
Wip1-Deficient Neutrophils Significantly Promote Intestinal Ischemia/Reperfusion Injury in Mice
9
Send Orders for Reprints to [email protected] 100 Current Molecular Medicine 2015, 15, 100-108 Wip1-Deficient Neutrophils Significantly Promote Intestinal Ischemia/Reperfusion Injury in Mice J. Du 1,§ , X. Shen 2,§ , Y. Zhao 3 , X. Hu 3 , B. Sun 3 , W. Guan *,2 , S. Li *,1 and Y. Zhao *,3 1 Department of General Surgery, General Hospital of Beijing Military Command, Beijing, China 2 Department of General Surgery, Affiliated Drum Tower Hospital of Nanjing University Medical School, Nanjing, China 3 State Key Laboratory of Biomembrane and Membrane Biotechnology, Institute of Zoology, Chinese Academy of Sciences, Beijing, China Abstract: Wip1 is a serine/threonine protein phosphatase which plays a critical role in neutrophil development and maturation. In the present study, we used a neutrophil- dependent model of intestinal ischemia/reperfusion (I/R) injury to identify the role of Wip1 in neutrophil function under the condition of oxidative stress and inflammation. Wip1- deficient mice displayed more severe intestinal I/R injury with increased infiltration of neutrophils and higher expression of chemokines like CXCL-1, CXCL-2 and CCL-2, as well as inflammatory cytokine like TNF-α and IL-17. Studies in Wip1KOWT full hematopoietic chimera mice showed that Wip1 intrinsically regulated the function of immune cells after intestinal I/R injury. Through adoptive transfer of neutrophils from WT mice or mice with deficiency of IL-17, IL-17/Wip1 or Wip1, we demonstrated that Wip1KO neutrophils produced more IL-17 and eventually led to more severe intestinal I/R injury. Thus, our findings identify Wip1 as an intrinsic negative regulator of neutrophil inflammation in intestinal I/R injury process. Keywords: Intestinal ischemia/reperfusion injury, neutrophils, IL-17A, Wip1. INTRODUCTION Intestinal ischemia/reperfusion (I/R) injury is a pathological condition which occurs after aortic surgery, liver and intestinal transplantation, and is characterized by an initial restriction of blood supply to intestine and the following restoration of perfusion and re- oxygenation [1]. Restoration of blood supply and re- oxygenation is frequently associated with an exacerbation of intestinal tissue injury and gut barrier dysfunction with increased bacterial translocation, local inflammatory response and infiltration of innate immune cells especially during the early phase of reperfusion [2]. Intestinal I/R injury can also cause systemic inflammatory responses that ultimately progress to multi-organ failure and possibly to death. *Address correspondence to these authors at the (Y. Zhao), Transplantation Biology Research Division, State Key Laboratory of Biomembrane and Membrane Biotechnology, Institute of Zoology, Chinese Academy of Sciences, Beichen West Road 1-5, Chaoyang District, Beijing 100101, China; Tel: (86) 10-64807302; Fax: (86) 10-64807313; E-mail: [email protected], (W. Guan) Department of General Surgery, Affiliated Drum Tower Hospital of Nanjing University Medical School, 321 Zhongshan Rd., Nanjing 210008, China; Tel: (86) 25-83105114; Fax: (86) 25-83304616; E-mail: [email protected] and (S. Li) Department of General Surgery, General Hospital of Beijing Military Command, No. 5 Nan Men Cang, Beijing 100700, China; Tel: (86) 10-66721188; Fax: (86) 10-66721188; E-mail: [email protected] § These authors contributed equally to this work as co-first authors. Neutrophils, which are one of the earliest innate immune cells recruited to the sites of infection and inflammatory response, are crucial for the pathophysiology of I/R injury [3]. Too many neutrophils promote uncontrolled inflammation and intestinal injury, and the depletion of redundant neutrophils is also implicated to ameliorate tissue injury in various I/R models including liver and heart I/R models [4-6]. It is also noteworthy that too few neutrophils may not allow for adequate tissue repair [2]. Thus, the accumulation of neutrophils after intestinal I/R should be tightly controlled to balance this. Although studies have implicated a role for complement C3a in mediating the mobilization of neutrophils [7] and chemokines such as KC and CXCL-2 in mediating their infiltration into intestine and lung [8], the precise molecular mechanisms that control the functions and migration of neutrophils after intestinal I/R are still needed to be settled. Wild-type p53-induced phosphatase 1 (Wip1, also called PP2Cδ) is a serine/threonine protein phosphatase belonging to the type 2Cδ protein phosphatases [9]. It is activated by various stresses and is involved in various cellular processes such as tumorigenesis and aging [10, 11]. Wip1 is also recognized as an oncogenic regulator of tumor repressor and DNA damage response by inhibiting several p53-dependent pathways, including ATM- CHK2–p53 and p38 mitogen-activated protein kinase (MAPK)–p53 pathways as well as NF-κB pathway [12- 1875-5666/15 $58.00+.00 © 2015 Bentham Science Publishers
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Wip1-Deficient Neutrophils Significantly Promote Intestinal Ischemia/Reperfusion Injury in Mice

Send Orders for Reprints to [email protected]
100 Current Molecular Medicine 2015, 15, 100-108
Wip1-Deficient Neutrophils Significantly Promote Intestinal Ischemia/Reperfusion Injury in Mice J. Du1,§, X. Shen2,§, Y. Zhao3, X. Hu3, B. Sun3, W. Guan*,2, S. Li*,1 and Y. Zhao*,3
1Department of General Surgery, General Hospital of Beijing Military Command, Beijing, China 2Department of General Surgery, Affiliated Drum Tower Hospital of Nanjing University Medical School, Nanjing, China 3State Key Laboratory of Biomembrane and Membrane Biotechnology, Institute of Zoology, Chinese Academy of Sciences, Beijing, China
Abstract: Wip1 is a serine/threonine protein phosphatase which plays a critical role in neutrophil development and maturation. In the present study, we used a neutrophil-dependent model of intestinal ischemia/reperfusion (I/R) injury to identify the role of Wip1 in neutrophil function under the condition of oxidative stress and inflammation. Wip1-deficient mice displayed more severe intestinal I/R injury with increased infiltration of neutrophils and higher expression of chemokines like CXCL-1, CXCL-2 and CCL-2, as well as inflammatory cytokine like TNF-α and IL-17. Studies in Wip1KOàWT full hematopoietic chimera mice showed that Wip1 intrinsically regulated the function of immune cells after intestinal I/R injury. Through adoptive transfer of neutrophils from WT mice or mice with deficiency of IL-17, IL-17/Wip1 or Wip1, we demonstrated that Wip1KO neutrophils produced more IL-17 and eventually led to more severe intestinal I/R injury. Thus, our findings identify Wip1 as an intrinsic negative regulator of neutrophil inflammation in intestinal I/R injury process.
Keywords: Intestinal ischemia/reperfusion injury, neutrophils, IL-17A, Wip1.
INTRODUCTION Intestinal ischemia/reperfusion (I/R) injury is a pathological condition which occurs after aortic surgery, liver and intestinal transplantation, and is characterized by an initial restriction of blood supply to intestine and the following restoration of perfusion and re-oxygenation [1]. Restoration of blood supply and re-oxygenation is frequently associated with an exacerbation of intestinal tissue injury and gut barrier dysfunction with increased bacterial translocation, local inflammatory response and infiltration of innate immune cells especially during the early phase of reperfusion [2]. Intestinal I/R injury can also cause systemic inflammatory responses that ultimately progress to multi-organ failure and possibly to death.
