
Heparanase of murine effector lymphocytes and neutrophils is not required for their diapedesis into...
12
The FASEB Journal • Research Communication Heparanase of murine effector lymphocytes and neutrophils is not required for their diapedesis into sites of inflammation Liat Stoler-Barak,* ,1 Ekaterina Petrovich,* ,1 Tegest Aychek,* Irina Gurevich,* Orna Tal,* Miki Hatzav,* Neta Ilan, † Sara W. Feigelson,* Guy Shakhar,* Israel Vlodavsky, † and Ronen Alon* ,2 *Department of Immunology, The Weizmann Institute of Science, Rehovot, Israel; and † Cancer and Vascular Biology Research Center, The Bruce Rappaport Faculty of Medicine, Technion, Haifa, Israel ABSTRACT Heparanase, the exclusive mammalian heparan sulfate-degrading enzyme, has been suggested to be utilized by leukocytes to penetrate through the dense basement membranes surrounding blood venules. Despite its established role in tumor cell invasion, heparanase function in leukocyte extravasation has never been dem- onstrated. We found that T H 1/T C 1-type effector T cells are highly enriched for this enzyme, with a 3.6-fold higher heparanase mRNA expression compared with naive lym- phocytes. Using adoptive transfer of wild-type and heparanase-deficient effector T cells into inflamed mice, we show that T-cell heparanase was not required for ex- travasation inside inflamed lymph nodes or skin. Leuko- cyte extravasation through acute inflamed skin vessels was also heparanase independent. Furthermore, neutrophils emigrated to the inflamed peritoneal cavity independently of heparanase expression on either the leukocytes or on the endothelial and mesothelial barriers, and overex- pression of the enzyme on neutrophils did not facilitate their emigration. However, heparanase absence signifi- cantly reduced monocyte emigration into the inflamed peritoneal cavity. These results collectively suggest that neither leukocyte nor endothelial heparanase is required for T-cell and neutrophil extravasation through inflamed vascular barriers, whereas this enzyme is required for op- timal monocyte recruitment to inflamed peritoneum.— Stoler-Barak, L., Petrovich, E., Aychek, T., Gurevich, I., Tal, O., Hatzav, M., Ilan, N., Feigelson, S. W., Shakhar, G., Vlodavsky, I., Alon, R. Heparanase of murine effector lymphocytes and neutrophils is not required for their diapedesis into sites of inflammation. FASEB J. 29, 000–000 (2015). www.fasebj.org Key Words: extracellular matrix • extravasation • heparan sulfate • leukocytes • postcapillary venules EXTRAVASATING LEUKOCYTES must pass through the sub- endothelial basement membrane on their way to various interstitial spaces (1). The basement membrane sur- rounding most postcapillary venules is highly enriched with extracellular matrix (ECM) proteoglycans (PGs) decorated with various glycosaminoglycans (GAGs) (2). Heparan sulfate (HS) GAGs are major GAG constituents of vascular basement membranes and are thought to play a key role in the self-assembly, insolubility, and barrier properties of the ECM (2–5). Recently, we reported the existence of very high HS densities in the basement membranes of both skin and lymph node blood vessels, and their associated pericytes and fibroblastic reticular cells, respectively (6). To invade through these complex networks, leukocytes, as well as other invasive cells, have been postulated to deploy various degradative enzymes, such as proteases, matrix metal- loproteinases (MMPs), and GAG-degrading enzymes in- cluding HS-degrading enzymes (2, 3, 5). Because of their high negative charge, HS PGs bind and present different growth factors, enzymes, and inflammatory cytokines and chemokines (4). In addition to reducing the barrier prop- erties of basement membranes, enzymatic degradation of HS moieties by extravasating leukocytes may also liberate these bound molecules and thus locally modulate their avail- abilityand functionality within the extravascular milieu. Modifications created by the cell surface endosulfatases, Sulf1 and Sulf2, which remove 6-O sulfates from HS dis- accharides, can alter HS structure (7, 8). However, HS degradation is mediated primarily by a single enzymatically active heparanase present in mammals, an endo-b-D- glucuronidase that cleaves HS side chains into fragments of 10–20 disaccharides (9). Heparanase expression is up- regulated in many malignant and inflammatory settings by multiple types of cells (9–14). Indeed, tumor cell invasion and metastasis are tightly regulated by heparanase, and the enzymatic activity of this key HS-degrading enzyme has Abbreviations: 3D, 3-dimensional; BSA, bovine serum albumin; AS, antisense; CFA, Complete Freund’s Adjuvant; CMTMR, (5-(and -6-)-(((4-chloromethyl)benzoyl)amino)tetramethylrhod- amine); DC, dendritic cell; ECM, extracellular matrix; FACS, fluorescence-activated cell sorter; GAG, glycosaminoglycan; HEV, high endothelial venule; hpa-tg, heparanase transgene; ( continued on next page) 1 These authors contributed equally to this work. 2 Correspondence: Department of Immunology, The Weizmann Institute of Science, Rehovot 76100, Israel. E-mail: ronen.alon@ weizmann.ac.il doi: 10.1096/fj.14-265447 This article includes supplemental data. Please visit http:// www.fasebj.org to obtain this information. 0892-6638/15/0029-0001 © FASEB 1 The FASEB Journal article fj.14-265447. Published online January 29, 2015.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Heparanase of murine effector lymphocytes and neutrophils is not required for their diapedesis into...

The FASEB Journal • Research Communication
Heparanase of murine effector lymphocytesand neutrophils is not required for their diapedesisinto sites of inflammation
Liat Stoler-Barak,*,1 Ekaterina Petrovich,*,1 Tegest Aychek,* Irina Gurevich,* Orna Tal,*Miki Hatzav,* Neta Ilan,† Sara W. Feigelson,* Guy Shakhar,* Israel Vlodavsky,†
and Ronen Alon*,2
*Department of Immunology, The Weizmann Institute of Science, Rehovot, Israel; and †Cancer andVascular Biology Research Center, The Bruce Rappaport Faculty of Medicine, Technion, Haifa, Israel
ABSTRACT Heparanase, the exclusive mammalianheparan sulfate-degrading enzyme, has been suggested tobe utilized by leukocytes to penetrate through the densebasementmembranes surrounding blood venules. Despiteits established role in tumor cell invasion, heparanasefunction in leukocyte extravasation has never been dem-onstrated. We found that TH1/TC1-type effector T cellsare highly enriched for this enzyme, with a 3.6-fold higherheparanase mRNA expression compared with naive lym-phocytes. Using adoptive transfer of wild-type andheparanase-deficient effector T cells into inflamed mice,we show that T-cell heparanase was not required for ex-travasation inside inflamed lymph nodes or skin. Leuko-cyte extravasation through acute inflamed skin vessels wasalso heparanase independent. Furthermore, neutrophilsemigrated to the inflamed peritoneal cavity independentlyof heparanase expression on either the leukocytes or onthe endothelial and mesothelial barriers, and overex-pression of the enzyme on neutrophils did not facilitatetheir emigration. However, heparanase absence signifi-cantly reduced monocyte emigration into the inflamedperitoneal cavity. These results collectively suggest thatneither leukocyte nor endothelial heparanase is requiredfor T-cell and neutrophil extravasation through inflamedvascular barriers, whereas this enzyme is required for op-timal monocyte recruitment to inflamed peritoneum.—Stoler-Barak, L., Petrovich, E., Aychek, T., Gurevich, I.,Tal, O., Hatzav, M., Ilan, N., Feigelson, S. W., Shakhar, G.,Vlodavsky, I., Alon, R. Heparanase of murine effectorlymphocytes and neutrophils is not required for theirdiapedesis into sites of inflammation. FASEB J.29, 000–000 (2015). www.fasebj.org
Key Words: extracellular matrix • extravasation • heparansulfate • leukocytes • postcapillary venules
EXTRAVASATING LEUKOCYTES must pass through the sub-endothelial basement membrane on their way to variousinterstitial spaces (1). The basement membrane sur-rounding most postcapillary venules is highly enrichedwith extracellular matrix (ECM) proteoglycans (PGs)decorated with various glycosaminoglycans (GAGs) (2).Heparan sulfate (HS)GAGsaremajorGAGconstituents ofvascular basement membranes and are thought to playa key role in the self-assembly, insolubility, and barrierproperties of the ECM (2–5).
Recently, we reported the existence of very high HSdensities in the basement membranes of both skin andlymph node blood vessels, and their associated pericytesand fibroblastic reticular cells, respectively (6). To invadethrough these complex networks, leukocytes, as well asother invasive cells, have been postulated to deploy variousdegradative enzymes, such as proteases, matrix metal-loproteinases (MMPs), and GAG-degrading enzymes in-cluding HS-degrading enzymes (2, 3, 5). Because of theirhigh negative charge, HS PGs bind and present differentgrowth factors, enzymes, and inflammatory cytokines andchemokines (4). In addition to reducing the barrier prop-erties of basement membranes, enzymatic degradation ofHS moieties by extravasating leukocytes may also liberatethese boundmolecules and thus locally modulate their avail-ability and functionality within the extravascular milieu.
