
Wind-induced natural ventilation of re-entrant bays in a high-rise building

875Q 2001 Estuarine Research Federation
Estuaries Vol. 24, No. 6A, p. 875–883 December 2001
Harmful Algal Blooms in the Chesapeake and Coastal Bays of
Maryland, USA: Comparison of 1997, 1998, and 1999 Events
PATRICIA M. GLIBERT1,*, ROBERT MAGNIEN2, MICHAEL W. LOMAS1,†, JEFFREY ALEXANDER1,CHUNLEI FAN1, ERIN HARAMOTO1, MARK TRICE1, and TODD M. KANA1
1 University of Maryland Center for Environmental Science, Horn Point Laboratory,P. O. Box 775, Cambridge, Maryland 21613
2 Maryland Department of Natural Resources, 580 Taylor Avenue D-2,Annapolis, Maryland 21401
ABSTRACT: Harmful algal blooms in the Chesapeake Bay and coastal bays of Maryland, USA, are not a new phenom-enon, but may be increasing in frequency and diversity. Outbreaks of Pfiesteria piscicida (Dinophyceae) were observedduring 1997 in several Chesapeake Bay tributaries, while in 1998, Pfiesteria-related events were not found but massiveblooms of Prorocentrum minimum (Dinophyceae) occurred. In 1999, Aureococcus anophagefferens (Pelagophyceae) devel-oped in the coastal bays in early summer in sufficient densities to cause a brown tide. In 1997, toxic Pfiesteria wasresponsible for fish kills at relatively low cell densities. In 1998 and 1999, the blooms of P. minimum and A. anophagefferenswere not toxic, but reached sufficiently high densities to have ecological consequences. These years differed in theamount and timing of rainfall events and resulting nutrient loading from the largely agricultural watershed. Nutrientloading to the eastern tributaries of Chesapeake Bay has been increasing over the past decade. Much of this nutrientdelivery is in organic form. The sites of the Pfiesteria outbreaks ranked among those with the highest organic loading ofall sites monitored bay-wide. The availability of dissolved organic carbon and phosphorus were also higher at sitesexperiencing A. anophagefferens blooms than at those without blooms. The ability to supplement photosynthesis withgrazing or organic substrates and to use a diversity of organic nutrients may play a role in the development and main-tenance of these species. For P. minimum and A. anophagefferens, urea is used preferentially over nitrate. Pfiesteria is agrazer, but also has the ability to take up nutrients directly. The timing of nutrient delivery may also be of criticalimportance in determining the success of certain species.
IntroductionMuch has been written about the links between
nutrient loading and phytoplankton blooms in theChesapeake Bay, USA. Emphasis has been placedon understanding the links between freshwaterflow, nutrient loading largely in the form of NO3
2,and the development of spring diatom blooms(e.g., Boynton et al. 1982; Malone et al. 1983; Fish-er et al. 1988, 1992; Glibert et al. 1995). Theseblooms have been of interest largely because therich phytoplankton biomass which develops fromthe nutrient pulse fuels high rates of microbial me-tabolism which eventually leads to oxygen deple-tion below the pycnocline (Boynton et al 1982; Se-liger et al. 1985; Malone et al. 1988; Malone 1992).It has been well documented that following thespring diatom blooms there is typically a rapidtransition to a cyanobacterial and microflagellate-dominated community in summer, but summerblooms have received much less attention. We now
* Corresponding author; fax: 410/221-8490; e-mail: [email protected].
† Current address: Bermuda Biological Station for Research,17 Biological Lane, St. Georges GE01, Bermuda.
recognize that some of these late spring and earlysummer plankton communities may be composedof harmful algal bloom (HAB) species. We also rec-ognize the importance of the tributaries and coast-al bays as sites where such blooms may develop(e.g., Lewitus et al. 1995; Burkholder and Glasgow1997).
During the summers of 1997, 1998, and 1999,outbreaks of harmful blooms occurred in severaltributaries of the Chesapeake and coastal bays ofMaryland. Each year was characterized by a bloomof a different species in a different location, withdifferent environmental consequences. Duringsummer 1997, outbreaks of Pfiesteria piscicida, andassociated fish kills, occurred in several easternshore tributaries of Chesapeake Bay (Maryland De-partment of Natural Resources 1998). Outbreaksof P. piscicida do not develop into near monospe-cific blooms, but are highly toxic (Burkholder etal. 1995). In 1998, a massive early summer bloomof Prorocentrum minimum occurred, and a second-ary mid-summer bloom of the same species oc-curred throughout many tributaries of the Bay. In1999, significant dinoflagellate blooms were notcommon, but a large brown tide due to Aureococcus

876 P. M. Glibert et al.
Fig. 1. Map of Chesapeake and Coastal Bays of Maryland, USA. Inset maps show the Choptank River, site of the 1998 Prorocentrumminimum bloom, the Pocomoke River, site of the 1997 Pfiesteria piscicida outbreaks, and the coastal bays, site of the 1999 Aureococcusanophagefferens brown tide blooms.
anophagefferens occurred along the coastal bays.Blooms of P. minimum and A. anophagefferens aretypically non-toxic to invertebrates, but cause en-vironmental damage through the development ofhigh biomass and resulting loss of seagrasses andshellfish (Dennison et al. 1989; Wikfors and Smo-lowicz 1995; Bricelj and Lonsdale 1997).
We compare and contrast these 3 blooms interms of environmental conditions and the physi-ological adaptations of these species which maycontribute to their success under these environ-mental conditions. More complete analyses of eachof these blooms are available or forthcoming(Maryland Department of Natural Resources 1998,1999; Lomas et al. in press); here, we underscorethe susceptibility of Chesapeake Bay to future out-breaks and highlight key environmental parame-ters that may need to be monitored and under-stood to better predict these outbreaks.