*Address correspondence to these authors at the (Y. Zhao), Transplantation Biology Research Division, State Key Laboratory of Biomembrane and Membrane Biotechnology, Institute of Zoology, Chinese Academy of Sciences, Beichen West Road 1-5, Chaoyang District, Beijing 100101, China; Tel: (86) 10-64807302; Fax: (86) 10-64807313; E-mail: [email protected], (W. Guan) Department of General Surgery, Affiliated Drum Tower Hospital of Nanjing University Medical School, 321 Zhongshan Rd., Nanjing 210008, China; Tel: (86) 25-83105114; Fax: (86) 25-83304616; E-mail: [email protected] and (S. Li) Department of General Surgery, General Hospital of Beijing Military Command, No. 5 Nan Men Cang, Beijing 100700, China; Tel: (86) 10-66721188; Fax: (86) 10-66721188; E-mail: [email protected] §These authors contributed equally to this work as co-first authors.
Neutrophils, which are one of the earliest innate immune cells recruited to the sites of infection and inflammatory response, are crucial for the pathophysiology of I/R injury [3]. Too many neutrophils promote uncontrolled inflammation and intestinal injury, and the depletion of redundant neutrophils is also implicated to ameliorate tissue injury in various I/R models including liver and heart I/R models [4-6]. It is also noteworthy that too few neutrophils may not allow for adequate tissue repair [2]. Thus, the accumulation of neutrophils after intestinal I/R should be tightly controlled to balance this. Although studies have implicated a role for complement C3a in mediating the mobilization of neutrophils [7] and chemokines such as KC and CXCL-2 in mediating their infiltration into intestine and lung [8], the precise molecular mechanisms that control the functions and migration of neutrophils after intestinal I/R are still needed to be settled. Wild-type p53-induced phosphatase 1 (Wip1, also called PP2Cδ) is a serine/threonine protein phosphatase belonging to the type 2Cδ protein phosphatases [9]. It is activated by various stresses and is involved in various cellular processes such as tumorigenesis and aging [10, 11]. Wip1 is also recognized as an oncogenic regulator of tumor repressor and DNA damage response by inhibiting several p53-dependent pathways, including ATM-CHK2–p53 and p38 mitogen-activated protein kinase (MAPK)–p53 pathways as well as NF-κB pathway [12-
1875-5666/15 $58.00+.00 © 2015 Bentham Science Publishers

Wip1 Down-Regulates Neutrophil Function Current Molecular Medicine, 2015, Vol. 15, No. 1 101
15]. Previously, we have shown that Wip1 negatively regulates granulocyte development and maturation [16]. The antibacterial and inflammatory function of neutrophils is also critically regulated by Wip1 as its deficiency in neutrophils contributed to the resistance to CXCL1-induced CXCR2 internalization and desensitization, and subsequently the enhanced migratory ability [17]. Since intestinal I/R injury is a pathophysiological process which includes oxidative stress and neutrophils-driven pathologies, we employed Wip1-deficient mice to investigate the role of Wip1 in intestinal I/R injury. We report that Wip1-deficient mice were more susceptible to intestinal I/R injury. Mechanistically, this worse I/R pathology resulted from the increased infiltration of neutrophils into intestine as depletion of neutrophils in Wip1KO mice ameliorated I/R injury significantly. Further studies showed that Wip1-deficient neutrophils displayed enhanced inflammatory activity through the increased expression of TNF-α and IL-17A in a cell-intrinsic manner, indicating that phosphatase Wip1 negatively masters neutrophil inflammatory response in intestinal I/R process.
MATERIALS AND METHODS Mice C57BL/6 (B6) and CD45.1+ B6 mice were purchased from Beijing University Experimental Animal Center (Beijing, China). Wip1 knock-out (Wip1KO) mice were kindly provided by the Key Laboratory of Human Diseases Comparative Medicine, the Ministry of Public Health (Beijing, China). Wip1KO mice have been described [12, 18] and backcrossed with B6 mice for more than 8 generations to the B6 genetic background in our lab. IL-17AKO (IL-17KO) mice were kindly provided by Prof. Zhinan Yin, Nankai University (Tianjin, China) [19]. Wip1KO and IL-17KO mice were mated to obtain IL-17/Wip1 double KO (DKO) mice. All mice were bred and maintained in specific pathogen-free conditions. Eight-to-12 weeks old sex-matched littermates were mainly used for experiments unless otherwise noted. Complete chimeras were generated by transferring 1-2x107 bone marrow cells (BMCs) from either WT or Wip1KO mice (CD45.2+) into lethally irradiated synegeneic CD45.1+ WT mice [20]. Experimental protocols were approved by the Animal Ethics Committee of the Institute of Zoology, Beijing, China.
Mouse Model of Intestinal Ischemia/Reperfusion (I/R) Male mice were subjected to sham surgery or 30 min of superior mesenteric artery (SMA) ischemia. After unclamping of the SMA, 0.5 ml warm saline was administered i.p. into recipients. Sham-operated mice underwent identical abdominal manipulations as mice subjected to intestinal I/R (laparotomy, intestinal retraction and positioning). Small intestine tissues were collected after sham operation or intestinal I/R for analysis of tissue injury, inflammation, and cytokine expression.
To deplete CD11b+Gr1+ cells in vivo, 0.25 mg depleting anti–Gr-1 mAb (RB6-8C5) was administered i.p. into recipient mice 3 days before intestinal I/R protocol. Depletion of neutrophils was confirmed by staining for circulating neutrophils with anti-Ly6G and anti-CD11b mAbs and assayed by flow cytometry (FCM).
Histological Analysis of Intestine I/R Injury Small intestine (jejunum and ileum) was washed in ice-cold PBS and fixed overnight in 10% formalin. After automated dehydration through a graded alcohol series, tissues were embedded in paraffin, sectioned at 5 microns and stained with hematoxylin-eosin (H&E). Intestinal H&E sections were graded for intestinal I/R-induced mucosal injury using the chiu score by a pathologist blinded to the samples as previously described [21]. In each section, 50–100 villi were graded on a six-tiered scale, as previously described [21]. Briefly, a score of 0 was assigned to a normal villus; villi with tip distortion were scored as 1; villi lacking goblet cells and containing Guggenheims' spaces were scored as 2; villi with patchy disruption of the epithelial cells were scored as 3; villi with exposed but intact lamina propria (LP) and epithelial cell sloughing were assigned a score of 4; villi in which the lamina propria was exuding were scored as 5; and finally, villi displaying hemorrhage or denudation were scored as 6.
Assessment of Neutrophil Infiltration in Intestine After Intestinal I/R The isolation of Lamina Propria (LP) cells in small intestines was performed as previously described [22]. Briefly, small intestines were cut into pieces 0.5 cm in length and shaken at 250 rpm for 30 min at 37oC in Hanks’ balanced-salt solution (Life Technologies) supplemented with 5% (vol/vol) FBS (CellGro) containing 2 mM EDTA for two times. Cell suspensions were passed through a strainer and the remaining intestinal tissues was washed and shaken for 30 min at 37oC in RPMI 1640 plus 5% (vol/vol) FBS and type IV collagenase (1mg/ml; Sigma). Cell suspensions were collected and passed through a strainer and were further enriched by centrifugation at room temperature at 500 g for 20 min in 40%/70% Percoll (GE Healthcare) in RPMI-1640. All the cells were incubated with mAbs to CD45, CD11b and Ly6G and the neutrophils were assessed for the expression of these markers by FCM.
