
Postharvest heat and conditioning treatments activate different molecular responses and reduce...
11
RESEARCH PAPER Postharvest heat and conditioning treatments activate different molecular responses and reduce chilling injuries in grapefruit Margarita Sapitnitskaya 1 , Pilar Maul 2 , Gregory T. McCollum 2 , Charles L. Guy 3 , Batia Weiss 1 , Alon Samach 4 and Ron Porat 1, * 1 Department of Postharvest Science, ARO, the Volcani Center, PO Box 6, Bet Dagan 50250, Israel 2 USDA, USHRL, 2001 S. Rock Rd, Ft Pierce, FL 34945, USA 3 Department of Environmental Horticulture, University of Florida, Gainesville, FL 32611, USA 4 Institute of Plant Sciences and Genetics, Faculty of Agricultural, Food and Environmental Quality Sciences, the Hebrew University of Jerusalem, PO Box 12, Rehovot 76100, Israel Received 5 April 2006; Accepted 15 May 2006 Abstract A combination of hot water (a rinse at 62 8C for 20 s) and conditioning (pre-storage at 16 8C for 7 d) treat- ments synergistically reduced chilling injury develop- ment in grapefruit (Citrus paradisi, cv. ‘Star Ruby’) during cold storage at 2 8C, suggesting that the treatments may activate different chilling tolerance responses. To study the molecular mechanisms in- volved, chilling- and conditioning-responsive genes were isolated by polymerase chain reaction (PCR) cDNA subtraction, cDNA libraries were constructed from hot water- and conditioning-treated fruit, and cDNA sequencing was used to identify putative stress- responsive and chilling tolerance genes. PCR cDNA subtraction revealed the identification of 17 chilling- responsive and heat- and conditioning-induced genes, and the expression patterns of 11 additional stress- related genes, antioxidant defensive genes, and genes encoding enzymes involved in membrane lipid mod- ifications were characterized. It was found that hot water and conditioning treatments had little effect on gene expression by themselves, but rather had a priming effect, and enabled the fruit to activate their defence responses after subsequent exposure to chilling. RNA gel blot hybridizations revealed that the expression patterns of eight genes, including HSP19-I, HSP19-II, dehydrin, universal stress protein (USP), EIN2, 1,3;4-b-D-glucanase, and superoxide dismutase (SOD), were specifically regulated by the heat treat- ment, and four genes, including fatty acid desaturase2 (FAD2) and lipid transfer protein (LTP), were specifi- cally regulated by the conditioning treatment. Further- more, four more genes were identified, including a translation initiation factor (SUI1), a chaperonin, and alcohol dehydrogenase (ADH), that were commonly regulated by both heat and conditioning treatments. According to these data, it is suggested that pre-storage heat and conditioning treatments may enhance fruit chilling tolerance by activating different molecular mechanisms. The hot water treatment activates mainly the expression of various stress-related genes, whereas the conditioning treatment activates mainly the ex- pression of lipid membrane modification enzymes. Key words: Chilling, conditioning, grapefruit, hot water, post- harvest, stress. Introduction Many plant species, especially those of tropical and sub- tropical origin, are severely injured or killed by exposure to low non-freezing temperatures (Lyons, 1973; Lynch, * To whom correspondence should be addressed. E-mail: [email protected] Abbreviations: ADH, alcohol dehydrogenase; ASP, ascorbate peroxidase; CAT, catalase; CI, chilling injury; DIP, dehydration-induced protein; EST, expressed sequence tag; FAD, fatty acid desaturase; HSP, heat shock protein; HWB, hot water rinsing and brushing; LTP, lipid transfer protein; SOD, superoxide dismutase; USP, universal stress protein Journal of Experimental Botany, Vol. 57, No. 12, pp. 2943–2953, 2006 doi:10.1093/jxb/erl055 Advance Access publication 14 August, 2006 ª The Author [2006]. Published by Oxford University Press [on behalf of the Society for Experimental Biology]. All rights reserved. For Permissions, please e-mail: [email protected] by guest on December 1, 2016 http://jxb.oxfordjournals.org/ Downloaded from
Transcript of Postharvest heat and conditioning treatments activate different molecular responses and reduce...

RESEARCH PAPER
Postharvest heat and conditioning treatments activatedifferent molecular responses and reduce chillinginjuries in grapefruit
Margarita Sapitnitskaya1, Pilar Maul2, Gregory T. McCollum2, Charles L. Guy3, Batia Weiss1,
Alon Samach4 and Ron Porat1,*
1 Department of Postharvest Science, ARO, the Volcani Center, PO Box 6, Bet Dagan 50250, Israel2 USDA, USHRL, 2001 S. Rock Rd, Ft Pierce, FL 34945, USA3 Department of Environmental Horticulture, University of Florida, Gainesville, FL 32611, USA4 Institute of Plant Sciences and Genetics, Faculty of Agricultural, Food and Environmental Quality Sciences,the Hebrew University of Jerusalem, PO Box 12, Rehovot 76100, Israel
Received 5 April 2006; Accepted 15 May 2006
Abstract
A combination of hot water (a rinse at 62 8C for 20 s)
and conditioning (pre-storage at 16 8C for 7 d) treat-
ments synergistically reduced chilling injury develop-
ment in grapefruit (Citrus paradisi, cv. ‘Star Ruby’)
during cold storage at 2 8C, suggesting that the
treatments may activate different chilling tolerance
responses. To study the molecular mechanisms in-
volved, chilling- and conditioning-responsive genes
were isolated by polymerase chain reaction (PCR)
cDNA subtraction, cDNA libraries were constructed
from hot water- and conditioning-treated fruit, and
cDNA sequencing was used to identify putative stress-
responsive and chilling tolerance genes. PCR cDNA
subtraction revealed the identification of 17 chilling-
responsive and heat- and conditioning-induced genes,
and the expression patterns of 11 additional stress-
related genes, antioxidant defensive genes, and genes
encoding enzymes involved in membrane lipid mod-
ifications were characterized. It was found that hot
water and conditioning treatments had little effect on
gene expression by themselves, but rather had a
priming effect, and enabled the fruit to activate their
defence responses after subsequent exposure to
chilling. RNA gel blot hybridizations revealed that the
expression patterns of eight genes, including HSP19-I,
HSP19-II, dehydrin, universal stress protein (USP),
EIN2, 1,3;4-b-D-glucanase, and superoxide dismutase
(SOD), were specifically regulated by the heat treat-
ment, and four genes, including fatty acid desaturase2
(FAD2) and lipid transfer protein (LTP), were specifi-
cally regulated by the conditioning treatment. Further-
more, four more genes were identified, including a
translation initiation factor (SUI1), a chaperonin, and
alcohol dehydrogenase (ADH), that were commonly
regulated by both heat and conditioning treatments.
According to these data, it is suggested that pre-storage
heat and conditioning treatments may enhance fruit
chilling tolerance by activating different molecular
mechanisms. The hot water treatment activates mainly
the expression of various stress-related genes, whereas
the conditioning treatment activates mainly the ex-
pression of lipid membrane modification enzymes.
