
Coordinated action of NSF and PKC regulates GABAB receptor signaling efficacy
Upload
independentCategory
view
0download
0
Cell Biology International 1999, Vol. 23, No. 3, 195–201Article No. cbir.1999.0343, available online at http://www.idealibrary.com on
VASOPRESSIN STIMULATES TYROSINE PHOSPHORYLATION BY ACTIVATIONOF PKC IN THE RAT SMOOTH MUSCLE CELL LINE, A-10
CARLOS B. GONZALEZ*, CARLOS E. REYES, CARLOS D. FIGUEROA, VALERIA BARRA andSILVIA TRONCOSO
Department of Physiology and Department of Histology and Pathology, Faculty of Medicine,Universidad Austral of Chile, Valdivia, Chile
Received 16 September 1998; accepted 12 January 1999
Arginine vasopressin (AVP)-induced tyrosine phosphorylation was studied in a rat smoothmuscle cell line, A-10, by western blotting, using a monoclonal antibody against phospho-tyrosine. AVP stimulated the phosphorylation of several cellular proteins of molecular mass60–130 kDa in a time- and dose-dependent manner. Phosphorylation was mediated largely byV1 receptor subtype since it was inhibited by selective V1 antagonist and was only partiallyelicited by the V2 agonist, desmopressin. Heterotrimeric G-proteins seemed to be involved in thephosphorylation mechanism because fluoraluminates, an activator of heterotrimeric G-proteins(and thus an uncoupler of the receptor–G-protein interaction) inhibited the AVP-inducedphosphorylation. The protein kinase C (PKC) inhibitors: staurosporine, H7 and GF109203Xare able to block the AVP-stimulated phosphorylation. The last of these has been shown to beone of the most selective inhibitors of PKC. These results indicate that PKC is upstream ofthe phosphorylation, a motion which is supported by the fact that the AVP-stimulatedphosphorylation was downregulated by phorbol esters. � 1999 Academic Press
K: vasopressin; receptors; protein phosphorylation; protein kinase C; vascular smooth muscle;peptide hormones.A: AVP: arginine-vasopressin; PKC: protein kinase C; IP3: inositol triphosphate; DAG:diacylglycerol; H7: (1-(5-isoquinolinesulfonyl)-2-methylpiperazine, HCL); GF109203X:bisindolylmaleimide; Pdbu: phorbol-12,13-dibutyrate; PGF2�: prostaglandin F2�.
*To whom correspondence should be addressed at Department ofPhysiology, Universidad Austral de Chile, Valdivia, Chile.
INTRODUCTION
Phosphorylation of proteins, on tyrosine, serineor threonine residues, has been considered animportant transduction mechanism in a wide rangeof physiological effects of hormones, includingregulation of cell proliferation (Johnson andVaillaucourt, 1994). Many peptide hormonereceptors which posses a single transmembranedomain, such as growth factor receptors, haveintrinsic tyrosine kinase activity (Kazlauskas,1994). Other peptide receptors possessing a singletransmembrane domain but, in turn, lackingintrinsic tyrosine kinase activity, as with cytokinereceptors, are able to stimulate cytoplasmic tyro-sine kinase, many of which are encoded by proto-
1065–6995/99/030195+07 $30.00/0
oncogenes (Sato and Miyajima, 1994). Recently,many workers have shown that G-protein-coupledreceptors are able to stimulate tyrosine phos-phorylation in a variety of cellular proteins bymechanisms which are mostly unknown (Zackaryet al., 1991a,b, 1992; Watanabe et al., 1994;Leeb-Lunberg and Song, 1991; Force et al., 1991;Cazaubon et al., 1993; Quarles et al., 1993).
AVP is a peptide hormone which has a variety ofactions through its binding to at least two types ofreceptors, the V1 and the V2. The V2 type iscoupled to adenylate cyclase and increases intra-cellular cyclic AMP, whereas the V1 receptor isassociated to phospholipase C and, in turn,increases intracellular IP3 and DAG, and henceactivates PKC. Several peptide hormones, forexample bombesin, bradykinin, endothelins andAVP have been found to be potent mitogenic
� 1999 Academic Press

196 Cell Biology International, Vol. 23, No. 3, 1999
factors (Rozengurt and Sinnet-Smith, 1983; Wolland Rozengurt, 1988), but the mechanism by whichthis action is carried out remains unclear. Thisaction of AVP is especially important since thishormone is a potent vasoconstrictor and has beenimplicated in the pathogenesis of some types ofhypertension (Mohring et al., 1981; Cowley et al.,1981).