Immunohistochemistry Staining Neutrophil infiltration in the small intestine after I/R was also determined with immunohistochemistry as described previously with a mAb against PMN (NIMP-R14, Thermo) [4, 23]. A primary isotype control mAb (MCA1212, AbD Serotec) was used as a negative control in all experiments. Sham control and experimental intestinal tissues stained for neutrophils were scored for neutrophil infiltration [24]. Positive staining cells were counted in 20 fields at 200X and the average was calculated and plotted [24].

102 Current Molecular Medicine, 2015, Vol. 15, No. 1 Du et al.
mAbs and FCM For FCM analysis of surface markers, cells were stained with Abs in PBS containing 0.1% (wt/vol) BSA and 0.1% NaN3. Anti-CD45R/B220 (RA3-6B2), anti-CD11b (M1/70), anti-Gr-1 (RB6-8C5), anti-CD45.1 (A20), and anti-CD45.2 (104) were purchased from eBioscience. Anti-CD11b (M1/70) was purchased from BD Biosciences. Anti-CD45 mAb (30-F11) was from Biolegend. Intracellular IL-17A (TC11-18H10.1) expression was analyzed by FCM according to the Biolegend’s instructions. Cells were acquired on a FACS Calibur (Becton Dickinson, CA) or Beckman Coulter Epics XL, and data were analyzed with CellQuest software. Cell numbers of various populations were calculated by multiplication of the total cell number by the percentages of each individual population from the same mouse, followed by averaging.
Enzyme Linked Immunosorbent Assay (ELISA) for Intestinal IL-17A Small intestine IL-17A levels were measured with mouse specific IL-17A ELISA kit according to the manufacturer’s instructions (eBiosciences, San Diego, CA). Tissues were homogenized in ice-cold RIPA buffer (150 mM NaCl, 50 mM Tris-HCl, 1mM EDTA, and 1% Triton-X [pH 7.4]) and samples processed for mouse specific ELISA kits.
Isolation of Neutrophils from BMCs and Adoptively Transfer Neutrophils from bone marrow were isolated were fractionated on a Percoll gradient [16]. The enriched neutrophil fraction was recovered at the interface of 65% and 72% Percoll. The purity of the enriched neutrophils was >80%. Neutrophils were sorted by high speed cell sorter (MoFlo, Beckman Coulter) using anti-Gr1/CD11b mAbs for parallel experiments between control and Wip1KO mice. The purity of the sorted neutrophils was usually more than 98%. The sorted WT, Wip1KO, IL-17KO and IL-17/Wip1DKO PMNs (1X106 cells/mouse, in 500 µl PBS) were adoptively transferred to naïve IL-17KO mice by i.v. injection 1 hour prior to intestinal I/R surgery.
RNA and Protein Analysis Frozen tissue samples were homogenized in ice-cold lysis buffer containing 10 mM HEPES, 2 mM EDTA (pH 8), 5 mM DTT, 1 mM Pefabloc, and 1 tablet of mixture of proteinase inhibitors (Roche). Real-time PCR was performed with probe sets from Applied Biosystems and normalized against the endogenous control gene HPRT. The primers used in the present study were summarized in Table 1.
Statistical Analysis All data are presented as the mean+SD. Student’s unpaired t-test for comparison of means was used to
compare groups. A P value less than 0.05 was considered to be statistically significant. Table 1. Primers used for real-time PCR.
Primers Sequence (Sense/Antisense)
CXCL-2 5’-CCAAGGGTTGACTTCAAGAAC-3’ 5’-AGCGAGGCACATCAGGTACG-3’
CCL-2 5’-ACCTGCTGCTACTCATTCAC-3’ 5’-TTGAGGTGGTTGTGGAAAAG-3’
CXCL-1 5’-GCACCCAAACCGAAGTCATAG-3’ 5’-AGAAGCCAGCGTTCACCAGA-3’
ICAM-1 5’-GTCCGCTGTGCTTTGAGAA-3’ 5’-GGTCCTTGCCTACTTGC-3’
TNF-α 5'-TACTGAACTTCGGGGTGATTGGTCC-3' 5'-CAGCCTTGTCCCTTGAAGAGAACC-3'
IL-17A 5'-TCCAGAAGGCCCTCAGACTA-3' 5'-ACACCCACCAGCATCTTCTC-3'
HPRT 5'-AGTACAGCCCCAAAATGGTTAAG-3' 5'-CTTAGGCTTTGTATTTGGCTTTTC-3'
CXCL-1, Chemokine (C-X-C motif) ligand 1; CCL-2, Chemokine (C-C motif) ligand 2; CXCL-2, Chemokine (C-X-C motif) ligand 2; ICAM-1, intercellular adhesion molecule-1; TNF-α, tumor necrosis factor-alpha; IL-17, interleukin-17.
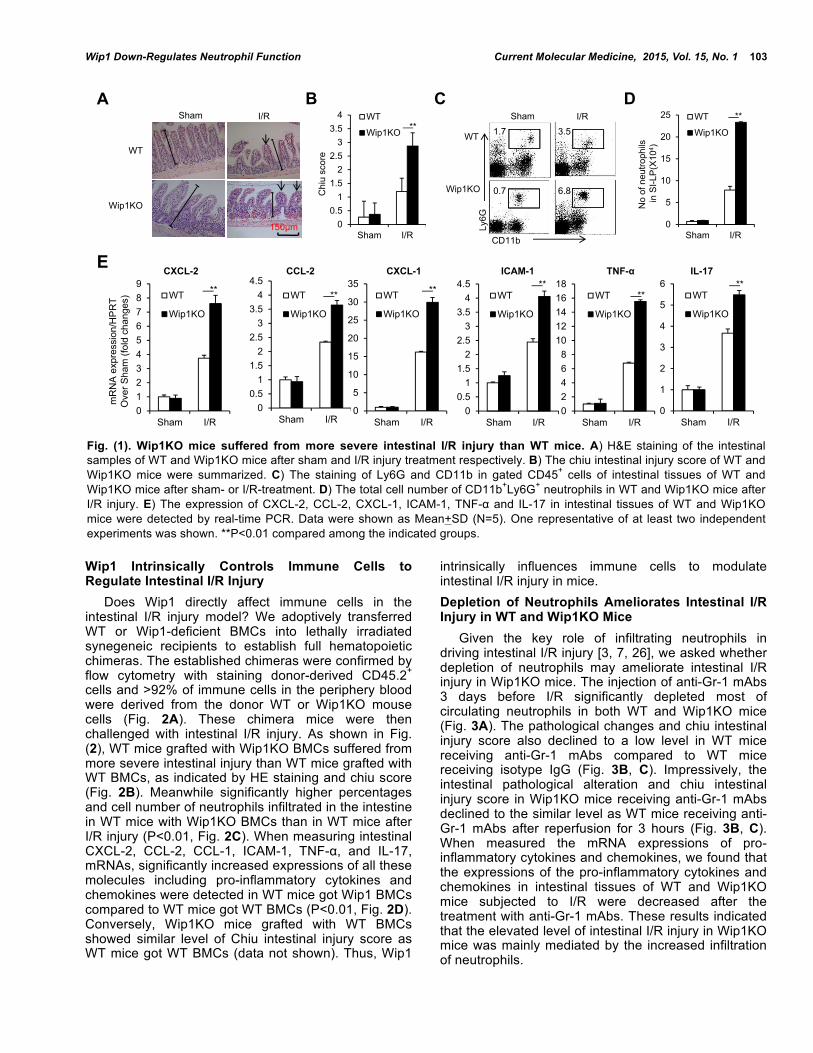
RESULTS Wip1-Deficient Mice Show Severe Intestinal I/R Injury We first determined whether Wip1KO mice were more susceptible to intestinal I/R injury. Wip1KO and WT mice were subjected to intestinal I/R, and the degree of intestinal injury after reperfusion for 3 hours were examined. As shown in Fig. (1), Wip1KO mice developed severe necrotic damage to the intestinal mucosa with sloughing of the villous tips and lifting of the epithelium from the LP, development of sub-epithelial Gruenhagen’s spaces (Fig. 1A). Chiu intestinal injury score in Wip1KO mice was also significantly higher than that in WT mice (P<0.01, Fig. 1B). It is also notable that there was higher percentage of neutrophils in SI-LP of Wip1KO mice (Fig. 1C), and the number of neutrophils was also significantly increased in Wip1KO mice than WT mice (P<0.01, Fig. 1D). Myeloperoxidase (MPO) is an abundant heme protein in neutrophils, with potent vascular inflammatory properties [25]. MPO levels were significantly elevated in WT mice subjected to I/R and increased further with Wip1 deficiency (data not shown). Next, we measured pro-inflammatory cytokines and chemokines mRNAs responsible for the infiltration of neutrophils and inflammatory damage in the intestine with real-time PCR. We found that the local expression of CXCL-2, CCL-2, CXCL-1, ICAM-1, TNF-α and IL-17 in I/R intestine were all significantly increased in Wip1KO mice than those in WT mice (P<0.01, Fig. 1E). These results collectively indicated that Wip1KO mice were more susceptible to intestinal I/R injury.