Key words: Chilling, conditioning, grapefruit, hot water, post-
harvest, stress.
Introduction
Many plant species, especially those of tropical and sub-tropical origin, are severely injured or killed by exposureto low non-freezing temperatures (Lyons, 1973; Lynch,
* To whom correspondence should be addressed. E-mail: [email protected]: ADH, alcohol dehydrogenase; ASP, ascorbate peroxidase; CAT, catalase; CI, chilling injury; DIP, dehydration-induced protein; EST,expressed sequence tag; FAD, fatty acid desaturase; HSP, heat shock protein; HWB, hot water rinsing and brushing; LTP, lipid transfer protein; SOD,superoxide dismutase; USP, universal stress protein
Journal of Experimental Botany, Vol. 57, No. 12, pp. 2943–2953, 2006
doi:10.1093/jxb/erl055 Advance Access publication 14 August, 2006
ª The Author [2006]. Published by Oxford University Press [on behalf of the Society for Experimental Biology]. All rights reserved.For Permissions, please e-mail: [email protected]
by guest on Decem
ber 1, 2016http://jxb.oxfordjournals.org/
Dow
nloaded from

1990). Exposure of these chilling-sensitive plants to lowgrowth temperatures often results in numerous cellularand metabolic dysfunctions, such as altered respirationrates, impaired photosynthetic activity, and changes in mem-brane permeability (Lyons, 1973; Graham and Patterson,1982; Allen and Ort, 2001).
In citrus, as with many other horticultural commodities,chilling sensitivity also imposes a major limitation onthe postharvest handling of the fruits, since, for somecultivars, such as grapefruit, it necessitates storage at re-latively high temperatures of 11–13 8C (Kader and Arpaia,2002). Exposure of grapefruit to lower temperatures of0–8 8C results in the development of chilling injuries (CIs),which appear as dark sunken lesions (pitting) of collapsedtissues throughout the peel surface (Porat, 2004) (Fig. 1).In previous studies, it was found that pre-storage heatand temperature conditioning treatments enhanced chillingtolerance in citrus fruits and reduced the developmentof CI during postharvest storage (Porat et al., 2000a).The recommended low temperature conditioning treatmentto reduce CI in citrus fruit includes exposure to 16 8Cfor 7 d before transfer to low temperatures (Hatton andCubbedge, 1982, 1983). In addition, it was previouslyshown that a short pre-storage hot water rinsing andbrushing (HWB) treatment at 62 8C for 20 s also effectivelyreduced CI development (Porat et al., 2000a, b).
As compared with plant responses to other abioticstresses, such as freezing, drought, salinity, and heat, onlylittle is known as yet regarding the molecular basis ofchilling tolerance, or of the signal transduction networks
involved in its acquisition. Nevertheless, it was previouslyreported that activation of genes encoding enzymes in-volved in membrane lipid modifications, antioxidant de-fensive genes, and possibly other known stress-responsivegenes may be involved in the enhancement of chillingtolerance. For example, Arabidopsis plants mutated invarious fatty acid desaturase (FAD) genes (fad2, fad5,and fad6) became chilling sensitive (Somerville andBrowse, 1991; Miquel et al., 1993), whereas overexpres-sion of FAD genes, or of other genes, such as acetyltrans-ferases and phospholipid translocases, which increasemembrane fluidity, correspondingly increase chilling toler-ance (Murata et al., 1992; Ishizaki-Nishizawa et al.,1996; Gomez et al., 2000). In addition, it was demon-strated that overexpression of antioxidant defensive genes,such as ascorbate peroxidase (APX), superoxide dismut-ase (SOD), and catalase (CAT), increased chilling toler-ance (Van Breusegem et al., 1999; Payton et al., 2001),whereas repression of CAT gene expression reduced it(Kerdnaimongkol and Woodson, 1999). Finally, severalreports have suggested that various stress genes, usuallyrelated to other types of stress responses, may also beinvolved in the acquisition of chilling tolerance. For in-stance, it was suggested that particular heat shock pro-teins (HSPs) (Sabehat et al., 1998; Sung et al., 2001) anddehydrin genes (Ismail et al., 1999) may also be involvedin conferring chilling tolerance on plants.
In previous studies, it was shown that application ofthe pre-storage HWB treatment at 62 8C for 20 s, whicheffectively increased chilling tolerance, also induced theexpression of various stress-related genes in grapefruitflavedo tissue (the outer coloured layer of the peel). Forexample, it was found that HWB followed by storage at2 8C increased the expression of two dehydrin genes(COR15 and cpDHN), four HSPs (HSP18-I, HSP18-II,HSP22, and HSP70), and cNHX1, a sodium proton anti-port gene known to be involved in salt tolerance in plants(Porat et al., 2002a, b, 2004; Rozenzvieg et al., 2004).Furthermore, other studies showed that a high tempera-ture heat conditioning treatment at 37 8C for 3 d signifi-cantly increased chilling tolerance in ‘Fortune’ mandarinsand correspondingly induced the expression of the WRKYand TFIIB transcription factors and of several stressgenes, such as chitinases and cell wall hydrolysis enzymes(Sanchez-Ballesta et al., 2003).
In the present study, it was found that postharvest lowtemperature conditioning and heat treatments activatedapparent different chilling tolerance-responsive pathwaysin grapefruit and synergistically prevented the develop-ment of CI. Furthermore, by using two different molecularapproaches—construction of polymerase chain reaction(PCR) cDNA subtraction libraries and conduction of agrapefruit flavedo expressed sequence tag (EST) sequenc-ing project—new chilling-responsive and HWB- andconditioning-induced genes were identified that are linked
Fig. 1. CI damage (pitting) in grapefruit. The chilling damage ismanifested as dark sunken areas of collapsed tissue (pitting) all overthe peel surface.
2944 Sapitnitskaya et al.
by guest on Decem
ber 1, 2016http://jxb.oxfordjournals.org/
Dow
nloaded from

with the acquisition of chilling tolerance in citrus. In all, itwas shown that postharvest heat and conditioningtreatments activate different molecular mechanisms,which suggests that several different defensive pathwaysagainst low temperature stress may be involved in theacquisition of chilling tolerance in fruits.
Materials and methods
Plant material and CI evaluation
Grapefruits (Citrus paradisi, cv. ‘Star Ruby’) were obtained fromcommercial orchards and used on the day after harvest. Afterwards,the fruits were stored at 2 8C for 8 or 14 weeks under a relativehumidity of ;90%. Each treatment included four boxes, eachcontaining 30 fruit (total of 120 fruit per treatment). The experimentswere performed twice, during two consecutive seasons. For CIevaluations, fruit were sorted into four categories according to theirCI severity: none (score 0; no pitting), slight (score 1; a few scatteredpits), moderate (score 2; pitting covering up to 30% of the fruitsurface), and severe (score 3; extensive pitting covering >30% of thefruit surface). The CI index was determined for each treatment bymultiplying the number of fruits in each category by their score, andthen dividing this sum by the total number of fruits assessed.
