
Central mineralocorticoid receptors are indispensable for corticosterone-induced impairment of...
10
CENTRAL MINERALOCORTICOID RECEPTORS ARE INDISPENSABLE FOR CORTICOSTERONE-INDUCED IMPAIRMENT OF MEMORY RETRIEVAL IN RATS M. KHAKSARI, A. RASHIDY-POUR* AND A. A. VAFAEI Laboratory of Learning and Memory, Department and Research Cen- ter of Physiology, Semnan University of Medical Sciences, P.O. Box 39195-163, Semnan, Iran Abstract—Previous studies indicated that stress levels of glucocorticoid hormones (cortisol in humans, and corticoste- rone in rodents) induce impairment of long-term memory retrieval, but the underlying mechanisms (genomic or non- genomic) are not clear. To clarify this issue, we investigated the involvement of brain corticosteroid receptors and protein synthesis in the corticosterone-induced impairment of mem- ory retrieval. Young rats were trained in the water maze task with six trials per day for 6 consecutive days. Retention of the spatial training was assessed 24 h after the last training session with a 60-s probe trial. Experiments included intra- ventricular injections of anisomycin, a specific protein syn- thesis inhibitor or specific antagonists for both types of cor- ticocosteroid receptors (mineralocorticoid receptor, MR, and glucocorticoids receptor, GR) before corticosterone admin- istration shortly before retention testing. The results showed that administration of anisomycin did not change the corti- costerone response. Administration of the MR, but not GR, antagonist blocked the corticosterone-induced response dose dependently. These findings provide evidence for the view that glucocorticoids impair memory retrieval through nongenomic mechanisms involving an interaction with cen- tral MRs. © 2007 IBRO. Published by Elsevier Ltd. All rights reserved. Key words: mineralocorticoid receptor, glucocorticoid recep- tor, spironolactone, RU 38486, anisomycin, memory retrieval. Neural mechanisms of memory retrieval are susceptible to disruption by glucocorticoids, the main stress hormones released from the adrenal cortex in a variety of stressful situations. In addition to well-known effects of glucocorti- coids on memory acquisition and consolidation (Lupien and McEwen, 1997; Belanoff et al., 2001; Roozendaal 2002), an increasing body of knowledge indicates that stress and glucocorticoids also impair retrieval of previ- ously acquired information including contextual/spatial in- formation in animals (de Quervain et al., 1998; Pakdel and Rashidy-Pour, 2006; Sajadi et al., 2006 and 2007) and declarative/episodic information in humans (de Quervain et al., 2000; Kuhlmann et al., 2004). However, the mech- anisms underlying such glucocorticoids effects on memory retrieval are not known. Classically, glucocorticoids modulate gene expression by an interaction with intracellular nuclear receptors, which act as ligand-dependent transcription factors. There are two intracellular receptors for glucocorticoids: the miner- alocorticoid receptor (MR) and the glucocorticoid receptor (GR). GRs have a low affinity for corticosterone and be- come occupied only during stress and at the circadian peak, when circulating levels of glucocorticoids are high. In contrast, MRs have a 10-fold higher affinity for corticoste- rone and are almost saturated under basal conditions. (Reul and de Kloet, 1985). In addition to genomic action, recent evidence indicates that glucocorticoids also induce rapid neural effects through nongenomic mechanisms in- volving membrane-associated receptors (for review see Makara and Haller, 2001). Three criteria have been established for differentiat- ing genomic and nongenomic actions of glucocorticoids. These are temporal, the MR/GR independence criterion, and the genomic independence criterion. In general, glu- cocorticoids effects that occur in a time frame incompatible with the genomic mechanism, are not affected by blockade of MR/GR, and are resistant to protein synthesis blockade can be mediated by nongenomic mechanisms (for review see Makara and Haller, 2001). However, others have re- ported nongenomic effects of glucocorticoids via mem- brane MR and/or GR activation (Tasker et al., 2006; Karst et al., 2005). Glucocorticoid effects on memory retrieval happen too fast to be mediated via interaction with intracellular recep- tors and may involve interaction with membrane receptors and several neurotransmitter systems in the brain (Rashidy- Pour et al. 2004, Pakdel and Rashidy-Pour 2006, Roozendaal et al., 2004b, Evans et al., 2000, Orchinik et al., 1992, Hua and Chen, 1989). In a recent study (Sajadi et al., 2006), we demonstrated that intrahippocampal infu- sion of anisomycin, an inhibitor of protein synthesis, did not block glucocorticoid impairment of memory retrieval, sug- gesting an involvement of a nongenomic mechanism in mediating this glucocorticoid response. To further clarify this issue, the present study was designed to define the role of the brain corticosteroid receptors and protein syn- thesis in the effects of systemic glucocorticoids on retrieval of long-term memory in the water maze (WM). For this purpose different groups of rats were injected corticosterone (CORT) with or without prior i.c.v. infusions of anisomycin (ANI) or antagonists for both types of corti- costeroid receptors before CORT administration shortly *Corresponding author. Tel: 98-231-3332082; fax: 98-231-3331551. E-mail address: [email protected] (A. Rashidy-Pour). Abbreviations: ANI, anisomycin; ANOVA, analysis of variance; CORT, corticosterone; GR, glucocorticoid receptor; MR, mineralocorticoid re- ceptor; SAL, saline; SPR, spironolactone; VEH, vehicle; WM, water maze. Neuroscience 149 (2007) 729 –738 0306-4522/07$30.000.00 © 2007 IBRO. Published by Elsevier Ltd. All rights reserved. doi:10.1016/j.neuroscience.2007.08.016 729
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Central mineralocorticoid receptors are indispensable for corticosterone-induced impairment of...