Wip1 Down-Regulates Neutrophil Function Current Molecular Medicine, 2015, Vol. 15, No. 1 103
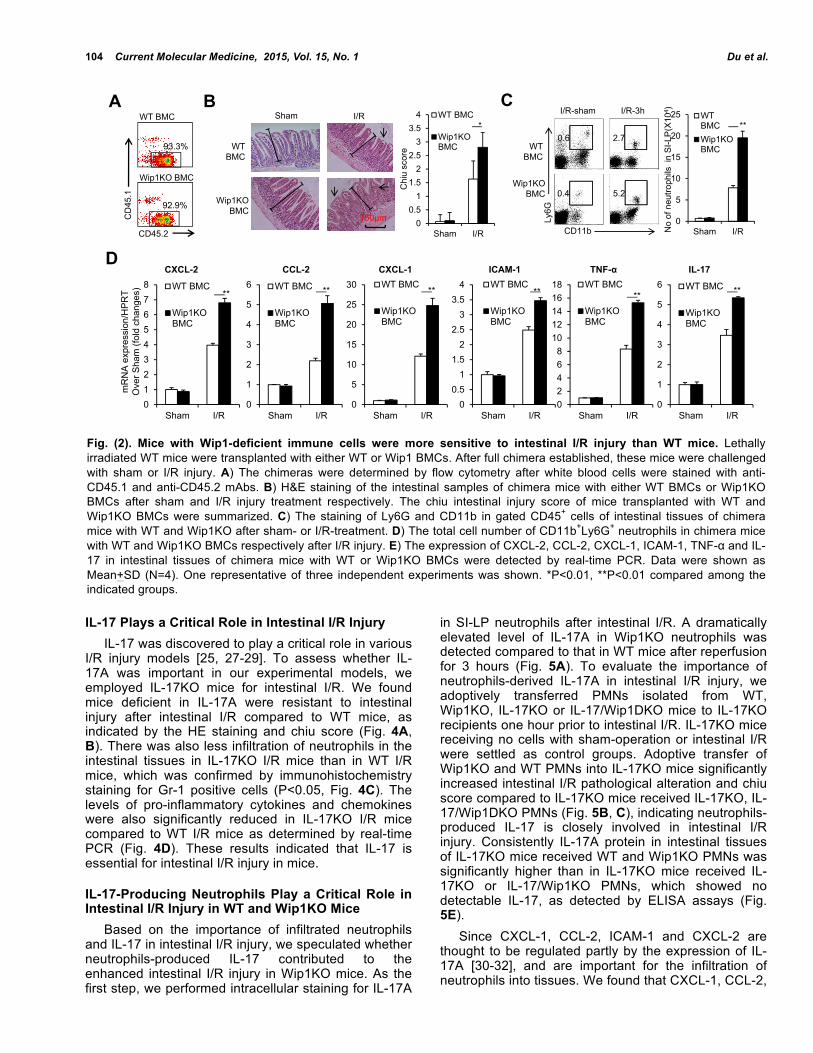
Wip1 Intrinsically Controls Immune Cells to Regulate Intestinal I/R Injury Does Wip1 directly affect immune cells in the intestinal I/R injury model? We adoptively transferred WT or Wip1-deficient BMCs into lethally irradiated synegeneic recipients to establish full hematopoietic chimeras. The established chimeras were confirmed by flow cytometry with staining donor-derived CD45.2+ cells and >92% of immune cells in the periphery blood were derived from the donor WT or Wip1KO mouse cells (Fig. 2A). These chimera mice were then challenged with intestinal I/R injury. As shown in Fig. (2), WT mice grafted with Wip1KO BMCs suffered from more severe intestinal injury than WT mice grafted with WT BMCs, as indicated by HE staining and chiu score (Fig. 2B). Meanwhile significantly higher percentages and cell number of neutrophils infiltrated in the intestine in WT mice with Wip1KO BMCs than in WT mice after I/R injury (P<0.01, Fig. 2C). When measuring intestinal CXCL-2, CCL-2, CCL-1, ICAM-1, TNF-α, and IL-17, mRNAs, significantly increased expressions of all these molecules including pro-inflammatory cytokines and chemokines were detected in WT mice got Wip1 BMCs compared to WT mice got WT BMCs (P<0.01, Fig. 2D). Conversely, Wip1KO mice grafted with WT BMCs showed similar level of Chiu intestinal injury score as WT mice got WT BMCs (data not shown). Thus, Wip1
intrinsically influences immune cells to modulate intestinal I/R injury in mice. Depletion of Neutrophils Ameliorates Intestinal I/R Injury in WT and Wip1KO Mice Given the key role of infiltrating neutrophils in driving intestinal I/R injury [3, 7, 26], we asked whether depletion of neutrophils may ameliorate intestinal I/R injury in Wip1KO mice. The injection of anti-Gr-1 mAbs 3 days before I/R significantly depleted most of circulating neutrophils in both WT and Wip1KO mice (Fig. 3A). The pathological changes and chiu intestinal injury score also declined to a low level in WT mice receiving anti-Gr-1 mAbs compared to WT mice receiving isotype IgG (Fig. 3B, C). Impressively, the intestinal pathological alteration and chiu intestinal injury score in Wip1KO mice receiving anti-Gr-1 mAbs declined to the similar level as WT mice receiving anti-Gr-1 mAbs after reperfusion for 3 hours (Fig. 3B, C). When measured the mRNA expressions of pro-inflammatory cytokines and chemokines, we found that the expressions of the pro-inflammatory cytokines and chemokines in intestinal tissues of WT and Wip1KO mice subjected to I/R were decreased after the treatment with anti-Gr-1 mAbs. These results indicated that the elevated level of intestinal I/R injury in Wip1KO mice was mainly mediated by the increased infiltration of neutrophils.