In the present work, we investigated tyrosinephosphorylation induced by AVP in a smoothmuscle cell line derived from rat aorta.
MATERIAL AND METHODS
Tissue culture
The smooth muscle cell line A-10 (ATCC CRL1476) was cultured to subconfluency on 35-mmdishes in Dulbecco modified Eagle medium con-taining 10% fetal calf serum. Cells were starved16 h before the experiments and then the appropri-ate amount of peptides was added prior to incu-bation for the indicated time. The treatmentwas stopped by addition of 100 �l of ice-coldRIPA buffer (150 m NaCl, 50 m Tris/HClpH 8.0, 1% deoxycholic acid, 1% Nonidet P40,0.1% SDS, 4 m EDTA, 1 m Na3VO4, 250 �g/mlp-nitrophenyl phosphate, 1 m phenylmethyl-sulphonyl fluoride, 1 �g/ml leupeptin, 1 �g/mlpepstatin and 1 �g/ml aprotinin). Cells werescraped, transferred to a microfuge tube and soni-cated. Proteins were precipitated with 10% TCAand resuspended in electrophoresis sample buffercontaining 1 m Na3VO4. In some experimentscells were incubated with 5 m NaF or 10 � AlCl3or the mixture of 5 m NaF and 10 � AlCl3 for30 min or with 50 n phorbol-12,13-dibutyrate(PdBu) for 15 min or 500 n PdBu for 18 h. Inother experiments, cells were incubated with thefollowing PKC inhibitors: 100 � staurosporine(Calbiochem), 20 � H-7 (Sigma) and 1 �GF109203X (Calbiochem).
Western blotting
Equal amounts of protein were loaded in each laneof a SDS polyacrylamide gel and separated byelectrophoresis at constant voltage for 3 h. Proteinswere then electrotransferred to nitrocellulose filters,using 0.05% SDS in the transfer buffer composedby 20 m Tris-glycine and 20% methanol. Filterswere incubated with the 4G10 anti-phospho-tyrosine biotin-labeled monoclonal antibody (UBI)at a concentration of 1 �g/�l, washed with TTBS
(20 m Tris-HCl pH 7.6, 500 m NaCl, 0.05%Tween 20) and then incubated with a strepto-avidin–biotinylated alkaline phosphatase-complex(BioRad). After washing, the alkaline phosphatasewas detected by incubation with 0.3 mg/ml ofnitroblue tetrazolium and 0.15 mg/ml of 5-bromo-4-chloro-3-indolyl phosphate in 0.1 Tris-HClpH 9.5, 5 m MgCl2.
Immunofluorescence
A-10 cells grown on coverslips in DMEM wereincubated with 100 n AVP for 0–30 min, afterwhich they were washed and fixed with 4% para-formaldehyde, permeabilized with cold (�20�C)methanol and incubated for 2 h with the anti-phosphotyrosine monoclonal antibody at a concen-tration of 5 �g/ml. Coverslips were washed andincubated with rabbit anti-mouse antibody dilutedat 1:500 and then with a fluorescein-labeled goatanti-rabbit IgG (1:30).
RESULTS AND DISCUSSION
Incubation of A-10 cells with 100 n AVP fordifferent times followed by western blot analysisusing an anti-phosphotyrosine monoclonal anti-body showed that AVP increased the phosphoryl-ation of proteins in the range of 60–130 kDa (Fig.1). Several proteins which are selectively phospho-rylated by AVP and other peptide hormones suchas angiotensin II have been identified, for examplea 68-kDa protein called paxillin, the p125 focaladhesion kinase and 42–44-kDa mitogen-activatedprotein kinases (MAPK) (Zackary et al., 1992;Cazaubon et al., 1993; Leduc and Meloche, 1995;Granot et al., 1993).
