
Helium-Ion-Induced Radiation Damage in LiNbO_3 Thin Film Electro-Optic Modulators
Upload
independentCategory
view
2download
0
Psychopharmacology (2005) 182: 116–127DOI 10.1007/s00213-005-0064-x
ORIGINAL INVESTIGATION
Eric W. Fish . Joseph F. DeBold . Klaus A. Miczek
Escalated aggression as a reward: corticosterone and GABAAreceptor positive modulators in mice
Received: 1 February 2005 / Accepted: 6 May 2005 / Published online: 13 August 2005# Springer-Verlag 2005
Abstract Rationale: Individuals seek out the opportunityto fight, but the mechanisms behind this positivelyreinforcing effect of aggression have yet to be understood.Objectives: The aims of this study were to (1) describebehavioral and corticosterone elevations that occur inaggressive mice conditioned to respond for the opportu-nity to fight another mouse, (2) determine if corticosteroneelevations are necessary for operant responding andescalated aggression, and (3) determine if corticosteroneelevations alter the aggression-heightening effects of γ-aminobutyric acid (GABA)A receptor positive modulators.Methods and results: Aggressive male CFW mice wereconditioned to respond under the control of a fixed-interval 10-min (FI10) schedule that reinforced their oper-ant behavior by the presentation of an intruder mouse intotheir home cage. After the FI10, aggressive behavior wasca. 75% higher than the species-typical levels of fightingand plasma corticosterone was more than twice as highafter briefly fighting and/or responding on the FI10 sched-ule. Inhibition of corticosterone synthesis by metyrapone(30–100 mg/kg) reduced both conditioned responding as
well as the aggressive behavior after the FI. Although thebenzodiazepine midazolam (0.3–3 mg/kg) heightened spe-cies-typical aggressive behavior, it did not increase thehigh level of aggression engendered by the FI schedule.However, midazolam (0.3 mg/kg) and the neurosteroid al-lopregnanolone (17 mg/kg) both heightened aggressionwhen given after corticosterone synthesis inhibition bymetyrapone (56 mg/kg). Conclusions: These data suggestthat corticosterone elevations are required for respondingthat is motivated by aggressive behavior and for escalatedaggression that follows this responding. Corticosterone ele-vations also appear to inhibit the aggression heighteningeffect of GABAA receptor positive modulators.
Keywords Benzodiazepines . Neurosteroids . Alcohol .HPA activity . Reinforcement schedule . Anxiety . Intruder
Introduction
In 1942, Ginsburg and Allee (1942) showed that prevailingin an aggressive confrontation increases the probability thatan individual will behave aggressively in the future. Thisobservation began the systematic study of the reinforcingeffects of aggression in laboratory animals. Animals aremore likely to approach locations where fights have pre-viously occurred and will occur again (Lagerspetz 1964;Roberts and Keiss 1964; Tellegen et al. 1969; Legrand1970; Tellegen and Horn 1972; Kelsey and Cassidy 1976;Meisel and Joppa 1994; Martinez et al. 1995). However,the clearest evidence that aggression can be reinforcingderives from studies in which birds, fish, mice, rats, andsquirrel monkeys perform responses that are reinforced bythe opportunity for aggression (Thompson 1964, 1969;Azrin et al. 1965; van Hemmel and Myer 1970; Chereket al. 1973; Connor 1974; Fish et al. 2002). When CFWmice responded under the control of a fixed-interval 10-min(FI10) schedule reinforced by the opportunity for aggres-sion, we noted that they were much more aggressive thanwas typical for this strain (Fish et al. 2002). The first ob-jective of the present experiments was to characterize the
E. W. Fish . J. F. DeBold . K. A. MiczekDepartment of Psychology, Tufts University,Medford, MA, USA
K. A. MiczekDepartments of Psychiatry, Pharmacology, and Neuroscience,Tufts University,Boston, MA, USA
E. W. FishDepartment of Neuroscience, Douglas Hospital ResearchCentre, McGill University,Montreal, Quebec, Canada
E. W. Fish (*)Department of Neuroscience,Douglas Hospital Research Centre,6875 Blvd LaSalle,Montreal, Quebec, H4H 1R3, Canadae-mail: [email protected]

escalated aggressive behavior that occurs after the com-pletion of the demands of the FI10 schedule.
Performance of a reinforcement schedule can be highlyarousing and individuals show signs of increased sympa-thetic activation, such as piloerection and motor stimula-tion including “pussyfooting” and “mincing” (Scott 1966;Falk 1971, 1977). These behaviors are also expressed dur-ing aggressive interactions, particularly in the period thatprecedes the initiation of attack (Scott 1958; Grant andMackintosh 1963; Krsiak 1975). Several adjunctive be-haviors such as schedule-induced polydipsia and attackstowards a cage mate have been observed during FI per-formance (Falk 1966; Cherek and Heistad 1971). Neuro-biologically, performance of conditioned behaviors isassociated with catecholaminergic activation (Lewy andSeiden 1972; Emmett-Oglesby et al. 1978; Salamone et al.1994; Ito et al. 2000; Roitman et al. 2004) and hypotha-lamic–pituitary–adrenal axis activity that are perhaps in-volved in both the performance of the behavior (Bronsonand Desjardins 1982; Piazza and Lemoal 1997; Merali etal. 1998; Tomie et al. 2002) and the anticipation of ascheduled reward (Coover et al. 1971; Krieger 1974;Dantzer et al. 1980; Deroche et al. 1997; Mantsch andGoeders 1999; Goeders 2002). The elevated corticoste-rone levels may also affect aggressive behavior; low tomoderate acute changes are associated with increasedaggression in rats (Haller et al. 2000a,b; Kruk et al. 2004),whereas higher, longer-lasting changes are associatedwith decreased aggression in mice (Leshner and Johnson1974; Leshner and Politch 1979). The second objective ofthese experiments was to determine whether respondingfor the opportunity to confront an opponent elevatesplasma corticosterone levels and, if so, whether theseelevations are necessary for the escalation of aggression.
In addition to an activating or permissive role for cor-ticosterone, heightened aggression is thought to be the re-sult of reduced serotonergic activity and enhanced positiveallosteric modulation of the GABAA receptor complex(e.g., Miczek et al. 1995, 2002). Positive allosteric modu-lators of GABAA receptors, such as benzodiazepines,neurosteroids, and alcohol, are among the few agents thatreliably increase aggression in laboratory animals (e.g.,Miczek 1974; Cole and Wolf 1970; Miczek and O’Donnell1980; Ferrari et al. 1997; Miczek et al. 1998; Fish et al.2001; Gourley et al. 2005). Allopregnanolone, a 5α-re-duced metabolite of progesterone, has been shown to in-crease species-typical levels of aggression when given atdoses of 10 and 17 mg/kg (Fish et al. 2001). However, itseffects on escalated aggression after the FI were blunted, inthat the 10-mg/kg dose produced a small increase whereasthe 17-mg/kg dose was ineffective (Fish et al. 2002). Thethird objective of the current study was twofold: first, tocompare the effects of the benzodiazepine midazolam onspecies-typical aggression and aggression that was height-ened by the schedule performance; second, to determine ifblocking the corticosterone rise facilitated GABAA posi-tive modulator effects on aggression.