Fig. (1). Wip1KO mice suffered from more severe intestinal I/R injury than WT mice. A) H&E staining of the intestinal samples of WT and Wip1KO mice after sham and I/R injury treatment respectively. B) The chiu intestinal injury score of WT and Wip1KO mice were summarized. C) The staining of Ly6G and CD11b in gated CD45+ cells of intestinal tissues of WT and Wip1KO mice after sham- or I/R-treatment. D) The total cell number of CD11b+Ly6G+ neutrophils in WT and Wip1KO mice after I/R injury. E) The expression of CXCL-2, CCL-2, CXCL-1, ICAM-1, TNF-α and IL-17 in intestinal tissues of WT and Wip1KO mice were detected by real-time PCR. Data were shown as Mean+SD (N=5). One representative of at least two independent experiments was shown. **P<0.01 compared among the indicated groups.
WT
Wip1KO
Sham I/R
CD11b
Ly6G
1.7 3.5
6.80.7
No
of n
eutro
phils
in S
I-LP
(X10
4 )
A DCB
E
00.5
11.5
22.5
33.5
4
Sham I/R
WT
Wip1KO
0
5
10
15
20
25
Sham I/R
WT
Wip1KO
Chi
u sc
ore
0123456789
Sham I/R
WT
Wip1KO
00.5
11.5
22.5
33.5
44.5
Sham I/R
WT
Wip1KO
00.5
11.5
22.5
33.5
44.5
Sham I/R
WT
Wip1KO
02468
1012141618
Sham I/R
WT
Wip1KO
0
5
10
15
20
25
30
35
Sham I/R
WT
Wip1KO
0
1
2
3
4
5
6
Sham I/R
WT
Wip1KO
CXCL-2 CCL-2 CXCL-1 ICAM-1 TNF-α IL-17
mR
NA
exp
ress
ion/
HP
RT
Ove
r Sha
m (f
old
chan
ges)
Sham I/R
WT
Wip1KO
****
********** **
150μm
104 Current Molecular Medicine, 2015, Vol. 15, No. 1 Du et al.
IL-17 Plays a Critical Role in Intestinal I/R Injury IL-17 was discovered to play a critical role in various I/R injury models [25, 27-29]. To assess whether IL-17A was important in our experimental models, we employed IL-17KO mice for intestinal I/R. We found mice deficient in IL-17A were resistant to intestinal injury after intestinal I/R compared to WT mice, as indicated by the HE staining and chiu score (Fig. 4A, B). There was also less infiltration of neutrophils in the intestinal tissues in IL-17KO I/R mice than in WT I/R mice, which was confirmed by immunohistochemistry staining for Gr-1 positive cells (P<0.05, Fig. 4C). The levels of pro-inflammatory cytokines and chemokines were also significantly reduced in IL-17KO I/R mice compared to WT I/R mice as determined by real-time PCR (Fig. 4D). These results indicated that IL-17 is essential for intestinal I/R injury in mice.
IL-17-Producing Neutrophils Play a Critical Role in Intestinal I/R Injury in WT and Wip1KO Mice Based on the importance of infiltrated neutrophils and IL-17 in intestinal I/R injury, we speculated whether neutrophils-produced IL-17 contributed to the enhanced intestinal I/R injury in Wip1KO mice. As the first step, we performed intracellular staining for IL-17A
in SI-LP neutrophils after intestinal I/R. A dramatically elevated level of IL-17A in Wip1KO neutrophils was detected compared to that in WT mice after reperfusion for 3 hours (Fig. 5A). To evaluate the importance of neutrophils-derived IL-17A in intestinal I/R injury, we adoptively transferred PMNs isolated from WT, Wip1KO, IL-17KO or IL-17/Wip1DKO mice to IL-17KO recipients one hour prior to intestinal I/R. IL-17KO mice receiving no cells with sham-operation or intestinal I/R were settled as control groups. Adoptive transfer of Wip1KO and WT PMNs into IL-17KO mice significantly increased intestinal I/R pathological alteration and chiu score compared to IL-17KO mice received IL-17KO, IL-17/Wip1DKO PMNs (Fig. 5B, C), indicating neutrophils-produced IL-17 is closely involved in intestinal I/R injury. Consistently IL-17A protein in intestinal tissues of IL-17KO mice received WT and Wip1KO PMNs was significantly higher than in IL-17KO mice received IL-17KO or IL-17/Wip1KO PMNs, which showed no detectable IL-17, as detected by ELISA assays (Fig. 5E). Since CXCL-1, CCL-2, ICAM-1 and CXCL-2 are thought to be regulated partly by the expression of IL-17A [30-32], and are important for the infiltration of neutrophils into tissues. We found that CXCL-1, CCL-2,
Fig. (2). Mice with Wip1-deficient immune cells were more sensitive to intestinal I/R injury than WT mice. Lethally irradiated WT mice were transplanted with either WT or Wip1 BMCs. After full chimera established, these mice were challenged with sham or I/R injury. A) The chimeras were determined by flow cytometry after white blood cells were stained with anti-CD45.1 and anti-CD45.2 mAbs. B) H&E staining of the intestinal samples of chimera mice with either WT BMCs or Wip1KO BMCs after sham and I/R injury treatment respectively. The chiu intestinal injury score of mice transplanted with WT and Wip1KO BMCs were summarized. C) The staining of Ly6G and CD11b in gated CD45+ cells of intestinal tissues of chimera mice with WT and Wip1KO after sham- or I/R-treatment. D) The total cell number of CD11b+Ly6G+ neutrophils in chimera mice with WT and Wip1KO BMCs respectively after I/R injury. E) The expression of CXCL-2, CCL-2, CXCL-1, ICAM-1, TNF-α and IL-17 in intestinal tissues of chimera mice with WT or Wip1KO BMCs were detected by real-time PCR. Data were shown as Mean+SD (N=4). One representative of three independent experiments was shown. *P<0.01, **P<0.01 compared among the indicated groups.
WT BMC
Wip1KO BMC
I/R-sham I/R-3h
CD11b
Ly6G
0.6 2.7
0.4 5.2
012345678
Sham I/R
WT BMC
Wip1KOBMC
0
1
2
3
4
5
6
Sham I/R
WT BMC
Wip1KOBMC
0
5
10
15
20
25
30
Sham I/R
WT BMC
Wip1KOBMC
00.5
11.5
22.5
33.5
4
Sham I/R
WT BMC
Wip1KOBMC
02468
1012141618
Sham I/R
WT BMC
Wip1KOBMC
0
1
2
3
4
5
6
Sham I/R
WT BMC
Wip1KOBMC
CXCL-2 CCL-2 CXCL-1 ICAM-1 TNF-α IL-17
mR
NA
exp
ress
ion/
HP
RT
Ove
r Sha
m (f
old
chan
ges)
0
5
10
15
20
25
Sham I/R
WTBMCWip1KOBMC
No
of n
eutro
phils
in
SI-L
P(X
104 )
D
CB**
****** ** ** **
00.5
11.5
22.5
33.5
4
Sham I/R
WT BMC
Wip1KOBMC
Chi
u sc
ore
*Sham I/R
WT BMC
Wip1KOBMC
A
150μm
WT BMC
Wip1KO BMC
CD
45.1
CD45.2
93.3%
92.9%

Wip1 Down-Regulates Neutrophil Function Current Molecular Medicine, 2015, Vol. 15, No. 1 105
CXCL-2 and ICAM-1 mRNA levels were significantly higher in IL-17KO mouse recipients received Wip1KO PMNs than those in IL-17KO recipients received WT PMNs, while there were no significant differences among IL-17KO recipients received no cells, IL-17KO PMNs, and Wip1/IL-17DKO PMNs (Fig. 5F). In addition, more ICAM and TNF-α mRNA expression was detected in IL-17KO mice received Wip1KO PMNs than mice received WT PMNs (P<0.01, Fig. 5F). We also found there were higher MPO levels in intestinal tissues in IL-17KO recipients received WT or Wip1KO PMNs compared to IL-17KO mice received no cells, IL-17KO PMNs or WT PMNs (data not shown). Thus, these results suggested that Wip1KO neutrophils likely produced more IL-17 to promote the infiltration of neutrophils in the intestinal tissues, and eventually led to more severe intestinal injury after intestinal I/R.