Modifications created by the cell surface endosulfatases,Sulf1 and Sulf2, which remove 6-O sulfates from HS dis-accharides, can alter HS structure (7, 8). However, HSdegradation ismediatedprimarily by a single enzymaticallyactive heparanase present in mammals, an endo-b-D-glucuronidase that cleavesHSside chains into fragmentsof10–20 disaccharides (9). Heparanase expression is up-regulated inmanymalignant and inflammatory settings bymultiple types of cells (9–14). Indeed, tumor cell invasionandmetastasis are tightly regulated byheparanase, and theenzymatic activity of this key HS-degrading enzyme has
Abbreviations: 3D, 3-dimensional; BSA, bovine serum albumin;AS, antisense; CFA, Complete Freund’s Adjuvant; CMTMR,(5-(and -6-)-(((4-chloromethyl)benzoyl)amino)tetramethylrhod-amine); DC, dendritic cell; ECM, extracellular matrix; FACS,fluorescence-activated cell sorter; GAG, glycosaminoglycan;HEV, high endothelial venule; hpa-tg, heparanase transgene;
(continued on next page)
1 These authors contributed equally to this work.2 Correspondence: Department of Immunology, TheWeizmann
Institute of Science, Rehovot 76100, Israel. E-mail: [email protected]: 10.1096/fj.14-265447This article includes supplemental data. Please visit http://
www.fasebj.org to obtain this information.
0892-6638/15/0029-0001 © FASEB 1
The FASEB Journal article fj.14-265447. Published online January 29, 2015.

beendirectly implicated in the ability of numerous types ofcancer cells to extravasate blood vessels (15–18).
In contrast to these nonhematopoietic cells, the roles ofheparanase expressed by different leukocytes in their in-vasive properties in general, and extravasation across ven-ular barriers in particular, remain elusive. It has beenshown that neutrophils, monocytes, and various types ofactivated T lymphocytes express heparanase and displayenzymatic activity of HS degradation ex vivo (19–24),suggesting a potential role for heparanase-mediated HS-degrading activity in leukocyte diapedesis. High hep-aranase expression was also identified in rat CD4+ T cellsinfiltrating the CNS during clinical disease (25). Func-tional heparanase is also expressed by mature dendriticcells (DCs) (26) and was implicated in DC motility andintravasation into skin draining lymphatic vessels and theirentry into lymph nodes (27, 28).
These studies collectively suggest that heparanase mayplay a key role in the ability of infiltrating leukocytes to crossbasement membrane barriers at sites of inflammation. Toaddress this question, we followed the in vivo migratorycapacities of multiple types of leukocytes derived from ei-thernormalorheparanase-deficient [heparanase-knockout(Hpse-KO)] mice (29) or heparanase-overexpressingheparanase transgene (hpa-tg) mice (30). We show thatT-cell and neutrophil heparanase is not required for extrav-asation and crossing through basement membranes inmultiple sites of inflammation. Neutrophil extravasationthrough acutely inflamed skin and mesothelial barrierssurrounding the inflamed peritoneal cavity took place in-dependently of heparanase activities, whereas monocytemigration across mesothelial barriers was heparanase de-pendent. Our results are a first direct demonstration thatheparanase-mediated degradation of theHS-rich basementmembranes is not a rate-limiting step in effector T-cell andneutrophil emigration through the basement membranesof inflamed blood vessels.
MATERIALS AND METHODS
Antibodies and reagents
Anti-mouse CD3e (145-2C11), anti-mouse CD28 (35.51), and PEanti-mouse CD11a (aL) (M17/4) were purchased from eBio-science (San Diego, CA, USA). Alexa 647-labeled goat anti-rat,CellTracker Violet, and (5-(and -6-)-(((4-chloromethyl)benzoyl)amino)tetramethylrhodamine) (CMTMR)were purchased fromInvitrogen (Grand Island, NY,USA). Anti-HSmonoclonalmouseIgM (10E4) was purchased from Seikagaku Corporation (Tokyo,Japan) and from USBiological Life Sciences (Salem, MA, USA).Goat anti-mouseVE-cadherinwas fromR&DSystems (Minneapolis,MN, USA). Rat anti-CD45 (30F11) was purchased from SantaCruz Biotechnology (Dallas, TX, USA). Rabbit anti-PECAM-1(platelet endothelial cell adhesion molecule 1; ab28364) andPE anti-mouse 7/4 (7/4) were purchased from Abcam(Cambridge,UnitedKingdom). Pacific Blue anti-mouseCD45(30F11), FITC anti-mouse/human CD11b (aM) (M1/70),APC/Cy7 anti-mouse Ly6G (1A8), PE anti-mouse CD18 (b2)
(M18/2), anti-mouse CD49d (a4) (9C10), anti-mouse integrinb7 (FIB504), anti-mouseCD29 (b1) (HMb1-1), APCanti-mouseCD62L (MEL-14), APC anti-mouse CXCR3 (CXCR3-173), Ar-menian hamster IgG isotype control (HTK888), PE rat IgG2aisotype control (RTK2758), and PE rat IgG1 isotype control(RTK2071)were all fromBioLegend (SanDiego,CA,USA). PE-conjugated goat anti-human IgG, Fcg Fragment Specific, Alexa647-labeled donkey anti-rat IgG, and Alexa 647-labeled goatanti-Armenian hamster IgG were from Jackson ImmunoR-esearch Laboratories (West Grove, PA, USA). PE anti-mouseCD162 (PSGL1) (2PH1), mouse P-selectin-IgG-Fc fusion pro-tein, and blood-lysing solution were obtained from BD Bio-sciences (San Jose, CA, USA). Complete Freund’s Adjuvant(CFA) was from Difco Laboratories (Detroit, MI, USA). Zymo-san was purchased from Sigma-Aldrich (St. Louis, MO, USA).
Animals
Wild-type (WT) mice, heparanase-deficient mice (Hpse-KO)(29), heparanase-overexpressing transgenic mice (hpa-tg) (30),and intercellular adhesion molecule (ICAM)-1/2 double-deficient mice (31) were of C57BL/6J background. For thegeneration of chimeric mice, C57BL/6 recipient mice were ir-radiatedwithg radiationat950rad fromacesiumsource.Twenty-four hours later, recipient mice were intravenously injected with5 3 106 fresh bone marrow cells containing white blood cells(counted after red cell lysis) obtained from hpa-tg or control WTmice. Chimeric mice were used 6 weeks after bone marrowtransplantation. All animal procedures were approved by the An-imal Research Committee at the Weizmann Institute of Science.
Preparation of effector T cells
Spleens were removed from 8- to 12-week-old mice. T cells werepurified over a Novamed column (Novamed, Jerusalem, Israel)and stimulated on plates coated with 1 mg/ml anti-CD3 and5 mg/ml anti-CD28 for 48 hours, followed by expansion for 5additional days with 10 U/ml of IL-2.
Induction of lymph node and skin inflammation
Inflammation within the popliteal lymph nodes was induced bysubcutaneous injection of CFA (2.5 mg/ml, 50–100 ml) to boththe footpad and flank, 48 hours before adoptive transfer experi-ments. Skin inflammation in the flank was induced 48 hours be-fore adoptive transfer by multizone intradermal CFA injections(2.5 mg/ml, 10 ml per injection), as described previously (32).Acute skin inflammation was induced in theflank 6 hours beforeskinremovalandfixation,by intradermalCFAinjection(2.5mg/ml,10 ml per injection).
Analysis of T-cell accumulation in lymph nodes, blood,and spleen
WT or heparanase-deficient effector T cells (107 cells) were eachlabeled with either CMTMR (10 mM for 30 minutes) or 2,3,6,7-tetrahyfro-9-bromomethyl-1H,5H-quinolozino-(9,1-gh)-coumarin(CellTracker Violet; 5 mM for 20 minutes), mixed at a 1:1 ratio,and injected intravenously to WT recipient mice. Mice wereanesthetized 2 hours after injection, bled, and differently labeledT cells were enumerated by fluorescence-activated cell sorter(FACS). To determine T-cell accumulation in popliteal lymphnodes and spleen, single-cell suspensions were prepared by me-chanical disruption, as described previously (33).Reversal of thesecell trackerdyes didnot alter the relative entry of the labeledT-cellpopulations.
(continued from previous page)Hpse-KO, heparanase knockout; HS, heparan sulfate; ICAM,intercellular adhesion molecule; MMP, matrix metallopro-teinase; PECAM-1, platelet endothelial cell adhesion molecule 1;PG, proteoglycan; S, sense; WT, wild-type
2 Vol. 29 May 2015 STOLER-BARAK ET AL.The FASEB Journal x www.fasebj.org

Intravital microscopic live imaging of effector T cellsentering inflamed lymph nodes
To enumerate the number of differently labeled injected T cellsthat accumulated either intravascularly or extravascularly inpopliteal lymph nodes, we used 2-photon microscopy. Lymphnodes of live recipient mice were imaged 2 hours after in-travenous injection of labeled effector T cells and Qtracker 655(Live Technologies, Carlsbad, CA, USA) quantum dots using anUltimaMultiphotonMicroscope attached to an upright platform(Olympus BX61WI; Tokyo, Japan) equipped with a 320 (NA0.95) water-immersion objective. An excitation wavelength of850 nmwas used for simultaneous imaging of CellTracker Violet,CMTMR, and Qtracker 655. To create a typical time-lapse se-quence, a 50-mm-thick section was scanned at 5 mmZ steps every47 seconds. Cell movement was analyzed in 3-dimensional (3D)usingVolocity 4 software (PerkinElmer,Waltham,MA,USA). Forcalculation of cell velocity anddisplacement rate, the coordinatesof each cell were recorded and tracked over time.
