
Forebrain mineralocorticoid receptor overexpression enhances memory, reduces anxiety and attenuates...
11
Forebrain mineralocorticoid receptor overexpression enhances memory, reduces anxiety and attenuates neuronal loss in cerebral ischaemia Maggie Lai, 1 Karen Horsburgh, 2 Sung-Eun Bae, 1 Roderick N. Carter, 1 Dirk J. Stenvers, 1 Jill H. Fowler, 2 Joyce L. Yau, 1 Celso E. Gomez-Sanchez, 3 Megan C. Holmes, 1 Christopher J. Kenyon, 1 Jonathan R. Seckl 1 and Malcolm R. Macleod 1 1 Endocrinology Unit, Centre for Cardiovascular Science, University of Edinburgh, Queen’s Medical Research Institute, 47 Little France Crescent, Edinburgh EH16 4TJ, UK 2 Centre for Neuroscience Research, University of Edinburgh, 1, George Square, Edinburgh EH8 9LS, UK 3 Division of Endocrinology, University of Mississippi Medical Center, 2500 North State Street, Jackson, Mississippi 39216, USA Keywords: behaviour, mouse, neuroprotection, stress, transgenic Abstract The nuclear mineralocorticoid receptor (MR), a high-affinity receptor for glucocorticoids, is highly expressed in the hippocampus where it underpins cognitive, behavioural and neuroendocrine regulation. Increased neuronal MR expression occurs early in the response to cellular injury in vivo and in vitro and is associated with enhanced neuronal survival. To determine whether increased neuronal MR might be causal in protecting against ischaemic damage in vivo we generated a forebrain-specific MR-overexpressing transgenic mouse (MR-Tg) under the control of the CamKII alpha promoter, and subjected mice to transient cerebral global ischaemia induced by bilateral common carotid artery occlusion for 20 min. We also separately assessed the effects of MR overexpression on hypothalamic–pituitary–adrenal (HPA) axis activity and cognitive and affective functions in noninjured animals. Our results showed that MR-Tg mice had significantly reduced neuronal death following transient cerebral global ischaemia compared to wild-type littermates. This effect was not associated with alterations in basal or poststress HPA axis function or in arterial blood pressure. MR-Tg mice also demonstrated improved spatial memory retention, reduced anxiety and altered behavioural response to novelty. The induction of neuronal MR appears to offer a protective response which has potential therapeutic implications in cerebral ischaemia and cognitive and affective disorders. Introduction Glucocorticoid hormones exert a broad range of actions in the brain including autoregulation of the stress response via feedback upon the hypothalamic–pituitary–adrenal (HPA) axis, as well as effects on mood, learning and memory (de Kloet et al., 1998). Importantly, glucocorticoids also have a profound impact on both the survival and death of neurons (Abraham et al., 2001). Their actions are mediated by two related intracellular receptors: the high-affinity mineralocorticoid receptor (MR) and the lower affinity glucocorticoid receptor (GR), both of which act as ligand-dependent transcription factors to regulate the expression of target genes. GRs are widely expressed in most neurons and glia whereas MRs are largely restricted to neurons of the limbic system with predominant expression in the hippocampus (Reul & de Kloet, 1985). Chronic overactivation of GR by consistently high levels of circulating glucocorticoid endangers (primarily hippocampal) neurons, making them vulnerable to neurotoxic insults (Reagan & McEwen, 1997; Sapolsky, 2000), disturbing neuroendocrine control and impairing cognition and behaviour. In contrast the consequences of increased MR activation in the CNS have yet to be explored. Current evidence suggests that predominant MR activation, as opposed to concomitant activation of MR and GR together, can trigger distinct and even opposite responses in, for example, neuronal excitability (Joels, 2001) and cognitive function (de Kloet et al., 1999). We recently showed that cellular stress rapidly increases MR mRNA and protein (but not GR) in rat primary hippocampal and, more surprisingly, cortical cultures. This induction of MR is protective because blocking the MR with spironolactone promotes cell death (Macleod et al., 2003). Increased MR expression was also observed in vivo in rat hippocampus following transient global ischaemia under hypothermic conditions and, similarly, spironolactone administration promoted neuronal death (Macleod et al., 2003). Furthermore, MR mRNA levels are higher in human post-mortem hippocampal neurons in patients who suffered transient global ischaemia due to cardiac arrest in the weeks leading up to their death (S. E. Bae, M. Lai and M. R. Macleod, unpublished observations). Similarly, a shift in the balance presumably towards MR activation following GR blockade also protects neurons from ischaemic insult (Krugers et al., 2000). Thus increased MR may represent an endogenous survival response to neuronal injury. However, while elevated MR levels are associated with increased neuronal survival, causality has yet to be established. In this study we have generated a forebrain-specific MR-over- expressing transgenic mouse model (MR-Tg) to address the hypothe- sis that increased neuronal MR expression protects against damage caused by transient global cerebral ischaemia. We have also exploited this genetic approach of increased MR function to further examine the Correspondence: Dr Maggie Lai, as above. E-mail: [email protected] Received 17 October 2006, revised 14 December 2006, accepted 18 January 2007 European Journal of Neuroscience, Vol. 25, pp. 1832–1842, 2007 doi:10.1111/j.1460-9568.2007.05427.x ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing Ltd
Transcript of Forebrain mineralocorticoid receptor overexpression enhances memory, reduces anxiety and attenuates...

Forebrain mineralocorticoid receptor overexpressionenhances memory, reduces anxiety and attenuatesneuronal loss in cerebral ischaemia
Maggie Lai,1 Karen Horsburgh,2 Sung-Eun Bae,1 Roderick N. Carter,1 Dirk J. Stenvers,1 Jill H. Fowler,2 Joyce L. Yau,1
Celso E. Gomez-Sanchez,3 Megan C. Holmes,1 Christopher J. Kenyon,1 Jonathan R. Seckl1 and Malcolm R. Macleod1
1Endocrinology Unit, Centre for Cardiovascular Science, University of Edinburgh, Queen’s Medical Research Institute, 47 LittleFrance Crescent, Edinburgh EH16 4TJ, UK2Centre for Neuroscience Research, University of Edinburgh, 1, George Square, Edinburgh EH8 9LS, UK3Division of Endocrinology, University of Mississippi Medical Center, 2500 North State Street, Jackson, Mississippi 39216, USA
Keywords: behaviour, mouse, neuroprotection, stress, transgenic
Abstract
The nuclear mineralocorticoid receptor (MR), a high-affinity receptor for glucocorticoids, is highly expressed in the hippocampuswhere it underpins cognitive, behavioural and neuroendocrine regulation. Increased neuronal MR expression occurs early in theresponse to cellular injury in vivo and in vitro and is associated with enhanced neuronal survival. To determine whether increasedneuronal MR might be causal in protecting against ischaemic damage in vivo we generated a forebrain-specific MR-overexpressingtransgenic mouse (MR-Tg) under the control of the CamKII alpha promoter, and subjected mice to transient cerebral globalischaemia induced by bilateral common carotid artery occlusion for 20 min. We also separately assessed the effects of MRoverexpression on hypothalamic–pituitary–adrenal (HPA) axis activity and cognitive and affective functions in noninjured animals.Our results showed that MR-Tg mice had significantly reduced neuronal death following transient cerebral global ischaemiacompared to wild-type littermates. This effect was not associated with alterations in basal or poststress HPA axis function or in arterialblood pressure. MR-Tg mice also demonstrated improved spatial memory retention, reduced anxiety and altered behaviouralresponse to novelty. The induction of neuronal MR appears to offer a protective response which has potential therapeutic implicationsin cerebral ischaemia and cognitive and affective disorders.
