
High PKC a and Low E-Cadherin Expression Contribute to High Migratory Activity of Colon Carcinoma...
10
Molecular Biology of the Cell Vol. 12, 1973–1982, July 2001 High PKC a and Low E-Cadherin Expression Contribute to High Migratory Activity of Colon Carcinoma Cells Kai Masur,* Kerstin Lang, Bernd Niggemann, Kurt S. Zanker, and Frank Entschladen Institute of Immunology, Witten/Herdecke University, 58448 Witten, Germany Submitted December 27, 2000; Revised April 9, 2001; Accepted April 26, 2001 Monitoring Editor: Richard Hynes The protein kinase C (PKC) is a family of serine/threonine kinases that are key regulatory enzymes involved in growth, differentiation, cytoskeletal reorganization, tumor promotion, and migration. We investigated the functional involvement of PKC isotypes and of E-cadherin in the regulation of the locomotion of six human colon-adenocarcinoma cell lines. The different levels of the PKC a and the E-cadherin expression have predictable implications in the spontaneous locomotory activity. With the use of PKC a–specific inhibitors (safingol, Go6976) as well as the PKC d–specific inhibitor rottlerin, we showed that only PKC a plays a major role in the regulation of tumor cell migration. The results were verified by knocking out the translation of PKC isozymes with the use of an antisense oligonucleotide strategy. After stimulation with phorbol ester we observed a translocation and a colocalization of the activated PKC a at the plasma membrane to the surrounding extracellular matrix. Furthermore, we investigated the functional involvement of E-cadherin in the locomotion with the use of a blocking antibody. A high level of PKC a expression together with a low E-cadherin expression was strongly related to a high migratory activity of the colon carcinoma cells. This correlation was independent of the differentiation grade of the tumor cell lines. INTRODUCTION Cell migration is an essential step for embryonic develop- ment, wound healing, immune response, and tumor cell migration, that is, invasion and metastasis (Horwitz and Parsons, 1999). However, the transduction pathways that guide signals into the cell leading to migration are poorly understood. Different families of cell surface receptors are required to transduce external signals (e.g., from the ECM) for cell migration. Receptors of the families of integrins, cadherins, and selectins are mediating cell– cell interactions as well as cell–ECM contacts (Maaser et al., 1999). These adhesive interactions are important for migration because they also regulate intracellular signal transduction pathways (Clark and Brugge, 1995). Alterations in the expression of integrins and cadherins have been associated with changes in the migratory activity and phenotype of cells (Hynes, 1992; Filardo et al., 1996; Huttenlocher et al., 1998; Rigot et al., 1998). It is known that PKC mediates migration via integrins (Rigot et al., 1998; Kiley et al., 1999; Ng et al., 1999) and is involved in the signaling of serpentine receptors as well as growth factor receptors (Entschladen and Zanker, 2000). Treatment with the phorbol ester phobol-12-myristate-13- acetate (PMA) increased the rate of cell division and induced cell migration, but the naturally occurring activators of pro- tein kinase C (PKC) in vivo are the diacylglycerols (DAGs) and arachidonic acid, which are provided by phospho- lipases (Clemens et al., 1992). The PKC is a family of serine/threonine kinases consisting of at least 11 isoenzymes divided into 3 subfamilies (Hof- mann, 1997). These 3 groups have different characteristics that serve for classification: classic PKC isozymes: a, bI, bII, g; novel PKCs: d, e, u, h; and atypical PKCs: l, j, i. The isotypes are classified according to their requirements for calcium ions and phospholipids (e.g., phosphatidylserine and DAG) or phorbol esters (e.g., PMA) for activation. Only the classic PKC isotypes possess a binding site for calcium ions. This region is lacking in the novel and atypical PKCs, whereas the atypical PKCs are also lacking the binding site for DAG and phorbol esters. The PKC isozymes contain an amino-terminal regulatory domain and a carboxy-terminal catalytic domain, which are linked by a hinge region. It is * Corresponding author. E-mail address: [email protected]. Abbreviations used: AO, antisense oligonucleotides; CC, cal- phostin C; DAG, diacylglycerol; ECM, extracellular matrix; MARCKS, myristoylated, alanine-rich C kinase substrate; PI, propidium iodide; PKC, protein kinase C; PMA, phorbol-12- myristate-13-acetate. © 2001 by The American Society for Cell Biology 1973
-
Upload
inp-greifswald -
Category
Documents
-
view
3 -
download
0
Transcript of High PKC a and Low E-Cadherin Expression Contribute to High Migratory Activity of Colon Carcinoma...

Molecular Biology of the CellVol. 12, 1973–1982, July 2001
High PKC a and Low E-Cadherin ExpressionContribute to High Migratory Activity of ColonCarcinoma CellsKai Masur,* Kerstin Lang, Bernd Niggemann, Kurt S. Zanker, and FrankEntschladen
Institute of Immunology, Witten/Herdecke University, 58448 Witten, Germany
Submitted December 27, 2000; Revised April 9, 2001; Accepted April 26, 2001Monitoring Editor: Richard Hynes
The protein kinase C (PKC) is a family of serine/threonine kinases that are key regulatoryenzymes involved in growth, differentiation, cytoskeletal reorganization, tumor promotion, andmigration. We investigated the functional involvement of PKC isotypes and of E-cadherin in theregulation of the locomotion of six human colon-adenocarcinoma cell lines. The different levels ofthe PKC a and the E-cadherin expression have predictable implications in the spontaneouslocomotory activity. With the use of PKC a–specific inhibitors (safingol, Go6976) as well as thePKC d–specific inhibitor rottlerin, we showed that only PKC a plays a major role in the regulationof tumor cell migration. The results were verified by knocking out the translation of PKC isozymeswith the use of an antisense oligonucleotide strategy. After stimulation with phorbol ester weobserved a translocation and a colocalization of the activated PKC a at the plasma membrane tothe surrounding extracellular matrix. Furthermore, we investigated the functional involvement ofE-cadherin in the locomotion with the use of a blocking antibody. A high level of PKC aexpression together with a low E-cadherin expression was strongly related to a high migratoryactivity of the colon carcinoma cells. This correlation was independent of the differentiation gradeof the tumor cell lines.
INTRODUCTION
Cell migration is an essential step for embryonic develop-ment, wound healing, immune response, and tumor cellmigration, that is, invasion and metastasis (Horwitz andParsons, 1999). However, the transduction pathways thatguide signals into the cell leading to migration are poorlyunderstood. Different families of cell surface receptors arerequired to transduce external signals (e.g., from the ECM)for cell migration. Receptors of the families of integrins,cadherins, and selectins are mediating cell–cell interactionsas well as cell–ECM contacts (Maaser et al., 1999). Theseadhesive interactions are important for migration becausethey also regulate intracellular signal transduction pathways(Clark and Brugge, 1995). Alterations in the expression ofintegrins and cadherins have been associated with changesin the migratory activity and phenotype of cells (Hynes,1992; Filardo et al., 1996; Huttenlocher et al., 1998; Rigot et al.,
1998). It is known that PKC mediates migration via integrins(Rigot et al., 1998; Kiley et al., 1999; Ng et al., 1999) and isinvolved in the signaling of serpentine receptors as well asgrowth factor receptors (Entschladen and Zanker, 2000).Treatment with the phorbol ester phobol-12-myristate-13-acetate (PMA) increased the rate of cell division and inducedcell migration, but the naturally occurring activators of pro-tein kinase C (PKC) in vivo are the diacylglycerols (DAGs)and arachidonic acid, which are provided by phospho-lipases (Clemens et al., 1992).