Analysis of effector T-cell accumulation and extravasationthrough skin vessels
Differently labeled T cells were coinjected intravenously as de-scribed above.Micewere killed 6 hours later, and either inflamedskin tissues (CFA injection spots) or noninflamed tissues wereremoved and immediately fixed in 4% paraformaldehyde.Whole-mount sampleswere stainedwith rat anti-mousePECAM-1in 0.3% Triton X-100/0.5% bovine serum albumin (BSA) over-night followed by Alexa 647-labeled goat anti-rat antibody.Immunostained tissues were imaged with a 2-photonmicroscopewith an excitation wavelength of 850 nm for simultaneous imag-ing of CellTracker Violet, CMTMR, and Alexa 647 at 320 mag-nification. A 50-mm-thick section was scanned at 3 mm Z steps.Intravascular or extravascular dye-labeled T cells were enumer-ated in 3D in multiple fields with Volocity 4 software.
Immunohistochemistry of popliteal lymph nodes andCD45-positive cells in the skin
Popliteal lymph node was removed 48 hours post-CFA injection,whereas acutely inflamed skin flank tissue was removed 6 hoursafter CFA injection. All tissues were fixed in 4% paraformal-dehyde and embedded in paraffin. Four-micrometer sectionswere subjected to target retrieval, and deparaffinization was per-formed using the target retrieval solution (pH 9; Dako, Pro-duktionsvej, Denmark) for 20 minutes at 95°C. All sections weresimilarly blocked with 10% horse serum, incubated with primaryantibody at 4°C overnight (HS 1:50, VE-cadherin 1:50, PECAM-11:50, or CD45 1:20), washed, and incubated with secondary an-tibody (1:100) and the nuclear dye Hoechst for 40 minutes atroom temperature. Sections were imaged by a Delta-Vision mi-croscope (Applied Precision, Issaquah, WA, USA).
Zymosan-induced peritoneal inflammation
Mice were bled and then injected intraperitoneally with 1 mgzymosan in0.2mlPBSor control solution(0.2mlPBS).Micewereinjected intraperitoneally with 6 ml washing solution for perito-neal lavage (3 mM EDTA in PBS) 2 hours (neutrophils) or4 hours (monocytes) after zymosan administration. The ab-domen was massaged, and free fluid was recovered from theperitoneal cavity. Blood and peritoneal lavage samples werestained for neutrophil or monocyte FACS analysis. Neutrophils
were defined as CD11bhigh and Ly6Ghigh. Monocytes weredefined as Ly6Glow and 7/4high (34).
RT-PCR
Total RNA was extracted from naive or effector T cells, usinga High Pure RNA Isolation Kit (Roche, Basel, Switzerland). Thereverse transcription reaction was performed using the HighCapacity cDNA Reverse Transcription Kit (Applied Biosystems,Foster City, CA, USA) with thermal cycling conditions accordingto the manufacturer’s instructions. The primer sequences wereheparanase [sense (S), 59-CTT GCT GTC CAA CAC CTT G-39;antisense (AS), 59-GGC TGA CAG GCC CAA TTT A-39), MMP-2(S, 59-AGC GTG AAG TTT GGA AGC AT-39; AS, 59-CAC ATCCTT CAC CTG GTG TG-39), MMP-9 (S, 59-AGA CGA CAT AGACGGCAT CC-39; AS, 59-GTG GTT CAG TTG TGG TGG TG-39),MMP-14 (S, 59-GCC TGG AACATT CTA ACG A-39; AS, 59-CTTTGTGGGTGACCC TGACT-39), andMMP-25 (S, 59-GCTGACTCG CTA TGG CTA CC-39; AS, 59-GTC ATT GGG TCC ATTTGT CC-39]. Amplification mixtures contained 23 LightCycler480 SYBRGreenmaster I (Roche). RT-PCRwas performed usinga LightCycler 480 (Roche) under the following conditions: 45amplification cycles of denaturation at 95°C for 10 seconds andannealing and extension at 60°C for 40 seconds. Relative mRNAexpression was normalized to the ribosomal protein L32.
Heparanase activity assay
Preparation of Na235SO4-labeled ECM-coated 35mm dishes and
determination of heparanase activity were performed as de-scribed in detail elsewhere (35). Briefly, naive or effector T cells(107 cells) derived from WT or heparanase-deficient mice weresubjected to 3 cycles of freeze/thaw in 1 ml reaction buffer[20mMphosphate-citratebuffer (pH6.2)containing1mMDTT,1mMCaCl2, and50mMNaCl]and incubation(24hours at 37°C)with 35S-labeled ECM. The incubation medium was centrifuged(20,0003 g at 4°C for 1minute), and the supernatant containing35S-labeled HS degradation fragments was analyzed by gel filtra-tion on a Sepharose CL-6B column. Typically, fragments releasedby heparanase are eluted at fractions 10–20 (29).
P-selectin-IgG-Fc fusion protein staining
EffectorT cells or fresh bonemarrow cells werewashedoncewithstandard stainingmedia (DMEM, 0.1%BSA, and 0.1%NaAzide)or with EDTA media (DMEM, 0.1% BSA, 0.1% Na Azide, and5mMEDTA). A total of 33 105 cells were stained with 30mg/mlmouse P-selectin-IgG-Fc fusion protein diluted in 100ml of eitherstandard or EDTA media for 30 minutes on ice. For neutrophilstaining in bone marrow cells, APC/Cy7 anti-Ly6G antibody wasincluded. A total of 400ml standard or EDTAmedia was added tothe cell suspension, and cells were spun at 4003 g for 5 minutesand stained for 30 minutes on ice with 2 mg/ml PE-conjugatedgoat anti-human IgG antibody, in either the standard or EDTA-containing media. Cells were washed once in the relevant mediaand analyzed by flow cytometry.
Statistical analysis
All data are reportedasmeanvalues6 SEMandwere analyzedwithPrism5 software (GraphPad Software, Incorporated, La Jolla,CA,USA) using Student’s t test. Coinjected differently labeled T cellsthat accumulated in individual fields of view were compared bya paired Student’s t test. Comparisons were deemed significantfor 2-tailed P values,0.05.
HEPARANASE IS DISPENSABLE FOR LEUKOCYTE EMIGRATION 3
RESULTS
Effector T cells up-regulate heparanase expressionand exhibit high heparanase activity
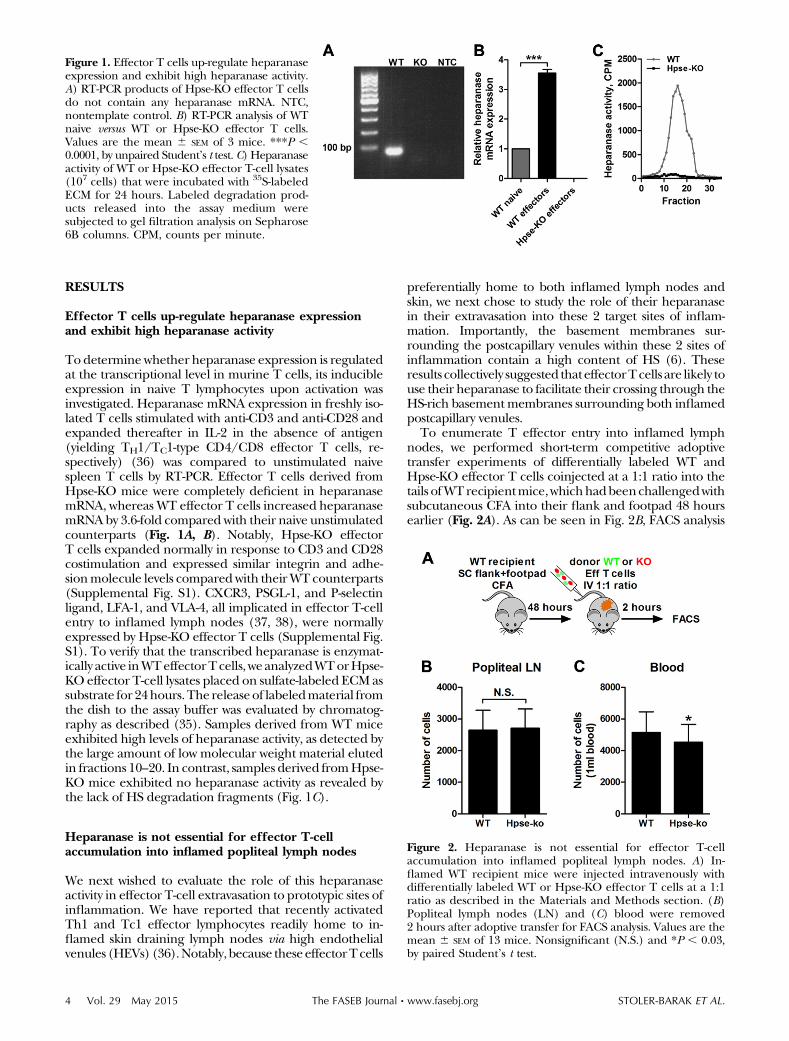
To determinewhether heparanase expression is regulatedat the transcriptional level in murine T cells, its inducibleexpression in naive T lymphocytes upon activation wasinvestigated. Heparanase mRNA expression in freshly iso-lated T cells stimulated with anti-CD3 and anti-CD28 andexpanded thereafter in IL-2 in the absence of antigen(yielding TH1/TC1-type CD4/CD8 effector T cells, re-spectively) (36) was compared to unstimulated naivespleen T cells by RT-PCR. Effector T cells derived fromHpse-KO mice were completely deficient in heparanasemRNA, whereas WT effector T cells increased heparanasemRNAby 3.6-fold compared with their naive unstimulatedcounterparts (Fig. 1A, B). Notably, Hpse-KO effectorT cells expanded normally in response to CD3 and CD28costimulation and expressed similar integrin and adhe-sionmolecule levels comparedwith theirWT counterparts(Supplemental Fig. S1). CXCR3, PSGL-1, and P-selectinligand, LFA-1, and VLA-4, all implicated in effector T-cellentry to inflamed lymph nodes (37, 38), were normallyexpressed by Hpse-KO effector T cells (Supplemental Fig.S1). To verify that the transcribed heparanase is enzymat-ically active inWTeffectorTcells, weanalyzedWTorHpse-KO effector T-cell lysates placed on sulfate-labeled ECMassubstrate for 24hours.The releaseof labeledmaterial fromthe dish to the assay buffer was evaluated by chromatog-raphy as described (35). Samples derived from WT miceexhibited high levels of heparanase activity, as detected bythe large amount of low molecular weight material elutedin fractions 10–20. In contrast, samples derived fromHpse-KO mice exhibited no heparanase activity as revealed bythe lack of HS degradation fragments (Fig. 1C).
