
Increases in sulphated glycoprotein-2 mRNA levels in the rat brain after transient forebrain...
15
Molecular Brain Research, 18 (1993) 163-177 163 © 1993 Elsevier Science Publishers B.V. All rights reserved 0169-328x/93/$06.00 BRESM 70588 Increases in sulphated glycoprotein-2 mRNA levels in the rat brain after transient forebrain ischemia or partial mesodiencephalic hemitransection Michele Zoli a Francesco Ferraguti a,, Isabella Zini a Saverio Bettuzzi b and Luigi F. Agnati a " Institute of Human Physiology and b Biological Chemistry, University ofModena, Modena (Italy) (Accepted 24 November 1992) Key words: Sulphated glycoprotein-2iTransient forebrain ischemia; Mesodiencephalic hemitransection; Cell death; In situ hybridization; Central nervous system; Rat Sulphated glycoprotein-2, thought to be involved in programmed cell death in peripheral organs, has been detected at increased levels in brain degenerative states. In this paper we have investigated the regional and cellular expression of this protein during development of brain lesion. With this aim sulphated glycoprotein-2 mRNA levels were studied in models of ischemic (transient forebrain ischemia) or mechanical (partial mesodiencephalic hemitransection) brain injuries using in situ hybridization histochemistry. Marked increases in sulphated glycoprotein-2 mRNA were observed in lesioned brains in both models. In addition, we report a shift in the regional distribution of positive cells in both lesion models 1-7 days post-lesion. After transient forebrain ischemia, sulphated glycoprotein-2 mRNA increases were always localized in selectively vulnerable regions (caudate-putamen, hippocampal formation), showing a temporal change in the pattern of intraregional distribution from less to more lesioned parts. In the case of mechanical lesion at 1 day, increased labelling had a widespread distribution on the lesioned side and was also observed on the intact side near the midline. In contrast, at 7 days increased labelling was restricted to regions directly lesioned (either areas whose input and/or output connections were severed by the transection or areas which were directly affected by the mechanical lesion). Analysis at the cellular level revealed that at both time intervals and in both lesion models most cell bodies overlain by dense clusters of specific grains were non-neuronal cells. The distribution patterns and their change over time suggest that at least some of these cells are inflammatory and phagocytic cells. The majority of degenerating neuronal cells after ischemia did not show increased levels of sulphated glycoprotein-2 mRNA. However, seven days after hemitransection and at all time intervals after transient ischemia, some cells clearly identifiable as neurones exhibited increased sulphated glycoprotein-2 mRNA levels. INTRODUCTION Sulphated glycoprotein-2 (SGP-2), a heterodimer composed of 34 and 47 kDA subunits, can be detected in a variety of tissues and animals (including birds and mammals). In the rat, SGP-2 has been shown to be equivalent to clusterin 17 and to testosterone-repressed prostate message-27'25. The bovine homologue of SGP-2 has been isolated from adrenal chromaffin cell gran- ules and named glycoprotein II135. In the human, a protein with high homology to rat SGP-2 has been isolated in different tissues and named serum and seminal plasma protein SP-40,40 22, complement cytoly- sis inhibitor z° and apolipoprotein j12. Sulphated glyco- protein-2 protein and/or mRNA have been detected in most rat tissues being particularly concentrated in the urogenital system (prostate and testicle, where it is the major component of Sertoli cell secretions; ref. 10) 2. In normal rat brain, SGP-2 mRNA has been detected by both Northern blot 2 and in situ hybridiza- tion histochemistry ts. The widespread distribution of SGP-2 in several tissues and organisms argues for an important role for this protein. Although its physiological function remains un- known, a variety of data suggest that in peripheral tissues increased levels of SGP-2 are related to pro- grammed cell death 3'4'7'8. Marked increases in SGP-2 transcript levels have been demonstrated in models of Correspondence: L.F. Agnati, Istituto di Fisiologia Umana, Universita' di Modena, Via Campi 287, 41100 Modena, Italy. Fax: (39) 59-367372. * Present address: Glaxo Research Laboratories, Verona, Italy.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Increases in sulphated glycoprotein-2 mRNA levels in the rat brain after transient forebrain...

Molecular Brain Research, 18 (1993) 163-177 163 © 1993 Elsevier Science Publishers B.V. All rights reserved 0169-328x/93/$06.00
BRESM 70588
Increases in sulphated glycoprotein-2 mRNA levels in the rat brain after transient forebrain ischemia
or partial mesodiencephalic hemitransection
M i c h e l e Z o l i a F r a n c e s c o F e r r a g u t i a , , I s a b e l l a Z i n i a S a v e r i o B e t t u z z i b a n d L u i g i F . A g n a t i a
" Institute of Human Physiology and b Biological Chemistry, University ofModena, Modena (Italy)
(Accepted 24 November 1992)
Key words: Sulphated glycoprotein-2i Transient forebrain ischemia; Mesodiencephalic hemitransection; Cell death; In situ hybridization; Central nervous system; Rat
Sulphated glycoprotein-2, thought to be involved in programmed cell death in peripheral organs, has been detected at increased levels in brain degenerative states. In this paper we have investigated the regional and cellular expression of this protein during development of brain lesion. With this aim sulphated glycoprotein-2 mRNA levels were studied in models of ischemic (transient forebrain ischemia) or mechanical (partial mesodiencephalic hemitransection) brain injuries using in situ hybridization histochemistry. Marked increases in sulphated glycoprotein-2 mRNA were observed in lesioned brains in both models. In addition, we report a shift in the regional distribution of positive cells in both lesion models 1-7 days post-lesion. After transient forebrain ischemia, sulphated glycoprotein-2 mRNA increases were always localized in selectively vulnerable regions (caudate-putamen, hippocampal formation), showing a temporal change in the pattern of intraregional distribution from less to more lesioned parts. In the case of mechanical lesion at 1 day, increased labelling had a widespread distribution on the lesioned side and was also observed on the intact side near the midline. In contrast, at 7 days increased labelling was restricted to regions directly lesioned (either areas whose input and/or output connections were severed by the transection or areas which were directly affected by the mechanical lesion). Analysis at the cellular level revealed that at both time intervals and in both lesion models most cell bodies overlain by dense clusters of specific grains were non-neuronal cells. The distribution patterns and their change over time suggest that at least some of these cells are inflammatory and phagocytic cells. The majority of degenerating neuronal cells after ischemia did not show increased levels of sulphated glycoprotein-2 mRNA. However, seven days after hemitransection and at all time intervals after transient ischemia, some cells clearly identifiable as neurones exhibited increased sulphated glycoprotein-2 mRNA levels.
INTRODUCTION
Sulpha ted glycoprotein-2 (SGP-2), a he te rod imer
composed of 34 and 47 k D A subunits , can be detec ted
in a variety of tissues and animals ( including birds and
mammals) . In the rat, SGP-2 has b e e n shown to be
equivalent to cluster in 17 and to tes tos terone-repressed
prostate message-27'25. The bovine homologue of SGP-2
has been isolated from adrena l chromaff in cell gran-
ules and n a m e d glycoprotein II135. In the human , a
pro te in with high homology to rat SGP-2 has been
isolated in different tissues and n a m e d serum and
seminal p lasma prote in SP-40,40 22, complemen t cytoly-
sis inhibi tor z° and apol ipoprote in j12. Sulphated glyco-
prote in-2 pro te in a n d / o r m R N A have been detected
in most rat tissues being part icularly concen t ra ted in
the urogeni ta l system (prostate and testicle, where it is
the major c ompone n t of Sertoli cell secretions; ref.
10) 2. In normal rat brain, SGP-2 m R N A has been
detected by both Nor the rn blot 2 and in situ hybridiza-
t ion histochemistry ts. The widespread dis t r ibut ion of
SGP-2 in several tissues and organisms argues for an
impor tan t role for this protein.