The tyrosine phosphorylation of these proteinsgradually increased up to 30 min and decreasedthereafter (Fig. 1a,b). As shown by immunoblot-ting, phosphorylation was clearly proportional tothe AVP concentration (Fig. 2). Concentrations aslow as 0.1 n were able to trigger the mechanism oftyrosine phosphorylation. It is noteworthy thatplasma concentrations of AVP vary from 10�17 to10�10 (Breyer and Ando, 1994). These resultsclearly show that AVP is able to stimulate thetyrosine phosphorylation of proteins in a time- anddose-dependent fashion.
The kinetics of phosphorylation proved similarto that elicited by bradykinin, bombesin andendothelins in fibroblast, that is there was anincrease in tyrosine phosphorylation, reaching themaximum between 15 and 30 min and decreasing

Cell Biology International, Vol. 23, No. 3, 1999 197
Fig. 1. Time-course kinetics of AVP-induced tyrosine phosphorylation of A-10 cells. Western immunoblot of cellular extractsfrom A-10 cells treated with 100 n AVP for the indicated times. Tyrosine phosphorylation was detected with a monoclonalantibody against phosphotyrosine (a). Densitometric analysis of the major phosphorylated bands (p130, p125, p68) was carriedout and the increment in the phosphorylation with respect to the control is presented. The biotinylated molecular weight markers(MWM) are: �-galactosidase (116,000), phosphorylase b (97,000), BSA (66,000), ovoalbumin (45,000), carbonic anhydrase B(31,000) and soybean trypsin inhibitor (21,500).
Fig. 2. Dose–response relationship of AVP-induced phosphorylation. Western blot analysis of A-10 cells extract treated withdifferent doses of AVP for 15 min (a). Densitometric analysis of the major bands was performed and the relative intensities arepresented as a percentage of the maximum (b).
thereafter. However, the kinetics of phosphoryl-ation evoked by thrombin, which is Ca2+ depen-dent, was different; the peak response was found tooccur within 1–2 min after treatment of muscle cellswith thrombin and it returned to basal levels withina further 5–10 min (Offermanns et al., 1993).
In non-stimulated cells, immunofluorescencemicroscopy using the anti-phosphotyrosine mono-clonal antibody showed the phosphorylatedproteins as thick spicules associated with theplasma membrane, apparently at the sites of celladhesion. The incubation of A-10 cells with AVP

198 Cell Biology International, Vol. 23, No. 3, 1999
Fig. 3. Immunofluorescence microscopy using a monoclonal phosphotyrosine antibody on A-10 cells without treatment (a) andtreated with 100 n AVP for 30 min (b). Note the overall increase of immunofluorescence intensity in the treated cells.
Fig. 4. Receptor specificity of AVP-stimulated tyrosine phosphorylation. Western blot of extracts from cells untreated or treatedwith 100 n AVP or 100 n AVP in the presence of 1 � Phaa,D-Tyr(Me)2,Arg6,Tyr9(NH2)AVP, a selective V1 antagonist (AT),or with 100 n desmopressin, a V2 agonist (dDAVP) (a). Densitometric analysis of the p68, p125 and p130 was carried out andexpressed as a fold increase of the control (b).
increased the overall immunofluorescence, whichbecame more diffuse than in non-stimulated cells(Fig. 3). The presence of immunoreactive phospho-tyrosine in relation to the focal adhesion sites mightbe associated with the ability of AVP to increasephosphorylation of the p125 focal adhesion kinase(Zackary et al., 1992).
In order to determine the receptor subtypeinvolved in phosphorylation the V1 selectiveantagonist Phaa, D-Tyr(Me)2, Arg6, Tyr9
(NH2)AVP was used (a kind gift of Professor M.Manning, Toledo, Ohio, U.S.A.). This analogue
almost completely blocked the increase of phos-phorylation of p130 and p125 induced by AVP. Onthe other hand, the V2 agonist dDAVP was unableto stimulate the phosphorylation of these proteins(Fig. 4). These results suggest that the tyrosinephosphorylation cascade is triggered by the V1receptor in smooth muscle cells.