The PKC is a family of serine/threonine kinases consistingof at least 11 isoenzymes divided into 3 subfamilies (Hof-mann, 1997). These 3 groups have different characteristicsthat serve for classification: classic PKC isozymes: a, bI, bII,g; novel PKCs: d, e, u, h; and atypical PKCs: l, j, i. Theisotypes are classified according to their requirements forcalcium ions and phospholipids (e.g., phosphatidylserineand DAG) or phorbol esters (e.g., PMA) for activation. Onlythe classic PKC isotypes possess a binding site for calciumions. This region is lacking in the novel and atypical PKCs,whereas the atypical PKCs are also lacking the binding sitefor DAG and phorbol esters. The PKC isozymes contain anamino-terminal regulatory domain and a carboxy-terminalcatalytic domain, which are linked by a hinge region. It is
* Corresponding author. E-mail address: [email protected] used: AO, antisense oligonucleotides; CC, cal-phostin C; DAG, diacylglycerol; ECM, extracellular matrix;MARCKS, myristoylated, alanine-rich C kinase substrate; PI,propidium iodide; PKC, protein kinase C; PMA, phorbol-12-myristate-13-acetate.
© 2001 by The American Society for Cell Biology 1973

important for the activation of the catalytic domain to openthe hinge region after removal of the pseudosubstrate regionfrom the catalytic site (Bruins and Epand, 1995). The regu-latory domain of classic PKC isozymes consists of two do-mains, C1 and C2. The C1 region is responsible for DAG andphorbol ester binding, whereas the C2 region mediates thebinding of calcium and negatively charged phosphatidylser-ine. The PKC requires acidic phospholipids for its activityand in the presence of activators the enzyme has the highestbinding affinity for membranes containing phosphatidylser-ine. The activation of PKC by phosphorylation of serine andthreonine residues is controlled by several modes, for exam-ple, autophosphorylation (Flint et al., 1990) as well as phos-phorylation by other PKC isotypes (transphosphorylation;Pears et al., 1992).
The presence of several isozymes in one cell and differen-tial activation or inhibition by different stimuli suggest thateach PKC isotype is involved in the regulation of differentfunctions and has a unique role in the cell. However, thebiological significance of the heterogeneity in the PKC fam-ily is not clear (Radominska-Pandya et al., 2000). The patternof PKC isotype expression is distinct between different tis-sues (Nishizuka, 1988) and differs even in the same tissue. InT lymphocytes the PKC isotype expression depends on theactivation state of the cells (Corrigan et al., 1995). In macro-phages the PKC d isotype is known to phosphorylate pleck-strin, which remains phosphorylated 60 min after phagocy-tosis (Brumell et al., 1999). The myristoylated, alanine-richC-kinase substrate (MARCKS) is also a conversant PKCsubstrate in fibroblasts and brain cells. The phosphorylationof MARCKS is found not to be specific for a special PKCisotype: conventional (c)PKC b1, novel (n)PKC d, and PKC eefficiently phosphorylated the MARCKS protein in vitro(Herget et al., 1995).
MATERIALS AND METHODS
Cells and Cell CultureAll cell lines used in this study were human colon carcinoma cellsobtained from the DSMZ (Deutsche Sammlung von Mikroorganis-men und Zellkulturen, Braunschweig, Germany) except SW 480 andSW 620, which were obtained from American Type Culture Collec-tion (ATCC, Rockville, MD). The differentiation grades of the tumorcell lines were taken from the data sheets of the supplying compa-nies. The poorly differentiated (grade III-IV) SW 480 cells and themetastatic SW 620 cell line were grown in antibiotic-free LeibovitzL-15 medium (PAA Laboratories GmbH, Linz, Austria), supple-mented with 10% FCS in a humidified atmosphere without CO2addition (Leibovitz et al., 1976). The moderately differentiated(grade II) Colo 320 cells were grown in RPMI 1640 medium (LifeTechnologies, Karlsruhe, Germany), supplemented with 10% FCSand 1% penicillin/streptomycin in a humidified 5% CO2 atmo-sphere (Quinn et al., 1979). The well-differentiated (grade I) HT 29cells were grown in McCoy’s 5A medium (PAA LaboratoriesGmbH), supplemented with 10% FCS and 1% penicillin/streptomy-cin in a humidified 5% CO2 atmosphere (von Kleist et al., 1975). Themoderately to poorly differentiated (grade II-III) SW 403 cells weregrown in Dulbecco’s MEM medium (PAA Laboratories GmbH) andthe poorly differentiated (grade III) SW 948 cells were grown inLeibovitz L-15 medium, both supplemented with 10% FCS and 1%penicillin/streptomycin in a humidified 5% CO2 atmosphere (Lei-bovitz et al., 1976).
Preparation of Three-dimensional ECM LatticesCultured cells were harvested with the use of a trypsin/EDTAsolution. An amount of 6 3 104 cells was mixed with 150 ml bufferedliquid collagen (pH 7.4, 1.63 mg/ml collagen type I; Collagen Cor-poration, Fremont, CA) containing minimal essential medium (Sig-ma, Deisenhofen, Germany) as well as the investigated substances:PMA, calphostin C (CC), rottlerin, safingol, and Go6976 (all pro-vided by Calbiochem, Bad Soden, Germany). This mixture wasloaded into self-constructed chambers as described previously(Friedl et al., 1995; Entschladen et al., 1997) with minor modifica-tions.
Three-dimensional Cell Migration AssayAfter polymerization of the collagen, the chambers were sealed, andcell locomotion within the three-dimensional collagen lattice wasrecorded by time-lapse videomicroscopy at 37°C for 12 h. For anal-ysis of the migratory activity, 30 cells of each sample were randomlyselected, and two-dimensional projections of paths were digitized asx/y-coordinates in 20-min intervals by computer-assisted cell track-ing. For the analysis of the displacement (i.e., the part of cells thatmoved within the observation period) 40 cells were randomly se-lected, and whether they developed migratory activity within thewhole observation period of 12 h was evaluated.
Flow CytometryThe viability of the cells was investigated by flow cytometry, sub-sequent to the migration experiments. Because the cells had beenincorporated into a collagen gel, a collagenase digestion (collage-nase type I; Collagen Corporation, Fremont, CA) for 20 min at 37°Cwas performed before flow cytometry. Propidium iodide (PI) wasused to distinguish nonviable from viable cells at a wavelength of488 nm. PI was added to a final concentration of 2 mg/ml. PI-negative (viable) cells were gated because of morphology and lackof fluorescence. The expression of E-cadherin also was measured byflow cytometry. The mouse mAb against human E-cadherin (clone67A4) was purchased from Coulter-Immunotech (Krefeld, Germa-ny). A secondary fluorescein isothiocyanate–conjugated goat anti-mouse antibody obtained from Jackson ImmunoResearch Labora-tories (West Grove, PA) was used for detection. The meanfluorescence intensity of specifically bound E-cadherin was mea-sured compared with the binding of an isotypic control mouseantibody (Coulter-Immunotech).
ImmunoblottingThe total amount of all classic and novel PKC isozymes (a, b, g, d,e, u, and h) was analyzed by immunoblotting as described previ-ously (Entschladen et al., 1997). Colon carcinoma cells (4 3 105 to 8 3105) were lysed in Laemmli sample buffer (Laemmli, 1970) andincubated for 10 min at 95°C. Proteins were separated with the useof PAGE according to the method of Laemmli and were transferredto an Immobilion-P membrane (Millipore, Bedford, MA) followedby blocking of the membranes with 5% dry milk powder (1 h, 20°C).After incubation of the membrane (2 h, 20°C) with the primarymonoclonal antibodies against the various PKC isotypes (purchasedfrom Transduction Laboratories, Lexington, KY), the membranewas washed vigorously with PBS-Tween. Subsequently, the mem-brane was first incubated with a peroxidase-linked anti-mouse an-tibody (1 mg/ml, 2 h, 20°C) and then with a chemiluminescencesubstrate (2 min, 20°C; Boehringer Mannheim). The chemilumines-cence signal was detected by exposure to a Kodak X-OMAT AR filmsheet (Sigma). Staining intensities were analyzed with a 300-dpi,8-bit flatbed scanner and quantified with the use of National Insti-tutes of Health Image software version 1.57 (Bethesda, MD).