Al though its physiological funct ion remains un-
known, a variety of data suggest that in per iphera l
tissues increased levels of SGP-2 are related to pro- g rammed cell death 3'4'7'8. Marked increases in SGP-2
t ranscr ipt levels have been demons t ra t ed in models of
Correspondence: L.F. Agnati, Istituto di Fisiologia Umana, Universita' di Modena, Via Campi 287, 41100 Modena, Italy. Fax: (39) 59-367372. * Present address: Glaxo Research Laboratories, Verona, Italy.

164
apoptosis ~'4°'41 in adult tissues (such as prostate during the regression phase following castration 2'4'7 and cul- tured thymocytes after exposure to high levels of gluco- corticoids3), and in developmental cell death (such as of interdigital tissues during ontogenetic regression 7).
Interestingly, increased SGP-2 mRNA, or related protein, levels have been observed in several central neurodegenerative states or experimental brain lesions, namely in the brains of scrapie-infected hamsters ~3, in the hippocampal formation of Alzheimer's and Pick's demented patients ~327, in brain tumors and epileptic foci ~r and in rat striatum 3~' and hippocampus 27 after excitotoxic lesions and transient ischemia 28. Most of these brain alterations have been associated with states of astrogliosis based on parallel increases in glial fibril- lary acidic protein (GFAP), a marker of astrocytic reaction. A direct demonstration of increased SGP-2 transcript levels in astrocytes has in fact been obtained in striatal GFAP-positive astrocytes 3 months after an excitotoxic lesion 3~ and in rat hippocampus following entorhinal cortex lesion z4. Sulphated glycoprotein-2 overexpression in astrocytes (which do not degenerate after lesion) puts into question a strict link between apoptosis and SGP-2 in neurodegeneration. In addi- tion, SGP-2 mRNA is not expressed in neurons during developmental death 15. However, all these data strongly support an important role of this protein in neurode- generation.
Therefore, it seemed of interest to further investi- gate the role of SGP-2 in models of brain lesions. With this aim, we evaluated SGP-2 mRNA levels at regional and cellular levels by means of in situ hybridization histochemistry using a 35S-labelled cRNA probe in various regions of rat brain in two established models of brain lesion, i.e., transient forebrain ischemia in- duced by four-vessel occlusion and partial mesodien- cephalic hemitransection. Several time intervals were investigated to unravel possible changes in the type of cells expressing SGP-2 mRNA and their regional dis- tribution during the development of the lesions.
Partial mesodiencephalic hemitransection severs the lateral portion of nigrostriatal and strionigral pathways, at the same time creating a penetrating lesion around the site of blade insertion (involving parts of cerebral cortex, lateral mesencephalon and ventral hippocam- pus) 1'19"43'44 (and Grimaldi and Zoli, unpublished ob-
servations). The four-vessel occlusion model induces transient ischemia of the forebrain, typically inducing neuronal cell death in the so-called selectively vulnera- ble regions (i.e. CAI hippocampal field, lateral cau- date-putamen, and, to a lesser extent, neocortex and thalamus) 38"3'~'42. Ischemic neuronal death in these ar- eas is not immediate but rather occurs 24-48 h follow-
ing the insult (maturation phenomenon39). It was, then, of particular interest to study the possible expression of SGP-2 mRNA in degenerating neurons during or after the maturation of the ischemic lesion.
M A T E R I A L S A N D M E T H O D S
Preparation of radioactit'e probes Radioactive sense and antisense RNA probes were transcribed
from a 304 bp AatII/Pt:uIl SGP2 cDNA fragment (from position 775 to position 1080) derived from pSB283 that had been subcloned in the transcription vector pGEM 7z (Promega Biotech). Transcrip- tion by T7 RNA polymerase (Boehringer, Mannheim, FRG) of the BamHI linearized plasmid pGEM 7z-304 bp SGP2 produced sense RNA, whereas transcription by SP6-RNA polymerase (Boehringer, Mannheim, FRG) of the Apal linearized pGEM 7z-304 bp SGP2 produced antisense RNA complementary to SGP2 mRNA. Both the sense and antisense probes were labelled with 12.5/~mol of [~SS]UTP (1000 Ci /mmol , Amersham, Buckinghamshire, UK). Transcription was carried out for 2 h at 37°C. The template was digested with RNAse-free DNAse 1 (Boehringer, Mannheim, FRG). The labelled probes were then partially purified by phenol extraction and ethanol precipitation. The pellets were dried under vacuum, redissolved in 50 #1 of 100 mM dithiothreitol (DTT) and stored in small aliquots at
20 °C until used.
Animals Adult male specific pathogen-free Sprague-Dawley rats (200 g
b.wt.) were used. They were kept under standardized temperature, humidity and lighting conditions (lights on at 08.00 h and off at 20.00 h) and had free access to water and food.
Experimental lesion models Transient forebrain ischemia. The four-vessel occlusion method of Pulsinelli and Brierley 3s was used to induce transient forebrain ischemia (see also ref. 42). In this model, the vertebral arteries are permanently occluded by cauterization one day before the transient occlusion of carotid arteries. In order to obtain forebrain ischemia, carotid arteries were clamped with microclips for 30 rain. Then, the carotid clips were removed and the animals were allowed to recover. During the ischemic period, the animals were observed clinically for their level of consciousness, the presence or absence of righting and corneal reflexes, ability to walk and to climb, and electroencephalo- graphic (EEG) activity. Only those animals immediately losing their righting reflex, being unresponsive for 20-30 min after bilateral carotid occlusion, with an isoelectric EEG within 2 -3 rain after carotid artery occlusion and without recovery throughout the is- chemic period, were studied. A full description of the animal model of transient forebrain ischemia and its characterization as regards neuronal damage, cerebrovascular blood cell distribution, neurologi- cal tests, behavioural parameters and arterial blood pressure mea- surements is given in Zini et al. 42. Four animal groups were consid- ered (n = 4 ra ts /group): unoperated rats, and rats sacrificed 1, 2 and 7 days after ischemic insult. Mesodiencephalie hemitransection. Partial hemitransection was per- formed under deep ketamine hydrochloride (Ketalar, Parke-Davis, 100 mg /kg , i.p.) anaesthesia according to a previously published procedure TM. Briefly, a thin metal blade was inserted in the right cerebral hemisphere at the level of bregma with an inclination of 20 ° in order to reach the nigrostriatal pathway at the mesodiencephalic junction (approximately bregma level - 4 . 3 mm according to Paxinos and Watson37). Three animal groups were considered ( n = 3 rats /group): unoperated rats, and hemitransected rats sacrificed 1 and 7 days after the operation.
hz situ hybridization procedure On the day of sacrifice the animals were deeply anaesthetized
with ketamine hydrochloride (100 mg /kg , i.p.) and killed by intracar- diac perfusion with 100 ml of ice-cold saline. Brains were rapidly

dissected out and frozen. Fourteen-/zm-thick coronal sections were cut on a cryostat. In the case of transient forebrain ischemia cuts were taken at bregma levels 1.2 mm (striatal level) and - 3 . 0 mm (hippocampal level) 37. Striatum and hippocampal formation are the regions showing the most consistent and marked loss of neurones in the four-vessel occlusion model of brain ischemia 39'42. In the case of partial hemitransection, sections were taken at bregma levels 1.2 mm (around 1.0 mm anterior to the site where the blade is inserted in the cortex) and - 4.8 mm (around 0.5 mm posterior to the site where the blade reaches the ventral surface of the brain) 37. The sections were fixed in 4% paraformaldehyde in phospate buffered saline and processed for in situ hybridization according to a previously pub- lished procedure 14'44. Film and emulsion autoradiography were per- formed with tritium-sensitive films (Amersham, Buckinghamshire, UK) and autoradiographic emulsion (K5, Ilford, UK), respectively. Sections exposed to autoradiographic emulsion were developed after 45-60 days of exposure and counterstained with Nissl staining. For a characterization of the probe used, see Bettuzzi et al. 5. The regional and cellular distribution of SGP-2 m R N A in brains of normal rats was in good agreement with that observed by other groups (see, in particular, ref. 15). In order to allow comparisons between lesioned and control tissues, all specimens (ischemic animals and respective controls, or hemitransected animals and respective controls) were processed together (same fixative, same labelled probe, same emul- sion and same exposure time).