Materials and methods
Animals
Male CFW mice (n=28; Charles River Breeding Labs,Wilmington, MA, USA) weighed 23–25 g upon arrival andwere housed as “residents” with a female of the samestrain. Over the course of the experiments, the mice gained,on average, between 10 and 15 g. The pairs were housed inclear polycarbonate cages (46×24×15 cm). These cages arelarger than those typically housing paired mice (i.e.,28×17×14 cm) because in a previous experiment usingthe same testing conditions (Fish et al. 2002), we found thatlarger cages were necessary to avoid injury to the“intruders.” Wood shavings lined the floor and Purinarodent chow and tap water were freely available throughthe stainless steel wire lid covering the cage. Except forweekly cage changes, the pairs were not disturbed for 3weeks. Pups born to these pairs were weaned at 3 weeks ofage. “Intruders” were male CFW mice (n=56) that werehoused ten to a cage. The vivarium was maintained at 21±1°C, humidity 30–40%, and a 12-h light/dark photocycle(lights on at 12:00). The Guide for the Care and Use ofLaboratory Animals (http://www.nap.edu/readingroom/books/labrats/) and procedures approved by Tufts Uni-versity’s Institutional Care and Use Committee (IACUC)were followed while caring for the mice.
Apparatus
At the time of testing, an operant conditioning panel(24×15 cm) was inserted into the center of the mouse’scage and secured to the side walls by thumbscrews (Miczekand de Almeida 2001; Fish et al. 2002). The left and rightside of the panel contained a nose-poke sensor and astimulus light that was illuminated to indicate the activeside. Each response briefly turned off the nose-pokesensor’s light. When reinforcement was available, a houselight in the center of the panel was illuminated. Duringexperimental sessions, a polycarbonate lid covered thecage. The panel was connected via a Med Associatesinterface to a personal computer running Windows MedPCsoftware. An experimenter used a specialized keyboard anda PC running The Observer software (Noldus, the Nether-lands) to code the frequency and duration of aggressivebehaviors (i.e., tail rattle, pursuit, sideways threat, andattack bite) and nonaggressive behaviors (i.e., grooming,rearing, walking, and contact with the intruder). Inexperiment III the final 2 min of the FI were analyzed forthe duration of walking, rearing, grooming, time spent nearthe panel (ca. 5 cm) and the duration of time the mouse’snose was in the panel to assess possible sedative effects ofthe drug treatments.
117

Procedure
After 3 weeks of cohabitation with the female, the residentmales were screened for aggressive behaviors. The femaleand any pups were removed from the home cage and agroup-housed male “intruder” mouse was introduced(Miczek and O’Donnell 1978). The confrontation lasted5 min after the first attack bite or, if no attack bite occurred,5 min only. An experimenter counted the frequency ofattack bites using a hand counter. Six confrontations wereconducted at 24-h intervals to allow the levels of aggres-sion toward the intruder to stabilize (Winslow and Miczek1984). By the sixth session, every resident vigorouslyattacked the intruders within 5 s of its presentation. Afterthe initial screenings, the levels of aggression across testdays did not appear to vary systematically with the age ofthe pups in the cage. Each intruder was used repeatedlywith the same resident for approximately a dozen 5-minconfrontations. When new intruders were presented to theresident, one or two 5-min confrontations were conductedto allow the residents to habituate to the new intruders(Winslow and Miczek 1984).
In the next phase of the experiment, the mice wereconditioned to perform a nose-poke task that was rein-forced according to a fixed-interval (FI) schedule. The firstnose poke in the assigned active hole (counterbalancedacross individuals) was reinforced after the interval hadelapsed. On the first day of conditioning, the FI was 30 sand over the next 11 days, the FI was increased to 10 min.When the mouse completed the behavioral demands for theFI schedule, the stimulus lights dimmed, the house lightturned on and an experimenter introduced an intruder intothe cage. After 12 s of vigorous attacks, the house lightdimmed, the intruder was removed, and the session ended.The 12-s duration was selected because it corresponds tothe length of one species-typical bout of sideways threats,attack bites, and tail rattles. Moreover, because of the needfor daily confrontations, a single bout of attacks minimizedthe potential for injury to the intruder and fatigue to theresident. After the fight, the panel was removed and thefemale partner was returned. The next session was con-ducted 24 h later. Five to seven test sessions were con-ducted per week until the mice demonstrated stable ratesof responding that were predominantly in the active holeand when the pattern of responding accelerated towardthe end of the FI (“scalloped”). This pattern was quantifiedusing the index of curvature (Fry et al. 1960), ranging from−0.75, if all the responses were made in first quarter of theinterval, to +0.75, if the responses were made in the finalquarter. If responses occurred equally across all four quar-ters, the index of curvature was 0. In the present experiment,curvature values exceeded +0.30 and were consistent withthose from the previous report (Fish et al. 2002).
Experiment I: schedule-heightened andspecies-typical aggression
Seven mice were conditioned to complete the requirementsof the FI10 schedule to be reinforced by the opportunity foraggression, as above. After response rates and patternsstabilized, schedule-heightened and species-typical aggres-sive behaviors were compared on two occasions in an A–B–A–B design. Species-typical aggression was elicited bypresenting the intruder into the resident’s cage on a daywhen the mice did not perform the nose-poke response,whereas schedule-heightened aggression occurred whenthe intruder was presented immediately after the FI10schedule. All confrontations lasted 5 min from the firstattack bite. Rather than a single bout of aggression, theselonger confrontations allowed the possibility of detectingchanges (both increases and decreases) in aggressive be-haviors. To minimize injury to the intruders, each 5-minconfrontation was separated by 72 h.
Experiment II: plasma corticosterone
Eleven resident male CFW mice were conditioned toperform the FI10 schedule reinforced by the opportunityfor aggression. At the time of testing, their mean rate ofactive responses and index of curvature were 4.1 responses/min and 0.44, respectively. Once a week, blood was takenfrom each mouse after the following events: (1) baseline,when the mice neither nose-poked nor fought; (2) fight,when the mice did not nose-poke, but briefly (ca. 12 s)confronted an intruder; (3) FI, when the mice completedthe FI, but did not fight; and (4) FI plus fight, when themice completed the FI and briefly confronted the intruder.Samples were taken either immediately or 30 min after theevent. All testing occurred 1–3 h before the onset of thelight cycle, at the same time the mice had been conditionedto perform the FI.
Experiment III: blockade of corticosterone synthesis
Ten residents were conditioned as described above. Beforethe drug-administration phase, the mice were habituated toan injection with saline given 1 h before the start of the FI.Schedule-heightened aggression was then verified bycomparing aggression after the FI to aggression in a 5-min species-typical confrontation (i.e., one not preceded bythe FI). Every third day, the mice were given injections ofdifferent doses of the corticosterone synthesis inhibitormetyrapone (30–100 mg/kg i.p.) or saline vehicle 60 minbefore the start of the FI and then reinforced by a 5-minconfrontation with the intruder. The order of drug treat-
118

ments was counterbalanced across mice. On all other days,the mice performed the FI and were reinforced by a 12-sfight.
Experiment IV: midazolam, FI responding,schedule-heightened and species-typical aggression
Experiment IV compared the effects of midazolam onschedule-heightened and species-typical levels of aggres-sion. The seven mice described in experiment I werehabituated to an i.p. saline injection given 10 min beforethe start of the FI. When measuring species-typical aggres-sion (i.e., in a confrontation that had not been preceded bynose poking), midazolam was given 20 min before theconfrontation. When measuring schedule-heightened ag-gression, saline or midazolam (0.3–3.0 mg/kg) was given10 min before the start of the FI (20 min before theconfrontation).