To investigate whether G-protein is involved inthe mechanism of tyrosine phosphorylation weincubated cells with AlF�
4 a specific activator ofheterotrimeric G-proteins. AlF�
4 was generated byaddition of 5 m NaF and 10 � AlCl . The AlF�
3 4
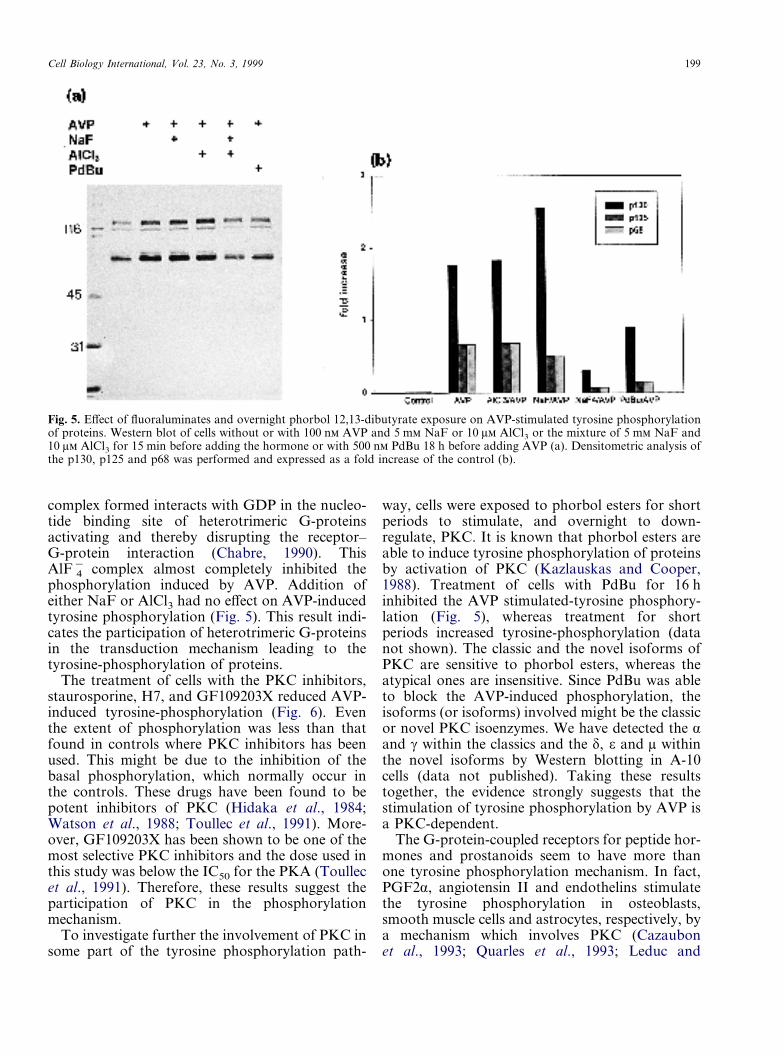
Cell Biology International, Vol. 23, No. 3, 1999 199
Fig. 5. Effect of fluoraluminates and overnight phorbol 12,13-dibutyrate exposure on AVP-stimulated tyrosine phosphorylationof proteins. Western blot of cells without or with 100 n AVP and 5 m NaF or 10 � AlCl3 or the mixture of 5 m NaF and10 � AlCl3 for 15 min before adding the hormone or with 500 n PdBu 18 h before adding AVP (a). Densitometric analysis ofthe p130, p125 and p68 was performed and expressed as a fold increase of the control (b).
complex formed interacts with GDP in the nucleo-tide binding site of heterotrimeric G-proteinsactivating and thereby disrupting the receptor–G-protein interaction (Chabre, 1990). ThisAlF�
4 complex almost completely inhibited thephosphorylation induced by AVP. Addition ofeither NaF or AlCl3 had no effect on AVP-inducedtyrosine phosphorylation (Fig. 5). This result indi-cates the participation of heterotrimeric G-proteinsin the transduction mechanism leading to thetyrosine-phosphorylation of proteins.
The treatment of cells with the PKC inhibitors,staurosporine, H7, and GF109203X reduced AVP-induced tyrosine-phosphorylation (Fig. 6). Eventhe extent of phosphorylation was less than thatfound in controls where PKC inhibitors has beenused. This might be due to the inhibition of thebasal phosphorylation, which normally occur inthe controls. These drugs have been found to bepotent inhibitors of PKC (Hidaka et al., 1984;Watson et al., 1988; Toullec et al., 1991). More-over, GF109203X has been shown to be one of themost selective PKC inhibitors and the dose used inthis study was below the IC50 for the PKA (Toullecet al., 1991). Therefore, these results suggest theparticipation of PKC in the phosphorylationmechanism.
