
Gonadotropin-releasing hormone agonist blocks anxiogenic-like and depressant-like effect of...
12
Gonadotropin-releasing hormone agonist blocks anxiogenic-like and depressant-like effect of corticotrophin-releasing hormone in mice S.N. Umathe a, * , P.S. Bhutada a , N.S. Jain b , N.R. Shukla a , Y.R. Mundhada a , P.V. Dixit a a Department of Pharmaceutical Sciences, Rashtrasant Tukadoji Maharaj Nagpur University, Nagpur 440 033, Maharashtra, India b J. L. C. College of Pharmacy, New Nandanvan Layout, Nagpur 440 009, Maharashtra, India Received 3 April 2008; accepted 18 April 2008 Available online 3 June 2008 Abstract Corticotrophin-releasing factor (CRF) is reported to inhibit the release of gonadotropin-releasing hormone (GnRH). In addition to the endocrine effects, GnRH is reported to influence the behavior via its neuronal interactions. We therefore, hypothesized that anxiety and depression produced by CRF could be also subsequent to the decrease in GnRH. To support such possibility, we inves- tigated the influence of GnRH agonists on CRF or CRF antagonist induced changes in social interaction time in social interaction test, and immobility time in forced swim test in mice, as the indices for anxiety and depression, respectively. Results indicated that GnRH agonists [leuprolide (20–80 ng/mouse, i.c.v.), or D-Trp-6-LHRH (40–160 ng/mouse, i.c.v.)] dose dependently increased social interaction time and decreased immobility time indicating anxiolytic- and antidepressant-like effect, respectively. Such effects of GnRH agonists were even evident in castrated mice, which suggest that these effects were unrelated to their endocrine influence. Administration of CRF (0.1 and 0.3 nmol/mouse, i.c.v.) produced just opposite effects as that of GnRH agonist on these parame- ters. Further, it was seen that pretreatment with leuprolide (10 or 20 ng/mouse, i.c.v.) or D-Trp-6-LHRH (20 or 40 ng/mouse, i.c.v.) dose dependently antagonized the effects of CRF (0.3 nmol/mouse, i.c.v.) in social interaction and forced swim test. CRF antagonist [a-Helical CRF (9–41), (1 or 10 nmol/mouse, i.c.v.)] was found to exhibit anxiolytic- and antidepressant-like effect, and its sub- effective dose (0.1 nmol/mouse, i.c.v.) when administered along with sub-threshold dose of leuprolide (10 ng/mouse, i.c.v.), or D-Trp-6-LHRH (20 ng/mouse, i.c.v.) also produced significant anxiolytic- and antidepressant-like effect. These observations suggest reciprocating role of GnRH in modulating the CRF induced anxiogenic- and depressant-like effects. Ó 2008 Elsevier Ltd. All rights reserved. Keywords: Luteinizing hormone-releasing hormone; Corticotrophin-releasing factor; HPA axis; HPG axis; Stress; Anxiety; Depression; Leuprolide; D-Trp-6-LHRH; Castration 1. Introduction Gonadotropin-releasing hormone (GnRH) is mostly known for its endocrine influence on reproductive sys- tem. However, some studies in the recent past suggested its additional role in modulating the behavior. In humans, GnRH or its receptor agonist increases alert- ness, reduces anxiety and fatigue (McAdoo et al., 1978; Soysal et al., 2001), and therefore such agents are clinically used in anxiety related disorders such as obsessive–compulsive disorder, premenstrual dysphoria and postmenopausal depression (West and Hillier, 1994; Eriksson, 2000; Bixo et al., 2001; Breckwoldt and Keck, 2002; Studd, 2006). These evidences point some role of GnRH in the modulation of anxiety. Studies conducted in human subjects using various psychological tests reported an antidepressant-like effect 0143-4179/$ - see front matter Ó 2008 Elsevier Ltd. All rights reserved. doi:10.1016/j.npep.2008.04.005 * Corresponding author. Tel.: +91 712 2500324; fax: +91 712 2500355. E-mail address: [email protected] (S.N. Umathe). www.elsevier.com/locate/npep Available online at www.sciencedirect.com Neuropeptides 42 (2008) 399–410 Neuropeptides
Transcript of Gonadotropin-releasing hormone agonist blocks anxiogenic-like and depressant-like effect of...

Available online at www.sciencedirect.com
www.elsevier.com/locate/npep
Neuropeptides 42 (2008) 399–410
Neuropeptides
Gonadotropin-releasing hormone agonist blocks anxiogenic-likeand depressant-like effect of corticotrophin-releasing hormone in mice
S.N. Umathe a,*, P.S. Bhutada a, N.S. Jain b, N.R. Shukla a,Y.R. Mundhada a, P.V. Dixit a
a Department of Pharmaceutical Sciences, Rashtrasant Tukadoji Maharaj Nagpur University, Nagpur 440 033, Maharashtra, Indiab J. L. C. College of Pharmacy, New Nandanvan Layout, Nagpur 440 009, Maharashtra, India
Received 3 April 2008; accepted 18 April 2008Available online 3 June 2008
Abstract
Corticotrophin-releasing factor (CRF) is reported to inhibit the release of gonadotropin-releasing hormone (GnRH). In additionto the endocrine effects, GnRH is reported to influence the behavior via its neuronal interactions. We therefore, hypothesized thatanxiety and depression produced by CRF could be also subsequent to the decrease in GnRH. To support such possibility, we inves-tigated the influence of GnRH agonists on CRF or CRF antagonist induced changes in social interaction time in social interactiontest, and immobility time in forced swim test in mice, as the indices for anxiety and depression, respectively. Results indicated thatGnRH agonists [leuprolide (20–80 ng/mouse, i.c.v.), or D-Trp-6-LHRH (40–160 ng/mouse, i.c.v.)] dose dependently increased socialinteraction time and decreased immobility time indicating anxiolytic- and antidepressant-like effect, respectively. Such effects ofGnRH agonists were even evident in castrated mice, which suggest that these effects were unrelated to their endocrine influence.Administration of CRF (0.1 and 0.3 nmol/mouse, i.c.v.) produced just opposite effects as that of GnRH agonist on these parame-ters. Further, it was seen that pretreatment with leuprolide (10 or 20 ng/mouse, i.c.v.) or D-Trp-6-LHRH (20 or 40 ng/mouse, i.c.v.)dose dependently antagonized the effects of CRF (0.3 nmol/mouse, i.c.v.) in social interaction and forced swim test. CRF antagonist[a-Helical CRF (9–41), (1 or 10 nmol/mouse, i.c.v.)] was found to exhibit anxiolytic- and antidepressant-like effect, and its sub-effective dose (0.1 nmol/mouse, i.c.v.) when administered along with sub-threshold dose of leuprolide (10 ng/mouse, i.c.v.), orD-Trp-6-LHRH (20 ng/mouse, i.c.v.) also produced significant anxiolytic- and antidepressant-like effect. These observations suggestreciprocating role of GnRH in modulating the CRF induced anxiogenic- and depressant-like effects.� 2008 Elsevier Ltd. All rights reserved.
Keywords: Luteinizing hormone-releasing hormone; Corticotrophin-releasing factor; HPA axis; HPG axis; Stress; Anxiety; Depression; Leuprolide;D-Trp-6-LHRH; Castration
1. Introduction
Gonadotropin-releasing hormone (GnRH) is mostlyknown for its endocrine influence on reproductive sys-tem. However, some studies in the recent past suggestedits additional role in modulating the behavior. In
0143-4179/$ - see front matter � 2008 Elsevier Ltd. All rights reserved.
doi:10.1016/j.npep.2008.04.005
* Corresponding author. Tel.: +91 712 2500324; fax: +91 7122500355.
