
Highly sulphated galactan from Halymenia durvillei (Halymeniales, Rhodophyta), a red seaweed of...
6
International Journal of Biological Macromolecules 45 (2009) 140–145 Contents lists available at ScienceDirect International Journal of Biological Macromolecules journal homepage: www.elsevier.com/locate/ijbiomac Highly sulphated galactan from Halymenia durvillei (Halymeniales, Rhodophyta), a red seaweed of Madagascar marine coasts T.A. Fenoradosoa a,b , C. Delattre a , C. Laroche a , A. Wadouachi c , V. Dulong d , L. Picton d , P. Andriamadio b , P. Michaud a,∗ a Laboratoire de Génie Chimique et Biochimique, Université Blaise Pascal, Polytech Clermont-Ferrand, 24 avenue des Landais, 63174 Aubière Cedex, France b Laboratoire de Chimie des Substances Naturelles, Université d’Antsiranana (201), Antsiranana, Madagascar c Laboratoire des Glucides UMR 6219, Université de Picardie Jules Verne, 33 rue Saint-Leu, F-80039 Amiens, France d Laboratoire Polymères Biopolymères Surfaces CNRS FRE 3101, Université de Rouen, Bd Maurice de Broglie, 76821 Mont Saint Aignan Cedex, France article info Article history: Received 13 March 2009 Received in revised form 20 April 2009 Accepted 22 April 2009 Available online 3 May 2009 Keywords: Galactan Halymenia Carrageenan abstract Halymenia durvillei is a red seaweed with a great potential as sulphated galactan producer collected in the coastal waters of small island of Madagascar (Nosy-be in Indian Ocean). To elucidate the structure of its polysaccharide, NMR ( 1 H and 13 C), FTIR, HPAEC and different colorimetric methods were carried out. It has been shown that this polysaccharide, consisted mainly of galactose, was branched by xylose and galactose in minor amounts. Arabinose and fucose were also detected. This galactan was found highly sulphated (42%, w/w) and pyruvylated (1.8%, w/w). Analysis of glycosidic linkages by CPG-MS and 13 C NMR indicated that the polysaccharide has the defining linear backbone of alternating 3--d-galactopyranosyl units and 4-linked -l/d-galactopyranosyl residues. 3,6-Anhydrogalactose units have been also detected in minor quantity. This -carrageenan like polysaccharide has shown original sulphatation patterns with 2-O (26%) or 2/6-O (58%) sulphated 3-linked -d-galactopyranosyl units and 6-O (19%) or 2/6-O (47%) 4-linked -l/d-galactopyranosyl residues. © 2009 Elsevier B.V. All rights reserved. 1. Introduction Red algae (Rhodophyta) are characterized by their content on non-fibrillar and sulphated polysaccharides such as carrageenans, agars and complex sulphated galactans which are the main com- ponents of their cell walls [1,2]. These polysaccharides, well known for their gelling and thickening properties [2,3], are made up of lin- ear chains of alternating 3-linked -galactopyranosyl and 4-linked -galactopyranosyl residues. The 3-linked units always belong to d series whereas the 4-linked ones may have the d or l configu- rations, often occurring as a 3,6-anhydrogalactopyranosyl moiety after enzymatic or alkaline treatment [4]. Many rhodophycean galactans have been described depending to their structural fea- tures. Generally the large carrageenan family is obtained from different species of Rhodophyta including Gigartina, Chondrus cris- pus, Eucheuma and Hypnea [5]. They are variably sulphated on C-2 and C-4 of the 3-linked -d-galactopyranosyl unit and have a part of 4-linked -d-galactopyranoses in 3,6-anhydrogalactopyranosyl configuration. Note that this residue can carry sulphate hemiester groups on position C-2 and/or C-6 [5,6]. Agar is a complex polysac- charide made up of a neutral fraction called agarose (or agaran) ∗ Corresponding author. Tel.: +33 0473407425; fax: +33 0473407402. E-mail address: [email protected] (P. Michaud). and a anionic agaropectin. It is extracted from marine red algae as Gelidium, Gracilaria, Hypnea, Pterocladia and Petrocladiella. The main difference between agarose and carrageenans are a 4-linked -galactopyranose in l configuration and a low sulphate content. Moreover methyl groups may be present at up to 20% of the 3- linked units [1,2]. Finally, can be classified as “complex galactans” linear backbone of galactoses arranged partially with the previ- ously described alternating sequence and ramified by other hexoses and/or pentoses. These polysaccharides can be also methylated, sul- phated and may carry pyruvic acid. -d/l-Galactopyranoses linked through positions C-1 and C-4 can be also chemically and enzy- matically converted to 3,6-anhydro forms. Chemical structures of these polysaccharides are very complex and indeed one view would be that there is no structure in the sense that agar or car- rageenan has a structure. Complex galactans have been extracted, purified and characterized notably from different genera of the order Halymeniales (Cryptonemiales) such as: Grateloupia, Pachy- menia, Phyllymenia and Aeodes [7–13]. Most of them carry sulphate at C-2 of the -d-galactose residue and could be assimilated to car- rageenans belonging to family. However, they differ from them by their amounts of 3,6-anhydrogalactose, the presence of l-galactose, the hexose and pentose substituents and methylated galactoses. Our study focuses on the determination of the main structural characteristics of a complex sulphated galactan extracted from Halymenia durvillei (a red seaweed of the family Cryptonemiaceae, 0141-8130/$ – see front matter © 2009 Elsevier B.V. All rights reserved. doi:10.1016/j.ijbiomac.2009.04.015
-
Upload
u-picardie -
Category
Documents
-
view
6 -
download
0
Transcript of Highly sulphated galactan from Halymenia durvillei (Halymeniales, Rhodophyta), a red seaweed of...