Postharvest HWB and conditioning treatments
Hot water treatments at 62 8C for 20 s were applied by rinsing the fruitas they moved along a set of brush rollers, as described previously(Porat et al., 2000a, b). The temperature conditioning treatment wasapplied by keeping the fruit at 16 8C in a separate storage room for7 d, and then transferring them to continuous storage at 2 8C (Hattonand Cubbedge, 1982, 1983; Porat et al., 2000a).
Isolation of RNA
Total RNA was isolated from grapefruit flavedo by phenol/chloro-form extraction and precipitation with LiCl according to standardprocedures (Sambrook et al., 1992). Poly(A)+ RNA was isolatedfrom total RNA by using the PolyATract mRNA Isolation SystemsKit (Promega, Madison, WI, USA).
Construction of cDNA libraries and sequence analysis
HWB- and conditioning-induced grapefruit flavedo cDNA librarieswere constructed from 5 lg of poly(A)+ RNA isolated 24 h afterHWB or immediately following the conditioning treatment (7 d at16 8C). The cDNA libraries were constructed with the ZAP-cDNA synthesis kit (Strategene, La Jolla, CA, USA) according tothe manufacturer’s instructions.
Sequencing was performed by the dideoxynucleotide chaintermination method (Sanger et al., 1977), and sequence analysiswas done with the BLAST computer programs (Altschul et al., 1997).
PCR cDNA subtraction analysis
PCR cDNA subtraction was performed with the CLONTECHPCR-Select� cDNA Subtraction Kit (Clontech, Palo Alto, CA,USA) according to the manufacturer’s instructions. In general, thesubtraction analysis was performed twice with cDNA synthesizedfrom 2 lg of poly(A)+ RNA isolated from either control (driver)or treated (tester) fruits immediately after the conditioning treatment(7 d at 16 8C) and after two more weeks in cold storage (7 d at 168C+2 weeks at 2 8C). The final subtracted cDNA fragments werecloned into the pGEM-T Easy vector (Promega, Madison, WI, USA).
RNA gel blot analysis
Total RNA (10 lg per lane) was separated on a 1% formaldehyde geland blotted onto a Hybond-N+ membrane (Amersham PharmaciaBiotech, Little Chalfont, Bucks, UK). Afterwards, blots were hy-bridized with the various cDNA probes, labelled to high specificactivity by random priming (Biological Industries Ltd, Israel) with50 mCi of [32P]dCTP at 42 8C in UltraHyb buffer solution (AmbionInc., Austin, TX, USA). After a 16 h hybridization period, theblots were washed three times for 20 min at 63 8C with 0.53 SSC and0.1% SDS, and exposed to an autoradiography film at –80 8Cfor 1–2 d.
Results
Heat and conditioning treatments synergisticallyreduce CI
CI is a major postharvest physiological disorder in citrusfruit, and causes considerable economic losses. It is mani-fested as dark sunken areas of collapsed tissue (pitting) allover the peel surface (Fig. 1). In the present study, it wasfound that HWB and conditioning treatments applied sepa-rately effectively reduced the development and severity ofCI by 85% and 83%, respectively, after 8 weeks of stor-age at 2 8C. After a much longer period of 14 weeks at2 8C, the CI index was reduced by 64% and 56%, re-spectively, (Fig. 2). However, it was found that the com-bined application of the two treatments was much moreeffective than the application of either treatment alone, andsignificantly reduced the development of CI by 98% and93% after 8 weeks and 14 weeks, respectively (Fig. 2).
0.0
0.5
1.0
1.5
2.0
2.5
3.0
Ch
illin
g in
ju
ry in
dex (0-3)
0.0
0.5
1.0
1.5
2.0
2.5
3.0
Ch
illin
g in
ju
ry in
dex (0-3)
Control Conditioning HWB HWB +Conditioning
A After 8 weeks at 2°C
B After 14 weeks at 2°C
Fig. 2. Effects of HWB and conditioning treatments on the developmentof CI in grapefruit. The CI percentages were measured after 8 weeks (A)and 14 weeks (B) of storage at 2 8C. Data are means of four replicationsper treatment, each containing 30 fruits (total of 120 fruit per treatment).
Heat and conditioning synergistically prevent chilling injuries 2945
by guest on Decem
ber 1, 2016http://jxb.oxfordjournals.org/
Dow
nloaded from

Isolation of chilling-, heat-, and conditioning-responsivegenes by PCR cDNA subtraction analysis
In order to identify genes that might be involved in theacquisition of chilling tolerance of grapefruit, two PCRcDNA subtraction libraries were constructed with RNAisolated from the peel of control fruits at time zero (drivercDNA) and from the peel of conditioned fruits at twodifferent time points: immediately after conditioning(7 d at 16 8C) or after two additional weeks in cold storage(7 d at 16 8C+2 weeks at 2 8C) (tester cDNAs). Sequenc-ing, nucleotide BLAST searches, and confirmation by RNAgel blot hybridization analysis of ;70 randomly chosenPCR cDNA fragments from each subtracted cDNA libraryrevealed the identities of 17 cDNAs that were specific-ally induced either by chilling itself (exposure to 2 8C) orfollowing application of the hot water or conditioningtreatments (Table 1).
Only one clone (S8, similar to an unknown proteinfrom tomato) was isolated from the first subtracted library,which was constructed immediately after the conditioningtreatment, whereas all the other cDNAs were isolated fromthe second cDNA subtracted library, which was con-structed after two additional weeks of exposure to 2 8C.Among these identified cDNAs, seven clones (S18, S26,S28, S41, S49, S78, and S80) were homologous to othergenes with known functions, whereas the other 10 wereeither novel or similar to other genes with unknown func-tions (Table 1).
Isolation of grapefruit stress-related, antioxidantdefence and lipid modification genes by ESTsequencing
In an effort to identify, in grapefruit, stress-responsive genes,antioxidant defensive genes, and genes encoding enzymesinvolved in membrane lipid modifications that may beinfluenced by the HWB and conditioning treatments, two
grapefruit flavedo cDNA libraries were constructed, thefirst 24 h after application of the HWB treatment, and thesecond immediately after the conditioning treatment (7 d at16 8C), and an EST sequencing project was initiated atthe US Horticulture Research Laboratory Genomic Centerat Ft Pierce, FL, USA. After ;1550 randomly chosen, dif-ferent cDNAs from the HWB-treated cDNA library and;550 cDNAs from the conditioned cDNA library had beensequenced, the expression patterns of 11 different cDNAswere evaluated following exposure of the fruits to the heatand conditioning treatments and subsequent cold storage(Table 2). These chosen cDNAs comprised six stress-responsive genes—alcohol dehydrogenase (ADH) (anaer-obic stress), dehydrin, and a dehydration-induced protein(DIP) (drought stress), HSP19-I and HSP19-II (heat stress),and universal stress protein (USP) (general stress respon-sive)—three antioxidant defensive genes—APX, CAT, andSOD—and two genes encoding the lipid modificationenzymes—FAD2 and lipid transfer protein (LTP) (Table 2).