CFR
M
Lt3
Agrrgtsowssvttgitcadvntr
Kt
Ndrsca2sofRde
*EAccm
Neuroscience 149 (2007) 729–738
0d
ENTRAL MINERALOCORTICOID RECEPTORS ARE INDISPENSABLEOR CORTICOSTERONE-INDUCED IMPAIRMENT OF MEMORY
ETRIEVAL IN RATSar
bata(cpcr(rrvM
iTacwocspbe
ftaPRaesbgmtrto
co
. KHAKSARI, A. RASHIDY-POUR* AND A. A. VAFAEI
aboratory of Learning and Memory, Department and Research Cen-er of Physiology, Semnan University of Medical Sciences, P.O. Box9195-163, Semnan, Iran
bstract—Previous studies indicated that stress levels oflucocorticoid hormones (cortisol in humans, and corticoste-one in rodents) induce impairment of long-term memoryetrieval, but the underlying mechanisms (genomic or non-enomic) are not clear. To clarify this issue, we investigatedhe involvement of brain corticosteroid receptors and proteinynthesis in the corticosterone-induced impairment of mem-ry retrieval. Young rats were trained in the water maze taskith six trials per day for 6 consecutive days. Retention of thepatial training was assessed 24 h after the last trainingession with a 60-s probe trial. Experiments included intra-entricular injections of anisomycin, a specific protein syn-hesis inhibitor or specific antagonists for both types of cor-icocosteroid receptors (mineralocorticoid receptor, MR, andlucocorticoids receptor, GR) before corticosterone admin-
stration shortly before retention testing. The results showedhat administration of anisomycin did not change the corti-osterone response. Administration of the MR, but not GR,ntagonist blocked the corticosterone-induced responseose dependently. These findings provide evidence for theiew that glucocorticoids impair memory retrieval throughongenomic mechanisms involving an interaction with cen-ral MRs. © 2007 IBRO. Published by Elsevier Ltd. All rightseserved.
ey words: mineralocorticoid receptor, glucocorticoid recep-or, spironolactone, RU 38486, anisomycin, memory retrieval.
eural mechanisms of memory retrieval are susceptible toisruption by glucocorticoids, the main stress hormoneseleased from the adrenal cortex in a variety of stressfulituations. In addition to well-known effects of glucocorti-oids on memory acquisition and consolidation (Lupiennd McEwen, 1997; Belanoff et al., 2001; Roozendaal002), an increasing body of knowledge indicates thattress and glucocorticoids also impair retrieval of previ-usly acquired information including contextual/spatial in-ormation in animals (de Quervain et al., 1998; Pakdel andashidy-Pour, 2006; Sajadi et al., 2006 and 2007) andeclarative/episodic information in humans (de Quervaint al., 2000; Kuhlmann et al., 2004). However, the mech-
Corresponding author. Tel: �98-231-3332082; fax: �98-231-3331551.-mail address: [email protected] (A. Rashidy-Pour).bbreviations: ANI, anisomycin; ANOVA, analysis of variance; CORT,orticosterone; GR, glucocorticoid receptor; MR, mineralocorticoid re-
ceptor; SAL, saline; SPR, spironolactone; VEH, vehicle; WM, wateraze.
306-4522/07$30.00�0.00 © 2007 IBRO. Published by Elsevier Ltd. All rights reseroi:10.1016/j.neuroscience.2007.08.016
729
nisms underlying such glucocorticoids effects on memoryetrieval are not known.
Classically, glucocorticoids modulate gene expressiony an interaction with intracellular nuclear receptors, whichct as ligand-dependent transcription factors. There arewo intracellular receptors for glucocorticoids: the miner-locorticoid receptor (MR) and the glucocorticoid receptorGR). GRs have a low affinity for corticosterone and be-ome occupied only during stress and at the circadianeak, when circulating levels of glucocorticoids are high. Inontrast, MRs have a 10-fold higher affinity for corticoste-one and are almost saturated under basal conditions.Reul and de Kloet, 1985). In addition to genomic action,ecent evidence indicates that glucocorticoids also induceapid neural effects through nongenomic mechanisms in-olving membrane-associated receptors (for review seeakara and Haller, 2001).
Three criteria have been established for differentiat-ng genomic and nongenomic actions of glucocorticoids.hese are temporal, the MR/GR independence criterion,nd the genomic independence criterion. In general, glu-ocorticoids effects that occur in a time frame incompatibleith the genomic mechanism, are not affected by blockadef MR/GR, and are resistant to protein synthesis blockadean be mediated by nongenomic mechanisms (for reviewee Makara and Haller, 2001). However, others have re-orted nongenomic effects of glucocorticoids via mem-rane MR and/or GR activation (Tasker et al., 2006; Karstt al., 2005).
Glucocorticoid effects on memory retrieval happen tooast to be mediated via interaction with intracellular recep-ors and may involve interaction with membrane receptorsnd several neurotransmitter systems in the brain (Rashidy-our et al. 2004, Pakdel and Rashidy-Pour 2006,oozendaal et al., 2004b, Evans et al., 2000, Orchinik etl., 1992, Hua and Chen, 1989). In a recent study (Sajadit al., 2006), we demonstrated that intrahippocampal infu-ion of anisomycin, an inhibitor of protein synthesis, did notlock glucocorticoid impairment of memory retrieval, sug-esting an involvement of a nongenomic mechanism inediating this glucocorticoid response. To further clarify
his issue, the present study was designed to define theole of the brain corticosteroid receptors and protein syn-hesis in the effects of systemic glucocorticoids on retrievalf long-term memory in the water maze (WM).
For this purpose different groups of rats were injectedorticosterone (CORT) with or without prior i.c.v. infusionsf anisomycin (ANI) or antagonists for both types of corti-
osteroid receptors before CORT administration shortlyved.

btri
A
AAwafptEa
S
Am(aTsar�lskcv
D
CdtT(AiRss0iTebSF
I
T(twbcdAm
W
Apedw
6rhofitaAcctrted
S
TarwqCdTdi
E
TC
iafiti(A
slnotswC(aPgrmS(
M. Khaksari et al. / Neuroscience 149 (2007) 729–738730
efore retention testing. Animals were trained in the WMask with six trials per day for 6 consecutive days andetention testing was carried out 24 h later. All drugs werenjected between 30 and 60 min before retention testing.