Ha
TPa
b
c
d
a
ARRAA
KGHC
1
napfe�dragtdpaocgc
0d
International Journal of Biological Macromolecules 45 (2009) 140–145
Contents lists available at ScienceDirect
International Journal of Biological Macromolecules
journa l homepage: www.e lsev ier .com/ locate / i jb iomac
ighly sulphated galactan from Halymenia durvillei (Halymeniales, Rhodophyta),red seaweed of Madagascar marine coasts
.A. Fenoradosoaa,b, C. Delattrea, C. Larochea, A. Wadouachic, V. Dulongd, L. Pictond,
. Andriamadiob, P. Michauda,∗
Laboratoire de Génie Chimique et Biochimique, Université Blaise Pascal, Polytech Clermont-Ferrand, 24 avenue des Landais, 63174 Aubière Cedex, FranceLaboratoire de Chimie des Substances Naturelles, Université d’Antsiranana (201), Antsiranana, MadagascarLaboratoire des Glucides UMR 6219, Université de Picardie Jules Verne, 33 rue Saint-Leu, F-80039 Amiens, FranceLaboratoire Polymères Biopolymères Surfaces CNRS FRE 3101, Université de Rouen, Bd Maurice de Broglie, 76821 Mont Saint Aignan Cedex, France
r t i c l e i n f o
rticle history:eceived 13 March 2009eceived in revised form 20 April 2009ccepted 22 April 2009vailable online 3 May 2009
a b s t r a c t
Halymenia durvillei is a red seaweed with a great potential as sulphated galactan producer collected inthe coastal waters of small island of Madagascar (Nosy-be in Indian Ocean). To elucidate the structure ofits polysaccharide, NMR (1H and 13C), FTIR, HPAEC and different colorimetric methods were carried out.It has been shown that this polysaccharide, consisted mainly of galactose, was branched by xylose and
eywords:alactanalymeniaarrageenan
galactose in minor amounts. Arabinose and fucose were also detected. This galactan was found highlysulphated (42%, w/w) and pyruvylated (1.8%, w/w). Analysis of glycosidic linkages by CPG-MS and 13C NMRindicated that the polysaccharide has the defining linear backbone of alternating 3-�-d-galactopyranosylunits and 4-linked �-l/d-galactopyranosyl residues. 3,6-Anhydrogalactose units have been also detectedin minor quantity. This �-carrageenan like polysaccharide has shown original sulphatation patterns with2-O (26%) or 2/6-O (58%) sulphated 3-linked �-d-galactopyranosyl units and 6-O (19%) or 2/6-O (47%)
ranos
4-linked �-l/d-galactopy. Introduction
Red algae (Rhodophyta) are characterized by their content onon-fibrillar and sulphated polysaccharides such as carrageenans,gars and complex sulphated galactans which are the main com-onents of their cell walls [1,2]. These polysaccharides, well knownor their gelling and thickening properties [2,3], are made up of lin-ar chains of alternating 3-linked �-galactopyranosyl and 4-linked-galactopyranosyl residues. The 3-linked units always belong toseries whereas the 4-linked ones may have the d or l configu-
ations, often occurring as a 3,6-anhydrogalactopyranosyl moietyfter enzymatic or alkaline treatment [4]. Many rhodophyceanalactans have been described depending to their structural fea-ures. Generally the large carrageenan family is obtained fromifferent species of Rhodophyta including Gigartina, Chondrus cris-us, Eucheuma and Hypnea [5]. They are variably sulphated on C-2nd C-4 of the 3-linked �-d-galactopyranosyl unit and have a part
f 4-linked �-d-galactopyranoses in 3,6-anhydrogalactopyranosylonfiguration. Note that this residue can carry sulphate hemiesterroups on position C-2 and/or C-6 [5,6]. Agar is a complex polysac-haride made up of a neutral fraction called agarose (or agaran)∗ Corresponding author. Tel.: +33 0473407425; fax: +33 0473407402.E-mail address: [email protected] (P. Michaud).