Experiment V: blockade of corticosterone synthesisand GABAA receptor positive modulators
The 10 mice from experiment III were injected on certaintest days with metyrapone (56 mg/kg) 1 h before the start ofthe FI and then given a saline injection or midazolam(0.3 mg/kg) 10 min before the FI. After this, the same micereceived metyrapone with either beta-cyclodextrin orallopregnanolone (17 mg/kg) 15 min before the FI. Thissingle dose of allopregnanolone was selected because inprior experiments, it heightened species-typical (Fish et al.2001), but not schedule-heightened aggression (Fish et al.2002). Aggression was measured for 5 min after comple-tion of the FI. After completing this phase, blood sampleswere taken from these mice to verify the effects of thefollowing treatments on corticosterone levels: (1) salineinjection; (2) saline and completion of the FI10; (3) 56 mg/kg metyrapone and completion of the FI; (4) 56 mg/kgmetyrapone and 0.3 mg/kg midazolam and completion ofthe FI.
Blood sampling and radioimmunoassay
Mice were rapidly anesthetized using isoflurane andcapillary tubes (OD 1.5 mm, VWR International, Willard,OH, USA) were inserted into the retro-orbital sinus tocollect ca. 100 μl of blood. The collection procedure, fromthe isoflurane administration until the mouse awoke, lastedless than 1.5 min. The blood was centrifuged and plasmawas separated and stored at −20°C until radioimmunoas-say. Corticosterone was measured using an 125I kit (ICNPharmaceuticals, Orangeburg, NY, USA) with standardsbetween 25 and 1,000 ng/ml and a 4–7% interassayvariation. To validate the sampling procedure, an initialpilot study found that the levels of plasma corticosterone
sampled from the retro-orbital sinus of an anesthetizedmouse immediately after a 5-min social defeat were similarto those sampled from trunk blood after decapitation (ca200 ng/ml) (Miczek et al. 1999).
Drugs
Allopregnanolone (5α-pregnan-3α-ol-2-one, Steraloids Inc.,Newport, RI, USA) was suspended via sonication in a 20%hydroxypropyl–betacyclodextrin solution (Research Bio-chemicals Inc., Natick, MA, USA). Midazolam maleate(8-chloro-6-(2-fluorophenyl)-1-methyl-4H-imidazo[1,5-a][1,4] and metyrapone (2-methyl-1,2-di(3-pyridyl)-1-pro-panone) were purchased from Sigma (St Louis, MO, USA)and dissolved in saline. The drugs were injected intraper-itoneally in a volume of 1 ml/100 g body weight using a27 G needle. For rapid anesthesia, isoflurane (1-chloro-2,2,2-trifluoroethyl difluoromethyl ether; Henry Schein,Melville, NY, USA) was administered as vapor.
Data analysis
Species-typical levels of fighting were compared toschedule-heightened aggression via paired t tests. A two-way repeated-measures ANOVA was used to determinewhether changes in corticosterone levels depended on timeand the FI/fighting condition and to compare the effectsof midazolam on species-typical and schedule-heightenedaggression. A one-way repeated-measures ANOVA wasused to determine whether metyrapone dose-dependentlyaffected operant responding and schedule-heightened ag-gression. This test was also used to compare the effects ofmidazolam and allopregnanolone after metyrapone treat-ment to those of metyrapone alone. Between-subjectsANOVAs compared the effects of midazolam and allo-pregnanolone after metyrapone to those of thesemodulatorsalone. P values less than 0.05 were considered statisticallysignificant.
Results
Experiment I. Species-typical vs schedule-heightenedaggression
Responding under the control of the FI schedule increasedlevels of aggression toward the intruder. Attack bites [t(6)=2.7, p=0.04; Fig. 1], sideways threats [t(6)=2.8, p=0.03;Fig. 1], and tail rattles [t(6)=4.6, p=0.004; Fig. 1] weresignificantly more frequent when the confrontations fol-lowed the FI compared to when confrontations occurred inthe absence of responding reinforced by aggressive behav-ior (Fig. 1). There was no effect of the FI schedule perfor-mance on the frequency of pursuit or on the nonaggressivebehaviors walking, rearing, or grooming (Table 1).
119

Experiment II: plasma corticosterone
Levels of corticosterone depended on the time of thesample and whether the mice engaged in aggression-reinforced responding and/or fought (F3,30=3.8, p=0.02;Fig. 2). Post hoc analysis revealed that performance underthe control of the FI elevated corticosterone above bothbaseline (p=0.03) and the brief fight alone (p=0.03). Thirtyminutes after completing the FI requirements and brieflyfighting, corticosterone levels were significantly higherthan baseline (p<0.001) and the FI alone (p=0.03) condi-tions. At this time, corticosterone was higher after the brieffight (p=0.054).
Experiment III: blockade of corticosterone synthesis
FI responding
The corticosterone synthesis inhibitor metyrapone dose-dependently altered operant responding and motor behav-iors during the FI. When given at the 100-mg/kg dose(p<0.05), metyrapone reduced the rate of responses in theactive hole (F3,27=16.5, p<0.001; Fig. 3a, Table 2), thepercentage of total responses that were made in the activehole (F3,27=3.4, p=0.03; Table 2), and the duration of timethat the mice spent sniffing the panel (F3,27=29.6, p<0.001;Table 2). There was also a trend for metyrapone to reduceresponding in the inactive hole (F3,27=2.6, p=0.073;Table 2). There were differences in the time spent nearthe panel (F3,27=4.0, p=0.02; Table 2) and the duration ofwalking (F3,27=4.6, p=0.01; Table 2), but no single doseof metyrapone was significantly different from vehicle.Metyrapone did not affect the duration of grooming orrearing or the index of curvature.
Schedule-heightened aggression
Metyrapone dose-dependently reduced the escalated ag-gressive behavior that followed the FI performance. Attackbites (F3,27=16.6, p<0.001; Fig. 3b) were less frequent aftertreatment with all the metyrapone doses. Sideways threats(F3,27=26.9, p<0.001; Fig. 3b) and pursuits (F3,27=8.9,p<0.001; Table 3) were lower after the 30-and 100-mg/kgdoses. Tail rattles (F3,27=7.2, p=0.001; Table 3) were lowerafter the 30-mg/kg dose. Metyrapone also lowered thenonaggressive behaviors walking (F3,27=12.0, p<0.001;Table 3) and rearing (F3,27=3.8, p=0.02; Table 3) but notgrooming. The 30- and 100-mg/kg doses reduced walking,but no single dose reduced rearing significantly from thevehicle treatment.
Experiment IV: midazolam, species-typical andschedule-heightened aggression
Schedule-heightened aggression was confirmed by thedisplay of higher levels of attack bites (t6=2.8, p=0.03;Fig. 4a, inset), sideways threats (t6=2.8, p=0.03; Fig. 4b,inset) and tail rattles (t6=4.6, p=0.004, data not shown) butnot pursuit. To accommodate the differences in baselinerate of behavior, these data were normalized to a percent ofthe respective vehicle treatments. The effects of midazolamon attack bites (F3,18=4.5, p=0.02; Fig. 4a) and sidewaysthreats (F3,18=5.8, p=0.006; Fig. 4b), but not tail rattles orpursuits, depended on whether the confrontation occurredafter responding that was reinforced by fighting or not. Posthoc Tukey’s tests determined that the 0.3- and 1.0-mg/kgdose of midazolam significantly increased species-typical,but not schedule-heightened, attack bites and sidewaysthreats. The 3.0-mg/kg dose decreased attack bites, side-ways threats, and the duration of walking (F3,18=13.3,p<0.001; data not shown) and these effects did not depend
Table 1 Behaviors during the confrontation that were not affectedby the demands of the FI10 schedule
Confrontation Species-typical Schedule-heightened
Pursuits 5.4±1.3 7.8±2.3Walk (s) 80.6±8.8 95.4±2.9Rear (s) 16.5±4.4 14.6±4.2Groom (s) 26.2±6.7 22.2±3.0
Data for each behavior are mean±SEM
Fig. 1 A comparison between species-typical and schedule-heightened aggressive behaviors. Mean (± SEM, vertical lines)frequency of tail rattles (left bars), attack bites (center bars), andsideways threats (right bars) after two kinds of aggressive con-frontations. The open bars represent the data from the species-typ-ical levels that result when a resident confronts an intruder in thehome cage. The filled gray bars represent data from the schedule-heightened levels that result when the resident has completed theFI10 schedule of reinforcement. Asterisks (*) indicate a higherfrequency relative to the species-typical level
120

on whether the confrontation was species typical or sched-ule heightened. No dose of midazolam decreased groomingor rearing significantly from saline vehicle.