K. Masur et al.
Molecular Biology of the Cell1974
Incubation with Antisense OligonucleotidesThe PKC (a, b, g, and d) isotype–specific phosphorothiolated anti-sense oligonucleotides (AO) and the control AO have been designedand manufactured by Biognostic GmbH (Goettingen, Germany).The efficiency of these AO to inhibit PKC isotype expression in SW480 colon carcinoma cells was successfully shown by Hochegger etal. (1999). After preparation of a 100 mM stock solution of each (AO),an amount of 3 3 105 cells was incubated in a 5 mM solution (24–36h, 37°C). The uptake of the oligonucleotides was checked by theaddition of fluorescein isothiocyanate-labeled control AO in testsamples with the use of flow cytometry and confocal laser scanmicroscopy for detection. To assess the effectiveness of the expres-sion of the blocking AO, an immunoblot was performed as de-scribed above.
Confocal Laser Scan MicroscopyFor immunofluorescence staining of the PKC a isoenzyme, 50 ml ofa suspension of 1 3 105 colon carcinoma cells in PBS or PBScontaining 50 ng/ml PMA was mixed with 100 ml buffered collagen,and the solution was transferred onto a coverslip. After 30 min ofpolymerization of the collagen matrix, cells were fixed with 3.7%paraformaldehyde (15 min, 20°C) and subsequently were perme-abilized with 0.5% Triton X-100 (10 min, 20°C). Thereafter, thesamples were incubated with 10 mg/ml (2 h, 20°C) of monoclonalmouse anti–PKC a antibody (purchased from Transduction Labo-ratories). After washing with PBS, the samples were incubated (2 h,20°C) with 10 mg/ml a Rhodamine Red–conjugated AffiniPure FabFragment of a goat anti-mouse antibody (Dianova, Hamburg, Ger-many). After an additional washing step, the coverslips were in-verted and mounted on slides. Confocal laser scanning microscopywith the use of a Leica TCS 4D microscope (Leica, Inc., Heidelberg,Germany) was performed as previously described (Friedl et al.,1997).
RESULTS
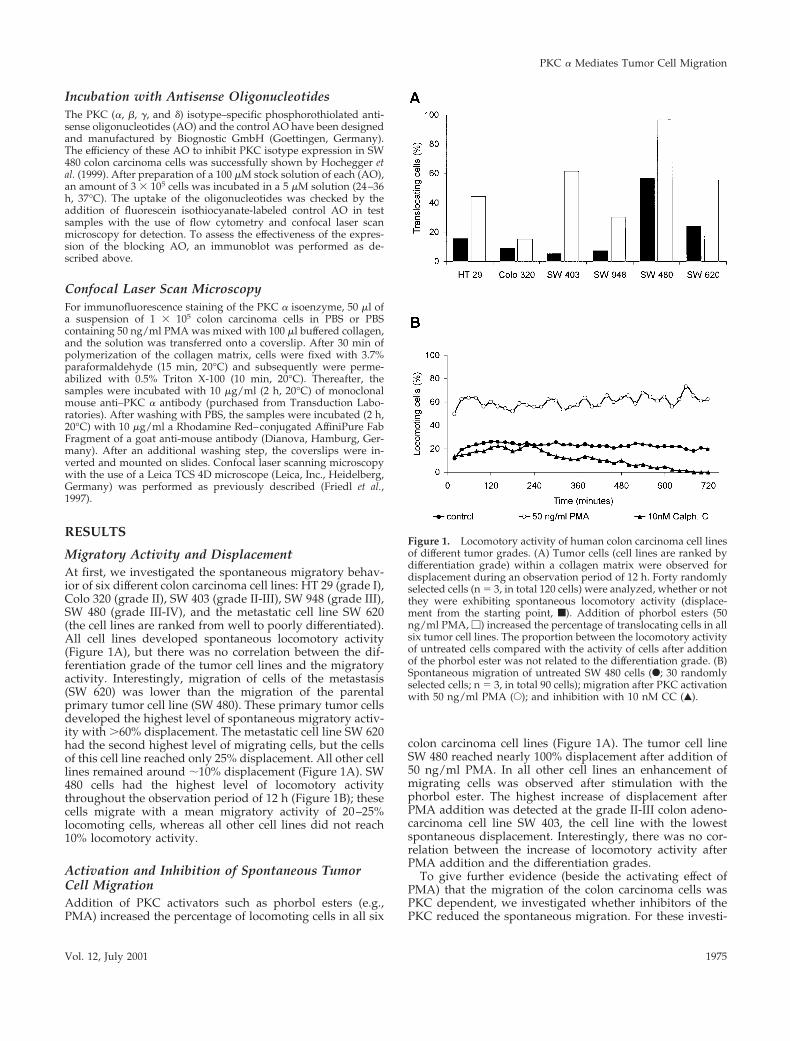
Migratory Activity and DisplacementAt first, we investigated the spontaneous migratory behav-ior of six different colon carcinoma cell lines: HT 29 (grade I),Colo 320 (grade II), SW 403 (grade II-III), SW 948 (grade III),SW 480 (grade III-IV), and the metastatic cell line SW 620(the cell lines are ranked from well to poorly differentiated).All cell lines developed spontaneous locomotory activity(Figure 1A), but there was no correlation between the dif-ferentiation grade of the tumor cell lines and the migratoryactivity. Interestingly, migration of cells of the metastasis(SW 620) was lower than the migration of the parentalprimary tumor cell line (SW 480). These primary tumor cellsdeveloped the highest level of spontaneous migratory activ-ity with .60% displacement. The metastatic cell line SW 620had the second highest level of migrating cells, but the cellsof this cell line reached only 25% displacement. All other celllines remained around ;10% displacement (Figure 1A). SW480 cells had the highest level of locomotory activitythroughout the observation period of 12 h (Figure 1B); thesecells migrate with a mean migratory activity of 20–25%locomoting cells, whereas all other cell lines did not reach10% locomotory activity.
Activation and Inhibition of Spontaneous TumorCell MigrationAddition of PKC activators such as phorbol esters (e.g.,PMA) increased the percentage of locomoting cells in all six
colon carcinoma cell lines (Figure 1A). The tumor cell lineSW 480 reached nearly 100% displacement after addition of50 ng/ml PMA. In all other cell lines an enhancement ofmigrating cells was observed after stimulation with thephorbol ester. The highest increase of displacement afterPMA addition was detected at the grade II-III colon adeno-carcinoma cell line SW 403, the cell line with the lowestspontaneous displacement. Interestingly, there was no cor-relation between the increase of locomotory activity afterPMA addition and the differentiation grades.
To give further evidence (beside the activating effect ofPMA) that the migration of the colon carcinoma cells wasPKC dependent, we investigated whether inhibitors of thePKC reduced the spontaneous migration. For these investi-
Figure 1. Locomotory activity of human colon carcinoma cell linesof different tumor grades. (A) Tumor cells (cell lines are ranked bydifferentiation grade) within a collagen matrix were observed fordisplacement during an observation period of 12 h. Forty randomlyselected cells (n 5 3, in total 120 cells) were analyzed, whether or notthey were exhibiting spontaneous locomotory activity (displace-ment from the starting point, f). Addition of phorbol esters (50ng/ml PMA, M) increased the percentage of translocating cells in allsix tumor cell lines. The proportion between the locomotory activityof untreated cells compared with the activity of cells after additionof the phorbol ester was not related to the differentiation grade. (B)Spontaneous migration of untreated SW 480 cells (F; 30 randomlyselected cells; n 5 3, in total 90 cells); migration after PKC activationwith 50 ng/ml PMA (E); and inhibition with 10 nM CC (Œ).
PKC a Mediates Tumor Cell Migration
Vol. 12, July 2001 1975
gations we first used the PKC-specific inhibitor CC, which isnot PKC isotype specific. CC inhibits all DAG-requiringPKC isozymes (Bruns et al., 1991). With the use of the SW 480cells, we found a total inhibition of migratory activity (Fig-ure 1B). Ten hours after addition of 10 nM CC the locomo-tion of the cells stopped completely. This reduction was notdue to cell death, as assessed by flow cytometry.