E-mail address: [email protected] (S.N. Umathe).
humans, GnRH or its receptor agonist increases alert-ness, reduces anxiety and fatigue (McAdoo et al.,1978; Soysal et al., 2001), and therefore such agentsare clinically used in anxiety related disorders such asobsessive–compulsive disorder, premenstrual dysphoriaand postmenopausal depression (West and Hillier,1994; Eriksson, 2000; Bixo et al., 2001; Breckwoldtand Keck, 2002; Studd, 2006). These evidences pointsome role of GnRH in the modulation of anxiety.
Studies conducted in human subjects using variouspsychological tests reported an antidepressant-like effect

400 S.N. Umathe et al. / Neuropeptides 42 (2008) 399–410
of GnRH (German and Stampfer, 1979; Soulairac et al.,1983). Similar effect has been demonstrated in rats usingconditioning and forced swim test (Massol et al., 1989;Jain and Subhedar, 1993). Moreover, GnRH is reportedto increase the binding sites of imipramine—a clinicallyused antidepressant (Bernardi et al., 1990). It is alsoshown that treatment with antidepressant such as ami-triptyline increases the number of GnRH neurons (Jainand Subhedar, 1993). These evidences indicate some ill-defined role of GnRH in depression.
Recent studies demonstrated that GnRH agonistattenuates anxiety related marble-burying behavior(Uday et al., 2007; Umathe et al., 2008b), ethanol-with-drawal induced hyperexcitability (Umathe et al., 2008a),and stress-induced immunosuppression (Bobyntsev andSever’yanova, 2002; Umathe et al., 2008c). Anxiety,depression, and immunosuppression are some of theimperative characteristic features of stress response tofear provoking stimuli, and corticotrophin-releasing fac-tor (CRF) administration produces similar effects(Irwin, 1993; Todorovic et al., 2005). Therefore, CRFis considered to be the key molecule in processing thestress-induced responses.
Hypothalamic–pituitary–gonadal (HPG) axis playsan interactive role in controlling stress-activated centralfear mechanisms (Rubinow et al., 2005; Van Honket al., 2005; Hermans et al., 2007). Castrated animalsexhibit higher levels of anxiety and depression (Ber-nardi et al., 1990; Maayan et al., 2006), and low levelsof GnRH/GnRH mRNA (Rudenstein et al., 1979; Kal-ra, 1985; Rothfeld et al., 1987; Park et al., 1988;Emanuele et al., 1996). In addition, GnRH positivelyregulates CRF binding protein (Westphal and Seas-holtz, 2005), which binds to CRF with an equal orgreater affinity than the CRF receptors, and hence sug-gested to be an important modulator of CRF activity(Sutton et al., 1995). Moreover, CRF is known to inhi-bit GnRH release, mRNA expression and transcription(Petraglia et al., 1987; Williams et al., 1990; Chenet al., 1992; Tellam et al., 1998; Kinsey-Jones et al.,2006), and glucocorticoids inhibit the release of lutein-izing hormone from pituitary, and secretion of estrogenand progesterone from ovary. These effects are respon-sible for ‘hypothalamic amenorrhea’ of stress, which isobserved in anxiety and depression (Chrousos et al.,1998).
In view of these evidences, it was contemplated thatstress-induced behavioral changes could be also relatedto inhibition of GnRH by CRF. To support this hypoth-esis, we studied the influence of GnRH agonists, such asleuprolide and D-Trp-6-LHRH, on CRF induced anxi-ety and depression in mice by using social interactionand forced swim test, respectively. In addition, the roleof peripheral sex hormones in the per se behavioraleffects of GnRH agonists was studied in castratedmice.
2. Materials and methods
2.1. Animals
The experimental procedures were in strict accor-dance with the guidelines approved by the InstitutionalAnimal Ethics Committee constituted for the purposeof control and supervision of experimental animalsunder Ministry of Environment and Forests, Govern-ment of India, New Delhi, India. Subjects were younghealthy male Swiss mice (24–28 g), born and reared inthe animal house of Department of Pharmaceutical Sci-ences, Rashtrasant Tukadoji Maharaj Nagpur Univer-sity, Nagpur from a stock originally purchased fromNational Institute of Nutrition, Hyderabad, India. Micewere group housed (7–9 per cage) in opaque polypropyl-ene cages and maintained at 25 ± 2 �C under 12:12 hlight/dark cycle (07:00–19:00 h) with rodent chow (Tri-murti Feeds, Nagpur, India) and water ad libitum. Ani-mals were naive to drug treatment and experimentationat the beginning of all studies. Each experimental grouphad a separate set of 7–9 animals.
2.2. Drugs and solutions
Leuprolide acetate, D-Trp-6-LHRH, CRF (human/rat) and CRF antagonist [a-Helical CRF (9–41)] (SigmaAldrich, St. Louis, USA) was dissolved in artificial cere-brospinal fluid (aCSF) having composition 0.2 M NaCl,0.02 M NaH2CO3, 2 mM KCl, 0.5 mM KH2PO4,1.2 mM CaCl2, 1.8 mM MgCl2, 0.5 mM Na2SO4, and5.8 mM D-glucose. The selection of the doses was basedon our preliminary observations, and the previousreports (Matsumoto et al., 1997; Pelleymounter et al.,2004; Umathe et al., 2008a).
2.3. Intracerebroventricular (i.c.v.) injection
The i.c.v. cannulation was carried out as describedearlier (Umathe et al., 2008a,c). In brief, mice were anes-thetized with ketamine (100 mg/kg, i.m.) and xylazine(5 mg/kg, i.m.) combination. A guide cannula (24gauge) was stereotaxically implanted with the stereo-taxic coordinates from Paxinos and Franklin (AP�0.82 mm; ML +1.5 mm and DV +2.0 mm; related tobregma). The guide cannula was secured to the skullusing mounting screws (Plastics One Inc., USA) anddental cement (Dental Products of India, Mumbai,India). A stainless steel dummy cannula was used toocclude the guide cannula when not in use. The animalswere then allowed to recover for a week under the coverof cefotaxim (50 mg/kg, s.c.), during which they werehabituated to the experimental protocols to minimizenonspecific stress. Injections were made using a Hamil-ton microliter syringe (Hamilton, Nevada, USA) con-nected to internal cannula (24 gauge) by polyethylene

S.N. Umathe et al. / Neuropeptides 42 (2008) 399–410 401
tubing and a volume of 2.0 ll was administered over aperiod of 1 min into the right lateral ventricle. The injec-tion cannula was left in place for further 1 min beforebeing slowly withdrawn to avoid back flow. At the endof all i.c.v. experiments, dilute India ink was injected(2 ll, i.c.v.) and animals were killed immediately. Only5–10% of the mice were eliminated because of inaccuratecannula placement or injection leakage.
2.4. Castration
Castration was carried out 3 weeks prior to the exper-iment, as per the procedure described earlier (Umatheet al., 2008a,c). In brief, mouse was anesthetized withketamine (100 mg/kg, i.m.) and xylazine (5 mg/kg,i.m.) combination. After disinfecting scrotal skin with10% povidone–iodine scrub followed by 70% alcoholwipe, a small scrotal incision was made to reveal the tes-tes, which were then removed along with surroundingfatty tissues. Later, the incision was closed with chromiccatgut (5–0). Sham castrated animals only underwentsimilar surgery except removal of testes.
2.5. Social interaction test
Social interaction test was carried out by a methoddescribed earlier (Egashira et al., 2007). On day 1 and2, each mouse was subjected to a neutral cage (opaqueplastic box: 34 � 22 � 19 cm), for 5 min to acclimatize.On day 3 (test day), two unfamiliar mice were placedtogether in neutral cage for 5 min, and their behavioralinteraction was recorded by a video-camera placed1.7 m above the cage. The social interaction of a treatedanimal with untreated one was evaluated in terms ofcumulative time spent in behaviors such as sniffing, adja-cent lying, following, crawling under/over partner, andmutual grooming in 5 min. The experimenter remainedoutside the chamber during testing. After each test thefecal matter from the cage was removed and the cagewas cleaned with damp cotton soaked with alcohol.The analysis of the recording was carried out by atrained experimenter who was unaware about treatmentidentity. Increase in social interaction is considered as ananxiolytic effect.
