
the motor cortical excitability in neurological disorders: a
review. Cerebellum. 2005;4:218–23.
82. Lu CL, Wu YT, Yeh TC, Chen LF, Chang FY, Lee SD,
et al. Neuronal correlates of gastric pain induced by fundus
distension: a 3T-fMRI study. Neurogastroenterol Motil.
2004;16:575–87.
83. Gracely RH, Geisser ME, Giesecke T, Grant MA, Petzke F,
Williams DA, et al. Pain catastrophizing and neural
responses to pain among persons with fibromyalgia. Brain.
2004;127(Pt 4):835–43.
270 D. Borsook et al.

84. Ogino Y, Nemoto H, Inui K, Saito S, Kakigi R, Goto F.
Inner experience of pain: imagination of pain while viewing
images showing painful events forms subjective pain
representation in human brain. Cereb Cortex.
2007;17(5):1139–46.
85. Keltner JR, Furst A, Fan C, Redfern R, Inglis B, Fields HL.
Isolating the modulatory effect of expectation on pain
transmission: a functional magnetic resonance imaging
study. J Neurosci. 2006;26:4437–43.
86. Yoo SS, Teh EK, Blinder RA, Jolesz FA. Modulation of
cerebellar activities by acupuncture stimulation: evidence
from fMRI study. Neuroimage. 2004;22:932–40.
87. Ito M. Bases and implications of learning in the cerebellum –
adaptive control and internal model mechanism. Prog Brain
Res. 2005;148:95–109.
88. Johansen JP, Fields HL. Glutamatergic activation of anterior
cingulate cortex produces an aversive teaching signal. Nat
Neurosci. 2004;7:398–403.
89. Seymour B, O’Doherty JP, Koltzenburg M, Wiech K,
Frackowiak R, Friston K, et al. Opponent appetitive-
aversive neural processes underlie predictive learning of pain
relief. Nat Neurosci. 2005;8:1234–40.
90. Bower JM. Control of sensory data acquisition. Int Rev
Neurobiol. 1997;41:489–513.
91. Apps R, Garwicz M. Anatomical and physiological founda-
tions of cerebellar information processing. Nat Rev
Neurosci. 2005;6:297–311.
92. Eccles J, Ito M, Szentagothai J. The cerebellum as a
neuronal machine. New York: Springer-Verlag, 1967.
93. Ohyama T, Nores WL, Murphy M, Mauk MD. What the
cerebellum computes. Trends Neurosci. 2003;26:222–7.
94. Tesche CD, Karhu JJ. Anticipatory cerebellar responses
during somatosensory omission in man. Hum Brain Mapp.
2000;9:119–42.
95. Tesche CD, Karhu J. Somatosensory evoked magnetic fields
arising from sources in the human cerebellum. Brain Res.
1997;744:23–31.
96. Ploghaus A, Tracey I, Gati JS, Clare S, Menon RS,
Matthews PM, et al. Dissociating pain from its anticipation
in the human brain. Science. 1999;284(5422):1979–81.
97. Restuccia D, Valeriani M, Barba C, Le Pera D, Capecci M,
Filippini V, et al. Functional changes of the primary
somatosensory cortex in patients with unilateral cerebellar
lesions. Brain. 2001;124(Pt 4):757–68.
98. Basbaum AI. Distinct neurochemical features of acute and
persistent pain. Proc Natl Acad Sci USA. 1999;96:7739–43.
99. Woolf CJ, Mannion RJ. Neuropathic pain: aetiology,
symptoms, mechanisms, and management. Lancet. 1999;
353(9168):1959–64.
100. Woolf CJ, Salter MW. Neuronal plasticity: increasing the
gain in pain. Science. 2000;288(5472):1765–9.
101. Thach WT, Bastian AJ. Role of the cerebellum in the control
and adaptation of gait in health and disease. Prog Brain Res.
2004;143:353–66.
102. Bower JM. Is the cerebellum sensory for motor’s sake, or
motor for sensory’s sake: the view from the whiskers of a rat?