Analysis of in situ hybridization preparations Regional analysis of SGP-2 labelling has been carried out on both
film and emulsion autoradiographic preparations. For analysis at the cellular level, only cells covered by clear-cut clusters of grains were considered to be positive. Positive cells were identified according to the following objective criterium. For each type of cell in each area analyzed, 50 cells randomly chosen in 2 control animals were mea- sured. Manual grain counting was performed at the light microscope. For each type of cell, a threshold was chosen equal to the mean control group value + 3 S.D., and cells above this threshold were considered as overexpressing SGP-2 mRNA. Even if a rather sharp distinction between positive and negative (i.e., control) cells was obtained (see Table I), a limited number of cells with slightly increased SGP-2 m R N A content may have been overlooked.
The density of positive cells in lesioned tissues was assessed in ischemic animals. Each positive cell was attributed to one of the main classes of cells recognizable with Nissl staining (neuronal cells, non-neuronal cells, ventricular and pia mater ependymal cells). Man- ual cell counts were performed by two researchers unaware of t reatment groups in caudate-putamen and hippocampal formation of ischemic rats. In caudate-putamen, circular fields of 0.4 mm diame- ter were sampled (2 measurements per animal, n = 4 animals /group) . In hippocampal formation, a rectangular field with a base of 0.4 mm and spanning the entire CA1 field was sampled (2 measurements per animal, n = 4 animals /group) .
After partial hemitransection, the amount of SGP-2 m R N A la- belling per region was rated relative to labelling in the same region of control animals according to the following scale: - = no increased labelling, + = weakly increased labelling, + = moderately increased labelling, + + = strongly increased labelling. A quantitative assess- ment of cellular SGP-2 m R N A content was performed only for large neurones of the substantia nigra. In this region, clearly recognizable neuronal cells (i.e. cells with cytoplasmic staining and cell body diameter larger than 20 /zm) were considered. Grain counting was performed by means of an automatic image analyzer IBAS I-II (Zeiss Kontron, Munich) 43. Square fields of 4096/xm 2 were selected. Cells were interactively encircled and their areas and grain counts mea- sured.
R E S U L T S
The pattern of neuronal and non-neuronal labelling that we observed in control animals was in fairly good agreement with previous papers (see, e.g., ref. 15).
165
Sulphated glycoprotein-2 mRNA was distributed in a relatively uniform fashion in central neurones of con- trol rats (data not shown). Some neuronal populations, such as neurones of the pars compacta of the substan- tia nigra and the preoptic hypothalamic nucleus ~5, ex- hibited higher baseline levels of labelling, possibly due to the large size of their cell bodies. In control animals, no detectable labelling was observed in most non-neu- ronal cell populations with the notable exception of choroid plexa and ventricular and pia mater ependymal cells which exhibited the highest levels of transcript in normal brain 15'z4.
Following both mechanical and ischemic lesions, a number of cells exhibited clear-cut increases in SGP-2 mRNA levels (see Table I and high magnification photomicrographs throughout the paper). In this pa- per, we have mapped out the distribution of these cells, for which we use the term 'positive cells'.
Transient forebrain ischemia Both localization and development in time of the
lesions after 30 min of 4-vessel occlusion were in agree- ment with previous observations in the same animal model 38'39'4z. Areas of neuronal degeneration were al-
ways observed in selectively vulnerable regions, i.e., lateral caudate-putamen and CA1 hippocampal forma- tion. In lateral caudate-putamen, the lesion was easily recognizable as an area with markedly decreased num- bers of neuronal cells visualized by Nissl staining al- ready at 1 day post-ischemia. In the hippocampal for- mation, no neuronal cell loss was observed 1 day after the insult. On the other hand, at 2 and 7 days most pyramidal neurones disappeared and increase of non- neuronal cells was evident in the CA1 hippocampal field. In the CA3 field and dentate gyrus, signs of neuronal degeneration were minor and inconstant at 1, 2, and 7 days, whereas marked non-neuronal cell in- creases were present. In addition, at all time intervals examined, some animals exhibited patches of neuronal cell loss in neocortex and in the ventrolateral part of the thalamus.
At all levels analyzed, markedly increased SGP-2 mRNA levels were observed exclusively in regions where signs of neuronal degeneration were also pre- sent. Neuronal lesions and increased SGP-2 labelling uniformly have been detected together in both cau- date-putamen and dorsal hippocampal formation of ischemic animals. This was also true for lesions in neocortex and ventrolateral thalamus. However, as these latter lesions were inconstant, we focussed our analysis on lateral caudate-putamen and hippocampal formation.

166
TABLE I
Grain counting of SGP-2 mRNA labelling in several cell types after transient forebrain ischemia
The mean (_+S.D.) number of grains per cell in caudate-putamen and CA1 hippocampal field in control animals, and in rats sacrificed 1 and 7 days after 30 min forebrain ischemia are given. For each control group, 50 cells randomly chosen in 2 animals were measured. Cells above a threshold corresponding to the mean control group value + 3 S.D. were considered as overexpressing SGP-2 mRNA. In lesioned animals 30 positive cells randomly chosen in 2 animals were measured.
Control SGP-2 overexpressing cells cells
1 day 7 days
A. Caudate-putamen Neurons 2.7_+1.8 21.8_+ 3.7 23.4_+4.6 Non-neuronal cells 0.5_+1.0 12.1_+ 4.4 12.1-+2.3
B. Hippocampal formation Pyramidal neurons 7.6 _+ 2.2 - - Interneurons 11.6-+3.8 66.3-+15.6 54.5_+5.9 Non-neuronal cells 1.3 + 1.5 32.3 _+ 6.7 23.9 + 4.0
Caudate-putamen. A temporal shift in the intraregional distribution of increased SGP-2 mRNA labelling was observed. At 1 and 2 days, positive cells were only observed in various grey and white matter regions surrounding the area of neuronal cell disappearance, such as ventral and medial portions of caudate-puta-
men, the nuc. accumbens, the lateral septum and the corpus callosum (Fig. 1A). Instead, 7 days after the lesion, the pattern of increased SGP-2 mRNA labelling was complementary to that observed at 1 and 2 days, as positive cells were exclusively observed inside the area of neuronal loss (Fig. 1B).
At cellular level, both neuronal and non-neuronal cells appeared to express increased levels of SGP-2 mRNA. However, the intraregional distribution and the proportion of positive cells in each cellular popula- tion varied at the different times examined. One and 2 days after ischemic insult, at the medial and ventral borders of the area of neuronal loss, the majority of cells showing clear-cut clusters of grains corresponded to non-neuronal cells (Fig. 2A), representing only a portion of the overall non-neuronal cell population (around 17%, see Table II). In addition, a few positive neuronal cells (around 8%, see Table II), scattered among many negative neurones were detected in the same area (Fig. 2A). At 7 days post-ischemia, positive cells were exclusively concentrated in the area of neu- ronal cell loss. Positive cells were both non-neuronal and neuronal, again constituting only a fraction of the respective overall cell populations (Fig. 2B). However, at this time interval the proportion of positive cells in the two groups was inverted with respect to the earlier
TABLE II
Density of SGP-2 mRNA overexpressing cells after transient forebrain ischemia in caudate-putamen and hippocampal field CA1
The mean (.+ S.E.M.) number of positive cells (i.e., cells expressing increased levels of SGP-2 mRNA) per sampled field in caudate-putamen and the various layers of CA1 hippocampal field 1 and 7 days after 30 rain forebrain ischemia is given. The values are also given as percent of the respective cell population. In caudate-putamen, circular fields of 0.4 mm of diameter were sampled (2 measurements per animal, n = 4 animals /group) . Measurements were performed at 1 day in fields of the medial caudate-putamen (med) and at 7 days in fields of the lateral caudate-putamen (lat). In hippocampal formation, a rectangular field with a base of 0.4 mm and spanning the entire CA1 field was sampled (2 measurements per animal, n = 4 animals /group) . For further details, see text. Mean total number of cells per field: Caudate-putamen. Neurones: 1 day, 110.0_+ 9.5; 7 days, 21.6_+ 0.7. Non-neuronal cells: 1 day, 82.8 _+ 5.5; 7 days, 102.4-+ 2.4. CA1 field s tratum lacunosum-moleculare. Neurones: 1 day, 4.7_+0.2; 7 days, 5.2_+0.6. Non-neuronal cells: 1 day, 72.0-+ 1.6; 7 days, 102.6-+3.3. CA1 field stratum radiatum. Neurones: 1 day, 4.6 _+ 0.2; 7 days, 5.4 _+ 0.3. Non-neuronal cells: 1 day, 27.2 _+ 1.6; 7 days, 82.6 _+ 4.8. CA1 field s tratum pyramidale. Neurones: 1 day, 83.8 _+ 1.6; 7 days, 26.2-+2.7. Non-neuronal cells: 1 day, 17.6-+ 1.1; 7 days, 69.4_+2.7. CAI field stratum oriens. Neurones: 1 day, 5.6-+0.2; 7 days, 5.4+0.6. Non-neuronal cells: 1 day, 45.0-+ 1.8; 7 days, 62.2_+4.4.