Introduction
Glucocorticoid hormones exert a broad range of actions in the brainincluding autoregulation of the stress response via feedback upon thehypothalamic–pituitary–adrenal (HPA) axis, as well as effects onmood, learning and memory (de Kloet et al., 1998). Importantly,glucocorticoids also have a profound impact on both the survival anddeath of neurons (Abraham et al., 2001). Their actions are mediated bytwo related intracellular receptors: the high-affinity mineralocorticoidreceptor (MR) and the lower affinity glucocorticoid receptor (GR),both of which act as ligand-dependent transcription factors to regulatethe expression of target genes. GRs are widely expressed in mostneurons and glia whereas MRs are largely restricted to neurons of thelimbic system with predominant expression in the hippocampus (Reul& de Kloet, 1985). Chronic overactivation of GR by consistently highlevels of circulating glucocorticoid endangers (primarily hippocampal)neurons, making them vulnerable to neurotoxic insults (Reagan &McEwen, 1997; Sapolsky, 2000), disturbing neuroendocrine controland impairing cognition and behaviour. In contrast the consequencesof increased MR activation in the CNS have yet to be explored.Current evidence suggests that predominant MR activation, asopposed to concomitant activation of MR and GR together, can
trigger distinct and even opposite responses in, for example, neuronalexcitability (Joels, 2001) and cognitive function (de Kloet et al.,1999).We recently showed that cellular stress rapidly increases MR
mRNA and protein (but not GR) in rat primary hippocampal and,more surprisingly, cortical cultures. This induction of MR is protectivebecause blocking the MR with spironolactone promotes cell death(Macleod et al., 2003). Increased MR expression was also observedin vivo in rat hippocampus following transient global ischaemia underhypothermic conditions and, similarly, spironolactone administrationpromoted neuronal death (Macleod et al., 2003). Furthermore, MRmRNA levels are higher in human post-mortem hippocampal neuronsin patients who suffered transient global ischaemia due to cardiacarrest in the weeks leading up to their death (S. E. Bae, M. Lai and M.R. Macleod, unpublished observations). Similarly, a shift in thebalance presumably towards MR activation following GR blockadealso protects neurons from ischaemic insult (Krugers et al., 2000).Thus increased MR may represent an endogenous survival response toneuronal injury. However, while elevated MR levels are associatedwith increased neuronal survival, causality has yet to be established.In this study we have generated a forebrain-specific MR-over-
expressing transgenic mouse model (MR-Tg) to address the hypothe-sis that increased neuronal MR expression protects against damagecaused by transient global cerebral ischaemia. We have also exploitedthis genetic approach of increased MR function to further examine the
Correspondence: Dr Maggie Lai, as above.E-mail: [email protected]
Received 17 October 2006, revised 14 December 2006, accepted 18 January 2007
European Journal of Neuroscience, Vol. 25, pp. 1832–1842, 2007 doi:10.1111/j.1460-9568.2007.05427.x
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing Ltd

role of MR in HPA axis regulation, cognition and behaviour, studiesthus far constrained to investigations by pharmacological means.
Materials and methods
Generation of MR transgenic mice
A haemagglutinin (HA) epitope tag was inserted in-frame into the N-terminus of the full-length human MR cDNA (Arriza et al., 1987). TheHA-MR cDNAwas subcloned into pNN265 (Mayford et al., 1996) tocreate pNN-HA-MR. Using this plasmid, expression of functional MRwas verified by measurement of 3H aldosterone (GE Healthcare UKLtd, Little Chalfont, UK) binding in transfected rat B103 cells.Nonspecific binding was measured by addition of a 500-fold excess ofcold aldosterone. Cell lysates underwent rapid filtration onto filtermatspresoaked in polyethylenimine (0.3%) and retained radioactivity wasdetermined by liquid scintillation counting (Perkin-Elmer, Beacons-field, UK). A Not I fragment containing the HA-MR cDNA flanked by5¢ ⁄ 3¢ introns and simian virus 40 polyadenylation signal was insertedinto pMM403 immediately downstream of the mouse 8.5-kb calcium–calmodulin-dependent protein kinase IIa (CaMKIIa) promoter (May-ford et al., 1996). Transgenic mice were generated by pronuclearinjection of the 12.5-kb CaMKIIa-HA-MR transgenic construct intoC57BL ⁄ 6J · CBA embryos (Babraham Institute, Cambridge, UK).Ten founder mice were generated and mouse lines were established bybreeding founders and their progeny to C57BL ⁄ 6J mice.
Genotyping
Animals positive for the transgene were identified by PCR of isolatedtail DNA using the primer pair 5¢-ATG TAC CCA TAC GAC GTCCC-3¢ at position 1 of the ATG in the HA epitope tag and 5¢-CACAGG ACT AGA ACA CCT GC-3¢ at position +294 of human MRcDNA, multiplexed with mouse hypoxanthine phosphoribosyl-transf-erase (HPRT) as a positive control for the quality of the genomic DNAsample using primer pair 5¢-GCT GGT GAA AAG GAC CTC T-3¢and 5¢-CAC AGG ACT AGA ACA CCT GC-3¢. Transgenic micewere further identified by Southern blot analysis of Afl II-digestedgenomic DNA. Blots were hybridised with a 32P-labelled DNAfragment comprising the full length transgene. The copy number of theintegrated transgene was estimated based on the comparison ofintensity of radioactive signal from nontransgenic genomic DNAspiked with known quantities of transgene.
Transgenic mice were maintained as hemizygotes. All mousemanipulations were conducted in strict accordance with the guidelinesof the UK Home Office Animals (Scientific Procedures) Act, 1986.Animals were maintained under conditions of controlled lighting(lights on 07.00–19.00 h) and temperature (22 �C) with free access tochow and water. All experiments were performed blind to genotype onmale mice using littermate wild-type mice as controls from F3generation at 3–6 months of age.
MR expression analyses
Coronal sections (8 lm) of fresh-frozen brains were processed forin situ hybridization using 35S-UTP (GE Healthcare)-labelled RNAprobes complementary to mRNA for human MR (Arriza et al., 1987)or rat GR (Miesfeld et al., 1986) at an annealing temperature of 50 �Cfor 16 h. Sections were washed, dehydrated and exposed toautoradiographic film for up to 5 days. Densitometric analyses ofautoradiographs were carried out using an MCID image analysissystem (MCID Research Imaging, Canada).
MR protein was detected by immunohistochemical staining ofparaffin-embedded brain sections (8 lm) using a mouse monoclonalantirat MR antibody (rMR-6G1) directed against amino acids 1–18 ofthe rat MR (Gomez-Sanchez et al., 2006) at 1 : 200 dilution followedby incubation with biotin-conjugated donkey antimouse IgG (1 : 1000dilution; Jackson ImmunoResearch, West Grove, PA, USA). Sectionswere processed using the Vector Elite Kit (Vector Laboratories,Peterborough, UK) and MR immunoreactivity was visualized usingdiaminobenzidine (Vector Laboratories).
Corticosterone measurement
For basal measurement, blood samples were collected by tail nick at08.00 h. In a separate cohort of animals, HPA axis response to acutestress was measured by collecting blood samples following decapit-ation immediately or 50 min after a 10-min period in a restraint tube.This procedure was carried out between 08.00 and 11.00 h. Plasmacorticosterone in duplicate was analysed using an in-house specificradioimmunoassay as described previously (Holmes et al., 1997) andmodified for microtitre plate scintillation proximity assay (GEHealthcare) with inter- and intra-assay variations of < 10%. At thetime of killing, adrenals, kidneys, thymus and heart were dissected,cleaned of fat and weighed.
Blood pressure measurement
Systolic blood pressure was measured by an automated tail-cuffplethysmography method as previously described (Evans et al., 1994).