During the FI, the rate of responding in the active hole(F3,18=3.3, p=0.04; data not shown) and the percentage ofreinforced responding (F3,18=3.0, p=0.05) were affected bymidazolam, but no single dose was significantly differentfrom vehicle. The 0.3-mg/kg dose produced a smallincrease in the rate of responding, whereas the 3.0-mg/kgdose produced a small decrease in the rate of responding.Midazolam affected neither curvature, the rate of respond-ing in the nonreinforced hole, nor the percentage of re-sponses on the nonreinforced side.
Experiment V: midazolam, allopregnanolone,escalated aggression, and corticosterone
After treatment with the 56-mg/kg dose of metyrapone, the0.3-mg/kg dose of midazolam and the 17-mg/kg dose ofallopregnanolone increased the frequency of attack bites(F3,27=14.3, p<0.001; Fig. 5) and sideways threats (F3,27=9.3, p<0.001; Fig. 5) significantly above the vehicle base-line. Midazolam also increased the frequency of pursuit(F3,27=7.2, p=0.001; data not shown) and there was a trendfor allopregnanolone to reduce tail rattles (F3,27=2.4, p=0.08; data not shown). Walking (F3,27=7.2, p<0.001; datanot shown) was increased by allopregnanolone, but neitherdrug affected rearing or grooming. An additional analysiswas performed to explore metyrapone’s modulation ofmidazolam’s and allopregnanolone’s effects on aggressivebehaviors. Data from previous experiments in which the
0.3-mg/kg dose of midazolam (experiment IV) or the 17-mg/kg dose of allopregnanolone (Fish et al. 2002) weregiven alone were compared to the current experiment inwhich these compounds were given after treatment withmetyrapone. Attack bites (F3,33=4.5, p=0.009; Fig. 5) andsideways threats (F3,33=5.6, p=0.003; Fig. 5) were sig-nificantly greater when allopregnanolone and midazolamwere given after metyrapone treatment thanwhen theyweregiven alone.
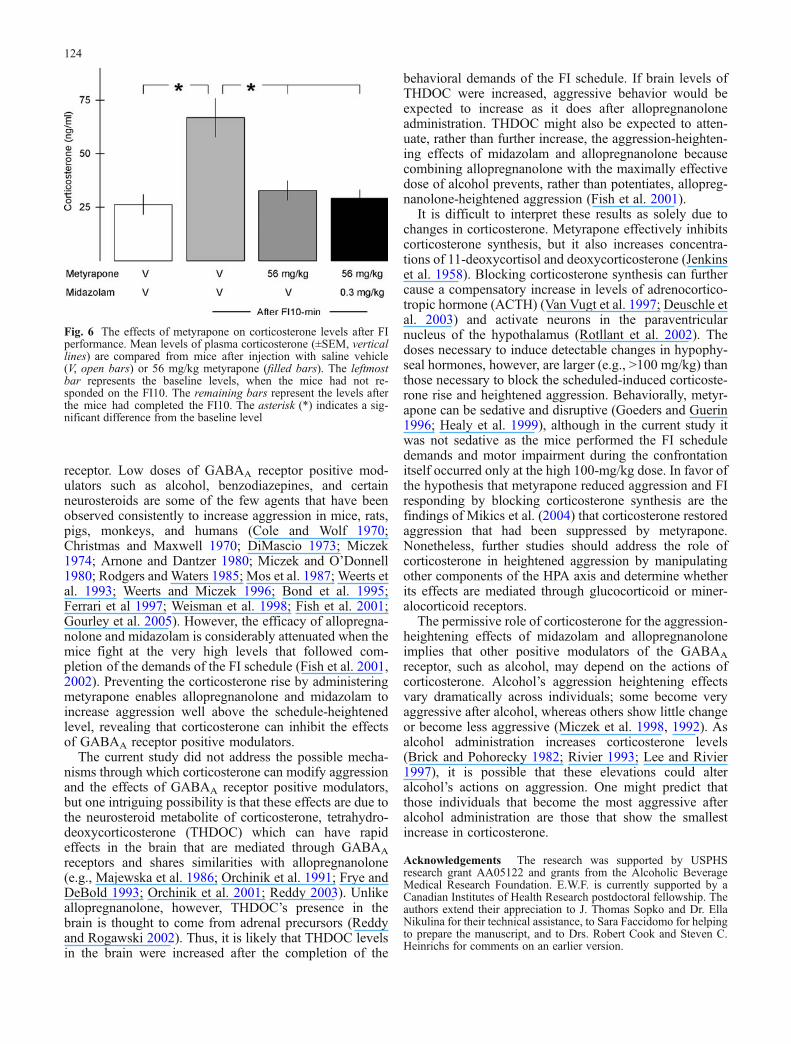
Corticosterone levels (F3,27=15.2, p<0.001; Fig. 6) weresignificantly increased after completion of the FI10performance and injection of saline but not after pretreat-ment with 56-mg/kg metyrapone.
Discussion
Animals will work for the opportunity to engage in anaggressive confrontation. As mice complete the require-ments of an FI10 schedule to be reinforced by access to anopponent, they are much more aggressive during thesubsequent confrontation than in the absence of thisbehavioral demand. The escalated aggression is similar tothe increases that occur after other behavioral manipula-tions, such as attack “priming” (Potegal 1984; Potegal andTenbrink 1984) or social instigation (Fish et al. 1999), andomission of scheduled reinforcement (i.e., “frustration”;Dollard et al. 1939; de Almeida and Miczek 2002).Concurrent with the increase in aggression, the mice showa small but significant rise in corticosterone, and theybecome less sensitive to the aggression heightening effectsof GABAA receptor positive modulators, midazolam and
Fig. 2 The effects of FI performance and fighting on corticoste-rone levels. The mean (±SEM, vertical lines) levels of plasma corti-costerone (ng/ml) are shown from resident mice immediately, or30 min after four different events. a The baseline levels at the time ofday when mice are conditioned to respond on the FI10 schedule, b thelevels after a brief attack flurry, c the levels after completing the
FI, but when no fight has occurred, and d the levels that occurafter completing the FI and engaging in a brief fight. The openbars represent corticosterone levels when the mice have not re-sponded on the FI10, and the filled gray bars represent cortico-sterone levels after completion of the FI10. Asterisks (*) indicatelevels that are significantly different
121

allopregnanolone. The changes in aggressive behaviorappear to critically depend on the corticosterone risebecause blockade of corticosterone synthesis reduces theschedule-heightened aggression and restores heightenedaggression after midazolam and allopregnanolone. Thepresent experiments highlight a dual role of corticosteronein anticipatory changes of aggression and in attenuating theactions of GABAA receptor positive modulators.