Characterization of chilling-, heat-, and conditioning-responsive genes by RNA gel blot hybridizations
In order to characterize the possible involvement of thevarious isolated genes in heat- and conditioning-inducedchilling tolerance responses in grapefruit, their expressionpatterns were evaluated in response to the application of eachtreatment alone, without subsequent cold storage. It can beseen that among the 28 cDNAs tested, expression patternsof only 13 cDNAs were affected by the applications of theHWB or the conditioning treatment by themselves (Fig. 3).Among these, the expression patterns of three genes wereinduced by either treatment (S34, Al-induced protein, andS70), the expression of five was specifically induced byHWB (HSP19-I, HSP19-II, TRX, S50, and S84), and twowere specifically induced by the conditioning treatment(S8 and FAD2) (Fig. 3). Furthermore, it was found that the
Table 1. List of chilling-inducible, HWB-responsive, and conditioning-responsive grapefruit flavedo cDNAs identified by PCR cDNAsubtraction analysis
Clone name GenBank accession number Homology to known genes E-value
S4 DQ056320 Hypothetical protein, A. thaliana (BAB10956.1) 9e-17S8 DQ056321 Clone 132586R, Lycopersicon esculentum (BT013727) 4e-04S18 AY271308 Thioredoxin h, C. paradisi 0.0S26 DQ056322 Lipase class 3-like, Oryza sativa (BAD35707.1) 8e-27S28 DQ056323 SUI1 protein, Coffea arabica (CAD58629.1) 6e-33S34 DQ056324 Unknown proteinS41 DQ056325 Chaperonin, putative, A. thaliana (NP200461.2) 4e-38S43 DQ056326 Unknown proteinS44 DQ056327 Unknown proteinS49 DQ056328 Al-induced protein, Gossypium hirsutum (AAQ74889.1) 4e-45S50 DQ056329 Unknown proteinS59 DQ056330 Unknown proteinS62 DQ056331 Unknown proteinS70 DQ056333 Unknown proteinS78 DQ056334 EIN2, Petunia3hybrida (AAR08678.1) 2e-29S80 DQ056335 Putative endo-1,3;4-b-D-glucanase, Oryza sativa (AAU10802.1) 1e-55S84 DQ056336 Unknown protein
2946 Sapitnitskaya et al.
by guest on Decem
ber 1, 2016http://jxb.oxfordjournals.org/
Dow
nloaded from

expression patterns of two genes were specifically down-regulated by HWB (S8 and DIP), and those of anothertwo were specifically down-regulated following the condi-tioning treatment (EIN2 and dehydrin) (Fig. 3).
Following exposure of the fruit to low temperature(storage at 2 8C), it was found that the expression patternsof nine genes were affected by exposure to chilling itself,no matter whether they had received the pre-storage hotwater or conditioning treatments that enhance chillingtolerance in fruits (Fig. 4). Among these, the expressionpatterns of seven genes were up-regulated by chilling(TRX, lipase, CAT, S4, S43, S50, and S62), and two weredown-regulated by chilling (S44 and DIP) (Fig. 4). Thesenine chilling-responsive genes are probably part of thefruit’s basal low-temperature defensive system, but not partof the HWB- or conditioning-induced chilling tolerancepathways.
In contrast to these general chilling-responsive genes,several genes were identified that were specifically regu-lated by cold storage that followed the application of theHWB or the conditioning treatment, but not by cold storagealone (Fig. 5). Among these, eight genes were identifiedthat were specifically regulated by HWB followed by coldstorage (Fig. 5A), four by conditioning followed by coldstorage (Fig. 5B), four regulated by either HWB or byconditioning, followed by cold storage (Fig. 5C), and twospecifically regulated by the combination of HWB andconditioning, followed by cold storage (Fig. 5D). TheHWB-specific genes included up-regulation of five genes(dehydrin, HSP19-I, HSP19-II, USP, and S84) and down-regulation of three (EIN2, 1,3;4-b-D-glucanase, and SOD)(Fig. 5A); the conditioning-specific genes included up-regulation of four genes (S8, S59, FAD2, and LTP) (Fig.5B); the common HWB- and conditioning-regulated genesincluded up-regulation of four genes (translation initiationfactor SUI1, S34, chaperonin, and ADH); and the combinedHWB and conditioning-regulated genes included up-regu-lation of two genes (Al-induced protein and S70) (Fig. 5D).
Interestingly, it was found that the induction of mostgenes that were specifically induced by either the HWBor conditioning treatment during cold storage required
Table 2. List of stress-responsive genes, antioxidant defensive genes, and genes encoding enzymes involved in membrane lipidmetabolism that were identified following the initiation of grapefruit flavedo EST sequencing projects from HWB- and conditioning-induced cDNA libraries
Clone name GenBank accession number Homology to known genes E-value
Stress-related cDNAsADH DN957723 Class III ADH, glutathione-dependent formaldehyde
dehydrogenase, A. thaliana (CAA57973)0.0
Dehydrin AY160772 Dehydrin, C. paradisi (AY160772) 0.0DIP DN957704 Dehydration-induced protein, Gossypium hirsutum (AAL67991) e-122HSP19-I AY261672 Heat shock protein 19 class I, C. paradisi (AY261672) 0.0HSP19-II AY261671 Heat shock protein 19 class II, C. paradisi (AY261671) 0.0USP DN796887 Universal stress protein, A. thaliana (NP567770) 8e-10Antioxidant defensive cDNAsAPX DN797057 Ascorbate peroxidase, I. batatas (AAP42501.1) e-116CAT DN797949 Catalase, Prunus persica (CAB56850.1) e-149SOD DN796979 Superoxide dismutase, Para rubber tree (S39492) e-105Lipid modification cDNAsFAD2 DN958297 Delta-12 fatty acid desaturase, Tropaeolum majus (AY780572) 5e-54LTP DN957734 Lipid transfer protein, C. sinensis (AAM21292.1) 5e-32
Fig. 3. Effects of HWB and conditioning treatments on the mRNAlevels of putative chilling tolerance genes in grapefruit flavedo. Each lanecontained 10 lg of total RNA isolated 0, 6, 12, and 24 h after HWB at20 8C or 0, 1, 2, 3, 4, 5, 6, or 7 d into the conditioning treatment at 16 8C.Ethidium bromide staining of the RNA gel was used to ensure equalloading among lanes.
Heat and conditioning synergistically prevent chilling injuries 2947
by guest on Decem
ber 1, 2016http://jxb.oxfordjournals.org/
Dow
nloaded from

further exposure to chilling, but these genes were not in-duced by the application of either treatment by itselfwithout subsequent cold storage (Table 3). For example,among the eight different HWB-regulated genes, onlyHSP19-I and HSP19-II were induced by exposure to hotwater alone, and, among the four conditioning-regulatedgenes, only S8 and FAD2 were affected by exposure tothe conditioning treatment alone (Figs 3, 5; Table 3).Among the HWB and conditioning commonly regulatedgenes found, only clone S34 was induced by exposure toeither treatment alone without subsequent cold storage(Figs 3, 5; Table 3). Thus, the HWB and condition-ing treatments mainly had a ‘priming’ effect, and enabledthe fruits to activate most of their defence responses afterthe subsequent exposure to chilling.
