
Generation of human artificial chromosomes expressing naturally controlled guanosine triphosphate...
12
© Blackwell Science Limited Genes to Cells (2002) 7 , 1021–1032 1021 Blackwell Science, Ltd Oxford, UK GTC Genes to Cells 1356-9597 © Blackwell Science Ltd October 2002 7 10 Original Article Human artificial chromosomes carrying GCH1 M Ikeno et al. Generation of human artificial chromosomes expressing naturally controlled guanosine triphosphate cyclohydrolase I gene Masashi Ikeno 1,2 , Hidehito Inagaki 1 , Keiko Nagata 1,2 , Miwa Morita 1,2 , Hiroshi Ichinose 1 and Tuneko Okazaki 1,2 * 1 Institute for Comprehensive Medical Science, Fujita Health University, Toyoake, Aichi 470-1192, Japan 2 Core Research for Evolutional Science and Technology, Japan Science and Technology Corporation, Kawaguchi, Saitama 332-0012, Japan Abstract Background : Human artificial chromosomes (HACs) are generated from the precursor DNA constructs containing α -satellite DNA with CENP-B boxes, and the process could be used for the incorporation of large genes in the HACs. Guanosine triphosphate cyclohydrolase I (GCH1) is the first and rate-limiting enzyme for the biosynthesis of tetrahydrobiopt- erin, the essential co-factor of aromatic amino acid hydroxylases and nitric oxide synthase. Results : We constructed HACs carrying a 180 kb genome segment encoding the human GCH1 gene and its control region from the bacterial artificial chromosome (BAC) with the GCH1 segment by co-transfection with the α -satellite DNA-containing BAC to a human fibroblast cell line. Two cell lines carrying a HAC with GCH1 genes were obtained. Both HACs were composed of multiple copies of precursor BACs and were maintained stably in human and mouse cell lines. The GCH1 activities of the HAC-carrying human fibroblast cell lines were elevated but still highly sensitive to IFN- γ induction, mimicking the response of the gene expression from the authentic chromosomal genes. Conclusion : These HACs will provide a useful system for analysis of the complex regulatory circuit of the GCH1 gene in vivo and also function as a tool for gene delivery in animal models or in therapeutic trials. Introduction Mitotically stable human artificial chromosomes (HACs), several mega-base pairs in size, are frequently generated de novo in the human fibroblast cell line, HT1080, upon introduction of precursor DNA constructs in either linear (YAC) or circular form (BAC or PAC (p1-derived artificial chromosome)) containing several tens of kilo- bases of human α -satellite (alphoid) DNA with frequent CENP-B boxes (Ikeno et al . 1998; Masumoto et al . 1998; Henning et al . 1999; Ebersole et al . 2000). Since essential kinetochore proteins are detected on such HACs, the input α -satellite arrays are capable of assembling a de novo active centromere/kinetochore structure similar to that of authentic human chromosomes (Ikeno et al . 1994, 1998; Masumoto et al . 1998; Henning et al . 1999; Ebersole et al . 2000; Ando et al . 2002). Since HACs duplicate once every cell cycle utilizing cellular protein factors, they also contain replication origin(s) in the alphoid sequence. Linear HACs made from the alphoid-YAC with telomere sequences acquired a functional telomere structure at the ends of the HACs, but circular HACs made from BAC or PAC had no telomere structure (Ikeno et al . 1998; Ebersole et al . 2000). Treating human diseases by gene therapy is a challeng- ing and promising field. Although we now have at hand tens of thousands of genes by which we might be able to cure defective human genes or to characterize in detail their function and regulation, the major obstacle still lies in the development of effective gene delivery techn- ology. Presently available vectors for mammalian cells are mainly derived from small viruses (Mineta et al . 1995; Fisher et al . 1997; Pfeifer & Verma 2001). Although they have the advantage of highly efficient transduction of the genes of interest (transgenes), their cloning capacity is limited.They are too small to include large genome seg- ments with tissue-specific regulatory regions. Moreover, Communicated by: Fumio Hanaoka * Correspondence : E-mail: [email protected]
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Generation of human artificial chromosomes expressing naturally controlled guanosine triphosphate...

© Blackwell Science Limited
Genes to Cells (2002)
7
, 1021–1032
1021
Blackwell Science, LtdOxford, UKGTCGenes to Cells1356-9597© Blackwell Science LtdOctober 2002710Original ArticleHuman artificial chromosomes carrying GCH1M Ikeno et al.