We concluded from these experiments concerning the ac-tivation and inhibition of the PKC that this enzyme plays acentral role in the regulation of migration. We found thatmigration was controlled either by classic or by novel PKCisotypes, because PMA enhanced the migratory activity.Therefore, we excluded an involvement of the atypical PKCisozymes l, j, and i.
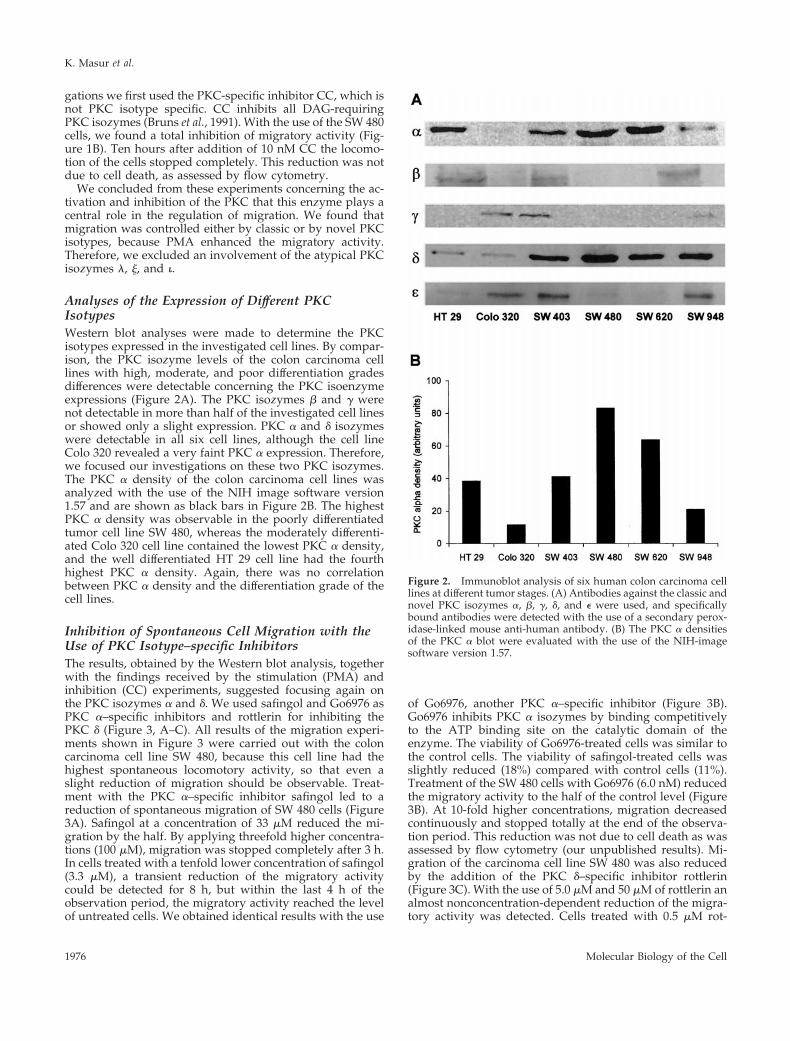
Analyses of the Expression of Different PKCIsotypesWestern blot analyses were made to determine the PKCisotypes expressed in the investigated cell lines. By compar-ison, the PKC isozyme levels of the colon carcinoma celllines with high, moderate, and poor differentiation gradesdifferences were detectable concerning the PKC isoenzymeexpressions (Figure 2A). The PKC isozymes b and g werenot detectable in more than half of the investigated cell linesor showed only a slight expression. PKC a and d isozymeswere detectable in all six cell lines, although the cell lineColo 320 revealed a very faint PKC a expression. Therefore,we focused our investigations on these two PKC isozymes.The PKC a density of the colon carcinoma cell lines wasanalyzed with the use of the NIH image software version1.57 and are shown as black bars in Figure 2B. The highestPKC a density was observable in the poorly differentiatedtumor cell line SW 480, whereas the moderately differenti-ated Colo 320 cell line contained the lowest PKC a density,and the well differentiated HT 29 cell line had the fourthhighest PKC a density. Again, there was no correlationbetween PKC a density and the differentiation grade of thecell lines.
Inhibition of Spontaneous Cell Migration with theUse of PKC Isotype–specific InhibitorsThe results, obtained by the Western blot analysis, togetherwith the findings received by the stimulation (PMA) andinhibition (CC) experiments, suggested focusing again onthe PKC isozymes a and d. We used safingol and Go6976 asPKC a–specific inhibitors and rottlerin for inhibiting thePKC d (Figure 3, A–C). All results of the migration experi-ments shown in Figure 3 were carried out with the coloncarcinoma cell line SW 480, because this cell line had thehighest spontaneous locomotory activity, so that even aslight reduction of migration should be observable. Treat-ment with the PKC a–specific inhibitor safingol led to areduction of spontaneous migration of SW 480 cells (Figure3A). Safingol at a concentration of 33 mM reduced the mi-gration by the half. By applying threefold higher concentra-tions (100 mM), migration was stopped completely after 3 h.In cells treated with a tenfold lower concentration of safingol(3.3 mM), a transient reduction of the migratory activitycould be detected for 8 h, but within the last 4 h of theobservation period, the migratory activity reached the levelof untreated cells. We obtained identical results with the use
of Go6976, another PKC a–specific inhibitor (Figure 3B).Go6976 inhibits PKC a isozymes by binding competitivelyto the ATP binding site on the catalytic domain of theenzyme. The viability of Go6976-treated cells was similar tothe control cells. The viability of safingol-treated cells wasslightly reduced (18%) compared with control cells (11%).Treatment of the SW 480 cells with Go6976 (6.0 nM) reducedthe migratory activity to the half of the control level (Figure3B). At 10-fold higher concentrations, migration decreasedcontinuously and stopped totally at the end of the observa-tion period. This reduction was not due to cell death as wasassessed by flow cytometry (our unpublished results). Mi-gration of the carcinoma cell line SW 480 was also reducedby the addition of the PKC d–specific inhibitor rottlerin(Figure 3C). With the use of 5.0 mM and 50 mM of rottlerin analmost nonconcentration-dependent reduction of the migra-tory activity was detected. Cells treated with 0.5 mM rot-
Figure 2. Immunoblot analysis of six human colon carcinoma celllines at different tumor stages. (A) Antibodies against the classic andnovel PKC isozymes a, b, g, d, and e were used, and specificallybound antibodies were detected with the use of a secondary perox-idase-linked mouse anti-human antibody. (B) The PKC a densitiesof the PKC a blot were evaluated with the use of the NIH-imagesoftware version 1.57.