Transient global ischaemia
Mice used for this experiment were all between the ages of 3 and4 months. Mice were initially anaesthetized with 3% halothane in 70%N2O and 30% O2, and maintained on 1.5% halothane during surgery.Transient global ischaemia was induced by bilateral common carotidartery occlusion for 20 min as previously described (Kelly et al.,2001). Body temperature was maintained at 37 �C throughout andanimals were allowed to recover in a thermally controlled incubatorfor 2 h before returning to their home cages. Mice were anaesthetizedunder halothane and killed 72 h postischaemia by perfusion-fixationwith 4% paraformaldehyde. Brains were dissected and prepared forparaffin embedment. Sections (6 lm) were cut and stained withhaematoxylin and eosin. The number of morphologically normalneurons and neurons showing features of ischaemic damage, definedby intense haematoxylin-stained pyknotic nuclei surrounded byeosinophilic cytoplasm, were counted in five randomly selected fieldswithin the CA-1 and CA-2 regions of the hippocampus and in thecaudate nucleus of each brain hemisphere using a 1-cm2 grid at ·400magnification by an investigator blinded to genotype and treatment.The percentage of ischaemic neurons was calculated for each areaof interest.
Behavioural testing
All behavioural tests were performed during the light periodbetween 08.00 and 13.00 h. Animals were singly housed at least72 h prior to testing and on the test day were acclimatised to theexperimental room for at least 30 min before each test. Allbehavioural tests were recorded using the Limelight� video trackingsystem and Actimetrics software (Actimetrics Inc., IL, USA) toenable full analysis.
Increased mineralocorticoid receptor is protective 1833
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 25, 1832–1842

Open-field test
Exploratory behaviour was assessed as previously described (Holmeset al., 2006) by placing animals individually into an open-field box(60 · 60 · 30 cm) marked off into 25 equal squares. The outersquares forming the perimeter of the field are considered lessanxiogenic than the squares in the inner zone. The number ofcrossings, time spent and overall distance travelled (movement of allfour limbs into a new square) into each new square was measured overa 5-min period. Total movement was considered a reflection of generalactivity whilst relative movement in the inner zone of the boxcorrelated with the anxiety state of the animal.
Light–dark exploration test
Anxiety-related behaviour was also assessed in the light–darkexploration test using a two-compartment box (40 · 9.5 · 18 cm;Ugo Basile Biological Research, Comerio, VA, Italy), one light(considered to be a more anxiogenic environment) and one dark ofequal size, separated by a wall with a sliding door. Mice were placedin the dark compartment and allowed to acclimatise for 90 s. Thesliding door was opened and latency to enter the light compartment,the subsequent total time spent in each compartment and the totalnumber of entries into each compartment in a 5-min trial wasrecorded.
Fig. 1. Generation and identification of MR-Tg mice. (a) Diagram showing the 12.5-kb transgenic construct used for microinjection. Afl II restriction site relevantfor Southern analysis is indicated. (b) Ligand binding capacity of PC12 cells overexpressing MR after transfection with pNN-HA-MR transgenic vector or emptypNN265 vector and incubation of cells with 5 nm
3[H] aldosterone in the presence or absence of excess cold label (2.5 lm). Extent of ligand binding is expressed asmean ± SEM dpm ⁄ mg protein; n ¼ 6 independent experiments. (c) Southern blot of genomic DNA from heterozygous MR-Tg mice (Lanes 1–4). Numbers abovethe lanes represent the ID of a particular founder from which the transgenic lines were established. Lane 5, negative DNA from nontransgenic littermate; Lane 6,negative DNA spiked with two copies of transgene; Lane 7, negative DNA spiked with 20 copies of transgene. (d) In situ hybridization analysis of MR mRNAexpression in wild-type (WT) and transgenic (MR-Tg) mouse forebrain. Representative autoradiographic images show sections taken from bregma +0.86 to)5.80 mm. LS, lateral septum; St, striatum; Hi, hippocampus; Cx, cortex; Am, amygdala; VHi, ventral hippocampus; Ce, cerebellum; S, sense strand riboprobeshown at the hippocampal level.
1834 M. Lai et al.
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 25, 1832–1842
Novel object recognition task
In another test of exploratory behaviour, mice were placed into theopen-field box containing two identical objects and allowed toexplore for 5 min. The time spent sniffing, touching and orienting toeach object was recorded. After a 1-h delay, one object (A) wasremoved and replaced with a new object (B) and the time spent ateach object was recorded over 5 min. Similarly, after a 3-h and 24-hdelay one familiar object was removed and replaced by a new object(C, 3-h; D, 24-h) and the time spent at each object was againrecorded over 5 min. The familiar object remained in the samecorner throughout all test sessions. Exploration of each object wasscored when the animal’s nose was within 2 cm of the object orwhen it reared against the object. The investigator was blinded toboth treatment and object novelty. Object type and location werefully randomized between the two groups. The objects used were atextured translucent glass, a light bulb stuck to a piece of cardboard,a green synthetic pear stuck to a piece of cork, and a blueearthenware cup placed upside down.
Water-maze test
Behaviour associated with learning and memory was tested byassessing the performance of MR-Tg and wild-type control mice inthe Morris water maze. Details of the water-maze experiments havebeen described elsewhere (Yau et al., 2001) with minor modifica-tions made. Briefly, mice were given four training trials per day for5 days, from a random start location with a hidden platform in afixed location. For each training trial, mice had a maximum of 90 sswim time to mount the platform and were guided there manually if90 s elapsed. Latency to escape from the water was considered ameasure of learning ability. At the end of day 5, mice were given a90-s probe trial in which the platform had been removed and the
time and direction of swim search was considered to reflect memoryretention.
Data analysis
Data were expressed as group means ± SEM. Statistical analyses wereperformed using one- or two-way anova followed where appropriatewith a post hoc Tukey’s test. For the novel object recognition test andwater-maze test, data were analysed using repeated-measures anova,and for open-field and light–dark exploration tests the nonparametricMann–Whitney U-test was used. Values were considered statisticallysignificant at P < 0.05.
Results
Generation of mice overexpressing MR in the forebrain
Transgenic mice overexpressing MR (MR-Tg) were generated usingthe previously characterized CamKIIa promoter to direct expression ofHA-tagged human MR cDNA specifically to the forebrain. Additionof the HA epitope tag did not affect the ligand-binding properties ofMR as B103 cells transfected with HA-MR under the control of aCMV promoter exhibited high levels of [3H] aldosterone binding thatwas displaced by a 500-fold excess cold ligand; B103 cells transfectedwith control vector did not produce detectable [3H] aldosteronebinding (Fig. 1b). Pronuclear injection of purified transgenic constructinto fertilized mouse ova (C57BL ⁄ 6J · CBA) resulted in 10 trans-genic founders from 44 mice identified by PCR analysis of genomicDNA. All 10 founders were bred with wild-type C57BL ⁄ 6J mice;Southern blot and PCR analysis of genomic DNA from the progenydemonstrated that four out of the 10 founders transmitted the transgenein various copies (Fig. 1c) and these were used to establishindependent lines. Breeding of transgenic F1–F3 with wild-type
1-AC1-AC
2-AC 2-AC
3-AC3-AC
GDGD
SLSL
xCxC
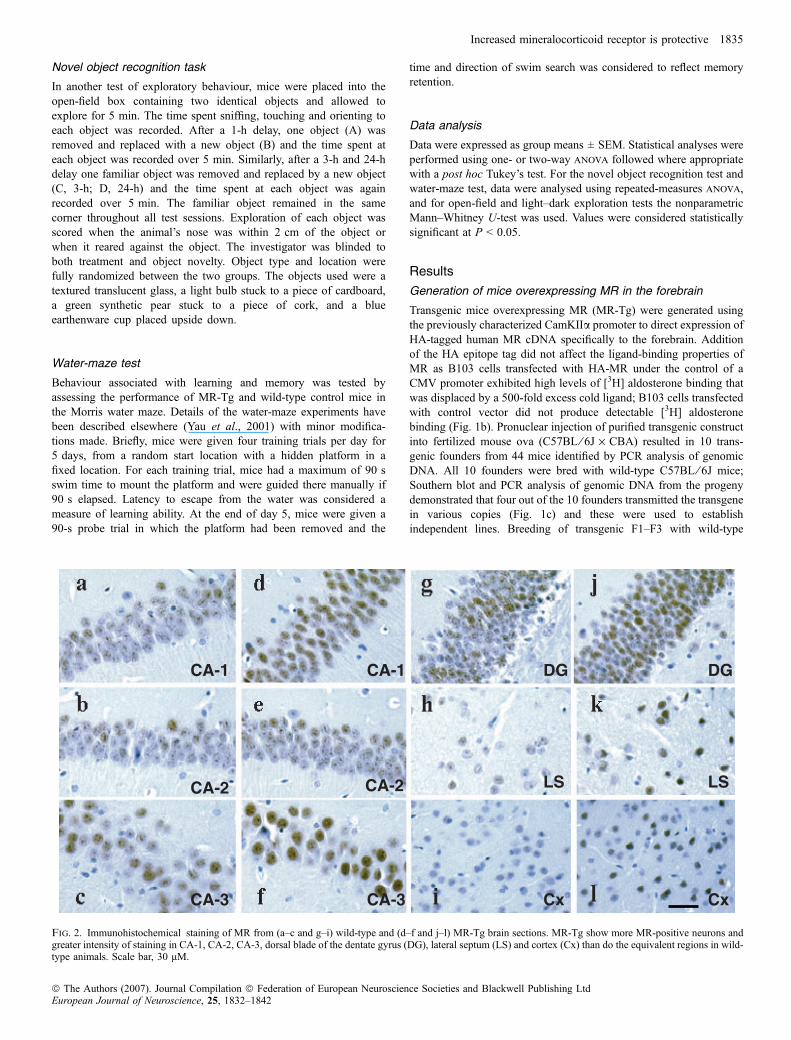
Fig. 2. Immunohistochemical staining of MR from (a–c and g–i) wild-type and (d–f and j–l) MR-Tg brain sections. MR-Tg show more MR-positive neurons andgreater intensity of staining in CA-1, CA-2, CA-3, dorsal blade of the dentate gyrus (DG), lateral septum (LS) and cortex (Cx) than do the equivalent regions in wild-type animals. Scale bar, 30 lM.