Generation of human artificial chromosomes expressing naturally controlled guanosine triphosphate cyclohydrolase I gene
Masashi Ikeno
1,2
, Hidehito Inagaki
1
, Keiko Nagata
1,2
, Miwa Morita
1,2
, Hiroshi Ichinose
1
and Tuneko Okazaki
1,2
*
1
Institute for Comprehensive Medical Science, Fujita Health University, Toyoake, Aichi 470-1192, Japan
2
Core Research for Evolutional Science and Technology, Japan Science and Technology Corporation, Kawaguchi, Saitama 332-0012, Japan
Abstract
Background
:
Human artificial chromosomes (HACs)are generated from the precursor DNA constructscontaining
αααα
-satellite DNA with CENP-B boxes,and the process could be used for the incorporationof large genes in the HACs. Guanosine triphosphatecyclohydrolase I (GCH1) is the first and rate-limitingenzyme for the biosynthesis of tetrahydrobiopt-erin, the essential co-factor of aromatic amino acidhydroxylases and nitric oxide synthase.
Results
:
We constructed HACs carrying a 180 kbgenome segment encoding the human GCH1 geneand its control region from the bacterial artificialchromosome (BAC) with the GCH1 segment byco-transfection with the
αααα
-satellite DNA-containing
BAC to a human fibroblast cell line. Two cell linescarrying a HAC with GCH1 genes were obtained.Both HACs were composed of multiple copies ofprecursor BACs and were maintained stably inhuman and mouse cell lines. The GCH1 activities ofthe HAC-carrying human fibroblast cell lines wereelevated but still highly sensitive to IFN-
γγγγ
induction,mimicking the response of the gene expression fromthe authentic chromosomal genes.
Conclusion
:
These HACs will provide a useful systemfor analysis of the complex regulatory circuit ofthe GCH1 gene
in vivo
and also function as a toolfor gene delivery in animal models or in therapeutictrials.
Introduction
Mitotically stable human artificial chromosomes (HACs),several mega-base pairs in size, are frequently generated
de novo
in the human fibroblast cell line, HT1080, uponintroduction of precursor DNA constructs in eitherlinear (YAC) or circular form (BAC or PAC (p1-derivedartificial chromosome)) containing several tens of kilo-bases of human
α
-satellite (alphoid) DNA with frequentCENP-B boxes (Ikeno
et al
. 1998; Masumoto
et al
. 1998;Henning
et al
. 1999; Ebersole
et al
. 2000). Since essentialkinetochore proteins are detected on such HACs, the input
α
-satellite arrays are capable of assembling a
de novo
activecentromere/kinetochore structure similar to that ofauthentic human chromosomes (Ikeno
et al
. 1994, 1998;Masumoto
et al
. 1998; Henning
et al
. 1999; Ebersole
et al
. 2000; Ando
et al
. 2002). Since HACs duplicate
once every cell cycle utilizing cellular protein factors,they also contain replication origin(s) in the alphoidsequence. Linear HACs made from the alphoid-YACwith telomere sequences acquired a functional telomerestructure at the ends of the HACs, but circular HACsmade from BAC or PAC had no telomere structure(Ikeno
et al
. 1998; Ebersole
et al
. 2000).Treating human diseases by gene therapy is a challeng-
ing and promising field. Although we now have at handtens of thousands of genes by which we might be able tocure defective human genes or to characterize in detailtheir function and regulation, the major obstacle still liesin the development of effective gene delivery techn-ology. Presently available vectors for mammalian cells aremainly derived from small viruses (Mineta
et al
. 1995;Fisher
et al
. 1997; Pfeifer & Verma 2001). Although theyhave the advantage of highly efficient transduction of thegenes of interest (transgenes), their cloning capacity islimited. They are too small to include large genome seg-ments with tissue-specific regulatory regions. Moreover,
Communicated by:
Fumio Hanaoka*
Correspondence
: E-mail: [email protected]
GTC_580.fm Page 1021 Monday, September 9, 2002 7:09 PM
M Ikeno
et al.
