
Herpes simplex virus antigens directly activate NK cells via TLR2, thus facilitating their...
13
The Journal of Immunology Herpes Simplex Virus Antigens Directly Activate NK Cells via TLR2, Thus Facilitating Their Presentation to CD4 T Lymphocytes Min Kim,* ,† Naomi R. Osborne,* ,† Weiguang Zeng, ‡ Heather Donaghy,* Kay McKinnon, x David C. Jackson, ‡ and Anthony L. Cunningham* ,† NK cells infiltrate human herpetic lesions, but their role has been underexplored. HSV can stimulate innate immune responses via surface TLR2, which is expressed on monocyte-derived dendritic cells (DCs) and NK cells. In this study, UV-inactivated HSV1/2 and immunodominant HSV2 glycoprotein D peptides conjugated to the TLR2 agonist dipalmitoyl-S-glyceryl cysteine stimulated CD4 T lymphocyte IFN-g responses within PBMCs or in coculture with monocyte-derived DCs. NK cells contributed markedly to the PBMC responses. Furthermore, NK cells alone were activated directly by both Ags, also upregulating HLA-DR and HLA-DQ and then they activated autologous CD4 T lymphocytes. Using Transwells, Ag-stimulated NK cells and CD4 T lymphocytes were shown to interact through both cell-to-cell contact and cytokines, differing in relative importance in different donors. A distinct immunological synapse between Ag-stimulated NK cells and CD4 T lymphocytes was observed, indicating the significance of their cell-to-cell contact. A large proportion (57%) of NK cells was also in contact with CD4 T lymphocytes in the dermal infiltrate of human recurrent herpetic lesions. Thus, NK cells stimulated by TLR2-activating HSV Ags can present Ag alone or augment the role of DCs in vitro and perhaps in herpetic lesions or draining lymph nodes. In addition to DCs, NK cells should be considered as targets for adjuvants during HSV vaccine development. The Journal of Immunology , 2012, 188: 4158–4170. I n cutaneous herpes simplex lesions of humans and in mu- rine models, keratinocytes, dendritic cells (DCs), and infil- trating lymphocytes, especially HSV-specific CD4 and CD8 T lymphocytes, were shown to play a central role in control- ling primary and recurrent HSV infections (1). In human recur- rent lesions, monocytes and CD4 T lymphocytes infiltrate first, followed by CD8 T lymphocytes, which appear to clear HSV in- fection (1–3). HSV infection of keratinocytes in vitro and in vivo induces the secretion of a sequence of chemokines and cytokines (first IFN-b and b chemokines and then IL-12, followed by IL-1b and IL-6) (4). The b chemokines probably attract monocytes and CD4 and CD8 T lymphocytes into lesions. IFN-a/b and IL-12 may entrain Th1 patterns of cytokine response from HSV Ag- stimulated CD4 (and CD8) T lymphocytes, especially IFN-g. IFN-a and IFN-g synergize to inhibit infection of keratinocytes after transmission from axon termini (5). HSV1/2 downregulate MHC class I expression by keratinocytes. This is reversed by IFN-g, mainly from early infiltrating CD4 T lymphocytes, thus allowing CD8 T lymphocytes to recognize infected keratinocytes (1, 4). IFN-g also stimulates MHC class II (MHC II) expression on keratinocytes, allowing recognition by CD4 T lymphocytes. Thus, Th1, rather than Th2, patterns of response are important for im- mune and vaccine control of HSV (6). Furthermore, resident memory CD8 T lymphocytes can be induced to proliferate directly in (murine) skin without the requirement to migrate to the lymph nodes. This requires DC interactions with CD4 and CD8 T lym- phocytes (7). Infrequent “sentinel” CD4 and CD8 T lympho- cytes are present in the dermis between lesions. Residual dermal DCs, plasmacytoid DCs (pDCs), and CD4 and CD8 T lympho- cytes persist in the dermis between lesions. CD8 T lymphocytes are most superficial, with some present at the dermo–epidermal junction (8). Mucocutaneous HSV infection is usually confined to the epi- dermal layer. Therefore, Langerhans cells (LCs) are likely to be involved in initial HSV Ag uptake (9), as shown by older murine studies (10). However, recent studies in mice demonstrated that migrating dermal CD103 + , langerin-expressing DCs are the major transporters of HSV Ag out of skin and, together with resident CD8 + DCs, are the major presenters of HSVAg to CD8 T lymphocytes in lymph nodes (11, 12), probably after Ag transfer from LCs (13). No human equivalent of the CD103 + dermal DC subset has been de- scribed, nor has a similar sequence of events, although it was shown that HSV-infected monocyte-derived DCs (MDDCs) undergo apo- ptosis and can be taken up by bystander uninfected MDDCs, which can then present HSV Ag to CD8 T lymphocytes (14). More recently, the important role of other innate immune ef- fectors, both humoral and cellular, such as macrophages, pDCs, NK cells, and gd T lymphocytes has been re-emphasized, either in direct immune control or via modulation of adaptive immune responses (15, 16). Recently, pDCs were demonstrated to infiltrate the upper dermis of recurrent herpes lesions where they show *Centre for Virus Research, Westmead Millennium Institute, Westmead, New South Wales 2145, Australia; † Sydney Medical School, University of Sydney, Sydney, New South Wales 2006, Australia; ‡ Department of Microbiology and Immunology, Uni- versity of Melbourne, Parkville, Victoria 3010, Australia; and x Parramatta Sexual Health Clinic, Parramatta, New South Wales 2150, Australia Received for publication December 9, 2011. Accepted for publication February 16, 2012. This work was supported by the National Health and Medical Research Council of Australia (Program Grant 358399 and Project Grant 632638). Address correspondence and reprint requests to Prof. Anthony L. Cunningham, West- mead Millennium Institute, Darcy Road, Westmead, NSW 2145, Australia. E-mail address: [email protected] Abbreviations used in this article: DC, dendritic cell; gD, HSV glycoprotein D; gD2, HSV2 glycoprotein D; gD2-24-4, HSV2 glycoprotein D peptide 24-4; gD2-30-5, HSV2 glycoprotein D peptide 30-5; hTLR2, human TLR2; IS, immunological syn- apse; LC, Langerhans cell; MDDC, monocyte-derived dendritic cell; MHC II, MHC class II; MOI, multiplicity of infection; Pam2Cys, dipalmitoyl-S-glyceryl cysteine; pDC, plasmacytoid dendritic cell; RH10, RPMI 1640 supplemented with 10% human AB serum; RT, room temperature; UV-HSV, UV-inactivated HSV. Copyright Ó 2012 by The American Association of Immunologists, Inc. 0022-1767/12/$16.00 www.jimmunol.org/cgi/doi/10.4049/jimmunol.1103450
Transcript of Herpes simplex virus antigens directly activate NK cells via TLR2, thus facilitating their...

The Journal of Immunology
Herpes Simplex Virus Antigens Directly Activate NK Cells viaTLR2, Thus Facilitating Their Presentation to CD4 TLymphocytes
Min Kim,*,† Naomi R. Osborne,*,† Weiguang Zeng,‡ Heather Donaghy,* Kay McKinnon,x
David C. Jackson,‡ and Anthony L. Cunningham*,†
NK cells infiltrate human herpetic lesions, but their role has been underexplored. HSV can stimulate innate immune responses via
surface TLR2, which is expressed on monocyte-derived dendritic cells (DCs) and NK cells. In this study, UV-inactivated HSV1/2
and immunodominant HSV2 glycoprotein D peptides conjugated to the TLR2 agonist dipalmitoyl-S-glyceryl cysteine stimulated
CD4 T lymphocyte IFN-g responses within PBMCs or in coculture with monocyte-derived DCs. NK cells contributed markedly to
the PBMC responses. Furthermore, NK cells alone were activated directly by both Ags, also upregulating HLA-DR and HLA-DQ
and then they activated autologous CD4 T lymphocytes. Using Transwells, Ag-stimulated NK cells and CD4 T lymphocytes were
shown to interact through both cell-to-cell contact and cytokines, differing in relative importance in different donors. A distinct
immunological synapse between Ag-stimulated NK cells and CD4 T lymphocytes was observed, indicating the significance of their
cell-to-cell contact. A large proportion (57%) of NK cells was also in contact with CD4 T lymphocytes in the dermal infiltrate of
human recurrent herpetic lesions. Thus, NK cells stimulated by TLR2-activating HSVAgs can present Ag alone or augment the
role of DCs in vitro and perhaps in herpetic lesions or draining lymph nodes. In addition to DCs, NK cells should be considered as
targets for adjuvants during HSV vaccine development. The Journal of Immunology, 2012, 188: 4158–4170.
In cutaneous herpes simplex lesions of humans and in mu-rine models, keratinocytes, dendritic cells (DCs), and infil-trating lymphocytes, especially HSV-specific CD4 and CD8
T lymphocytes, were shown to play a central role in control-ling primary and recurrent HSV infections (1). In human recur-rent lesions, monocytes and CD4 T lymphocytes infiltrate first,followed by CD8 T lymphocytes, which appear to clear HSV in-fection (1–3). HSV infection of keratinocytes in vitro and in vivoinduces the secretion of a sequence of chemokines and cytokines(first IFN-b and b chemokines and then IL-12, followed by IL-1band IL-6) (4). The b chemokines probably attract monocytes andCD4 and CD8 T lymphocytes into lesions. IFN-a/b and IL-12may entrain Th1 patterns of cytokine response from HSV Ag-stimulated CD4 (and CD8) T lymphocytes, especially IFN-g.IFN-a and IFN-g synergize to inhibit infection of keratinocytesafter transmission from axon termini (5). HSV1/2 downregulate
MHC class I expression by keratinocytes. This is reversed byIFN-g, mainly from early infiltrating CD4 T lymphocytes, thusallowing CD8 T lymphocytes to recognize infected keratinocytes(1, 4). IFN-g also stimulates MHC class II (MHC II) expression onkeratinocytes, allowing recognition by CD4 T lymphocytes. Thus,Th1, rather than Th2, patterns of response are important for im-mune and vaccine control of HSV (6). Furthermore, residentmemory CD8 T lymphocytes can be induced to proliferate directlyin (murine) skin without the requirement to migrate to the lymphnodes. This requires DC interactions with CD4 and CD8 T lym-phocytes (7). Infrequent “sentinel” CD4 and CD8 T lympho-cytes are present in the dermis between lesions. Residual dermalDCs, plasmacytoid DCs (pDCs), and CD4 and CD8 T lympho-cytes persist in the dermis between lesions. CD8 T lymphocytesare most superficial, with some present at the dermo–epidermaljunction (8).Mucocutaneous HSV infection is usually confined to the epi-
dermal layer. Therefore, Langerhans cells (LCs) are likely to beinvolved in initial HSV Ag uptake (9), as shown by older murinestudies (10). However, recent studies in mice demonstrated thatmigrating dermal CD103+, langerin-expressing DCs are the majortransporters of HSVAg out of skin and, together with resident CD8+
DCs, are the major presenters of HSVAg to CD8 T lymphocytes inlymph nodes (11, 12), probably after Ag transfer from LCs (13). Nohuman equivalent of the CD103+ dermal DC subset has been de-scribed, nor has a similar sequence of events, although it was shownthat HSV-infected monocyte-derived DCs (MDDCs) undergo apo-ptosis and can be taken up by bystander uninfected MDDCs, whichcan then present HSVAg to CD8 T lymphocytes (14).More recently, the important role of other innate immune ef-
fectors, both humoral and cellular, such as macrophages, pDCs,NK cells, and gd T lymphocytes has been re-emphasized, either indirect immune control or via modulation of adaptive immuneresponses (15, 16). Recently, pDCs were demonstrated to infiltratethe upper dermis of recurrent herpes lesions where they show
*Centre for Virus Research, Westmead Millennium Institute, Westmead, New SouthWales 2145, Australia; †Sydney Medical School, University of Sydney, Sydney, NewSouth Wales 2006, Australia; ‡Department of Microbiology and Immunology, Uni-versity of Melbourne, Parkville, Victoria 3010, Australia; and xParramatta SexualHealth Clinic, Parramatta, New South Wales 2150, Australia
Received for publication December 9, 2011. Accepted for publication February 16,2012.