141-8130/$ – see front matter © 2009 Elsevier B.V. All rights reserved.oi:10.1016/j.ijbiomac.2009.04.015
yl residues.© 2009 Elsevier B.V. All rights reserved.
and a anionic agaropectin. It is extracted from marine red algaeas Gelidium, Gracilaria, Hypnea, Pterocladia and Petrocladiella. Themain difference between agarose and carrageenans are a 4-linked�-galactopyranose in l configuration and a low sulphate content.Moreover methyl groups may be present at up to 20% of the 3-linked units [1,2]. Finally, can be classified as “complex galactans”linear backbone of galactoses arranged partially with the previ-ously described alternating sequence and ramified by other hexosesand/or pentoses. These polysaccharides can be also methylated, sul-phated and may carry pyruvic acid. �-d/l-Galactopyranoses linkedthrough positions C-1 and C-4 can be also chemically and enzy-matically converted to 3,6-anhydro forms. Chemical structuresof these polysaccharides are very complex and indeed one viewwould be that there is no structure in the sense that agar or car-rageenan has a structure. Complex galactans have been extracted,purified and characterized notably from different genera of theorder Halymeniales (Cryptonemiales) such as: Grateloupia, Pachy-menia, Phyllymenia and Aeodes [7–13]. Most of them carry sulphateat C-2 of the �-d-galactose residue and could be assimilated to car-rageenans belonging to � family. However, they differ from them by
their amounts of 3,6-anhydrogalactose, the presence of l-galactose,the hexose and pentose substituents and methylated galactoses.Our study focuses on the determination of the main structuralcharacteristics of a complex sulphated galactan extracted fromHalymenia durvillei (a red seaweed of the family Cryptonemiaceae,

l of Bio
oostde[
2
2
o2espw
2
tafigapfvla
at1Tcp3w
2
ampDAaf
2
Dldcf
pCp
T.A. Fenoradosoa et al. / International Journa
rder Halymeniales) collected in the coastal waters of small islandf Madagascar (Nosy-be in Indian Ocean). The genus Halymeniapecies durvillei present four varieties, but the taxonomic interrela-ionship among the four varieties (var. formosa, var. ceylanica, var.enudata and var. edentata) remained unresolved, although appar-ntly encompassed within the morphological range of H. durvillei14].
. Materials and methods
.1. Material
The red seaweed H. durvillei was collected in the coastal watersf small island of Madagascar (Nosy-be in Indian Ocean) in August006. Thallus was cleaned manually with sea water to eliminatepiphytes and then sun-dried at ambient temperature before to betored in aerated bags in a shaded and ventilate site. Before eacholysaccharide extraction, the sample was washed abundantly withater and dried during 30 h at 65 ◦C.
.2. Extraction
Extraction of the polysaccharide with water (pH 6.0) at a soluteo solvent ratio of 1:20 (w/v) was conducted at 100 ◦C for 4 h. Sep-ration of the residue from the extract was performed by filtrationrstly through glass filter G-1 (100–160 �m) and secondly throughlass filter G-2 (40–100 �m). The residue was briefly washed withdditional distilled water and the wash was collected to maximizeolysaccharide recovery. The solution was centrifuged at 15,000 × gor 30 min and the supernatant was precipitated with ethanol (3ol.). This step was repeated twice. The polysaccharide was col-ected by centrifugation at 15,000 × g for 30 min, dialyzed overnightgainst water and freeze-dried.
Extraction of polysaccharide with 250 mL of 0.3 M NaOH waspplied to 25 g of dried algae, at 90 ◦C during 4 h. The medium washen diluted with 250 mL of water and filtered through glass filter G-(100–160 �m) and secondly through glass filter G-2 (40–100 �m).he solution was centrifuged at 15,000 × g for 30 min and pre-ipitated with ethanol (3 vol.). This step was repeated twice. Theolysaccharide was collected by centrifugation at 15,000 × g for0 min, dialyzed overnight against water and freeze-dried. This stepas repeated twice.
.3. Solvolytic desulphation
The polysaccharide was converted into the pyridinium saltsnd the solvolytic desulphation was performed according to theethod described by Falshaw and Furneaux [15]. Briefly, the sam-
le (20 mg) was dissolved in 8 mL of a mixture of anhydrousMSO–MeOH–pyridine (89:10:1, v/v/v) and heated at 100 ◦C for 4 h.fter cooling at room temperature, distilled water (5 mL) was addednd the mixture was dialyzed overnight against distilled water andreeze-dried.
.4. Composition analysis
Sulphur content was determined by the turbidimetric method ofodgson and Price [16]. 3,6-Anhydrogalactose content was estab-
ished according to Yaphe and Arsenault [17] using fructose as stan-ard. The presence of pyruvic acetal was determined after polysac-haride hydrolysis by reaction with 2,4-dinitrophenylhydrazone
ollowing the procedure outlined by Sloneker and Orentas [18].The monosaccharide composition was determined using high-erformance anion exchange chromatography (HPAEC), on aarbopac PA-1 analytical column (4 mm × 250 mm). Detection waserformed with a pulsed amperometric ED50 detector (Dionex
logical Macromolecules 45 (2009) 140–145 141
Corp., Sunnyvale, CA). Twenty-five microliters of sample wereinjected. Each carbohydrate concentration was established afterintegration of respective areas [Chromeleon management system(Dionex)] and comparison with standard curves obtained with allrelevant monosaccharide standards (Sigma). To investigate the neu-tral monosaccharides, the elution has been achieved isocraticallywith 16 mM NaOH at a flow rate of 1 mL min−1. On the other hand,to elucidate the acidic sugars composition of all samples, a gradientof 160 mM NaOH (solvent A) and 600 mM ammonium acetate in160 mM NaOH (solvent B) was applied at a flow rate of 1 mL min−1.The gradient contained four steps (expressed in percent B in A): 0%during 10 min; 0–100% from 10 to 40 min; 100% from 40 to 45 min;100–0% from 45 to 50 min.