Positive neuronal cells per field Positive non-neuronal cells per field
n % n %
A. Caudate-putamen 1 day (reed. part) 9.2_+ 1.4 8.2 14.4_+0.9 17.5 7 days (lat. part) 6.8_+0.9 31.2 5,8+ 1.0 5.7
B. Hippocampal field CA1 Stratum lacunosum-moleculare
1 day 0.9.+0.3 18.4 40.4+ 1.5 56.1 7 days 4.1 .+0.5 78.5 23.2_+3.1 22.6
Stratum radiatum 1 day 0.7_+0.2 15.6 13.0_+ 1.2 47.7 7 days 4.0 -+ 0.3 74.6 8.2 + 0.6 10.0
Stratum pyramidale 1 day 1.5 +_ 0.5 1.8 8.7 -+ 0.6 49.7 7 days 9.5 _+ 1.1 36.2 10.2 _+ 1.4 14.7
Stratum oriens 1 day 1.0.+0.3 17.9 19.7_+ 1.2 43.8 7 days 3.7 _+ 0.4 69.8 9.7 _+ 0.7 15.7

time interval, with around 30% of surviving neurones being positive and only around 6% of non-neuronal ceils (see Table II). At all time intervals analyzed, positive neurones were medium-sized. No clearly la- belled large neurones were observed, even at 7 days post-ischemia (Fig. 2B) in the area of cell loss where they constitute a relatively higher proportion of surviv- ing neuronal cells. Hippocampal formation. At all time intervals, many ceils showing increased SGP-2 labelling were observed in all subdivisions of the dorsal hippocampal forma- tion, being more concentrated in the CA1 field. In this region, both at 1 and 7 days post-lesion the labelling was higher in the stratum lacunosum-moleculare than in other layers (Fig. 3A,B).
A detailed analysis at cellular level was performed in the highly vulnerable CA1 region. In this region, at 1 day many non-neuronal ceils in the stratum lacunosum-moleculare were strongly positive (Fig. 4A). Positive non-neuronal cells were also present in the stratum oriens (Fig. 5A). In the stratum pyramidale, where at this time interval pyramidal neurones still constitute a continuous strip, positive non-neuronal cells were mostly localized on the border of the layer. In all layers of CA1 field, positive ceils represented around 50% of the overall non-neuronal cell popula- tion (see Table II). Scattered neurones also appeared to be positive (Figs. 4A and 5A), representing 15-20% of the total number of neurones in non-pyramidal cell layers, but only a very limited fraction in pyramidal layer (around 2%, see Table II). At 2 and 7 days, an increasing number of neurones appeared clearly posi- tive, paralleled by a decrease in the number of positive non-neuronal cells (see Table II). The increase in the density of positive neuronal cells was particularly prominent (around 70% of the respective overall popu- lation, see Table II) in interneuronal populations of strata lacunosum-moleculare, radiatum and oriens (Fig. 4B).
Partial mesodiencephalic hemitransection After hemitransection, SGP-2 mRNA showed
marked accumulation in several brain regions both rostral and caudal to the site of blade insertion. At 1 day, rostral to the lesion, SGP-2 transcript increases were observed in most cerebral regions of the lesioned hemisphere (see Table III and Figs. 6B, 7A and 8B). The ependyma of the third ventricle (Figs. 7A and 8B) and pia mater covering the cerebral hemisphere also showed increased labelling. In addition, a slight in- crease of SGP-2 mRNA was observed in the medial portion of the intact hemisphere at the levels analyzed. Interestingly, in the caudate-putamen the increased
167
T A B L E III
Regional distribution of SGP-2 mRNA labelling after partial mesodien- cephalic hemitransection
R e g i o n a l scores a r e re la t ive to S G P - 2 m R N A labe l l ing in the s a m e r e g i o n o f c o n t r o l an imals : - = n o i n c r e a s e d label l ing, _+ = weak ly i n c r e a s e d label l ing , + = m o d e r a t e l y i n c r e a s e d label l ing , + + = s t rong ly i n c r e a s e d label l ing , s.l.m. = s t r a t u m l a c u n o s u m - m o l e c u l a r e , s.o. = s t r a t u m or iens , s.p. = s t r a t u m p y r a m i d a l e , s.r. = s t r a t u m r a d i a - turn.
Cerebral region 1 day after 7 days after lesion lesion
A. R e g i o n s ros t r a l to the site o f b l a d e inse r t ion N e o c o r t e x
l ayer I + + layers I I - V + + l ayer VI + +
C i n g u l a t e co r t ex + _+ C o r p u s c a l l o s u m + + + S e p t u m + + C a u d a t e - p u t a m e n
s u b v e n t r i c u l a r + - subca l losa l _+ + + c e n t r a l + +
Nuc . a c c u m b e n s - - T u b e r c u l u m o l f a c t o r i u m -
B. R e g i o n s c a u d a l to t he site o f b l a d e in se r t i on N e o c o r t e x
l ayer I + + layers I I - V + + l ayer VI + +
C i n g u l a t e co r t ex - - C o r p u s c a l l o s u m + + D o r s a l h i p p o c a m p u s
C A 1 f ield s.o. + + C A 1 f ield s.p. + + C A 1 f ield s.r. + + C A 1 f ield s.l.m. + + + d e n t a t e gy rus + -
V e n t r a l h i p p o c a m p u s + + + D o r s a l m e s e n c e p h a l o n _+ + V e n t r a l m e s e n c e p h a l o n + + +
labelling was consistently observed in the subventricu- lar region, and more inconstantly in the overall striatal mass (Fig. 7A). Also caudal to the lesion increase in SGP-2 labelling was rather widespread in the lesioned side (see Table III and Figs. 9B and 10B). Labelling was particularly prominent in an ill-defined area in the ventrolateral mesencephalon (which includes the sub- stantia nigra, both pars compacta and pars reticulata (Fig. 10B), the pyramidal tract and an area dorsal to the substantia nigra), and the ventral hippocampus. On the intact side, a slight but significant increase in labelling was observed in the corpus callosum and in the dorsal hippocampus.
At 7 days, elevated SGP-2 mRNA labelling became more restricted to directly lesioned brain areas: either areas whose input and/or output connections were severed by the transection (such as lateral portion of caudate-putamen and substantia nigra pars compacta

and reticulata after sectioning of the lateral parts of Within caudate-putamen, positive cells were prefer-
the nigrostriatal and strionigral pathways) or areas
which were directly affected by the mechanical lesion
(such as cerebral cortex around the site of blade inser-
tion, lateral mesencephalon and ventral hippocampus).