This work was supported by the National Health and Medical Research Council ofAustralia (Program Grant 358399 and Project Grant 632638).
Address correspondence and reprint requests to Prof. Anthony L. Cunningham, West-mead Millennium Institute, Darcy Road, Westmead, NSW 2145, Australia. E-mailaddress: [email protected]
Abbreviations used in this article: DC, dendritic cell; gD, HSV glycoprotein D; gD2,HSV2 glycoprotein D; gD2-24-4, HSV2 glycoprotein D peptide 24-4; gD2-30-5,HSV2 glycoprotein D peptide 30-5; hTLR2, human TLR2; IS, immunological syn-apse; LC, Langerhans cell; MDDC, monocyte-derived dendritic cell; MHC II, MHCclass II; MOI, multiplicity of infection; Pam2Cys, dipalmitoyl-S-glyceryl cysteine;pDC, plasmacytoid dendritic cell; RH10, RPMI 1640 supplemented with 10% humanAB serum; RT, room temperature; UV-HSV, UV-inactivated HSV.
Copyright� 2012 by TheAmericanAssociation of Immunologists, Inc. 0022-1767/12/$16.00
www.jimmunol.org/cgi/doi/10.4049/jimmunol.1103450

evidence of IFN-a production and are in contact with activatedNK cells and lymphocytes (17). They produce IFN-a in responseto stimulation with HSV, to limit viral spread (18–21), and canpresent Ag to naive effector and regulatory lymphocytes (21–24).Like DCs, NK cells can bridge the innate and adaptive immune
responses. In humans, there are two main NK subsets: the first andwell-characterized CD56dimCD16+ NKs make up .95% of thosein blood and readily lyse malignant or virally infected cells but arelow cytokine secretors. In contrast, the more recently describedCD56brightCD162 NK cells produce large amounts of cytokines,such as TNF-a and IFN-g, upon stimulation, but they only acquirecytotoxicity after prolonged activation. DCs and NK cells oftencolocalize and are able to interact both at sites of inflammationand in lymph nodes (25, 26). In DC–NK cell interactions, cyto-kines, such as IL-12, IFN-a, and IL-15, are important, but directcell-to-cell contacts also seem to be essential in promoting full NKactivation by DCs (27). The CD56brightCD162 subset can also actas accessory APCs via upregulated HLA-DR and secretion ofIFN-g, thus enhancing CD4 T lymphocyte responses (28, 29).Interactions among DCs, NK cells, and CD4 T lymphocytes canenhance such responses (22, 30). IFN-g production by NK cellscan be induced by IL-12 secreted by TLR-stimulated DCs (31).Cross-talk between DCs and NK cells in viral infection was re-ported (32). However, it has not been fully explored how the trioof NK cells, DCs, and CD4 T lymphocytes interact with eachother, especially physically.TLRs on the surface and in endosomes of dendritic, NK, and other
innate immune cells are key sensors of viral pathogens leading toactivation of the innate and adaptive immune responses. Human LCsexpress TLRs 1, 2, 3, 5, 6, and 10 (33, 34); dermal DCs andMDDCsexpress TLRs 1, 2, 3, 4, 6, 8, and 10 (23); and NK cells expressTLRs 1, 2, 3, 5, 6, 7, 8, and 9 (31, 35). Some strains of HSV1 andHSV2, as well as human CMV, interact with TLR2 at the surfaceof many cell types via an unknown mechanism, as well as withTLR9 in the endosomes (particularly of pDCs) via unmethylatedCpGs in viral DNA. UV-inactivated HSV2 requires initial interac-tion with TLR2 prior to full activation via TLR9, whereas infecti-ous HSV2 can interact directly with TLR9 (18). IFN-a productionby pDCs via TLR9 (18–20) also activates NK cells, which, in turn,contribute to the Th1 adaptive immune response (36). TLRs 2and 9 also synergize in control of HSV in the mouse brain (37).HSV glycoprotein D (gD) is a highly immunogenic molecule
containing both B and T cell epitopes capable of inducing neu-tralizing Abs and CD4 T lymphocyte Th1 responses in the majorityof the human outbred population. Therefore, it has been used asan immunogen in human vaccine trials. However, gD alone doesnot appear to interact with surface TLRs on innate immune cells(38). In consort studies of couples discordant for genital herpes, avaccine consisting of HSV2 gD (gD2) and the TLR4 agonist d-monophosphoryl lipid A showed 73–74% efficacy in preventinggenital herpes in HSV-seronegative women whose partners hadgenital herpes (39), but apparently this was not confirmed in im-munizing a general population of HSV-seronegative women (40).The reasons for these differences are unclear and indicate the needfor increased understanding of the human immune response andcontrol of HSV infection, including innate immunity and its keyrole in directing the adaptive immune response.Recently, a high density of promiscuous immunodominant
peptide epitopes in gD2 for CD4 T lymphocytes was identified inour laboratory (41). The identified immunodominant gD2 peptide24-4 (gD2-24-4) and gD2 peptide 30-5 (gD2-30-5) promiscuouslybound to a broad range of HLA-DR molecules and were recog-nized by both HSV1- and HSV2-infected individuals, providingan explanation for the broad T cell recognition in humans. In
general, short peptides are weak immunogens, so they need to beadministered with adjuvants. To enhance the immunogenicity ofour gD2 peptides, they were conjugated to dipalmitoyl-S-glycerylcysteine (Pam2Cys) (42), which binds specifically to and stim-ulates via TLR2 and its heterodimeric partner TLR6 and inducesmaturation in DCs (43–46) (some HSV strains also target TLR2naturally).In preliminary experiments, we determined that these lipopep-
tides markedly enhanced HSV-specific CD4 T lymphocyte IFN-gresponses. Therefore, we examined the mechanism of these re-sponses and then compared them with those of endogenousTLR2 stimulation by whole HSV as UV-inactivated Ag. Initi-ally, the HSV2 186 strain, previously shown to stimulate se-quentially via TLR2 and 9, was used, followed by an HSV1 strainalso shown to stimulate via TLR2. Both types of Ags, UV-inactivated HSV (UV-HSV) and Pam2Cys–gD2 peptides, arealso known to stimulate maturation of human MDDCs (47, 48).Because of the prominent infiltration of NK cells into herpeticlesions, we also examined the role of NK cells in modulating theimmune response of CD4 T lymphocytes to these Ags. We foundthat NK cells contributed significantly to the unseparated PBMCIFN-g responses to HSV Ag and enhanced MDDC–CD4T lymphocyte responses to UV-HSV and Pam2Cys-conjugatedHSV peptides. Furthermore, NK cells alone were capable of di-rect presentation of these HSVAgs to CD4 T lymphocytes in theabsence of DCs. The mechanism was determined to be throughboth direct NK cell–CD4 T lymphocyte physical interaction andcytokines. These studies are relevant to such DC–NK cell–CD4T lymphocyte interactions in pairs and triplets in herpes lesions, aswell as to adjuvant choice and their mechanisms of action in HSVvaccine development.
Materials and MethodsSynthesis of gD2 peptides
gD2-24-4 and -30-5 were synthesized using Fmoc chemistry, as previouslydescribed (42). The amino acid sequences of the immunodominant CD4T lymphocyte epitopes in gD2 were QQGVTVDSIGML and PEDPED-SALLED for peptides gD2-24-4 and gD2-30-5, respectively. The twoPam2Cys-conjugated gD peptides (Pam2Cys–gD2-24-4 and Pam2Cys–gD2-30-5) were synthesized by covalently coupling of Pam2Cys lipidmoiety through two serine residues to each parental peptide (42). Forcarboxyfluorescein conjugation onto gD2-30-5 and Pam2Cys–gD2-30-5,the N-terminal of the Fmoc group was removed following the last acyla-tion reaction, and 5(6)-carboxyfluorescein (Fluka Biochmika) was thencoupled to the exposed amino group using a 4-fold molar excess in thepresence of equimolar amounts of O-benzotriazole-N,N,N9,N9-tetramethyl-uronium-hexafluorophosphate (Merck) and 1-hydroxybenzotriazole(Merck) and a 1.5-fold excess of diisopropylethylamine (Fluka Biochmika)for 4 h. All peptides were cleaved from the solid support, and side chainprotecting groups were simultaneously removed before being purifiedand characterized, as described previously (42). All peptide sampleswere dissolved at a concentration of 10 mM in DMSO and kept at280˚C.
PBMC preparation
Blood was collected from HSV1- and/or HSV2-seropositive healthy donorsand donor serotype-matched HSV1 or HSV2 Ags in individual experiments.Informed consent was obtained from all of the blood donors, and the studywas approved by the Ethics Committee for Sydney Western Area HealthService Research. PBMCs were prepared by Ficoll-Hypaque (GE Health-care Biosciences) density gradient.
Depletion of CD8+ and CD56+ lymphocytes from PBMCs
To examine the immunogenicity of Pam2Cys–gD2 peptides and whetherPam2Cys–gD2 peptides target only CD4 T lymphocytes as intended, CD8T lymphocytes and CD56+ NK cells were positively depleted using CD8and CD56 MicroBeads (Miltenyi Biotec), respectively. The same startingnumber of PBMCs was used as undepleted PBMC control for CD8 de-pletion as well as for CD56 depletion. After cell depletion, the resulting
The Journal of Immunology 4159

cells for each depletion were resuspended in an equal volume of RPMI1640 (Invitrogen) supplemented with 10% human AB serum (RH10;Sigma-Aldrich) to a final concentration of 1 3 106 cells/ml based on thenumber of undepleted PBMCs. Cells were cultured for 40 h in a 96-wellELISPOT plate with 5 mg/ml PHA, UV-HSV1 (strain F) or UV-HSV2(strain 186) at multiplicity of infection (MOI) 0.5, 10 mM peptide, orDMSO (Sigma-Aldrich) as negative control. UV-HSV was prepared aspreviously described (14). Viral infectivity was not observed after UVinactivation by plaque assay. To measure the immune response after theincubation, IFN-g ELISPOT was performed, as previously described (41).
TLR2-binding assay using HEK-Blue TLR2 cells
To investigate the mechanism of UV-HSV binding to NK cells, TLR2 wastested as one of the receptor candidates for UV-HSV. HEK-Blue humanTLR2 (hTLR2) cells and HEK-Blue Null cells (parental cell line of HEK-Blue hTLR2 cells) were purchased from InvivoGen. They were culturedand used for assays, according to the manufacturer’s instructions. Beforethe assay, TLR2 expression on both cells was examined by flow cytometryusing FITC-conjugated hTLR2 mAb. Pam2CSK4 (10 mM; InvivoGen),Pam2Cys–gD2-30-5 (10 mM), UV-HSV1 (MOI 0.5), UV-HSV2 (MOI0.5), and water and TLR7/8 agonist Resiquimod (5 mg/ml; InvivoGen) asnegative control were added to the cells and cultured for 20 h at 37˚C, 5%CO2. Supernatants were collected for measuring NF-kB–induced secretedembryonic alkaline phosphatase resulting from TLR2 binding by testedstimuli and incubated with QUANTI-BLUE medium (InvivoGen) for 1 h at37˚C. After the incubation, OD was measured at 650 nm (Victor2TM 1420Multilabel Counter; PerkinElmer).