DISCUSSION Wip1 has long been recognized as an oncogene in tumorigenesis because of its over-expression in human
tumors and a profound tumor-resistant phenotype of Wip1-dificient mice [33-35]. Recent years, Wip1 has also been identified to play an important role in several physiological processes including neurogenesis and aging [17, 33]. Our recent studies have demonstrated the importance of Wip1 in immune responses [16, 36]. Wip1 critically regulates granulocyte development and maturation [16], as well as the anti-bacteria and inflammatory function of neutrophils in mouse models [17]. In the present study we found that Wip1KO mice developed much more severe intestinal I/R injury after 30 min of SMA occlusion and 3 hr of reperfusion when compared to WT mice. Through establishing full hematopoietic chimeras, we further showed that Wip1 intrinsically regulate immune cells to impact intestinal I/R injury. Importantly, we provide evidence that Wip1KO mice produced more IL-17A after intestinal I/R, which drove more infiltration of neutrophils into intestinal tissues, and eventually led to more severe intestinal I/R injury in Wip1KO mice. I/R injury has long been recognized as sterile inflammation [4], while it is a completely different story
Fig. (3). Depletion of neutrophils significantly decreased intestinal I/R injury in Wip1KO mice. WT and Wip1KO mice were injected with depleting anti-Gr-1 mAb 3 days before I/R injury as described in materials and methods. A) Anti-Gr-1 mAb efficiently depleted CD11b+Ly6G+ cells in WT and Wip1KO mice. B) H&E staining of the intestinal samples of WT and Wip1KO mice after sham and I/R injury treatment respectively. C) The chiu intestinal injury score of WT and Wip1KO mice were summarized. D) The expression of CXCL-2, CCL-2, CXCL-1, ICAM-1, TNF-α and IL-17 in intestinal tissues of WT and Wip1KO mice were detected by real-time PCR. Data were shown as Mean+SD (N=4). One representative of two independent experiments was shown. *P<0.05, **P<0.01 compared among the indicated groups.
0123456789 Sham
I/R
0
1
2
3
4
5
6
7 ShamI/R
00.5
11.5
22.5
33.5
44.5 Sham
I/R
0
5
10
15
20
25 Sham
I/R
0
1
2
3
4
5
6
7 ShamI/R
0
5
10
15
20
25 Sham
I/R
CXCL-2 CCL-2 CXCL-1 ICAM-1 TNF-α IL-17
mR
NA
exp
ress
ion/
HP
RT
Ove
r Sha
m (f
old
chan
ges)
00.5
11.5
22.5
33.5 Sham
I/R
Chi
u sc
ore
B
D
A IgG Anti-Gr-1 mAb
CD11b
Ly6G
WT
Wip1KO
15.1
25.4
0.2
0.1
20.5
51.1
10.4
39.8 Sham I/R
WT
Wip1KO
CAnti-Gr-1 mAb pre-treated mice
** **
**
**
**
**
125μm

106 Current Molecular Medicine, 2015, Vol. 15, No. 1 Du et al.
when referred to intestinal I/R as there localized various microorganisms such as commensal bacteria and pathogens [37]. Bacteria translocation is a common feature of intestinal I/R injury which may enhance intestinal I/R injury. Hence, intestinal I/R injury is more complex than those in other organs and tissues. As depletion of neutrophils ameliorated intestinal I/R injury in both of WT and Wip1KO mice, better control of the infiltration of neutrophils may be of central importance for the process of intestinal I/R injury. Too many neutrophils may promote uncontrolled inflammation and damage intestinal tissues, while too few may not allow for adequate immunity against bacteria translocation. Indeed, we discovered increased infiltration of neutrophils in Wip1KO mice subjected to intestinal I/R when compared to WT mice, as well as more severe intestinal I/R injury with elevated levels of pro-inflammatory cytokines and chemokine such as TNF-α, CXCL-2, CCL-2, CCL-1 and IL-17. Our previous studies showed that Wip1KO neutrophils expressed high levels of CXCR2, the ligand for CXCL-1/CXCL-2, than WT cells. Thus, the higher expressions of CXCR2 in Wip1KO neutrophils would drive more neutrophils into intestinal I/R tissues. Similar results were obtained in full hematopoietic chimeras in which WT mice grafted with Wip1KO BMCs. Therefore, Wip1 intrinsically regulates neutrophil inflammation and
migration and subsequently impacts intestinal I/R injury in mice. IL-17A is identified to play a critical role in numerous I/R models through regulating cell apoptosis and the infiltration of granulocytes [25, 38, 39]. IL-17A is also suggested to stable chemokines mRNAs [40] and regulate the function of the potential inflammatory mediator matrix metalloproteinases 3 (MMP3) to promote I/R injury [25]. In the present study, we found that Wip1KO neutrophils produced more IL-17 than WT neutrophils. Adoptive transfer of Wip1KO neutrophils into IL-17KO mice caused significantly more severe I/R damage than transfer of WT neutrophils into IL-17KO mice. Importantly, adoptive transfer of IL-17/Wip1DKO neutrophils failed to do so in IL-17KO mice. Thus, the increased IL-17 production by Wip1KO neutrophils might contribute to the enhanced intestinal I/R injury in mice. The expression of IL-17 in neutrophils have been identified in various models such as fungal infection [41], liver I/R injury [4], and liver fibrosis [42]. IL-23 signal pathway was thought to be of central importance in regulating the expression of IL-17 in neutrophils. Using intestinal I/R models and Wip1KO mice, we showed that Wip1 negatively regulated the expression of IL-17 in neutrophils. Further studies will be needed to shed light on how Wip1 regulate the IL-17 expression in neutrophils.
Fig. (4). IL-17KO mice were more resistant to intestinal I/R injury than WT mice. A) H&E staining of the intestinal samples of WT and IL-17KO mice after sham and I/R injury treatment, respectively. B) The chiu intestinal injury score of WT and IL-17KO mice were summarized. C) The cell number of neutrophils in intestinal tissues of WT and Wip1KO mice after I/R injury. D) The expression of CXCL-2, CCL-2, CXCL-1, ICAM-1, and TNF-α in intestinal tissues of WT and IL-17KO mice were detected by real-time PCR. Data were shown as Mean+SD (N=4). One representative of at least two independent experiments was shown. *P<0.05, **P<0.01 compared between the indicated groups.