In the latter case, the distribution of positive cells did
not respect the boundaries between anatomical areas
but rather followed the relatively variable localization
of the lesion.
entially distributed in the subcallosal region and almost
absent in the subventricular region (Fig. 7B). In the
ventral half of the brain, a high intensity of labelling
was observed in areas surrounding tissue loss in the
ventrolateral part of mesencephalon (Fig. lOC3, and in
adjacent hippocampal and limbic cortical regions.
At both days 1 and 7. the vast majority of well-de-
fined clusters of grains were observed to overlie non-
Fig. I. Dark-field microphotographs showing in situ hybridization with antisense SGP-2 RNA in dorsal caudate-putamen 1 (A) and 7 (B) days after 30’ forebrain ischemia. Note the redistribution of increased SGP-2 mRNA levels from outside to inside the highly lesioned lateral part of caudate-putamen between I and 7 days after the ischemic insult. Section thickness = 14 pm. Bregma level 1.2 mm according to Paxinos and
Watson atlas?‘. Bar = 500 pm. cc, corpus callosum.

169
Fig. 2. High-power bright-field microphotographs showing the hybridization signal for SGP-2 mRNA in the lesioned caudate-putamen 1 (A) and 7 (B) days after 30 min forebrain ischemia. At 1 day an area within the mildly lesioned medial caudate-putamen is shown, whereas at 7 days an area inside the highly lesioned lateral caudate-putamen is shown. Positive neurones and non-neuronal cells are indicated with an arrow or an arrowhead, respectively. At day 1, and to a lesser extent at 7 days, several neuronal and non-neuronal cells showing control levels of SGP-2
labelling are visible. Bar = 25 ~m.
neu rona l cells in all the areas with increased labelling.
Positive cells were part icularly dense in cortical layer I,
in s t ra tum lacunosum-molecu la re of h ippocampal field
CA1 and in dorsal corpus cal losum at 1 day. At day 7,
many positive cells were localized in tissue su r round ing
the areas of cell loss in the ventra l mesencepha lon .
High levels of labell ing were also presen t in the
ependymal cells of the lateral ventricles, the choroid
plexa and the pia mater .
At day 7 as opposed to day 1, some neurona l cells in
lesioned areas, such as the substant ia nigra, pars com-
pacta (Fig. l l B ) and reticulata, ventral tegmenta l area
(Fig. l l A ) , c auda t e -pu t amen (Fig. 12) and cerebral
cortex were overlain by discrete clusters of grains.
Fig. 3. Dark-field microphotographs showing in situ hybridization with antisense SGP-2 RNA in CA1 hippocampal field 1 (A) and 7 (B) days after 30 rain forebrain ischemia. Section thickness = 14 p,m. Bregma level -3.0 mm according to Paxinos and Watson atlas 37. Bar = 150 ~m.
DG, dentate gyrus; LM, stratum lacunosum-moleculare; OR, stratum oriens; P, stratum pyramidale; RA, stratum radiatum.

170
Fig. 4. High-power bright-field microphotographs showing the hy- bridization signal for SGP-2 mRNA in the stratum lacunosum- moleculare of CA1 hippocampal field 1 (A) and 7 (B) days after 30 rain forebrain ischemia. Positive neurones and non-neuronal cells are indicated with an arrow or an arrowhead, respectively. Bar = 25 #m.
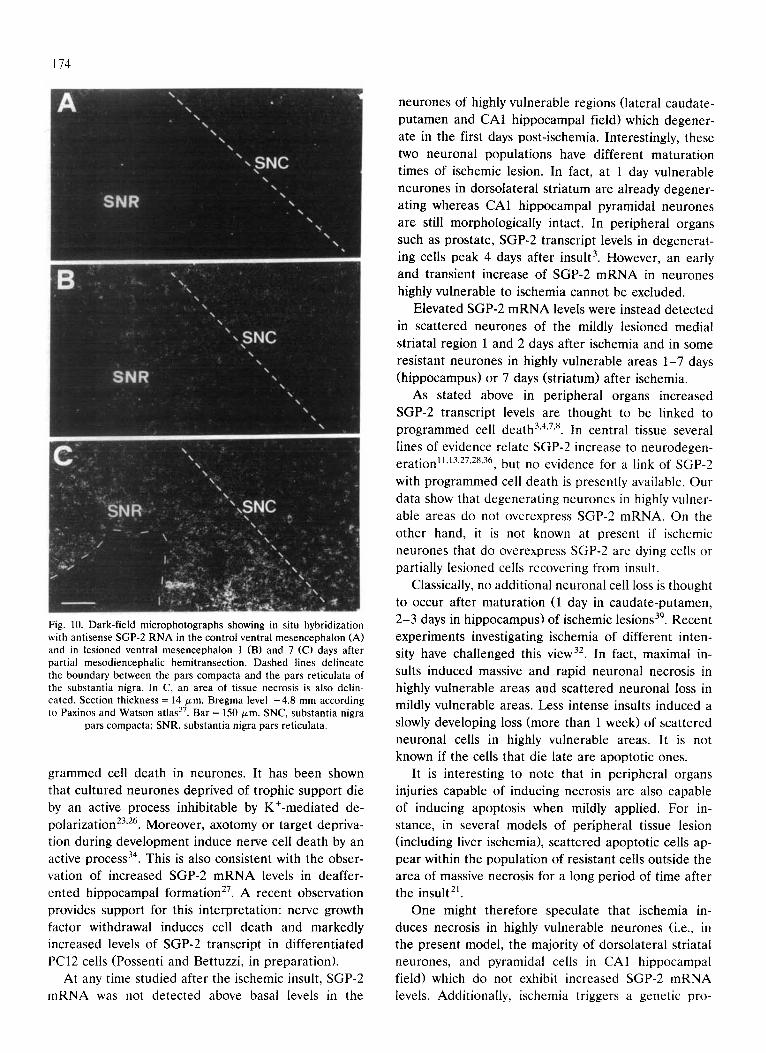
Based on the morphologic criterion of cytoplasmic Nissl staining, these cells could be unambiguously iden- tified as neuronal. In the caudate-putamen, positive neurones were preferentially localized in the lateral part, and in the substantia nigra they were localized in the periphery of the pars compacta and in the pars reticulata.
Manual counts of large neuronal cells in substantia nigra and ventral tegmental area showed that only a
few cells (around 5 cells/section of ventral mesen- cephalon) contained a high density of grains on the lesioned side 7 days after the operation. Computer-as- sisted grain counting performed on large nigral neu- rones showed that these cells contained almost three times more grains (143.3 _ 20.2 grains/cell , cell size = 417.3 + 76.0 /zm 2) than other nigral cells of the le- sioned side (55.0 _+ 4.4 grains/cell , cell size = 487.9 _+ 26.1 ~m2). These latter cells, in turn, were not signifi- cantly different from nigral cells of the lesioned side 1 day post-lesion, on the intact side 1 and 7 days post-le- sion, and in control animals.
D I S C U S S I O N
We have investigated the cellular and regional dis- tributions of SGP-2 m R N A in rat brain following two types of experimental Lesion. After partial mesodien- cephalic hemitransection, we observed a temporal pro- gression in the pattern of increased SGP-2 transcript levels from a relatively diffuse labelling in the lesioned and, to a lower extent, intact hemispheres to a more restricted distribution in directly lesioned areas. Simi- larly, after ischemic lesion, SGP-2 m R N A increase progressively shifts from least to most lesioned areas
(though it remains restricted to selectively vulnerable regions). This was particularly evident in striatum, where a redistribution of positive cells from medial
caudate-putamen and nuc. accumbens to lateral cau- date-putamen was observed.
Thus, in contrast with previous studies where ele- vated levels of SGP-2 m R N A or protein were re- stricted to lesioned areas L<24"27"36, we report a temporal
redistribution of positive ceils in both models exam- ined. The redistribution of cells exhibiting increased
Fig. 5. High-power bright-field microphotographs showing the hy- bridization signal for SGP-2 mRNA in the strata pyramidale and oriens of CA1 hippocampal field 1 (A) and 7 (B) days after 30 min forebrain ischemia. Positive neurones and non-neuronal cells are indicated with an arrow or an arrowhead, respectively. In the stratum pyramidale at 1 day pyramidal neurons show control levels of SGP-2
mRNA. Bar = 25 p.m.