Generation of MDDCs
Monocytes were isolated from PBMCs using CD14 MicroBeads (MiltenyiBiotec), according to the manufacturer’s instructions. Isolated CD14+
monocytes were incubated at a cell concentration of 5 3 105 cells/ml for5–6 d in RH10 supplemented with IL-4 (250 U/ml; Biosource) and GM-CSF (200 U/ml; Biosource) at 37˚C, 5% CO2. After the incubation, cellmorphology was examined under the microscope, and flow cytometry wasperformed to check the phenotypes of immature MDDCs. To culture withNK cells and/or CD4 T lymphocytes in RH10 media in ELISPOT plate, theratio of MDDCs, NK cells, and CD4 T lymphocytes was kept at 1:1:10.
Isolation and culture of NK cells and CD4 T lymphocytes
NK cells and CD4 T lymphocytes were negatively isolated using EasySephuman NK Enrichment kit and EasySep human CD4 T Enrichment kit(StemCell Technologies), according to the manufacturer’s instructions.RH10 was used to culture cells at a concentration of 106 cells/ml. ForIFN-g ELISPOT, cells were cultured for 40 h in a 96-well ELISPOT plate(Millipore), with or without peptide. For the rest of the experiments, NKcells and CD4 T lymphocytes were cocultured overnight with 10 mMpeptides or with DMSO as negative control. The ratio of NK cells/CD4T lymphocytes for coculture was 1:4, unless otherwise indicated.
Coculture of HSV Ag-pulsed NK cells with CD4 T lymphocytesin Transwells
To investigate whether NK cells activate CD4 T lymphocytes by cell-to-cellcontact after being stimulated with peptides, 0.4-mM pore-sized Transwells(Corning) were used for cell culture to block cell migration. Purified NKcells were placed with purified CD4 T lymphocytes in the lower chamberor placed in the upper chamber with CD4 T lymphocytes in the lowerchamber. CD4 T lymphocytes were cultured in the lower chamber, with orwithout NK cells. Peptide (10 mM) and DMSO of equivalent volume asnegative control were added to the well in which NK cells were placed,because NK cells were suspected APCs. Maximum cell number in theupper chamber was 13 105 in 100 ml RH10, and that in the lower chamberwas 6 3 105 in 600 ml RH10. Cells were incubated overnight at 37˚C, 5%CO2. The following day, cells were collected separately from the bottom orupper chamber for flow cytometry.
Flow cytometry
To check the phenotype of immature DCs, allophycocyanin–anti-CD14,PE–anti-CD86, and PE–Cy7–anti-HLA-DR Abs were purchased from BD.For cell depletion, allophycocyanin–anti-CD8 (BD), FITC–anti-CD3 (BDPharmingen), and PE–anti-CD56 (Miltenyi Biotec) Abs were used to as-sess the efficiency of CD8 and CD56 cell depletion. To measure the ac-tivation of NK cells and CD4 T lymphocytes, the early activation markerCD69 was labeled with allophycocyanin–anti-CD69 Ab (BD Pharmingen).CD4 T lymphocytes labeled with PE–Cy7–anti-CD4 Ab (BD) were ob-
tained by gating on live lymphocytes and then CD4 and CD3 double-positive cells. NK cells were gated on live lymphocytes and then CD56+
CD32 cells. The purity of NK and CD4 T lymphocytes was .98%.PerCP–anti-HLA-DR (BD) or PE–Cy7–anti-HLA-DR (BioLegend) andFITC–anti-HLA-DQ (BioLegend) Abs were used to assess MHC II ex-pression on stimulated NK cells. PE–Cy7–anti-TLR2 (eBioscience) wasused to determine whether TLR2 is expressed on MHC II-expressing NKcells. Because of the low percentage of CD56brightCD162 NK cells andpeptide-specific CD4 memory T cells, $200,000 cells were acquired persample on a BD FACSCanto. Data were analyzed by FlowJo software.
Preparation of peptide-specific CD4 T cell line
To increase the frequency of immunological synapse (IS) formation be-tween Pam2Cys–gD2-30-5–stimulated NK cells and CD4 T lymphocytes,a gD2-30-5–specific CD4 T cell line was generated. PBMCs of HSV1–infected individuals were pulsed with 10 mM gD2-30-5 in 100 ml serum-free RPMI 1640 for 1 h at 37˚C, 5% CO2 and cocultured with autologousPBMCs at a 1:2 ratio at a cell concentration of 13 106 cells in RPMI 1640supplemented with 10% FBS, penicillin, and streptomycin (50 U/ml and50 mg/ml, respectively; Gibco-BRL). After 1 wk, cells were restimulatedwith irradiated (3000 rad) peptide-sensitized autologous PBMCs, as above.Restimulated cells were cultured in RPMI 1640 supplemented with 10%FBS, IL-2 (10 U/ml; Roche), and penicillin/streptomycin at 1 3 106 cells/ml. Restimulation was repeated once a week until enough cells were ob-tained. Medium was replenished twice a week.
Detection and quantification of NK cell–CD4 T lymphocyteconjugates
To confirm that NK cells can directly contact CD4 T lymphocytes, purifiedand stimulated NK cells were cocultured with purified CD4 T lymphocytesor gD2-30-5–specific CD4 T cell line as follows. Purified NK cells werepulsed with 10 mM Pam2Cys–gD2-30-5 or UV-HSV1 at MOI 0.5, orwithout Ag as negative control, in 100 ml serum-free RPMI 1640 for 1 h at37˚C, 5% CO2. After the incubation, cells received a 103 volume ofserum-free RPMI 1640 and were centrifuged for 5 min at 300 3 g. Afterdecanting the supernatant, NK cell pellet was resuspended and added toCD4 T lymphocytes at a 2:1 ratio. The combined NK cells and CD4T lymphocytes were briefly centrifuged at 10,000 3 g and incubated for 5min at 37˚C. After the conjugation, cells were gently resuspended; serum-free RPMI 1640 was added to the cells, and they were loaded onto theprepared Cell Tak (BD)-coated cover slips through “gummy squares”(eight-well flexiPERM removable and reusable TC chambers; ISC Bio-Express). Cells were centrifuged for 3 min at 300 3 g. The gummy squarewas removed before cells were fixed and labeled.
Immunofluorescence microscopy
Cells loaded on cover slips were fixed with 3% paraformaldehyde (Pro-SciTech) for 20 min at room temperature (RT) and then neutralized with 50mM NH4Cl three times for 3 min each. Cells were permeabilized with0.05% Triton X-100 (Sigma-Aldrich) for 1 min at RT and washed oncewith PBS. After permeabilization, cells were left for 15 min in PBS sup-plemented with 1% fish skin gelatin (Sigma-Aldrich) and 0.02% saponin(Sigma-Aldrich) and then for 30 min in Image-iT FX signal enhancer(Invitrogen). For visualizing ISs, the actin-binding molecule talin was la-beled with mouse anti-human talin mAb (Abcam) as primary Ab for 30min at RT and then with Alexa Fluor 633-conjugated goat anti-mouse IgG(Molecular Probes) for 30 min as secondary Ab. For the secondary stainingto differentiate NK cells and CD4 T lymphocytes, mouse anti-human CD4Ab (BD Pharmingen) was used to label CD4 T lymphocytes for 30 min,followed by Alexa Fluor 546-conjugated goat anti-mouse IgG (MolecularProbes) for 30 min. Mouse anti-human IgG (Sigma-Aldrich) was used asprimary Ab for negative control staining for primary and secondarystaining. All washes between steps were carried out with 10% goat serum(Sigma-Aldrich) in PBS. After staining, ProLong Gold Antifade reagent(Invitrogen) was added to the stained cells, and glass slides were mountedonto cell-loaded coverslips. Slides were visualized through an 3100 1.35NA oil-immersion lens with an inverted Olympus IX-70 microscope(DeltaVision Image Restoration Microscope; Applied Precision/Olympus)and a Photometrics CoolSnap QE camera.
Immunostaining of histologic sections of herpetic lesionbiopsies
Punch biopsies were taken from recurrent genital herpes lesions #4 d afteronset. Samples were obtained under a protocol approved by the ethicscommittee for Sydney West Area Health Service. Written informed con-
4160 HSV Ag-STIMULATED NK–CD4 T CELL INTERACTIONS
sent was obtained prior to sample donation. The biopsies were snap-frozenin OCT, cut into 5-mm sections, and placed on slides. Slides were kept at280˚C until required for immunofluorescent staining. Slides were fixedwith ice-cold methanol/acetone (1:1) prior to staining. Abs used for mi-croscopy were mouse mAb for CD56 (1:200; Invitrogen) and goat poly-clonal Ab for CD4 (1:100; R&D Systems). Slides were incubated at 37˚Cwith primary Abs for 45 min. Following washing, the secondary Abs AlexaFluor 546 donkey anti-goat (1:400; Molecular Probes) and Alexa Fluor488 donkey anti-mouse (1:400; Molecular Probes) were added for 30 minat 37˚C. Excess secondary Ab was washed away, cells were mounted usingProLong Gold Antifade supplemented with DAPI (Invitrogen), coverslipswere applied, and slides were refrigerated until imaged with an OlympusIX-70 microscope.
Statistical analysis
Outcome variables were log transformed to stabilize the variance prior toanalysis. Repeated measures ANOVAwith multiple pair-wise comparisonswas used to assess the effects of treatment within a group. SPSS version 20was used to analyze data. A 5% significance level with two-sided tests wasused throughout.
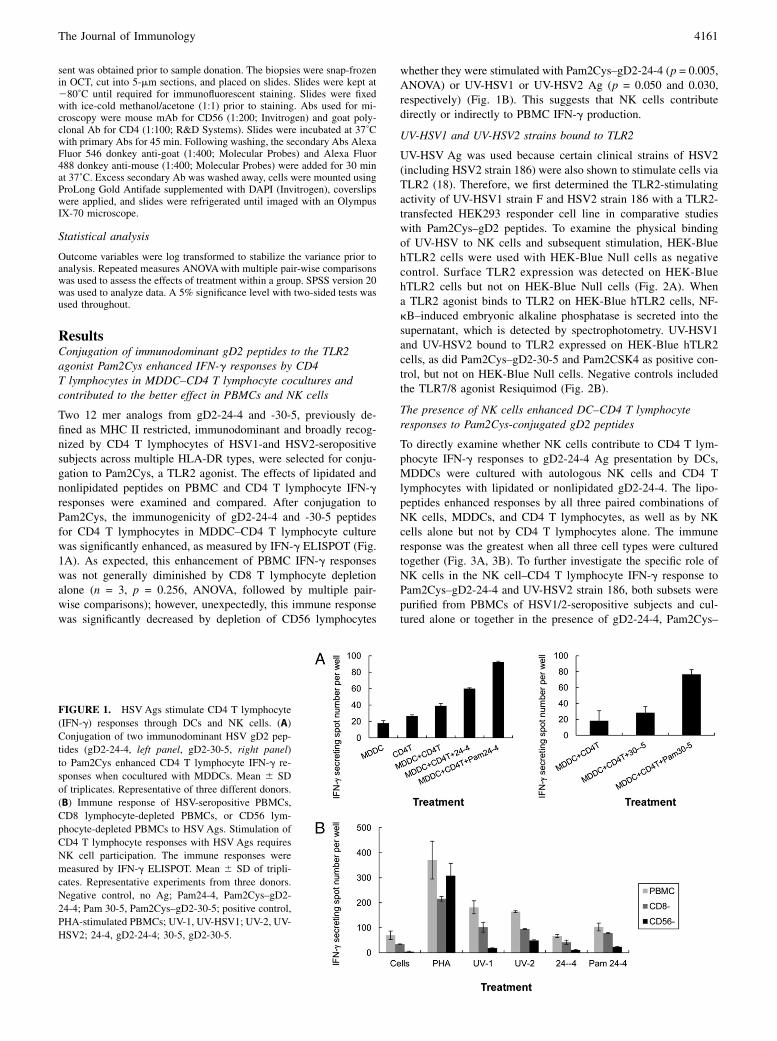
ResultsConjugation of immunodominant gD2 peptides to the TLR2agonist Pam2Cys enhanced IFN-g responses by CD4T lymphocytes in MDDC–CD4 T lymphocyte cocultures andcontributed to the better effect in PBMCs and NK cells
Two 12 mer analogs from gD2-24-4 and -30-5, previously de-fined as MHC II restricted, immunodominant and broadly recog-nized by CD4 T lymphocytes of HSV1-and HSV2-seropositivesubjects across multiple HLA-DR types, were selected for conju-gation to Pam2Cys, a TLR2 agonist. The effects of lipidated andnonlipidated peptides on PBMC and CD4 T lymphocyte IFN-gresponses were examined and compared. After conjugation toPam2Cys, the immunogenicity of gD2-24-4 and -30-5 peptidesfor CD4 T lymphocytes in MDDC–CD4 T lymphocyte culturewas significantly enhanced, as measured by IFN-g ELISPOT (Fig.1A). As expected, this enhancement of PBMC IFN-g responseswas not generally diminished by CD8 T lymphocyte depletionalone (n = 3, p = 0.256, ANOVA, followed by multiple pair-wise comparisons); however, unexpectedly, this immune responsewas significantly decreased by depletion of CD56 lymphocytes
whether they were stimulated with Pam2Cys–gD2-24-4 (p = 0.005,ANOVA) or UV-HSV1 or UV-HSV2 Ag (p = 0.050 and 0.030,respectively) (Fig. 1B). This suggests that NK cells contributedirectly or indirectly to PBMC IFN-g production.