EXPERIMENTAL PROCEDURES
nimals
dult male Wistar rats (250–300 g) were used in this study.nimals were housed five per cage (60�40�20 cm) in a roomith natural light cycle and constant temperature (24�2 °C). Foodnd water were available ad libitum. All experiments were per-ormed between 10:00 and 13:00 h during the light cycle. Allrocedures were conducted in agreement with the National Insti-utes of Health Guide for care and use of laboratory animals.very effort was made to minimize the number of animals usednd their suffering.
urgical procedure
pproximately 7 days prior to initiation of the behavioral experi-ents, the rats were anesthetized with ketamine hydrochloride
70 mg/kg, i.p.). The rats were fixed in a stereotaxic apparatus,nd a midline incision of the skin in the cranial region was made.he skull was dried and cleaned of fascias. A sterile stainless-teel guide cannula (22 gauge, 12 mm) was inserted below durabove the right cerebral ventricle at the following coordinateselative to the bregma: AP �0.8 mm; L�1.6 mm (midline); DV3.4 from dura; with nose bar �3.30 mm below the interaural
ines (Paxino and Watson, 1998). The cannula was affixed to thekull with dental acrylic; a stylet was inserted into the cannula toeep it patent. At the end of experiments, correct placement of theannula was verified by analysis of dye dispersion throughout theentricular system following i.c.v. injection of 5 �l Methylene Blue.
rugs
ORT (Sigma-Aldrich Chemie Gmbh, Steinheim, Germany) wasissolved in propylene glycol (1 mg/kg) and was injected intraperi-oneally at a volume of 1 ml/kg 30 min before retention testing.his dose of drug was chosen according to our recent studiesPakdel and Rashidy-Pour, 2006; Sajadi et al., 2006, and 2007).NI (Sigma), a specific protein synthesis inhibitor, was dissolved
n saline and was infused (i.c.v.) at doses of 187.5 or 450 �g/5 �l.U 38486, a specific GR antagonist, and spironolactone (SPR), apecific MR antagonist (both purchased from Sigma) were dis-olved in absolute ethanol (5 mg/ml) and subsequently diluted in.9% saline to a final concentrations of 75 or 150 ng/ 5 �l for i.c.v
njections. For SPR, a lower dose of 37.5 ng/ 5 �l was also used.hese doses of corticosteroid receptors antagonists or ANI, andven lower doses of these drugs, were sufficient to interfere withehavioral responses (Oitzl and deKloet, 1992; Oitzl et al.,1994;andi et al., 1996, Meiri and Rosenblum, 1998; Robinson andranklin, 2007).
nfusion procedures
he i.c.v. infusions of the drugs were performed over 5 min1 �l/min) through an injection needle (30 gauge, 14 mm) attachedo a 10-�l Hamilton syringe via polyethylene tubing. The needleas equipped with a stopper limiting the depth of insertion to 2 mmeyond the tip of the cannula. The injection needle remained in theannula for 1 min following the infusion in order to maximizeiffusion away from the needle tip and to minimize dorsal diffusion.NI or corticosteroid receptor antagonists were injected i.c.v. 60
in before retention testing (30 min before CORT injection). (M task and spatial training procedures
detailed description of the apparatus and tracking system wasresented in our previous reports (Sajadi et al., 2006, 2007; Sarihit al., 2000). In brief, the WM was a blue circular pool (140 cm iniameter and 50 cm high) filled to a 25-cm depth with 20�2 °Cater.
For spatial training, rats were subjected to six trials a session forconsecutive days. Twenty-four hours prior to the start of training,
ats were allowed to swim 3 min in the pool containing no platform forabituation. On each trial, the rat was placed into the water from onef the four cardinal points of the compass (N, E, S, W), which variedrom trial to trial in a quasirandom order. The rat had to swim untilt found and climbed onto the escape platform. Rats were guidedo the platform if they failed to locate it within 60 s. The rat wasllowed to stay on the platform for 30 s as the intertrial interval.fter the last trial, the rat was towel dried and placed in a holdingage under a heating lamp before it was returned to the homeage. Performance of each rat was tested 24 h after the finalraining day in a probe trial (60 s), during which the platform wasemoved. The parameters measured from the probe test wereime spent (%) in the target and opposite quadrants, number ofntrances in the target quadrant of the pool, and total swimistance.
tatistical analysis
he training data were analyzed with a one-way analysis of vari-nce (ANOVA) with the six blocks (each consisted of six trials) asepeated measures. Quadrant search times on the probe testere analyzed with a three-way ANOVA with target and oppositeuadrants as repeated measures and i.c.v. drugs and systemicORT injections both as between-subject variables. Swim pathata on the probe test were analyzed with two-way ANOVA.ukey’s post hoc test was performed to determine the source ofetected significances. Values of P�0.05 were considered signif-
cant.
xperiment 1
his experiment examined the effects of i.c.v. injections of ANI onORT-induced impairment of memory retrieval.
Methods. Rats were randomly divided into six groups (n�8n each group) and trained and tested by procedures describedbove. On the test day, the animals received two injections: therst i.c.v. injection was given 60 min prior to retention testing andhe second systemic injection was given 30 min after the firstnjection. The eight pairs of injections were saline (SAL) � vehicleVEH); SAL�CORT, ANI (187.5 �g) � VEH, ANI (450 �g) � VEH,NI (187.5 �g) � CORT, and ANI (450 �g) � CORT.
Results. All rats learned to locate the platform during the 6uccessive days of training as indicated by decreasing escapeatencies as training progressed (F5,282�105.08, P�0.0001, dataot shown). The mean escape latencies of the last day of trainingf all rats were around 5–6 s. Figure 1A illustrates the effects ofreatments on retention performance as assessed by the timepent in the target and opposite quadrants. A three-way ANOVAith quadrant as repeated measures showed a significant effect ofORT (F1,42�8.67, P�0.005), and no significant effect of ANI
F2,42�0.57, P�0.57) on quadrant search times during probe testnd no significant CORT�ANI�quadrant interaction (F2,42�1.61,�0.21). Post hoc comparisons indicated that control animalsiven SAL�VEH spent significantly more time in the target quad-ant than in the opposite quadrant (P�0.01), indicating goodemory for the location of platform. Animals treated withAL�CORT spent significantly less time in the target quadrant
P�0.01) and simultaneously more time in the opposite quadrant
P�0.01) in comparison with the SAL�VEH group. The i.c.v.