Partially methylated alditol acetates were prepared by themethod of Ciucanu and Kerek [19] and analysed as described byRay and Lahaye [20] on an Agilent 6890-plus (Agilent technolo-gies) gas chromatograph coupled with a 5973 N Mass Spectrometer(Agilent technologies) working at 70 eV. Chromatography was per-formed on a an apolar CP-SIL-5CB (Chrompack) capillary column(60 m × 0.25 mm id) using the following programme temperature:from 50 to 120 ◦C at 20 ◦C min−1 followed by a step from 120 to240 ◦C (10 min hold) at 4 ◦C min−1. The helium total flow rate was0.9 mL min−1.
2.5. Spectroscopic analysis
For NMR analysis, the dried samples were dissolved in D2O(20–30 mg/500 �L). The NMR spectra of the solutions wererecorded at 80 ◦C using a Bruker Advance 300 spectrometer of300 MHz equipped with 13C/1H dual probe.
Fourier-transform infrared (FTIR) spectra of polysaccharide wererecorded on a Nicolet 380 FT-IR instrument (Thermoelectron Cor-poration). Dried polysaccharides samples (1 mg) were dispersed in100 mg of anhydrous KBr and pressed. The IR spectra were recordedat room temperature in the wavenumber range of 400–4000 cm−1
and referenced against air. A total of 32 scans were averaged foreach sample at 4 cm−1 resolution.
2.6. Rheological measurements
Flow measurements were performed using a controlled stressrheometer AR 2000 (TA instrument) with a Peltier temperature con-trol device. All the measurements were performed with a solventtrap to prevent any evaporation of the solvent. The used geometrywas a cone with diameter of 40 mm and angle of 2◦. Polymer solu-tions were prepared by dissolving the solid polymer in milli-Q water(pure water or with the suitable ionic strength, i.e. NaCl 0.3 M or KCl0.3 M) under vigorous stirring for 24 h at 25 ◦C. The concentration(Cp) was fixed at 20 g L−1 in the so-called semi-dilute regime wherepolymer chains are largely entangled. The solution was left on therheometer plate for 10 min before starting the experiment in orderto allow the recovery of its rheological properties after placing it onthe geometry. Flow experiment were conducted from 0.1 to 300 Pain 10 min (up to higher shear rates plot) and from 300 to 0.1 Pa in10 min (down to lower shear rates plot) in order to check a possiblethixotropy.
The Cross equation [21] (1) is a suitable rheological model clas-sically used for such entangled polymer solution:
� − �∞�0 − �∞
= 11 + (K�)m (1)
�0 is the Newtonian viscosity at low shear rates, �∞ the Newtonianviscosity at high shear rates, K the consistency (s) and m the rateindex.
In our experiments, �∞ was not accessible. So, we have usedthe derivative Williamson model (2) which consists in considering

142 T.A. Fenoradosoa et al. / International Journal of Biological Macromolecules 45 (2009) 140–145
d alka
�r
St
3
3
ewaoyo
3
bihdwwadrtta
to 3,6-anhydrogalactose residues was detected for the threepolysaccharides but its very low intensity even in the case ofthe alkali-modified one confirmed the molar ratios previouslyobtained. More, as seen from the FTIR spectra, the alkaline extrac-tion only slightly modified H. durvillei galactan. The diagnostic
Table 1Sugar composition (mol%) of polysaccharide from Halymenia durvillei extracted inaqueous and alkaline conditions and desulphated.
PN1 PN2 PDS1 PDS2
Sulphatea 44 43 n.d. n.d.Pyruvatea 1.8 1.7 1.7 1.6NSa 40 48 72 85Fucb 2.1 1.6 2 1.7Xylb 2.8 1.82 2.7 2.3Galb 58.2 57.3 64 65AnGalb 6.2 9.8 – –Glcb 16 14.75 16 15Arab 15 15.5 15.3 16
Fig. 1. FTIR spectra of desulphated (A), native (B) an
∞ as negligible according to �0 and �. This approximation seemseasonable when looking at our results:
� − �0
�0= 1
1 + (K�)m (2)
uch model permits the comparison between different systems inerms of �0, K and m.
. Results and discussion
.1. Isolation of galactan sulphated
The polysaccharide from H. durvillei was submitted to aqueousxtractions in neutral and alkaline conditions. Alkaline extractionas tested to increase the yield of polysaccharide even if alter-
tions of monosaccharide composition and substitution were oftenbserved [22]. After alcoholic precipitation and freeze-drying theields obtained were 18% and 15% (w/w) for respectively the aque-us and alkaline conditions.