UV-HSV1 and UV-HSV2 strains bound to TLR2
UV-HSV Ag was used because certain clinical strains of HSV2(including HSV2 strain 186) were also shown to stimulate cells viaTLR2 (18). Therefore, we first determined the TLR2-stimulatingactivity of UV-HSV1 strain F and HSV2 strain 186 with a TLR2-transfected HEK293 responder cell line in comparative studieswith Pam2Cys–gD2 peptides. To examine the physical bindingof UV-HSV to NK cells and subsequent stimulation, HEK-BluehTLR2 cells were used with HEK-Blue Null cells as negativecontrol. Surface TLR2 expression was detected on HEK-BluehTLR2 cells but not on HEK-Blue Null cells (Fig. 2A). Whena TLR2 agonist binds to TLR2 on HEK-Blue hTLR2 cells, NF-kB–induced embryonic alkaline phosphatase is secreted into thesupernatant, which is detected by spectrophotometry. UV-HSV1and UV-HSV2 bound to TLR2 expressed on HEK-Blue hTLR2cells, as did Pam2Cys–gD2-30-5 and Pam2CSK4 as positive con-trol, but not on HEK-Blue Null cells. Negative controls includedthe TLR7/8 agonist Resiquimod (Fig. 2B).
The presence of NK cells enhanced DC–CD4 T lymphocyteresponses to Pam2Cys-conjugated gD2 peptides
To directly examine whether NK cells contribute to CD4 T lym-phocyte IFN-g responses to gD2-24-4 Ag presentation by DCs,MDDCs were cultured with autologous NK cells and CD4 Tlymphocytes with lipidated or nonlipidated gD2-24-4. The lipo-peptides enhanced responses by all three paired combinations ofNK cells, MDDCs, and CD4 T lymphocytes, as well as by NKcells alone but not by CD4 T lymphocytes alone. The immuneresponse was the greatest when all three cell types were culturedtogether (Fig. 3A, 3B). To further investigate the specific role ofNK cells in the NK cell–CD4 T lymphocyte IFN-g response toPam2Cys–gD2-24-4 and UV-HSV2 strain 186, both subsets werepurified from PBMCs of HSV1/2-seropositive subjects and cul-tured alone or together in the presence of gD2-24-4, Pam2Cys–
FIGURE 1. HSV Ags stimulate CD4 T lymphocyte
(IFN-g) responses through DCs and NK cells. (A)
Conjugation of two immunodominant HSV gD2 pep-
tides (gD2-24-4, left panel, gD2-30-5, right panel)
to Pam2Cys enhanced CD4 T lymphocyte IFN-g re-
sponses when cocultured with MDDCs. Mean 6 SD
of triplicates. Representative of three different donors.
(B) Immune response of HSV-seropositive PBMCs,
CD8 lymphocyte-depleted PBMCs, or CD56 lym-
phocyte-depleted PBMCs to HSVAgs. Stimulation of
CD4 T lymphocyte responses with HSV Ags requires
NK cell participation. The immune responses were
measured by IFN-g ELISPOT. Mean 6 SD of tripli-
cates. Representative experiments from three donors.
Negative control, no Ag; Pam24-4, Pam2Cys–gD2-
24-4; Pam 30-5, Pam2Cys–gD2-30-5; positive control,
PHA-stimulated PBMCs; UV-1, UV-HSV1; UV-2, UV-
HSV2; 24-4, gD2-24-4; 30-5, gD2-30-5.
The Journal of Immunology 4161

gD2-24-4, or UV-HSV2. However, the NK cell–CD4 T lymphocytecombination again produced more IFN-g than NK cells alone andin the absence of other APCs (Fig. 3C). Thus, NK cells produceIFN-g alone and probably stimulate CD4 T lymphocytes to enhancetheir production with both UV-HSV2 and Pam2Cys–gD2-24-4.
Both Pam2Cys–gD2 peptide and UV-HSV directly activatedNK cells
Whether NK cells are activated by Pam2Cys–gD2 peptide or UV-HSV Ag was determined by measuring expression of the earlyactivation marker CD69 on the surface of NK cells, after stimu-lation by these HSV Ags. Both types of Ags directly and specif-ically activated NK cells, similar to the IFN-g responses (by
ELISPOT) (Fig. 4 compared with Fig. 3). Both CD56brightCD162
and CD56dimCD16+ NK cell subsets were activated, but activationwas greater in the former (data not shown).
Pam2Cys–gD2 peptide and UV-HSV Ag-pulsed NK cellsactivated CD4 T lymphocytes
To confirm the activation of CD4 T lymphocytes by NK cells pulsedwith Pam2Cys–gD2 peptide and UV-HSV, the expression of CD69on CD4 T lymphocytes was examined by flow cytometry aftercoculturing with Ag-stimulated NK cells. The activation of CD4T lymphocytes was significantly enhanced by Pam2Cys conju-gation of the gD2 peptides (Fig. 5A) and by UV-HSV1 stimulation(Fig. 5C), although there was considerable individual variation
FIGURE 2. Binding of UV-HSV1 and UV-
HSV2 to TLR2. (A) HEK-Blue Null cells show
no expression of TLR2 on the surface, whereas
HEK-Blue hTLR2 cells do. (B) HEK-Blue Null
cells and HEK-Blue hTLR2 cells were stimu-
lated with Pam2CSK4 (10 mM), Pam2Cys–gD2-
30-5 (10 mM), UV-HSV1 (MOI 0.5), UV-HSV2
(MOI 0.5), and water and TLR7/8 agonist Resi-
quimod (5 mg/ml) as negative control for 20 h at
37˚C, 5% CO2. NF-kB–induced secreted embry-
onic alkaline phosphatase activity in culture su-
pernatants was measured using QUANTI-BLUE
medium and spectrophotometry (Victor2TM 1420
Multilabel Counter). OD was checked at 650 nm.
*p , 0.05, versus negative control.
FIGURE 3. NK cells augment HSVAg-induced IFN-g responses of CD4 T lymphocytes alone or with MDDCs. (A) Immune responses of MDDCs, NK
cells, and CD4 T lymphocytes in different combinations (MDDC–CD4 T, MDDC–CD4 T–NK, NK alone, and NK–CD4 T lymphocytes) to gD2 peptide,
with or without Pam2Cys. The response was measured by IFN-g ELISPOT. The predetermined ratio of MDDCs, NK cells, and CD4 T lymphocytes was
1:1:10. (B) Immune responses of MDDC–CD4 T, MDDC–NK cell, and MDDC–NK cell–CD4 T lymphocytes to gD2 peptides, with or without Pam2Cys
conjugation. (C) NK cells alone and NK cell–CD4 T lymphocytes were cultured in the presence or absence of HSVAgs and in the absence of MDDCs. The
immune response was measured by IFN-g ELISPOT. Mean 6 SD of triplicates. Representative experiments from three donors in (A)–(C). Pam24-4,
Pam2Cys–gD2-24-4; 24-4, gD2-24-4.
4162 HSV Ag-STIMULATED NK–CD4 T CELL INTERACTIONS
(Fig. 5). CD4 T lymphocytes were not directly activated by the Ag(Fig. 5B), as also shown in Fig. 3B.
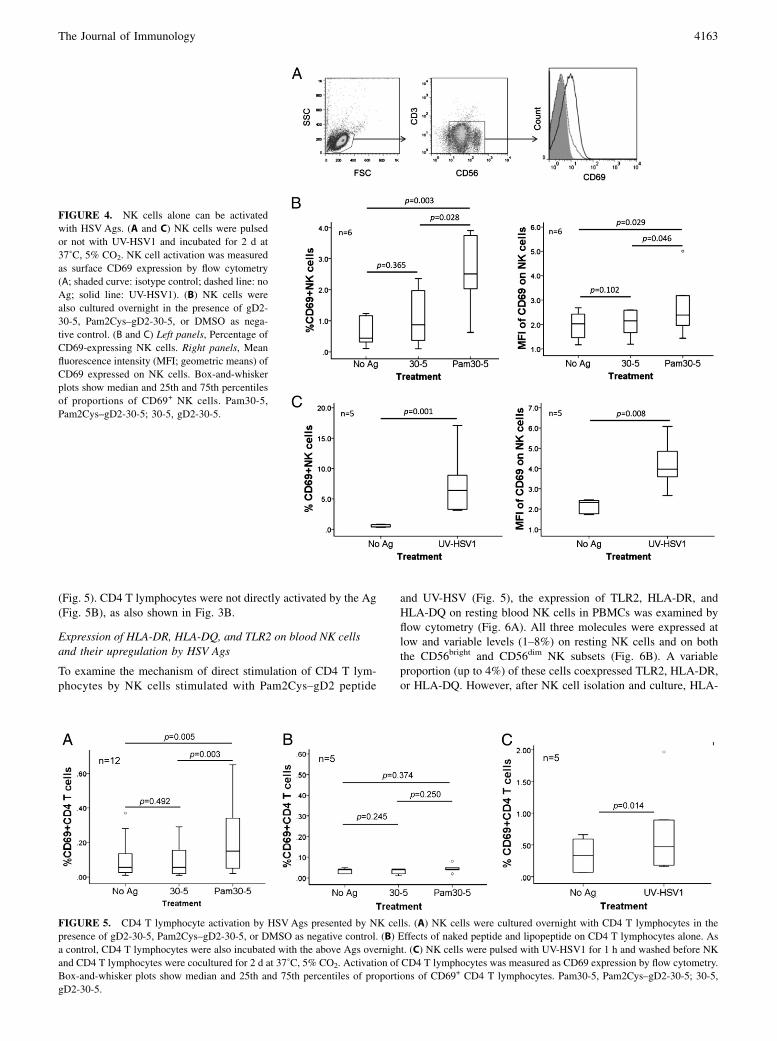
Expression of HLA-DR, HLA-DQ, and TLR2 on blood NK cellsand their upregulation by HSV Ags
To examine the mechanism of direct stimulation of CD4 T lym-phocytes by NK cells stimulated with Pam2Cys–gD2 peptide
and UV-HSV (Fig. 5), the expression of TLR2, HLA-DR, andHLA-DQ on resting blood NK cells in PBMCs was examined byflow cytometry (Fig. 6A). All three molecules were expressed atlow and variable levels (1–8%) on resting NK cells and on boththe CD56bright and CD56dim NK subsets (Fig. 6B). A variableproportion (up to 4%) of these cells coexpressed TLR2, HLA-DR,or HLA-DQ. However, after NK cell isolation and culture, HLA-
FIGURE 4. NK cells alone can be activated
with HSV Ags. (A and C) NK cells were pulsed
or not with UV-HSV1 and incubated for 2 d at
37˚C, 5% CO2. NK cell activation was measured
as surface CD69 expression by flow cytometry
(A; shaded curve: isotype control; dashed line: no
Ag; solid line: UV-HSV1). (B) NK cells were
also cultured overnight in the presence of gD2-
30-5, Pam2Cys–gD2-30-5, or DMSO as nega-
tive control. (B and C) Left panels, Percentage of
CD69-expressing NK cells. Right panels, Mean
fluorescence intensity (MFI; geometric means) of
CD69 expressed on NK cells. Box-and-whisker
plots show median and 25th and 75th percentiles
of proportions of CD69+ NK cells. Pam30-5,
Pam2Cys–gD2-30-5; 30-5, gD2-30-5.