171
SGP-2 mRNA levels from least to most lesioned areas after brain lesion may have been overlooked in previ- ous studies in which the time course of the lesion was not investigated. It must, however, be mentioned that both models studied here cause massive lesions of brain tissue and, perhaps more importantly, rupture of the hematoencephalic barrier.
Temporal shift in the distribution of non-neuronal cells overexpressing SGP-2 mRNA suggests an initial appearance of a reactive cell population followed by its migration towards the lesioned areas. Alternatively, a subsequent activation rather than (or in addition to) migration of positive cells can be hypothesized.
The identification of at least part of positive non- neuronal cells with migratory cells (phagocytic and other inflammatory cells) is indirectly supported by several lines of evidence. For instance, a temporal shift in the distribution of phagocytes (resident microglia and monocytes deriving from blood vessels, meninges and cerebral ventricles, refs. 6, 16) from the periphery of the lesion (1 day post-lesion) to the lesioned area (2-7 days post-lesion) was shown after penetrating wound lesions of rat cortex 16. Accordingly, after hemi- transection, positive non-neuronal cells appear initially in proximity of traffic routes to the brain, i.e., around the ventricles (in periventricular caudate-putamen, septum and corpus callosum) and the meninges (corti-
cal layer I), and subsequently (7 days post-lesion) in- side lesioned regions. In the case of ischemia, available evidence indicates that 7 days after the insult the area of neuronal necrosis, where we detected markedly ele- vated SGP-2 mRNA levels, is devoid of astrocytes but contain increased number of phagocytes 18'31'33'42 (and Grimaldi and Zoli, unpublished observations).
However, in the absence of counterstaining with specific astroglial and phagocytic markers, we have not unambiguously identified positive non-neuronal cells in lesioned cerebral tissues. Experiments are in progress to characterize non-neuronal cell populations express- ing SGP2 mRNA after ischemic and mechanical le- sions.
Other non-neuronal cells, e.g., choroid plexa, ven- tricular and pia mater ependymas, exhibit increased levels of SGP-2 mRNA in our study. It should be noted that these cells, as other proliferating epithelia (e.g., prostatic epithelium, refs. 3, 5) exhibit high levels of SGP-2 mRNA in the basal state ~5. Increased SGP-2 mRNA levels might tentatively be related to increased cell turnover induced by the lesion 3°.
Though the majority of ceils covered by discrete clusters of autoradiographic grains were non-neuronal, also some neuronal populations contained increased SGP-2 transcript levels after both experimental lesions. Previous reports of increased SGP-2 protein in neu-
Fig. 6. Dark-field microphotographs showing in situ hybridization with antisense SGP-2 R N A in the control cerebral cortex (A) and in lesioned cerebral cortex 1 (B) and 7 (C) days after partial mesodiencephalic hemitransection. Section thickness = 14 ttm. Bregma level 1.2 mm according
to Paxinos and Watson atlas 37. Note the increase of SGP-2 m R N A levels mainly in layer I at 1 day and more diffusely at 7 days. Bar = 150 ~m.


A C
173
Fig. 8. Dark-field microphotographs showing in situ hybridization with antisense SGP-2 RNA in the control dorsal septum/dorsomedial caudate-putamen (A) and in lesioned dorsal septum/dorsomedial caudate-putamen 1 (B) and 7 (C) days after partial mesodiencephalic hemitransection. Section thickness = 14 tzm. Bregma level 1.2 mm according to Paxinos and Watson atlas 37. Note the increase of SGP-2 mRNA
levels in subventricular caudate-putamen at 1 day, which is no longer present at 7 days. Bar = 150 ~m.
nections, such as caudate-putamen and substantia ni- gra, exhibit increased levels of SGP-2 mRNA (7 days post-lesion). These neurons may represent cells axo-
tomized or deafferented and hence deprived of trophic support. Several lines of evidence suggest that axotomy and/or deprivation of trophic support can induce pro-
C •
Fig. 9. Dark-field microphotographs showing in situ hybridization with antisense SGP-2 RNA in the control CA1 hippocampal field (A) and in lesioned CA1 hippocampal field 1 (B) and 7 (C) days after partial mesodiencephalic hemitransection. Dashed lines delineate the boundaries of
the pyramidal layer. Section thickness = 14 p.m. Bregma level - 4 . 8 mm according to Paxinos and Watson atlas 37. Bar = 50/.~m.
174
Fig. 10. Dark-field microphotographs showing in situ hybridization with antisense SGP-2 RNA in the control ventral mesencephalon (A) and in lesioned ventral mesencephalon 1 (B) and 7 (C) days after partial mesodiencephalic hemitransection. Dashed lines delineate the boundary between the pars compacta and the pars reticulata of the substantia nigra. In C, an area of tissue necrosis is also delin- eated. Section thickness = 14/zm. Bregma level -4.8 mm according to Paxinos and Watson atlas 37. Bar = 150 ~m. SNC, substantia nigra
pars compacta; SNR, substantia nigra pars reticulata.
grammed cell death in neurones. It has been shown that cultured neurones deprived of trophic support die by an active process inhibitable by K--media ted de- polarization 23'26. Moreover, axotomy or target depriva- tion during development induce nerve cell death by an active process 34. This is also consistent with the obser- vation of increased SGP-2 m R N A levels in deaffer- ented hippocampal formation 27. A recent observation
provides support for this interpretation: nerve growth factor withdrawal induces cell death and markedly increased levels of SGP-2 transcript in differentiated PC12 cells (Possenti and Bettuzzi, in preparation).
At any time studied after the ischemic insult, SGP-2 m R N A was not detected above basal levels in the
neurones of highly vulnerable regions (lateral caudate- putamen and CA1 hippocampal field) which degener- ate in the first days post-ischemia. Interestingly, these two neuronal populations have different maturat ion times of ischemic lesion. In fact, at 1 day vulnerable neurones in dorsolateral striatum are already degener- ating whereas CA1 hippocampal pyramidal neurones are still morphologically intact. In peripheral organs such as prostate, SGP-2 transcript levels in degenerat- ing cells peak 4 days after insult 3. However, an early and transient increase of SGP-2 m R N A in neurones highly vulnerable to ischemia cannot be excluded.
Elevated SGP-2 m R N A levels were instead detected in scattered neurones of the mildly lesioned medial striatal region 1 and 2 days after ischemia and in some resistant neurones in highly vulnerable areas 1-7 days (hippocampus) or 7 days (striatum) after ischemia.
As stated above in peripheral organs increased SGP-2 transcript levels are thought to be linked to programmed cell death 3'4'7's. In central tissue several
lines of evidence relate SGP-2 increase to neurodegen- eration 11'13'27'2~'36, but no evidence for a link of SGP-2
with programmed cell death is presently available. Our data show that degenerating neurones in highly vulner- able areas do not overexpress SGP-2 mRNA. On the
other hand, it is not known at present if ischemic neurones that do overexpress SGP-2 are dying cells or partially lesioned ceils recovering from insult.
Classically, no additional neuronal cell loss is thought to occur after maturation (1 day in caudate-putamen, 2-3 days in hippocampus) of ischemic lesions 39. Recent experiments investigating ischemia of different inten- sity have challenged this view 32. In fact, maximal in-
suits induced massive and rapid neuronal necrosis in highly vulnerable areas and scattered neuronal loss in mildly vulnerable areas. Less intense insults induced a slowly developing loss (more than 1 week) of scattered neuronal cells in highly vulnerable areas. It is not known if the cells that die late are apoptotic ones.
It is interesting to note that in peripheral organs injuries capable of inducing necrosis are also capable of inducing apoptosis when mildly applied. For in- stance, in several models of peripheral tissue lesion (including liver ischemia), scattered apoptotic cells ap- pear within the population of resistant cells outside the area of massive necrosis for a long period of time after the insult 21.