Genes to Cells (2002)
7
, 1021–1032
© Blackwell Science Limited
1022
transgenes are usually stably maintained only afterrandom integration into host-cell chromosomes, geneexpression from which is usually unpredictable (mostlysuppressed) and not under the control of the authenticregulatory region of the genes. Even worse, this step hasthe potential to induce unfavourable mutagenesis.
In contrast, HACs have the capacity to accommodatea large transgene with a controlling region in excess of100 kb of DNA. HACs containing transgenes aregenerated
de novo
from a precursor construct with boththe transgene and an alphoid array (Mejia
et al
. 2001), orfrom precursor constructs containing an alphoid array andthe transgene in separate entities (Grimes
et al
. 2001). Thus,HACs may be used not only as vectors in therapeuticapplications but also as model systems useful in analysesof tissue or organ specific regulation of gene expressionthat would only be possible with large genome segments.
Considering not only basic studies into gene regulationbut also future therapeutic applications, we constructed
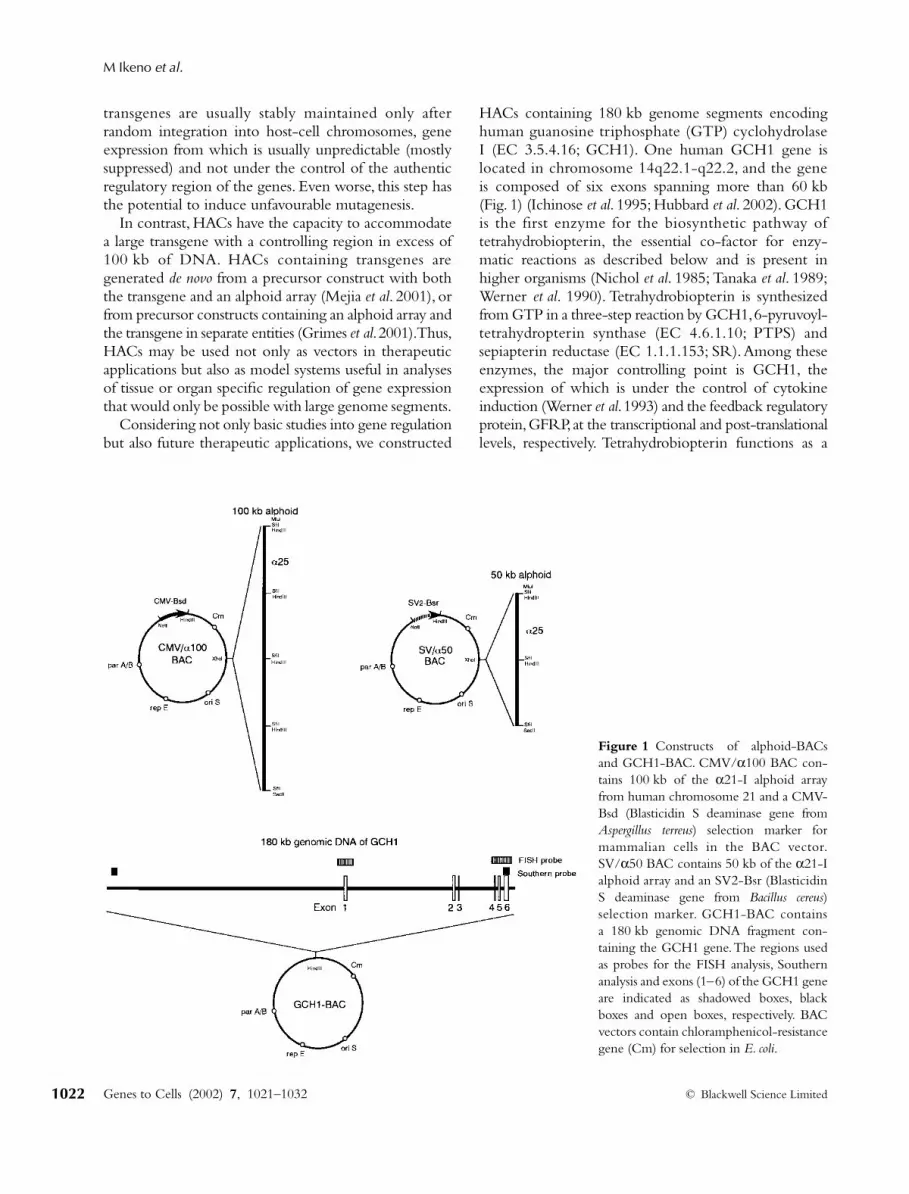
HACs containing 180 kb genome segments encodinghuman guanosine triphosphate (GTP) cyclohydrolaseI (EC 3.5.4.16; GCH1). One human GCH1 gene islocated in chromosome 14q22.1-q22.2, and the geneis composed of six exons spanning more than 60 kb(Fig. 1) (Ichinose
et al
. 1995; Hubbard
et al
. 2002). GCH1is the first enzyme for the biosynthetic pathway oftetrahydrobiopterin, the essential co-factor for enzy-matic reactions as described below and is present inhigher organisms (Nichol
et al
. 1985; Tanaka
et al
. 1989;Werner
et al
. 1990). Tetrahydrobiopterin is synthesizedfrom GTP in a three-step reaction by GCH1, 6-pyruvoyl-tetrahydropterin synthase (EC 4.6.1.10; PTPS) andsepiapterin reductase (EC 1.1.1.153; SR). Among theseenzymes, the major controlling point is GCH1, theexpression of which is under the control of cytokineinduction (Werner
et al
. 1993) and the feedback regulatoryprotein, GFRP, at the transcriptional and post-translationallevels, respectively. Tetrahydrobiopterin functions as a