FIGURE 5. CD4 T lymphocyte activation by HSVAgs presented by NK cells. (A) NK cells were cultured overnight with CD4 T lymphocytes in the
presence of gD2-30-5, Pam2Cys–gD2-30-5, or DMSO as negative control. (B) Effects of naked peptide and lipopeptide on CD4 T lymphocytes alone. As
a control, CD4 T lymphocytes were also incubated with the above Ags overnight. (C) NK cells were pulsed with UV-HSV1 for 1 h and washed before NK
and CD4 T lymphocytes were cocultured for 2 d at 37˚C, 5% CO2. Activation of CD4 T lymphocytes was measured as CD69 expression by flow cytometry.
Box-and-whisker plots show median and 25th and 75th percentiles of proportions of CD69+ CD4 T lymphocytes. Pam30-5, Pam2Cys–gD2-30-5; 30-5,
gD2-30-5.
The Journal of Immunology 4163

DR and HLA-DQ expression decreased, reducing the proportionof TLR2+MHC II+ NK cells (M. Kim and N. Osborne, unpub-lished observations).The expression of HLA-DR and HLA-DQ molecules on the
surface of cultured NK cells was then examined, after stimulationwith both types of HSV Ags, and shown to be significantly in-creased (Fig. 7). When NK cell subsets were examined, the lev-els of HLA-DR and HLA-DQ and the proportion of MHC II-expressing NK cells were significantly increased and to a greaterdegree by lipopeptide on the CD56bright CD162 NK subset com-pared with the CD56dim CD16+ NK subset (Fig. 7A).
Pam2Cys–gD2 peptide and UV-HSV Ag-stimulated NK cellsactivated CD4 T lymphocyte responses through cell-to-cellcontact and cytokine effects
To determine whether CD4 T lymphocytes needed to directlycontact NK cells to be activated in an Ag-dependent manner, theywere cultured separately or together in the same compartment inTranswells. Because NK cells were postulated to be APCs, thepeptidewas always added to the chamber containingNK cells. Afterovernight culture, cells were harvested and labeled for flowcytometry. In all five donors, CD69 expression was enhanced toa greater degree on CD4 T lymphocytes when they were coculturedwith NK cells in the same chamber (Fig. 8A); there was an overallsignificant decrease in expression when the two cell types wereseparated, although this showed marked interdonor variation inmagnitude (1.69-fold decrease; 95% confidence interval, 1.20–2.37; p = 0.013, paired t test) (Fig. 8B). There was no significantreduction in naked peptide-stimulated NK cell–CD4 responses byseparate cultures of the two cultures in Transwells (p = 0.273,paired t test). Separate cultures of NK cell and CD4 T lymphocyteshad no consistent or significant effect on NK cell activation (Fig.
8A), indicating that there was little feedback effect of activatedCD4 T lymphocytes on activation of NK cells.Thus, cell-to-cell contact between NK and CD4 T lymphocytes
varied between donors with regard to the magnitude of its im-portance for CD4 T lymphocyte activation, being complemented bythe humoral (or cytokine) effects (similar results were observedwith UV-HSV Ag stimulation; data not shown).
NK cells formed ISs with CD4 T lymphocytes after stimulationwith Pam2Cys–gD2 peptide and UV-HSV1
To examine the physical contact between NK cells and CD4T lymphocytes, highly purified NK cells were pulsed withPam2Cys–gD2-30-5, UV-HSV1 Ag, or controls for 1 h and thenwashed prior to examining for conjugate formation with CD4T lymphocytes. After 5 min of coculture, NK cells and CD4T lymphocytes were fixed/permeabilized and labeled for immu-nofluorescence microscopy. A very low background level of ho-motypic and heterotypic cell-conjugate formation was observedbetween NK cells and CD4 T lymphocytes in control cultures(i.e., CD4–CD4, NK–NK, and NK–CD4 T lymphocyte conjugatesformed) (Fig. 9A). However, the proportion of NK cell–CD4T lymphocyte conjugates in the Pam2Cys–gD2-30-5–stimulatedcocultures was significantly greater than in controls. As expected,these proportions were very low in the lipopeptide-stimulated NKcells. Therefore, to increase the frequency of IS formation be-tween NK cells and CD4 T lymphocytes, gD2-30-5–stimulatedCD4 T cell lines were generated. As a result, ISs were morefrequent and recognized by polarization of the actin-bindingmolecule talin and also the CD4 molecule to the CD4 T lym-phocyte–NK cell contact region with the Pam2Cys–gD2-30-5–stimulated CD4 T cell line (1 IS/104 cell pair for gD2-specificpeptide). Similar ISs were observed in UV-HSV1–pulsed NK
FIGURE 6. Coexpression of HLA-DR,
HLA-DQ, and TLR2 on fresh unstimulated
NK cells in PBMCs. (A) Cells were gated on
live lymphocytes (left panel), then on CD56+
CD32 cells (middle panel), and then on
resting cells expressing TLR2, HLA-DR, or
HLA-DQ (far right panels). The near right
panels show isotype controls. TLR2, HLA-
DR and DQ were often coexpressed on the
same NK cells. (B) Proportion of CD56bright
or CD56dim NK subsets expressing TLR2,
HLA-DR, HLA-DQ, or combinations.
4164 HSV Ag-STIMULATED NK–CD4 T CELL INTERACTIONS

cells with CD4 T lymphocytes (1 IS/3 3 105 cell pair) (Fig. 9B,9C, respectively). Although the cells partnering CD4 T lympho-
cytes were of similar size and CD42 and, therefore, very un-
likely to be contaminating DCs, this possibility was excluded
by their quantification in the purified NK population. Contami-
nating CD11c+lin2 myeloid DCs or BDCA2+lin2 pDCs were
undetectable in the 2% of non-CD56+CD32 NK cells (as were
CD19+ B lymphocytes; i.e., a maximum concentration ,1026). If
the maximum concentrations of Ag-specific CD4 T lymphocytes
were ,5 3 1022 in the cell lines and ,2 3 1023 in UV-HSV
Ag-stimulated CD4 T lymphocytes, then the frequency of any
IS formed by potential contaminating cells with CD4 T lym-
phocytes was calculated to be .3 logs lower than that actually
observed.
NK cells and CD4 T lymphocytes were in contact in themononuclear dermal infiltrates in recurrent herpetic skinlesions
To determine whether these direct NK cell–CD4 T lymphocyteinteractions may occur in vivo, biopsies from three recurrentherpetic lesions at #4 d after appearance were stained for the twocell types. CD4 T lymphocytes were highly abundant in the in-filtrate. CD56+ NK cells were also detected, with up to 27 NKcells/field of view (340 magnification). For each biopsy, thenumber of CD56+ NK cells was counted in six to eight fields ofview. An average of 57.5% of the CD56+ NK cells was found to becontacting a CD4 T lymphocyte in the lesions (Fig. 10). In normalskin control biopsies, no infiltrating CD4 T lymphocytes or CD56+
NK cells were observed in the dermis.
FIGURE 7. MHC II expression by NK cells is upregulated with HSV Ags. (A) NK cells were incubated with CD4 T lymphocytes overnight in the
presence of gD2-30-5, Pam2Cys–gD2-30-5, or DMSO as negative control. The proportion of HLA-DR–expressing cells (Aa) and HLA-DQ–expressing
cells (Ab) (total NK cells [left panels], CD56bright NK subset [middle panels], and CD56dim NK subset [right panels]) were measured by flow cytometry. (B)
UV-HSV1–stimulated NK cells were incubated with CD4 T lymphocytes for 2 d at 37˚C, 5% CO2, and the proportion of NK cells expressing HLA-DR (left
panel) or HLA-DQ (right panel) was measured by flow cytometry. Medium was used as negative control for UV-HSV1. Box-and-whisker plots show
median and 25th and 75th percentiles. Pam30-5, Pam2Cys–gD2-30-5; 30-5, gD2-30-5.
The Journal of Immunology 4165

DiscussionBoth CD4 and CD8 T lymphocytes are key effectors in humancutaneous herpetic lesions and persist as sentinels between lesions:CD8 T lymphocytes at the dermo–epidermal junction and CD4T lymphocytes deeper in the dermis (3, 49). They show differentkinetics of recruitment into clinical recurrent lesions and appar-ently different, but overlapping, functions. Early in inflammation,CD4 T lymphocytes are the major IFN-g–producing cells, al-though they are capable of cytolytic function against infectedkeratinocytes in vitro (1). IFN-g plays an important role in HSV-infected sites by restoring MHC class I downregulation of HSV-infected cells and upregulating MHC II molecules, allowing thelater-infiltrating CD8 T lymphocytes to recognize and clear in-fected cells. However, they also can secrete IFN-g. Recently, theload of HSV2 shed in vaginal secretions was shown to be inverselyproportional to the density of lesional CD8 T lymphocytes, al-though this could just represent the density of all key immuneeffectors (3).NK cells are also present in the dermis of cutaneous herpetic
lesions, in contact with other infiltrating immune cells, especiallypDCs (17), as well as in cells expanded from genital sites of HSV2infection ex vivo (2). An important role for NK cells in HSV in-fection was suggested previously: Dalloul et al. (50) observedsevere HSV2 infection in patients with myelodysplasia and un-detectable NK cells and pDCs in the blood. Nandakumar et al.(51) demonstrated that mice lacking or depleted of NK cells weresusceptible to HSV-induced lesions and that NK cell supplemen-tation enhanced the function of anti-HSV CD8 T lymphocytes.Nevertheless, the role of NK cells in human HSV immunology hasbeen underinvestigated, despite the likely contribution of both NKcell subsets to control of HSV. The minor CD56brightCD162
subset produces IFN-g, and the major CD56dimCD16+ subset iscytotoxic. In this study, we determined that NK cells may play
a pivotal role in interacting with CD4 T lymphocytes, as well asDCs, to augment their response to HSV Ag and, specifically, to
gD2 peptides, especially after TLR2 stimulation on both DCs and
NK cells.In general, short peptides, such as the immunodominant gD2 12
mers, gD2-24-4 and gD2-30-5 used in this study, are poor im-
munogens and need to be administered with adjuvants to max-
imize their immunogenicity. Such adjuvants are often TLR ago-
nists, administered concurrently with Ag, which act on DCs and
macrophages via TLRs (e.g., d-monophosphoryl lipid A and CpG
oligonucleotides) (52). However, the TLR2 agonist Pam2Cys acts
as a self-adjuvant when directly conjugated to peptides. It then
binds specifically to TLR2 on APCs and directs peptide uptake by
the same cells (53–55). After uptake of these in PBMCs, pre-
sumably by TLR2-expressing NK cells, monocytes, and myeloid
DCs, Pam2Cys conjugation to the two gD2 peptides markedly
enhanced CD4 T lymphocyte IFN-g responses, as shown by IFN-g
ELISPOT. As expected from previous reports indicating that CD4
T lymphocytes are the main IFN-g producers in this setting (1, 56,
57), CD8 T lymphocyte depletion did not affect these responses.