2.6. Forced swim test
Forced swim test was carried out by a methoddescribed earlier (Porsolt et al., 1977). Mice were placedfor 6 min in a glass cylinder (height: 35 cm; diameter:17 cm) filled with water (25 ± 1 �C) to a depth of25 cm. The water depth was adjusted so that the animalsmust swim or float without their hind limbs or tailtouching the bottom. During testing, individual mousewas placed in the cylinder for 6 min, and the duration
of immobility (the time during which mouse made onlythe small movements necessary to keep their headsabove water) was scored. As suggested by Porsolt, onlythe data scored during the last 4 min were analyzed andpresented.
2.7. Locomotor activity
Locomotor activity was assessed in actophotometer(Techno, Lucknow), having a circular arena of 40 cm,equipped with three infrared beams and photo-cells con-nected to digital counter. Locomotor activity wasexpressed in terms of total number of counts of lightbeams interruptions in 30 min.
2.8. Influence of leuprolide, D-Trp-6-LHRH, CRF, and
CRF antagonist in the social interaction and forced swim
test
Leuprolide (5, 10, 20, 40, or 80 ng/mouse) or D-Trp-6-LHRH (10, 20, 40, 80, or 160 ng/mouse) or CRF(0.01, 0.1, or 0.3 nmol/mouse) or CRF antagonist (0.1,1, or 10 nmol/mouse) or aCSF (2 ll/mouse) was admin-istered i.c.v. to mice. Twenty minutes after these admin-istrations, individual mouse was subjected to eithersocial interaction test or forced swim test, and time spentin social interaction, and immobility time in forced swimtest was recorded as described above. Separate group ofmice (n = 7–9) was employed for each treatment, doseand test.
2.9. Influence of leuprolide, D-Trp-6-LHRH, CRF, and
CRF antagonist on locomotor activity
Twenty minutes after leuprolide (5, 10, 20, 40, or80 ng/mouse, i.c.v.) or D-Trp-6-LHRH (10, 20, 40, 80,or 160 ng/mouse, i.c.v.) or CRF (0.01, 0.1, or0.3 nmol/mouse, i.c.v.) or CRF antagonist (0.1, 1, or10 nmol/mouse, i.c.v.) or aCSF (2 ll/mouse, i.c.v.) treat-ment, mice were tested in actophotometer as describedabove and number of light beam interruptions in30 min was recorded as locomotor count. Separategroup of mice (n = 7–9) was employed for each treat-ment and dose.
2.10. Influence of GnRH agonists on the effect of CRF inthe social interaction and forced swim test
Separate groups of mice (n = 7–9) were treated withleuprolide (5, 10, or 20 ng/mouse, i.c.v.) or D-Trp-6-LHRH (10, 20, or 40 ng/mouse, i.c.v.), 10 min beforeCRF (0.3 nmol/mouse, i.c.v.) administration. Ten min-utes thereafter, individual mouse was subjected to eithersocial interaction or forced swim test, and related indiceswere recorded as described above.
402 S.N. Umathe et al. / Neuropeptides 42 (2008) 399–410
2.11. Influence of GnRH agonists co-administered with
CRF antagonist in sub-effective doses in the social
interaction and forced swim test
Separate set of mice (n = 7–9) were treated concomi-tantly with CRF antagonist (0.1 nmol, i.c.v.), and leu-prolide (10 ng/mouse, i.c.v.) or D-Trp-6-LHRH (20 ng/mouse, i.c.v.), and 20 min after these administration,individual mouse was subjected to either social interac-tion or forced swim test, and related indices wererecorded as described above.
2.12. Influence of leuprolide and D-Trp-6-LHRH on the
social interaction and forced swim test in castrated mice
Leuprolide (40 ng/mouse) or D-Trp-6-LHRH (80 ng/mouse) was administered i.c.v. in castrated and shamcastrated mice. Control group received aCSF (2 ll/mouse, i.c.v.). Twenty minutes after administration,individual mouse was subjected to either social interac-tion test or forced swim test and time spent in socialinteraction and immobility time in forced swim testwas recorded. Separate groups of mice (n = 6–7), wereused for each test, drug and dose.
2.13. Data analysis
Data was analyzed using either one-way analysis ofvariance (ANOVA) followed by Dunnett’s/Newman-
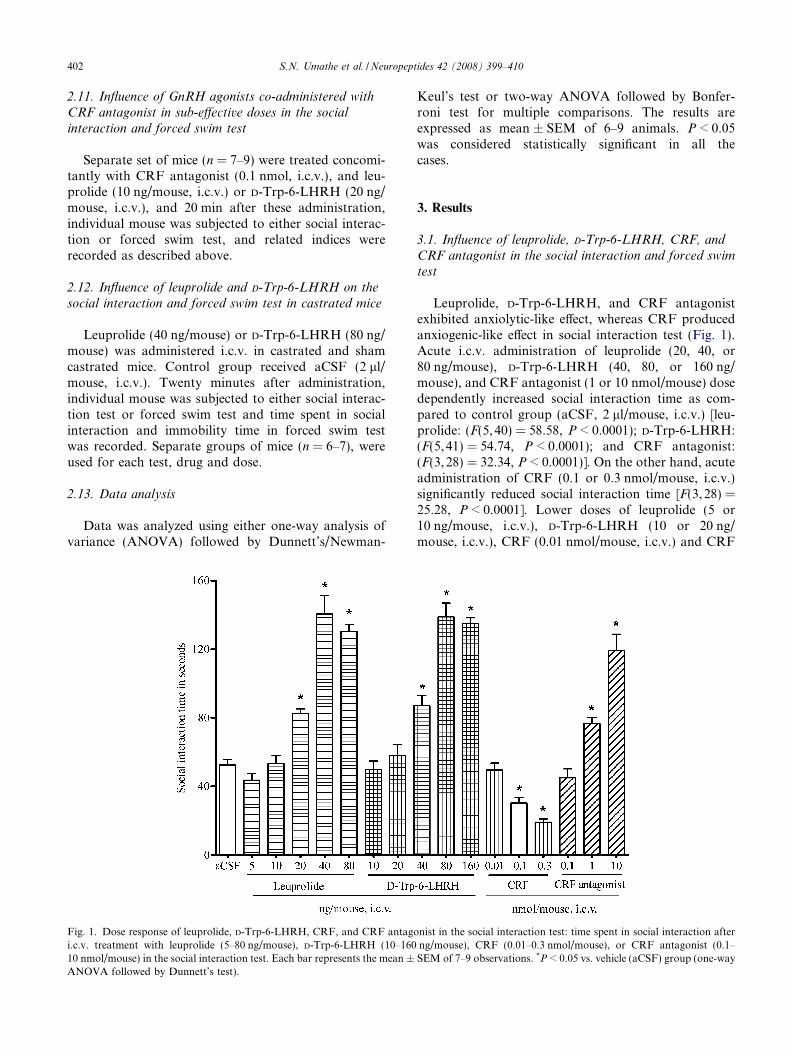
Fig. 1. Dose response of leuprolide, D-Trp-6-LHRH, CRF, and CRF antagoi.c.v. treatment with leuprolide (5–80 ng/mouse), D-Trp-6-LHRH (10–16010 nmol/mouse) in the social interaction test. Each bar represents the mean ±ANOVA followed by Dunnett’s test).
Keul’s test or two-way ANOVA followed by Bonfer-roni test for multiple comparisons. The results areexpressed as mean ± SEM of 6–9 animals. P < 0.05was considered statistically significant in all thecases.