Figure 1 Constructs of alphoid-BACsand GCH1-BAC. CMV/α100 BAC con-tains 100 kb of the α21-I alphoid arrayfrom human chromosome 21 and a CMV-Bsd (Blasticidin S deaminase gene fromAspergillus terreus) selection marker formammalian cells in the BAC vector.SV/α50 BAC contains 50 kb of the α21-Ialphoid array and an SV2-Bsr (BlasticidinS deaminase gene from Bacillus cereus)selection marker. GCH1-BAC containsa 180 kb genomic DNA fragment con-taining the GCH1 gene. The regions usedas probes for the FISH analysis, Southernanalysis and exons (1–6) of the GCH1 geneare indicated as shadowed boxes, blackboxes and open boxes, respectively. BACvectors contain chloramphenicol-resistancegene (Cm) for selection in E. coli.
GTC_580.fm Page 1022 Monday, September 9, 2002 7:09 PM

Human artificial chromosomes carrying GCH1
© Blackwell Science Limited
Genes to Cells (2002)
7
, 1021–1032
1023
natural co-factor of the aromatic amino acid hydrox-ylases, phenylalanine hydroxylase (EC 1.14.16.2; PAH),tyrosine hydroxylase (EC 1.14.16.3; TH), the first andrate-limiting enzyme of dopamine synthesis, tryptophan5-hydroxylase (EC 1.14.16.4; TPH), serotonin biosyn-thesis. Tetrahydrobiopterin is also essential for all threeforms of nitric oxide synthase (NOS) (Kaufman 1993).Decreases in GCH1 activity, tetrahydrobiopterin leveland/or TH activity causes a dopamine deficiency in thenigrostriatum dopamine neurones and provokes severalwell-known clinical symptoms, such as hereditary dopa-responsive dystonia (DRD/Segawa’s syndrome) (Ichi-nose
et al
. 1994) or parkinsonism. Thus, HACs carryingthe GCH1 gene with the authentic regulatory regionwould almost certainly prove useful for compensatingfor the defects in the GCH1 gene as well as facilitatinga close study of the complex regulatory mechanism ofGCH1 gene expression
in vivo
.In this study, we co-transfected the GCH1-BAC
containing a 180 kb genomic region covering the entireGCH1 gene and its upstream control region and thealphoid-BAC containing a 50 kb or 100 kb alphoidarray to the HT1080 cells, and generated HACs con-taining multiple copies of GCH1 genes. We were ableto show that the HACs are stably maintained both inhuman and mouse cells without selection. GCH1 activ-ity arising from human fibroblast cells with a GCH1-HAC is elevated, but when interferon-
γ
(IFN-
γ
) is addedto the culture medium, GCH1 activity is further up-regulated, as is observed with the gene expression fromthe host cell chromosomes. These results show that theGCH1-HAC will not only provide a good system forstudy of the complex regulatory mechanism of theGCH1 gene in cells and bodies but will also be useful asa gene delivery tool for animal models or in therapeutic
trials. The HAC vector system will open the simple andgeneral way to the delivery of very large genes with anoriginal regulatory region.