K. Masur et al.
Molecular Biology of the Cell1976
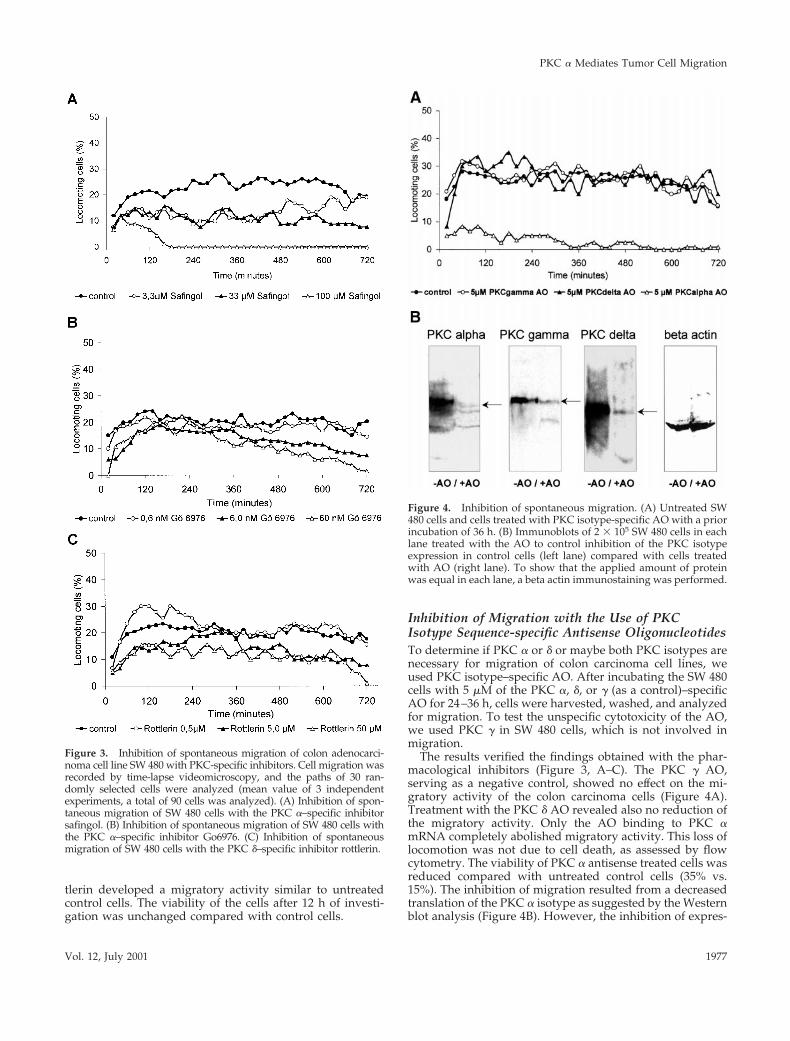
tlerin developed a migratory activity similar to untreatedcontrol cells. The viability of the cells after 12 h of investi-gation was unchanged compared with control cells.
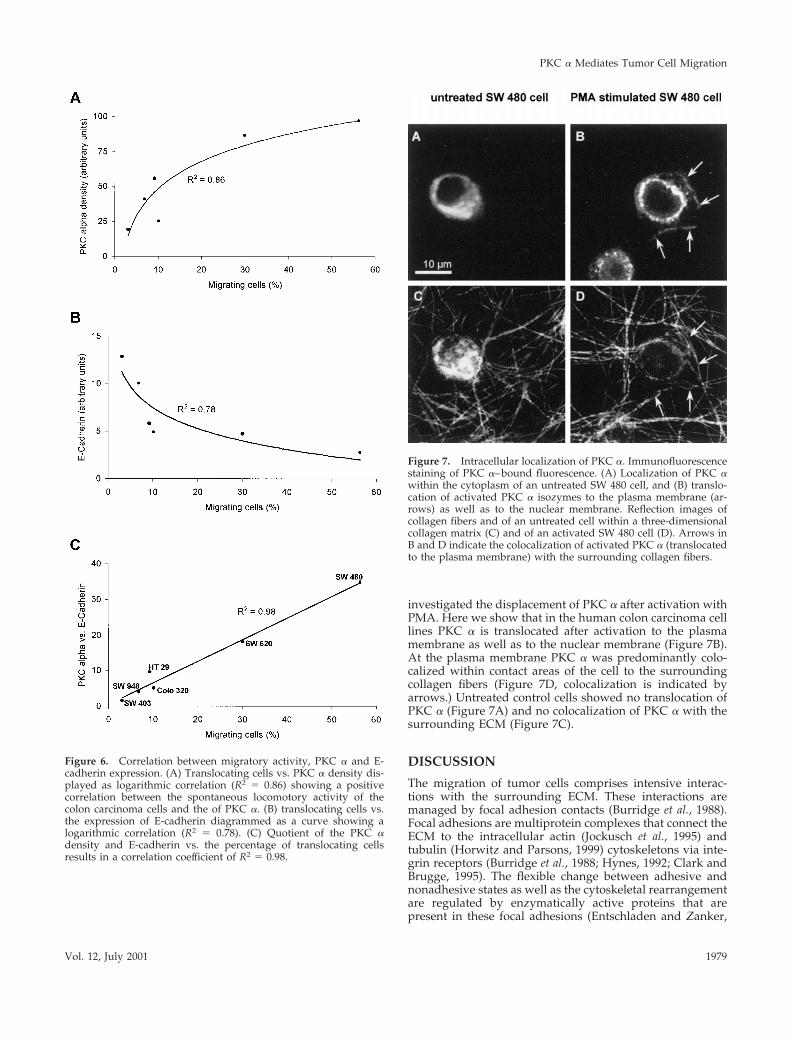
Inhibition of Migration with the Use of PKCIsotype Sequence-specific Antisense OligonucleotidesTo determine if PKC a or d or maybe both PKC isotypes arenecessary for migration of colon carcinoma cell lines, weused PKC isotype–specific AO. After incubating the SW 480cells with 5 mM of the PKC a, d, or g (as a control)–specificAO for 24–36 h, cells were harvested, washed, and analyzedfor migration. To test the unspecific cytotoxicity of the AO,we used PKC g in SW 480 cells, which is not involved inmigration.
The results verified the findings obtained with the phar-macological inhibitors (Figure 3, A–C). The PKC g AO,serving as a negative control, showed no effect on the mi-gratory activity of the colon carcinoma cells (Figure 4A).Treatment with the PKC d AO revealed also no reduction ofthe migratory activity. Only the AO binding to PKC amRNA completely abolished migratory activity. This loss oflocomotion was not due to cell death, as assessed by flowcytometry. The viability of PKC a antisense treated cells wasreduced compared with untreated control cells (35% vs.15%). The inhibition of migration resulted from a decreasedtranslation of the PKC a isotype as suggested by the Westernblot analysis (Figure 4B). However, the inhibition of expres-
Figure 3. Inhibition of spontaneous migration of colon adenocarci-noma cell line SW 480 with PKC-specific inhibitors. Cell migration wasrecorded by time-lapse videomicroscopy, and the paths of 30 ran-domly selected cells were analyzed (mean value of 3 independentexperiments, a total of 90 cells was analyzed). (A) Inhibition of spon-taneous migration of SW 480 cells with the PKC a–specific inhibitorsafingol. (B) Inhibition of spontaneous migration of SW 480 cells withthe PKC a–specific inhibitor Go6976. (C) Inhibition of spontaneousmigration of SW 480 cells with the PKC d–specific inhibitor rottlerin.
Figure 4. Inhibition of spontaneous migration. (A) Untreated SW480 cells and cells treated with PKC isotype-specific AO with a priorincubation of 36 h. (B) Immunoblots of 2 3 105 SW 480 cells in eachlane treated with the AO to control inhibition of the PKC isotypeexpression in control cells (left lane) compared with cells treatedwith AO (right lane). To show that the applied amount of proteinwas equal in each lane, a beta actin immunostaining was performed.
PKC a Mediates Tumor Cell Migration
Vol. 12, July 2001 1977

sion was not completely with neither AO. Therefore, wecould not exclude residual side-effects of PKC isotypes otherthan the PKC a.
Analysis of the E-Cadherin Expression and Influenceon Cell MigrationWe also investigated the expression levels of E-Cadherin inall six colon carcinoma cell lines (Figure 5A). The metastaticcell line SW 620 and the grade III cell line SW 480 containedno detectable amounts of E-cadherin, whereas the cell linesHT 29, Colo 320, SW 948, and SW 403 expressed differentlevels of this adhesion molecule. With a mean fluorescenceof 2.77, the cell line SW480 showed the lowest E-cadherinexpression compared with the mean fluorescence intensityof 10.03 for the SW 948 cell lines and 12.86 for the SW 403 cellline. Analyzing the expression of specifically bound antibod-ies of each cell line compared the unspecific binding of anisotypic control, only these two cell lines, SW 948 and SW403, revealed markedly higher expression of the E-cadherin–specific antibody (14.3% and 15.4%, E-cadherin-positivecells, respectively; Figure 5A). To give evidence for a func-tional dependence between displacement and E-cadherincontent, we investigated the migratory activity by addingmonoclonal blocking antibodies against E-cadherin. The SW403 and SW 948 cell lines expressed high levels of E-cadherinbut showed a spontaneous migratory activity of ,10% lo-comoting cells. With the use of a E-cadherin–blocking anti-body to prevent the development and rearrangement of cellclusters, we were able to increase the quantity of locomotoryactive cells to 36% in SW 948 cells and to 20% in SW 403 cells(Figure 5B). Screen shots of untreated SW 948 cells as well asof cells treated with the blocking antibody show that thisantibody prevents the development of cell clusters, andtherefore, increased single cell motility was observable.