3. Results
3.1. Influence of leuprolide, D-Trp-6-LHRH, CRF, and
CRF antagonist in the social interaction and forced swim
test
Leuprolide, D-Trp-6-LHRH, and CRF antagonistexhibited anxiolytic-like effect, whereas CRF producedanxiogenic-like effect in social interaction test (Fig. 1).Acute i.c.v. administration of leuprolide (20, 40, or80 ng/mouse), D-Trp-6-LHRH (40, 80, or 160 ng/mouse), and CRF antagonist (1 or 10 nmol/mouse) dosedependently increased social interaction time as com-pared to control group (aCSF, 2 ll/mouse, i.c.v.) [leu-prolide: (F(5, 40) = 58.58, P < 0.0001); D-Trp-6-LHRH:(F(5,41) = 54.74, P < 0.0001); and CRF antagonist:(F(3,28) = 32.34, P < 0.0001)]. On the other hand, acuteadministration of CRF (0.1 or 0.3 nmol/mouse, i.c.v.)significantly reduced social interaction time [F(3, 28) =25.28, P < 0.0001]. Lower doses of leuprolide (5 or10 ng/mouse, i.c.v.), D-Trp-6-LHRH (10 or 20 ng/mouse, i.c.v.), CRF (0.01 nmol/mouse, i.c.v.) and CRF
nist in the social interaction test: time spent in social interaction afterng/mouse), CRF (0.01–0.3 nmol/mouse), or CRF antagonist (0.1–SEM of 7–9 observations. *P < 0.05 vs. vehicle (aCSF) group (one-way
S.N. Umathe et al. / Neuropeptides 42 (2008) 399–410 403
antagonist (0.1 nmol/mouse, i.c.v.) were devoid of anyeffect (P > 0.05).
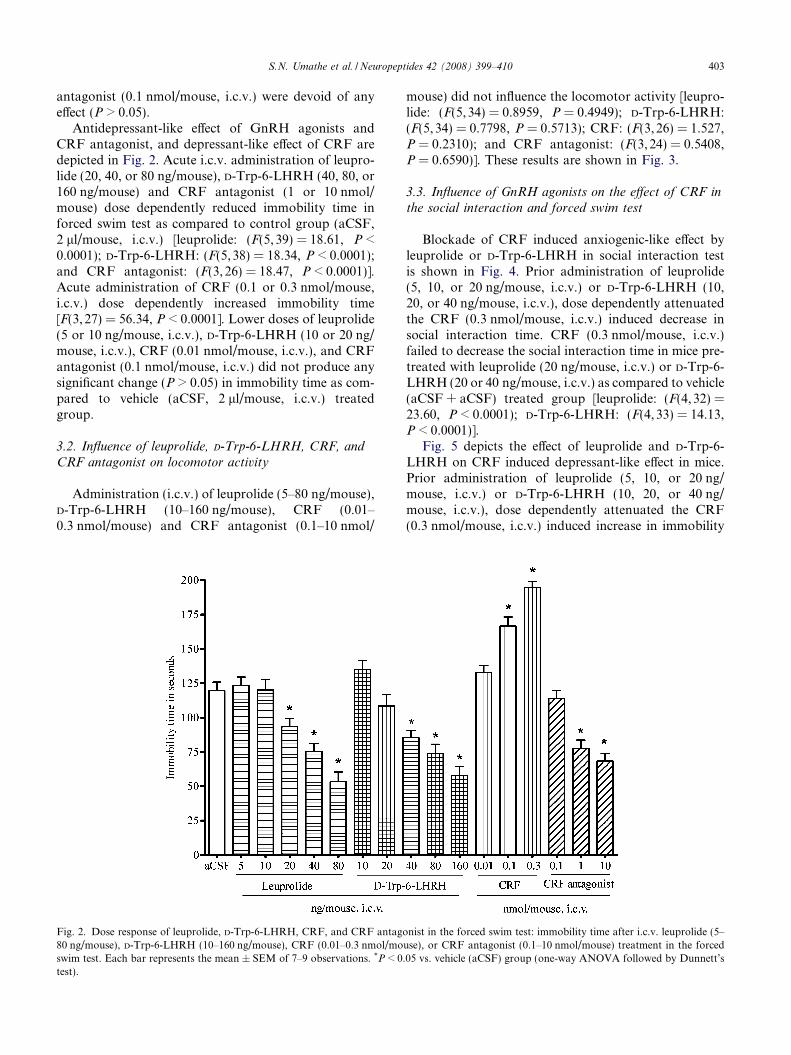
Antidepressant-like effect of GnRH agonists andCRF antagonist, and depressant-like effect of CRF aredepicted in Fig. 2. Acute i.c.v. administration of leupro-lide (20, 40, or 80 ng/mouse), D-Trp-6-LHRH (40, 80, or160 ng/mouse) and CRF antagonist (1 or 10 nmol/mouse) dose dependently reduced immobility time inforced swim test as compared to control group (aCSF,2 ll/mouse, i.c.v.) [leuprolide: (F(5, 39) = 18.61, P <0.0001); D-Trp-6-LHRH: (F(5,38) = 18.34, P < 0.0001);and CRF antagonist: (F(3, 26) = 18.47, P < 0.0001)].Acute administration of CRF (0.1 or 0.3 nmol/mouse,i.c.v.) dose dependently increased immobility time[F(3, 27) = 56.34, P < 0.0001]. Lower doses of leuprolide(5 or 10 ng/mouse, i.c.v.), D-Trp-6-LHRH (10 or 20 ng/mouse, i.c.v.), CRF (0.01 nmol/mouse, i.c.v.), and CRFantagonist (0.1 nmol/mouse, i.c.v.) did not produce anysignificant change (P > 0.05) in immobility time as com-pared to vehicle (aCSF, 2 ll/mouse, i.c.v.) treatedgroup.
3.2. Influence of leuprolide, D-Trp-6-LHRH, CRF, and
CRF antagonist on locomotor activity
Administration (i.c.v.) of leuprolide (5–80 ng/mouse),D-Trp-6-LHRH (10–160 ng/mouse), CRF (0.01–0.3 nmol/mouse) and CRF antagonist (0.1–10 nmol/
Fig. 2. Dose response of leuprolide, D-Trp-6-LHRH, CRF, and CRF antago80 ng/mouse), D-Trp-6-LHRH (10–160 ng/mouse), CRF (0.01–0.3 nmol/moswim test. Each bar represents the mean ± SEM of 7–9 observations. *P < 0.test).
mouse) did not influence the locomotor activity [leupro-lide: (F(5,34) = 0.8959, P = 0.4949); D-Trp-6-LHRH:(F(5, 34) = 0.7798, P = 0.5713); CRF: (F(3, 26) = 1.527,P = 0.2310); and CRF antagonist: (F(3, 24) = 0.5408,P = 0.6590)]. These results are shown in Fig. 3.
3.3. Influence of GnRH agonists on the effect of CRF in
the social interaction and forced swim test
Blockade of CRF induced anxiogenic-like effect byleuprolide or D-Trp-6-LHRH in social interaction testis shown in Fig. 4. Prior administration of leuprolide(5, 10, or 20 ng/mouse, i.c.v.) or D-Trp-6-LHRH (10,20, or 40 ng/mouse, i.c.v.), dose dependently attenuatedthe CRF (0.3 nmol/mouse, i.c.v.) induced decrease insocial interaction time. CRF (0.3 nmol/mouse, i.c.v.)failed to decrease the social interaction time in mice pre-treated with leuprolide (20 ng/mouse, i.c.v.) or D-Trp-6-LHRH (20 or 40 ng/mouse, i.c.v.) as compared to vehicle(aCSF + aCSF) treated group [leuprolide: (F(4,32) =23.60, P < 0.0001); D-Trp-6-LHRH: (F(4, 33) = 14.13,P < 0.0001)].