Results
Generation of HAC containing the GCH1 genomic locus
α
21-I alphoid, consisting of an 11mer higher orderrepeat unit derived from human chromosome 21 (Ikeno
et al
. 1994), is able to generate a HAC efficiently whenit is introduced into HT1080 cells (Ikeno
et al
. 1998).We generated HACs containing a GCH1 genomic locuswith naturally regulated gene expression, utilizingalphoid-BACs and GCH1-BAC. The BACs used in thisstudy are shown in Fig. 1. CMV/
α
100 contains 100 kbof an
α
21-I alphoid array and a CMV-Bsd as a selectablemarker, and SV/
α
50 contains 50 kb of an
α
21-I alphoidarray and a SV2-Bsr selection marker. The GCH1-BACwas obtained from a BAC library (Genome systems)and has a 180 kb genomic DNA fragment containingthe GCH1 gene. We co-transfected either one of thealphoid-BACs and the GCH1-BAC in a 1: 1 molecularratio into HT1080 cells by lipofection and isolatedBlasticidin S (BS) resistant cell lines after 10 days. To detectthe presence of HAC as an extrachromosomal element,the BS-resistant cell lines were analysed by FISH usingboth
α
21-I alphoid DNA and BAC vector as probes.One out of 16 transformed cell lines obtained by the co-transfection of CMV/
α
100 and GCH1-BAC, and threeout of 17 cell lines obtained by the co-transfection ofSV/
α
50 and GCH1-BAC, contained one copy of HACper nuclei in more than 95% of the inspected cells(Table 1). In the remaining cell lines, introduced BACs
Table 1 The fates of co-transfected BACs in the transformed cell lines
Input BACsBSr cell lines inspected
Alphoid and BAC signals on
HAC Chromosome Non HAC with GCH1
CMV/α100 BAC 16 1 4 11 1 (HT/GCH2-10)+GCH1-BAC
SV/α50 BAC 17 3 13 1 1 (HT/GCH5-18)+GCH1-BAC
BS-resistant cell lines obtained by co-transfection of GCH1-BAC plus CMV/α100 BAC or SV/α50 BAC were analysed by FISH. ‘HAC’indicates cell lines with an artificial chromosome detected both with α21-I alphoid DNA and BAC vector probes. One copy of HACwas detected in more than 95% of the inspected metaphase spread of these cell lines. In the remaining cell lines, introduced BACs wereeither integrated into the chromosomes of HT1080 (Chromosome) or the signals were undetectable (Non) by FISH analysis. ‘HAC withGCH1’ indicates the cell lines carrying a HAC with signals for the GCH1 gene.
GTC_580.fm Page 1023 Monday, September 9, 2002 7:09 PM