Prog Brain Res. 1997;114:463–96.
103. Fox PT, Raichle ME, Thach WT. Functional mapping of
the human cerebellum with positron emission tomography.
Proc Natl Acad Sci USA. 1985;82(21):7462–6.
104. Kim SG, Ugurbil K, Strick PL. Activation of a cerebellar
output nucleus during cognitive processing. Science.
1994;265(5174):949–51.
105. Hsieh JC, Stahle-Backdahl M, Hagermark O, Stone-
Elander S, Rosenquist G, Ingvar M. Traumatic nociceptive
pain activates the hypothalamus and the periaqueductal
gray: a positron emission tomography study. Pain. 1996;64:
303–14.
106. Casey KL, Minoshima S, Morrow TJ, Koeppe RA.
Comparison of human cerebral activation pattern during
cutaneous warmth, heat pain, and deep cold pain. J
Neurophysiol. 1996;76:571–81.
107. Svensson P, Minoshima S, Beydoun A, Morrow TJ,
Casey KL. Cerebral processing of acute skin and muscle
pain in humans. J Neurophysiol. 1997;78:450–60.
108. Xu X, Fukuyama H, Yazawa S, Mima T, Hanakawa T,
Magata Y, et al. Functional localization of pain perception in
the human brain studied by PET. Neuroreport. 1997;8:
555–9.
109. Derbyshire SW, Jones AK. Cerebral responses to a continual
tonic pain stimulus measured using positron emission
tomography. Pain. 1998;76:127–35.
110. Iadarola MJ, Berman KF, Zeffiro TA, Byas-Smith MG,
Gracely RH, Max MB, et al. Neural activation during acute
capsaicin-evoked pain and allodynia assessed with PET.
Brain. 1998;121(Pt 5):931–47.
111. May A, Kaube H, Buchel C, Eichten C, Rijntjes M,
Juptner M, et al. Experimental cranial pain elicited by
capsaicin: a PET study. Pain. 1998;74:61–6.
112. Paulson PE, Minoshima S, Morrow TJ, Casey KL. Gender
differences in pain perception and patterns of cerebral
activation during noxious heat stimulation in humans. Pain.
1998;76:223–9.
113. Becerra LR, Breiter HC, Stojanovic M, Fishman S,
Edwards A, Comite AR, et al. Human brain activation
under controlled thermal stimulation and habituation to
noxious heat: an fMRI study. Magn Reson Med.
1999;41:1044–57.
114. Coghill RC, Sang CN, Maisog JM, Iadarola MJ. Pain
intensity processing within the human brain: a bilateral,
distributed mechanism. J Neurophysiol. 1999;82:1934–43.
115. Peyron R, Garcia-Larrea L, Gregoire MC, Costes N,
Convers P, Lavenne F, et al. Haemodynamic brain
responses to acute pain in humans: sensory and attentional
networks. Brain. 1999;122(Pt 9):1765–80.
116. Becerra L, Breiter HC, Wise R, Gonzalez RG, Borsook D.
Reward circuitry activation by noxious thermal stimuli.
Neuron. 2001;32:927–46.
117. Casey KL, Morrow TJ, Lorenz J, Minoshima S. Temporal
and spatial dynamics of human forebrain activity during heat
pain: analysis by positron emission tomography. J
Neurophysiol. 2001;85:951–9.
118. Coghill RC, Gilron I, Iadarola MJ. Hemispheric lateraliza-
tion of somatosensory processing. J Neurophysiol.
2001;85:2602–12.
119. Bingel U, Quante M, Knab R, Bromm B, Weiller C,
Buchel C. Subcortical structures involved in pain processing:
evidence from single-trial fMRI. Pain. 2002;99:313–21.
120. Derbyshire SW, Jones AK, Creed F, Starz T, Meltzer CC,
Townsend DW, et al. Cerebral responses to noxious thermal
stimulation in chronic low back pain patients and normal
controls. Neuroimage. 2002;16:158–68.