.2. Composition of polysaccharide
The two polysaccharides were analysed for their compositionefore and after desulphation by solvolysis in dimethyl sulphox-
de. This method was chosen because it has shown previouslyigher recovery compared to other such as methanol–HCl and auto-esulphation [11]. The general expectation was that the extractould be a complex galactan based on galactose as major sugar,ith a range of substituent including sulphate esters, and pyruvic
cetal (Table 1). Glucose, xylose, arabinose and fucose were also
etected but not uronic acids. We noted the high sulphate esteratio of the native polysaccharide and the presence of anhydrogalac-ose leading us to suspect the complex sulphated galactan nature ofhis polysaccharide extract from H. durvillei. The weak increase ofnhydrogalactose molar ratio after an alkaline extraction confirmedli-modified (C) galactans from Halymenia durvillei.
the impact of this treatment on the polysaccharide composition.However no significant decrease of sulphate esters was observedas currently described [23]. The native polysaccharide obtained bywater extraction was used in the following study.
3.3. FTIR spectroscopic analysis
The FTIR spectra (Fig. 1) of the native (A) and alkali-extracted (B)polysaccharides contained strong absorption bands at 1253 cm−1
indicative of S O stretching vibration of sulphate groups [23,24].Another minor band at 1375 cm−1 was also indicative of the sul-phate ester substitution [25]. The signal at 933 cm−1, attributable
NS: neutral sugar; PN1: polysaccharide native (water extract); PN2: polysaccha-ride native (sodium hydroxide extract); PDS1: polysaccharide desulphated (waterextract); PDS2: polysaccharide desulphated (sodium hydroxide extract).
a Percent weight of polysaccharide dry weight.b Mol percent of neutral sugars.
l of Biological Macromolecules 45 (2009) 140–145 143
rgeobba8bagilclbeowtgp
3
fO(rcagwh1tpftostikhsp
Table 2Methylation analysis of the native polysaccharide (PN) and the desulphated (PDS)galactan from Halymenia durvillei.
Derivative PN (mol%) PDS (mol%)
3-Linked residues2,4,6-Me3-Gal 2.3 424,6-Me2-Gal 17 4.7
4-Me-Gal 27 –
4-Linked residues2,3,6-Me3-Gal 11 472,3-Me2-Gal 6 13-Me-Gal 15 1.72,3,6-Me3-Glc 3.1 3.3
Undefined residues2-Me-Gal 9.5 1.3Gal 9.2 –
T.A. Fenoradosoa et al. / International Journa
egion of the FTIR spectra between 930 and 800 cm−1 of nativealactan contained one major absorption band at 825 cm−1. How-ver, on the spectrum of alkali treated polysaccharide it wasbserved two absorption bands at 835 and 823 cm−1, proba-ly not resoluted in the spectrum of native galactan. These twoands were respectively attributed to equatorial sulphate esterst O-2 and O-6 of galactose residues respectively. The band at20 cm−1 is effectively well known in IR spectra of �-carrageenansecause of its diagnostic sulphate ester substitution at O-6 of 4-nd 3-linked galactose residues [26]. However, alkalin-treatmentsenerally decrease its intensity and increase proportionally thentensity of the absorption band at 933 cm−1 by conversion of 4-inked 6-O-sulphated galactoses in 3,6-anhydrogalactose. So weonclude that H. durvillei produced an alkalin resistant pyruvy-ated galactan 2-O and 6-O-sulphated. No significative absorptionands around 850 and 805 cm−1 indicated the lack of axial sulphatester substitutions at the O-4 of a 3-linked galactose and at the O-2f a 4-linked 3,6-anhydrogalactose respectively [27]. In the sameay, the very low intensity of absorption at 900 cm−1 suggested
he polysaccharide contained minor ratio of unsulphated 3-linkedalactopyranoses and/or 6-O methyl galactopyranose and/or unsul-hated residues bearing pyruvate acetal at O-4 and O-6 [28].