Increased mineralocorticoid receptor is protective 1835
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 25, 1832–1842

C57BL ⁄ 6J resulted in the expected Mendelian ratio of offspring(MR-Tg, 45 ⁄ 89; WT, 44 ⁄ 89). Heterozygous MR-Tg mice wereviable, fertile and physically indistinguishable from wild-types,showing normal weight gain and development (Table 1). Gross brainmorphology appeared normal in MR-Tg.
Localization of transgene mRNA and protein expression
In situ hybridization analyses using a human MR cRNA proberevealed strong hybridization signals in the hippocampus, cortex,striatum, lateral septum and basal lateral nucleus of amygdala(Fig. 1d, Table 1) in three lines; the fourth line showed much lowertransgene mRNA levels and was not studied further. As expected,endogenous MR mRNA was also detected by the human cRNAprobe in wild-type controls and was largely restricted to thehippocampus with undetectable levels in the cortex and striatum(Fig. 1d). Typically with this promoter, MR-Tg was not detected incerebellum, hypothalamus or peripheral tissues such as liver, lungand muscle in any line (data not shown). Densitometric analyses ofautoradiographs showed that, within the hippocampal region ofMR-Tg mice, CA-2 neurons showed the greatest increase in MRexpression (Table 1). MR protein was expressed in forebrain asdetected by immunohistochemistry; increased MR was observed inthe CA-1 (Fig. 2d), CA-2 (Fig. 2e), CA-3 (Fig. 2f) and dentategyrus (DG; Fig. 2j) regions of the hippocampus in MR-Tg micecompared with control mice (CA-1, Fig. 2a; CA-2, Fig. 2b; CA-3,Fig. 2c; DG, Fig. 2g). More strikingly in MR-Tg mice, neurons inthe lateral septum (Fig. 2k) and cortex (Fig. 2l) were stronglyimmunoreactive for MR, whereas in control mice MR protein wasbarely detectable in these regions (lateral septum, Fig. 2h; cortex,Fig. 2i). All three high-expressing lines were systematicallycharacterized and showed similar HPA axis and behaviouralphenotypes, suggesting that these effects related to the expressionof the transgene rather than the site of transgene integration. Line5207 was used for further investigation with nontransgeniclittermates used as controls.
MR-Tg displayed normal HPA axis activity
Previous pharmacological studies have implicated hippocampal MR inthe control of both basal and poststress corticosterone levels (VanHaarst et al., 1997; Spencer et al., 1998). Altered HPA axis activitymight impact on the response to cerebral ischaemia through activationof endogenous MR or GR. However, we found no difference in basalcorticosterone levels (Fig. 3a) between wild types and their MR-Tglittermates (WT, 35.2 ± 6.2 nmol ⁄ L; MR-Tg, 33.9 ± 2.6 nmol ⁄ L;n ¼ 8). Similarly, there was no difference between MR-Tg andcontrols in the plasma corticosterone response to 10-min restrain stress(WT, 366.6 ± 37.9 nmol ⁄ L; MR-Tg, 310.4 ± 15.8 nmol ⁄ L; n ¼ 6)and its recovery 60 min later (WT, 125.3 ± 16.4 nmol ⁄ L; MR-Tg,98.1 nmol ⁄ L ± 25.6; n ¼ 6; Fig. 3b). In support of the normal long-term HPA axis function in MR-Tg mice, there were no differences inadrenal or thymus weight compared to controls (Table 1). The normalHPA axis function in MR-Tg did not appear to be due tocompensatory changes in GR, as GR mRNA levels were similar inMR-Tg and control hippocampus (CA-1, CA-3) and hypothalamicparaventricular nucleus (n ¼ 4; Table 1).
Table 1. Biological parameters of male wild-type (WT) and MR-Tg mice at3 months of age
Parameter (Units�) WT MR-Tg n P
WeightsBody (g) 30.4 ± 0.5 30.7 ± 1.3 40 n.s.Adrenal (lg ⁄ g) 10.6 ± 1.3 11.5 ± 0.5 12 n.s.Thymus (mg ⁄ g) 2.2 ± 0.2 2.2 ± 0.1 12 n.s.Heart (mg ⁄ g) 5.1 ± 0.2 4.8 ± 0.2 12 n.s.Kidney (mg ⁄ g) 7.2 ± 0.3 6.5 ± 0.3 12 n.s.
Basal expressionGR mRNAPVN (%) 100.0 ± 1.3 101.4 ± 1.8 6 n.s.CA-1 (%) 100.0 ± 14.1 92.7 ± 8.9 6 n.s.CA-3 (%) 100.0 ± 16.7 95.5 ± 14.6 6 n.s.
MR mRNACA-1 (%) 100.0 ± 1.2 401.2 ± 3.3 6 < 0.01CA-2 (%) 100.0 ± 12.4 684.7 ± 1.7 6 < 0.001CA-3 (%) 100.0 ± 2.7 336.7 ± 4.9 6 < 0.01Cortex (%) 100.0 ± 2.6 3543.6 ± 9.5 6 < 0.01Striatum (%) 100.0 ± 7.5 1224.4 ± 15.5 6 < 0.001BLA (%) 100.0 ± 5.8 1982.9 ± 10.4 6 < 0.001
�Organ weights are expressed as a ratio of body weight. Values are mean ±SEM.MR andGRmRNA expression was measured by densitometric analysis ofautoradiographs. P-values are vs. wild-type controls; n.s., not significant. BLA,basolateral nucleus of amygdala; PVN, hypothalamic paraventricular nucleus.
Fig. 3. Effect of transgene expression on (a) basal circadian trough plasmacorticosterone levels; (b) plasma corticosterone levels in response to 10 minof acute restraint stress taken immediately and 50 min after the end of restraint.TG, transgenic (MR-Tg). Values are the mean ± SEM; n ¼ 6–8. There were nosignificant differences in these HPA axis responses between groups.