Materials and Methods
The sites in Maryland waters with HAB out-breaks from 1997–1999 include several tributariesof the Chesapeake Bay and coastal bays (Fig. 1).Although several eastern shore tributaries were im-pacted by the Pfiesteria events in 1997, and the P.minimum events of 1998, only two will receive at-tention here: the Pocomoke River and the Chop-tank River. These tributaries drain largely agricul-tural lands with relatively little (, 5%) direct ni-trogen input from point sources (Maryland De-partment of Natural Resources 1998). From 1985–1996, nitrogen inputs to these rivers increased by24% and 54%, respectively (Maryland Departmentof Natural Resources 1998). The coastal bays areaffected by heavily developed seaside resort towns,inland agricultural and rural areas, and a nationalpark.
Samples in the Pocomoke River in 1997 were

Comparison of Harmful Blooms in the Chesapeake Bay 877
Fig. 2. Average flow of the Susquehanna River (n) and meanwater temperature (l) for 1997, 1998, and 1999 for the Ches-apeake Bay system. Gray areas indicate time periods of the re-spective HAB outbreaks.
collected as part of the Maryland Department ofNatural Resources routine monitoring programand their more intensive monitoring following thefirst reports of fish kills. Samples were collectedfrom just below the surface and transported in thedark, on ice, to the laboratory for filtration andsubsequent analysis. Water temperature, salinity,and light penetration were also recorded at eachsampling site. Protocols for sample collection weresimilar in 1999, but intensity of sampling was in-creased through the National Oceanic and Atmo-spheric Administration-sponsored Ecology andOceanography of Harmful Algal Blooms (ECO-HAB) program.
In the Choptank River in 1998, samples were col-lected using an acid-washed bucket from the dockof the Horn Point Laboratory. Samples were re-turned within minutes to the laboratory for filtra-tion and analysis. In the coastal bays during 1999,small boats, docks, and access ramps were used toobtain surface samples, using buckets or 1 liter bot-tles. Samples were transported to the laboratory assoon as possible and within 2 h.
All samples were filtered through precombustedGF/F filters and retained for chlorophyll and par-ticulate carbon and nitrogen analysis. Chlorophyllwas measured fluorometrically (Parsons et al.1984). Upon filtration, the filtrate was also re-tained for subsequent nutrient analyses. Inorganicnutrients (NO3
2, NO22, NH4
1, PO432) were ana-
lyzed using standard autoanalyzer methods. Sam-ples for total dissolved nitrogen (TDN) were ana-lyzed using persulfate oxidation (early summer1997) or an Antek Instruments high temperatureoxidation instrument, which has been intercali-brated with UV oxidation and persulfate oxidationmethods (Bronk et al. 2000). Concentrations ofdissolved organic nitrogen (DON) were calculatedby subtracting all dissolved inorganic nitrogen(DIN) concentrations from the TDN results. Con-centrations of dissolved organic carbon (DOC)were analyzed on a Shimadzu TOC instrument.
In the Choptank River, additional experimentson the rates of nitrogen uptake by the P. minimumdominated plankton assemblage were conducted.These experiments were done by the addition of15N-labeled nitrogen substrates, in concentrationswhich ranged from , 10% to . 50% of the am-bient substrate concentration, depending on theexperiment being conducted. The substrates usedalso varied with experiment and included NO3
2,NH4
1, and urea. All incubations were conductedin polypropylene tubes, for a duration of , 1 hunder simulated in situ irradiance and tempera-ture conditions. Isotopic analysis was done using aNuclide 30 608 sector mass spectrometer using pro-tocols described in Glibert et al. (1991).
Monthly flow data was obtained from U.S. Geo-logical Survey, and mean water column tempera-tures were from the database of the ChesapeakeBay Observing System.
Results and DiscussionFRESHWATER FLOW
The years from 1997–1999 had considerably dif-ferent freshwater flow conditions (Fig. 2). Thesprings of 1997 and 1998 were unusually wet,whereas the spring of 1999 was relatively dry.Freshwater flow in the spring of 1998 was 2–2.53higher than that during the comparable period of1999. Freshwater flow remained relatively highthrough the month of July in 1998, whereas in theother 2 years, the freshwater flow during July andAugust was , 700 m3 s21. The considerable fresh-

878 P. M. Glibert et al.
TABLE 1. Ranges of environmental parameters for the tribu-taries and coastal bays of Maryland during harmful algal bloomsfor the month and year indicated. na indicates not available.
Parameter
August 1997P. piscicida
outbreaks inPocomoke
River
May 1998P. minimum
outbreaks inChoptank
River
June 1999A. anopha-
gefferensoutbreaks incoastal bays
SalinityTemperature (8C)Chlorophyll (mg 121)DIN (mM)
2–925.0–27.018.0–30.0
,0.05–28.6
5.5–7.015.7–23.082.8–7651.11–4.19
24.4–31.520.0–28.019.0–44.00.88–2.44
DIP (mM)DOC (mM)DON (mM)DOP (mM)
0.33–1.67683–91739.0–43.00.57–0.73
,0.05–0.07375–87019.4–27.7
na
0.01–0.31316–45625.0–70.0*0.9–1.78
* For this sampling period, the mean DON was 26.55 6 2.14for 10 observations; only one sample had a concentration of 70.This value was not used in the estimates of DOC:DON ratios.
water flow in 1997 correlated with water tempera-tures that were below the long-term average,whereas the dry spring of 1999 resulted in earlysummer temperatures that were up to 58C warmerthan the other years (Fig. 2).
DESCRIPTIONS OF THE BLOOM EVENTS
The August 1997 P. piscicida outbreak in the Po-comoke River was identified following the obser-vation of dead and lesioned fish near the mouthof the river. Smaller fish kills and lesion events hadbeen reported from the prior fall and spring, butwere not positively confirmed to be a consequenceof Pfiesteria or Pfiesteria-like organisms. SubsequentPfiesteria-related fish kills were reported throughOctober 1997 in the Chicamicomico River andKings Creek, also eastern shore tributaries (Mary-land Department of Natural Resources 1998).