.4. Linkage analysis
Methylation analysis of desulphated polysaccharide extractedrom H. durvillei revealed large amounts of 2,4,6- and 2,3,6-tri--methyl galactoses in the ratio 42:47, indicating the presence of
1,3)- and (1,4)-linked galactopyranoses respectively (Table 2). Thisesult confirms the existence of galactose majority in the polysac-haride extracted. In literature, the polysaccharides from red algaere described as having regularly alternating units of 4-�-d/l-alactopyranoses and 3-�-d-galactopyranoses [1]. Relying on this,e can reasonably assume that the polymer of Halymenia was aighly sulphated galactan branched. Indeed, small proportions of-linked xylose and 1-linked galactopyranose have been attributedo non-sulphated single terminal residues indicative of a branchedolysaccharide (around 5%) because their molar ratios were similaror native and desulphated polysaccharide. The presence of 2,3,6-ri-O-methyl glucopyranose could be associated to minor amountsf (1,4)-linked glucopyranoses or to residual floridean starch. Theecond hypothesis was the most likely because the floridean ishe principal storage glucan of red algae, often reported to remain
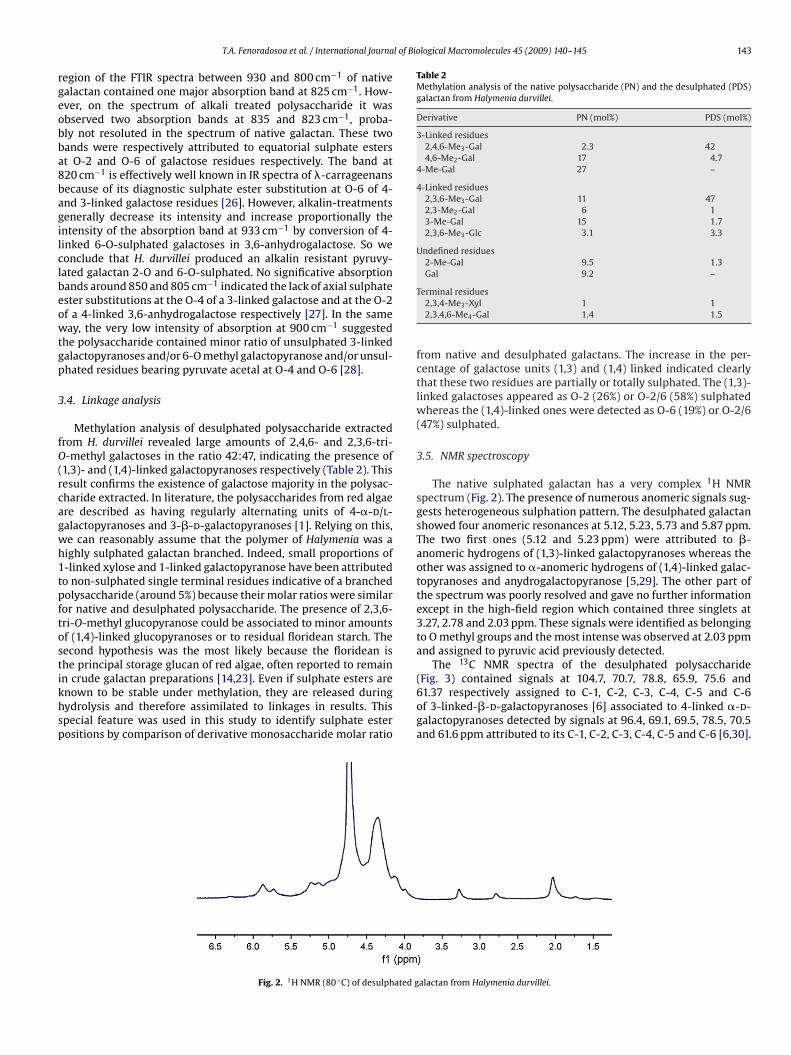
n crude galactan preparations [14,23]. Even if sulphate esters arenown to be stable under methylation, they are released duringydrolysis and therefore assimilated to linkages in results. Thispecial feature was used in this study to identify sulphate esterositions by comparison of derivative monosaccharide molar ratioFig. 2. 1H NMR (80 ◦C) of desulphated g
Terminal residues2,3,4-Me3-Xyl 1 12,3,4,6-Me4-Gal 1.4 1.5
from native and desulphated galactans. The increase in the per-centage of galactose units (1,3) and (1,4) linked indicated clearlythat these two residues are partially or totally sulphated. The (1,3)-linked galactoses appeared as O-2 (26%) or O-2/6 (58%) sulphatedwhereas the (1,4)-linked ones were detected as O-6 (19%) or O-2/6(47%) sulphated.
3.5. NMR spectroscopy
The native sulphated galactan has a very complex 1H NMRspectrum (Fig. 2). The presence of numerous anomeric signals sug-gests heterogeneous sulphation pattern. The desulphated galactanshowed four anomeric resonances at 5.12, 5.23, 5.73 and 5.87 ppm.The two first ones (5.12 and 5.23 ppm) were attributed to �-anomeric hydrogens of (1,3)-linked galactopyranoses whereas theother was assigned to �-anomeric hydrogens of (1,4)-linked galac-topyranoses and anydrogalactopyranose [5,29]. The other part ofthe spectrum was poorly resolved and gave no further informationexcept in the high-field region which contained three singlets at3.27, 2.78 and 2.03 ppm. These signals were identified as belongingto O methyl groups and the most intense was observed at 2.03 ppmand assigned to pyruvic acid previously detected.
The 13C NMR spectra of the desulphated polysaccharide
(Fig. 3) contained signals at 104.7, 70.7, 78.8, 65.9, 75.6 and61.37 respectively assigned to C-1, C-2, C-3, C-4, C-5 and C-6of 3-linked-�-d-galactopyranoses [6] associated to 4-linked �-d-galactopyranoses detected by signals at 96.4, 69.1, 69.5, 78.5, 70.5and 61.6 ppm attributed to its C-1, C-2, C-3, C-4, C-5 and C-6 [6,30].alactan from Halymenia durvillei.