Heparanase is not essential for effector T-cellaccumulation into inflamed popliteal lymph nodes
We next wished to evaluate the role of this heparanaseactivity in effector T-cell extravasation to prototypic sites ofinflammation. We have reported that recently activatedTh1 and Tc1 effector lymphocytes readily home to in-flamed skin draining lymph nodes via high endothelialvenules (HEVs)(36).Notably, because theseeffectorTcells
preferentially home to both inflamed lymph nodes andskin, we next chose to study the role of their heparanasein their extravasation into these 2 target sites of inflam-mation. Importantly, the basement membranes sur-rounding the postcapillary venules within these 2 sites ofinflammation contain a high content of HS (6). Theseresults collectively suggested thateffectorTcells are likely touse their heparanase to facilitate their crossing through theHS-rich basementmembranes surrounding both inflamedpostcapillary venules.
To enumerate T effector entry into inflamed lymphnodes, we performed short-term competitive adoptivetransfer experiments of differentially labeled WT andHpse-KO effector T cells coinjected at a 1:1 ratio into thetails ofWTrecipientmice,whichhadbeenchallengedwithsubcutaneous CFA into their flank and footpad 48 hoursearlier (Fig. 2A). As can be seen in Fig. 2B, FACS analysis
Figure 1. Effector T cells up-regulate heparanaseexpression and exhibit high heparanase activity.A) RT-PCR products of Hpse-KO effector T cellsdo not contain any heparanase mRNA. NTC,nontemplate control. B) RT-PCR analysis of WTnaive versus WT or Hpse-KO effector T cells.Values are the mean 6 SEM of 3 mice. ***P ,0.0001, by unpaired Student’s t test. C) Heparanaseactivity of WT or Hpse-KO effector T-cell lysates(107 cells) that were incubated with 35S-labeledECM for 24 hours. Labeled degradation prod-ucts released into the assay medium weresubjected to gel filtration analysis on Sepharose6B columns. CPM, counts per minute.
Figure 2. Heparanase is not essential for effector T-cellaccumulation into inflamed popliteal lymph nodes. A) In-flamed WT recipient mice were injected intravenously withdifferentially labeled WT or Hpse-KO effector T cells at a 1:1ratio as described in the Materials and Methods section. (B)Popliteal lymph nodes (LN) and (C) blood were removed2 hours after adoptive transfer for FACS analysis. Values are themean 6 SEM of 13 mice. Nonsignificant (N.S.) and *P , 0.03,by paired Student’s t test.
4 Vol. 29 May 2015 STOLER-BARAK ET AL.The FASEB Journal x www.fasebj.org

revealed similar numbers of Hpse-KO and WT effectorT cells recovered in inflamed popliteal lymph nodes 2 hpostadoptive transfer. Because the numbers of Hpse-KOeffectorT cells recovered in the blood shortly after transferwere only slightly reduced (Fig. 2C), these experimentscollectively suggest that the absence of heparanase on ef-fectorT cells does not alter their recirculationpatterns andaccumulation in inflamed lymph nodes.
T-cell heparanase is not essential for basementmembrane crossing and interstitial motility of effectorT cells inside inflamed lymph nodes
Similar levels of T-cell accumulation in lymph nodesreported by FACS-based assays could reflect normal stick-ing and extravasation through lymph node HEVs but de-fective crossing of the basement membrane of thesepostcapillary venules. Indeed, many T cells often reside inperivascular HEV compartments without entering thelymph node parenchyma (39). We thus reasoned thata fraction of heparanase-deficient effector T cells may failto cross the basement membrane barrier surroundingthese vessels. To check this possibility, we repeated the
competitive adoptive transfer experiments described intheprevious section(Fig. 3A)but followedby2-photon live-imaging microscopy the diapedesis through the denseECM HS and extravascular motility of these differentT cells inside inflamed lymph nodes (Fig. 3B and Supple-mentalVideo1).As canbeappreciated inFig. 3C–H, T-cell-expressed heparanase was not required for lymphocyteextravasation, crossing of the HEV basement membrane,entry to the lymph node parenchyma, or for interstitialmotility inside the inflamed lymph nodes.
The heparanase independence of effector T-cell emi-gration across the HS-rich basement membrane ofinflamed HEVs could reflect insufficient functionality ofthis enzyme in these T cells. We therefore next askedwhether robust overexpression of the enzyme in theseT cells will provide these lymphocytes with gain of functionin terms of accumulation in inflamed lymph nodes aspredicted from their high ECM degradation activity (30).T cells derived from heparanase transgenic mice normallyexpanded in response to CD3 and CD28 costimulationand transcribed 1 order of magnitude higher levels ofheparanase than WT effector T cells (Fig. 4A). Notably,these T cells expressed similar adhesion molecule levelscompared to their WT counterparts (data not shown).
Figure 3. T-cell heparanase is not essential for basement membrane crossing and interstitial motility of effector T cells insideinflamed lymph nodes. A) Inflamed WT recipient mice were injected intravenously with labeled WT or Hpse-KO effector T cellsat a 1:1 ratio. Migration to inflamed popliteal lymph nodes was observed 2 hours after adoptive transfer and intravenous injectionof Qtracker 655 quantum dots by 2-photon live microscopy. B) Representative field of view of popliteal lymph node with WT(green) and Hpse-KO (red) effector T cells. Field dimensions are 500 3 500 mm2. In the right inset, the green dot depicts WTeffector T cell and the broken white circle depicts 10 mm distance from the inflamed vessel wall immunostained for ECM HS(red). Endothelial cells are marked by VE-cadherin (cyan) staining. (C) Number of cells extravasating the vessels and (D)percentage of emigrating cells that extravasated at least 10 mm away from the vessel wall calculated out of only extravasated cells.Measured values are the mean 6 SEM of 8 fields from 5 mice. (E) WT and (F) Hpse-KO cell trajectories determined at 47 secondintervals. 3D paths tracking T-cell movement are depicted as if emanating from a common starting point. Scatter plots illustratingthe (G) velocities and (H) displacement rates of 26 individual T-cell tracks. Nonsignificant (N.S.), by paired Student’s t test.
HEPARANASE IS DISPENSABLE FOR LEUKOCYTE EMIGRATION 5

Interestingly, competitive adoptive transfer of equal num-bers of differentially labeledWTandhpa-tg effector T cells(Fig. 4B) revealed that despite the much higher hepar-anasemRNA levels in the transgenic T cells, their ability toaccumulate in WT inflamed lymph nodes shortly aftertransfer was only marginally increased relative to WT ef-fector T cells, although at a statistically significant manner(Fig. 4C), with similar numbers of WT and hpa-tg effectorT cells recovered from the circulation (Fig. 4D). Thus,a forced overexpression of theHS-degrading enzyme doesnot further increase the seemingly high and heparanase-independent intrinsic ability ofWTeffector T cells to enterinflamed lymph nodes.
T-cell heparanase is dispensable for T-cellextravasation through inflamed skin vessels andcrossing of their basement membranes
The composition and barrier properties of basementmembranes surrounding HEVs differ from those of flatendothelial cells withinnonlymphoidpostcapillary venules(40), but both vessels are surrounded by a dense layer ofHS-enriched ECM (6). We therefore next wished to in-vestigate the role of T-cell heparanase in the emigration ofeffector Th1 and Tc1 cells into postcapillary venules ofinflamed skin. We recently reported that these lympho-cytes can enter CFA inflamed flank tissue within 4–6 hourspostadoptive transfer (36). To accurately determine the
accumulation, extravasation, and basement membranecrossing of heparanase-deficient and WT effector T cells,we coinjected these differentially labeled effector cells ata 1:1 ratio into the tails ofWT recipientmice that had beenchallenged intradermally with CFA into their flank48 hours earlier (Fig. 5A). We performed high-resolutionT-cell-tracking analysis on whole-mounted skin samplesstained with the blood vessel marker PECAM-1 (Fig. 5B).Notably, the amounts of effector T cells found to extrava-sate CFA inflamed skin vessels ofWT recipientmice and tosuccessfully cross the basement membrane of these vesselsand enter the interstitial dermal space were all in-distinguishable between Hpse-KO and WT T cells (Fig.5C–E). Collectively, these results indicate that heparanasedeficiency does not impair the ability of effector T cells toextravasate and penetrate through the HS-enriched base-ment membranes of inflamed skin vessels on their way tothe interstitial space.