Correlations between Migration Activity, PKC aDensity, and E-Cadherin ExpressionIn essence, the PKC a density of the six tumor cell linesshowed a positive correlation (R2 5 0.88) to the migratoryactivity (Figure 6A). Cell lines with a high PKC a densityshowed a high rate of translocating cells. In turn, the cellsurface expression of E-cadherin displayed a negative cor-relation (R2 5 0.78) to the quantity of migrating cells (Figure6B), meaning that cells with a high amount of E-cadherinshowed a limited displacement. Because not only the PKC adensity was decisive for migration but also the content ofE-cadherin, we calculated the quotient of the PKC a densityand the E-cadherin content of the six colon carcinoma celllines. This quotient correlates to the migratory activity of thecell lines and gives a nearly linear correlation (R2 5 0.98)(Figure 6C). In conclusion, the capability of the differenthuman tumor cells for migratory activity is dependent onthe expression of E-cadherin expressed on the cell surface aswell as on the content of PKC a.
Translocation of the Activated PKC a to theMembranesTranslocation of the activated PKC from the cytoplasm tothe membranes has been shown by various groups (Haller etal., 1998; Almholt et al., 1999; Gschwend et al., 2000). We
Figure 5. Determination of the E-cadherin expression of the colon car-cinoma cell lines analyzed by flow cytometry. (A) Graphs show theexpression of E-cadherin compared with the isotypic control (MFI, meanfluorescence intensity; M1, E-cadherin–positive cells compared with themarker region of the isotypic control adjusted to 99%). (B) Increase of thepercentage of translocating SW 403 and SW 948 cells treated with E-cadherin–blocking antibodies compared with untreated cells (f). Size ofscreen shots 400 3 600 mm2. Arrows, nonlocomotory cell clusters.
K. Masur et al.
Molecular Biology of the Cell1978
investigated the displacement of PKC a after activation withPMA. Here we show that in the human colon carcinoma celllines PKC a is translocated after activation to the plasmamembrane as well as to the nuclear membrane (Figure 7B).At the plasma membrane PKC a was predominantly colo-calized within contact areas of the cell to the surroundingcollagen fibers (Figure 7D, colocalization is indicated byarrows.) Untreated control cells showed no translocation ofPKC a (Figure 7A) and no colocalization of PKC a with thesurrounding ECM (Figure 7C).
DISCUSSION
The migration of tumor cells comprises intensive interac-tions with the surrounding ECM. These interactions aremanaged by focal adhesion contacts (Burridge et al., 1988).Focal adhesions are multiprotein complexes that connect theECM to the intracellular actin (Jockusch et al., 1995) andtubulin (Horwitz and Parsons, 1999) cytoskeletons via inte-grin receptors (Burridge et al., 1988; Hynes, 1992; Clark andBrugge, 1995). The flexible change between adhesive andnonadhesive states as well as the cytoskeletal rearrangementare regulated by enzymatically active proteins that arepresent in these focal adhesions (Entschladen and Zanker,
Figure 6. Correlation between migratory activity, PKC a and E-cadherin expression. (A) Translocating cells vs. PKC a density dis-played as logarithmic correlation (R2 5 0.86) showing a positivecorrelation between the spontaneous locomotory activity of thecolon carcinoma cells and the of PKC a. (B) translocating cells vs.the expression of E-cadherin diagrammed as a curve showing alogarithmic correlation (R2 5 0.78). (C) Quotient of the PKC adensity and E-cadherin vs. the percentage of translocating cellsresults in a correlation coefficient of R2 5 0.98.
Figure 7. Intracellular localization of PKC a. Immunofluorescencestaining of PKC a–bound fluorescence. (A) Localization of PKC awithin the cytoplasm of an untreated SW 480 cell, and (B) translo-cation of activated PKC a isozymes to the plasma membrane (ar-rows) as well as to the nuclear membrane. Reflection images ofcollagen fibers and of an untreated cell within a three-dimensionalcollagen matrix (C) and of an activated SW 480 cell (D). Arrows inB and D indicate the colocalization of activated PKC a (translocatedto the plasma membrane) with the surrounding collagen fibers.
PKC a Mediates Tumor Cell Migration
Vol. 12, July 2001 1979

2000). Among them are the PKC isotypes, which have re-peatedly and convincingly been shown to be associated withfocal adhesions: the PKC a is associated with focal adhesionsin rat embryo fibroblasts (Jaken et al., 1989; Liao et al., 1994).Adams et al. (Adams et al., 1999) established the matrix-initiated, PKC-dependent regulation of fascin phosphoryla-tion at serine 39 as a mechanism whereby matrix adhesion iscoupled to the organization of cytoskeletal structure.
The translocation of PKC a to the plasma membrane andthe direct colocalization with the surrounding collagen fi-bers (arrows in Figure 7, B, D, and F) is a further indicationof the involvement of PKC isozymes in the rearrangement ofthe cytoskeleton. Chapline et al. (Chapline et al., 1998) alsoshowed that PKCs directly interact with a group of substrateproteins, STICKs (substrates that interact with C-kinase).These STICKs are involved in cytoskeletal remodeling,shown by immunostaining of actin depending on the phos-phorylation state. Furthermore Ng et al. (1999) providedevidence for a key regulatory role of PKC isozymes for theb1 integrin traffic in migrating human breast carcinomacells. Kiley et al. (Kiley et al., 1999) pointed toward an in-volvement of the PKC d in tumor progression and cytoskel-etal remodeling. Barry and Chritchley (1994) found out thatPKC d plays a central role in the regulation of focal adhesioncontacts of Swiss 3T3 fibroblasts. TNF-a was shown to con-tribute to insulin resistance in rat adipocytes by altering PKCb translocation from the membrane to cytosol (Miura et al.,1999). Therefore, PKC isotypes have distinct functions in theregulation of different cellular signaling pathways.
We present here strong evidence for an involvement of thePKC a isotype in the regulation of colon carcinoma cellmigration with the use of pharmacological inhibitors andgenetic antisense oligonucleotide tools. The reduced migra-tory activity after treatment with the PKC d–specific inhibi-tor rottlerin is probably due to a cross-reaction inhibition ofthe PKC a, because the results derived with PKC d AOs didnot support a functional role of this isotype in cell migrationof colon carcinoma cell lines. At least, there was still aresidual expression of each PKC isotype after treatment withthe specific AO. Therefore, as mentioned before, we cannotcompletely exclude a minimal side effect of the PKC d on theregulation of migration; however, the results clearly showthe prominent role of the PKC a. Furthermore, the PKC aexpression is positively correlated to the migratory activityof tumor cells but not to the differentiation grades. Frey et al.(1997) have shown in an elegant way, that PKC a in non-transformed intestinal epithelial cells plays an importantrole by regulating the growth via modulation of Cip/Kipfamily cyclin-dependent kinase inhibitors and the retino-blastoma suppressor protein. Thus, the PKC a is a keyenzyme in transformed and untransformed cells of the in-testinal epithelium with respect to growth and migrationregulation. However, downstream in the signal transductionpathway regulating the migratory activity, other PKC iso-types might be involved that need an activation by PKCa–dependent pathways. Such a functional link has beenshown for the integrin phosphorylation by the PKC z inneutrophil granulocytes (Laudanna et al., 1998; Entschladenand Zanker, 2000).