144 T.A. Fenoradosoa et al. / International Journal of Biological Macromolecules 45 (2009) 140–145
ed (B)
Itbiabts[dsaatis
botatw
3
t
measurements in the semi-dilute regime of concentration, i.e. inthe entangled regime. 20 g L−1 solutions of the native polysaccha-ride were prepared in either NaCl 0.3 M or in KCl 0.3 M in order tofollow their flow properties. Results are reported in Fig. 4. No strong
Fig. 3. 13C NMR native (A) and desulphat
n the �-anomeric region, a significant signal at 101.2 ppm revealedhe presence of the C-1 of 4-linked �-l-galactopyranoses proba-ly associated to a 3-linked-�-d-galactopyranoses whom C-1 was
dentified at 103.9 ppm. The C-5 signal of this �-l-galactopyranoset 72.3 ppm confirmed its presence. The 105.0 ppm resonance coulde attributed to C-1 �-d-xylopyranosyl single stubs linked to a posi-ion of high mobility, possibly the C-6 of �-d-galactopyranosyl ashown by signal of substituted hydroxymethyl group at 69.7 ppm29]. The attribution of other C from d-xylopyranosyl residue wasifficult in regard to the spectra complexity. The lack of specificignals for the C-1 of 4-linked 3,6-anhydro-�-d-galactopyranosest 94.7 ppm [6] reinforced the previous FTIR results and the lowmounts of this sugar detected in the polysaccharide composi-ion. The spectra of native and desulphated galactans showed twontensive signals around 61.5 ppm indicated the presence of unsub-tituted primary alcoholic groups in galactopyranosyl residues.
Signals at 25.79 and 101.7 were attributed to pyruvic methyl car-on and pyruvic acetal carbon respectively [28]. This pyruvylationf this galactan was reinforced by the signal at 65.88 ppm, specific ofhe C-6 and C-5 of a pyruvated �-d-galactopyranose unit [31–33]. Inddition, the high signal at 101.6 ppm for the native galactan spec-ra was attributed to C-1 of a galactopyranose 2-O-sulphated. Thereere also evident absorptions at 66.97 ppm (C-6 sulphate) [34].
.6. Rheological measurements
The native sulphated galactan from H durvillei presents a struc-ure which may be quite similar to that of some carrageenans. As
polysaccharide from Halymenia durvillei.
a consequence, we have conducted some preliminary rheological
Fig. 4. Flow curves (25 ◦C) of the native sulphated galactan (20 g L−1) in NaCl (circles)and in KCl 0.3 M (square). Up to higher shear rates (full symbols), down to lower shearrates (open symbols). Bold lines: fit of Williamson Model.

T.A. Fenoradosoa et al. / International Journal of Bio
Table 3Williamson parameters for the native sulphated galactan at 20 g L−1 in NaCl and KCl0.3 M extracted from the “up to higher shear rates” plots.
NK
dcmstaeoTcocs�afl
4
ycWhdmbdattp(aatiped
[
[[
[[[
[[[[[[
[[
[[
[
[[
[249–265.
�0 (Pa s) K (s) m
aCl 0.3 M 3.4 0.08 0.7Cl 0.3 M 3.9 0.10 0.7
ifferences appear between flows of the polymer at the same con-entration in both salts at the same ionic strength. The Williamsonodel applied on these results (Table 3) indicates that the ‘zero
hear’ viscosity (i.e. �0) is slightly higher in potassium chloridehan in sodium chloride. Other parameters from Williamson modelre quite similar whatever the used salt. Nevertheless, this differ-nce seems not so significant and do not indicate the formationf a structured liquid (pseudogel) in the presence of potassium.hese solutions have been brought at 80 ◦C during 10 min and thenooled at ambient temperature. In these conditions, any pseudogelsr networks of native sulphated galactan have been obtained as itan be observed with the �- or �-carrageenan [3]. In fact the nativeulphated galactan seems to be quite rheologically similar to the-carrageenan with a classical and good thickening ability. Fig. 4lso indicates that any thixotropy occurs during the up and downow scans.
. Conclusion
This study is the first description and partial structural anal-sis of a polysaccharide extracted from H. durvillei, a red algaeollected in the marine coasts of Madagascar (Indian Ocean).e have shown that this polysaccharide is a pyruvylated and
ighly sulphated galactan predominantly composed of 3-linked �--galactopyranoses and 4-linked-�-d/l-galactopyranoses havinginor amounts of 3,6-anhydrogalactopyranoses. Glucose, ara-
inose, xylose and fucose have also been detected but only-xylopyranoses andd-galactopyranoses branched units have beenssigned to the polysaccharide structure. The (1,3)-linked galac-oses appeared as O-2 (26%) and O-2/6 (58%) sulphated whereashe (1,4)-linked ones were detected as O-6 (19%) or O-2/6 (47%) sul-hated. This result differs from those previously published where1,3)-linked galactoses O-2/6 sulphated have not been describedt your knowledge. So the polysaccharide from H durvillei may bessimilated to a �-carrageenan polymer pyruvylated with some of
hese 3-linked galactopyranoses 2/6 (47%) O-sulphated. Next stud-es will deal with the physicochemical characterisation of solutionroperties of the polysaccharide from H durvillei by means of sizexclusion chromatography coupled on-line with both static andynamic light scattering, viscometry and rheology.[[
[
logical Macromolecules 45 (2009) 140–145 145
Acknowledgement
This work was supported by the Agence Universitaire de la Fran-cophonie.