Fiot(q
M. Khaksari et al. / Neuroscience 149 (2007) 729–738 731
ig. 1. Effects of i.c.v. injections of anisomycin on corticosterone-induced impairment of long-term memory retrieval in a water maze task. The i.c.v.nfusions of both doses of anisomycin (187.5 or 450 �g/5 �l) 60 min before retention testing did not block the impairing effects of systemic injectionf corticosterone (1 mg/kg, i.p.) 30 min before retention testing on probe trial retention performance. (A, B) The columns represent the mean (�SEM)
ime spent (%) in target (T) and opposite (O) quadrants and number of entrances in the target quadrant of the pool in the 60-s probe test, respectively.C) Total swim path (mean�SEM) in centimeters. * P�0.01 compared with time in training quadrant; ● P�0.01 compared with the corresponding
uadrant in vehicle group that received same treatment; # P�0.01 compared with the vehicle group.
isPr
qtssiaegosi
pPPa
E
T3
iVRtaE
slnrar((taCqqTapc
qtsiiaegosi
pP(
fp
E
TC
eC(nncu
slnsaqCittdnt(dqC
ttaebtt(7ei
pPPmm
TbtraiMivn
M. Khaksari et al. / Neuroscience 149 (2007) 729–738732
nfusions of the different doses of ANI alone did not affect the timepent in the target and opposite quadrants in the control group.retreatment of ANI also did not change the CORT-induced
esponse.The pattern of results for number of entrances into the target
uadrant (Fig. 2B) during the probe test was similar to that for theime spent in the target quadrant. A two-way ANOVA revealed aignificant effect of CORT (F1,42�43.55, P�0.0001), lack of aignificant effect of ANI (F2,42�1.8, P�0.18), and no significant
nteraction between both factors (F2,42�2.44, P�0.1). Systemicdministration of CORT significantly decreased the number ofntrances into the target quadrant in comparison with the controlroup (P�0.01), and this effect was not blocked by i.c.v. injectionsf ANI. The above results show that the impairing effects ofystemic CORT on memory retrieval were not blocked by i.c.v.nfusions of ANI.
Two-way analysis of total swim paths (Fig. 1C) during therobe test indicated no significant effects of CORT (F1,42�2.98,�0.1), ANI (F2,42�0.03, P�0.96), or interaction (F2,42�0.26,�0.76). These findings indicate that motor performance was notffected by any of the experimental manipulations.
xperiment 2
his experiment examined the effects of i.c.v. injections of RU8486 on CORT-induced impairment of memory retrieval.
Methods. Rats were randomly divided into six groups (n�7n each group). The six pairs of injections were VEH�VEH;EH�CORT, RU 38486 (75 ng)�VEH, RU38486 (150ng)�VEH,U38486 (75 ng)�CORT, and RU38486 (150 ng)�CORT. The
raining and testing procedures were the same as mentionedbove. The injection schedule was the same as that reported inxperiment 1.
Results. All rats learned to locate the platform during the 6uccessive days of training as indicated by decreasing escapeatencies as training progressed (F5,246�74.54, P�0.0001, dataot shown). Figure 2A illustrates the effects of treatments onetention performance as assessed by the time spent in the targetnd opposite quadrants. A three-way ANOVA with quadrant asepeated measures showed a significant effect of CORTF1,36�8.91, P�0.005), lack of a significant effect of RU38486F2,36�2.51, P�0.095) on quadrant search times during the probeest, and lack of a significant RU 38486�CORT�quadrant inter-ction (F2,36�0.39 P�0.67). Post hoc comparisons indicated thatORT-treated animals spent significantly less time in the targetuadrant (P�0.01) and simultaneously more time in the oppositeuadrant (P�0.01) when compared with that of the control group.he i.c.v. infusions of the different doses of RU 38486 alone did notffect the time spent in the target and opposite quadrants in com-arison to the control group. Pretreatment of RU38486 also did nothange the CORT-induced response.
The pattern of results for number of entrances into the targetuadrant (Fig. 2B) during the probe test was similar to that for theime spent in the target quadrant. A two-way ANOVA revealed aignificant effect of CORT (F1,36�5.26, P�0.04), lack of a signif-cant effect of RU38486 (F2,36�4.07, P�0.28), and no significantnteraction between both factors (F2,36�2.08, P�0.14). Systemicdministration of CORT significantly decreased the number ofntrances into the target quadrant in comparison with the controlroup (P�0.01), and this effect was not blocked by i.c.v. injectionsf RU38486. These results indicate that the impairing effects ofystemic CORT on memory retrieval were not blocked by i.c.v.nfusions of RU38486.
Two-way analysis of total swim paths (Fig. 2C) during therobe test indicated no significant effects of CORT (F1,36�1.85,�0.19), RU 38486 (F �1.2, P�0.31), or interaction
2,36F2,36�2.31, P�0.12). These findings indicate that retention per- c
ormance was not affected by any cross disturbances in motorerformance induced by experimental manipulations.
xperiment 3
his experiment examined the effects of i.c.v. injections of SPR onORT-induced impairment of memory retrieval.
Methods. The data of the control group from the secondxperiment given VEH�VEH and from the group given VEH plusORT were used in this experiment. The subsequent treatments
n�7 in each group) were: SPR (37.5 ng)�VEH, SPR (75g)�VEH, SPR (150 ng)�VEH, SPR (37.5 ng)�CORT, SPR (75g)�CORT, SPR (150 ng)�CORT. The training and testing pro-edures were the same as mentioned above. The injection sched-le was the same as that reported in Experiment 1.
Results. All rats learned to locate the platform during the 6uccessive days of training as indicated by decreasing escapeatencies as training progressed (F5,336�126.88, P�0.0001, dataot shown). A three-way ANOVA with quadrant as repeated mea-ures showed a significant effect of CORT (F1,48�4.08, P�0.05)nd lack of a significant effect of SPR (F3,48�1.3, P�0.28) onuadrant search times during the probe test, but a significantORT�SPR�quadrant interaction (F3,48�4.55; P�0.007). The
.c.v. infusions of the different doses of SPR alone did not affecthe time spent in the target and opposite quadrants in comparisono the control group, but SPR blocked the effects of CORT, doseependently (Fig. 3A). Animals given doses of 75 and 150 ng, butot 37.5 ng SPR plus CORT spent significantly more time in theraining (P�0.01) and less time in the opposite quadrantsP�0.01) than rats given SAL�CORT. Moreover, no significantifferences were found in the time spent in the target and oppositeuadrants between groups of SPR (75 or 150 ng) together withORT with either VEH or SPR (75 or 150 ng) plus VEH.