One might therefore speculate that ischemia in- duces necrosis in highly vulnerable neurones (i.e., in the present model, the majority of dorsolateral striatal neurones, and pyramidal cells in CA1 hippocampal field) which do not exhibit increased SGP-2 m R N A levels. Additionally, ischemia triggers a genetic pro-

175
Fig. 11. High-power bright-field microphotographs showing the hybridization signal for SGP-2 mRNA in the lesioned ventral tegmental area (A) and substantia nigra pars compacta (B) 7 days after partial mesodiencephalic hemitransection. A few positive neurones, indicated with an arrow,
are intermingled with several neurons showing control levels of SGP-2 labelling. Bar = 25/zm.
gram for cell death in some resistant a n d / o r mildly lesioned neurones (i.e., in the present model, ventro- medial striatal neurones, surviving neurones in dorso-
Fig. 12. High-power bright-field microphotographs showing the hy- bridization signal for SGP-2 mRNA in the lesioned caudate-putamen 1 (A) and 7 (B) days after partial mesodiencephalic hemitransection. Positive neurones and non-neuronal cells are indicated with an arrow or an arrowhead, respectively. At both 1 a n d 7 days, several neuronal and non-neuronal cells showing control levels of SGP-2 labelling are
visible. Bar = 25/~m.
lateral striatum, interneurones in CA1 hippocampal field), which exhibit increased levels of SGP-2 mRNA.
On the other hand, neurones with increased SGP-2 mRNA levels may be cells resistant to ischemic insult and synthesizing SGP-2 for purposes unrelated to pro- grammed cell death. There are in the literature at least two situations in central nervous system where in- creases in SGP-2 transcript levels and programmed cell death are not associated: SGP-2 is not expressed in neuronal populations during developmental cell death but rather after maturation is completed~5; after lesion, SGP-2 mRNA levels are markedly increased in astro- cytes, which are thought not to undergo degeneration but rather hypertrophy and proliferation 24'36. Sul-
phated glycoprotein-2 may then be a protein (or a family of homologous proteins) with multiple func- tional roles in different tissues and physiopathological conditions (for a thorough discussion, see ref. 15), whose function in brain degenerative states is unre- lated to programmed cell death s . Based on the pro- posed peripheral actions of SGP-2-1ike molecules, some speculations can be drawn about SGP-2 role in central tissue after lesion. In particular, SGP-2 may have a protective role against lesion-induced cytolytic agents a n d / o r be necessary for repair of damaged ceils. The former proposal is supported by the evidence that a human protein related to rat SGP-2 can inhibit com- plement-mediated cytolysis 2°'22. Sulphated glycopro-

176
tein-2 may contribute to neuronal cell repair processes by functioning as a lipid transport protein ~2. Interest- ingly, an increased expression of apolipoproteins has been observed in glial cells after injury of peripheral nerves 5.
Conclusions 1. Sulphated glycoprotein-2 mRNA levels are
markedly increased in heterogeneous populations of cells, including neuronal and non-neuronal cells, after both ischemic and mechanical lesions.
2. There is a temporal redistribution of positive cells from least to most lesioned areas, suggesting their identification with a population of migratory cells.
3. Sulphated glycoprotein-2 mRNA levels are not increased in highly vulnerable neurones which degen- erate in the first days after transient ischemia.
4. Sulphated glycoprotein-2 mRNA levels are in- creased in some neurones in mildly lesioned areas and in part of surviving neurons in highly lesioned areas after transient ischemia.
Acknowledgements. This paper has been supported by a MURST grant. We thank Dr. Joseph Addison Hill Jr. for careful reading and illuminating comments on the manuscript.
R E F E R E N C E S
I Agnati, L.F., Fuxe, K., Calza', L., Benfenati, F., Cavicchioli, L., Toffano, G. and Goldstein, M., Gangliosides increase the survival of lesioned nigral dopamine neurons and favour the recovery of dopaminergic synaptic function of rats by collateral sprouting, Acta Physiol. Scand., 119 (1983) 347-363.
2 Bettuzzi, S., Hiipakka, R.A., Gilna, P. and Liao, S., Identification of an androgen-repressed mRNA in rat ventral prostate as coding for sulphated glycoprotein 2 by cDNA cloning and sequence analysis, Biochem. J., 257 (1989) 293-296.
3 Bettuzzi, S., Troiano, L., Davalli, P., Tropea, F., Ingletti, M.C., Grassilli, E., Monti, D., Corti, A. and Franceschi, C., In vivo accumulation of sulfated glycoprotein-2 mRNA in rat thymocytes upon dexamethasone-induced cell death, Biochem. Biophys. Res. Commun., 175 (1991) 810 815.
4 Bettuzzi, S., Zoli, M., Ferraguti, F., Ingletti, M.C., Agnati, L.F. and Corti, A., Regional and cellular distribution within the rat prostate of two mRNA species undergoing opposite regulation by androgens, J. Endocrinol., 132 (1992) 361-367.
5 Boyles, J.K., Zoellner, C.D., Anderson, L.J., Kosik, L.M., Pitas, R.E., Weisgraber, K.H., Hui, D.Y., Mahley, R.W., Gebicke- Haerter, P.J., Ignatius, M.L. and Shooter, E.M., Apolipoprotein E associated with astrocytic glia of the central nervous system and with nonmyelinating gila of the peripheral nervous system, J. Clin. Ira,est., 83 (1989) 1015-1031.
6 Brierley, J.B. and Brown, A.W., The origin of lipid phagocytes in the central nervous system: I. The intrinsic microglia. Z Comp. Neurol., 211 (1982) 397-406.
7 Buttyan, R., Olsson, C.A., Pintar, J., Chang, C., Bandyk, M., Ng, P. and Sawezuk, t.S., Induction of TRPM-2 gene in cells udergo- ing programmed death, MoL Cell. Biol., 9 (1989) 3473-3481.
8 Buttyan, R., Genetic response of prostate cells to androgen deprivation: insights into the molecular mechanisms of apoptosis. In L.D Tomei and F.O. Cope (Eds.), Apoptosis. The Molecular Basis of Cell Death, Cold Spring Harbor Laboratory Press, New York, 1991, pp. 157-173.
9 Clarke, P.G.H., Developmental cell death: morphological diver- sity and multiple mechanisms, Anat. Embryol., 181 (1990) 195- 213.
10 Collard, M.W. and Griswold, M.D., Biosyntesis and molecular cloning of sulfated glycoprotein 2 secreted by rat Sertoli cells, Biochemistry, 26 (1987) 3297-3303.
11 Danik, M., Chabot, J.G., Mercier, C., Denabid, A.L. Chauvin, C., Quirion, R. and Suh, M., Human gliomas and epileptic foci express high levels of a mRNA related to rat testicular sulfated glycoprotein 2, a purported marker of cell death, Proc. Natl. Acad. Sci. USA 88 (1991) 8577-8581.
12 De Silva, H.V., Harmony, J.A.K., Stuart, W.D., Gil, C.M. and Robbins, J., Apolipoprotein J: structure and tissue distribution. Biochemisto', 29 (199(I) 5380-5389.
13 Duguid, J.R., Bohmont, C.W., Liu, N. and Tourtellotte, W.W., Changes in brain gene expression shared by scrapie and Alzheimer disease, Proc. Natl. Acad. Sci. USA, 86 (1989) 7260-7264.
14 Ferraguti, F., Zoli, M., Aronsson, M., Agnati, L.F.. Goldstein, M., Filer, D. and Fuxe, K., Distribution of glutamic acid decar- boxylase messenger RNA containing nerve cell populations of the male rat brain, J. Chem. Neuroanat., 3 (1990) 377-396.
15 Garden, G.A., Bothwell, M. and Rubel, E.W., Lack of correspon- dence between mRNA expression for a putative cell death molecule (SGP-2) and neuronal death in the central nervous system, J. Neurobiol., 22 (199l) 590-604.