Fig. 5 depicts the effect of leuprolide and D-Trp-6-LHRH on CRF induced depressant-like effect in mice.Prior administration of leuprolide (5, 10, or 20 ng/mouse, i.c.v.) or D-Trp-6-LHRH (10, 20, or 40 ng/mouse, i.c.v.), dose dependently attenuated the CRF(0.3 nmol/mouse, i.c.v.) induced increase in immobility
nist in the forced swim test: immobility time after i.c.v. leuprolide (5–use), or CRF antagonist (0.1–10 nmol/mouse) treatment in the forced05 vs. vehicle (aCSF) group (one-way ANOVA followed by Dunnett’s

Fig. 3. Effect of leuprolide, D-Trp-6-LHRH, CRF, and CRF antagonist on locomotor activity: locomotor activity after i.c.v. leuprolide (5–80 ng/mouse), D-Trp-6-LHRH (10–160 ng/mouse), CRF (0.01–0.3 nmol/mouse), or CRF antagonist (0.1–10 nmol/mouse) treatment. Each bar representsthe mean ± SEM of 7–9 observations (one-way ANOVA followed by Dunnett’s test).
Fig. 4. Effect of leuprolide or D-Trp-6-LHRH on CRF induced anxiogenic-like effect in the social interaction test: mice were treated with leuprolide(5, 10, or 20 ng/mouse, i.c.v.), or D-Trp-6-LHRH (10, 20, or 40 ng/mouse, i.c.v.), prior to CRF (0.3 nmol/mouse), and time spent in social interactionwas recorded. Each bar represents the mean ± SEM of 7–9 observations. **P < 0.001, *P < 0.01 vs. control group (aCSF + aCSF); ##P < 0.001,#P < 0.05 vs. (aCSF + CRF) treated group (one-way ANOVA followed by Newman-Keul’s test).
404 S.N. Umathe et al. / Neuropeptides 42 (2008) 399–410
time. CRF (0.3 nmol/mouse, i.c.v.) failed to increase theimmobility time in mice pretreated with leuprolide
(20 ng/mouse, i.c.v.) or D-Trp-6-LHRH (40 ng/mouse,i.c.v.) as compared to vehicle (aCSF + aCSF) treated

Fig. 5. Effect of GnRH agonists on CRF induced depressant-like effect in the forced swim test: mice were treated with leuprolide (5, 10, or 20 ng/mouse, i.c.v.) or D-Trp-6-LHRH (10, 20, or 40 ng/mouse, i.c.v.), prior to CRF (0.3 nmol/mouse), and immobility time in forced swim test wasrecorded. Each bar represents the mean ± SEM of 7–9 observations. **P < 0.001, *P < 0.01 vs. control group (aCSF + aCSF); ##P < 0.001 vs.(aCSF + CRF) treated group (one-way ANOVA followed by Newman-Keul’s test).
S.N. Umathe et al. / Neuropeptides 42 (2008) 399–410 405
group [leuprolide: (F(4,31) = 37.41, P < 0.0001); D-Trp-6-LHRH: (F(4,33) = 36.16, P < 0.0001)].
3.4. Influence of GnRH agonists co-administered with
CRF antagonist in the social interaction and forced swim
test
Co-administration of GnRH agonists and CRFantagonist in their respective sub-effective doses pro-duced significant anxiolytic- and antidepressant-likeeffect. Leuprolide (10 ng/mouse, i.c.v.) or D-Trp-6-LHRH (20 ng/mouse, i.c.v.) with CRF antagonist(0.1 nmol/mouse, i.c.v.), significantly increased socialinteraction time [leuprolide: (F(3, 26) = 24.50, P <0.0001); D-Trp-6-LHRH: (F(3,25) = 15.03, P < 0.0001)](Fig. 6), and decreased immobility time [leuprolide:(F(3, 26) = 18.62, P < 0.0001); D-Trp-6-LHRH:(F(3, 25) = 15.90, P < 0.0001)] (Fig. 7).
3.5. Influence of leuprolide and D-Trp-6-LHRH on the
social interaction and forced swim test in castrated mice
Fig. 8 depicts the effect of GnRH agonists in cas-trated mice. Castrated mice per se exhibited anxiogenic-and depressant-like effect in social interaction andforced swim test, respectively. Treatment of castratedmice with leuprolide (40 ng/mouse, i.c.v.) or D-Trp-6-LHRH (80 ng/mouse, i.c.v.), significantly increased
social interaction time [castration effect: (F(1,30) =38.64, P < 0.0001); and treatment effect: (F(2,30) =54.32, P < 0.0001)]. Result shown in Fig. 9 depicts theeffect of GnRH agonists on immobility time in forcedswim test. Leuprolide (40 ng/mouse, i.c.v.) or D-Trp-6-LHRH (80 ng/mouse, i.c.v.) significantly decreasedimmobility time [castration effect: (F(1, 30) = 89.77,P < 0.0001); and treatment effect: (F(2, 30) = 80.79,P < 0.0001)].
4. Discussion
The present investigations demonstrated that i.c.v.administration of GnRH agonists produced anxiolytic-and antidepressant-like effect in mice, when tested,respectively, in social interaction and forced swim test.In accordance with the earlier reports, i.c.v. administra-tion of CRF exhibited anxiogenic- and depressant-likebehavior (Britton et al., 1985, 1988; Pelleymounteret al., 2004; Swiergiel et al., 2007), and CRF antagonistproduced anxiolytic- and antidepressant-like effect(Holsboer, 1999; Hodgson et al., 2007). In addition, pre-treatment with sub-effective dose of GnRH agonists sig-nificantly antagonized, while minimum effective dosecompletely blocked CRF induced anxiogenic- anddepressant-like behavior, which indicate that GnRHnegatively modulated CRF induced behavioral effects.

Fig. 6. Effect of co-administration of sub-effective doses of GnRH agonists and CRF antagonist in the social interaction test: mice were treated withleuprolide (10 ng/mouse, i.c.v.), or D-Trp-6-LHRH (20 ng/mouse, i.c.v.) with CRF antagonist (0.1 nmol/mouse, i.c.v.), and time spent in socialinteraction was recorded. Each bar represents the mean ± SEM of 7–9 observations. *P < 0.001 vs. (CRF antagonist + aCSF), and (aCSF + GnRHagonist) treated group (one-way ANOVA followed by Dunnett’s test).
Fig. 7. Effect of co-administration of sub-effective doses of GnRH agonists and CRF antagonist in the forced swim test: mice were treated withleuprolide (10 ng/mouse, i.c.v.), or D-Trp-6-LHRH (20 ng/mouse, i.c.v.) with CRF antagonist (0.1 nmol/mouse), and immobility time was recorded.Each bar represents the mean ± SEM of 7–9 observations. *P < 0.001 vs. (CRF antagonist + aCSF), and (aCSF + GnRH agonist) treated group(one-way ANOVA followed by Dunnett’s test).
406 S.N. Umathe et al. / Neuropeptides 42 (2008) 399–410
It was interesting to note that GnRH agonists and CRFantagonist, when co-administered in their respective
sub-effective doses produced significant anxiolytic- andantidepressant-like effect. None of these treatments per

Fig. 8. Effect of GnRH agonists in castrated mice on the socialinteraction test: castrated and sham castrated mice were treated withleuprolide (40 ng/mouse, i.c.v.) or D-Trp-6-LHRH (80 ng/mouse,i.c.v.), or vehicle (2 ll/mouse, i.c.v.), and time spent in socialinteraction was recorded. Each bar represents the mean ± SEM of6–7 observations. *P < 0.05 vs. vehicle group (sham castrated);#P < 0.05 vs. vehicle group (castrated) (two-way ANOVA followedby Bonferroni test).