Notably, this apparently normal ability of heparanase-deficient T cells to cross HS-rich basement membranes ofboth lymph node HEVs and peripheral skin vessels andenter the interstitial spaces of these tissues could not beexplained by compensatory proteolytic mechanisms be-cause the transcription levels of themajorMMPs expressedby effector T cells were not significantly elevated (Supple-mental Fig. S2).
Heparanase is not essential for the entry of leukocytesinto acutely inflamed skin
Thus far, our analysis was limited to effector T cells. Be-cause heparanase is also expressed by other leukocytes, aswell as by endothelial and nonhematopoietic stromal cells(23), we next wished to investigate if the combined activi-ties of the enzyme expressed by all these cells are necessaryfor the emigration of endogenous blood-borne CD45-positive cells (i.e., leukocytes) into acutely CFA inflamedskin. To this end, WT or heparanase-deficient mice wereinjected intradermally with CFA into theirflank. Infiltrateswereassessedat 6hours afterCFAadministration (Fig. 6A),a time period sufficient to attract multiple migrating leu-kocytes (Fig. 6B), primarily circulating neutrophils, andmonocytes to the site of inflammation (41, 42). FACSanalysis of blood-circulating leukocytes revealed that atsteady state, Hpse-KO mice have more circulating CD45leukocytes (Fig. 6C). High-resolution cell-tracking analysisof skin sections derived from eitherWT orHpse-KOCFAinflamed skin stained with the blood vessel markerPECAM-1 and CD45 revealed massive and comparableleukocyte infiltration into the skin interstitium inducedby the CFA stimulus in bothWT and Hpse-KOmice (Fig.6B, D). Importantly, the fractions of extravasating leu-kocytes and the extent these leukocytes migrated awayfrom the inflamed vessels they have crossed were practi-cally identical inWTandHpse-KO skin (Fig. 6E, F).Theseresults therefore suggest that despite an apparentregulatory role of the enzyme in the steady state of blood-circulating leukocytes, both leukocyte and endothelial-derived heparanase activities appear dispensable forleukocyte crossing of the denseHS layer surrounding theskin blood vessels (6) and for their successful extravasa-tion into the tissue.
Figure 4. Heparanase overexpression in effector T cellsslightly increases their accumulation in inflamed popliteallymph nodes. A) RT-PCR analysis of WT versus hpa-tg effectorT cells. Values are the mean 6 SEM of 3 mice. ***P , 0.0003,by unpaired Student’s t test. B) Inflamed WT recipient micewere injected intravenously with labeled WT or hpa-tg effectorT cells at a 1:1 ratio. (C) Popliteal lymph nodes (LN) and (D)blood were collected 2 hours after adoptive transfer for FACSanalysis. Values are the mean 6 SEM of 8 mice. Nonsignificant(N.S.) and **P , 0.003, by paired Student’s t test.
6 Vol. 29 May 2015 STOLER-BARAK ET AL.The FASEB Journal x www.fasebj.org

Heparanase is dispensable for neutrophil entry intozymosan-inflamed peritoneum but contributes tomonocyte entry into this compartment
Our results in the skin and lymph node inflammationmodels suggested that blood-borne heparanase-deficientleukocytes normally cross HS-rich vascular barriers inmultiple inflammatory settings. Zymosan-induced perito-nitis is another classic readout for integrin-mediated neu-trophil extravasation during acute inflammatory response(34, 43). In response to zymosan administration (Fig. 7A),blood-borne neutrophils extravasate through postcapillaryvenules surrounding the inflamed peritoneal cavity andthen cross through the mesothelial barrier. We reasonedthat combined leukocyte and endothelial and/ormesothelial-based heparanase activities (23) may facili-tate the ability of blood-borne neutrophils to enter thezymosan-inflamed peritoneal cavity. Notably, neutro-phils from Hpse-KO bone marrow expressed similar b2integrins and P-selectin ligand levels compared withWT (Supplemental Fig. S3). Consistent with the resultsshown in Fig. 6C, the number of baseline circulatingneutrophils in Hpse-KO mice was twice the number ofcirculating neutrophils in WTmice (Fig. 7B, right inset).The number of neutrophils recovered in the inflamedperitoneal cavity in Hpse-KOmice was also doubled (Fig.
7B, C). In contrast, and consistent with the literature(43), significantly reduced numbers of neutrophils (3.4-fold) were recovered from the inflamed peritoneal cavityof ICAM-1 and ICAM-2 double-deficient mice despitea dramatically elevated number of circulating neu-trophils in these mice (Supplemental Fig. S4). Thus,heparanase activities on both the emigrating neutrophilsand on the multiple barriers they cross (i.e., vascular andmesothelial) are dispensable for the ability of neutrophilsto cross these barriers.
Heparanase expressed by monocytes was suggested todegrade ECM HS during in vitro transmigration (44). Wetherefore used a similar zymosan-induced peritoneum in-flammationmodel to investigate the role of heparanase inmonocyte emigration to the peritoneal cavity (Fig. 7D).Importantly, the number of blood-circulating monocytesin Hpse-KO mice was comparable to the number of cir-culating monocytes in WT mice (Fig. 7E, right inset).However, the number of monocytes recovered in theinflamed peritoneal cavity in Hpse-KO mice was reducedby 2-fold at the earliest time point after zymosan challengeat which significant monocyte emigration could be detec-ted (Fig. 7E, F) (34). Therefore, heparanase activity is re-quired for monocyte emigration, but not for neutrophilemigration across heparanase-deficient vascular and me-sothelial barriers.
Figure 5. Heparanase is dispensable for effector T-cell extravasation through inflamed skin vessels and crossing of their basementmembrane. A) Inflamed WT recipient mice were injected intravenously with labeled WT or Hpse-KO effector T cells at a 1:1ratio. The inflamed flank was removed 6 hours after adoptive transfer, followed by fixation and whole-mount staining forPECAM-1. Z sections were acquired using a 2-photon microscope. IHC, immunohistochemistry. B) Representative field showingWT (red) or Hpse-KO (green) effectors and PECAM-1 (blue) staining. White cells are endogenous autofluorescent cells. Fieldsize, 500 mm. Scale bar, 90 mm. C) Number of cells per field. D) Percentage of intravascular and extravascular cells per mouse. E)Percentage of emigrating T cells that extravasated at least 10 mm from the vessel wall calculated out of only extravasated cells permouse. Values are the mean 6 SEM of (C) 55 fields or (D and E) 4 mice. Nonsignificant (N.S.), by paired Student’s t test.
HEPARANASE IS DISPENSABLE FOR LEUKOCYTE EMIGRATION 7

Heparanase overexpression in neutrophils does notfacilitate their entry into zymosan-inflamed peritoneum
In a previous part (Fig. 4C), we found that overexpressionofheparanase on effector T cells marginally increased theiremigration into inflamed lymph nodes. Although hepar-anase activity was not required for neutrophil entry into theinflamed peritoneum (Fig. 7B), we hypothesized that be-cause this entry involves neutrophil crossing of multiplebarriers, a forced overexpression of heparanase on theseleukocytes may facilitate their emigration into the inflamedperitoneal cavity. To that end, we generated chimeric micethat express normal heparanase on all their radiation-resistant cells but are reconstituted with bone marrowexpressing either normal or overexpressed heparanase lev-els (herein WT chimera and hpa-tg chimera, respectively).We then used these 2 chimericmice to compare neutrophilexit frombloodandentry into the inflamedperitonealcavityat the earliest time point of the inflammatory process, i.e.,2 hours after zymosan challenge (34) (Fig. 8A). Surprisingly,neutrophil emigration to the inflamed peritoneal cavity wascomparable in theWTandhpa-tg chimeras (Fig. 8B). Theseresults further suggest that even when overexpressed, hep-aranase does not accelerate the emigration of neutrophilsfrom the circulation to the inflamed peritoneal cavity.
DISCUSSION
Vascular and lymphatic basement membranes are a sheetof interconnecting macromolecules, including laminins
and collagen IV that are physically stabilized by cross-linking GAGs, primarily HS-decorated PGs, such as perle-can, generated by cells of the vascular wall. The basementmembrane provides structural support and signalingproperties to its neighboring cells (1).