References
[1] M. Rinaudo, in: J.P. Kamerling, G.J. Boons, Y.C. Lee, A. Suzuki, N. Taniguchi, A.G.V.Voragen (Eds.), Comprehensive Glycoscience, vol. 2, Elsevier, Oxford, 2007, pp.691–735.
[2] S.H. Knutsen, D.E. Myslabodski, B. Larsen, A.I. Usov, Bot. Mar. 37 (1994) 163–169.
[3] F. van de Velde, G.A. De Ruiter, in: A. Steinbüchel, S. DeBaets, E.J. VanDamme(Eds.), Biopolymers (vol6) Polysaccharide II Polysaccharides from Eukaryotes,Wiley–VCH, Weinheim, 2002, pp. 245–274.
[4] T.J. Painter, in: G.O. Aspinall (Ed.), The Polysaccharides, vol. 2, Academic Press,New York, 1983, pp. 195–285.
[5] V.L. Campo, F. Kawano, D.B. Silva Junior, I., Carbohydr. Polym. 77 (2009)167–180.
[6] F. van de Velde, S.H. Knutsen, A.I. Usov, H.R. Rollema, A.S. Cerezo, Trends FoodSci. Technol. 13 (2002) 73–92.
[7] I.J. Miller, R. Falshaw, R.H. Furneaux, Carbohydr. Res. 268 (1995) 219–232.[8] S. Hirase, C. Araki, K. Watanabe, Bull. Chem. Soc. Jpn. 40 (1967) 1445–1448.[9] A.J. Farrant, J.R. Nunn, H. Parolis, Carbohydr. Res. 19 (1971) 161–168.10] A.J. Farrant, J.R. Nunn, H. Parolis, Carbohydr. Res. 25 (1972) 283–292.
[11] K. Chattopadhyay, C.G. Mateu, P. Mandal, C.A. Pujol, E.B. Damonte, B. Ray, Phy-tochemistry 68 (2007) 1428–1435.
12] J.R. Nunn, H. Parolis, Carbohydr. Res. 14 (1970) 145–150.13] A.J.R. Allsobrook, J.R. Nunn, H. Parolis, Carbohydr. Res. 16 (1971)
71–78.14] S. Kawaguchi, S. Shimada, T. Abe, R. Terada, Coast. Mar. Sci. 30 (2006) 201–208.15] R. Falshaw, R.H. Furneaux, Carbohydr. Res. 307 (1998) 325–331.16] K.S. Dodgson, R.G. Price, Biochem. J. 84 (1962) 106–110.
[17] W. Yaphe, G.P. Arsenault, Anal. Biochem. 13 (1965) 143–148.18] J.H. Sloneker, D.G. Orentas, Nature 194 (1962) 478–479.19] I. Ciucanu, F. Kerek, Carbohydr. Res. 131 (1984) 209–217.20] B. Ray, M. Lahaye, Carbohydr. Res. 274 (1995) 313–318.21] M.M. Cross, J. Colloid Sci. 20 (1965) 417–423.22] C.W. Greer, W. Yaphe, Bot. Mar. 27 (1984) 473–478.23] M.-L. Liao, A. Chiovitti, S.L.A. Munro, D.J. Craik, G.T. Kraft, A. Munro, D.J. Craik,
G.T. Kraft, A. Bacic, Carbohydr. Res. 296 (1996) 237–247.24] A.G. Lloyd, K.S. Dodgson, R.B. Price, F.A. Rose, BBA 46 (1961) 108–115.25] R. Tuvikene, K. Truus, M. Robal, T. Pehk, T. Kailas, M. Vaher, T. Paalme, Carbohydr.
Res. 344 (2009) 788–794.26] T. Chopin, B.F. Kerin, R. Mazerolle, Phycol. Res. 47 (1999) 167–188.27] J. Prado-Fernandez, J.A. Rodriguez-Vasquez, E. Tojo, J.M. Andrade, Anal. Chim.
Acta 480 (2003) 23–37.28] A. Chiovitti, A. Bacic, D.J. Craik, G.T. Kraft, M.-L. Liao, R. Falshaw, R.H. Furneaux,
Carbohydr. Res. 310 (1998) 77–83.29] A.I. Usov, M.I. Bilan, A.S. Shashkoov, Carbohydr. Res. 300 (1997) 93–102.30] B. Matsuhiro, A.F. Conte, E.B. Damonte, A.A. Kolender, M.C. Matulewicz, E.G.
Mejías, C.A. Pujol, E.A. Zúniga, Carbohydr. Res. 340 (2005) 2392–2402.31] M. Lahaye, W. Yaphe, M.T. Phan Viet, C. Rochas, Carbohydr. Res. 190 (1989)
32] M. Lahaye, C. Rochas, Hydrobiologia 221 (1991) 137–148.33] A. Chiovitti, A. Bacic, D.J. Craik, S.L.A. Munro, G.T. Kraft, M.L. Liao, Carbohydr.
Res. 299 (1997) 229–243.34] M.E.R. Duarte, M.D. Noseda, M.A. Cardoso, S. Tulio, A.S. Cerezo, Carbohydr. Res.
337 (2002) 1137–1144.




