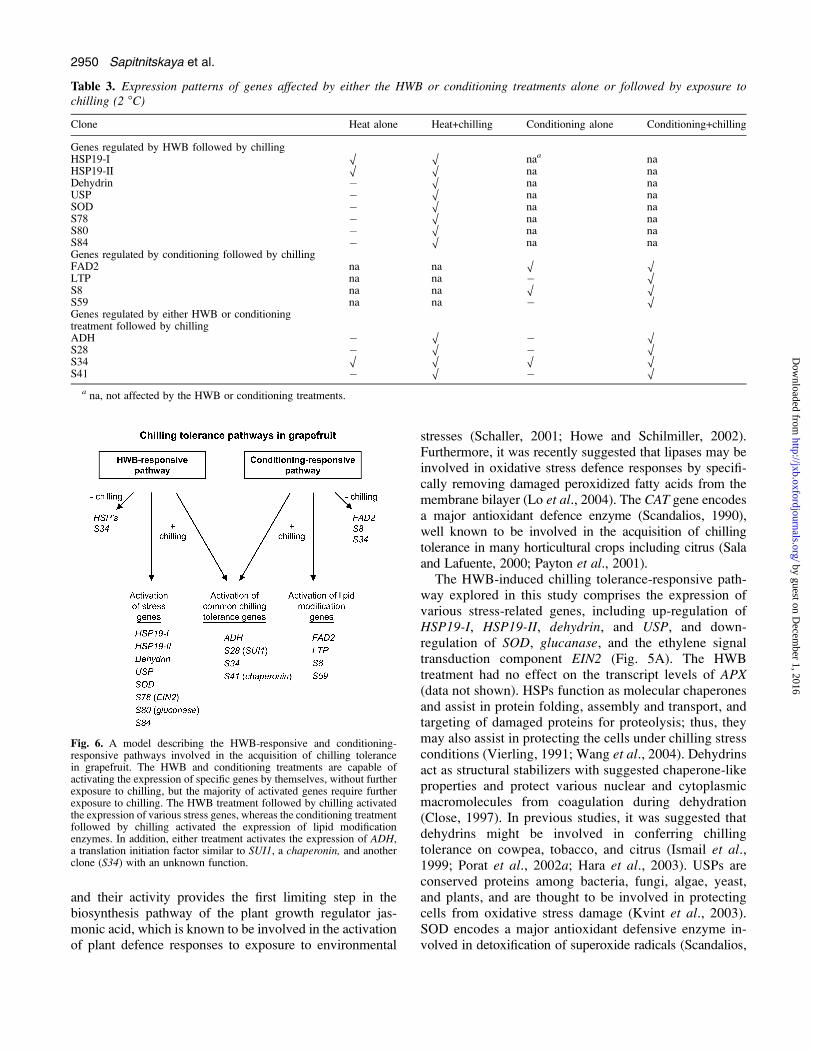
Overall, these RNA hybridization studies revealed thatpre-storage hot water and conditioning treatments specifi-cally activated different sets of genes in grapefruit peeltissue during cold storage (Fig. 6). Moreover, it can be seenthat the HWB treatment mainly effected the expression ofvarious stress-related genes (such as HSP19-I, HSP19-II,dehydrin, USP, SOD, EIN2, and glucanase), whereas theconditioning treatment mainly activated the expressionof genes encoding enzymes involved in membrane lipidmodifications (such as FAD2 and LTP) (Fig. 6). In additionto heat-specific and conditioning-specific gene expres-sion pathways, both treatments also commonly activatedanother set of genes (including SUI1, ADH, and chaperonin)
which may have a common role in conferring fruitchilling tolerance (Fig. 6).
Discussion
In this study, it was found that the combined applicationof HWB and conditioning treatments was much moreeffective in reducing the development of CI in grapefruitthan the application of each treatment alone, which sug-gests that the two treatments activated different chilling-stress defensive pathways (Fig. 1). This finding appears tobe a novel observation that a synergistic effect betweentwo different treatments can increase chilling tolerance.
Overall, nine chilling-responsive genes were identifiedin grapefruit, whose expression patterns were affected byexposure to chilling, independently of the treatments thatthe fruits had received after harvest (Fig. 4). These chilling-responsive genes included four known genes, TRX, lipase,CAT, and DIP, and five genes of unknown function (Fig. 4).It is suggested that these chilling-responsive genes may bepart of a basal low temperature defensive pathway of thefruits, and are probably involved in their natural attemptto cope with exposure to chilling. Among these genes,TRX participates in redox regulation of selected target pro-teins, including oxidative defensive enzymes, such asperoxiredoxins (Schurmann and Jacquot, 2000). Lipasescatalyse the hydrolysis of fatty acids from phospholipids,
Fig. 4. Effects of chilling, HWB, and conditioning treatments on the mRNA levels of putative chilling tolerance genes in grapefruit flavedo. Each lanecontained 10 lg of total RNA isolated after 0, 1, 2, 3, 4, 6, or 9 weeks of exposure to 2 8C. Ethidium bromide staining of the RNA gel was used to ensureequal loading among lanes.
2948 Sapitnitskaya et al.
by guest on Decem
ber 1, 2016http://jxb.oxfordjournals.org/
Dow
nloaded from

Fig. 5. Effects of chilling, HWB, and conditioning treatments on the mRNA levels of putative chilling tolerance genes in grapefruit flavedo. Each lanecontained 10 lg of total RNA isolated after 0, 1, 2, 3, 4, 6, or 9 weeks of exposure to 2 8C. The various genes were classified into different groupsaccording to their expression patterns: (A), genes specifically regulated by HWB; (B), genes specifically regulated by conditioning; (C), genes regulatedby either HWB or conditioning, (D), genes specifically regulated by the combination of HWB and conditioning. Ethidium bromide staining of the RNAgel was used to ensure equal loading among lanes.
Heat and conditioning synergistically prevent chilling injuries 2949
by guest on Decem
ber 1, 2016http://jxb.oxfordjournals.org/
Dow
nloaded from
and their activity provides the first limiting step in thebiosynthesis pathway of the plant growth regulator jas-monic acid, which is known to be involved in the activationof plant defence responses to exposure to environmental
stresses (Schaller, 2001; Howe and Schilmiller, 2002).Furthermore, it was recently suggested that lipases may beinvolved in oxidative stress defence responses by specifi-cally removing damaged peroxidized fatty acids from themembrane bilayer (Lo et al., 2004). The CAT gene encodesa major antioxidant defence enzyme (Scandalios, 1990),well known to be involved in the acquisition of chillingtolerance in many horticultural crops including citrus (Salaand Lafuente, 2000; Payton et al., 2001).