Mesenchymal cells such as fibroblasts and invasive tu-mor cells were demonstrated to employ multiple pro-teolytic and GAG-degrading enzymes to invade and crossbasementmembranes (45). Among these GAG-degradingmachineries, heparanase, the sole mammalian HS-degrading endo-b-D-glucuronidase, was indicated to facil-itate tumor cell invasion andmigration (15–18, 46).Unlikein these nonhematopoietic cells, the function of proteasesand GAG-degrading enzymes in enabling different leuko-cytes that express them to extravasate venular barriers hasbeen unclear. A first indication for a role of heparanase inleukocyte invasion has been provided in works addressingthe role of this enzyme in DCs migrating from peripheraltissues such as the skin to draining lymph nodes (27).These leukocytes must utilize complex interstitial motilitythrough the ECM of multiple barriers. Heparanase ex-pressed byDCs andmonocytes was found to degrade ECMHS during in vitro transmigration (26, 44). An in vivotransmigration assay confirmed these results by report-ing defective motility and intravasation of heparanase-deficient DCs into skin draining lymphatic vessels andreduced entry into draining lymph nodes (27). Moreover,a significant reduction of FITC-internalized migratingDCs from the skin was observed in the lymph nodes ofheparanase-deficient mice compared with WT mice (28).
Figure 6. Heparanase is not essential for leukocyte entry into inflamed skin. A) Inflamed flank tissue was removed from WT orHpse-KO mice, followed by fixation and paraffin embedding. B) Representative fields showing CD45-positive leukocytes (red)and PECAM-1 (green) staining. Field dimensions are 3003 300 mm2. Scale bars, 20 mm. C) Blood FACS analysis for CD45-positivecells. Values are the mean 6 SEM of 6 mice. D) Number of extravasated CD45-positive cells in the skin per field. E) Percentage ofextravasated cells in inflamed skin per field. F) Percentage of emigrating cells that extravasated at least 10 mm away from the inflamedvessel wall calculated out of only extravasated cells. Images were acquired using a Delta-Vision microscope. Values are the mean6 SEM
of 44 fields from 6 mice per group. N.S., nonsignificant. **P , 0.005 and ***P , 0.0001, by unpaired Student’s t test.
8 Vol. 29 May 2015 STOLER-BARAK ET AL.The FASEB Journal x www.fasebj.org

Despite thesepublished studies, and theevidence forup-regulated heparanase on leukocytes in inflammatory set-tings (25), direct in vivo evidence for promigratory roles ofheparanase expressed by T cells, neutrophils, and mono-cytes known to cross vascular and epithelial HS-enrichedbarriers has not been reported to date. The putative con-tributions of heparanase for lymphocyte adhesion andmigration were based primarily on heparanase blockingwith heparin (47), a blocker of heparanase as well as ofotherHS-bindingmolecules (48).We thereforedecided todirectly assess the contribution of heparanase to leukocytediapedesis using either genetic ablation or overexpressionof the enzyme in multiple types of leukocytes. We hencechose to investigate the contribution of heparanase to themigratory properties of 3 different leukocytes, known to
express heparanase, effector T cells, neutrophils, andmonocytes (19–24). Our results clearly indicate that hep-aranase is not required for the invasion of neutrophils or Teffector cells through theHS-richbarriers surrounding theinflamedvenularwalls indifferent inflamed tissues, suchaslymph node, skin, or peritoneum. Furthermore, over-expression of the enzyme did not facilitate neutrophil ex-travasation and crossing of both venular and epithelialbarriers of the inflamed peritoneum, measured at earliesttime points of inflammation. In the same model, thecombined deficiency of heparanase on both circulatingneutrophils and on inflamed stromal cells also did notresult in any noticeable impairment of neutrophil extra-vasation from blood and their crossing through the mul-tiple vascular and mesothelial barriers surrounding the
Figure 7. Heparanase is dispensable for neutro-phil entry into zymosan-inflamed peritoneumbut contributes to monocyte entry into thiscompartment. A) WT or Hpse-KO mice werebled and injected with zymosan or control PBS.At 2 hours postinjection, peritoneal lavage wasextracted for neutrophil FACS analysis. B)Number of peritoneal neutrophils. Values arethe mean 6 SEM of 4–6 mice. The right insetshows WT or Hpse-KO blood FACS analysis forcirculating neutrophils. Values are the mean6 SEM
of 4 mice. *P, 0.02 and **P, 0.009, by unpairedStudent’s t test. C) Representative FACS analysisof peritoneal lavage neutrophils. Neutrophilswere defined as CD11bhigh and Ly6Ghigh. D)WT or Hpse-KO mice were bled and injectedwith zymosan or control PBS. At 4 hours post-injection, peritoneal lavage was extracted andmonocyte numbers were determined by FACSanalysis. E) Number of peritoneal monocytes.Values are the mean 6 SEM of 4–12 mice. Theright inset shows WT or Hpse-KO blood FACSanalysis for circulating monocytes. Values arethe mean6 SEM of 10–12 mice. **P, 0.005 and***P , 0.0007, by unpaired Student’s t test. F)Representative FACS analysis of peritoneallavage monocytes. Monocytes were defined asLy6Glow and 7/4high.
HEPARANASE IS DISPENSABLE FOR LEUKOCYTE EMIGRATION 9

peritoneal cavity. Interestingly, however, the combineddeficiency of heparanase on both circulating monocytesand on inflamed peritoneal vascular and mesothelial cellsresulted in a significant impairment of monocyte emigra-tion and crossing through these multiple barriers. Never-theless, althoughalluding toa specializedroleofheparanasein monocyte migration, these results do not allow us to dif-ferentiate between the contribution of monocyte hepar-anase to thebloodextravasation stepper se, as opposed to thesequential transepithelial migration step involved in mono-cyteentry to theperitoneal cavity. Future studieson the rolesof monocyte heparanase in extravasation from blood andtransepithelial migration will be required to validate aspecific contributionofheparanase tomonocyte crossingthroughHS-rich venular basementmembranes as opposedto monocyte migration through other HS-containingbasement membranes and interstitial spaces.
Although heparanase is not essential for neutrophil andT effector cell invasion and migration across inflamedvascular barriers, it could play an important role as a tran-scription factor in these leukocytes. For instance, it wasfound that the enzyme enters the nucleus of activatedT lymphocytes and regulates the transcription of severalinducible immune response genes by controlling histoneH3 methylation patterns (49). Along this line, the in-creasednumberof circulating leukocytes inHpse-KOmiceraises apossible regulatory roleof this enzyme in the steady-state number of blood-circulating leukocytes (50). Inaddition, enzymatically inactive proheparanase was impli-cated in leukocyte integrin-mediated adhesion to the ECM(51). Proheparanase is stored in intracellular vesicles to-gether with its processing enzyme, cathepsin L, whichyields the active form of the enzyme (52). In light of thenormal ability of heparanase-deficient neutrophils andT cells to cross HS barriers in vivo, it is therefore possiblethat neither neutrophils nor effector T cells release theirenzymatically active enzyme from these stores during theirencounter of and migration through blood vessels. Onceresiding in inflamed tissues, these leukocytes may be ca-pable of releasing the processed enzyme in response toadditional cytokine signals that arenonencounteredby theextravasating leukocytes. Furthermore, catalytically activeheparanase canoptimally degrade itsHS substrateat acidicpH within sites of inflammation, whereas at physiologicpH, there is little or no heparanase activity (15). Thus,
rather than facilitating T-cell and neutrophil invasion, themajor functions of leukocyte- and stromal-derived hepar-anase once released and rendered enzymatically active inthe inflammatory milieu may involve the liberation of HS-bound growth factors and cytokines to be made availablefor tissue remodeling, wound healing, angiogenesis, andinflammation resolution (53). In addition, heparanasemay either down-regulate an ongoing inflammatory re-sponse by releasing a trisulfated HS-bound disaccharidethat inhibits TNF production by T cells and macrophages(54) or up-regulate inflammation by releasing other HSfragments that bind TLR-4 (55). Moreover, it was foundthat heparanase overexpression increases the retention ofhematopoietic stem cells in the bone marrow (50). Ourpreliminary results support such a key role of heparanase.Indeed,chimericmiceexpressingnormalheparanase failedto undergo reconstitution by heparanase-deficient bonemarrow and died within 5–6 weeks posttransplantation(data not shown).
Collectively, our results rule out a general role of hep-aranase in T-cell and neutrophil invasion and diapedesisthrough prototypic blood vessels despite the high HScontent of the basement membranes surrounding thesevessels. These surprising data contrast the contribution ofthisenzyme to invasionofDCsacross lymphatic vessels (27)and of inflammatory monocytes across peritoneal vascularand epithelial barriers (this study). Future studies shouldaim at further dissection of alternativemachineries used byneutrophils and T cells to cross vascular basement mem-branes as well as other ECMbarriers. These leukocytesmayuse alternative barrier-crossing strategies apparently notused by mesenchymal cells, DCs, and monocytes for cross-ing various basement membranes. Indeed, the heparanaseindependence of T-cell and neutrophil invasion revealedby us can be explained by the exceptional ability of theseleukocytes to identify and squeeze through basementmembranediscontinuities without an apparent need touseany proteolytic or GAG-degrading enzymatic machineriesto degrade these ECM-enriched barriers (1, 56).