1836 M. Lai et al.
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 25, 1832–1842

Forebrain MR overexpression did not affect blood pressure
Differences in arterial blood pressure might confound observeddifferences in the response to cerebral ischaemia. Aldosterone (but notcorticosterone) acts centrally via MR to elevate blood pressure. Suchaldosterone-selective MR are not found in hippocampus and forebrainbut are believed to be located in circumventricular and brainstemstructures ‘protected’ from corticosterone by 11b-hydroxysteroiddehydrogenase type 2 (Gomez-Sanchez, 2004). To assess the effectsof forebrain MR overexpression on blood pressure we measuredsystolic blood pressure using tail-cuff plethysmography; there were nosignificant differences between groups (wild-type,104.5 ± 1.3 mmHg; MR-Tg, 103.8 ± 2.7 mmHg; n ¼ 6). Moreover,heart and kidney weights were similar in MR-Tg and wild-typelittermates (Table 1), also indicating no long-term differences in bloodpressure.
Injury following transient global cerebral ischaemia was attenu-ated by overexpression of MR in forebrain neurons
To test whether increased neuronal MR expression per se has aneuroprotective effect in vivo we used a model of transient globalischaemia (20-min bilateral common carotid artery occlusion) inwhich neurons of the CA-1 and CA-2 hippocampal regions and thecaudate nucleus are particularly susceptible to damage. There were nodifferences in mortality between MR-Tg (0) mice and wild-typecontrols (0) within 72 h after global ischaemia and reperfusion.Quantitative analysis of ischaemic neurons showed that MR-Tg micewere significantly protected from the effects of transient globalcerebral ischaemia, demonstrating a significantly reduced proportionof ischaemic neurons compared to wild-type controls (Fig. 4; two-wayanova, genotype effect: WT, 50.6 ± 3.7%; MR-Tg, 33.4 ± 3.9%,
n ¼ 12; F1,63 ¼ 10.4, P ¼ 0.002). Post hoc comparisons indicatedthat this effect was most marked in the CA-2 region (WT,66.2 ± 6.0%; MR-Tg, 42.5 ± 7.8%; F1,21 ¼ 5.87, P ¼ 0.02) com-pared to CA-1 (WT, 27.2 ± 5.7%; MR-Tg, 15.0 ± 4.4%) and caudatenucleus (WT, 58.4 ± 7.9%; MR-Tg, 42.6 ± 6.6%).
MR-Tg showed reduced anxiety-related behaviour
Pharmacological manipulations of central MR signalling havesuggested that the receptor modulates anxiety-related behaviour(Bitran et al., 1998; Smythe et al., 1997). We therefore observedMR-Tg and wild-type littermates in different aversive environments.In the open-field test MR-Tg made significantly more entries into(WT, 4.5 ± 0.9%, n ¼ 20; MR-Tg, 9.6 ± 1.8%, n ¼ 20; P ¼ 0.05,Fig. 5a) and spent more time in (WT, 4.9 ± 2.2 s; MR-Tg,12.5 ± 3.2 s, P ¼ 0.03; Fig. 5b) the ‘anxiogenic’ central zone thanwild-type littermates. There was no difference in general locomotoractivity (WT 409.1 ± 37.9 cm, n ¼ 20; MR-Tg, 515.6 ± 69.8 cm,n ¼ 20). MR-Tg also showed a decreased anxiogenic response in thelight–dark exploration test, demonstrating significantly reducedlatency to enter the light box (WT, 69.3 ± 21.4 s, n ¼ 12; MR-Tg,30.3 ± 9.9 s, n ¼ 12; P ¼ 0.02; Fig. 5c) and a trend to more entriesinto the light box than wild-type mice (WT, 13.4 ± 2.5; MR-Tg,19.8 ± 2.2; P ¼ 0.07; Fig. 5d).
MR-Tg showed altered reactivity in novel object recognition task
In a separate cohort of mice we measured differences in exploratoryactivity when confronted with both a novel object and a familiar object.There was no overall difference in total exploration time between wild-type and MR-Tg mice; however, MR-Tg mice showed a significant
Fig. 4. Effect of transgene expression on the response to transient global cerebral ischaemia. Results represent the percentage ischaemic neuronal damage. Valuesare the mean ± SEM. WT, n ¼ 12; MR-Tg, n ¼ 11. (a) Overall ischaemic damage was significantly lower in MR-Tg mice (TG) than in wild-type controls (two-wayanova, genotype effect: F1,63 ¼ 10.4, P ¼ 0.002). Post hoc analysis indicated significant differences between groups (P ¼ 0.02) in the CA-2 hippocampal region.(b) Histological sections show the extent of ischaemic neuronal damage in the CA-2 pyramidal cell layer of MR-Tg (upper panel) and WT (lower panel). *P < 0.05.
Increased mineralocorticoid receptor is protective 1837
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 25, 1832–1842

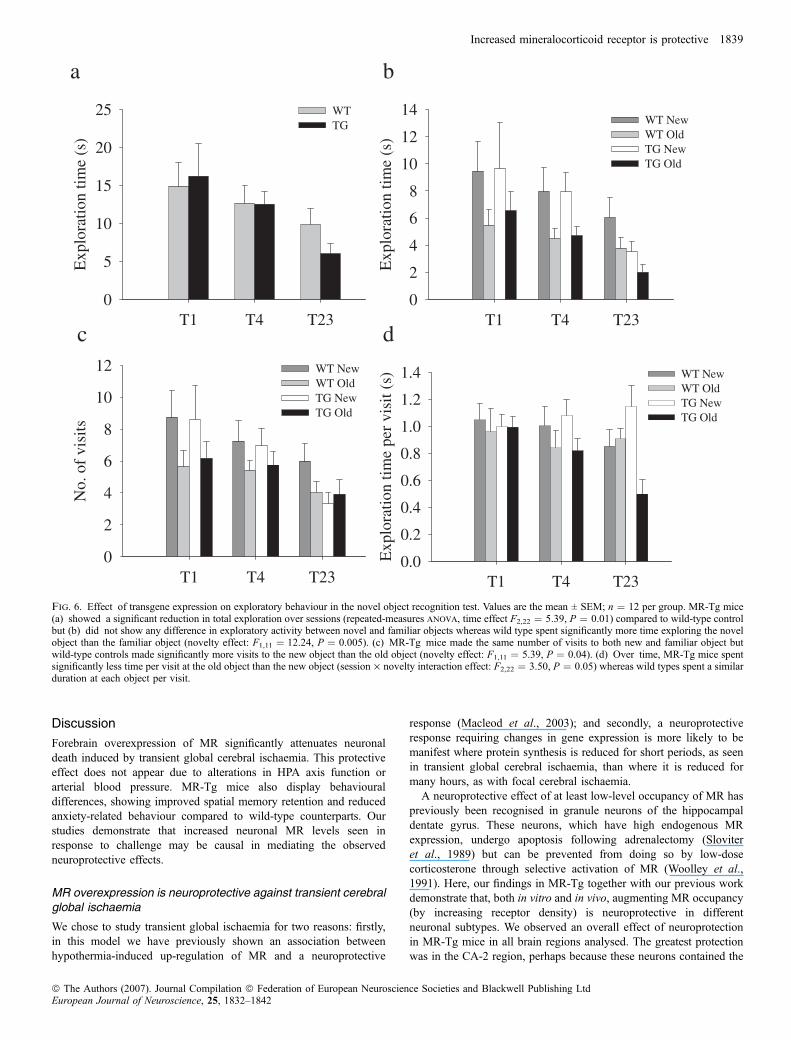
reduction in exploration over sessions compared to wild-type controls(WT, F2,22 ¼ 2.05, P ¼ 0.15; MR-Tg, F2,22 ¼ 5.39, P ¼ 0.01;Fig. 6a). When analysed together, wild-type and MR-Tg mice showedincreased exploration of the novel object compared to familiar object(F1,22 ¼ 13.05, P ¼ 0.002; Fig. 6b). However, when genotypes wereanalysed separately, wild-type controls displayed significantly greaterexploration of the novel object than the familiar object (F1,11 ¼ 12.24,P ¼ 0.005) whereas MR-Tg mice did not show this preference(F1,11 ¼ 3.71, P ¼ 0.08). We also measured the number of visits madeto each object and the duration of each visit: wild-type controlsconsistently made more visits to the novel object than to the familiarobject (F1,11 ¼ 5.39, P ¼ 0.04) whereas MR-Tg mice made the sameamount of visits to both objects (F1,11 ¼ 1.15, P ¼ 0.3; Fig. 6c).However, whilst wild types spent a similar duration per visit to eachobject (F1,11 ¼ 0.406, P ¼ 0.54), MR-Tg mice significantly spent lesstime per visit at the familiar object than the novel object (F1,11 ¼ 15.38,P ¼ 0.002), this also being significant over time (session · noveltyinteraction: WT, F2,22 ¼ 0.78, P ¼ 0.47; MR-Tg, F2,22 ¼ 3.50,P ¼ 0.05; Fig. 6d), findings compatible with differences in behaviouralreaction to novelty between MR-Tg and control mice.