0
0.5
1
1.5
2
2.5
Sham I/R
WT
IL-17KO
02468
1012141618
Sham I/R
WT
IL-17KO
00.5
11.5
22.5
33.5
44.5
Sham I/R
WT
IL-17KO
00.5
11.5
22.5
33.5
44.5
Sham I/R
WT
IL-17KO
02468
1012141618
Sham I/R
WT
IL-17KO
0
0.5
1
1.5
2
2.5
Sham I/R
WT
IL-17KO
012345678
Sham I/R
WT
IL-17KO
CXCL-2 CCL-2 CXCL-1 ICAM-1 TNF-α
Sham I/R
IL-17KO
WT
Chu
sco
re
No
of in
flitra
ted
neut
roph
ils
mR
NA
exp
ress
ion/
HP
RT
Ove
r Sha
m (f
old
chan
ges)
D
A CB
**
**
**
**
**
***
125μm

Wip1 Down-Regulates Neutrophil Function Current Molecular Medicine, 2015, Vol. 15, No. 1 107
In summary, Wip1 deficiency in neutrophils caused more neutrophil infiltration and enhanced inflammatory cytokine production like TNF-α and IL-17 in intestinal I/R injury. In addition, the increased IL-17 expression in Wip1KO neutrophils might contribute to the enhanced intestinal I/R damage in Wip1KO mice. Nevertheless, the phosphatase Wip1 negatively regulates neutrophil inflammatory activation and migration in the intestinal I/R injury mouse model. Wip1, which is an intrinsic negative master for neutrophils, may be a promising target for therapeutic intervention to modulate neutrophil function on purpose.
CONFLICT OF INTEREST The authors confirm that this article content has no conflict of interest.
ACKNOWLEDGEMENTS The authors wish to thank Dr. Tao Yang for his kind review of the manuscript, Mrs. Jing Wang, Mr. Yabing Liu, and Mrs. Xiaoqiu Liu for their expert technical assistance, Mrs. Jianxia Peng for her excellent laboratory management, and Mr. Baisheng Ren for his outstanding animal husbandry. This work was supported by grants from the National Natural Science Foundation for General and Key Programs (No. 81000189, J.D.; C81130055, C81072396, Y.Z.), the National Basic Research Program of China (2010CB945301 and 2011CB710900, Y.Z.), and China Postdoctoral Science Foundation (No. 2014M552695).
REFERENCES [1] Hart ML, Jacobi B, Schittenhelm J, Henn M, Eltzschig HK.
Cutting Edge: A2B Adenosine receptor signaling provides potent
Fig. (5). Wip1-deficient neutrophils promote intestinal I/R injury through higher expression of IL-17A. A) Neutrophils were isolated from SI-LP of WT or Wip1KO I/R mice and were then treated with medium or PMA/Ionomycin for 5 hrs, the expression of IL-17 in neutrophils were then determined by intracellular staining FCM. Wip1KO neutrophils produced more IL-17 than WT neutrophils. (B-C) IL-17KO mice were i.v. injected with WT, Wip1KO, IL-17/Wip1DKO and IL-17KO PMNs respectively 1 hr before I/R injury. The intestinal pathological changes and cytokines were detected. B) H&E staining of the intestinal samples of IL-17KO mice with IL-17KO, WT, IL-17/Wip1DKO and Wip1KO PMNs after I/R injury were shown. C) The chiu intestinal injury score of IL-17KO mice with IL-17KO, WT, IL-17/Wip1DKO and Wip1KO PMNs were summarized. D) The cell number of neutrophils in the intestinal tissues of IL-17KO mice with IL-17KO, WT, IL-17/Wip1DKO and Wip1KO PMNs after I/R injury. E) The protein levels of IL-17A in the intestinal tissues of IL-17KO mice with IL-17KO, WT, IL-17/Wip1DKO and Wip1KO PMNs were detected by ELISA assays. F) The expression of CXCL-2, CCL-2, CXCL-1, ICAM-1 and TNF-α in the intestinal tissues of IL-17KO mice with IL-17KO, WT, IL-17/Wip1DKO and Wip1KO PMNs were detected by real-time PCR. Data were shown as Mean+SD (N=4). One representative of two independent experiments was shown. *P<0.05, **P<0.01 compared among the indicated groups.
0
0.5
1
1.5
2
2.5
3
3.5Sham
I/R
0
1
2
3
4
5
6Sham
I/R
0
0.5
1
1.5
2
2.5
3
3.5Sham
I/R
0
1
2
3
4
5
6
7
8Sham
I/R
0
2
4
6
8
10
12
14
16Sham
I/R
CXCL-2 CCL-2 CXCL-1 ICAM-1 TNF-α
Control Stimulated
WT
Wip1KO
Ly6G
IL-1
7A 0.5 1.3
1.0 7.2 0
5
10
15
20 Sham
I/Rm
RN
A e
xpre
ssio
n/H
PR
TO
ver S
ham
(fol
d ch
ange
s)
No
of n
eutro
phils
Wip1KOIL-17/Wip1 DKO
WT IL-17KO
0
0.5
1
1.5
2
2.5 Sham
IR
Chi
u sc
ore
F
A DCB
05
101520253035
IL-1
7A e
xpre
ssio
n (p
g/m
g pr
otei
n)
E**
** **
** ** **
* **
150μm

108 Current Molecular Medicine, 2015, Vol. 15, No. 1 Du et al.
protection during intestinal ischemia/reperfusion injury. J Immunol 2009; 182(7): 3965-8.
[2] Eltzschig HK, Eckle T. Ischemia and reperfusion--from mechanism to translation. Nat Med 2011; 17(11): 1391-401.
[3] Kubes P, Hunter J, Granger DN. Ischemia/reperfusion-induced feline intestinal dysfunction: importance of granulocyte recruitment. Gastroenterology 1992; 103(3): 807-12.
[4] Tan Z, Jiang R, Wang X, et al. RORgammat+IL-17+ neutrophils play a critical role in hepatic ischemia-reperfusion injury. J Mol Cell Biol 2013; 5(2): 143-6.
[5] Mantovani A, Cassatella MA, Costantini C, Jaillon S. Neutrophils in the activation and regulation of innate and adaptive immunity. Nat Rev Immunol 2011; 11(8): 519-31.
[6] Kohler D, Straub A, Weissmuller T, et al. Phosphorylation of vasodilator-stimulated phosphoprotein prevents platelet-neutrophil complex formation and dampens myocardial ischemia-reperfusion injury. Circulation 2011; 123(22): 2579-90.
[7] Wu MC, Brennan FH, Lynch JP, et al. The receptor for complement component C3a mediates protection from intestinal ischemia-reperfusion injuries by inhibiting neutrophil mobilization. Proc Natl Acad Sci U S A 2013; 110(23): 9439-44.
[8] Sadik CD, Kim ND, Luster AD. Neutrophils cascading their way to inflammation. Trends Immunol 2011; 32(10): 452-60.
[9] Fiscella M, Zhang HL, Fan SJ, et al. Wip1, a novel human protein phosphatase that is induced in response to ionizing radiation in a p53-dependent manner. Proc Natl Acad Sci U S A 1997; 94(12): 6048-53.
[10] Le Guezennec X, Bulavin DV. WIP1 phosphatase at the crossroads of cancer and aging. Trends Biochem Sci 2010; 35(2): 109-14.
[11] Salminen A, Kaarniranta K. Control of p53 and NF-kappaB signaling by WIP1 and MIF: role in cellular senescence and organismal aging. Cell Signal 2011; 23(5): 747-52.