The HWB-induced chilling tolerance-responsive path-way explored in this study comprises the expression ofvarious stress-related genes, including up-regulation ofHSP19-I, HSP19-II, dehydrin, and USP, and down-regulation of SOD, glucanase, and the ethylene signaltransduction component EIN2 (Fig. 5A). The HWBtreatment had no effect on the transcript levels of APX(data not shown). HSPs function as molecular chaperonesand assist in protein folding, assembly and transport, andtargeting of damaged proteins for proteolysis; thus, theymay also assist in protecting the cells under chilling stressconditions (Vierling, 1991; Wang et al., 2004). Dehydrinsact as structural stabilizers with suggested chaperone-likeproperties and protect various nuclear and cytoplasmicmacromolecules from coagulation during dehydration(Close, 1997). In previous studies, it was suggested thatdehydrins might be involved in conferring chillingtolerance on cowpea, tobacco, and citrus (Ismail et al.,1999; Porat et al., 2002a; Hara et al., 2003). USPs areconserved proteins among bacteria, fungi, algae, yeast,and plants, and are thought to be involved in protectingcells from oxidative stress damage (Kvint et al., 2003).SOD encodes a major antioxidant defensive enzyme in-volved in detoxification of superoxide radicals (Scandalios,
Table 3. Expression patterns of genes affected by either the HWB or conditioning treatments alone or followed by exposure tochilling (2 8C)
Clone Heat alone Heat+chilling Conditioning alone Conditioning+chilling
Genes regulated by HWB followed by chillingHSP19-I O O naa naHSP19-II O O na naDehydrin � O na naUSP � O na naSOD � O na naS78 � O na naS80 � O na naS84 � O na naGenes regulated by conditioning followed by chillingFAD2 na na O OLTP na na � OS8 na na O OS59 na na � OGenes regulated by either HWB or conditioningtreatment followed by chillingADH � O � OS28 � O � OS34 O O O OS41 � O � O
a na, not affected by the HWB or conditioning treatments.
Fig. 6. A model describing the HWB-responsive and conditioning-responsive pathways involved in the acquisition of chilling tolerancein grapefruit. The HWB and conditioning treatments are capable ofactivating the expression of specific genes by themselves, without furtherexposure to chilling, but the majority of activated genes require furtherexposure to chilling. The HWB treatment followed by chilling activatedthe expression of various stress genes, whereas the conditioning treatmentfollowed by chilling activated the expression of lipid modificationenzymes. In addition, either treatment activates the expression of ADH,a translation initiation factor similar to SUI1, a chaperonin, and anotherclone (S34) with an unknown function.
2950 Sapitnitskaya et al.
by guest on Decem
ber 1, 2016http://jxb.oxfordjournals.org/
Dow
nloaded from

1990; Van Breusegem et al., 1999), and glucanases areinvolved in cell wall softening and pathogen defenceresponses (Kauffmann et al., 1987; Brummel et al., 2004).In many horticultural commodities including citrus,ethylene enhances the development of CI during lowtemperature storage (Lafuente et al., 2001; Porat, 2004).Interestingly, in the present study, it was found that HWBdown-regulated the expression of the EIN2 gene, a trans-ducer of the ethylene response (Alonso et al., 1999), and,thus, may have inhibited the development of ethylene-promoted CI.
The identified conditioning-induced chilling tolerance-responsive pathway comprises the expression of FAD2and LTP, which encode enzymes involved in lipid metab-olism. The FAD2 gene encodes a microsomal 18:1 de-saturase that is required for survival of Arabidopsis plantsunder chilling temperatures (Miquel et al., 1993). Thepossible involvement of the FAD2 gene in conditioning-induced chilling tolerance responses is further supportedby other reports that conditioning treatments increasedfatty acid desaturation and chilling tolerance in tobaccoand grapefruit (Nordby et al., 1987; Kodama et al.,1995). LTPs are involved in membrane biogenesis andthe transport of phospholipids (Somerville and Browse,1991; Kader, 1996). In addition, LTPs are induced in plantcells upon exposure to biotic and abiotic stresses; insome cases they were reported to exhibit antifreeze activityand, thus, were hypothesized to be involved in conferr-ing frost tolerance (Hon et al., 1995; Wu et al., 2004).Hereby, it is suggested that LTPs may also participatein conditioning-induced chilling tolerance responses ingrapefruit.
The group of genes induced by chilling that followseither the HWB or the conditioning treatment, but notby chilling alone most probably encode major chillingtolerance proteins required to protect the fruit upon expo-sure to low temperatures. These genes include S28, whichencodes a translation initiation factor similar to SUI1. Itwas reported that besides initiating transcription, theSUI1 protein may also be involved in repairing impairedmRNAs (Cui et al., 1999). In sum, it is possible thatthe SUI1 protein may be required to govern translationunder low temperature conditions. The HWB and con-ditioning treatments also increased the expression of achaperonin gene; these, like HSPs, are required to maintainprotein structure and function under stress conditions(Yamada et al., 2002; Wang et al., 2004). Finally, itwas found that both treatments increased the expres-sion of a specific glutathione-dependent formaldehydedehydrogenase (ADH, class III), known to be involved indetoxification of aldehydes (Martinez et al., 1996). Fur-ther support for the hypothesis that ADH activity may berequired for chilling tolerance comes from a recent studyof maize, which demonstrated that plants containingdouble-null mutations in adh1 and adh2 developed more
CI than wild-type plants, following exposure to 2 8C(Peters and Frankel, 2004).
Overall, a general model illustrating the various HWB-responsive and conditioning-responsive chilling tolerancepathways in grapefruit peel tissue is presented in Fig. 6.It can be seen that HWB and conditioning treatmentsactivated several stress defensive genes by themselves(HSP19-I, II, FAD2, S8, and S34), but the majority of theHWB- and conditioning-regulated genes were induced onlyfollowing subsequent exposure to chilling. Furthermore,the HWB chilling tolerance-responsive pathway activatedmainly expression of stress-related genes, whereas theconditioning chilling tolerance-responsive pathway acti-vated expression of genes involved in fatty acid lipidmetabolism (Fig. 6). It should be noted that in most casesthe combined application of both HWB and conditioningtreatments had similar effects on gene expression levels ascompared with each treatment alone (Fig. 5A–C). How-ever, the transcript levels of two clones, S49 and S70, werespecifically induced by the combined application of bothtreatments but not by each one of them alone (Fig. 5D).These observations may explain in part why the combinedapplication of HWB and conditioning together was moreeffective in reducing CI as compared with each treatmentalone (Fig. 2).
Finally, an important observation from the geneexpression studies is that the pre-storage hot water andconditioning treatments had only minor effects on geneexpression patterns by themselves, but rather had ‘poten-tiation’ or ‘priming’ effects, and enabled the fruit to acti-vate most of its defence responses after subsequentexposure to chilling (Table 3). Similar priming effectshave been reported previously regarding the mode ofaction of salicylic acid in promoting the induction ofHsp70/Hsc70 in response to heat in tomato, and inenhancing ion transport following elicitation in parsleycells (Crunje and Bornman, 1999; Katz et al., 2002).Furthermore, it was recently demonstrated that b-amino-butyric acid enhanced pathogen defence and abiotic stresstolerance in Arabidopsis and grapevine by priming theplants’ responses towards exposure to stresses (Hami-duzzaman et al., 2005; Ton et al., 2005). Finally, it hasbeen shown that exposure to short days potentiated lowtemperature-induced gene expression during cold acclim-ation in silver birch (Puhakainen et al., 2004). Thus,potentiation or priming effects may provide an importantmode of action in conferring plant stress tolerance by varioustreatments.