MR-Tg showed improved spatial memory retention
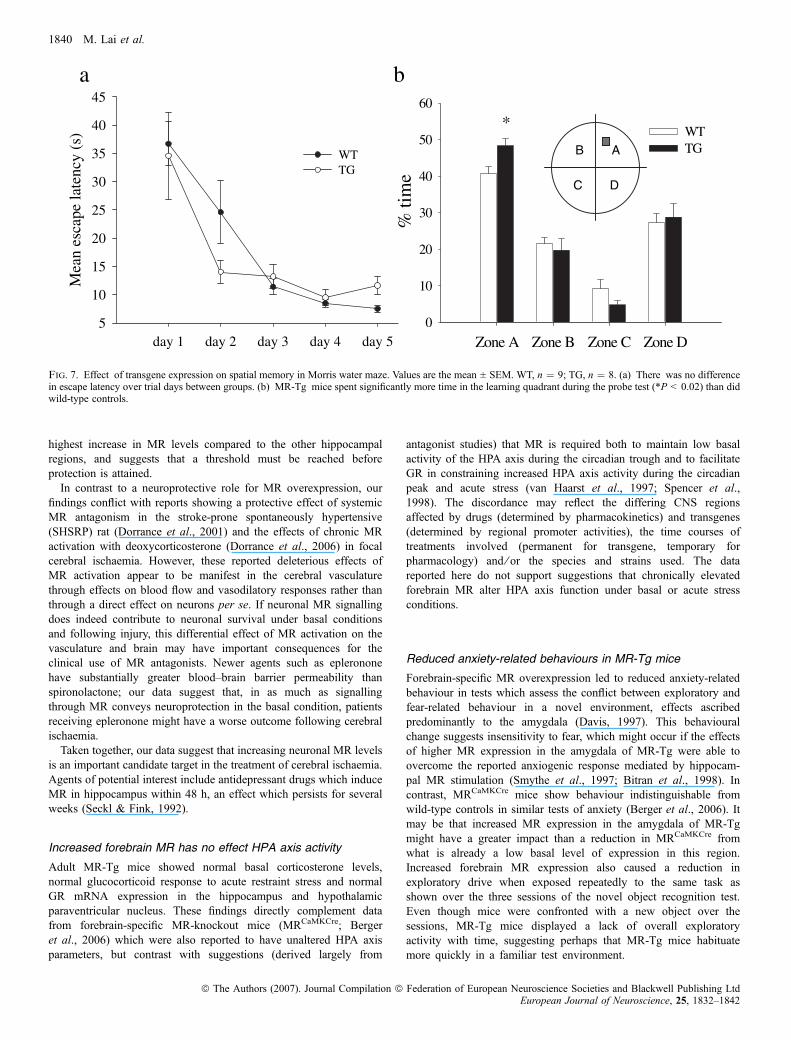
Both central MR antagonism (Oitzl & de Kloet, 1992; Yau et al.,1999) and loss of forebrain MR (Berger et al., 2006) impairmemory, particularly in tests of spatial memory retention, whilechronic GR antagonism often facilitates hippocampus-associatedmemory (Oitzl & de Kloet, 1992). We used the water maze andnovel object recognition to determine the effect of increasedforebrain MR on learning and memory. In the water maze, animalswere trained for 5 days (four trials per day) to find the hiddenplatform. Escape latency decreased in both wild-type and MR-Tg,with successive days of training indicating that spatial learning hadoccurred. This learning appeared to be at the same rate in bothwild-type and MR-Tg mice (Fig. 7a). One hour after the last trial,animals underwent a further 90-s trial without a hidden platform totest the retention of spatial memory; MR-Tg mice spent asignificantly greater percentage of time in the target quadrant thandid wild-type controls (WT, 40.6 ± 2.1%; MR-Tg, 48.3 ± 2.1%;F1,21 ¼ 6.93, P ¼ 0.02; Fig. 7b), indicating improved retention ofspatial memory.
Fig. 5. Effect of transgene expression on anxiety-related behaviour in (a and b) the open-field test and in (c and d) the light–dark exploration test. Values are themean ± SEM; n ¼ 12 per group. Analysis of behaviour in the open-field test showed that MR-Tg mice (a) entered (*P < 0.05) and (b) spent more time(**P < 0.03) in the central ‘anxiogenic zone’ than wild-type controls. In the light–dark exploration test MR-Tg mice showed (c) a decreased latency to enter thelight box (***P < 0.02) but (d) no difference in the time spent in the light box compared to wild-type controls.
1838 M. Lai et al.
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 25, 1832–1842
Discussion
Forebrain overexpression of MR significantly attenuates neuronaldeath induced by transient global cerebral ischaemia. This protectiveeffect does not appear due to alterations in HPA axis function orarterial blood pressure. MR-Tg mice also display behaviouraldifferences, showing improved spatial memory retention and reducedanxiety-related behaviour compared to wild-type counterparts. Ourstudies demonstrate that increased neuronal MR levels seen inresponse to challenge may be causal in mediating the observedneuroprotective effects.
MR overexpression is neuroprotective against transient cerebralglobal ischaemia
We chose to study transient global ischaemia for two reasons: firstly,in this model we have previously shown an association betweenhypothermia-induced up-regulation of MR and a neuroprotective
response (Macleod et al., 2003); and secondly, a neuroprotectiveresponse requiring changes in gene expression is more likely to bemanifest where protein synthesis is reduced for short periods, as seenin transient global cerebral ischaemia, than where it is reduced formany hours, as with focal cerebral ischaemia.A neuroprotective effect of at least low-level occupancy of MR has
previously been recognised in granule neurons of the hippocampaldentate gyrus. These neurons, which have high endogenous MRexpression, undergo apoptosis following adrenalectomy (Sloviteret al., 1989) but can be prevented from doing so by low-dosecorticosterone through selective activation of MR (Woolley et al.,1991). Here, our findings in MR-Tg together with our previous workdemonstrate that, both in vitro and in vivo, augmenting MR occupancy(by increasing receptor density) is neuroprotective in differentneuronal subtypes. We observed an overall effect of neuroprotectionin MR-Tg mice in all brain regions analysed. The greatest protectionwas in the CA-2 region, perhaps because these neurons contained the
Fig. 6. Effect of transgene expression on exploratory behaviour in the novel object recognition test. Values are the mean ± SEM; n ¼ 12 per group. MR-Tg mice(a) showed a significant reduction in total exploration over sessions (repeated-measures anova, time effect F2,22 ¼ 5.39, P ¼ 0.01) compared to wild-type controlbut (b) did not show any difference in exploratory activity between novel and familiar objects whereas wild type spent significantly more time exploring the novelobject than the familiar object (novelty effect: F1,11 ¼ 12.24, P ¼ 0.005). (c) MR-Tg mice made the same number of visits to both new and familiar object butwild-type controls made significantly more visits to the new object than the old object (novelty effect: F1,11 ¼ 5.39, P ¼ 0.04). (d) Over time, MR-Tg mice spentsignificantly less time per visit at the old object than the new object (session · novelty interaction effect: F2,22 ¼ 3.50, P ¼ 0.05) whereas wild types spent a similarduration at each object per visit.