During the Pfiesteria outbreak, salinity was , 10and temperatures ranged from 25–278C (Table 1).Chlorophyll concentrations up to 30 mg l21 werenoted, composed of a mixed assemblage of flagel-lates and diatoms, with maximum concentrationscoincident with the largest fish kill event (Mary-land Department of Natural Resources 1999). Con-centrations of DIN and dissolved inorganic phos-phorus (DIP), while variable, were generally in arange that would support at least a doubling ofphytoplankton biomass, and were not consideredto be limiting. Organic nutrients were also elevat-ed, with DOC concentrations ranging from 683–917 mM, and DON concentrations ranging from39–43 mM (Table 1). It has been suggested that lowdissolved oxygen (data not shown) in the regionimmediately upstream of the fish kill may have pre-vented the migrating menhaden from moving up-stream, and thus they stayed near the mouth of theriver (Maryland Department of Natural Resources1998). Elevated chlorophyll, combined with low
oxygen upstream, along with sufficient nutrients,may have promoted the development of toxic P.piscicida.
Environmental conditions in 1998 were suffi-ciently different from those present in 1997 thatphytoplankton communities developed quite dif-ferently. With significantly higher flow, there was achange in when and where on the rivers nutrientand chlorophyll maxima appeared. By May 1998,phytoplankton populations in many tributaries,and especially the Choptank River, had becomedominated by P. minimum in near-monospecificblooms, with chlorophyll concentrations rangingfrom 82–765 mg l21 in a highly patchy distribution(Table 1). Salinity remained , 7, while tempera-tures during the duration of the bloom (;20 d)ranged from 15.7–238C (Table 1). During the peakof the bloom, both DIN and DIP were found atlevels that were insufficient to support this bio-mass. Organic nutrients were abundant with DOCconcentrations ranging from 375–870 mM andDON concentrations ranging from 19.4–27.7 mM(Table 1). The unusually low salinities (, 10) andsurface temperatures (, 238C) combined withavailability of organic nutrients may have helpedto support this bloom. Similar conditions werefound during a large P. minimum bloom in the KielFjord (Kimor et al. 1985) and in Malta (Fudge1977). In the Choptank River, the bloom dissipatedby late May, but a smaller bloom (data not shown)occurred in early July following additional rain.
In 1999, the relative lack of spring rain may haveled to environmental conditions that favored thedevelopment of A. anophagefferens in the coastalbays. By mid June, A. anophagefferens was present insufficiently high densities (100,000–300,000 cellsl21) at selected sites in the coastal bays to be con-sidered a brown tide. Development of brown tideshas previously been correlated with years of lowrainfall (LaRoche et al. 1997). While lack of rain-fall, and its resulting effects on the relative avail-ability of inorganic and organic nitrogen, has prov-en to be a successful predictor of brown tides inPeconic Bay, New York, it has not been as success-ful in predicting brown tides in adjacent bays(Gobler 1999; Lomas et al. in press), implying thatadditional factors must be important in determin-ing the yearly success of A. anophagefferens. Thereis considerable evidence suggesting a role for or-ganic nutrients in supporting brown tide blooms(Dzurica et al. 1989; Berg et al. 1997; Lomas et al.in press), and the environmental characteristics ofthe Maryland coastal bays would be consistent withthis requirement. Concentrations of DIN and DIPduring the brown tide bloom were relatively low(Table 1) and, as in the case of the P. minimumbloom, not sufficient to support the biomass of
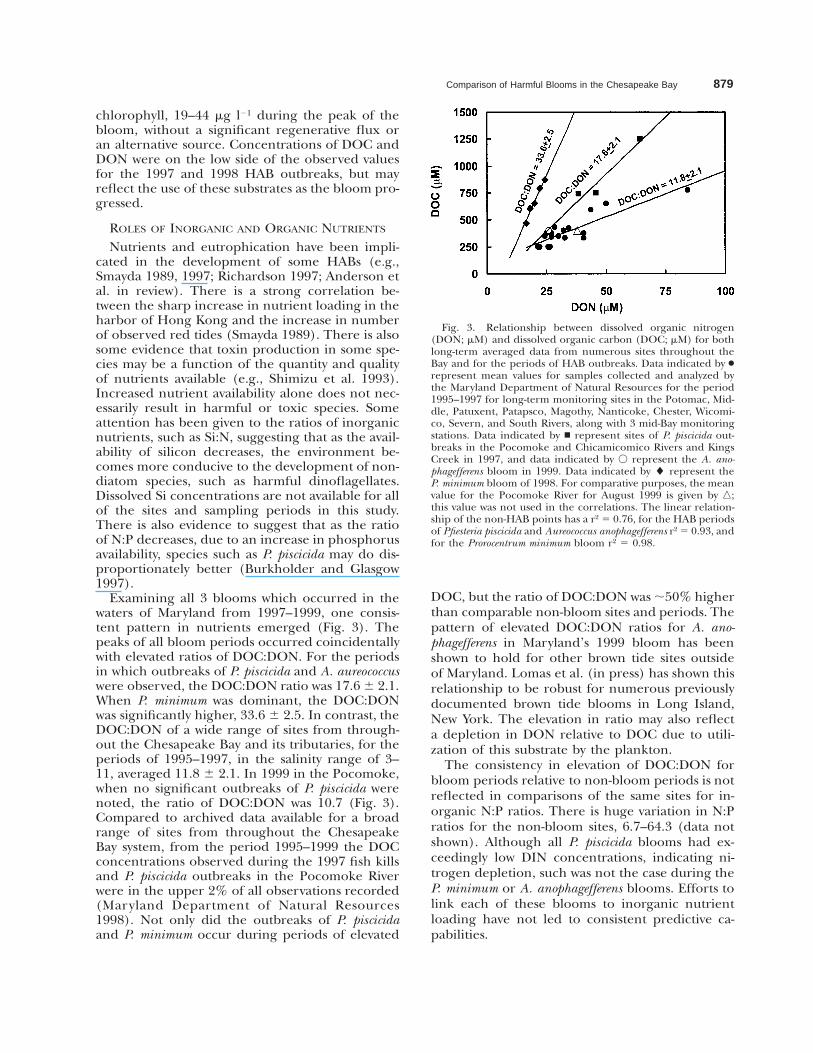
Comparison of Harmful Blooms in the Chesapeake Bay 879
Fig. 3. Relationship between dissolved organic nitrogen(DON; mM) and dissolved organic carbon (DOC; mM) for bothlong-term averaged data from numerous sites throughout theBay and for the periods of HAB outbreaks. Data indicated by ●represent mean values for samples collected and analyzed bythe Maryland Department of Natural Resources for the period1995–1997 for long-term monitoring sites in the Potomac, Mid-dle, Patuxent, Patapsco, Magothy, Nanticoke, Chester, Wicomi-co, Severn, and South Rivers, along with 3 mid-Bay monitoringstations. Data indicated by n represent sites of P. piscicida out-breaks in the Pocomoke and Chicamicomico Rivers and KingsCreek in 1997, and data indicated by V represent the A. ano-phagefferens bloom in 1999. Data indicated by l represent theP. minimum bloom of 1998. For comparative purposes, the meanvalue for the Pocomoke River for August 1999 is given by n;this value was not used in the correlations. The linear relation-ship of the non-HAB points has a r2 5 0.76, for the HAB periodsof Pfiesteria piscicida and Aureococcus anophagefferens r2 5 0.93, andfor the Prorocentrum minimum bloom r2 5 0.98.