The pattern of results for the number of entrances into thearget quadrant (Fig. 3B) during probe test was similar to that forhe timespent in the target quadrant. A two-way ANOVA revealedsignificant effect of CORT (F1,48�11.73, P�0.005), a significantffect of SPR (F3,48�5.23, P�0.004), and a significant interactionetween both factors (F3,48�3.29, P�0.03). Systemic administra-
ion of CORT decreased significantly the number of entrances intohe target quadrant in comparison with the control groupP�0.01), and this effect was blocked by i.c.v. injections of SPR at5- and 150-ng doses. These results show that the impairingffects of systemic CORT on memory retrieval were blocked by
.c.v. infusions of SPR in a dose-dependent manner.Two-way analysis of total swim paths (Fig. 3C) during the
robe test indicated no significant effects of CORT (F1,48�0.88,�0.36), SPR (F3,48�2.2, P�0.1), or interaction (F3,48�1.94,�0.14). These findings indicate that none of the experimentalanipulations induced any cross disturbances in motor perfor-ance during the probe trial.
DISCUSSION
he purpose of this study was to investigate the role ofrain protein synthesis and corticosteroid receptors in cor-icosterone-induced impairment of memory retrieval. Theesults of the present study indicate that administration ofnisomycin did not change the corticosterone-induced def-
cit in memory retrieval. Moreover, administration of theR, but not GR, antagonist blocked the corticosterone-
nduced response. These findings provide evidence for theiew that glucocorticoids impair memory retrieval throughongenomic mechanisms involving an interaction with
entral MRs.
Fict(q
M. Khaksari et al. / Neuroscience 149 (2007) 729–738 733
ig. 2. Effects of i.c.v. injections of RU38486 on corticosterone-induced impairment of long-term memory retrieval in a water maze task. The i.c.v.nfusions of both doses of RU38486 (75 or 150 ng/5 �l) 60 min before retention testing did not block the impairing effects of systemic injection oforticosterone (1 mg/kg, i.p.) 30 min before retention testing on probe trial retention performance. (A, B) The columns represent the mean (�SEM)ime spent (%) in target (T) and opposite (O) quadrants and number of entrances in the target quadrant of the pool in the 60-s probe test, respectively.C) Total swim path (mean�SEM) in centimeters. * P�0.01 compared with time in training quadrant; ● P�0.01 compared with the corresponding
uadrant in vehicle group that received same treatment; # P�0.01 compared with the vehicle group.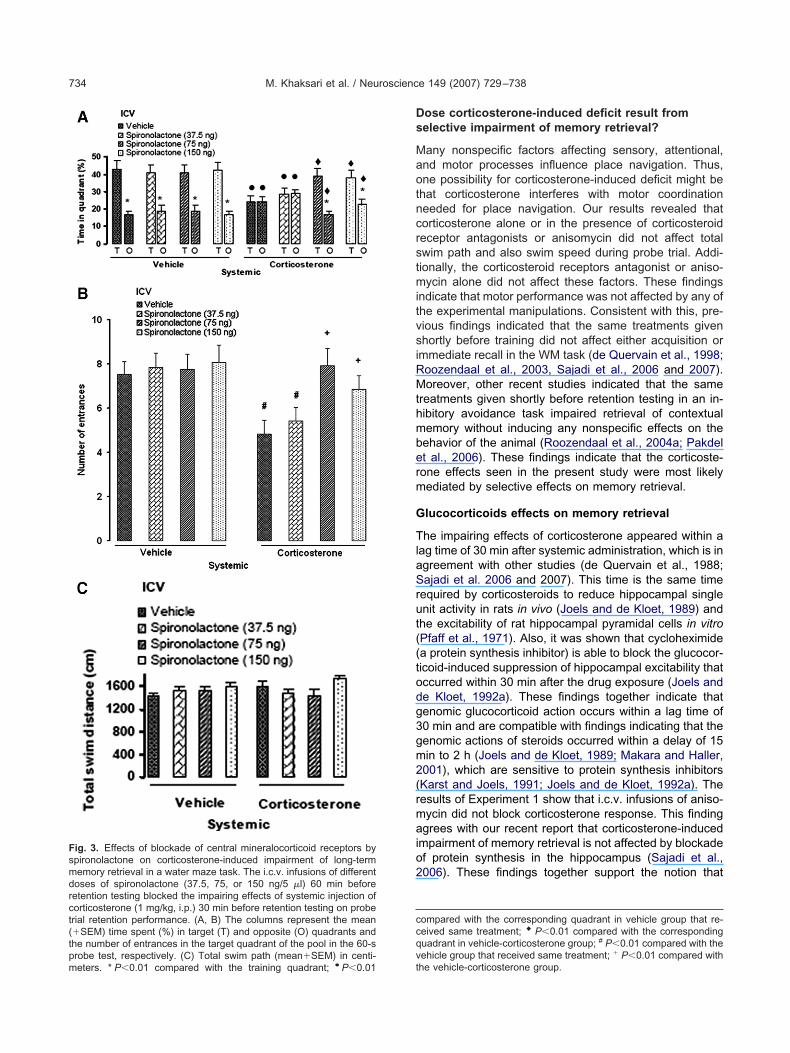
Ds
MaotncrstmitvsiRMthmberm
G
TlaSrut((todg3gm2(rmaio2
Fsmdrct(tprobe test, respectively. (C) Total swim path (mean�SEM) in centi-meters. * P�0.01 compared with the training quadrant; ● P�0.01
ccqvt
M. Khaksari et al. / Neuroscience 149 (2007) 729–738734
ose corticosterone-induced deficit result fromelective impairment of memory retrieval?