[12] Chew J, Biswas S, Shreeram S, et al. WIP1 phosphatase is a negative regulator of NF-kappa B signalling. Nat Cell Biol 2009; 11(5): 659-66.
[13] Lu XB, Nguyen TA, Moon SH, Darlington Y, Sommer M, Donehower LA. The type 2C phosphatase Wip1: An oncogenic regulator of tumor suppressor and DNA damage response pathways. Cancer Metast Rev 2008; 27(2): 123-35.
[14] Oliva-Trastoy M, Berthonaud V, Chevalier A, et al. The Wip1 phosphatase (PPM1D) antagonizes activation of the Chk2 tumour suppressor kinase. Oncogene 2007; 26(10): 1449-58.
[15] Bulavin DV, Phillips C, Nannenga B, et al. Inactivation of the Wip1 phosphatase inhibits mammary tumorigenesis through p38 MAPK-mediated activation of the p16(Ink4a)-p19(Arf) pathway. Nat Genet 2004; 36(4): 343-50.
[16] Liu GW, Hu XL, Sun B, et al. Phosphatase Wip1 negatively regulates neutrophil development through p38 MAPK-STAT1. Blood 2013; 121(3): 519-29.
[17] Sun B, Hu X, Liu G, et al. Phosphatase Wip1 Negatively Regulates Neutrophil Migration and Inflammation. J Immunol 2014; 192(3): 1184-95.
[18] Choi J, Nannenga B, Demidov ON, et al. Mice deficient for the wild-type p53-induced phosphatase gene (Wip1) exhibit defects in reproductive organs, immune function, and cell cycle control. Mol Cell Biol 2002; 22(4): 1094-105.
[19] Zhao N, Hao J, Ni Y, et al. Vgamma4 gammadelta T cell-derived IL-17A negatively regulates NKT cell function in Con A-induced fulminant hepatitis. J Immunol 2011; 187(10): 5007-14.
[20] Liu G, Duan K, Ma H, Niu Z, Peng J, Zhao Y. An instructive role of donor macrophages in mixed chimeras in the induction of recipient CD4(+)Foxp3(+) Treg cells. Immunol Cell Biol 2011; 89(8): 827-35.
[21] Edgerton C, Crispin JC, Moratz CM, et al. IL-17 producing CD4+ T cells mediate accelerated ischemia/reperfusion-induced injury in autoimmunity-prone mice. Clin Immunol 2009; 130(3): 313-21.
[22] Denning TL, Wang YC, Patel SR, Williams IR, Pulendran B. Lamina propria macrophages and dendritic cells differentially induce regulatory and interleukin 17-producing T cell responses. Nat Immunol 2007; 8(10): 1086-94.
[23] Xiao H, Heeringa P, Liu Z, et al. The role of neutrophils in the induction of glomerulonephritis by anti-myeloperoxidase antibodies. Am J Pathol 2005; 167(1): 39-45.
[24] Lapchak PH, Ioannou A, Rani P, et al. The role of platelet factor 4 in local and remote tissue damage in a mouse model of mesenteric ischemia/reperfusion injury. PLoS One 2012; 7(7): e39934.
[25] Gelderblom M, Weymar A, Bernreuther C, et al. Neutralization of the IL-17 axis diminishes neutrophil invasion and protects from ischemic stroke. Blood 2012; 120(18): 3793-802.
[26] Grisham MB, Hernandez LA, Granger DN. Xanthine oxidase and neutrophil infiltration in intestinal ischemia. Am J Physiol 1986; 251(4 Pt 1): G567-74.
[27] Loi P, Yuan Q, Torres D, et al. Interferon regulatory factor 3 deficiency leads to interleukin-17-mediated liver ischemia-reperfusion injury. Hepatology 2013; 57(1): 351-61.
[28] Lee HT, Kim M, Kim JY, et al. Critical role of interleukin-17A in murine intestinal ischemia-reperfusion injury. Am J Physiol Gastrointest Liver Physiol 2013; 304(1): G12-25.
[29] Wang X, Sun R, Wei H, Tian Z. High-mobility group box 1 (HMGB1)-Toll-like receptor (TLR)4-interleukin (IL)-23-IL-17A axis in drug-induced damage-associated lethal hepatitis: Interaction of gammadelta T cells with macrophages. Hepatology 2013; 57(1): 373-84.
[30] Li L, Huang L, Vergis AL, et al. IL-17 produced by neutrophils regulates IFN-gamma-mediated neutrophil migration in mouse kidney ischemia-reperfusion injury. J Clin Invest 2010; 120(1): 331-42.
[31] Brackett CM, Muhitch JB, Evans SS, Gollnick SO. IL-17 promotes neutrophil entry into tumor-draining lymph nodes following induction of sterile inflammation. J Immunol 2013; 191(8): 4348-57.
[32] Park SW, Kim M, Brown KM, D'Agati VD, Lee HT. Paneth Cell-Derived Interleukin-17A Causes Multiorgan Dysfunction After Hepatic Ischemia and Reperfusion Injury. Hepatology 2011; 53(5): 1662-75.
[33] Zhu YH, Bulavin DV. Wip1-dependent signaling pathways in health and diseases. Prog Mol Biol Transl Sci 2012; 106: 307-25.
[34] Harrison M, Li J, Degenhardt Y, Hoey T, Powers S. Wip1-deficient mice are resistant to common cancer genes. Trends Mol Med 2004; 10(8): 359-61.
[35] Lu X, Nguyen TA, Moon SH, Darlington Y, Sommer M, Donehower LA. The type 2C phosphatase Wip1: an oncogenic regulator of tumor suppressor and DNA damage response pathways. Cancer Metastasis Rev 2008; 27(2): 123-35.
[36] Sun LN, Li HR, Luo HY, et al. Phosphatase Wip1 is Essential for the Maturation and Homeostasis of Medullary Thymic Epithelial Cells in Mice. J Immunol 2013; 191(6): 3210-20.
[37] Yoshiya K, Lapchak PH, Thai TH, et al. Depletion of gut commensal bacteria attenuates intestinal ischemia/reperfusion injury. Am J Physiol Gastrointest Liver Physiol 2011; 301(6): G1020-30.
[38] Liao YH, Xia N, Zhou SF, et al. Interleukin-17A contributes to myocardial ischemia/reperfusion injury by regulating cardiomyocyte apoptosis and neutrophil infiltration. J Am Coll Cardiol 2012; 59(4): 420-9.
[39] Park SW, Kim M, Kim JY, et al. Paneth cell-mediated multiorgan dysfunction after acute kidney injury. J Immunol 2012; 189(11): 5421-33.
[40] Hartupee J, Liu C, Novotny M, Li X, Hamilton T. IL-17 enhances chemokine gene expression through mRNA stabilization. J Immunol 2007; 179(6): 4135-41.
[41] Taylor PR, Roy S, Leal SM Jr, et al. Activation of neutrophils by autocrine IL-17A-IL-17RC interactions during fungal infection is regulated by IL-6, IL-23, RORgammat and dectin-2. Nat Immunol 2014; 15(2): 143-51.
[42] Tan ZM, Qian XF, Jiang RQ, et al. IL-17A Plays a Critical Role in the Pathogenesis of Liver Fibrosis through Hepatic Stellate Cell Activation. J Immunol 2013; 191(4): 1835-44.
Received: July 11, 2014 Revised: September 16, 2014 Accepted: October 01, 2014




