To investigate further the involvement of PKC insome part of the tyrosine phosphorylation path-
way, cells were exposed to phorbol esters for shortperiods to stimulate, and overnight to down-regulate, PKC. It is known that phorbol esters areable to induce tyrosine phosphorylation of proteinsby activation of PKC (Kazlauskas and Cooper,1988). Treatment of cells with PdBu for 16 hinhibited the AVP stimulated-tyrosine phosphory-lation (Fig. 5), whereas treatment for shortperiods increased tyrosine-phosphorylation (datanot shown). The classic and the novel isoforms ofPKC are sensitive to phorbol esters, whereas theatypical ones are insensitive. Since PdBu was ableto block the AVP-induced phosphorylation, theisoforms (or isoforms) involved might be the classicor novel PKC isoenzymes. We have detected the �and � within the classics and the �, � and � withinthe novel isoforms by Western blotting in A-10cells (data not published). Taking these resultstogether, the evidence strongly suggests that thestimulation of tyrosine phosphorylation by AVP isa PKC-dependent.
The G-protein-coupled receptors for peptide hor-mones and prostanoids seem to have more thanone tyrosine phosphorylation mechanism. In fact,PGF2�, angiotensin II and endothelins stimulatethe tyrosine phosphorylation in osteoblasts,smooth muscle cells and astrocytes, respectively, bya mechanism which involves PKC (Cazaubonet al., 1993; Quarles et al., 1993; Leduc and

200 Cell Biology International, Vol. 23, No. 3, 1999
Meloche, 1995). In contrast, endothelins andangiotensin II enhance tyrosine phosphorylationby a PKC-dependent and -independent pathway inglomerular mesangial cells (Force et al., 1991).However, the stimulation of tyrosine phosphoryl-ation by bombesin, bradykinin, AVP and endo-thelins seems to be carried out by a mechanismwhich is independent of PKC and Ca2+ in Swiss 3T3fibroblast (Zackary et al., 1991b). PGF2� inducesthe phosphorylation in the same type of cellsthrough a Ca2+-dependent but PKC-independentpathway (Watanabe et al., 1994). Therefore, thereseem to be at least three pathways involved in tyro-sine phosphorylation of proteins by the seven trans-membrane domain receptors: one which involvesPKC, a second which uses Ca2+ and a third which isboth Ca2+ and PKC-independent. The predominantpathway for the V1 AVP receptor in smooth musclecells seems to be a pathway where PKC is upstreamof this tyrosine phosphorylation cascade.
ACKNOWLEDGEMENTS
We thank E.Oyarzun for technical assistance. Thiswork was supported by grants S 93-45 from theDireccion de Investigacion y Desarrollo, Universi-dad Austral de Chile, 1961171, 8980002 and2970018 from FONDECYT. The authors are
grateful to the Manuscript Presentation Service ofCBI for preparation of this article.
Fig. 6. Effect of PKC inhibitors on AVP-induced phosphorylation. Western blot of extracts from cells untreated (control) orincubated with AVP and with AVP and following pretreatment with staurosporine, H7 or GF109203x (a). Densitometric analysisof the p130, p125 and p68 shows the almost complete inhibition of the phosphorylation of these bands with the three PKCinhibitors (b).
REFERENCES
B MD, A Y, 1994. Hormonal signaling and regula-tion of salt and water transport in the collecting duct. AnnRev Physiol 56: 711–739.
C S, P PJ, S AD, C P-O,1993. Endothelins stimulate tyrosine phosphorylation andactivity of p42/mitogen-activated protein kinase in astro-cytes. Biochem J 293: 381–386.
C M, 1990. Aluminofluoride and beryllofluoride com-plexes: new phosphate analogs in enzymology. Trends Bio-chem Sci 15: 6–10.
C AW, J, C WC, Q EW, J, SMM, L HG, 1981. Vasopressin elevation in essen-tial hypertension and increased responsiveness to sodiumintake. Hypertension Dallas 3 (Suppl I): 93–100.
F T, K JM, A J, B IV, 1991.Endothelin, vasopressin and angiotensin II enhance tyrosinephosphorylation by protein kinase C-dependent and-independent pathways. J Biol Chem 266: 6650–6656.