Corticosterone elevations occur after many significantphysiological events including operant performance forseveral rewards, such as food and drug administration(Coover et al. 1971; Dantzer et al. 1980; Deroche et al.1997; Mantsch and Goeders 1999; Goeders 2002). Thepresent studies show that corticosterone level increasesduring operant responding and also as a consequence of avery brief fight. Baseline measurements of corticosterone(ca. 25 ng/ml) were obtained when the mice expected theconditioning session, which rules out temporal condition-ing (Krieger 1974) and circadian and ultradian rhythms(Haller et al. 2000a) as likely explanations for the rise.Rather, the increase appears to be mediated by at least thepresentation of the conditioning panel, and it is not knownwhether responding was necessary to elevate corticoste-rone. Previous studies have shown that corticosterone isinvolved in the maintenance of operant responding and hasits own reinforcing effects (Deroche et al. 1993; Piazzaet al. 1993). The present experiments also show that cor-ticosterone is involved in responding for aggression. The100-mg/kg dose of metyrapone reduced responding foraggression, but also reduced the duration of walking andsniffing the operant panel.Although these behavioral changesmay suggest motor impairment, metyrapone did not affectgrooming and rearing. Overall, the results are consistentwith the hypothesis that corticosterone has relatively rapideffects, which are necessary for the performance of re-inforced behaviors.
The largest change in corticosterone occurred 30 minafter the mice responded for aggression and engaged in thebrief fight. The function of corticosterone at this time pointwas not addressed in the current studies, but it may in-corporate long-term genomic mechanisms that are impor-tant in the recovery from a significant event like aggression(e.g., McEwen and Wingfield 2003). The rise in cortico-sterone during operant responding for the opportunity to
Fig. 3 The effects of blocking corticosterone synthesis on FIperformance and aggressive behaviors. Effects of different doses ofmetyrapone are compared to the saline vehicle (V) on FI10 respond-ing for the opportunity to confront an intruder mouse (a) and theaggressive behaviors that occur after completion of the interval (b).In a, the filled circles (left y axis) represent the mean rate ofresponding in the active hole and the filled squares (right y axis)represent the mean indices of curvature. In b, the filled upside downtriangles (left y axis) represent the mean frequency of attack bitesand the filled triangles represent the mean frequency of sidewaysthreats. Vertical lines represent±SEM. Asterisks (*) indicate levelsthat differ from the vehicle (V) treatment
Table 2 Effects of metyrapone on behaviors during the FI
Metyrapone dose (mg/kg)
Vehicle 30.0 56.0 100.0
Responses perminute, activehole
6.4±0.5 5.3±1.0 5.1±0.5 1.4±0.3
Responses perminute, inactivehole
1.0±0.2 1.1±0.3 0.9±0.2 0.4±0.1
% Active hole 87.6±1.8 84.9±2.6 85.5±2.2 76.1±4.3Index of curvature 0.36±0.01 0.35±0.06 0.35±0.03 0.29±0.07Groom (s) 3.3±0.8 4.4±1.3 4.7±2.2 6.2±2.7Rear (s) 40.1±2.3 42.9±4.1 43.9±3.6 35.3±3.6Walk (s) 25.5±1.4 27.2±2.7 20.6±1.7 32.3±4.6Near panel (s) 87.6±2.0 86.5±4.0 100.6±4.8 71.6±9.5Nose in hole (s) 19.6±1.2 16.3±1.9 17.3±1.4 7.0±1.8
Data for each behavior are mean±SEM. Emboldened values arestatistically significant from vehicle (p<0.05)
Table 3 Effects of metyrapone on behaviors during the confron-tation
Dose (mg/kg)
Vehicle 30.0 56.0 100.0
Pursuits 8.4±1.6 3.2±1.5 5.6±1.5 2.0±0.8Tail rattles 16.1±2.5 25.9±3.9 13.9±3.7 10.5±1.8Walk (s) 95.6±4.9 75.6±5.5 90.6±4.7 67.2±5.7Rear (s) 30.1±5.5 20.4±6.5 37.7±8.0 20.4±6.4Groom (s) 17.7±2.4 26.6±3.8 15.6±2.7 15.2±8.0
Data for each behavior are mean±SEM. Emboldened values arestatistically significant from vehicle (p<0.05)
122

fight appears to contribute to the high level of fighting thatoccurs in the subsequent confrontation because blockingthis rise with metyrapone reduced the escalated aggressionto the species-typical level (35 vs 17 bites/5 min). Theseresults are consistent with several lines of evidence indi-cating that corticosterone can acutely facilitate aggressivebehavior. Circadian and ultradian variations in aggressivebehavior (e.g., Sofia and Salama 1970) vary closely withcorticosterone levels in rats (Haller et al. 2000a,b) andhamsters (Landau 1975a,b). Systemic injection of cortico-sterone in rats facilitated aggression induced by electricalstimulation of the anterior hypothalamus (Kruk et al. 2004).Microinjection of cortisol (36 pg) or its precursor deoxy-corticosterone (33 ng) into this region also increased ag-gressive behaviors in male hamsters (Hayden-Hixson andFerris 1991), whereas higher concentrations of deoxycor-ticosterone were suppressive (Kohlert and Meisel 2001).Inhibition of corticosterone synthesis or blockade of themineralocorticoid receptor both reduce offensive aggres-sion in experienced rats (Haller et al. 2000a,b; Mikics et al.2004). In addition, α2 adrenergic antagonists increase
aggression in experienced rats by stimulating the release ofcorticosterone (Haller et al. 1994). Similar examples of highlevels of aggression in the presence of high levels ofcorticosterone have been shown following “frustration”(Coover et al. 1971; Dantzer et al. 1980) in mice lacking thegene for neural cell adhesion molecule (Stork et al. 1997)and rats after repeated experience with restraint stress(Wood et al. 2003). Together, these results indicate thatcorticosterone can facilitate aggressive behaviors, althoughthe precise mechanism for this effect is presently notknown.
A second effect of the increase in corticosterone is theinhibition of the aggression-heightening effects of theGABAA receptor positive modulators midazolam andallopregnanolone. This finding suggests that whereas acorticosterone rise facilitates aggression, it can also preventfurther escalations by positive modulation of the GABAA
Fig. 4 Midazolam’s effects on two kinds of aggressive behavior.Effects of different doses of midazolam on the schedule-heightenedaggressive behaviors that occur following completion of the FI10(filled bars and circles) are compared to its effects on the species-typical aggressive behaviors (open bars and circles). The data areexpressed as a percent of the saline vehicle (V) baseline. The un-transformed baseline data are portrayed in the inset figures. Top,mean (±SEM) frequency of attack bites; bottom, sideways threats.Asterisks (*) indicate a significant difference between species-typical and schedule heightened aggression
Fig. 5 The effects of GABAA receptor positive modulators aftertreatment with metyrapone. This figure portrays the effects ofallopregnanolone (Allo, 17 mg/kg) and midazolam (MDZ, 0.3 mg/kg) on the mean levels (± SEM, vertical lines) of scheduleheightened attack bites (a) and sideways threats (b). The open barsrepresent the mean frequency of behavior after treatment with thesaline or beta-cyclodextrin vehicle (V) and the filled bars representthe mean frequency of behavior after the GABAA receptor positivemodulators. The left bars portray previous results when allopregna-nolone and midazolam are given alone and the right bars representthe behaviors when allopregnanolone and midazolam are given afterprior treatment with 56 mg/kg metyrapone. Asterisks (*) indicate asignificant difference from the vehicle and respective drug treat-ments without prior metyrapone treatment
123
receptor. Low doses of GABAA receptor positive mod-ulators such as alcohol, benzodiazepines, and certainneurosteroids are some of the few agents that have beenobserved consistently to increase aggression in mice, rats,pigs, monkeys, and humans (Cole and Wolf 1970;Christmas and Maxwell 1970; DiMascio 1973; Miczek1974; Arnone and Dantzer 1980; Miczek and O’Donnell1980; Rodgers and Waters 1985; Mos et al. 1987; Weerts etal. 1993; Weerts and Miczek 1996; Bond et al. 1995;Ferrari et al 1997; Weisman et al. 1998; Fish et al. 2001;Gourley et al. 2005). However, the efficacy of allopregna-nolone and midazolam is considerably attenuated when themice fight at the very high levels that followed com-pletion of the demands of the FI schedule (Fish et al. 2001,2002). Preventing the corticosterone rise by administeringmetyrapone enables allopregnanolone and midazolam toincrease aggression well above the schedule-heightenedlevel, revealing that corticosterone can inhibit the effectsof GABAA receptor positive modulators.