Altogether, in this study, evidence for the existence oftwo different chilling tolerance-responsive pathways ingrapefruit has been uncovered, and it has been shown thatpre-storage HWB and conditioning treatments potentiatethe fruit peel tissue enabling it to respond adequately tochilling by enhanced expression of various low-temperaturedefensive genes (Table 3).
Heat and conditioning synergistically prevent chilling injuries 2951
by guest on Decem
ber 1, 2016http://jxb.oxfordjournals.org/
Dow
nloaded from

Acknowledgements
This research was supported by Research Grant No. IS-3499-03Rfrom BARD, the United States–Israel Binational AgriculturalResearch and Development fund.
References
Allen D, Ort D. 2001. Impacts of chilling temperatures onphotosynthesis in warm-climate plants. Trends in Plant Science6, 36–42.
Alonso J, Hirayama T, Roman G, Nourizadeh S, Ecker J. 1999.EIN2, a bifunctional transducer of ethylene and stress response inArabidopsis. Science 284, 2148–2152.
Altschul SF, Madden TL, Schiffer AA, Zhang J, Zhang Z,Miller W, Lipman DJ. 1997. Gapped BLAST and PSI-BLAST:a new generation of protein database search programs. NucleicAcids Research 25, 3389–3402.
Brummell DA, Dal Cin V, Crisosto CH, Labavitch JM. 2004. Cellwall metabolism during maturation, ripening and senescence ofpeach fruit. Journal of Experimental Botany 55, 2029–2039.
Close TJ. 1997. Dehydrins: a commonality in the response of plantsto dehydration and low temperature. Physiologia Plantarum100, 291–296.
Crunje MJ, Bornman L. 1999. Salicylic acid influences Hsp70/Hsc70 expression in Lycopersicon esculentum: dose- and time-dependent induction or potentiation. Biochemical and BiophysicalResearch Communications 265, 422–427.
Cui Y, Gonzales CI, Kinzy TG, Dinman JG, Peltz SW. 1999.Mutations in the MOF2/SUI1 gene affect both translation andnonsense-mediated mRNA decay. RNA 5, 794–804.
Gomez E, Jakobsen M, Axelsen K, Geisler M, Palmgren. 2000.Chilling tolerance in Arabidopsis involves ALA1, a member ofa new family of putative aminophospholipid translocases. ThePlant Cell 12, 2441–2454.
Graham D, Patterson B. 1982. Responses of plants to lownonfreezing temperatures: proteins, metabolism, and acclimation.Annuals Reviews of Plant Physiology 33, 47–72.
Hamiduzzaman MM, Jakab G, Barnavon L, Neuhaus JM,Mauch-Mani B. 2005. beta-Aminobutyric acid-induced resistanceagainst downy mildew in grapevines acts through potentiation ofcallose formation and jasmonic acid signaling. Molecular Plant–Microbe Interactions 18, 819–829.
HaraM, Terashima S, Fukaya T, Kubboi T. 2003. Enhancement ofcold tolerance and inhibition of lipid peroxidation by citrusdehydrin in transgenic tobacco. Planta 217, 290–298.
Hatton T, Cubbedge R. 1982. Conditioning Florida grapefruit toreduce chilling injury during low-temperature storage. Journal ofthe American Society of Horticultural Science 107, 57–60.
Hatton T, Cubbedge R. 1983. Temperature for prestorage condi-tioning of ‘Marsh’ grapefruit to prevent chilling injury at lowtemperatures. HortScience 18, 721–722.
Hon WC, Griffith M, Mlynarz A, Kwok YC, Chang P, YangDSC. 1995. Antifreeze proteins in winter rye are similar topathogenesis-related proteins. Plant Physiology 109, 879–889.
Howe G, Schilmiller A. 2002. Oxylipin metabolism in response tostress. Current Opinion in Plant Biology 5, 230–236.
Ishizaki-Nishizawa O, Fujii T, Azuma M, Sekiguchi K,Murata N, Ohtani T, Toguri T. 1996. Low temperature re-sistance of higher plants is significantly enhanced by nonspecificcyanobacterial desaturase. Nature Biotechnology 14, 1003–1006.
Ismail AM, Hall AE, Close TJ. 1999. Allelic variation of a dehydringene cosegregates with chilling tolerance during seedling emer-
gence. Proceedings of the National Academy of Sciences, USA96, 13566–13570.
Kader JC. 1996. Lipid-transfer proteins in plants. Annual Reviews ofPlant Molecular Biology 47, 627–654.
Kader AA, Arpaia ML. 2002. Postharvest handling systems:subtropical fruit. In: AA Kader, ed, Postharvest technology ofhorticultural crops. Oakland, CA: Regents of the University ofCalifornia, DNAR, 233–240.
Katz V, Fuchs A, Conrath U. 2002. Pretreatment with salicylic acidprimes parsley cells for enhanced ion transport following elicita-tion. FEBS Letters 520, 53–57.
Kauffmann S, Legrand M, Geoffroy P, Friting B. 1987. Thebiological function of pathogenesis-related proteins: 4 PR proteinsof tobacco have 1,3-beta-glucanase activity. EMBO Journal 6,3209–3212.
Kerdnaimongkol K, Woodson WR. 1999. Inhibition of catalase byantisense RNA increases susceptibility to oxidative stress andchilling injury in tomato plants. Journal of the American Societyof Horticultural Science 124, 330–336.
Kodama H, Hamada T, Horiguchi G, Nishimura G, Iba K. 1995.Fatty acid desaturation during chilling acclimation is one of thefactors involved in conferring low-temperature tolerance to youngtobacco leaves. Plant Physiology 107, 1177–1185.
Kvint K, Nachin L, Diez A, Nystrom T. 2003. The bacterialuniversal stress protein: function and regulation. Current Opinionin Microbiology 6, 140–145.
Lafuente MT, Zacarias L, Martınez-Tellez MA, Sanchez-Ballesta MT, Dupille E. 2001. Phenylalanine ammonia-lyaseas related to ethylene in the development of chilling symptomsduring cold storage of citrus fruits. Journal of Agriculture andFood Chemistry 49, 6020–6025.
Lo M, Taylor C, Wang L, Nowack L, Wang T-W, Thompson J.2004. Characterization of an ultraviolet B-induced lipase inArabidopsis. Plant Physiology 135, 947–958.
Lynch DV. 1990. Chilling injury in plants: the relevance ofmembrane lipids. In: Katterman F, ed. Environmental injury toplants. San Diego, CA: Academic Press, 17–34.
Lyons JM. 1973. Chilling injury in plants. Annual Reviews of PlantPhysiology 24, 445–466.
Martinez M, Achkor H, Persson B, Fernandez M, Shafqat J,Farres J, Jornvall H, Pares X. 1996. Arabidopsis formaldehydedehydrogenase: molecular properties of plant class III alcoholdehydrogenase provide further insights into the origins, structureand function of plant class P and liver class I alcohol dehyd-rogenases. European Journal of Biochemistry 241, 849–857.