chlorophyll, 19–44 mg l21 during the peak of thebloom, without a significant regenerative flux oran alternative source. Concentrations of DOC andDON were on the low side of the observed valuesfor the 1997 and 1998 HAB outbreaks, but mayreflect the use of these substrates as the bloom pro-gressed.
ROLES OF INORGANIC AND ORGANIC NUTRIENTS
Nutrients and eutrophication have been impli-cated in the development of some HABs (e.g.,Smayda 1989, 1997; Richardson 1997; Anderson etal. in review). There is a strong correlation be-tween the sharp increase in nutrient loading in theharbor of Hong Kong and the increase in numberof observed red tides (Smayda 1989). There is alsosome evidence that toxin production in some spe-cies may be a function of the quantity and qualityof nutrients available (e.g., Shimizu et al. 1993).Increased nutrient availability alone does not nec-essarily result in harmful or toxic species. Someattention has been given to the ratios of inorganicnutrients, such as Si:N, suggesting that as the avail-ability of silicon decreases, the environment be-comes more conducive to the development of non-diatom species, such as harmful dinoflagellates.Dissolved Si concentrations are not available for allof the sites and sampling periods in this study.There is also evidence to suggest that as the ratioof N:P decreases, due to an increase in phosphorusavailability, species such as P. piscicida may do dis-proportionately better (Burkholder and Glasgow1997).
Examining all 3 blooms which occurred in thewaters of Maryland from 1997–1999, one consis-tent pattern in nutrients emerged (Fig. 3). Thepeaks of all bloom periods occurred coincidentallywith elevated ratios of DOC:DON. For the periodsin which outbreaks of P. piscicida and A. aureococcuswere observed, the DOC:DON ratio was 17.6 6 2.1.When P. minimum was dominant, the DOC:DONwas significantly higher, 33.6 6 2.5. In contrast, theDOC:DON of a wide range of sites from through-out the Chesapeake Bay and its tributaries, for theperiods of 1995–1997, in the salinity range of 3–11, averaged 11.8 6 2.1. In 1999 in the Pocomoke,when no significant outbreaks of P. piscicida werenoted, the ratio of DOC:DON was 10.7 (Fig. 3).Compared to archived data available for a broadrange of sites from throughout the ChesapeakeBay system, from the period 1995–1999 the DOCconcentrations observed during the 1997 fish killsand P. piscicida outbreaks in the Pocomoke Riverwere in the upper 2% of all observations recorded(Maryland Department of Natural Resources1998). Not only did the outbreaks of P. piscicidaand P. minimum occur during periods of elevated
DOC, but the ratio of DOC:DON was ;50% higherthan comparable non-bloom sites and periods. Thepattern of elevated DOC:DON ratios for A. ano-phagefferens in Maryland’s 1999 bloom has beenshown to hold for other brown tide sites outsideof Maryland. Lomas et al. (in press) has shown thisrelationship to be robust for numerous previouslydocumented brown tide blooms in Long Island,New York. The elevation in ratio may also reflecta depletion in DON relative to DOC due to utili-zation of this substrate by the plankton.
The consistency in elevation of DOC:DON forbloom periods relative to non-bloom periods is notreflected in comparisons of the same sites for in-organic N:P ratios. There is huge variation in N:Pratios for the non-bloom sites, 6.7–64.3 (data notshown). Although all P. piscicida blooms had ex-ceedingly low DIN concentrations, indicating ni-trogen depletion, such was not the case during theP. minimum or A. anophagefferens blooms. Efforts tolink each of these blooms to inorganic nutrientloading have not led to consistent predictive ca-pabilities.

880 P. M. Glibert et al.
Fig. 4. Specific uptake (V h21) of nitrogen by Prorocentrumminimum dominated phytoplankton assemblages as a functionof substrate concentration for samples collected in the Chop-tank River, May 14, 1998. Uptake rates of NO3
2 are shown asopen circles, uptake rates of NH4
1 are shown as open squares,and uptake rates of urea are shown as solid squares.