With the use of phorbol esters as a positive control forinduction of migration, we found that the PKC-activatorPMA triggered the physical translocation of the PKC a
isozyme from the cytosol to the plasma membrane of coloncarcinoma cells as described by Almholt et al. (1999) in babyhamster kidney cells (Almholt et al., 1999). We could alsoshow a translocation of PKC a isozymes to the nuclearmembrane of SW 480 colon carcinoma cells, as described byHaller et al. in smooth muscle cells (Haller et al., 1998) and byWagner et al. in fibroblasts (Wagner et al., 2000). Jaken et al.(1989)support the viewpoint that the PKC a is involved inthe regulation of focal adhesion contacts.
Beside integrins, which are main constituents for the EC-M–cell interactions in focal adhesion, other cytoskeletal ad-hesion molecules are involved in adhesive processes relatedto tumor cell migration. E-cadherin is an important adhesionmolecule for cell–cell adhesions. The expression of an acti-vated PKC a isotype alters the functionality of E-cadherin(Batlle et al., 1998), which results in low cell aggregations.Gabbert et al. (1996) showed, for gastric cancer tissue spec-imens, that the tumor differentiation grade correlates withthe E-cadherin expression but not with the prognostic pa-rameters such as the depth of invasion, the lymph nodeinvolvement, and the vascular invasion.
Because Batlle et al. (1998) provided evidence for a regu-latory function of the PKC a in E-cadherin–mediated cell–cell interactions, we investigated the expression of E-cad-herin. Interestingly, the level of E-cadherin expression of thesix colon carcinoma cell lines was negatively correlated withthe migratory activity of the cells. The higher the PKC aexpression and the lower the E-cadherin expression was, thehigher was the migratory activity of the tumor cells, leadingto a strong linear correlation (R2 5 0.98).
Such a correlation between PKC a and E-cadherin expres-sion, and locomotory activity was not only found for cells ofdifferent colon carcinoma cells but also for three bladdercarcinoma cell lines (TCC-SUP, T 24, and HT 1376). The celllines TCC-SUP and T 24 show a high spontaneous migratoryactivity (85 and 65% locomoting cells, respectively), whereasthe HT 1376 cell line showed only minor locomotory activity(20% locomoting cells). Related to this, the two highly activecell lines exhibited a high expression level of PKC a, but noE-cadherin expression was detectable. In contrast the HT1376 cells had a tenfold lower expression of PKC a and ahigh amount of E-cadherin on the cell surface.
In summary, a high level of PKC a expression simulta-neously with a low E-cadherin level predicts an elevatedmigratory activity of colon carcinoma cells. However, thiscorrelation is independent of the differentiation grade of thetumor cells.
Our results suggest, that the PKC a and E-cadherin ex-pression of human intestinal cells underlies individual dif-ferences. As a consequence of these differences, the locomo-tory activity of human intestinal cells might differindividually. Maturating normal colon cells migrate fromthe crypta to the top of the villus. Cells with a high intrinsicPKC a and low E-cadherin expression would reach the topof the villus in a shorter period than cells with a low expres-sion of PKC a and a high E-cadherin expression. In case ofmutations and the development of a tumor, transformedcells with a high intrinsic PKC a and low E-cadherin expres-sion would be more motile, and the likelihood to builtmetastases at early stages of the tumor growth is greaterthan in cells with a low PKC a and a high E-cadherinexpression. However, the high motility of transformed cells
K. Masur et al.
Molecular Biology of the Cell1980

might be an advantage, because these cells from whichotherwise a solid tumor develops, quickly reach the top ofthe villus and are shed there by the peristaltic motion, exceptthat these tumor cells acquire concomitantly with cell mi-gration an invasive phenotype, entering the mucosa, sub-mucosa, and adjacent lymph nodes to form solid metastases.This body of work can be taken as a database for under-standing the processes that might underlie invasion andmetastasis. However, in addition to the herein describedinvolvements of the PKC a and E-cadherin, further signalingevents remain to be elucidated.
ACKNOWLEDGMENTS
We thank Britta Reubke-Gothe for excellent technical assistance.This work was supported by the Deutsche Krebshilfe, Bonn, Ger-many, and the Fritz-Bender-Foundation, Munich, Germany.
REFERENCES
Adams, J.C., Clelland, J.D., Collett, G.D., Matsumura, F., Yamashiro,S., and Zhang, L. (1999). Cell-matrix adhesions differentially regu-late fascin phosphorylation. Mol. Biol. Cell 10, 4177–4190.
Almholt, K., Arkhammar, P.O., Thastrup, O., and Tullin, S. (1999).Simultaneous visualization of the translocation of protein kinaseCalpha-green fluorescent protein hybrids and intracellular calciumconcentrations. Biochem. J. 337, 211–218.
Barry, S.T., and Critchley, D.R. (1994). The RhoA-dependent assem-bly of focal adhesions in Swiss 3T3 cells is associated with increasedtyrosine phosphorylation and the recruitment of both pp125FAKand protein kinase C-delta to focal adhesions. J. Cell Sci. 107, 2033–2045.
Batlle, E., Verdu, J., Dominguez, D., del Mont Llosas, M., Diaz, V.,Loukili, N., Paciucci, R., Alameda, F., and de Herreros, A.G. (1998).Protein kinase C-alpha activity inversely modulates invasion andgrowth of intestinal cells. J. Biol. Chem. 273, 15091–15098.
Bruins, R.H., and Epand, R.M. (1995). Substrate-induced transloca-tion of PKC-alpha to the membrane. Arch. Biochem. Biophys. 324,216–222.
Brumell, J.H., Howard, J.C., Craig, K., Grinstein, S., Schreiber, A.D.,and Tyers, M. (1999). Expression of the protein kinase C substratepleckstrin in macrophages: association with phagosomal mem-branes. J. Immunol. 163, 3388–3395.
Bruns, R.F., Miller, F.D., Merriman, R.L., Howbert, J.J., Heath, W.F.,Kobayashi, E., Takahashi, I., Tamaoki, T., and Nakano, H. (1991).Inhibition of protein kinase C by calphostin C is light-dependent.Biochem. Biophys. Res. Commun. 176, 288–293.
Burridge, K., Fath, K., Kelly, T., Nuckolls, G., and Turner, C. (1988).Focal adhesions: transmembrane junctions between the extracellularmatrix and the cytoskeleton. Annu. Rev. Cell Biol. 4, 487–525.
Chapline, C., Cottom, J., Tobin, H., Hulmes, J., Crabb, J., and Jaken,S. (1998). A major, transformation-sensitive PKC-binding protein isalso a PKC substrate involved in cytoskeletal remodeling. J. Biol.Chem. 273, 19482–19489.
Clark, E.A., and Brugge, J.S. (1995). Integrins and signal transduc-tion pathways: the road taken. Science 268, 233–239.
Clemens, M.J., Trayner, I., and Menaya, J. (1992). The role of proteinkinase C isoenzymes in the regulation of cell proliferation anddifferentiation. J. Cell Sci. 103, 881–887.
Corrigan, E., Kelleher, D., Feighery, C., and Long, A. (1995). Proteinkinase C isoform expression in CD45RA1 and CD45RO1 T lym-phocytes. Immunology 85, 299–303.
Entschladen, F., Niggemann, B., Zanker, K.S., and Friedl, P. (1997).Differential requirement of protein tyrosine kinases and proteinkinase C in the regulation of T cell locomotion in three-dimensionalcollagen matrices. J. Immunol. 159, 3203–3210.
Entschladen, F., and Zanker, K.S. (2000). Locomotion of tumor cells.a molecular comparison to migrating pre- and postmitotic leuko-cytes. J. Cancer Res. Clin. Oncol. 126, 671–681.