The current study did not address the possible mecha-nisms through which corticosterone can modify aggressionand the effects of GABAA receptor positive modulators,but one intriguing possibility is that these effects are due tothe neurosteroid metabolite of corticosterone, tetrahydro-deoxycorticosterone (THDOC) which can have rapideffects in the brain that are mediated through GABAA
receptors and shares similarities with allopregnanolone(e.g., Majewska et al. 1986; Orchinik et al. 1991; Frye andDeBold 1993; Orchinik et al. 2001; Reddy 2003). Unlikeallopregnanolone, however, THDOC’s presence in thebrain is thought to come from adrenal precursors (Reddyand Rogawski 2002). Thus, it is likely that THDOC levelsin the brain were increased after the completion of the
behavioral demands of the FI schedule. If brain levels ofTHDOC were increased, aggressive behavior would beexpected to increase as it does after allopregnanoloneadministration. THDOC might also be expected to atten-uate, rather than further increase, the aggression-heighten-ing effects of midazolam and allopregnanolone becausecombining allopregnanolone with the maximally effectivedose of alcohol prevents, rather than potentiates, allopreg-nanolone-heightened aggression (Fish et al. 2001).
It is difficult to interpret these results as solely due tochanges in corticosterone. Metyrapone effectively inhibitscorticosterone synthesis, but it also increases concentra-tions of 11-deoxycortisol and deoxycorticosterone (Jenkinset al. 1958). Blocking corticosterone synthesis can furthercause a compensatory increase in levels of adrenocortico-tropic hormone (ACTH) (Van Vugt et al. 1997; Deuschle etal. 2003) and activate neurons in the paraventricularnucleus of the hypothalamus (Rotllant et al. 2002). Thedoses necessary to induce detectable changes in hypophy-seal hormones, however, are larger (e.g., >100 mg/kg) thanthose necessary to block the scheduled-induced corticoste-rone rise and heightened aggression. Behaviorally, metyr-apone can be sedative and disruptive (Goeders and Guerin1996; Healy et al. 1999), although in the current study itwas not sedative as the mice performed the FI scheduledemands and motor impairment during the confrontationitself occurred only at the high 100-mg/kg dose. In favor ofthe hypothesis that metyrapone reduced aggression and FIresponding by blocking corticosterone synthesis are thefindings of Mikics et al. (2004) that corticosterone restoredaggression that had been suppressed by metyrapone.Nonetheless, further studies should address the role ofcorticosterone in heightened aggression by manipulatingother components of the HPA axis and determine whetherits effects are mediated through glucocorticoid or miner-alocorticoid receptors.
The permissive role of corticosterone for the aggression-heightening effects of midazolam and allopregnanoloneimplies that other positive modulators of the GABAA
receptor, such as alcohol, may depend on the actions ofcorticosterone. Alcohol’s aggression heightening effectsvary dramatically across individuals; some become veryaggressive after alcohol, whereas others show little changeor become less aggressive (Miczek et al. 1998, 1992). Asalcohol administration increases corticosterone levels(Brick and Pohorecky 1982; Rivier 1993; Lee and Rivier1997), it is possible that these elevations could alteralcohol’s actions on aggression. One might predict thatthose individuals that become the most aggressive afteralcohol administration are those that show the smallestincrease in corticosterone.
Acknowledgements The research was supported by USPHSresearch grant AA05122 and grants from the Alcoholic BeverageMedical Research Foundation. E.W.F. is currently supported by aCanadian Institutes of Health Research postdoctoral fellowship. Theauthors extend their appreciation to J. Thomas Sopko and Dr. EllaNikulina for their technical assistance, to Sara Faccidomo for helpingto prepare the manuscript, and to Drs. Robert Cook and Steven C.Heinrichs for comments on an earlier version.
Fig. 6 The effects of metyrapone on corticosterone levels after FIperformance. Mean levels of plasma corticosterone (±SEM, verticallines) are compared from mice after injection with saline vehicle(V, open bars) or 56 mg/kg metyrapone (filled bars). The leftmostbar represents the baseline levels, when the mice had not re-sponded on the FI10. The remaining bars represent the levels afterthe mice had completed the FI10. The asterisk (*) indicates a sig-nificant difference from the baseline level
124

References
Arnone M, Dantzer R (1980) Effects of diazepam on extinctioninduced aggression in pigs. Pharmacol Biochem Behav 13:27–30
Azrin NH, Hutchinson RR, McLaughlin R (1965) The opportunityfor aggression as an operant reinforcer during aversive stim-ulation. J Exp Anal Behav 8:171–180
Bond AJ, Curran HV, Bruce MS, O’Sullivan G, Shine P (1995)Behavioural aggression in panic disorder after 8 weeks’treatment with alprazolam. J Affect Disord 35:117–123
Brick J, Pohorecky LA (1982) Ethanol–stress interaction: biochem-ical findings. Psychopharmacology 77:81–84
Bronson FH, Desjardins C (1982) Endocrine responses to sexualarousal in male mice. Endocrinology 111:1286–1291
Cherek DR, Heistad GT (1971) Fixed-interval induced aggression.Psychon Sci 25:7–8
Cherek DR, Thompson T, Heistad GT (1973) Responding main-tained by the opportunity to attack during an interval food re-inforcement schedule. J Exp Anal Behav 19:113–123
Christmas AJ, Maxwell DR (1970) A comparison of the effects ofsome benzodiazepines and other drugs on aggressive and ex-ploratory behaviour in mice and rats. Neuropharmacology 9:17–29
Cole HF, Wolf HH (1970) Laboratory evaluation of aggressivebehavior of the grasshopper mouse (Onychomys). J PharmacolSci 59:969–971
Connor JL (1974) Waning and recovery of conspecific aggression inthe house mouse (Mus musculus L). J Comp Physiol Psychol87:215–227
Coover GD, Goldman L, Levine S (1971) Plasma corticosteroneincreases produced by extinction of operant behavior in rats.Physiol Behav 6:261–263
Dantzer R, Arnone M, Mormede P (1980) Effects of frustration onbehaviour and plasma corticosteroid levels in pigs. PhysiolBehav 24:1–4
de Almeida RMM, Miczek KA (2002) Aggression escalated bysocial instigation or by discontinuation of reinforcement(“frustration”) in mice: inhibition by anpirtoline-a 5-HT1Breceptor agonist. Neuropsychopharmacology 27:171–181
Deroche V, Piazza PV, Deminiere JM, Le Moal M, Simon H (1993)Rats orally self-administer corticosterone. Brain Res 622:315–320
Deroche V, Marinelli M, Le Moal M, Piazza PV (1997) Gluco-corticoids and behavioral effects of psychostimulants. II: co-caine intravenous self-administration and reinstatement dependon glucocorticoid levels. J Pharmacol Exp Ther 281: 1401–1407