any nonspecific factors affecting sensory, attentional,nd motor processes influence place navigation. Thus,ne possibility for corticosterone-induced deficit might behat corticosterone interferes with motor coordinationeeded for place navigation. Our results revealed thatorticosterone alone or in the presence of corticosteroideceptor antagonists or anisomycin did not affect totalwim path and also swim speed during probe trial. Addi-ionally, the corticosteroid receptors antagonist or aniso-ycin alone did not affect these factors. These findings
ndicate that motor performance was not affected by any ofhe experimental manipulations. Consistent with this, pre-ious findings indicated that the same treatments givenhortly before training did not affect either acquisition or
mmediate recall in the WM task (de Quervain et al., 1998;oozendaal et al., 2003, Sajadi et al., 2006 and 2007).oreover, other recent studies indicated that the same
reatments given shortly before retention testing in an in-ibitory avoidance task impaired retrieval of contextualemory without inducing any nonspecific effects on theehavior of the animal (Roozendaal et al., 2004a; Pakdelt al., 2006). These findings indicate that the corticoste-one effects seen in the present study were most likelyediated by selective effects on memory retrieval.
lucocorticoids effects on memory retrieval
he impairing effects of corticosterone appeared within aag time of 30 min after systemic administration, which is ingreement with other studies (de Quervain et al., 1988;ajadi et al. 2006 and 2007). This time is the same time
equired by corticosteroids to reduce hippocampal singlenit activity in rats in vivo (Joels and de Kloet, 1989) andhe excitability of rat hippocampal pyramidal cells in vitroPfaff et al., 1971). Also, it was shown that cycloheximidea protein synthesis inhibitor) is able to block the glucocor-icoid-induced suppression of hippocampal excitability thatccurred within 30 min after the drug exposure (Joels ande Kloet, 1992a). These findings together indicate thatenomic glucocorticoid action occurs within a lag time of0 min and are compatible with findings indicating that theenomic actions of steroids occurred within a delay of 15in to 2 h (Joels and de Kloet, 1989; Makara and Haller,001), which are sensitive to protein synthesis inhibitorsKarst and Joels, 1991; Joels and de Kloet, 1992a). Theesults of Experiment 1 show that i.c.v. infusions of aniso-ycin did not block corticosterone response. This findinggrees with our recent report that corticosterone-induced
mpairment of memory retrieval is not affected by blockadef protein synthesis in the hippocampus (Sajadi et al.,006). These findings together support the notion that
ompared with the corresponding quadrant in vehicle group that re-eived same treatment; � P�0.01 compared with the correspondinguadrant in vehicle-corticosterone group; # P�0.01 compared with the
�
ig. 3. Effects of blockade of central mineralocorticoid receptors bypironolactone on corticosterone-induced impairment of long-termemory retrieval in a water maze task. The i.c.v. infusions of differentoses of spironolactone (37.5, 75, or 150 ng/5 �l) 60 min beforeetention testing blocked the impairing effects of systemic injection oforticosterone (1 mg/kg, i.p.) 30 min before retention testing on proberial retention performance. (A, B) The columns represent the mean�SEM) time spent (%) in target (T) and opposite (O) quadrants andhe number of entrances in the target quadrant of the pool in the 60-s
ehicle group that received same treatment; P�0.01 compared withhe vehicle-corticosterone group.

gm
ibnwbstspetiwtinattsr
cstrHaKpo(lMl
Hi
AsPiceisdethvrw1tt
wmcirrsde
R
Ppvtriadetat3t9papw1anssncrptsttncuStdttbtwl9
M. Khaksari et al. / Neuroscience 149 (2007) 729–738 735
lucocorticoids impair memory retrieval via nongenomicechanisms.
In Experiment 2, we demonstrated that corticosterone-nduced impairment of memory retrieval was not affectedy blockade of GRs, further supporting an involvement ofongenomic mechanisms. This finding is in agreementith results of Oitzl and de Kloet (1992), showing thatlockade of GRs by i.c.v. administration of RU38486hortly before retention test did not influence memory re-rieval in the WM. But our finding is in contrast with a recenttudy indicating that infusions of a GR agonist into hip-ocampus impair spatial memory retrieval (Roozendaalt al., 2003). This discrepancy in results is likely due tohe different experimental procedures. For example, wenjected the GR antagonist into the ventricular system,hich might significantly bind central GRs located
hroughout the brain, while they infused the GR agonistnto the hippocampus. Moreover, blockade of GRs notecessarily producing opposing effects appeared withctivation of GRs. The results of Experiment 3 indicatedhat the effects of corticosterone seem to be mediatedhrough the classical MRs as evidenced by the ability ofpironolactone, an MR antagonist, to block corticoste-one-induced response.
The anatomical sites of interaction between systemicorticosterone and central MRs are not known. Numeroustudies using lesioning and pharmacological methods in-roduced the hippocampus as an important structure inetrieval of spatial memory in rats (Moser and Moser, 1998;olt and Maren, 1999; Steffenach et al., 2002). This areabundantly expresses both GRs and MRs (Reul and deloet,1985; Morimoto et al., 1996). Because the hip-ocampus plays an important role in mediating the effectsf glucocorticoid on retrieval of long-term memoryRoozendaal et al., 2003), in the present study it is moreikely that attenuation of the corticosterone response by
Rs antagonist may be at least due to blockade of MRsocated in the hippocampus.
ow does blockade of MR reverse corticosterone-nduced deficit in memory retrieval?
s aforementioned, MRs have a high affinity for cortico-terone and are almost saturated under basal conditions.revious findings indicate that this dose of corticosterone
nduces, 30 min after injection, plasma concentrations oforticosterone resembling moderate stress (de Quervaint al., 1988). Thus, it is difficult to explain how MR, which
s already occupied with low circulating levels of cortico-terone, is involved in this corticosterone response. Evi-ence from other studies may support our finding. Inter-stingly, a recent in vitro study reported that spironolac-
one is able to block the rapid effects of corticosterone onippocampal cells (Karst et al., 2005), indicating an in-olvement of MRs. In that study, the threshold corticoste-one concentration for rapid effects was about 10 nm,hereas previous electrophysiological studies showed that0- to 20-fold lower concentrations are enough to stimulatehe intracellular MRs (Joels and de Kloet, 1992b). Addi-
ionally, the effects of corticosterone on hippocampal cells fere also detected in the presence of a membrane-imper-eable BSA–corticosterone conjugate, suggesting that
orticosterone might act via membrane receptors. Thus, its possible that this lower affinity of MRs is involved in theapid effects of corticosterone on memory retrieval. Sucholes for MRs in the rapid effects of glucocorticoids onerotonergic transmission and locomotor activity wereemonstrated elsewhere (Meijer and de Kloet, 1995; Sandit al., 1996).