Miquel M, James D, Dooner H, Browse J. 1993. Arabidopsisrequires polyunsaturated lipids for low temperature survival.Proceedings of the National Academy of Sciences, USA 90,6208–6212.
Murata N, Ishizaki-Nishizawa O, Higashi S, Hayashi H,Tasaka Y, Nishida I. 1992. Genetically engineered alterationon the chilling sensitivity of plants. Nature 356, 710–713.
Nordby H, Purvis A, Yelenosky G. 1987. Lipids in peel ofgrapefruit and resistance to chilling injury during cold storage.HortScience 22, 915–917.
Payton P, Webb R, Kornyeyev D, Allen R, Holaday S. 2001.Protecting cotton photosynthesis during moderate chilling at highlight intensity by increasing chloroplastic antioxidant enzymeactivity. Journal of Experimental Botany 52, 2345–2354.
Peters J, Frenkel C. 2004. Relationship between alcohol dehyd-rogenase activity and low- temperature in two maize genotypes,Silverado F1 and Adh1– Adh2– doubly null. Plant Physiology andBiochemistry 42, 841–846.
Porat R. 2004. Reduction of chilling injury disorders in citrus fruit.In: Dris R, Niskanen R, eds. Crop management and postharvest
2952 Sapitnitskaya et al.
by guest on Decem
ber 1, 2016http://jxb.oxfordjournals.org/
Dow
nloaded from

handling of horticultural products. Vol. IV. Diseases and disordersof fruits and vegetables. New Delhi, India: Oxford & IBHPublishing Co. Pvt. Ltd., 225–236.
Porat R, Pasentsis K, Rozentzvieg D, Gerasopoulos D, Falara V,Samach A, Lurie S, Kanellis AK. 2004. Isolation of a dehydrincDNA from orange and grapefruit citrus fruit that is specificallyinduced by the combination of heat followed by chilling temper-atures. Physiologia Plantarum 120, 256–264.
Porat R, Pavoncello D, Ben-Hayyim G, Lurie S. 2002b. A heattreatment induced the expression of a Na+/H+ antiport gene(cNHX1) in citrus fruit. Plant Science 162, 957–963.
Porat R, Pavoncello D, Lurie S, McCollum TG. 2002a. Iden-tification of a grapefruit cDNA belonging to a unique class ofcitrus dehydrins and characterization of its expression patternsunder temperature stress conditions. Physiologia Plantarum 115,598–603.
Porat R, Pavoncello D, Peretz J, Ben-Yehoshua S, Lurie S.2000a. Effect of various heat treatments on the postharvestqualities of ‘Star Ruby’ grapefruit. Postharvest Biology andTechnology 18, 159–165.
Porat R, Pavoncello D, Peretz J, Weiss B, Daus A, Cohen L,Ben-Yehoshua S, Droby S, Lurie S. 2000b. Induction of resis-tance to Penicillium digitatum and chilling injury in ‘Star Ruby’grapefruit by a short hot rinse and brushing treatment. Journalof Horticultural Science and Biotechnology 75, 428–432.
Puhakainen T, Li CY, Boije-Malm M, Kangasjarvi J, Heino P,Palva ET. 2004. Short-day potentiation of low temperature-induced gene expression of a C-repeat-binding factor-controlledgene during cold acclimation in silver birch. Plant Physiology 136,4299–4307.
Rozenzvieg D, Elmaci C, Samach A, Lurie S, Porat R. 2004.Isolation of four heat shock protein cDNAs from grapefruit peeltissue and characterization of their expression in response to heatand chilling temperature stresses. Physiologia Plantarum 121,421–428.
Sabehat A, Lurie S, Weiss D. 1998. Expression of small heat shockproteins at low temperature: a possible role in protecting againstchilling injuries. Plant Physiology 117, 651–658.
Sala JM, Lafuente MT. 2000. Catalase enzyme activity related totolerance of mandarin fruits to chilling. Postharvest Biology andTechnology 20, 81–89.
Sambrook J, Fritsch EF, Maniatis T. 1992. Molecular cloning:a laboratory manual. Cold Spring Harbor, NY: Cold SpringHarbor Laboratory Press.
Sanchez-Ballesta MT, Lunch Y, Gosalbes MJ, Zacarias L,Granell A, Lafuente MT. 2003. A survey of genes differentiallyexpressed during long-term heat-induced chilling tolerance incitrus fruit. Planta 218, 65–70.
Sanger F, Nicklen S, Coulson AR. 1977. DNA sequencing withchain terminating inhibitors. Proceedings of the National Academyof Sciences, USA 74, 5463–5467.
Scandalios J. 1990. Response of plant antioxidant defense genes toenvironmental stress. Advances in Genetics 28, 1–41.
Schaller F. 2001. Enzymes of the biosynthesis of octadecanoid-derived signaling molecules. Journal of Experimental Botany 52,11–23.
Schurmann P, Jacquot J-P. 2000. Plant thioredoxin systemsrevisited. Annual Review of Plant Physiology and Plant MolecularBiology 51, 371–400.
Sommerville CR, Browse J. 1991. Plant lipids: metabolism,mutants, and membranes. Science 252, 80–87.
Sung DY, Vierling E, Guy CL. 2001. Comprehensive expressionprofile analysis of the Arabidopsis Hsp70 gene family. PlantPhysiology 126, 789–800.
Ton J, Jakab G, Toquin V, Flors V, Iavicoli A, Maeder MN,Metraux JP, Mauch-Mani B. 2005. Dissecting the beta-aminobutyric acid-induced priming phenomenon in Arabidopsis.The Plant Cell 17, 987–999.
Van Breusegem F, Slooten L, Stassart J, Botterman J, Moens T,Van Montagu M, Inze D. 1999. Effects of overproduction oftobacco MnSOD in maize chloroplasts on foliar tolerance tocold and oxidative stress. Journal of Experimental Botany 50,71–78.
Vierling E. 1991. The roles of heat shock proteins in plants. AnnualReview of Plant Physiology and Plant Molecular Biology 42,579–620.
Wang WX, Vinocur B, Shoseyov O, Altman A. 2004. Roleof plant heat-shock proteins and molecular chaperones inthe abiotic stress response. Trends in Plant Science 9,244–252.
Wu G, Robertson A, Liu X, Zheng P, Wilen R, Nesbitt N,Gusta L. 2004. A lipid transfer protein gene BG-14 is differen-tially regulated by abiotic stress, ABA, anisomycin, and sphigosinein bromegrass (Bromus inermis). Journal of Plant Physiology161, 449–458.
Yamada A, Sekiguchi M, Mimura T, Ozeki Y. 2002. The roleof plant CCT in salt- and osmotic-stress tolerance. Plant and CellPhysiology 43, 1043–1048.
Heat and conditioning synergistically prevent chilling injuries 2953
by guest on Decem
ber 1, 2016http://jxb.oxfordjournals.org/
Dow
nloaded from




