PHYSIOLOGICAL ADAPTATIONS
The elevation in DOC:DON for the 3 bloomperiods studied is not meant to necessarily implycause and effect. There are multiple possibilitiesfor this relationship, from direct stimulation ofthese blooms by an organic rich environment, tostimulation or correlation with other factors thatmay, in turn, have a stimulatory effect on HABspecies. There are physiological adaptations byeach of these species which may be significant inunderstanding why they may do especially well inorganic rich environments. P. piscicida is a het-erotrophic dinoflagellate with an ability to retainchloroplasts following the feeding of algal prey(Lewitus et al. 1999a,b). This ability, kleptochlo-roplasty, is becoming increasingly recognized indinoflagellates and some ciliates (Skovgaard1998; Stoecker 1998). Recent laboratory experi-ments with kleptochloroplastidic P. piscicida haveshown direct nitrogen uptake of NO3
2, NH41,
and glutamate at rates within the range of otherphytoplankton (Syrett et al. 1986; Antia et al.1991; Lomas and Glibert 1999). Using estimatesof ingestion of cryptophytes, along with reten-tion estimates per cryptophyte ingested (Lewitusand Caron 1990), Lewitus et al. (1999b) estimatethat nitrogen acquisition through uptake may ap-proach or exceed that obtained through grazing.Burkholder and Glasgow (1997) reported thattoxic P. piscicida can take up 14C-protein hydro-lysate. An ability to use organic nitrogen sub-strate may prove to be advantageous when inor-ganic nitrogen forms are depleted. For P. pisci-cida, it may also be of significance that crypto-phytes have been shown to have enhanced ratesof photosynthesis and growth upon organic ad-ditions (Lewitus et al. 1991; Lewitus and Kana1994), and as prey for P. piscicida may be pro-moted in systems such as the Pocomoke River.Toxic P. piscicida outbreaks have occurred in re-gions that have been highly affected by nutrients,including various North Carolina estuarine re-gions and Maryland’s eastern tributaries of Ches-apeake Bay (Burkholder et al. 1995; Lewitus etal. 1995; Burkholder and Glasgow 1997; Mary-land Department of Natural Resources 1998,1999). Clearly nutrients alone are not sufficientto promote these blooms and fish kills, as flush-ing, fish population dynamics, and other factorsare important in the development of a fish kill.As noted by Lewitus et al. (1999b), nutrients,both organic and inorganic, may promote P. pis-cicida through both direct (uptake) and indirect(trophodynamic interactions) pathways.
In contrast to P. piscicida, the dinoflagellate P.minimum is autotrophic, but it may also consume
prey heterotrophically under certain conditions(Stoecker et al. 1997). In the May 1998 P. mini-mum bloom, no grazing activity was detected bythis population (Stoecker unpublished data). Di-rect measurements of the kinetics of nitrogen up-take for the P. minimum dominated plankton com-munity (Fig. 4) revealed substantially higher ratesof NH4
1 and urea uptake than of NO32. Concen-
trations of NO32 remained at detectable concen-
trations throughout the course of the bloom, in-dicating that either demand for this substrate waslow or that rates of supply were on par with de-mand. The nitrogen required for the populationto undergo one doubling, based on the assump-tion that a healthy phytoplankton assemblage hasa particulate nitrogen:chlorophyll ratio of near1.0 (McCarthy and Nevins 1986), was in excess of100 mM N per doubling at the midpoint of thebloom. The preferential use of NH4
1 and ureaover NO3
2 indicates that these substrates weremore likely contributing to the nitrogen require-ments of the bloom, and to do so, must have hadhigh rates of regeneration. Rates of regenerationof NH4
1 and urea are not available for this period.Examination of the time course of mean urea con-centration in the water column of the easternshore rivers experiencing this P. minimum bloomshows that unusually high urea concentrationswere observed just prior to the bloom develop-ment (Fig. 5). These concentrations were nearly103 those for the same period in 1999 (Glibertunpublished data), and were correlated with theperiods of heavy rainfall noted in spring and early
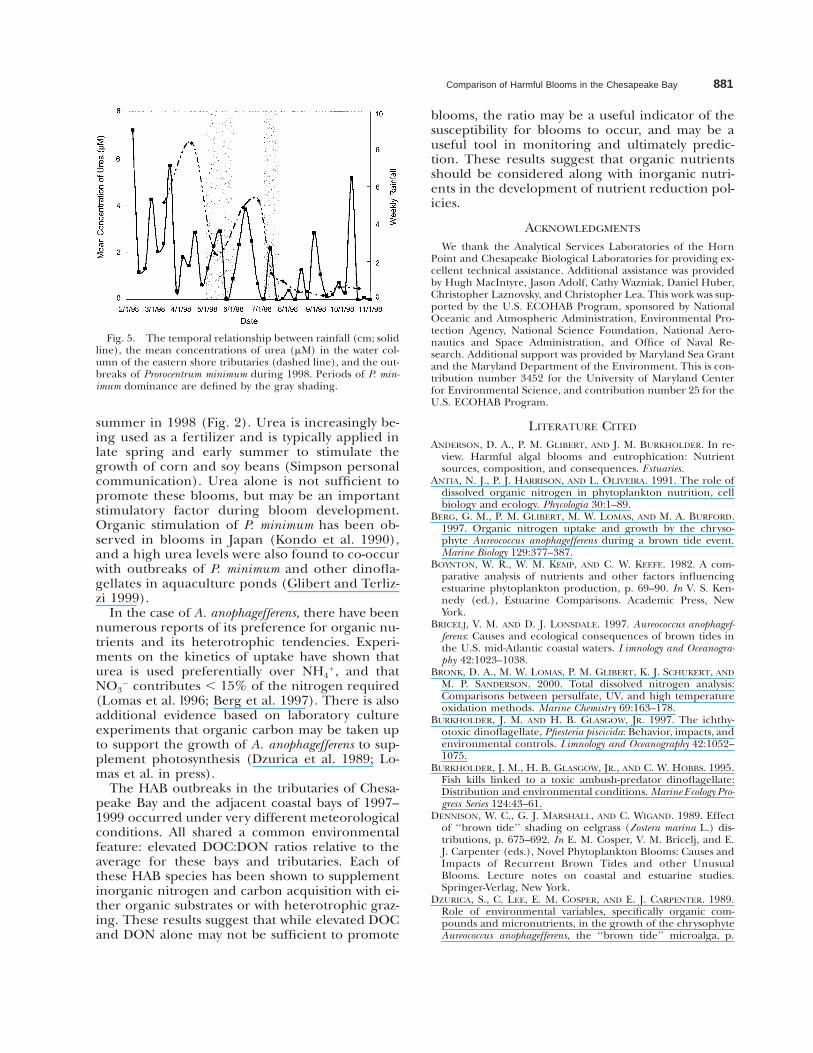
Comparison of Harmful Blooms in the Chesapeake Bay 881
Fig. 5. The temporal relationship between rainfall (cm; solidline), the mean concentrations of urea (mM) in the water col-umn of the eastern shore tributaries (dashed line), and the out-breaks of Prorocentrum minimum during 1998. Periods of P. min-imum dominance are defined by the gray shading.