G Y, E E, F H, V P V, WB, S RW, M J, 1993. Direct evidence for tyrosineand threonine phosphorylation and activation of mitogen-activated protein kinase by vasopressin in cultured ratvascular smooth muscle cells. J Biol Chem 268: 9564–9569.
H H, I M, K S, S Y, 1984.Isoquinolinesulfonamides, novel and potent inhibitors ofcyclic nucleotide dependent protein kinase and proteinkinase C. Biochemistry 23: 5036–5041.

Cell Biology International, Vol. 23, No. 3, 1999 201
J GL, V RR, 1994. Sequential proteinkinase reactions controlling cell growth and differentiation.Curr Opin Cell Biol 6: 230–238.
K A, 1994. Receptor tyrosine kinases and theirtargets. Curr Opin Genet Dev 4: 5–14.
K A, C JA, 1988. Protein kinase C mediatesplatelet-derived growth factor-induced tyrosine phosphory-lation of p42. J Cell Biol 106: 1395–1402.
L Y, M S, 1995. Angiotensin II stimulates tyrosinephosphorylation of the focal adhesion-associated proteinPaxillin in aortic smooth muscle cells. J Biol Chem 270:4401–4404.
L-L F, S X, 1991. Bradykinin and bombesinrapidly stimulate tyrosine phosphorylation of a 120-kDagroup of proteins in Swiss 3t3 cells. J Biol Chem 266:7746–7749.
M J, K J, S J, 1979. Studies on the role ofvasopressin in blood pressure control of spontaneouslyhypertensive rats with established hypertension (SHR,stroke-prone strain). J Cardiovasc Pharmacol 1: 593–608.
O S, B E, S G, 1993. ThrombinCa2+-dependently stimulates protein tyrosine phosphoryl-ation in BC3H1 muscle cells. Biochem J 290: 27–32.
Q LD, H DM, D G, M JP, 1993.Prostaglandin F2�-induced mitogenesis in MC3T3-E1 osteo-blasts; role of protein kinase-C-mediated tyrosinephosphorylation. Endocrinology 132: 1505–1513.
R E, S-S J, 1983. Bombesin stimulationof DNA synthesis and cell division in cultures of Swiss 3T3cells. Proc Natl Acad Sci USA 80: 2936–2940.
S N, M A, 1994. Multimeric cytokine receptors:common versus specific functions. Curr Opin Cell Biol 6:174–179.
T D, P P, C H, B P,G-P T, A M, B V, B P,
B E, L F, D L, C D,K J, 1991. The bisindolylmaleimide GF 109203Xis a potent and selective inhibitor of protein kinase C. J BiolChem 266: 15,771–15,781.
W T, N A, E D, H Y,T K, H Y, K M, K K,1994. Prostagladin F2� enhances tyrosine phosphorylationand DNA synthesis through phospholipase C-coupledreceptor via Ca2+-dependent intracellular pathway inNIH-3T3 cells. J Biol Chem 269: 17,619–17,625.
W S, MN J, S L, G P, 1988. Theaction of the protein kinase C inhibitor, staurosporine, onhuman platelets. Evidence against a regulatory role forprotein kinase C in the formation of inositol trisphosphateby thrombin. Biochem J 249: 345–350.
W PJ, R E, 1988. Two classes of antagonistinteract with receptors for the mitogenic neuropeptidesbombesin, bradykinin, and vasopressin. Growth Factors 1:75–83.
Z Y, G J, L W, S-S J,R E, 1991a. Bombesin, vasopressin and endo-thelin rapidly stimulate tyrosine phosphorylation in intactSwiss cells. J Biol Chem 88: 4577–4581.
Z Y, S-S J, R E, 1991b. Stimula-tion of tyrosine kinase activity in anti-phosphotyrosineimmune complexes of Swiss 3T3 cell lysates occurs rapidlyafter addition of bombesin, vasopressin, and endothelin tointact cells. J Biol Chem 266: 24,126–24,133.
Z Y, S-S J, R E, 1992.Bombesin, vasopressin and endothelin stimulation of tyro-sine phosphorylation in Swiss 3T3 cells. J Biol Chem 267:19,031–19,034.
Copyright © 2022 FDOKUMEN




