Fig. 9. Effect of GnRH agonists in castrated mice on the forced swimtest: castrated and sham castrated mice were treated with leuprolide(40 ng/mouse, i.c.v.), or D-Trp-6-LHRH (80 ng/mouse, i.c.v.) orvehicle (2 ll/mouse, i.c.v.), and immobility time was recorded. Eachbar represents the mean ± SEM of 6–7 observations. *P < 0.05 vs.vehicle group (sham castrated); #P < 0.05 vs. vehicle group (castrated)(two-way ANOVA followed by Bonferroni test).
S.N. Umathe et al. / Neuropeptides 42 (2008) 399–410 407
se influenced the locomotor activity in the employeddoses. Thus, GnRH and CRF induced behavioralchanges were independent of motor toxicity. The under-lying mechanisms for the observed behavioral interac-tion between GnRH and CRF are obscure.
Interestingly, stress, ethanol, and CRF have beenshown to inhibit the hypothalamic GnRH release andGnRH mRNA expression (Petraglia et al., 1987; Wil-liams et al., 1990; Chen et al., 1992; Rettori and McC-ann, 1997; Tellam et al., 1998; Lomniczi et al., 2000;Kim et al., 2003; Kinsey-Jones et al., 2006); GnRH ago-
nists are reported to prevent stress-induced immunosup-pression, ethanol-withdrawal induced hyperactivity andmarble-burying behavior in mice (Bobyntsev andSever’yanova, 2002; Umathe et al., 2008a,b). In line withthese reports, present study demonstrated significantblockade of CRF induced anxiogenic- and depressant-like effect by GnRH agonists, which suggest that GnRHcan counter the behavioral consequences of CRF. Anx-iolytic- and antidepressant-like effect seen after the co-administration of GnRH agonists and CRF antagonistin their respective sub-effective doses, further supportsthe reciprocating interaction between them.
Incidentally, GnRH has been demonstrated to posi-tively regulate the expression of CRF binding protein(Westphal and Seasholtz, 2005). Thus, increased bindingof CRF with its protein may be the possible reason forthe ineffectiveness of CRF to produce its behavioraleffects in presence of GnRH agonists. Immunocyto-chemical studies have demonstrated synaptic connec-tions between CRF and GnRH neurons in medialpreoptic area of hypothalamus (MacLusky et al.,1988), which indicate their cross talk in the related func-tions. Similarly, literature also documents the presenceof receptors for GnRH at amygdala, hippocampus,nucleus accumbens, septum, preoptic area, olfactorysystem, prepiriform cortex (Merchenthaler et al., 1984;Reubi and Maurer, 1984; Badr and Pelletier, 1987; Reu-bi et al., 1987; Jennes et al., 1988; Leblanc et al., 1988;Rance et al., 1994), and for CRF as well (De Souzaet al., 1985; De Souza, 1987). These neuroanatomicalsites are responsible for the behavior like anxiety anddepression. In view of these evidences, the ability ofGnRH to block the behavioral effects of CRF couldbe related to the interaction between their receptors atthese neuroanatomical regions.
Literature documents several evidences of neuromod-ulatory influence of GnRH. GnRH increases gamma-aminobutyric acid (GABA) in several brain regions(Pasha and Vijayan, 1992); and the agonists of theGABA/benzodiazepine receptor complex were shownto attenuate behavioral effects of stress/CRF (Brittonet al., 1985, 1988). In addition, antidopaminergics areknown to produce anxiolytic- and antidepressant-likeeffect (Simon et al., 1993; Ponnusamy et al., 2005;Yatham et al., 2005); and GnRH is reported to exhibitantidopaminergic-like activity (Kadar et al., 1992).GnRH is also known to modulate serotonergic system(Pan et al., 1988; Uday et al., 2007); and selective sero-tonin reuptake inhibitors are well established antide-pressants and reduce anxiety (Papakostas et al., 2008).Whether these effects of GnRH agonists are responsiblefor the observed anxiolytic- and antidepressant-likeeffect as well as for their ability to block the behavioraleffects of CRF, is quite intriguing.
GnRH agonists activate sex hormones synthesis inperiphery, which also influence stress related behavior

408 S.N. Umathe et al. / Neuropeptides 42 (2008) 399–410
such as anxiety and depression. Testosterone is reportedto exhibit anxiolytic- and antidepressant-like effect(Rubinow et al., 2005; Van Honk et al., 2005; Hermanset al., 2007; Kanayama et al., 2007). Therefore to under-stand the contribution of sex steroid in the observedeffects of GnRH agonists per se, the studies were carriedout in castrated mice. Results revealed that castrationper se produced anxiogenic- and depressant-like effect,which is well in accordance with earlier reports (Ber-nardi et al., 1990; Maayan et al., 2006). However, it isinteresting to note that GnRH agonists blocked the cas-tration induced anxiety and depression, suggesting thatthis effect of GnRH agonists may be due to their neuro-nal interactions. Incidentally, castrated animals arereported to exhibit low levels of GnRH/GnRH mRNAin hypothalamus (Rudenstein et al., 1979; Kalra, 1985;Rothfeld et al., 1987; Park et al., 1988; Emanueleet al., 1996).
When we attempted to study the interaction betweenGnRH and CRF in castrated mice, it was surprising tonote that administration of CRF (0.01–0.1 nmol/mouse,i.c.v.) failed to further augment the already higher levelof anxiety and depression evident in castrated mice; andthe higher dose of CRF (0.3 nmol/mouse, i.c.v.) pro-duced significant motor in-coordination (data notshown). The basal level of anxiety and depression exhib-ited by the castrated mice was comparable to the anxietyand depression induced by CRF (0.3 nmol/mouse, i.c.v.)administration in non-castrated group. Therefore, theemployed animal model was not viable to study theinteraction between CRF and GnRH in castrated mice.However, it is important to note that castrated animalsexhibit increased levels of CRF (Bingaman et al., 1994;Ogilvie and Rivier, 1997), which indirectly supportsthe interaction between GnRH and CRF.
In conclusion, central administration of GnRHblocked anxiogenic- and depressant-like effects ofCRF. These findings support the hypothesis that CRFand GnRH modulate the behavioral responses to stressin a counter regulating manner.
References
Badr, M., Pelletier, G., 1987. Characterization and autoradiographiclocalization of LHRH receptors in the rat brain. Synapse 1, 567–571.
Bernardi, M., Sandrini, M., Vergoni, A.V., Marrama, D., Tagliavini,S., Bertolini, A., 1990. Influence of gonadotropin-releasing hor-mone on castration-induced ‘depression’ in mice a behavioral andbinding study. Eur. J. Pharmacol. 187, 501–506.
Bingaman, E.W., Magnuson, D.J., Gray, T.S., Handa, R.J., 1994.Androgen inhibits the increases in hypothalamic corticotropin-releasing hormone (CRH) and CRH-immunoreactivity followinggonadectomy. Neuroendocrinology 59, 228–234.
Bixo, M., Allard, P., Backstrom, T., Mjorndal, T., Nyberg, S., Spigset,O., Sundstrom-Poromaa, I., 2001. Binding of [3H] paroxetine toserotonin uptake sites and of [3H] lysergic acid diethylamide to 5-
HT2A receptors in platelets from women with premenstrualdysphoric disorder during gonadotropin releasing hormone treat-ment. Psychoneuroendocrinology 26, 551–564.
Bobyntsev, I.I., Sever’yanova, L.A., 2002. Immunotropic effects ofgonadotropin-releasing hormone analog under conditions of emo-tional painful stress. Bull. Exp. Biol. Med. 133, 436–438.
Breckwoldt, M., Keck, C.H., 2002. Premenstrual syndrome. Ther.Umsch. 59, 183–187.
Britton, K.T., Morgan, J., Rivier, J., Koob, G.F., 1985. Chlordiaz-epoxide attenuates CRF-induced response suppression in theconflict test. Psychopharmacology 86, 170–174.