However, unexpectedly, CD56 lymphocyte depletion markedly
decreased such responses to Pam2Cys–gD2 peptides, as well as to
UV-HSV1 and UV-HSV2 shown to bind to TLR2 in a reporter cell
FIGURE 8. Pam2Cys–gD2 peptide-stimulated NK cells activated CD4
T lymphocytes through cell-to-cell contact and through cytokine effects.
CD4 T lymphocytes from five donors were cocultured with autologous NK
cells in the lower chamber of Transwells, or NK cells were cultured sep-
arately from CD4 T lymphocytes in the upper chamber. Pam2Cys–gD2-
24-4 or the control gD2-24-4 peptides were always added to the chamber
containing NK cells. The pore size of the Transwell membrane was 0.4
mM to inhibit cell migration between the upper and lower chambers. After
overnight culture, cells were harvested and labeled. Activation of both NK
cells (A) and CD4 T lymphocytes (B) was measured by CD69 expression.
Pam24-4, Pam2Cys–gD2-24-4; 24-4, gD2-24-4.
FIGURE 9. HSV Ag-stimulated NK cells and CD4 T lymphocytes in-
teract and form ISs. (A) NK–CD4 cell pairs without or before IS formation.
(B) Pam2Cys–gD2-30-5–stimulated NK cell–gD2-30-5–specific CD4 T
lymphocyte conjugates showing IS. (C) UV-HSV1–stimulated NK cell–
CD4 conjugates showing IS. Purified CD4 T lymphocytes from stimulated
autologous PBMCs were used in (A) and (C). A gD2-30-5–specific CD4 T
cell line was used in (B). NK cells were isolated from PBMCs and pulsed
with Pam2Cys–gD2-30-5 (B) or UV-HSV1 (C) for 1 h; Ag was washed out
prior to coculture with CD4 T lymphocytes for 5 min at 37˚C. (A) Non-
specific conjugate of NK and CD4 T lymphocytes. Talin was expressed on
both cells and was evenly distributed around the membrane, with more on
NK cells. (B and C) IS formed between NK and CD4 T lymphocyte
conjugates. Talin and CD4 were polarized at the contact site of NK and
CD4 T lymphocytes. Representative images of five ISs each observed in
three individuals (original magnification 3100). Scale bar, 5 mM (A, B);
20 mM (C). (D) Frequency of ISs between lipopeptide or UV-HSV1 Ag-
pulsed NK cells and CD4 T lymphocyte cells or cell lines.
4166 HSV Ag-STIMULATED NK–CD4 T CELL INTERACTIONS

line; this is important because not all HSV strains bind TLR2. Twopossible explanations for the IFN-g production were considered:direct IFN-g secretion by NK cells or interactions between DCs,NK cells, and CD4 T lymphocytes, resulting in the enhancementof CD4 T lymphocyte IFN-g production, or most likely, both mayoccur in combination.To examine the relative contributions of each of the three cell
types, they were studied in pairs and triplets after stimulation withthe gD2 lipopeptides and UV-HSV. The presence of NK cellssignificantly enhanced CD4 T lymphocyte responses primed byMDDCs stimulated with Pam2Cys–gD2-24-4 or by UV-HSV. Suchcross-talk by direct contact between DCs and NK cells (17, 58–61), as well as between CD4 T lymphocytes and NK cells throughOX40–OX40 ligand or 2B4–CD48 interactions (62, 63), has beendescribed, especially in murine systems. Fehniger et al. (64) alsoreported an interaction among DCs, NK cells, and CD4 T lym-phocytes through proinflammatory cytokines, such as IL-12,IFN-g, and IL-2. NK cells can mature DCs and induce the Th1transcription factor T-bet in CD4 T lymphocytes through IFN-gproduction (65, 66), both of which promote Th1 responses. Hannaet al. (28) reported that NK cells (especially the CD56brightCD162
subset) activated by PHA and IL-2 can present Ag, and they de-fined distinct proteomic and genomic profiles of the two NK cellsubsets (67, 68).In this study, we showed that isolatedNK and CD4T lymphocytes
in coculture could also respond to Pam2Cys–gD2 peptides and toUV-HSV by IFN-g production, but the NK cell–CD4 T lymphocyteinteraction produced significantly more IFN-g than NK cells alone.NK cells alone were directly activated, as shown by CD69 ex-pression and IFN-g production, reflecting expression of TLR2 onhuman NK cells (35, 69, 70) but not on resting CD4 T lymphocytes.Furthermore, CD4 T lymphocyte activation by HSVAg-pulsed NKcells was confirmed by upregulated CD69 expression, stronglysuggesting that both cell types were sources for IFN-g. We furthershowed that TLR2, HLA-DR, and HLA-DQ were expressed andalso coexpressed on a small proportion of unstimulated NK cells, as
well as both NK subsets (although MHC II expression decreased onculture in the absence of HSVAgs). Thus, HSVAg binding to TLR2could upregulate HLA-DR and HLA-DQ on the same NK cells orinduce their expression de novo to a greater extent on the CD56bright
CD162 NK subset.To determine whether direct contact between NK cell and CD4
T lymphocytes was required for CD4 T lymphocyte activation,a Transwell system was used. In all five donors, cell-to-cell contactenhanced CD4 T lymphocyte activation; although there was aconsiderable variation in the magnitude of this effect betweendonors, overall it was significant. Where the effect of contact wasminor, activation of CD4 T lymphocytes probably occurred throughNK cytokine production, in particular IFN-g, as previously de-scribed (64). Cytokines, such as IL-12 and IL-15 from DCs, IFN-gfrom NK cells, and IL-2 from CD4 T lymphocytes, may allcontribute to these interactions. There may be several reasons forthese differences in the relative importance of contact and cyto-kines between individual donors. These may include individualvariations in cytokine production, TLR2 expression on NK cells,or expression of cell-adhesion molecules between NK cells andCD4 T lymphocytes. Separation of NK cells and CD4 T lym-phocytes had little effect on HSV Ag/TLR2-induced NK activa-tion, arguing against a significant role for activated CD4 T lym-phocyte feedback on NK activation.To define the mechanism of Ag presentation to CD4 T lym-
phocytes by contacting NK cells pulsed with lipopeptide orwhole UV-HSVAg, their HLA-DR and HLA-DQ expression wasexamined. Both MHC II molecules were significantly upregulatedon NK cells, and HLA-DR expression was upregulated to thegreatest extent on the CD56brightCD162 NK cell subset (data notshown). Expression of MHC II molecules on human NK cells asmRNA and protein was reported (67, 68) but not previously shownto be upregulated by lipopeptide or a viral pathogen, althoughsimilar lipopeptides induce maturation and upregulate MHC II onDCs (71).Direct cell-to-cell contact between NK cells and CD4 T lym-
phocytes, resulting in Ag presentation, is likely to occur viaISs. MHC–peptide complexes reach a peak ∼5 min after cocultureand remain maximally concentrated for 10–20 min (72). Adhesionmolecules in the clusters increase the avidity of the APC–T cellinteraction, thus facilitating more efficient TCR engagement. Theyare polarized to the contact region between APCs and lymphocytes(73). The actin-binding molecule talin is one of the molecules thatpolarizes to ISs. In DC–NK cell ISs, F-action polymerizationoccurs within DCs, whereas in cytolytic NK cell–K562 conjugates,F-actin polymerization occurs in the NK cells (74). Talin polari-zation is present in ISs formed by noncytolytic and cytolytic NKcells but is transient in noncytolytic ISs (∼1 min) when interact-ing with target cells (75). If true for interactions with CD4T lymphocytes, this could lead to underestimates of ISs in oursystem. CD4 molecules are also involved in IS formation within 5min (76). In our study, ISs were observed as colocalization of CD4and talin at the contact region with the expected kinetics, 5 minafter coculture, of CD4 T lymphocytes and NK cells stimulatedwith Pam2Cys–gD2-30-5 or UV-HSVAg. To our knowledge, thisis the first report of physical contact between NK cells and CD4T lymphocytes through ISs.At all time points, significantly more NK–CD4 cell conjugates
were observed with Pam2Cys–gD2 peptide or UV-HSV Ag stim-ulation than in control unstimulated cultures. Low proportionsof NK–NK and CD4–CD4 cell conjugates were also found inPam2Cys–gD2 peptide-stimulated samples and are probably non-specific. As previously reported, unstimulated naive lymphocytes
FIGURE 10. Direct contact of CD4 T lymphocytes and NK cells in
dermal infiltrate of recurrent herpes lesions. NK cells (green) are juxta-
posed and in contact with CD4 T lymphocytes (red) in recurrent herpetic
lesions, biopsied 4 d after onset at 340 magnification (A) and 3100
magnification (B). Scale bar, 10 mM. (C) Mean proportion (6 SD) of NK
cells in contact with CD4 T lymphocytes as a ratio of total NK cells in six
to eight high-power fields (340)/biopsy from three donors.
The Journal of Immunology 4167

and DCs can form conjugates in the absence of Ag, which may playa major role in the maintenance of naive lymphocytes in vivo (77).Specific NK cell–CD4 T lymphocyte IS formation, shown by
talin and CD4 polarization to the IS, was rare and reflects the lowfrequency of circulating memory CD4 T lymphocytes specific forsingle gD2 peptides. The mean frequency of whole-HSV Ag-specific memory CD4 T lymphocytes is reported to be ∼0.2%of total circulating CD4 T lymphocytes (78). Less than 5% ofNK cells (probably mostly the CD56brightCD162 subset) were in-volved in ISs, also correlating with the proportion of NK cellsexpressing HLA-DR (data not shown). Calculation of the ap-proximate specificities of whole-HSV Ags for human T lym-phocytes in seropositive patients suggests that 1 in 10,000 cellsshould form specific ISs, consistent with our experimental re-sults. In an attempt to increase the frequency of IS formation,UV-HSV1 or a gD2-30-5–specific CD4 T cell line was used tostudy IS formation with NK cells. IS formation between UV-HSV1 or Pam2Cys–gD2 peptide-stimulated NK cells and CD4T lymphocytes was observed in different HSV-seropositive do-nors. The mechanism of processing of particulate viral Ags, likeHSV, by NK cells is unclear and is being investigated. Pre-sumably, this involves endocytosis, cathepsin-mediated peptidecleavage, and combination with MHC II molecules in vesicularcompartments, as with other APCs (79).Both CD56brightCD162 and CD56dimCD16+ NK cell subsets are
found in lung, liver, and skin and circulate through peripheralblood. However, the minor subset CD56brightCD162 NK cells aremore abundant in secondary lymphoid organs, accounting for upto 75% of NK cells in lymph nodes and 50% in the spleen (25).CD56brightCD162 NK cells are found in T cell-enriched areas inhuman lymph nodes and tonsils and, thus, can directly contactlymphocytes in their vicinity (29). Postinfection with Leish-mania, major IFN-g–secreting NK cells were found to contact thesame DCs as Ag-specific CD4 T lymphocytes in the lymph node(80). However, a direct interaction between NK cells and CD4T lymphocytes in response to pathogens has not been shownpreviously. In this study, we demonstrate a new role for NK cellsas APCs in the context of HSV peptide conjugated to Pam2Cysand with UV-HSV1 or UV-HSV2, and also show IS formationbetween NK and CD4 T lymphocytes. However, the exact mech-anism of HSV binding to TLR2 is unknown. With human CMV, itis via the major envelope glycoproteins, but that does not seem tobe the case for HSV1 or HSV2, and it also appears to be straindependent (18, 38).In summary, NK cell depletion markedly decreased IFN-g se-
cretion of PBMCs in response to an immunogenic HSV2 peptideconjugated to the TLR2 agonist Pam2Cys, and the presence of NKcells enhanced the resulting immune response to HSV Ags. NKcells alone responded to whole-HSV–inactivated Ags and HSVlipopeptides through TLR2 molecules, then stimulated NK cellsinduced CD4 T cell activation though cytokine and cell-to-cellcontact via MHC II molecules on NK cells. The significance oftheir physical contact was shown by the observation of IS for-mation between them in vitro and contact between them in re-current herpetic lesions in vivo.These findings indicate the importance of interactions between
DCs, NK cells, and CD4 T lymphocytes in pairs and triplets inhuman viral immunology using HSVas an example, as well as themultiple roles of NK cells that extend beyond cytotoxicity andcytokine production to Ag presentation. Furthermore, there arepractical consequences to developing vaccines for HSV. TLR2 (andother TLRs) on NK cells, as well as on DCs, must be considereda potential target for adjuvants to augment protective CD4, andperhaps CD8, T lymphocyte responses.