ole of MRs and GRs in memory processing
revious studies indicated that corticosteroid receptorslay differential roles in cognitive behavior. GRs are in-olved in the consolidation of recently acquired informa-ion, whereas MRs are essential for interpretation of envi-onmental stimuli and selection of the appropriate behav-oral response. This is based on studies that used spatialnd nonspatial/contextual tasks. Oitzl and de Kloet (1992)emonstrated that blockade of GRs and MRs alters differ-nt aspects of spatial learning and memory. They foundhat blockade of GRs, but not MRs, before or immediatelyfter training in the water maze, results in impaired reten-ion of spatial memory in rats. In contrast, GR blockade0–45 min before retrieval on day 2 did not change eitherhe swim pattern in the free swim trial (which was done0–120 min later) or the time necessary for finding thelatform (in day 3). In contrast, blockade of MRs did notffect latencies to find platform, but changed the searchattern in the free swim trial significantly. Similar resultsere found in studies on inhibitory avoidance responses of-day-old chicks (Sandi and Rose, 1994) and rats (Oitzl etl., 1993). In our study, administration of the MR antago-ist 60 min before retrieval test did not change the timepent in the quadrant target or the swim pattern. In fact, thewim pattern (Fig. 4) of animals receiving the MR antago-ist was essentially similar to that of the control group. Thisontrasts with the results of Oitzl and de Kloet (1992). Theeasons for this discrepancy may be reflected in the ex-erimental procedures. One important difference betweenhe two studies is the amount of training (6 days withix trails per day vs. 3 days with four trials per day) and,hus, the strength of memory. Previous studies indicatedhat the modulatory action of glucocorticoid receptor ago-ists or antagonists or manipulations of endogenousorticosteroid levels on memory process is dependentpon training intensity (Sandi et al., 1997; Cordero andandi, 1998). Thus, this difference in number of training
rials and the strength of memory may explain theseifferent effects in two studies. Another difference is
ime and treatment dose. We administrated the MR an-agonist spironolactone (37.5, 75, and 150 ng) 60 minefore the retrieval test on day 7 (24 h after the last
raining day) and tested their performance 60 min later,hile Oitzl and de Kloet (1992) administrated spirono-
actone (100 ng) after session 2 and tested animals0 –120 min later in the free swim trials. This may dif-
erentially affect memory retrieval.

Fl
M. Khaksari et al. / Neuroscience 149 (2007) 729–738736
ig. 4. Swimming pattern of rats in the free swim trial randomly chosen from each group. Arrow indicates the starting point; “T” shows the platform
ocation.
Fevmi(cwin(
ARIih
B
C
d
d
E
H
H
J
J
J
K
K
K
L
M
M
M
M
M
O
O
O
O
P
P
P
R
R
R
R
R
R
R
S
S
S
S
M. Khaksari et al. / Neuroscience 149 (2007) 729–738 737
CONCLUSIONS
indings from our laboratory and others provide importantvidence that glucocorticoids influence memory retrievalia rapid mechanisms that do not involve the genome. Theechanisms of such rapid effects of glucocorticoids may
nvolve activation of membrane glucocorticoid receptorspossibly a modified form of MR). In addition, glucocorti-oid effects on memory retrieval may involve an interactionith several neurotransmitter systems in the brain, includ-
ng the adrenergic (Roozendaal et al., 2004 a, b), dopami-ergic (Pakdel and Rashidy-Pour, 2006), and opioidergicRashidy-Pour et al., 2004; Sajadi et al., 2007) systems.
cknowledgments—This work was supported by grants from theesearch Office of Management and Planning Organization of
ran and Semnan University of Medical Sciences to Dr. A. Rash-dy-Pour. We thank Dr. B. Roozendaal for his critical reading andelpful comments on the manuscript.
REFERENCES
elanoff JK, Gross K, Yagar A, Schatzberg AF (2001) Corticosteroidsand cognition. J Psychiatr Res 35:127–145.
ordero MI, Sandi C (1998) A role for brain glucocorticoid receptors incontextual fear conditioning: dependence upon training intensity.Brain Res 786:11–17.
e Quervain DJF, Roozendaal B, McGaugh JL (1998) Stress andglucocorticoids impair retrieval of long term spatial memory. Nature394:787–790.
e Quervain DJF, Roozendaal B, Nitsch RM, McGaugh JL, Hock C(2000) Acute cortisone administration impairs retrieval of long termdeclarative memory in humans. Nat Neurosci 3:313–314.
vans SJ, Murray TF, Moore FJ (2000) Partial purification and bio-chemical characterization of a membrane glucocorticoid receptorfrom an amphibian brain. J Steroid Biochem Mol Biol 72:209–221.
olt W, Maren S (1999) Muscimol inactivation of the dorsal hippocam-pus impairs contextual retrieval of fear memory. J Neurosci19:9054–9062.
ua SY, Chen YZ (1989) Membrane receptor mediated electrophysi-ological effects of glucocorticoid on mammalian neurons. Endocri-nology 124:687–691.
oels M, de Kloet ER (1989) Effects of glucocorticoids and norepi-nephrine on the excitability in the hippocampus. Science 245:1502–1505.
oels M, de Kloet ER (1992a) Control of neuronal excitability bycorticosteroid hormones. Trends Neurosci 15:25–30.
oels M, de Kloet ER (1992b) Coordinative mineralocorticoid andglucocorticoid receptor-mediated control of responses to serotoninin rat hippocampus. Neuroendocrinology 55:344–350.
arst H, Berger S, Turiault M, Troncge F, Schutz G, Joels M (2005)Mineralocorticoid receptors are indispensable for nongenomicmodulation of hippocampal glutamate transmission by corticoste-rone. Proc Natl Acad Sci U S A 102:19204–19027.
arst H, Joels M (1991) The induction of corticosteroid actions onmembrane properties of hippocampal CA1 neurons requires pro-tein synthesis. Neurosci Lett 130:27–31.