Britton, K.T., Lee, G., Koob, G.F., 1988. Corticotropin releasingfactor and amphetamine exaggerate partial agonist properties ofbenzodiazepine antagonist Ro 15-1788 in the conflict test. Psycho-pharmacology 94, 306–311.
Chen, M.D., O’Byrne, K.T., Chiappini, S.E., Hotchkiss, J., Knobil, E.,1992. Hypoglycemic ‘‘stress” and gonadotropin-releasing hormonepulse generator activity in the rhesus monkey: role of the ovary.Neuroendocrinology 56, 666–673.
Chrousos, G.P., Torpy, D.J., Gold, P.W., 1998. Interactions betweenthe hypothalamic–pituitary–adrenal axis and the female reproduc-tive system: clinical implications. Ann. Intern. Med. 129, 229–240.
De Souza, E.B., 1987. Corticotropin-releasing factor receptors in therat central nervous system: characterization and regional distribu-tion. J. Neurosci. 7, 88–100.
De Souza, E.B., Insel, T.R., Perrin, M.H., Rivier, J., Vale, W.W.,Kuhar, M.J., 1985. Corticotropin-releasing factor receptors arewidely distributed within the rat central nervous system: anautoradiographic study. J. Neurosci. 5, 3189–3203.
Egashira, N., Tanoue, A., Matsuda, T., Koushi, E., Harada, S.,Takano, Y., Tsujimoto, G., Mishima, K., Iwasaki, K., Fujiwara,M., 2007. Impaired social interaction and reduced anxiety-relatedbehavior in vasopressin V1a receptor knockout mice. Behav. BrainRes. 178, 123–127.
Emanuele, N.V., Jurgens, J., La Paglia, N., Williams, D.W., Kelley,M.R., 1996. The effect of castration on steady state levels ofluteinizing hormone-releasing hormone (LHRH) mRNA andproLHRH processing: time course study utilizing semi-quantitativereverse transcription/polymerase chain reaction. J. Endocrinol.148, 509–515.
Eriksson, T., 2000. Antiandrogenic treatment for obsessive–compul-sive disorder. Am. J. Psychiat. 157, 483.
German, G.A., Stampfer, H.G., 1979. Hypothalamic releasing factorfor reactive depression. Lancet 13, 789.
Hermans, E.J., Putmanc, P., Baasa, J.M., Gecksa, N.M., Kenemansa,J.L., Honka, J.V., 2007. Exogenous testosterone attenuates theintegrated central stress response in healthy young women.Psychoneuroendocrinology 32, 1052–1061.
Hodgson, R.A., Higgins, G.A., Guthrie, D.H., Lu, S.X., Pond, A.J.,Mullins, D.E., Guzzi, M.F., Parker, E.M., Varty, G.B., 2007.Comparison of the V1b antagonist, SSR149415, and the CRF1antagonist, CP-154,526, in rodent models of anxiety and depres-sion. Pharmacol. Biochem. Behav. 86, 431–440.
Holsboer, F., 1999. The rationale for corticotropin-releasing hormonereceptor (CRH-R) antagonists to treat depression and anxiety. J.Psychiat. Res. 33, 181–214.
Irwin, M., 1993. Stress-induced immune suppression. Role of theautonomic nervous system. Ann. NY Acad. Sci. 697, 203–218.
Jain, M.R., Subhedar, N.K., 1993. Increase in number of LHRHneurones in septal-preoptic area of rats following chronic amitrip-tyline treatment: implication in antidepressant effect. Brain Res.604, 7–15.
Jennes, L., Dalati, B., Conn, P.M., 1988. Distribution of gonadotropinreleasing hormone agonist binding sites in the rat central nervoussystem. Brain Res. 452, 156–164.

S.N. Umathe et al. / Neuropeptides 42 (2008) 399–410 409
Kadar, T., Telegdy, G., Schally, A.V., 1992. Behavioral effects ofcentrally administered LHRH agonist in rats. Physiol. Behav. 51,601–605.
Kalra, P.S., 1985. Stimulation of hypothalamic LHRH levels andrelease by gonadal steroids. J. Steroid Biochem. 23, 725–731.
Kanayama, G., Amiaz, R., Seidman, S., Pope Jr., H.G., 2007.Testosterone supplementation for depressed men: current researchand suggested treatment guidelines. Exp. Clin. Psychopharmacol.15, 529–538.
Kim, J.H., Kim, H.J., Noh, H.S., Roh, G.S., Kang, S.S., Cho, G.J.,Park, S.K., Lee, B.J., Choi, W.S., 2003. Suppression by ethanol ofmale reproductive activity. Brain Res. 989, 91–98.
Kinsey-Jones, J.S., Li, X.F., Bowe, J.E., Lightman, S.L., O’Byrne,K.T., 2006. Corticotrophin-releasing factor type 2 receptor-medi-ated suppression of gonadotrophin-releasing hormone mRNAexpression in GT1-7 cells. Stress 4, 215–222.
Leblanc, P., Crumeyrolle, M., Latouche, J., Jordan, D., Fillion, G.,L’Heritier, A., Kordon, C., Dussaillant, M., Rostene, W., Haour,F., 1988. Characterization and distribution of receptors forgonadotropin releasing hormone in the rat hippocampus. Neuro-endocrinology 48, 482–488.
Lomniczi, A., Mastronardi, C.A., Faletti, A.G., Seilicovich, A., DeLaurentiis, A., McCann, S.M., Rettori, V., 2000. Inhibitorypathways and the inhibition of luteinizing hormone-releasinghormone release by alcohol. Proc. Natl. Acad. Sci. USA 97,2337–2342.
Maayan, R., Touati-Werner, D., Ram, E., Strous, R., Keren, O.,Weizman, A., 2006. The protective effect of frontal cortexdehydroepiandrosterone in anxiety and depressive models in mice.Pharmacol. Biochem. Behav. 85, 415–421.
MacLusky, N.J., Naftolin, F., Leranth, C., 1988. Immunocytochem-ical evidence for direct synaptic connections between corticotro-phin-releasing factor (CRF) and gonadotrophin-releasing hormone(GnRH)-containing neurons in the preoptic area of the rat. BrainRes. 439, 391–395.
Massol, J., Martin, P., Adessi, G., Peuch, A.J., 1989. Gonadotropin-releasing hormone (GnRH) as antidepressant: a psychopharmaco-logical animal study. Eur. J. Pharmacol. 160, 395–399.
Matsumoto, K., Ojima, K., Watanabe, H., 1997. Central corticotro-pin-releasing factor and benzodiazepine receptor systems areinvolved in the social isolation stress-induced decrease in ethanolsleep in mice. Brain Res. 753, 318–321.
McAdoo, B.C., Doering, C.H., Kraemer, H.C., Dessert, N., Brodie,H.K., Hamburg, D.A., 1978. A study of the effects of gonadotro-pin-releasing hormone on human mood and behavior. Psychosom.Med. 40, 199–209.
Merchenthaler, I., Gorcs, T., Setalo, G., Petrusz, P., Flerko, B., 1984.Gonadotropin-releasing hormone (GnRH) neurons and pathwaysin the rat brain. Cell Tissue Res. 237, 15–29.
Ogilvie, K.M., Rivier, C., 1997. Gender difference in hypothalamic–pituitary–adrenal axis response to alcohol in the rat: activationalrole of gonadal steroids. Brain Res. 766, 19–28.
Pan, J.T., Kow, L.M., Pfaff, D.W., 1988. Modulatory actions ofluteinizing hormone-releasing hormone on electrical activity ofpreoptic neurons in brain slices. Neuroscience 27, 623–628.