AcknowledgmentsWe thank Dr. Stuart G. Turville for training M.K. and N.R.O. for Delta-
Vision Microscopy and Fabienne Brillot-Turville for providing the proto-
col for conjugation experiments. We also thank Dr. Karen Byth-Wilson
for statistical analysis.
DisclosuresThe authors have no financial conflicts of interest. A.L.C. has served as
a consultant to GlaxoSmithKline on the development of vaccines for genital
herpes, but no commercial funding was received for this project.
References1. Cunningham, A. L., R. R. Turner, A. C. Miller, M. F. Para, and T. C. Merigan.
1985. Evolution of recurrent herpes simplex lesions. An immunohistologic study.J. Clin. Invest. 75: 226–233.
2. Koelle, D. M., C. M. Posavad, G. R. Barnum, M. L. Johnson, J. M. Frank, andL. Corey. 1998. Clearance of HSV-2 from recurrent genital lesions correlateswith infiltration of HSV-specific cytotoxic T lymphocytes. J. Clin. Invest. 101:1500–1508.
3. Schiffer, J. T., L. Abu-Raddad, K. E. Mark, J. Zhu, S. Selke, D. M. Koelle,A. Wald, and L. Corey. 2010. Mucosal host immune response predicts the se-verity and duration of herpes simplex virus-2 genital tract shedding episodes.Proc. Natl. Acad. Sci. USA 107: 18973–18978.
4. Mikloska, Z., and A. L. Cunningham. 1998. Herpes simplex virus type 1 gly-coproteins gB, gC and gD are major targets for CD4 T-lymphocyte cytotoxicity inHLA-DR expressing human epidermal keratinocytes. J. Gen. Virol. 79: 353–361.
5. Mikloska, Z., and A. L. Cunningham. 2001. Alpha and gamma interferons in-hibit herpes simplex virus type 1 infection and spread in epidermal cells afteraxonal transmission. J. Virol. 75: 11821–11826.
6. Mikloska, Z., M. Ruckholdt, I. Ghadiminejad, H. Dunckley, M. Denis, andA. L. Cunningham. 2000. Monophosphoryl lipid A and QS21 increase CD8T lymphocyte cytotoxicity to herpes simplex virus-2 infected cell proteins 4 and27 through IFN-gamma and IL-12 production. J. Immunol. 164: 5167–5176.
7. Wakim, L. M., J. Waithman, N. van Rooijen, W. R. Heath, and F. R. Carbone.2008. Dendritic cell-induced memory T cell activation in nonlymphoid tissues.Science 319: 198–202.
8. Zhu, J., F. Hladik, A. Woodward, A. Klock, T. Peng, C. Johnston, M. Remington,A. Magaret, D. M. Koelle, A. Wald, and L. Corey. 2009. Persistence of HIV-1receptor-positive cells after HSV-2 reactivation is a potential mechanism forincreased HIV-1 acquisition. Nat. Med. 15: 886–892.
9. Stingl, G., K. Tamaki, and S. I. Katz. 1980. Origin and function of epidermalLangerhans cells. Immunol. Rev. 53: 149–174.
10. Sprecher, E., and Y. Becker. 1992. Detection of IL-1 beta, TNF-alpha, and IL-6gene transcription by the polymerase chain reaction in keratinocytes, Langerhanscells and peritoneal exudate cells during infection with herpes simplex virus-1.Arch. Virol. 126: 253–269.
11. Allan, R. S., C. M. Smith, G. T. Belz, A. L. van Lint, L. M. Wakim, W. R. Heath,and F. R. Carbone. 2003. Epidermal viral immunity induced by CD8alpha+dendritic cells but not by Langerhans cells. Science 301: 1925–1928.
12. Zhao, X., E. Deak, K. Soderberg, M. Linehan, D. Spezzano, J. Zhu, D. M. Knipe,and A. Iwasaki. 2003. Vaginal submucosal dendritic cells, but not Langerhanscells, induce protective Th1 responses to herpes simplex virus-2. J. Exp. Med.197: 153–162.
13. Puttur, F. K., M. A. Fernandez, R. White, B. Roediger, A. L. Cunningham,W. Weninger, and C. A. Jones. 2010. Herpes simplex virus infects skin gammadelta T cells before Langerhans cells and impedes migration of infected Lang-erhans cells by inducing apoptosis and blocking E-cadherin downregulation. J.Immunol. 185: 477–487.
14. Bosnjak, L., M. Miranda-Saksena, D. M. Koelle, R. A. Boadle, C. A. Jones, andA. L. Cunningham. 2005. Herpes simplex virus infection of human dendriticcells induces apoptosis and allows cross-presentation via uninfected dendriticcells. J. Immunol. 174: 2220–2227.
15. Bukowski, J. F., and R. M. Welsh. 1986. The role of natural killer cells andinterferon in resistance to acute infection of mice with herpes simplex virus type1. J. Immunol. 136: 3481–3485.
16. Cheng, H., T. M. Tumpey, H. F. Staats, N. van Rooijen, J. E. Oakes, andR. N. Lausch. 2000. Role of macrophages in restricting herpes simplex virustype 1 growth after ocular infection. Invest. Ophthalmol. Vis. Sci. 41: 1402–1409.
17. Donaghy, H., L. Bosnjak, A. N. Harman, V.Marsden, S. K. Tyring, T. C.Meng, andA. L. Cunningham. 2009. Role for plasmacytoid dendritic cells in the immunecontrol of recurrent human herpes simplex virus infection. J. Virol. 83: 1952–1961.
18. Sato, A., M. M. Linehan, and A. Iwasaki. 2006. Dual recognition of herpessimplex viruses by TLR2 and TLR9 in dendritic cells. Proc. Natl. Acad. Sci.USA 103: 17343–17348.
19. Lund, J., A. Sato, S. Akira, R. Medzhitov, and A. Iwasaki. 2003. Toll-like re-ceptor 9-mediated recognition of Herpes simplex virus-2 by plasmacytoid den-dritic cells. J. Exp. Med. 198: 513–520.
20. Siegal, F. P., N. Kadowaki, M. Shodell, P. A. Fitzgerald-Bocarsly, K. Shah,S. Ho, S. Antonenko, and Y. J. Liu. 1999. The nature of the principal type 1interferon-producing cells in human blood. Science 284: 1835–1837.
21. Fonteneau, J. F., M. Larsson, A. S. Beignon, K. McKenna, I. Dasilva, A. Amara,Y. J. Liu, J. D. Lifson, D. R. Littman, and N. Bhardwaj. 2004. Human immu-
4168 HSV Ag-STIMULATED NK–CD4 T CELL INTERACTIONS

nodeficiency virus type 1 activates plasmacytoid dendritic cells and concomi-tantly induces the bystander maturation of myeloid dendritic cells. J. Virol. 78:5223–5232.
22. Moretta, L., G. Ferlazzo, C. Bottino, M. Vitale, D. Pende, M. C. Mingari, andA. Moretta. 2006. Effector and regulatory events during natural killer-dendriticcell interactions. Immunol. Rev. 214: 219–228.
23. Kadowaki, N., S. Antonenko, J. Y. Lau, and Y. J. Liu. 2000. Natural interferonalpha/beta-producing cells link innate and adaptive immunity. J. Exp. Med. 192:219–226.
24. Kawamura, K., N. Kadowaki, T. Kitawaki, and T. Uchiyama. 2006. Virus-stimulated plasmacytoid dendritic cells induce CD4+ cytotoxic regulatoryT cells. Blood 107: 1031–1038.
25. Strowig, T., F. Brilot, and C. Munz. 2008. Noncytotoxic functions of NK cells:direct pathogen restriction and assistance to adaptive immunity. J. Immunol. 180:7785–7791.
26. Moretta, A. 2005. The dialogue between human natural killer cells and dendriticcells. Curr. Opin. Immunol. 17: 306–311.
27. Ferlazzo, G., M. Pack, D. Thomas, C. Paludan, D. Schmid, T. Strowig, G. Bougras,W. A. Muller, L. Moretta, and C. Munz. 2004. Distinct roles of IL-12 and IL-15 inhuman natural killer cell activation by dendritic cells from secondary lymphoidorgans. Proc. Natl. Acad. Sci. USA 101: 16606–16611.
28. Hanna, J., T. Gonen-Gross, J. Fitchett, T. Rowe, M. Daniels, T. I. Arnon,R. Gazit, A. Joseph, K. W. Schjetne, A. Steinle, et al. 2004. Novel APC-likeproperties of human NK cells directly regulate T cell activation. J. Clin. Invest.114: 1612–1623.
29. Hanna, J., and O. Mandelboim. 2007. When killers become helpers. TrendsImmunol. 28: 201–206.
30. Adam, C., S. King, T. Allgeier, H. Braumuller, C. Luking, J. Mysliwietz,A. Kriegeskorte, D. H. Busch, M. Rocken, and R. Mocikat. 2005. DC-NK cellcross talk as a novel CD4+ T-cell-independent pathway for antitumor CTL in-duction. Blood 106: 338–344.
31. Hart, O. M., V. Athie-Morales, G. M. O’Connor, and C. M. Gardiner. 2005.TLR7/8-mediated activation of human NK cells results in accessory cell-dependent IFN-gamma production. J. Immunol. 175: 1636–1642.
32. Andrews, D. M., C. E. Andoniou, A. A. Scalzo, S. L. van Dommelen,M. E. Wallace, M. J. Smyth, and M. A. Degli-Esposti. 2005. Cross-talk betweendendritic cells and natural killer cells in viral infection. Mol. Immunol. 42: 547–555.
33. Megjugorac, N. J., H. A. Young, S. B. Amrute, S. L. Olshalsky, and P. Fitzgerald-Bocarsly. 2004. Virally stimulated plasmacytoid dendritic cells produce che-mokines and induce migration of T and NK cells. J. Leukoc. Biol. 75: 504–514.
34. Flacher, V., M. Bouschbacher, E. Verronese, C. Massacrier, V. Sisirak,O. Berthier-Vergnes, B. de Saint-Vis, C. Caux, C. Dezutter-Dambuyant,S. Lebecque, and J. Valladeau. 2006. Human Langerhans cells express a specificTLR profile and differentially respond to viruses and Gram-positive bacteria. J.Immunol. 177: 7959–7967.
35. Hornung, V., S. Rothenfusser, S. Britsch, A. Krug, B. Jahrsdorfer, T. Giese,S. Endres, and G. Hartmann. 2002. Quantitative expression of toll-like receptor1-10 mRNA in cellular subsets of human peripheral blood mononuclear cells andsensitivity to CpG oligodeoxynucleotides. J. Immunol. 168: 4531–4537.