121. Helmchen C, Mohr C, Erdmann C, Petersen D,
Nitschke MF. Differential cerebellar activation related to
perceived pain intensity during noxious thermal stimulation
in humans: a functional magnetic resonance imaging study.
Neurosci Lett. 2003;335:202–6.
122. Koyama T, McHaffie JG, Laurienti PJ, Coghill RC. The
single-epoch fMRI design: validation of a simplified para-
digm for the collection of subjective ratings. Neuroimage.
2003;19:976–87.
123. Strigo IA, Duncan GH, Boivin M, Bushnell MC.
Differentiation of visceral and cutaneous pain in the human
brain. J Neurophysiol. 2003;89:3294–303.
124. Ibinson JW, Small RH, Algaze A, Roberts CJ, Clark DL,
Schmalbrock P. Functional magnetic resonance imaging
studies of pain: an investigation of signal decay during and
across sessions. Anesthesiology. 2004;101:960–9.
125. Wager TD, Rilling JK, Smith EE, Sokolik A, Casey KL,
Davidson RJ, et al. Placebo-induced changes in FMRI in the
271Pain and the cerebellum

anticipation and experience of pain. Science. 2004;
303(5661):1162–7.
126. Wiech K, Seymour B, Kalisch R, Stephan KE,
Koltzenburg M, Driver J, et al. Modulation of pain
processing in hyperalgesia by cognitive demand.
Neuroimage. 2005;27:59–69.
127. Albuquerque RJ, de Leeuw R, Carlson CR, Okeson JP,
Miller CS, Andersen AH. Cerebral activation during
thermal stimulation of patients who have burning mouth
disorder: an fMRI study. Pain. 2006;122:223–34.
128. Kong J, White NS, Kwong KK, Vangel MG, Rosman IS,
Gracely RH, et al. Using fMRI to dissociate sensory
encoding from cognitive evaluation of heat pain intensity.
Hum Brain Mapp. 2006;27:715–21.
129. Seminowicz DA, Davis KD. Interactions of pain
intensity and cognitive load: the brain stays on task. Cereb
Cortex. 2007;17(6):1412–22.
130. Bantick SJ, Wise RG, Ploghaus A, Clare S, Smith SM,
Tracey I. Imaging how attention modulates pain in humans
using functional MRI. Brain. 2002;125(Pt 2):310–9.
131. Brooks JC, Nurmikko TJ, Bimson WE, Singh KD,
Roberts N. fMRI of thermal pain: effects of stimulus
laterality and attention. Neuroimage. 2002;15:293–301.
132. Smith KA, Ploghaus A, Cowen PJ, McCleery JM,
Goodwin GM, Smith S, et al. Cerebellar responses during
anticipation of noxious stimuli in subjects recovered from
depression. Functional magnetic resonance imaging study.
Br J Psychiatry. 2002;181:411–5.
133. Moriguchi Y, Decety J, Ohnishi T, Maeda M, Mori T,
Nemoto K, et al. Empathy and judging other’s pain: an
fMRI study of alexithymia. Cereb Cortex. 2006 Dec 5;
[Epub ahead of print].
134. Ducreux D, Attal N, Parker F, Bouhassira D. Mechanisms
of central neuropathic pain: a combined psychophysical and
fMRI study in syringomyelia. Brain. 2006;129(Pt 4):963–76.
135. Schweinhardt P, Glynn C, Brooks J, McQuay H, Jack T,
Chessell I, et al. An fMRI study of cerebral processing of
brush-evoked allodynia in neuropathic pain patients.
Neuroimage. 2006;32:256–65.
136. Witting N, Kupers RC, Svensson P, Jensen TS. A PET
activation study of brush-evoked allodynia in patients with
nerve injury pain. Pain. 2006;120:145–54.
137. Geha PY, Baliki MN, Chialvo DR, Harden RN, Paice JA,
Apkarian AV. Brain activity for spontaneous pain of
postherpetic neuralgia and its modulation by lidocaine patch
therapy. Pain. 2007;128:88–100.
272 D. Borsook et al.




