summer in 1998 (Fig. 2). Urea is increasingly be-ing used as a fertilizer and is typically applied inlate spring and early summer to stimulate thegrowth of corn and soy beans (Simpson personalcommunication). Urea alone is not sufficient topromote these blooms, but may be an importantstimulatory factor during bloom development.Organic stimulation of P. minimum has been ob-served in blooms in Japan (Kondo et al. 1990),and a high urea levels were also found to co-occurwith outbreaks of P. minimum and other dinofla-gellates in aquaculture ponds (Glibert and Terliz-zi 1999).
In the case of A. anophagefferens, there have beennumerous reports of its preference for organic nu-trients and its heterotrophic tendencies. Experi-ments on the kinetics of uptake have shown thaturea is used preferentially over NH4
1, and thatNO3
2 contributes , 15% of the nitrogen required(Lomas et al. l996; Berg et al. 1997). There is alsoadditional evidence based on laboratory cultureexperiments that organic carbon may be taken upto support the growth of A. anophagefferens to sup-plement photosynthesis (Dzurica et al. 1989; Lo-mas et al. in press).
The HAB outbreaks in the tributaries of Chesa-peake Bay and the adjacent coastal bays of 1997–1999 occurred under very different meteorologicalconditions. All shared a common environmentalfeature: elevated DOC:DON ratios relative to theaverage for these bays and tributaries. Each ofthese HAB species has been shown to supplementinorganic nitrogen and carbon acquisition with ei-ther organic substrates or with heterotrophic graz-ing. These results suggest that while elevated DOCand DON alone may not be sufficient to promote
blooms, the ratio may be a useful indicator of thesusceptibility for blooms to occur, and may be auseful tool in monitoring and ultimately predic-tion. These results suggest that organic nutrientsshould be considered along with inorganic nutri-ents in the development of nutrient reduction pol-icies.
ACKNOWLEDGMENTS
We thank the Analytical Services Laboratories of the HornPoint and Chesapeake Biological Laboratories for providing ex-cellent technical assistance. Additional assistance was providedby Hugh MacIntyre, Jason Adolf, Cathy Wazniak, Daniel Huber,Christopher Laznovsky, and Christopher Lea. This work was sup-ported by the U.S. ECOHAB Program, sponsored by NationalOceanic and Atmospheric Administration, Environmental Pro-tection Agency, National Science Foundation, National Aero-nautics and Space Administration, and Office of Naval Re-search. Additional support was provided by Maryland Sea Grantand the Maryland Department of the Environment. This is con-tribution number 3452 for the University of Maryland Centerfor Environmental Science, and contribution number 25 for theU.S. ECOHAB Program.
LITERATURE CITED
ANDERSON, D. A., P. M. GLIBERT, AND J. M. BURKHOLDER. In re-view. Harmful algal blooms and eutrophication: Nutrientsources, composition, and consequences. Estuaries.
ANTIA, N. J., P. J. HARRISON, AND L. OLIVEIRA. 1991. The role ofdissolved organic nitrogen in phytoplankton nutrition, cellbiology and ecology. Phycologia 30:1–89.
BERG, G. M., P. M. GLIBERT, M. W. LOMAS, AND M. A. BURFORD.1997. Organic nitrogen uptake and growth by the chryso-phyte Aureococcus anophagefferens during a brown tide event.Marine Biology 129:377–387.
BOYNTON, W. R., W. M. KEMP, AND C. W. KEEFE. 1982. A com-parative analysis of nutrients and other factors influencingestuarine phytoplankton production, p. 69–90. In V. S. Ken-nedy (ed.), Estuarine Comparisons. Academic Press, NewYork.
BRICELJ, V. M. AND D. J. LONSDALE. 1997. Aureococcus anophagef-ferens: Causes and ecological consequences of brown tides inthe U.S. mid-Atlantic coastal waters. Limnology and Oceanogra-phy 42:1023–1038.
BRONK, D. A., M. W. LOMAS, P. M. GLIBERT, K. J. SCHUKERT, AND
M. P. SANDERSON. 2000. Total dissolved nitrogen analysis:Comparisons between persulfate, UV, and high temperatureoxidation methods. Marine Chemistry 69:163–178.
BURKHOLDER, J. M. AND H. B. GLASGOW, JR. 1997. The ichthy-otoxic dinoflagellate, Pfiesteria piscicida: Behavior, impacts, andenvironmental controls. Limnology and Oceanography 42:1052–1075.