36. Morandi, B., G. Bougras, W. A. Muller, G. Ferlazzo, and C. Munz. 2006. NKcells of human secondary lymphoid tissues enhance T cell polarization via IFN-gamma secretion. Eur. J. Immunol. 36: 2394–2400.
37. Sørensen, L. N., L. S. Reinert, L. Malmgaard, C. Bartholdy, A. R. Thomsen, andS. R. Paludan. 2008. TLR2 and TLR9 synergistically control herpes simplexvirus infection in the brain. J. Immunol. 181: 8604–8612.
38. Reske, A., G. Pollara, C. Krummenacher, D. R. Katz, and B. M. Chain. 2008.Glycoprotein-dependent and TLR2-independent innate immune recognition ofherpes simplex virus-1 by dendritic cells. J. Immunol. 180: 7525–7536.
39. Stanberry, L. R., S. L. Spruance, A. L. Cunningham, D. I. Bernstein, A. Mindel,S. Sacks, S. Tyring, F. Y. Aoki, M. Slaoui, M. Denis, et al; GlaxoSmithKlineHerpes Vaccine Efficacy Study Group. 2002. Glycoprotein-D-adjuvant vaccineto prevent genital herpes. N. Engl. J. Med. 347: 1652–1661.
40. Cohen, J. 2010. Immunology. Painful failure of promising genital herpes vac-cine. Science 330: 304.
41. Kim, M., J. Taylor, J. Sidney, Z. Mikloska, N. Bodsworth, K. Lagios,H. Dunckley, K. Byth-Wilson, M. Denis, R. Finlayson, et al. 2008. Immuno-dominant epitopes in herpes simplex virus type 2 glycoprotein D are recognizedby CD4 lymphocytes from both HSV-1 and HSV-2 seropositive subjects. J.Immunol. 181: 6604–6615.
42. Zeng, W., S. Ghosh, Y. F. Lau, L. E. Brown, and D. C. Jackson. 2002. Highlyimmunogenic and totally synthetic lipopeptides as self-adjuvanting immuno-contraceptive vaccines. J. Immunol. 169: 4905–4912.
43. Akira, S., S. Uematsu, and O. Takeuchi. 2006. Pathogen recognition and innateimmunity. Cell 124: 783–801.
44. Kang, J. Y., X. Nan, M. S. Jin, S. J. Youn, Y. H. Ryu, S. Mah, S. H. Han, H. Lee,S. G. Paik, and J. O. Lee. 2009. Recognition of lipopeptide patterns by Toll-likereceptor 2-Toll-like receptor 6 heterodimer. Immunity 31: 873–884.
45. Jin, M. S., S. E. Kim, J. Y. Heo, M. E. Lee, H. M. Kim, S. G. Paik, H. Lee, andJ. O. Lee. 2007. Crystal structure of the TLR1-TLR2 heterodimer induced bybinding of a tri-acylated lipopeptide. Cell 130: 1071–1082.
46. Schenk, M., J. T. Belisle, and R. L. Modlin. 2009. TLR2 looks at lipoproteins.Immunity 31: 847–849.
47. Bettahi, I., X. Zhang, R. E. Afifi, and L. BenMohamed. 2006. Protective im-munity to genital herpes simplex virus type 1 and type 2 provided by self-adjuvanting lipopeptides that drive dendritic cell maturation and elicit a polar-ized Th1 immune response. Viral Immunol. 19: 220–236.
48. Mikloska, Z., L. Bosnjak, and A. L. Cunningham. 2001. Immature monocyte-derived dendritic cells are productively infected with herpes simplex virus type1. J. Virol. 75: 5958–5964.
49. Zhu, J., D. M. Koelle, J. Cao, J. Vazquez, M. L. Huang, F. Hladik, A. Wald, andL. Corey. 2007. Virus-specific CD8+ T cells accumulate near sensory nerveendings in genital skin during subclinical HSV-2 reactivation. J. Exp. Med. 204:595–603.
50. Dalloul, A., E. Oksenhendler, O. Chosidow, P. Ribaud, G. Carcelain, S. Louvet,P. Massip, P. Lebon, and B. Autran. 2004. Severe herpes virus (HSV-2) infectionin two patients with myelodysplasia and undetectable NK cells and plasmacytoiddendritic cells in the blood. J. Clin. Virol. 30: 329–336.
51. Nandakumar, S., S. N. Woolard, D. Yuan, B. T. Rouse, and U. Kumaraguru.2008. Natural killer cells as novel helpers in anti-herpes simplex virus immuneresponse. J. Virol. 82: 10820–10831.
52. Zauner, W., K. Lingnau, F. Mattner, A. von Gabain, and M. Buschle. 2001.Defined synthetic vaccines. Biol. Chem. 382: 581–595.
53. Warren, H. S., F. R. Vogel, and L. A. Chedid. 1986. Current status of immu-nological adjuvants. Annu. Rev. Immunol. 4: 369–388.
54. Ghielmetti, M., A. Reschner, M. Zwicker, and E. Padovan. 2005. Syntheticbacterial lipopeptide analogs: structural requirements for adjuvanticity. Immu-nobiology 210: 211–215.
55. Toth, I. 1994. A novel chemical approach to drug delivery: lipidic amino acidconjugates. J. Drug Target. 2: 217–239.
56. Koelle, D. M., H. Abbo, A. Peck, K. Ziegweid, and L. Corey. 1994. Direct re-covery of herpes simplex virus (HSV)-specific T lymphocyte clones from re-current genital HSV-2 lesions. J. Infect. Dis. 169: 956–961.
57. Cunningham, A. L., and T. C. Merigan. 1984. Leu-3+ T cells produce gamma-interferon in patients with recurrent herpes labialis. J. Immunol. 132: 197–202.
58. Brilot, F., T. Strowig, and C. Munz. 2008. NK cells interactions with dendriticcells shape innate and adaptive immunity. Front. Biosci. 13: 6443–6454.
59. Kassim, S. H., N. K. Rajasagi, B. W. Ritz, S. B. Pruett, E. M. Gardner,R. Chervenak, and S. R. Jennings. 2009. Dendritic cells are required for optimalactivation of natural killer functions following primary infection with herpessimplex virus type 1. J. Virol. 83: 3175–3186.
60. Moretta, A., E. Marcenaro, S. Parolini, G. Ferlazzo, and L. Moretta. 2008. NKcells at the interface between innate and adaptive immunity. Cell Death Differ.15: 226–233.
61. Andoniou, C. E., D. M. Andrews, and M. A. Degli-Esposti. 2006. Natural killercells in viral infection: more than just killers. Immunol. Rev. 214: 239–250.
62. Zingoni, A., T. Sornasse, B. G. Cocks, Y. Tanaka, A. Santoni, and L. L. Lanier.2004. Cross-talk between activated human NK cells and CD4+ T cells via OX40-OX40 ligand interactions. J. Immunol. 173: 3716–3724.
63. Assarsson, E., T. Kambayashi, J. D. Schatzle, S. O. Cramer, A. von Bonin,P. E. Jensen, H. G. Ljunggren, and B. J. Chambers. 2004. NK cells stimulateproliferation of T and NK cells through 2B4/CD48 interactions. J. Immunol. 173:174–180.
64. Fehniger, T. A., M. A. Cooper, G. J. Nuovo, M. Cella, F. Facchetti, M. Colonna,and M. A. Caligiuri. 2003. CD56bright natural killer cells are present in humanlymph nodes and are activated by T cell-derived IL-2: a potential new link be-tween adaptive and innate immunity. Blood 101: 3052–3057.
65. Afkarian, M., J. R. Sedy, J. Yang, N. G. Jacobson, N. Cereb, S. Y. Yang,T. L. Murphy, and K. M. Murphy. 2002. T-bet is a STAT1-induced regulator ofIL-12R expression in naıve CD4+ T cells. Nat. Immunol. 3: 549–557.
66. Lighvani, A. A., D. M. Frucht, D. Jankovic, H. Yamane, J. Aliberti,B. D. Hissong, B. V. Nguyen, M. Gadina, A. Sher, W. E. Paul, and J. J. O’Shea.2001. T-bet is rapidly induced by interferon-gamma in lymphoid and myeloidcells. Proc. Natl. Acad. Sci. USA 98: 15137–15142.
67. Wendt, K., E. Wilk, S. Buyny, J. Buer, R. E. Schmidt, and R. Jacobs. 2006. Geneand protein characteristics reflect functional diversity of CD56dim and CD56bright
NK cells. J. Leukoc. Biol. 80: 1529–1541.68. Hanna, J., P. Bechtel, Y. Zhai, F. Youssef, K. McLachlan, and O. Mandelboim.
2004. Novel insights on human NK cells’ immunological modalities revealed bygene expression profiling. J. Immunol. 173: 6547–6563.
69. Lauzon, N. M., F. Mian, R. MacKenzie, and A. A. Ashkar. 2006. The directeffects of Toll-like receptor ligands on human NK cell cytokine production andcytotoxicity. Cell. Immunol. 241: 102–112.
70. Martinez, J., X. Huang, and Y. Yang. 2010. Direct TLR2 signaling is critical forNK cell activation and function in response to vaccinia viral infection. PLoSPathog. 6: e1000811.
71. Nesburn, A. B., I. Bettahi, X. Zhang, X. Zhu, W. Chamberlain, R. E. Afifi,S. L. Wechsler, and L. BenMohamed. 2006. Topical/mucosal delivery of sub-unitvaccines that stimulate the ocular mucosal immune system. Ocul. Surf. 4: 178–187.
72. Vyas, Y. M., H. Maniar, and B. Dupont. 2002. Visualization of signaling path-ways and cortical cytoskeleton in cytolytic and noncytolytic natural killer cellimmune synapses. Immunol. Rev. 189: 161–178.
73. Abraham, C., J. Griffith, and J. Miller. 1999. The dependence for leukocytefunction-associated antigen-1/ICAM-1 interactions in T cell activation cannot beovercome by expression of high density TCR ligand. J. Immunol. 162: 4399–4405.
74. Barreira da Silva, R., C. Graf, and C. Munz. 2011. Cytoskeletal stabilization ofinhibitory interactions in immunologic synapses of mature human dendritic cellswith natural killer cells. Blood 118: 6487–6498.
75. Vyas, Y. M., H. Maniar, and B. Dupont. 2002. Cutting edge: differential seg-regation of the SRC homology 2-containing protein tyrosine phosphatase-1within the early NK cell immune synapse distinguishes noncytolytic from cy-tolytic interactions. J. Immunol. 168: 3150–3154.
The Journal of Immunology 4169

76. Monks, C. R., B. A. Freiberg, H. Kupfer, N. Sciaky, and A. Kupfer. 1998. Three-dimensional segregation of supramolecular activation clusters in T cells. Nature395: 82–86.
77. Revy, P., M. Sospedra, B. Barbour, and A. Trautmann. 2001. Functional antigen-independent synapses formed between T cells and dendritic cells. Nat. Immunol.2: 925–931.
78. Asanuma, H., M. Sharp, H. T. Maecker, V. C. Maino, and A. M. Arvin. 2000.Frequencies of memory T cells specific for varicella-zoster virus, herpes simplex
virus, and cytomegalovirus by intracellular detection of cytokine expression. J.Infect. Dis. 181: 859–866.
79. Watts, C. 2004. The exogenous pathway for antigen presentation on majorhistocompatibility complex class II and CD1 molecules. Nat. Immunol. 5: 685–692.
80. Bajenoff, M., B. Breart, A. Y. Huang, H. Qi, J. Cazareth, V. M. Braud,R. N. Germain, and N. Glaichenhaus. 2006. Natural killer cell behavior in lymphnodes revealed by static and real-time imaging. J. Exp. Med. 203: 619–631.
4170 HSV Ag-STIMULATED NK–CD4 T CELL INTERACTIONS




