The authors thank Dr. Jin-Ping Li (Department of MedicalBiochemistry and Microbiology, University of Uppsala, Uppsala,Sweden) for generating the heparanase-knockout mice. Theauthors also thank Yochai Wolf for intellectual assistance. R.A. isan incumbent of the Linda Jacobs Chair in Immune and Stem
Figure 8. Heparanase overexpression in neutro-phils does not increase their entry into zymosan-inflamed peritoneum of chimeric mice. A) WTor hpa-tg chimeric mice were bled and injectedwith zymosan or control PBS. At 2 hours post-injection, peritoneal lavage was extracted forneutrophil FACS analysis. B) Number of perito-neal neutrophils. Values are the mean 6 SEM of2–10 mice. The right inset shows blood FACSanalysis for circulating neutrophils in either WT orhpa-tg chimeric mice. Values are the mean 6 SEM
of 7–8 mice. N.S. nonsignificant. **P , 0.008, byunpaired Student’s t test.
10 Vol. 29 May 2015 STOLER-BARAK ET AL.The FASEB Journal x www.fasebj.org

Cell Research, and his research is supported by the German-Israeli Foundation, the Israel Science Foundation, the U.S.-IsraelBinational Science Foundation, the Flight Attendant MedicalResearch Institute Foundation, and the Minerva Foundation,Germany. The authors declare no conflicts of interest.
REFERENCES
1. Nourshargh, S., Hordijk, P. L., and Sixt, M. (2010) Breachingmultiple barriers: leukocyte motility through venular walls andthe interstitium. Nat. Rev. Mol. Cell Biol. 11, 366–378
2. Parish, C. R. (2006) The role of heparan sulphate ininflammation. Nat. Rev. Immunol. 6, 633–643
3. Parish, C. R., Hindmarsh, E. J., Bartlett, M. R., Staykova, M. A.,Cowden, W. B., and Willenborg, D. O. (1998) Treatment ofcentral nervous system inflammation with inhibitors of basementmembrane degradation. Immunol. Cell Biol. 76, 104–113
4. Bishop, J. R., Schuksz, M., and Esko, J. D. (2007) Heparansulphate proteoglycans fine-tune mammalian physiology. Nature446, 1030–1037
5. Simon Davis, D. A., and Parish, C. R. (2013) Heparan sulfate:a ubiquitous glycosaminoglycan with multiple roles in immunity.Front. Immunol. 4, 470
6. Stoler-Barak, L., Moussion, C., Shezen, E., Hatzav, M., Sixt, M.,and Alon, R. (2014) Blood vessels pattern heparan sulfategradients between their apical and basolateral aspects. PLoS One9, e85699
7. Morimoto-Tomita, M., Uchimura, K., Werb, Z., Hemmerich, S.,and Rosen, S. D. (2002) Cloning and characterization of twoextracellular heparin-degrading endosulfatases in mice andhumans. J. Biol. Chem. 277, 49175–49185
8. Sanderson, R. D., Yang, Y., Kelly, T., MacLeod, V., Dai, Y., andTheus, A. (2005) Enzymatic remodeling of heparan sulfateproteoglycans within the tumor microenvironment: growthregulation and the prospect of new cancer therapies. J. Cell.Biochem. 96, 897–905
9. Vlodavsky, I., Iozzo, R. V., and Sanderson, R. D. (2013)Heparanase: multiple functions in inflammation, diabetes andatherosclerosis. Matrix Biol. 32, 220–222
10. Elkin, M., Ilan, N., Ishai-Michaeli, R., Friedmann, Y., Papo, O.,Pecker, I., and Vlodavsky, I. (2001) Heparanase as mediator ofangiogenesis: mode of action. FASEB J. 15, 1661–1663
11. Zcharia, E., Zilka, R., Yaar, A., Yacoby-Zeevi, O., Zetser, A.,Metzger, S., Sarid, R., Naggi, A., Casu, B., Ilan, N., Vlodavsky, I.,and Abramovitch, R. (2005) Heparanase accelerates woundangiogenesis and wound healing in mouse and rat models.FASEB J. 19, 211–221
12. Arvatz, G., Shafat, I., Levy-Adam, F., Ilan, N., and Vlodavsky, I.(2011) The heparanase system and tumor metastasis: isheparanase the seed and soil? Cancer Metastasis Rev. 30, 253–268
13. Lerner, I., Hermano, E., Zcharia, E., Rodkin, D., Bulvik, R.,Doviner, V., Rubinstein, A. M., Ishai-Michaeli, R., Atzmon, R.,Sherman, Y., Meirovitz, A., Peretz, T., Vlodavsky, I., and Elkin, M.(2011) Heparanase powers a chronic inflammatory circuit thatpromotes colitis-associated tumorigenesis in mice. J. Clin. Invest.121, 1709–1721
14. Hermano, E., Lerner, I., and Elkin, M. (2012) Heparanaseenzyme in chronic inflammatory bowel disease and coloncancer. Cell. Mol. Life Sci. 69, 2501–2513
15. Goldshmidt, O., Zcharia, E., Abramovitch, R., Metzger, S.,Aingorn, H., Friedmann, Y., Schirrmacher, V., Mitrani, E., andVlodavsky, I. (2002) Cell surface expression and secretion ofheparanase markedly promote tumor angiogenesis andmetastasis. Proc. Natl. Acad. Sci. USA 99, 10031–10036
16. Vlodavsky, I., Goldshmidt, O., Zcharia, E., Atzmon, R.,Rangini-Guatta, Z., Elkin, M., Peretz, T., and Friedmann, Y.(2002) Mammalian heparanase: involvement in cancer metastasis,angiogenesis and normal development. Semin. Cancer Biol. 12,121–129
17. Ilan, N., Elkin, M., and Vlodavsky, I. (2006) Regulation, functionand clinical significance of heparanase in cancer metastasis andangiogenesis. Int. J. Biochem. Cell Biol. 38, 2018–2039
18. Nadir, Y., and Brenner, B. (2014) Heparanase multiple effects incancer. Thromb. Res. 133(Suppl 2), S90–S94
19. Matzner, Y., Bar-Ner, M., Yahalom, J., Ishai-Michaeli, R., Fuks, Z.,and Vlodavsky, I. (1985) Degradation of heparan sulfate in thesubendothelial extracellular matrix by a readily releasedheparanase from human neutrophils. Possible role in invasionthrough basement membranes. J. Clin. Invest. 76, 1306–1313
20. Vlodavsky, I., Eldor, A., Haimovitz-Friedman, A., Matzner, Y.,Ishai-Michaeli, R., Lider, O., Naparstek, Y., Cohen, I. R., andFuks, Z. (1992) Expression of heparanase by platelets andcirculating cells of the immune system: possible involvement indiapedesis and extravasation. Invasion Metastasis 12, 112–127
21. Naparstek, Y., Cohen, I. R., Fuks, Z., and Vlodavsky, I. (1984)Activated T lymphocytes produce a matrix-degrading heparansulphate endoglycosidase. Nature 310, 241–244
22. Fridman, R., Lider, O., Naparstek, Y., Fuks, Z., Vlodavsky, I., andCohen, I. R. (1987) Soluble antigen induces T lymphocytes tosecrete an endoglycosidase that degrades the heparan sulfate moietyof subendothelial extracellular matrix. J. Cell. Physiol. 130, 85–92
23. Bartlett, M. R., Underwood, P. A., and Parish, C. R. (1995)Comparative analysis of the ability of leucocytes, endothelial cellsand platelets to degrade the subendothelial basement membrane:evidence for cytokine dependence and detection of a novelsulfatase. Immunol. Cell Biol. 73, 113–124
24. Sewell, R. F., Brenchley, P. E., and Mallick, N. P. (1989) Humanmononuclear cells contain an endoglycosidase specific forheparan sulphate glycosaminoglycan demonstrable with the useof a specific solid-phase metabolically radiolabelled substrate.Biochem. J. 264, 777–783
25. de Mestre, A. M., Staykova, M. A., Hornby, J. R., Willenborg,D. O., and Hulett, M. D. (2007) Expression of the heparansulfate-degrading enzyme heparanase is induced in infiltratingCD4+ T cells in experimental autoimmune encephalomyelitisand regulated at the level of transcription by early growth re-sponse gene 1. J. Leukoc. Biol. 82, 1289–1300
26. Benhamron, S., Nechushtan, H., Verbovetski, I., Krispin, A.,Abboud-Jarrous, G., Zcharia, E., Edovitsky, E., Nahari, E., Peretz,T., Vlodavsky, I., and Mevorach, D. (2006) Translocation of activeheparanase to cell surface regulates degradation of extracellularmatrix heparan sulfate upon transmigration of mature monocyte-derived dendritic cells. J. Immunol. 176, 6417–6424
27. Benhamron, S., Reiner, I., Zcharia, E., Atallah, M., Grau, A.,Vlodavsky, I., and Mevorach, D. (2012) Dissociation betweenmature phenotype and impaired transmigration in dendriticcells from heparanase-deficient mice. PLoS One 7, e35602