BURKHOLDER, J. M., H. B. GLASGOW, JR., AND C. W. HOBBS. 1995.Fish kills linked to a toxic ambush-predator dinoflagellate:Distribution and environmental conditions. Marine Ecology Pro-gress Series 124:43–61.
DENNISON, W. C., G. J. MARSHALL, AND C. WIGAND. 1989. Effectof ‘‘brown tide’’ shading on eelgrass (Zostera marina L.) dis-tributions, p. 675–692. In E. M. Cosper, V. M. Bricelj, and E.J. Carpenter (eds.), Novel Phytoplankton Blooms: Causes andImpacts of Recurrent Brown Tides and other UnusualBlooms. Lecture notes on coastal and estuarine studies.Springer-Verlag, New York.
DZURICA, S., C. LEE, E. M. COSPER, AND E. J. CARPENTER. 1989.Role of environmental variables, specifically organic com-pounds and micronutrients, in the growth of the chrysophyteAureococcus anophagefferens, the ‘‘brown tide’’ microalga, p.

882 P. M. Glibert et al.
229–252. In E. M. Cosper, V. M. Bricelj, and E. J. Carpenter(eds.), Novel Phytoplankton Blooms: Causes and Impacts ofRecurrent Brown Tides and other Unusual Blooms. Lecturenotes on coastal and estuarine studies. Springer-Verlag, NewYork.
FISHER, T. R., L. W. HARDING, D. W. STANLEY, AND L. G. WARD.1988. Phytoplankton, nutrients, and turbidity in the Chesa-peake, Delaware, and Hudson estuaries. Estuarine, Coastal, andShelf Science 27:61–93.
FISHER, T. R., E. R. PEELE, J. W. AMMERMAN, AND L. W. HARDING.1992. Nutrient limitation of phytoplankton in ChesapeakeBay. Marine Ecology Progress Series 82:51–63.
FUDGE, H. 1977. The red tides of Malta. Marine Biology 39:381–386.
GLIBERT, P. M., D. J. CONLEY, T. R. FISHER, L. W. HARDING, JR.,AND T. C. MALONE. 1995. Dynamics of the 1990 winter/springbloom in Chesapeake Bay. Marine Ecology Progress Series 122:27–43.
GLIBERT, P. M., C. GARSIDE, J. A. FUHRMAN, AND M. R. ROMAN.1991. Time-dependent coupling of inorganic and organic ni-trogen uptake and regeneration in the plume of the Chesa-peake Bay estuary and its regulation by large heterotrophs.Limnology and Oceanography 36:895–909.
GLIBERT, P. M. AND D. E. TERLIZZI. 1999. Co-occurrence of ele-vated urea levels and dinoflagellate blooms in temperate es-tuarine aquaculture ponds. Applied and Environmental Micro-biology 65:5594–5596.
GOBLER, C. J. 1999. A biogeochemical investigation of Aureococ-cus anophagefferens blooms: Interactions with organic nutrientsand trace metals. Ph.D. Dissertation. State University of NewYork, Stony Brook, New York.
KIMOR, B., A. G. MOIGIS, V. DOHMS, AND C. STIENEN. 1985. Acase of mass occurrence of Prorocentrum minimum in the KielFjord. Marine Ecology Progress Series 27:209–215.
KONDO, K., Y. SEIKE, AND Y. DATE. 1990. Red tides in the brackishLake Nakanoumi (III). The stimulatory effects of organic sub-stances in the interstitial water of bottom sediments and inthe excreta from Skeletonema costatum on the growth of Proro-centrum minimum. Bulletin of Plankton Society of Japan 37:35–47.
LAROCHE, J., R. NUZZI, R. WATERS, K. WYMAN, P. G. FALKOWSKI,AND D. W. R. WALLACE. 1997. Brown tide blooms in LongIsland’s coastal waters linked to variability in groundwaterflow. Global Change Biology 3:397–410.
LEWITUS, A. J., J. M. BURKHOLDER, H. B. GLASGOW, JR., P. M.GLIBERT, B. M. WILLIS, K. C. HAYES, AND M. BURKE. 1999b.Mixotrophy and nitrogen uptake by Pfiesteria piscicida (Dino-phyceae). Journal of Phycology 35:1430–1437.
LEWITUS, A. J. AND D. A. CARON. 1990. Relative effects of nitro-gen or phosphorus depletion and light intensity on the pig-mentation, chemical composition, and volume of Pyrenomonassalina (Cryptophyceae). Marine Ecology Progress Series 61:171–181.
LEWITUS, A. J., D. A. CARON, AND K. R. MILLER. 1991. Effects oflight and glycerol on the organization of the photosyntheticapparatus in the facultative heterotroph Pyrenomonas salina(Cryptophyceae). Journal of Phycology 27:578–587.
LEWITUS, A. J., H. B. GLASGOW, JR., AND J. M. BURKHOLDER.1999a. Kleptoplastidy in the toxic dinoflagellate, Pfiesteria pis-cicida (Dinophyceae). Journal of Phycology 35:303–312.
LEWITUS, A. J., R. V. JESIEN, T. M. KANA, J. M. BURKHOLDER, H.B. GLASGOW, JR., AND E. MAY. 1995. Discovery of the ‘‘phan-tom’’ dinoflagellate in Chesapeake Bay. Estuaries 18:373–378.
LEWITUS, A. J. AND T. M. KANA. 1994. Responses of estuarinephytoplankton to exogenous glucose: Stimulation versus in-hibition of photosynthesis and respiration. Limnology andOceanography 39:182–188.
LOMAS, M. W. AND P. M. GLIBERT. 1999. Temperature regulationof nitrate uptake: A novel hypothesis about nitrate uptake andcool-water diatoms. Limnology and Oceanography 44:556–572.