Increased mineralocorticoid receptor is protective 1839
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 25, 1832–1842
highest increase in MR levels compared to the other hippocampalregions, and suggests that a threshold must be reached beforeprotection is attained.In contrast to a neuroprotective role for MR overexpression, our
findings conflict with reports showing a protective effect of systemicMR antagonism in the stroke-prone spontaneously hypertensive(SHSRP) rat (Dorrance et al., 2001) and the effects of chronic MRactivation with deoxycorticosterone (Dorrance et al., 2006) in focalcerebral ischaemia. However, these reported deleterious effects ofMR activation appear to be manifest in the cerebral vasculaturethrough effects on blood flow and vasodilatory responses rather thanthrough a direct effect on neurons per se. If neuronal MR signallingdoes indeed contribute to neuronal survival under basal conditionsand following injury, this differential effect of MR activation on thevasculature and brain may have important consequences for theclinical use of MR antagonists. Newer agents such as eplerononehave substantially greater blood–brain barrier permeability thanspironolactone; our data suggest that, in as much as signallingthrough MR conveys neuroprotection in the basal condition, patientsreceiving epleronone might have a worse outcome following cerebralischaemia.Taken together, our data suggest that increasing neuronal MR levels
is an important candidate target in the treatment of cerebral ischaemia.Agents of potential interest include antidepressant drugs which induceMR in hippocampus within 48 h, an effect which persists for severalweeks (Seckl & Fink, 1992).
Increased forebrain MR has no effect HPA axis activity
Adult MR-Tg mice showed normal basal corticosterone levels,normal glucocorticoid response to acute restraint stress and normalGR mRNA expression in the hippocampus and hypothalamicparaventricular nucleus. These findings directly complement datafrom forebrain-specific MR-knockout mice (MRCaMKCre; Bergeret al., 2006) which were also reported to have unaltered HPA axisparameters, but contrast with suggestions (derived largely from
antagonist studies) that MR is required both to maintain low basalactivity of the HPA axis during the circadian trough and to facilitateGR in constraining increased HPA axis activity during the circadianpeak and acute stress (van Haarst et al., 1997; Spencer et al.,1998). The discordance may reflect the differing CNS regionsaffected by drugs (determined by pharmacokinetics) and transgenes(determined by regional promoter activities), the time courses oftreatments involved (permanent for transgene, temporary forpharmacology) and ⁄ or the species and strains used. The datareported here do not support suggestions that chronically elevatedforebrain MR alter HPA axis function under basal or acute stressconditions.
Reduced anxiety-related behaviours in MR-Tg mice
Forebrain-specific MR overexpression led to reduced anxiety-relatedbehaviour in tests which assess the conflict between exploratory andfear-related behaviour in a novel environment, effects ascribedpredominantly to the amygdala (Davis, 1997). This behaviouralchange suggests insensitivity to fear, which might occur if the effectsof higher MR expression in the amygdala of MR-Tg were able toovercome the reported anxiogenic response mediated by hippocam-pal MR stimulation (Smythe et al., 1997; Bitran et al., 1998). Incontrast, MRCaMKCre mice show behaviour indistinguishable fromwild-type controls in similar tests of anxiety (Berger et al., 2006). Itmay be that increased MR expression in the amygdala of MR-Tgmight have a greater impact than a reduction in MRCaMKCre fromwhat is already a low basal level of expression in this region.Increased forebrain MR expression also caused a reduction inexploratory drive when exposed repeatedly to the same task asshown over the three sessions of the novel object recognition test.Even though mice were confronted with a new object over thesessions, MR-Tg mice displayed a lack of overall exploratoryactivity with time, suggesting perhaps that MR-Tg mice habituatemore quickly in a familiar test environment.
5yad4yad3yad2yad1yad
Mea
n es
cape
late
ncy
(s)
5
01
51
02
52
03
53
04
54
TWGT
DenoZCenoZBenoZAenoZ
% ti
me
0
01
02
03
04
05
06
TWGT
*
DC
AB
ba
Fig. 7. Effect of transgene expression on spatial memory in Morris water maze. Values are the mean ± SEM. WT, n ¼ 9; TG, n ¼ 8. (a) There was no differencein escape latency over trial days between groups. (b) MR-Tg mice spent significantly more time in the learning quadrant during the probe test (*P < 0.02) than didwild-type controls.
1840 M. Lai et al.
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 25, 1832–1842

Increased MR results in improved spatial memory performance
Efficient learning and memory performance is thought to depend onthe appropriate functioning of the HPA axis brought about bycoordinated activation of MR and GR (Oitzl & de Kloet, 1992).Studies using both specific MR- and GR-antagonists and transgenicmodels of reduced MR and GR signalling (Gass et al., 2001) indicatethat chronic changes in HPA axis activity interfere with cognitiveperformance. On the other hand, treatment with antidepressants whichrestore HPA axis function can prevent cognitive decline with ageing(Yau et al., 2002). Increased forebrain MR improved performance inthe probe test of the water maze, confirming a role for MR in memoryretention. Indeed, antidepressant-induced increases in hippocampalMR correlate with improved spatial memory retention whereaschronic (4 weeks) intracerebroventricular spironolactone infusionmediates a selective decline in probe test performance (Yau et al.,1999). Performance of MR-Tg mice in the object recognition test,however, suggests that MR expression may be more important forspatial than nonspatial memory.
Summary
In conclusion, using a transgenic mouse model with selectiveoverexpression of MR in the forebrain we provide novel evidencefor a neuroprotective role of increased MR against cerebral ischaemicdamage and we present data that provide additional support for animportant role of forebrain MR in mood and memory retention. Thesefindings provide further support for the development of treatmentsdirected at increasing brain MR expression and ⁄ or signalling, not onlyin cerebral ischaemia but also in conditions characterized by disordersof mood and memory.
Acknowledgements
This work was supported by a Royal Society of Edinburgh–Lloyds TSBFoundation Research Fellowship (M.L.), a Wellcome Trust University Award(K.H.) and a Wellcome Trust project grant (M.R.M.). We thank Jessie Hongand William Mungall for excellent technical assistance.
Abbreviations
CaMKIIa, calcium–calmodulin-dependent protein kinase IIa; GR, glucocorti-coid receptor; HA, haemagglutinin; HPA, hypothalamic–pituitary–adrenal;MR, mineralocorticoid receptor; MR-Tg, forebrain-specific MR-overexpressingtransgenic mice; WT, wild-type.
References
Abraham, I.M., Harkany, T., Horvath, K.M. & Luiten, P.G. (2001) Actions ofglucocorticoids on survival of nerve cells: promopting neurodegeneration ofneuroprotection? J. Neuroendocrinol., 13, 749–760.
Arriza, J.L., Weinberger, C., Cerelli, G., Glaser, T.M., Handelin, B.L.,Housman, D.E. & Evans, R.M. (1987) Cloning of human mineralocorticoidreceptor complementary DNA: structural and functional kinship with theglucocorticoid receptor. Science, 237, 268–275.
Berger, S., Wolfer, D.P., Selbach, O., Alter, H., Erdmann, G., Reichardt, H.M.,Chepkova, A.N., Welz, H., Hass, H.L., Lipp, H.-P. & Schutz, G. (2006) Lossof the limbic mineralocorticoid receptor impairs behavioural plasticity. Proc.Natl Acad. Sci. USA, 103, 195–200.