Filardo, E.J., Deming, S.L., and Cheresh, D.A. (1996). Regulation ofcell migration by the integrin beta subunit ectodomain. J. Cell Sci.,109, 1615–1622.
Flint, A.J., Paladini, R.D., and Koshland, D.E., Jr. (1990). Autophos-phorylation of protein kinase C at three separated regions of itsprimary sequence. Science 249, 408–411.
Frey, M.R., Saxon, M.L., Zhao, X., Rollins, A., Evans, S.S., and Black,J.D. (1997). Protein kinase C isozyme-mediated cell cycle arrestinvolves induction of p21(waf1/cip1) and p27(kip1) and hypophos-phorylation of the retinoblastoma protein in intestinal epithelialcells. J. Biol. Chem. 272, 9424–9435.
Friedl, P., Maaser, K., Klein, C.E., Niggemann, B., Krohne, G., andZanker, K.S. (1997). Migration of highly aggressive MV3 melanomacells in 3-dimensional collagen lattices results in local matrix reor-ganization and shedding of alpha2 and beta1 integrins and CD44.Cancer Res. 57, 2061–2070.
Friedl, P., Noble, P.B., Walton, P.A., Laird, D.W., Chauvin, P.J.,Tabah, R.J., Black, M., and Zanker, K.S. (1995). Migration of coordi-nated cell clusters in mesenchymal and epithelial cancer explants invitro. Cancer Res. 55, 4557–4560.
Gabbert, H.E., Mueller, W., Schneiders, A., Meier, S., Moll, R.,Birchmeier, W., and Hommel, G. (1996). Prognostic value of E-cadherin expression in 413 gastric carcinomas. Int. J. Cancer 69,184–189.
Gschwend, J.E., Fair, W.R., and Powell, C.T. (2000). Bryostatin 1induces prolonged activation of extracellular regulated protein ki-nases in, and apoptosis of LNCaP human prostate cancer cellsoverexpressing protein kinase calpha. Mol. Pharmacol. 57, 1224–1234.
Haller, H., Lindschau, C., Maasch, C., Olthoff, H., Kurscheid, D., andLuft, F.C. (1998). Integrin-induced protein kinase Calpha and Cep-silon translocation to focal adhesions mediates vascular smoothmuscle cell spreading. Circ. Res. 82, 157–165.
Herget, T., Oehrlein, S.A., Pappin, D.J., Rozengurt, E., and Parker,P.J. (1995). The myristoylated alanine-rich C-kinase substrate(MARCKS) is sequentially phosphorylated by conventional, noveland atypical isotypes of protein kinase C. Eur. J. Biochem. 233,448–457.
Hochegger, K., Partik, G., Schorkhuber, M., and Marian, B. (1999).Protein-kinase-C iso-enzymes support DNA synthesis and cell sur-vival in colorectal-tumor cells. Int. J. Cancer 83, 650–656.
Hofmann, J. (1997). The potential for isoenzyme-selective modula-tion of protein kinase C. FASEB J. 11, 649–669.
Horwitz, A.R., and Parsons, J.T. (1999). Cell migration—movin’on.Science 286, 1102–1103.
Huttenlocher, A., Lakonishok, M., Kinder, M., Wu, S., Truong, T.,Knudsen, K.A., and Horwitz, A.F. (1998). Integrin and cadherinsynergy regulates contact inhibition of migration and motile activ-ity. J. Cell Biol. 141, 515–526.
Hynes, R.O. (1992). Integrins: versatility, modulation, and signalingin cell adhesion. Cell 69, 11–25.
Jaken, S., Leach, K., and Klauck, T. (1989). Association of type 3protein kinase C with focal contacts in rat embryo fibroblasts. J. CellBiol., 109, 697–704.
PKC a Mediates Tumor Cell Migration
Vol. 12, July 2001 1981

Jockusch, B.M., Bubeck, P., Giehl, K., Kroemker, M., Moschner, J.,Rothkegel, M., Rudiger, M., Schluter, K., Stanke, G., and Winkler, J.(1995). The molecular architecture of focal adhesions. Annu. Rev.Cell Dev. Biol. 11, 379–416.
Kiley, S.C., Clark, K.J., Goodnough, M., Welch, D.R., and Jaken, S.(1999). Protein kinase C delta involvement in mammary tumor cellmetastasis. Cancer Res. 59, 3230–3238.
Laemmli, U.K. (1970). Cleavage of structural proteins during theassembly of the head of bacteriophage T4. Nature 227, 680–685.
Laudanna, C., Mochly-Rosen, D., Liron, T., Constantin, G., andButcher, E.C. (1998). Evidence of zeta protein kinase C involvementin polymorphonuclear neutrophil integrin-dependent adhesion andchemotaxis. J. Biol. Chem. 273, 30306–30315.
Leibovitz, A., Stinson, J.C., McCombs, W. B., 3rd, McCoy, C.E.,Mazur, K.C. and Mabry, N.D. (1976). Classification of human colo-rectal adenocarcinoma cell lines. Cancer Res. 36, 4562–4569.
Liao, L., Ramsay, K., and Jaken, S. (1994). Protein kinase C isozymesin progressively transformed rat embryo fibroblasts. Cell GrowthDiffer. 5, 1185–1194.
Maaser, K., Wolf, K., Klein, C.E., Niggemann, B., Zanker, K.S.,Brocker, E.B., and Friedl, P. (1999). Functional hierarchy of simul-taneously expressed adhesion receptors: integrin alpha2beta1 butnot CD44 mediates MV3 melanoma cell migration and matrix reor-ganization within three-dimensional hyaluronan-containing colla-gen matrices. Mol. Biol. Cell 10, 3067–3079.
Miura, A., Ishizuka, T., Kanoh, Y., Ishizawa, M., Itaya, S., Kimura,M., Kajita, K., and Yasuda, K. (1999). Effect of tumor necrosis factor-alpha on insulin signal transduction in rat adipocytes: relation to
PKCbeta and zeta translocation. Biochim. Biophys. Acta 1449, 227–238.
Ng, T., Shima, D., Squire, A., Bastiaens, P.I., Gschmeissner, S.,Humphries, M.J., and Parker, P.J. (1999). PKCalpha regulates beta1integrin-dependent cell motility through association and control ofintegrin traffic. EMBO J. 18, 3909–3923.
Nishizuka, Y. (1988). The molecular heterogeneity of protein kinaseC and its implications for cellular regulation. Nature 334, 661–665.
Pears, C., Stabel, S., Cazaubon, S., and Parker, P.J. (1992). Studies onthe phosphorylation of protein kinase C-alpha. Biochem. J. 283,515–518.
Quinn, L.A., Moore, G.E., Morgan, R.T., and Woods, L.K. (1979).Cell lines from human colon carcinoma with unusual cell products,double minutes, and homogeneously staining regions. Cancer Res.39, 4914–4924.
Radominska-Pandya, A., Chen, G., Czernik, P.J., Little, J.M.,Samokyszyn, V.M., Carter, C.A., and Nowak, G. (2000). Direct in-teraction of all trans-retinoic acid with protein kinase C. Implica-tions for PKC signaling and cancer therapy. J. Biol. Chem. 275,22324–22330.
Rigot, V., Lehmann, M., Andre, F., Daemi, N., Marvaldi, J., and Luis,J. (1998). Integrin ligation and PKC activation are required formigration of colon carcinoma cells. J. Cell Sci. 111, 3119–3127.
von Kleist, S., Chany, E., Burtin, P., King, M., and Fogh, J. (1975).Immunohistology of the antigenic pattern of a continuous cell linefrom a human colon tumor. J. Natl. Cancer Inst. 55, 555–560.
Wagner, S., Harteneck, C., Hucho, F., and Buchner, K. (2000). Anal-ysis of the subcellular distribution of protein kinase Calpha usingPKC-GFP fusion proteins. Exp. Cell Res. 258, 204–214.
K. Masur et al.
Molecular Biology of the Cell1982




