Deuschle M, Lecei O, Stalla GK, Landgraf R, Hamann B,Lederbogen F, Uhr M, Luppa P, Maras A, Colla M, Heuser I(2003) Steroid synthesis inhibition with ketoconazole and itseffect upon the regulation of the hypothalamus–pituitary–adrenal system in healthy humans. Neuropsychopharmacology28:379–383
DiMascio A (1973) The effects of benzodiazepines on aggression:reduced or increased? Psychopharmacologia 30:95–102
Dollard J, Doob L, Miller N, Mowrer O, Sears R (1939) Frustrationand aggression. Yale University Press, New Haven
Emmett-Oglesby MW, Lewy AJ, Albert LH, Seiden LS (1978) Roleof lever responding and water presentation in altering rat braincatecholamine metabolism. J Pharmacol Exp Ther 204:406–415
Falk JL (1966) Schedule-induced polydipsia as a function of fixedinterval length. J Exp Anal Behav 9:37–39
Falk JL (1971) The nature and determinants of adjunctive behavior.Physiol Behav 6:577–588
Falk JL (1977) The origin and functions of adjunctive behavior.Anim Learn Behav 5:325–335
Ferrari PF, Parmigiani S, Rodgers RJ, Palanza P (1997) Differentialeffects of chlordiazepoxide on aggressive behavior in malemice: the influence of social factors. Psychopharmacology134:258–265
Fish EW, Faccidomo S, Miczek KA (1999) Aggression heightenedby alcohol or social instigation in mice: reduction by the 5-HT1B receptor agonist CP-94,253. Psychopharmacology 146:391–399
Fish EW, Faccidomo S, DeBold JF, Miczek KA (2001) Alcohol,allopregnanolone and aggression in mice. Psychopharmacology153:473–483
Fish EW, DeBold JF, Miczek KA (2002) Aggressive behavior as areinforcer in mice: activation by allopregnanolone. Psycho-pharmacology 163:459–466
Fry W, Kelleher RT, Cook L (1960) A mathematical index ofperformance on fixed-interval schedules of reinforcement. JExp Anal Behav 3:193–199
Frye CA, DeBold JF (1993) 3-α-OH-DHP and 5-α-THDOCimplants to the ventral tegmental area facilitate sexual recep-tivity in hamsters after progesterone priming to the ventral medialhypothalamus. Brain Res 612:130–137
Ginsburg B, Allee WC (1942) Some effects of conditioning onsocial dominance and subordination in inbred strains of mice.Physiol Zool 15:485–506
Goeders NE (2002) The HPA axis and cocaine reinforcement.Psychoneuroendocrinology 27:13–33
Goeders NE, Guerin GF (1996) Role of corticosterone in intrave-nous cocaine self-administration in rats. Neuroendocrinology64(5):337–348
Gourley SL, DeBold JF, Yin W, Cook J, Miczek KA (2005)Benzodiazepines and heightened aggressive behavior in rats:reduction by GABAA/alpha1 receptor antagonists. Psychophar-macology (in press)
Grant EC, Mackintosh JH (1963) A comparison of the social pos-tures of some common laboratory rodents. Behaviour 21: 246–295
Haller J, Barna I, Kovacs JL (1994) Alpha2-adrenoceptor blockade,pituitary–adrenal hormones, and agonistic interactions in rats.Psychopharmacology 115:478–484
Haller J, Halasz J, Mikics E, Kruk MR, Makara GB (2000a)Ultradian corticosterone rhythm and the propensity to behaveaggressively in male rats. J Neuroendocrinol 12:937–940
Haller J, Millar S, van de SJ, de Kloet RE, Kruk MR (2000b) Theactive phase-related increase in corticosterone and aggressionare linked. J Neuroendocrinol 12:431–436
Hayden-Hixson DM, Ferris CF (1991) Cortisol exerts site-, context-and dose-dependent effects on agonistic responding in ham-sters. J Neuroendocrinol 3:613–622
Healy DG, Harkin A, Cryan JF, Kelly JP, Leonard BE (1999)Metyrapone displays antidepressant-like properties in preclini-cal paradigms. Psychopharmacology 145:303–308
Ito R, Dalley JW, Howes SR, Robbins TW, Everitt BJ (2000)Dissociation in conditioned dopamine release in the nucleusaccumbens core and shell in response to cocaine cues andduring cocaine-seeking behavior in rats. J Neurosci 20:7489–7495
Jenkins JS, Meakin JW, Nelson DH, Thorn GW (1958) Inhibition ofadrenal steroid 11-oxygenation in the dog. Science 128:478–480
Kelsey JE, Cassidy D (1976) The reinforcing properties of aggres-sive vs. nonaggressive social interactions in isolated male ICRmice (Mus musculus). Aggress Behav 2:275–284
Kohlert JG, Meisel RL (2001) Inhibition of aggression by proges-terone and its metabolites in female Syrian hamsters. AggressBehav 27:372–381
Krieger DT (1974) Food and water restriction shifts corticosterone,temperature, activity and brain amine periodicity. Endocrinol-ogy 95:1195–1201
Krsiak M (1975) Tail rattling in aggressive mice as a measure oftranquilizing activity of drugs. Act Nerv Super 17:225–226
Kruk MR, Halasz J, Weelis W, Haller J (2004) Fast positivefeedback between adrenocortical stress response and a brainmechanism involved in aggressive behavior. Behav Neurosci118:1062–1070
Lagerspetz K (1964) Studies on the aggressive behaviour of mice.Ann Acad Sci Fenn 131:1–131
125

Landau IT (1975a) Light–dark rhythms in aggressive behavior ofthe male golden hamster. Physiol Behav 14:767–774
Landau IT (1975b) Effects of adrenalectomy on rhythmic and non-rhythmic aggressive behavior in the male golden hamster.Physiol Behav 14:775–780
Lee S, Rivier C (1997) An initial, 3-day-long treatment with alcoholinduces a long-lasting phenomenon of selective tolerance inthe activity of the rat hypothalamic–pituitary–adrenal axis. JNeurosci 17:8856–8866
Legrand R (1970) Successful aggression as the reinforcer forrunway behavior of mice. Psychon Sci 20:303–305
Leshner AI, Johnson AE (1974) Effects of adrenalectomy on theaggressiveness of neonatally androgenized female mice.Physiol Behav 13:703–705
Leshner AI, Politch JA (1979) Hormonal control of submissivenessin mice: irrelevance of the androgens and relevance of thepituitary–adrenal hormones. Physiol Behav 22:531–534
Lewy AJ, Seiden LS (1972) Operant behavior changes norepineph-rine metabolism in rat brain. Science 175:454–456
Majewska MD, Harrison NL, Schwartz RD, Barker JL, Paul SM(1986) Steroid hormone metabolites are barbiturate-likemodulators of the GABA receptor. Science 232:1004–1007
Mantsch JR, Goeders NE (1999) Ketoconazole blocks the stress-induced reinstatement of cocaine-seeking behavior in rats:relationship to the discriminative stimulus effects of cocaine.Psychopharmacology 142:399–407
Martinez M, Guillen-Salazar F, Salvador A, Simon VM (1995)Successful intermale aggression and conditioned place pref-erence in mice. Physiol Behav 58:323–328
McEwen BS, Wingfield JC (2003) The concept of allostasis inbiology and biomedicine. Horm Behav 43:2–15
Meisel RL, Joppa MA (1994) Conditioned place preference infemale hamsters following aggressive or sexual encounters.Physiol Behav 56:1115–1118