Bitran, D., Shiekh, M., Dowd, J.A., Dugan, M.M. & Renda, P. (1998)Corticosterone is permissive to the anxiolytic effect that results from theblockade of hippocampal mineralocorticoid receptors. Pharmacol. Biochem.Behav., 60, 879–887.
Davis, M. (1997) Neurobiology of fear responses: the role of the amygdala.J. Neuropsychiatry Clin. Neurosci., 9, 382–402.
Dorrance, A.M., Osborn, H.L., Grekin, R. & Webb, R.C. (2001) Spironolactonereduces cerebral infarct size and EGF-receptor mRNA in stroke prone rats.Am. J. Physiol. Regul. Integr. Comp. Physiol., 281, R944–R950.
Dorrance, A.M., Rupp, N.C. & Noguiera, E.F. (2006) Mineralocorticoidreceptor activation causes cerebral vessel remodeling and exacerbates thedamage caused by cerebral ischemia. Hypertension, 47, 590–595.
Evans, A.L., Brown, W., Kenyon, C.J., Maxted, K.J. & Smith, D.C. (1994)Improved system for measuring systolic blood pressure in the conscious rat.Med. Biol. Eng. Comput., 32, 101–102.
Gass, P., Reichardt, H.M., Strekalova, T., Henn, F. & Tronche, F. (2001) Micewith targeted mutations of glucocorticoid and mineralocorticoid receptors:models of depression and anxiety? Physiol. Behav., 73, 811–825.
Gomez-Sanchez, E.P. (2004) Brain mineralocorticoid receptors: orchestratorsof hypertension and end-organ disease. Curr. Opin. Nephrol. Hypertension,13, 191–196.
Gomez-Sanchez, C.E., de Rodriguez, A.F., Romero, D.G., Estess, J., Warden,M.P., Gomez-Sanchez, M.T. & Gomez-Sanchez, E.P. (2006) Development ofa panel of monoclonal antibodies against the mineralocorticoid receptor.Endocrinology, 147, 1343–1348.
Holmes, M.C., Abrahamsen, C.T., French, K.L., Paterson, J.M., Mullins, J.J. &Seckl, J.R. (2006) The mother or the fetus? 11 beta hydroxysteroid-dehydrogenase type 2 null mice provide evidence for direct fetalprogramming of behaviour by endogenous glucocorticoids. J. Neurosci.,26, 3840–3844.
Holmes, M.C., French, K.L. & Seck1, J.R. (1997) Dysregulation of diurnalrhythms of serotonin 5HT-2C and corticosteroid receptor gene expression inthe hippocampus with food restriction and glucocorticoids. J. Neurosci., 17,4056–4065.
Joels, M. (2001) Corticosteroid actions in the hippocampus. J. Neuroendocr-inol., 13, 57–669.
Kelly, S., McCulloch, J. & Horsburgh, K. (2001) Minimal ischemic neuronaldamage and HSP70 expression in MF1 strain mice following bilateralcommon carotid artery occlusion. Brain Res., 914, 185–195.
de Kloet, E.R., Oitzl, M.S. & Joels, M. (1999) Stress and cognition: arecorticosteroids good or bad guys? Trends Neurosci., 22, 422–426.
de Kloet, E.R., Vreugdenhil, E., Oitzl, M.S. & Joels, M. (1998) Braincorticosteroid receptor balance in health and disease. Endocrine Rev., 19,269–301.
Krugers, H.J., Maslam, S., Korf, J., Joels, M. & Holsboer, F. (2000) Thecorticosterone synthesis inhibitor metyrapone prevents hypoxia ⁄ ischaemic-induced loss of synaptic function in the rat hippocampus. Stroke, 31, 1162–1172.
Macleod, M.R., Johansson, I.M., Soderstrom, I., Lai, M., Gido, G., Wieloch, T.,Seckl, J.R. & Olsson, T. (2003) Mineralocorticoid receptor expression andincreased survival following neuronal injury. Eur. J. Neurosci., 17, 1549–1555.
Mayford, M., Bach, M.E., Huang, Y.Y., Wang, L., Hawkins, R.D. & Kandel,E.R. (1996) Control of memory formation through regulated expression of aCamKII transgene. Science, 274, 1678–1683.
Miesfeld, R., Rusconi, S., Godowski, P.J., Maler, B.A., Okret, S., Wilkstrom,A.-C., Gustafsson, J.-A. & Yamamoto, K.R. (1986) Genetic complementa-tion of a glucocorticoid receptor deficiency by expression of a clonedreceptor cDNA. Cell, 49, 389–399.
Oitzl, M.S. & de Kloet, E.R. (1992) Selective corticosteroid antagonistsmodify specific aspects of spatial orientation learning. Behav. Neurosci.,106, 62–71.
Reagan, L.P. & McEwen, B.S. (1997) Controversies surrounding glucocorti-coid-mediated cell death in the hippocampus. J. Chem. Neuroanat., 13, 149–167.
Reul, J.M. & de Kloet, E.R. (1985) Two receptor systems for corticosterone inrat brain: microdistribution and differential occupation. Endocrinology, 117,2505–2511.
Sapolsky, R.M. (2000) Glucocorticoids and hippocampal atrophy in neuropsy-chiatric disorders. Arch. Gen. Psychiatry, 57, 925–930.
Seckl, J.R. & Fink, G. (1992) Antidepressants increase glucocorticoid andmineralocorticoid receptor messenger RNA expression. Neuroendocrinology,55, 621–626.
Sloviter, R.S., Valiquette, G., Abrams, G.M., Ronk, E.C., Sollas, A.L., Paul,L.A. & Neubort, S. (1989) Selective loss of hippocampal granule cells in themature rat brain after adrenalectomy. Science, 243, 535–538.
Smythe, J.W., Murphy, D., Timothy, C. & Costall, B. (1997) Hippocampalmineralocorticoid but not glucocorticoid receptors modulate anxiety-likebehaviour in rats. Pharmacol. Biochem. Behav., 56, 507–513.
Spencer, R.L., Kim, P.J., Kalman, B.A. & Cole, M.A. (1998) Evidence formineralocorticoid receptor facilitation of glucocorticoid receptor dependent
Increased mineralocorticoid receptor is protective 1841
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 25, 1832–1842

regulation of hypothalamic-pituitary-adrenal axis activity. Endocrinology,139, 2718–2726.
Van Haarst, A.D., Oitzl, M.S. & de Kloet, E.R. (1997) Facilitation of feedbackinhibition through blockade of glucocorticoid receptors in the hippocampus.Neurochem. Res., 22, 1323–1328.
Woolley, C.S., Gould, E., Sakai, R.R., Spencer, R.L. & McEwen, B.S. (1991)Effects of aldosterone or RU28362 treatment on adrenalectomy-induced celldeath in the dentate gyrus of the adult rat. Brain Res., 554, 312–315.
Yau, J.L., Noble, J., Hibberd, C., Rowe, W.B., Meaney, M.J., Morris, R.G. &Seckl, J.R. (2002) Chronic treatment with the antidepressant amitryptiline
prevents impairments in water maze learning in aged rats. J. Neurosci., 15,1436–1442.
Yau, J.L., Noble, J., Kenyon, C.J., Hibberd, C., Kotelevtsev, Y., Mullins, J.J.& Seckl, J.R. (2001) Lack of tissue glucocorticoid reactivation in11beta-hydroxysteroid dehydrogenase type 1 knockout mice amelioratesage-related learning impairments. Proc. Natl Acad. Sci. USA, 98, 4716–4721.
Yau, J.L., Noble, J. & Seckl, J.R. (1999) Continuous blockade of brainmineralocorticoid receptors impairs spatial learning in rats. Neurosci. Lett.,277, 45–48.
1842 M. Lai et al.
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 25, 1832–1842




