28. Poon, I. K., Goodall, K. J., Phipps, S., Chow, J. D., Pagler, E. B.,Andrews, D. M., Conlan, C. L., Ryan, G. F., White, J. A., Wong,M. K., Horan, C., Matthaei, K. I., Smyth, M. J., and Hulett, M. D.(2014) Mice deficient in heparanase exhibit impaired dendriticcell migration and reduced airway inflammation. Eur. J. Immunol.44, 1016–1030
29. Zcharia, E., Jia, J., Zhang, X., Baraz, L., Lindahl, U., Peretz, T.,Vlodavsky, I., and Li, J. P. (2009) Newly generated heparanaseknock-out mice unravel co-regulation of heparanase and matrixmetalloproteinases. PLoS One 4, e5181
30. Zcharia, E., Metzger, S., Chajek-Shaul, T., Aingorn, H., Elkin, M.,Friedmann, Y., Weinstein, T., Li, J. P., Lindahl, U., and Vlodavsky,I. (2004) Transgenic expression of mammalian heparanaseuncovers physiological functions of heparan sulfate in tissuemorphogenesis, vascularization, and feeding behavior. FASEB J.18, 252–263
31. Boscacci, R. T., Pfeiffer, F., Gollmer, K., Sevilla, A. I., Martin,A. M., Soriano, S. F., Natale, D., Henrickson, S., von Andrian,U. H., Fukui, Y., Mellado, M., Deutsch, U., Engelhardt, B., andStein, J. V. (2010) Comprehensive analysis of lymph node stroma-expressed Ig superfamily members reveals redundant and non-redundant roles for ICAM-1, ICAM-2, and VCAM-1 in lymphocytehoming. Blood 116, 915–925
32. Shulman, Z., Cohen, S. J., Roediger, B., Kalchenko, V., Jain, R.,Grabovsky, V., Klein, E., Shinder, V., Stoler-Barak, L., Feigelson,S. W., Meshel, T., Nurmi, S. M., Goldstein, I., Hartley, O.,Gahmberg, C. G., Etzioni, A., Weninger, W., Ben-Baruch, A., andAlon, R. (2011) Transendothelial migration of lymphocytesmediated by intraendothelial vesicle stores rather than byextracellular chemokine depots. Nat. Immunol. 13, 67–76
33. Zhang, J., Dong, Z., Zhou, R., Luo, D., Wei, H., and Tian, Z.(2005) Isolation of lymphocytes and their innate immunecharacterizations from liver, intestine, lung and uterus. Cell.Mol. Immunol. 2, 271–280
HEPARANASE IS DISPENSABLE FOR LEUKOCYTE EMIGRATION 11

34. Cash, J. L., White, G. E., and Greaves, D. R. (2009) Chapter 17.Zymosan-induced peritonitis as a simple experimental system forthe study of inflammation. Methods Enzymol. 461, 379–396
35. Vlodavsky, I., Friedmann, Y., Elkin, M., Aingorn, H., Atzmon, R.,Ishai-Michaeli, R., Bitan, M., Pappo, O., Peretz, T., Michal, I.,Spector, L., and Pecker, I. (1999) Mammalian heparanase: genecloning, expression and function in tumor progression andmetastasis. Nat. Med. 5, 793–802
36. Cohen, S. J., Gurevich, I., Feigelson, S. W., Petrovich, E., Moser,M., Shakhar, G., Fassler, R., and Alon, R. (2013) The integrincoactivator Kindlin-3 is not required for lymphocyte diapedesis.Blood 122, 2609–2617
37. Guarda, G., Hons, M., Soriano, S. F., Huang, A. Y., Polley, R.,Martın-Fontecha, A., Stein, J. V., Germain, R. N., Lanzavecchia,A., and Sallusto, F. (2007) L-selectin-negative CCR7—effectorand memory CD8+ T cells enter reactive lymph nodes and killdendritic cells. Nat. Immunol. 8, 743–752
38. Martın-Fontecha, A., Baumjohann, D., Guarda, G., Reboldi, A.,Hons, M., Lanzavecchia, A., and Sallusto, F. (2008) CD40L+ CD4+memory T cells migrate in a CD62P-dependent fashion intoreactive lymph nodes and license dendritic cells for T cellpriming. J. Exp. Med. 205, 2561–2574
39. Shakhar, G., Lindquist, R. L., Skokos, D., Dudziak, D., Huang,J. H., Nussenzweig, M. C., and Dustin, M. L. (2005) Stable T cell-dendritic cell interactions precede the development of bothtolerance and immunity in vivo. Nat. Immunol. 6, 707–714
40. Kim, S. H., Turnbull, J., and Guimond, S. (2011) Extracellularmatrix and cell signalling: the dynamic cooperation of integrin,proteoglycan and growth factor receptor. J. Endocrinol. 209,139–151
41. Maletto, B. A., Ropolo, A. S., Alignani, D. O., Liscovsky, M. V.,Ranocchia, R. P., Moron, V. G., and Pistoresi-Palencia, M. C.(2006) Presence of neutrophil-bearing antigen in lymphoidorgans of immune mice. Blood 108, 3094–3102
42. Cunha, T. M., and Verri, Jr., W. A. (2006) Neutrophils: are theyhyperalgesic or anti-hyperalgesic? J. Leukoc. Biol. 80, 727–728,author reply 729–730
43. Mulligan, M. S., Varani, J., Warren, J. S., Till, G. O., Smith, C. W.,Anderson, D. C., Todd III, R. F., and Ward, P. A. (1992) Roles ofbeta 2 integrins of rat neutrophils in complement- and oxygenradical-mediated acute inflammatory injury. J. Immunol. 148,1847–1857
44. Sasaki, N., Higashi, N., Taka, T., Nakajima, M., and Irimura, T.(2004) Cell surface localization of heparanase on macrophagesregulates degradation of extracellular matrix heparan sulfate.J. Immunol. 172, 3830–3835
45. Lu, C., Li, X. Y., Hu, Y., Rowe, R. G., and Weiss, S. J. (2010) MT1-MMP controls human mesenchymal stem cell trafficking anddifferentiation. Blood 115, 221–229
46. Edovitsky, E., Elkin, M., Zcharia, E., Peretz, T., and Vlodavsky, I.(2004) Heparanase gene silencing, tumor invasiveness, angiogenesis,and metastasis. J. Natl. Cancer Inst. 96, 1219–1230
47. Lider, O., Mekori, Y. A., Miller, T., Bar-Tana, R., Vlodavsky, I.,Baharav, E., Cohen, I. R., and Naparstek, Y. (1990) Inhibition ofT lymphocyte heparanase by heparin prevents T cell migrationand T cell-mediated immunity. Eur. J. Immunol. 20, 493–499
48. Bernfield, M., Gotte, M., Park, P. W., Reizes, O., Fitzgerald, M. L.,Lincecum, J., and Zako, M. (1999) Functions of cell surfaceheparan sulfate proteoglycans. Annu. Rev. Biochem. 68, 729–777
49. He, Y. Q., Sutcliffe, E. L., Bunting, K. L., Li, J., Goodall, K. J.,Poon, I. K., Hulett, M. D., Freeman, C., Zafar, A., McInnes, R. L.,Taya, T., Parish, C. R., and Rao, S. (2012) The endoglycosidaseheparanase enters the nucleus of T lymphocytes and modulatesH3 methylation at actively transcribed genes via the interplaywith key chromatin modifying enzymes. Transcription 3, 130–145
50. Spiegel, A., Zcharia, E., Vagima, Y., Itkin, T., Kalinkovich, A., Dar, A.,Kollet, O., Netzer, N., Golan, K., Shafat, I., Ilan, N., Nagler, A.,Vlodavsky, I., and Lapidot, T. (2008) Heparanase regulates retentionand proliferation of primitive Sca-1+/c-Kit+/Lin- cells via modula-tion of the bone marrow microenvironment. Blood 111, 4934–4943
51. Sotnikov, I., Hershkoviz, R., Grabovsky, V., Ilan, N., Cahalon, L.,Vlodavsky, I., Alon, R., and Lider, O. (2004) Enzymaticallyquiescent heparanase augments T cell interactions with VCAM-1and extracellular matrix components under versatile dynamiccontexts. J. Immunol. 172, 5185–5193
52. Abboud-Jarrous, G., Atzmon, R., Peretz, T., Palermo, C., Gadea,B. B., Joyce, J. A., and Vlodavsky, I. (2008) Cathepsin L is responsiblefor processing and activation of proheparanase through multiplecleavages of a linker segment. J. Biol. Chem. 283, 18167–18176
53. Parish, C. R., Freeman, C., and Hulett, M. D. (2001) Heparanase:a key enzyme involved in cell invasion. Biochim. Biophys. Acta1471, M99–M108
54. Lider, O., Cahalon, L., Gilat, D., Hershkoviz, R., Siegel, D.,Margalit, R., Shoseyov, O., and Cohen, I. R. (1995) A dis-accharide that inhibits tumor necrosis factor alpha is formed fromthe extracellular matrix by the enzyme heparanase. Proc. Natl. Acad.Sci. USA 92, 5037–5041
55. Goodall, K. J., Poon, I. K., Phipps, S., and Hulett, M. D. (2014)Soluble heparan sulfate fragments generated by heparanasetrigger the release of pro-inflammatory cytokines through TLR-4.PLoS One 9, e109596
56. Proebstl, D., Voisin, M. B., Woodfin, A., Whiteford, J., D’Acquisto,F., Jones, G. E., Rowe, D., and Nourshargh, S. (2012) Pericytessupport neutrophil subendothelial cell crawling and breaching ofvenular walls in vivo. J. Exp. Med. 209, 1219–1234
Received for publication October 3, 2014.Accepted for publication January 5, 2015.
12 Vol. 29 May 2015 STOLER-BARAK ET AL.The FASEB Journal x www.fasebj.org




