16 Giulian, D., Chen, J., lngeman, J.E., George, J.K. and Noponen, M., The role of mononuclear phagocytes in wound healing after traumatic injury to adult mammalian brain, J. Neurosci., 9 (1989) 4116 4429.
17 Grima, J., Zwain, 1., Lockshin. R.A., Bardin, C.W. and Cheng, C.Y., Diverse secretory patterns of clusterin by epididymis and prostate/seminal vescicles undergoing cell regression after or- chidectomy, Endocrinology, 126 (1990) 2989-2997.
18 Grimaldi, R., Zoli, M., Agnati, L.F., Ferraguti, F., Fuxc, K.. Toffano, G. and Zini, 1., Effects of transient forebrain ischemia on peptidergic neurons and astroglial cells: evidence for recovery of peptide immunoreactivities in neocortex and striatum but not hippocampal formation, Exp. Brain Res., 82 (19911) 123-136.
19 Janson, A.M.. Fuxe, K., Agnati, L.F., Kitayama, 1., Harfstrand. A., Andersson, K. and Goldstein, M., Chronic nicotine treatment counteracts the disappearance of tyrosine-hydroxylase-im- munoreactive nerve cell bodies, dendrites and terminals in the mesostriatal dopamine system of the male rat after partial hemi- transection, Brain Res., 455 (1988) 332-345.
20 Jenne, D.E. and Tschopp, J., Molecular structure and functional characterization of a human complement cytolysis inhibitor found in blood and seminal plasma: identity with sulfated glycoprotein 2, a constituent of rat testis fluid, Proc. Natl. Aead. Sci. USA, 86 (1989) 7123-7127.
21 Kerr, J.F.R. and Harmon, B.V., Definition and incidence of apoptosis: an historical perspective. In L.D Tomei and F.O. Cope (Eds.), Apoptosis. The Molecular Basis of Cell Death, Cold Spring Harbor Laboratory Press, New York, 1991, pp. 5-29.
22 Kirszbaum, L., Sharpe, J.A., Murphy, B., D'Apice, A.J.F., Clas- son, B., Hudson, P. and Walker, I.D., Molecular cloning and characterization of the novel human complement-associated pro- tein, SP-40,40: a link between the complement and reproductive systems. EMBO J., 8 (1989) 711-718.
23 Koike, T., Martin, D.P. and Johnson, E.M. jr., Role of Ca 2~ channels on the ability of membrane depolarization to prevent death induced by trophic-factor deprivation: evidence that levels of internal Ca z + determine nerve growth factor dependence of sympathetic ganglion cells, Proc. Natl. Acad. Sci. USA, 86 (1989) 6421-6425.
24 Lampert-Etchells, M., McNeill, T.H., Laping, N.J., Zarow, C.. Finch, C.E. and May, P.C., Sulfated glycoprotein-2 is increased in rat hippocampus following entorhinal cortex lesioning, Brain Res., 563 (19911 101-106.
25 Leger, J.G., Montpetit, M.L. and Tenniswood, M.P., Characteri- zation and cloning of androgen-repressed mRNAs from rat ven- tral prostate, Biochem. Biophys. Res. Commun,, 147 (1987) 196- 2(/3.

26 Martin, D.P., Schmidt, R.E., DiStefano, P.S., Lowry, O.H., Carter, S.G. and Johnson, E.M. Jr., Inhibitors of protein synthesis and RNA synthesis prevent neuronal death caused by Nerve Growth Factor deprivation, J. Cell Biol., 106 (1988) 829-844.
27 May, P.C., Lampert-Etchells, M., Johnson, S.A., Poirier, J., Mas- ters, J.N. and Finch, C.E., Dynamics of gene expression for a hippocampal glycoprotein elevated in Alzheimer's disease and in response to experimental lesions in rat, Neuron, 5 (1990) 831-839.
28 May, P.C., Clemens, J., Fuson, K.S., Smalstig, E.B. and Robison, P., Transient global ischemia induces expression of sulfated gly- coprotein-2 (SGP-2) in rodent brain, Soc. Neurosci. Abstr. 17 (1991) 573.1.
29 Meldrum, B. and Garthwaite, J., Excitatory amino acid neurotox- icity and neurodegenerative diseases, Trends Pharmacol. Sci., 12 (Suppl.) (1991) 54-62.
30 Morshead, C.M. and van der Kooy, D., Postmitotic death is the fate of constitutively proliferating cells in the subependymal layer of the adult mouse brain, J. Neurosci., 12 (1992) 249-256.
31 Myers, R., Manjil, L.G., Cullen, B.M., Price, G.W., Frackowiack, R.S. and Cremer, J.E., Macrophage and astrocyte populations in relation to 3H-PKlI195 binding in rat caudate-putamen and cortex following a local ischemic lesion, J. Cereb. Blood Flow Metab., 11 (1991) 314-322.
32 Nakano, S., Kogure, K. and Fujikura, H., Ischemia-induced slowly progressive neuronal damage in the rat brain, Neuroscience, 38 (1990) 115-124.
33 Nemeth, G., Cintra, A., Herb, J.-M., Ding, A., Goldstein, M., Agnati, L.F., Hoyer, S. and Fuxe, K., Changes in striatal dopamine neurochemistry and biochemistry after incomplete cerebral is- chemia in the rat, Exp. Brain Res., 86 (1991) 545-554.
34 Oppenheim, R.W., Prevette, D., Tytell, M. and Homma, S., Naturally occurring and induced neuronal death in the chick embryo in vivo requires protein and RNA synthesis: evidence for the role of cell death genes, Det,. Biol., 138 (1990) 104-113.
177
35 Palmer, D.J. and Christie, D.L., The primary structure of glyco- protein III from bovine adrenal medullary chromaffin granules, J. Biol. Chem., 265 (1990) 6617-6623.
36 Pasinetti, G.M. and Finch, C.E., Sulfated glycoprotein-2 (SGP-2) mRNA is expressed in rat striatal astrocytes following ibotenic acid lesions, Neurosci. Lett., 130 (1991) 1-4.
37 Paxinos, G. and Watson, C., The Rat Brain in Stereotaxic Coordi- nates, Academic Press, New York, 1982.
38 Pulsinelli, W.A. and Brierley, J.B., A new model of bilateral hemispheric ischemia in the unanesthetized rat, Stroke, 10 (1979) 267-272.
39 Pulsinelli, W.A., Brierley, J.B. and Plum, F., Temporal profile of neuronal damage in a model of transient forebrain ischemia, Ann. Neurol., 11 (1982) 491-498.
40 Server, A.C. and Mobley, W.C., Neuronal cell death and the role of apoptosis. In L.D Tomei and F.O. Cope (Eds.), Apoptosis. The Molecular Basis of Cell Death, Cold Spring Harbor Laboratory Press, New York, 1991, pp. 263-278.
41 Wyllie, A.H., Kerr, J.F.R. and Currie, A.R., Cell death: the significance of apoptosis, Int. Reu. Cytol., 68 (1980) 251-306.
42 Zini, I., Grimaldi, R., Merlo Pich, E., Zoli, M., Fuxe, K. and Agnati, L.F., Aspects of neural plasticity in the central nervous system - - V. Studies on a model of transient forebrain ischemia in male Sprague-Dawley rats, Neurochem. Int., 16 (1990) 451-468.
43 Zoli, M., Zini, I., Agnati, L.F., Guidolin, D., Ferraguti, F. and Fuxe, K., Aspects of neural plasticity in the central nervous system. I. Computer-assisted image analysis methods, Neu- rochem. Int., 16 (1990) 383-418.
44 Zoli, M., Bettuzzi, S., Ferraguti, F., Ingletti, M.C., Zini, 1., Fuxe, K., Agnati, L.F. and Corti, A., Regional increases in ornithine decarboxylase mRNA levels in the rat brain after partial mesodi- encephalic hemitransection as revealed by in situ hybridization histochemistry, Neurochem. Int., 18 (1990)347-352.




