uhlmann S, Kirschbaum C, Wolf OT (2004) Effects of oral cortisoltreatment in healthy young women on memory retrieval of negativeand neutral words. Neurobiol Learn Mem 83:158–162.
upien SJ, McEwen BS (1997) The acute effects of corticosteroids oncognition: Integration of animal and human model studies. BrainRes Rev 24:1–27.
akara GB, Haller J (2001) Non-genomic effects of glucocorticoids inthe neural system. Evidence, mechanisms and implication. Prog
Neurobiol 65:367–390.eijer OC, de Kloet ER (1995) A role for the mineralocorticoid receptorin a rapid and transient suppression of hippocampal 5-HT1A re-ceptor mRRA by corticosterone. J Neuroendocrinol 7:653–657.
eiri N, Rosenblum K (1998) Lateral ventricle injection of the proteinsynthesis inhibitor anisomycin impairs long-term memory in a spa-tial memory task. Brain Res 789:48–55.
orimoto M, Morita N, Ozawa H, Yokoyama K, Kawata M (1996)Distribution of glucocorticoid receptor immunoreactivity and mRNAin the rat brain: An immunohistochemical and in situ hybridizationstudy. Neurosci Res 26:235–269.
oser MB, Moser EI (1998) Distributed encoding and retrieval ofspatial memory in the hippocampus. J Neurosci 18:7535–7542.
itzl MS, de Kloet ER (1992) Selective corticosteroid antagonistsmodulates specific aspects of spatial orientation learning. BehavNeurosci 106:62–71.
itzl MS, Fluttert M, de Kloet ER (1994) The effect of corticosterone onreactivity to spatial novelty is mediated by central mineralocorticoidreceptors. Eur J Neurosci 6:1072–1079.
itzl MS, Josephy M, Spruijt BM (1993) An ACTH/MSH(4–9) analogcounteracts the behavioral effects of a mineralocorticoid receptorantagonist. Pharmcol Biochem Behav 44:447–450.
rchinik M, Murray TF, Franklin PH, Morre FL (1992) Guanyl nucleo-tides modulate binding to steroid receptors in neuronal mem-branes. Proc Natl Acad Sci U S A 89:3830–3834.
akdel R, Rashidy-Pour A (2006) Glucocorticoid-induced impairmentof long-term memory retrieval in rats: An interaction with dopamineD2 receptors. Neurobiol Learn Mem 85:300–306.
axino G, Watson C (1998) The rat brain in stereotaxic coordinates(4th ed). Orlando, FL: Academic Press.
faff DW, Silva MTA, Weiss JM (1971) Telemetered recording ofhormone effects on hippocampal neurons. Science 172:394–395.
ashidy-Pour A, Sagedghi H, Taherian AA, Vafaei AA, Fathollahi Y(2004) The effects of acute restraint stress and dexamethasone onretrieval of long term memory in rats: an interaction with opiatesystem. Behav Brain Res 154:193–198.
eul JMHM, de Kloet ER (1985) Two receptor systems for corticoste-rone in rat brain: Microdistribution and differential occupation. En-docrinology 117:2505–2512.
obinson MJF, Franklin KBJ (2007) Effects of anisomycin on consol-idation and reconsolidation of a morphine-conditioned place pref-erence. Behav Brain Res 178:146–153.
oozendaal B (2002) Stress and memory: Opposing effects of glu-cocorticoids on memory consolidation and memory retrieval. Neu-robiol Learn Mem 78:578–595.
oozendaal B, de Quervain DJ, Schelling G, McGaugh JL (2004a) Asystemically administered �-adrenoceptor antagonist blocks corti-costerone-induced impairment of contextual memory retrieval inrats. Neurobiol Learn Mem 81:150–154.
oozendaal B, Hahn EL, Nathan SV, de Quervain DJF, McGaugh JL(2004b) Glucocorticoid effects on memory retrieval require concur-rent noradrenergic activity in the hippocampus and basolateralamygdala. J Neurosci 24:8161–8169.
oozendaal B, Griffith QK, Buranday J, de Quervain DJF, McGaughJL (2003) The hippocampus mediates glucocorticoid induced im-pairment of spatial memory retrieval: Dependence on the basolat-eral amygdala. Proc Natl Acad Sci U S A 100:1328–1333.
andi C, Loscertales M, Guanza C (1997) Experience-dependentfacilitating effect of corticosterone on spatial memory formation inthe water maze. Eur J Neurosci 9:637–642.
andi C, Rose SPR (1994) corticosteroid receptor antagonists areamnesic for passive avoidance in day-old chicks. Eur J Neurosci6:1292–1297.
andi C, Venero C, Guaza C (1996) Novelty-related locomotor effectsof corticosterone in rats. Eur J Neurosci 8:794–800.
ajadi AA, Samaei SA, Rashidy-Pour A (2006) Intra-hippocampalmicroinjections of anisomycin did not block glucocorticoid-inducedimpairment of memory retrieval in rats: An evidence for non-
genomic effects of glucocorticoids. Behav Brain Res 173:158–162.
S
S
S
T
M. Khaksari et al. / Neuroscience 149 (2007) 729–738738
ajadi AA, Samaei SA, Rashidy-Pour A (2007) Intra-hippocampal micro-injections of naltrexone block glucocorticoid-induced impairment ofspatial memory retrieval in rats. Neuropharmacology 6:347–354.
arihi A, Motamedi F, Naghdi N, Rashidy-Pour A (2000) Lidocainereversible inactivation of the median raphe nucleus has no effecton reference memory but enhances working memory versions of
the Morris water maze. Brain Res 114:1–9.teffenach HA, Sloviter RS, Moser EI, Moser MB (2002) Impairedretention of spatial memory after transection of longitudinally ori-ented axons of hippocampal CA3 pyramidal cells. Proc Natl AcadSci U S A 99:3194–3198.
asker JG, Di S, Malcher-Lopes R (2006) Minireview: Rapid glucocor-ticoid signaling via membrane-associated receptors. Endocrinol-
ogy 147:5549–5556.(Accepted 13 August 2007)(Available online 14 August 2007)




