Papakostas, G.I., Trivedi, M.H., Alpert, J.E., Seifert, C.A., Krishen,A., Goodale, E.P., Tucker, V.L., 2008. Efficacy of bupropion andthe selective serotonin reuptake inhibitors in the treatment ofanxiety symptoms in major depressive disorder: a meta-analysis ofindividual patient data from 10 double-blind, randomized clinicaltrials. J. Psychiat. Res. 42, 134–140.
Park, Y., Park, S.D., Cho, W.K., Kim, K., 1988. Testosteronestimulates LHRH-like mRNA level in the rat hypothalamus. BrainRes. 451, 255–261.
Pasha, K.V., Vijayan, E., 1992. Acute and short-term effects ofintraventricular injection of somatostatin and LHRH on glutamateand GABA levels in rat brain. Biochem. Int. 26, 7–15.
Pelleymounter, M.A., Joppa, M., Ling, N., Foster, A.C., 2004.Behavioral and neuroendocrine effects of the selective CRF2receptor agonists urocortin II and urocortin III. Peptides 25,659–666.
Petraglia, F., Sutton, S., Vale, W., Plotsky, P., 1987. Corticotropin-releasing factor decreases plasma luteinizing hormone levels infemale rats by inhibiting gonadotropin-releasing hormone releaseinto hypophysial-portal circulation. Endocrinology 120, 1083–1088.
Ponnusamy, R., Nissim, H.A., Barad, M., 2005. Systemic blockade ofD2-like dopamine receptors facilitates extinction of conditionedfear in mice. Learn. Mem. 12, 399–406.
Porsolt, R.D., Le Pichon, M., Jalfre, M., 1977. Depression: a newanimal model sensitive to antidepressant treatments. Nature 266,730–732.
Rance, N.E., Young III, W.S., McMullen, N.T., 1994. Topography ofneurons expressing luteinizing hormone releasing hormone genetranscripts in the human hypothalamus and basal forebrain. J.Comp. Neurol. 339, 573–586.
Rettori, V., McCann, S.M., 1997. The mechanism of action of alcoholto suppress gonadotropin secretion. Mol. Psychiat. 2, 350–354.
Reubi, J.C., Maurer, R., 1984. Visualization of LHRH receptors in therat brain. Eur. J. Pharmacol. 106, 453–454.
Reubi, J.C., Palacios, J.M., Maurer, R., 1987. Specific luteinizing-hormone-releasing hormone receptor binding sites in hippocampusand pituitary: an autoradiographic study. Neuroscience 21, 847–856.
Rothfeld, J.M., Hejtmancik, J.F., Conn, P.M., Pfaff, D.W., 1987.LHRH messenger RNA in neurons in the intact and castrate malerat forebrain, studied by in situ hybridization. Exp. Brain Res. 67,113–118.
Rubinow, D.R., Roca, C.A., Schmidt, P.J., Danaceau, M.A., Putnam,K., Cizza, G., Chrousos, G., Nieman, L., 2005. Testosteronesuppression of CRH-stimulated cortisol in men. Neuropsycho-pharmacology 30, 1906–1912.
Rudenstein, R.S., Bigdeli, H., McDonald, M.H., Snyder, P.J., 1979.Administration of gonadal steroids to the castrated male ratprevents a decrease in the release of gonadotrophin-releasinghormone from the incubated hypothalamus. J. Clin. Invest. 63,262–267.
Simon, P., Panissaud, C., Costentin, J., 1993. Anxiogenic-like effectsinduced by stimulation of dopamine receptors. Pharmacol. Bio-chem. Behav. 45, 685–690.
Soulairac, M.M.A., Lambinet, H., Neuman, J.C., Schally, A.V., 1983.Action de la luteo-liberina (LHRH). Ann. Med. Psychol. 141, 876.
Soysal, M.E., Soysal, S., Vicdan, K., Ozer, S., 2001. A randomizedcontrolled trial of goserelin and medroxyprogesterone acetate inthe treatment of pelvic congestion. Hum. Reprod. 16, 931–939.
Studd, J., 2006. Ovariectomy for menstrual madness and premenstrualsyndrome-19th century history and lessons for current practice.Gynecol. Endocrinol. 22, 411–415.
Swiergiel, A.H., Zhou, Y., Dunn, A.J., 2007. Effects of chronicfootshock, restraint and corticotropin-releasing factor on freezing,ultrasonic vocalization and forced swim behavior in rats. Behav.Brain Res. 183, 178–187.
Sutton, S.W., Behan, D.P., Lahrichi, S.L., Kaiser, R., Corrigan, A.,Lowry, P., Potter, E., Perrin, M.H., Rivier, J., Vale, W.W., 1995.Ligand requirements of the human corticotropin-releasing factor-binding protein. Endocrinology 136, 1097–1102.
Tellam, D.J., Perone, M.J., Dunn, I.C., Radovick, S., Brennand, J.,Rivier, J.E., Catsro, M.G., Lovejoy, D.A., 1998. Direct regulationof GnRH transcription by CRF-like peptides in an immortalizedneuronal cell line. Neuroreport 9, 3135–3140.
Todorovic, C., Jahna, O., Tezvala, H., Hippela, C., Spiessa, J., 2005.The role of CRF receptors in anxiety and depression: implicationsof the novel CRF1 agonist cortagine. Neurosci. Biobehav. Rev. 29,1323–1333.

410 S.N. Umathe et al. / Neuropeptides 42 (2008) 399–410
Uday, G., Pravinkumar, B., Manish, W., Sudhir, U., 2007. LHRHantagonist attenuates the effect of fluoxetine on marble-buryingbehavior in mice. Eur. J. Pharmacol. 563, 155–159.
Umathe, S.N., Bhutada, P.S., Dixit, P.V., Jain, N.S., 2008a. Leupro-lide – a luteinizing hormone releasing hormone agonist attenuatesethanol withdrawal syndrome and ethanol-induced locomotorsensitization in mice. Neuropeptides (Epub ahead of print),doi:10.1016/j.npep.2007.12.006.
Umathe, S., Bhutada, P., Dixit, P., Shende V., 2008b. Increasedmarble-burying behavior in ethanol-withdrawal state: modulationby gonadotropin-releasing hormone agonist. Eur. J. Pharmacol.(Epub ahead of print).
Umathe, S.N., Dixit, P.V., Wanjari, M.M., Ullewar, M.P., 2008c.Leuprolide – a GnRH agonist, prevents restraint stress-inducedimmunosuppression via sex steroid-independent peripheral mech-anism in mice. Int. Immunopharmacol. 8, 71–79.
Van Honk, J., Peper, J.S., Schutter, D.J., 2005. Testosterone reducesunconscious fear but not consciously experienced anxiety: impli-cations for the disorders of fear and anxiety. Biol. Psychiat. 58,218–225.
West, C.P., Hillier, H., 1994. Ovarian suppression with the gonado-trophin-releasing hormone agonist goserelin (Zoladex) in manage-ment of the premenstrual tension syndrome. Hum. Reprod. 9,1058–1063.
Westphal, N.J., Seasholtz, A.F., 2005. Gonadotropin-releasing hor-mone (GnRH) positively regulates corticotropin-releasing hor-mone-binding protein expression via multiple intracellularsignaling pathways and a multipartite GnRH response element inalpha T3-1 cells. Mol. Endocrinol. 19, 2780–2797.
Williams, C.L., Nishihara, M., Thalabard, J.C., Grosser, P.M.,Hotchkiss, J., Knobil, E., 1990. Corticotropin-releasing factorand gonadotropin releasing hormone pulse generator activity in therhesus monkey. Neuroendocrinology 52, 133–137.
Yatham, L.N., Goldstein, J.M., Vieta, E., Bowden, C.L., Grunze, H.,Post, R.M., Suppes, T., Calabrese, J.R., 2005. Atypical antipsy-chotics in bipolar depression: potential mechanisms of action. J.Clin. Psychiat. 66 (Suppl. 5), 40–48.




