Merali Z, McIntosh J, Kent P, Michaud D, Anisman H (1998)Aversive and appetitive events evoke the release of corticotro-pin-releasing hormone and bombesin-like peptides at thecentral nucleus of the amygdala. J Neurosci 18:4758–4766
Miczek KA (1974) Intraspecies aggression in rats: effects of d-amphetamine and chlordiazepoxide. Psychopharmacologia 39:275–301
Miczek KA, de Almeida RMM (2001) Oral drug self-administrationin the home cage of mice: alcohol-heightened aggression andinhibition by the 5-HT1B agonist anpirtoline. Psychopharma-cology (Berl) 157:421–429
Miczek KA, O’Donnell JM (1978) Intruder-evoked aggression inisolated and nonisolated mice: effects of psychomotor stimu-lants and L-dopa. Psychopharmacology 57:47–55
Miczek KA, O’Donnell JM (1980) Alcohol and chlordiazepoxideincrease suppressed aggression in mice. Psychopharmacology69:39–44
Miczek KA, Weerts EM, Tornatzky W, DeBold JF, Vatne TM (1992)Alcohol and “bursts” of aggressive behavior: ethologicalanalysis of individual differences in rats. Psychopharmacology107:551–563
Miczek KA, Weerts EM, Vivian JA, Barros HM (1995) Aggression,anxiety and vocalizations in animals: GABA(A) and 5-HTanxiolytics. Psychopharmacology 121:38–56
Miczek KA, Barros HM, Sakoda L, Weerts EM (1998) Alcohol andheightened aggression in individual mice. Alcohol Clin ExpRes 22:1698–1705
Miczek KA, Nikulina E, Kream R, Carter G, Espejo E (1999)Behavioral sensitization to cocaine after a brief social defeatstress: c-fos expression in the PAG. Psychopharmacology 141:225–234
Miczek KA, Fish EW, DeBold JF, de Almeida RMM (2002) Socialand neural determinants of aggressive behavior: pharmacother-apeutic targets at serotonin, dopamine and γ-aminobutyric acidsystems. Psychopharmacology 163:434–458
Mikics E, Kruk MR, Haller J (2004) Genomic and nongenomiceffects of glucocorticoids on aggressive behavior in male rats.Psychoneuroendocrinology 29:618–635
Mos J, Olivier B, van der Poel AM (1987) Modulary actions ofbenzodiazepine receptor ligands on agonistic behaviour. Phys-iol Behav 41:265–278
Orchinik M, Murray TF, Moore FL (1991) A corticosteroid receptorin neuronal membranes. Science 252:1848–1851
Orchinik M, Carroll SS, Li YH, McEwen BS, Weiland NG (2001)Heterogeneity of hippocampal GABA(A) receptors: regulationby corticosterone. J Neurosci 21:330–339
Piazza PV, Lemoal M (1997) Glucocorticoids as a biologicalsubstrate of reward: physiological and pathophysiological im-plications. Brain Res Rev 25:359–372
Piazza PV, Deroche V, Deminiere JM, Maccari S, Le Moal M,Simon H (1993) Corticosterone in the range of stress-inducedlevels possesses reinforcing properties: implications for sensa-tion-seeking behaviors. Proc Natl Acad Sci U S A 90:11738–11742
Potegal M (1984) The persistence of attack satiation in femalegolden hamsters. Aggress Behav 10:303–307
Potegal M, Tenbrink L (1984) Behavior of attack-primed and attack-satiated female golden hamsters (Mesocricetus auratus). JComp Psychol 98:66–75
Reddy DS (2003) Is there a physiological role for the neurosteroidTHDOC in stress-sensitive conditions? Trends Pharmacol Sci24:103–106
Reddy DS, Rogawski MA (2002) Stress-induced deoxycorticoster-one-derived neurosteroids modulate GABA(A) receptor func-tion and seizure susceptibility. J Neurosci 22:3795–3805
Rivier C (1993) Female rats release more corticosterone than male inresponse to alcohol: influence of circulating sex steroids andpossible consequences for blood alcohol levels. Alcohol ClinExp Res 17:854–859
Roberts WW, Keiss H (1964) Attack elicited by hypothalamicstimulation in cats raised in social isolation. J Comp PhysiolPsychol 58:187–193
Rodgers RJ, Waters AJ (1985) Benzodiazepines and their antag-onists: a pharmacoethological analysis with particular referenceto effects on “aggression”. Neurosci Biobehav Rev 9:21–35
Roitman MF, Stuber GD, Phillips PEM, Wightman RM, Carelli RM(2004) Dopamine operates as a subsecond modulator of foodseeking. J Neurosci 24:1265–1271
Rotllant D, Ons S, Carrasco J, Armario A (2002) Evidence thatmetyrapone can act as a stressor: effect on pituitary–adrenalhormones, plasma glucose and brain c-fos induction. Eur JNeurosci 16:693–700
Salamone JD, Cousins MS, McCullough LH, Carriero DL,Berkowitz RJ (1994) Nucleus accumbens dopamine releaseincreases during instrumental level pressing for food but notfree food consumption. Pharmacol Biochem Behav 49:25–31
Scott JP (1958) Aggression. University of Chicago Press, ChicagoScott JP (1966) Agonistic behavior of mice and rats: a review. Am
Zool 6:683–701Sofia RD, Salama AI (1970) Circadian rhythm for experimentally-
induced aggressive behavior in mice. Life Sci 9:331–338Stork O, Welzl H, Cremer H, Schachner M (1997) Increased
intermale aggression and neuroendocrine response in micedeficient for the neural cell adhesion molecule (NCAM). Eur JNeurosci 9:1117–1125
Tellegen A, Horn JM (1972) Primary aggressive motivation in threeinbred strains of mice. J Comp Physiol Psychol 78:297–304
Tellegen A, Horn JM, Legrand RG (1969) Opportunity foraggression as a reinforcer in mice. Psychon Sci 14:104–105
Thompson T (1964) Visual reinforcement in fighting cocks. J ExpAnal Behav 7:45–49
Thompson T (1969) Aggressive behaviour of Siamese fighting fish.In: Garattini S, Sigg EB (eds) Aggressive behaviour. ExcerptaMedica, Amsterdam, pp 15–31
Tomie A, Silberman Y, Williams K, Pohorecky LA (2002)Pavlovian autoshaping procedures increase plasma corticoste-rone levels in rats. Pharmacol Biochem Behav 72:507–513
Van Hemel PE, Myer JS (1970) Satiation of mouse killing by rats inan operant situation. Psychon Sci 21:129–130
126

Van Vugt DA, Piercy J, Farley AE, Reid RL, Rivest S (1997)Leutinizing hormone secretion and corticotropin-releasingfactor gene expression in the paraventricular nucleus of rhesusmonkeys following cortisol synthesis inhibition. Endocrinology138:2249–2258
Weerts EM, Miczek KA (1996) Primate vocalizations during socialseparation and aggression: effects of alcohol and benzodiaze-pines. Psychopharmacology 127:255–264
Weerts EM, Tornatzky W, Miczek KA (1993) “Anxiolytic” and“anxiogenic” benzodiazepines and beta-carbolines: effects onaggressive and social behavior in rats and squirrel monkeys.Psychopharmacology 110:451–459
Weisman AM, Berman ME, Taylor SP (1998) Effects of clorazepate,diazepam, and oxazepam on a laboratory measurement ofaggression in men. Int Clin Psychopharmacol 13:183–188
Winslow JT, Miczek KA (1984) Habituation of aggressive behaviorin mice: a parametric study. Aggress Behav 10:103–113
Wood GE, Young LT, Reagan LP, McEwen BS (2003) Acute andchronic restraint stress alter the incidence of social conflict inmale rats. Horm Behav 43:205–213
127
Copyright © 2022 FDOKUMEN




