LOMAS, M. W., P. M. GLIBERT, G. M. BERG, AND M. BURFORD.1996. Characterization of nitrogen uptake by natural popu-lations of Aureococcus anophagefferens (Chrysophyceae) as afunction of incubation duration, substrate concentration,light, and temperature. Journal of Phycology 32:907–916.
LOMAS, M. W., P. M. GLIBERT, D. A. CLOUGERTY, D. E. HUBER, J.JONES, J. ALEXANDER, AND E. HARAMOTO. In press. Elevatedorganic nutrient ratios associated with brown tide blooms ofAureococcus anophagefferens (Pelagophyceae). Journal of Plank-ton Research.
MALONE, T. C. 1992. Effects of water column processes on dis-solved oxygen, nutrients, phytoplankton, and zooplankton, p.61–112. In D. A. Smith, M. Leffler, and G. Mackiernan (eds.),Oxygen Dynamics in the Chesapeake Bay. Maryland SeaGrant College, College Park, Maryland.
MALONE, T. C., L. H. CROCKER, S. E. PIKE, AND B. W. WENDLER.1988. Influences of river flow on the dynamics of phytoplank-ton production in a partially stratified estuary. Marine EcologyProgress Series 48:235–249.
MALONE, T. C., P. G. FALKOWSKI, T. S. HOPKINS, G. T. ROWE, AND
T. E. WHITLEDGE. 1983. Mesoscale response of diatom popu-lations to a wind event in the plume of the Hudson River.Deep-Sea Research 30:149–170.
MARYLAND DEPARTMENT OF NATURAL RESOURCES. 1998. Waterquality, habitat and biological conditions of river systems af-fected by Pfiesteria or Pfiesteria-like organisms in the LowerEastern Shore of Maryland. 1997 Summary. Tidewater Eco-system Assessment Division. Annapolis, Maryland.
MARYLAND DEPARTMENT OF NATURAL RESOURCES. 1999. Assess-ment of water quality, habitat and biological conditions relat-ed to fish health and Pfiesteria-like organisms in Maryland.Summary 1998. Tidewater Ecosystem Assessment Division.Annapolis, Maryland.
MCCARTHY, J. J. AND J. L. NEVINS. 1986. Utilization of nitrogenand phosphorus by primary producers in warm-core ring 82-B following deep convective mixing. Deep-Sea Research 33:1773–1788.
PARSONS, T. R., Y. MAITA, AND C. M. LALLI. 1984. A Manual ofChemical and Biological Methods for Seawater Analysis. Per-gamon Press, Elmsford, New York.
RICHARDSON, K. 1997. Harmful or exceptional phytoplanktonblooms in the marine ecosystem, p. 302–386. In J. H. S. Blax-ter and A. J. Southward (eds.), Advances in Marine Biology,Vol. 31. Academic Press, San Diego, California.
SELIGER, H. H., J. A. BOGGS, AND W. H. BIGGLEY. 1985. Cata-strophic anoxia in the Chesapeake Bay in 1984. Science 228:70–73.
SHIMIZU, Y., N. WATANABE, AND G. WRENSFORD. 1993. Biosynthe-sis of brevetoxins and heterotrophic metabolism in Gymnodin-ium breve, p. 351–357. In P. Lassus, G. Arzul, E. Erard-Le-Denn,P. Gentien, and C. Marcaillou (eds.), Harmful Marine AlgalBlooms. Lavoisier Publishing, Paris.
SKOVGAARD, A. 1998. Role of chloroplast retention in a marinedinoflagellate. Aquatic Microbial Ecology 15:293–301.
SMAYDA, T. J. 1989. Primary production and the global epidemicof phytoplankton blooms in the sea: A linkage?, p. 449–483.In E. M. Cosper, V. M. Bricelj, and E. J. Carpenter (eds.),Novel Phytoplankton Blooms: Causes and Impacts of Recur-rent Brown Tides and other Unusual Blooms. Lecture Noteson Coastal and Estuarine Studies. Springer-Verlag, New York.
SMAYDA, T. J. 1997. Harmful algal blooms: Their ecophysiologyand general relevance to phytoplankton blooms in the sea.Limnology and Oceanography 42:1137–1153.
STOECKER, D. K. 1998. Conceptual models of mixotrophy inplanktonic protests and some ecological and evolutionary im-plications. European Journal of Protistology 34:281–290.
STOECKER, D. K., A. LI, W. COATS, D. E. GUSTAFSON, AND M. K.NANNEN. 1997. Mixotrophy in the dinoflagellate Prorocentrumminimum. Marine Ecology Progress Series 152:1–12.

Comparison of Harmful Blooms in the Chesapeake Bay 883
SYRETT, P. J., K. J. FLYNN, C. J. MOLLOY, G. K. DIXON, A. M.PEPLINSKA, AND R. C. CRESWELL. 1986. Effects of nitrogen dep-rivation on rates of uptake of nitrogenous compounds by thediatom, Phaeodactylum tricornutum Bohlin. New Phytologist 102:39–44.
WIKFORS, G. H. AND R. M. SMOLOWITZ. 1995. Experimental andhistological studies of four life-history stages of the easternoyster, Crassostrea virginica, exposed to a cultured strain of thedinoflagellate Prorocentrum minimum. Biological Bulletin 188:313–328.
SOURCES OF UNPUBLISHED MATERIALS
SIMPSON, T. Personal Communication. State of Maryland, De-partment of Agriculture, The Wayne A. Crawley Jr. Building,50 Harry S. Truman Parkway, Annapolis, Maryland 21401.
STOECKER, D. Unpublished Data. Horn Point Laboratory, Uni-versity of Maryland Center for Environmental Science, P.O.Box 775, Cambridge, Maryland 21613.
Received for consideration, May 10, 2000Accepted for publication, April 4, 2001
Copyright © 2022 FDOKUMEN




