M Ikeno
et al.
Genes to Cells (2002)
7
, 1021–1032
© Blackwell Science Limited
1024
were either integrated into the chromosomes ofHT1080 or the signals were undetectable by FISHanalyses.
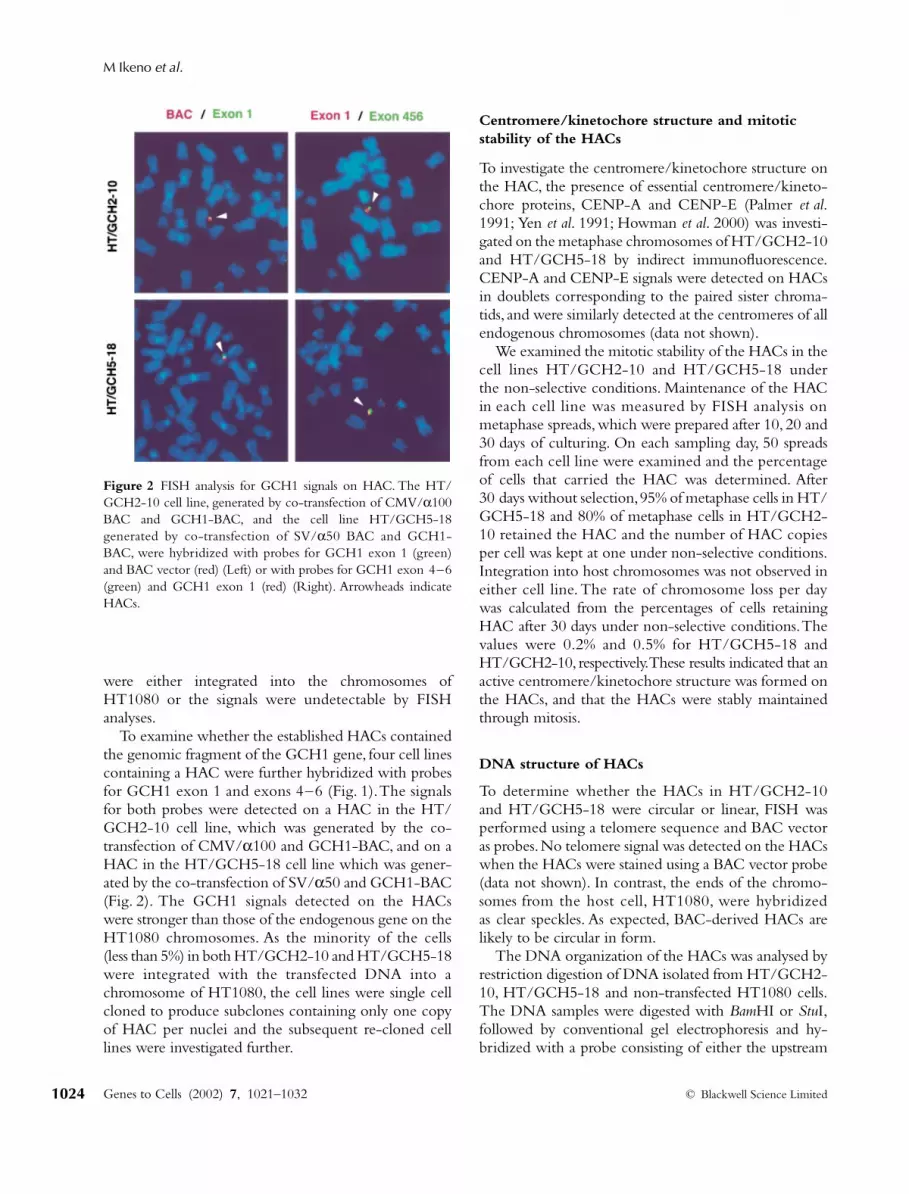
To examine whether the established HACs containedthe genomic fragment of the GCH1 gene, four cell linescontaining a HAC were further hybridized with probesfor GCH1 exon 1 and exons 4–6 (Fig. 1). The signalsfor both probes were detected on a HAC in the HT/GCH2-10 cell line, which was generated by the co-transfection of CMV/
α
100 and GCH1-BAC, and on aHAC in the HT/GCH5-18 cell line which was gener-ated by the co-transfection of SV/
α
50 and GCH1-BAC(Fig. 2). The GCH1 signals detected on the HACswere stronger than those of the endogenous gene on theHT1080 chromosomes. As the minority of the cells(less than 5%) in both HT/GCH2-10 and HT/GCH5-18were integrated with the transfected DNA into achromosome of HT1080, the cell lines were single cellcloned to produce subclones containing only one copyof HAC per nuclei and the subsequent re-cloned celllines were investigated further.
Centromere/kinetochore structure and mitotic stability of the HACs
To investigate the centromere/kinetochore structure onthe HAC, the presence of essential centromere/kineto-chore proteins, CENP-A and CENP-E (Palmer
et al
.1991; Yen
et al
. 1991; Howman
et al
. 2000) was investi-gated on the metaphase chromosomes of HT/GCH2-10and HT/GCH5-18 by indirect immunofluorescence.CENP-A and CENP-E signals were detected on HACsin doublets corresponding to the paired sister chroma-tids, and were similarly detected at the centromeres of allendogenous chromosomes (data not shown).
We examined the mitotic stability of the HACs in thecell lines HT/GCH2-10 and HT/GCH5-18 underthe non-selective conditions. Maintenance of the HACin each cell line was measured by FISH analysis onmetaphase spreads, which were prepared after 10, 20 and30 days of culturing. On each sampling day, 50 spreadsfrom each cell line were examined and the percentageof cells that carried the HAC was determined. After30 days without selection, 95% of metaphase cells in HT/GCH5-18 and 80% of metaphase cells in HT/GCH2-10 retained the HAC and the number of HAC copiesper cell was kept at one under non-selective conditions.Integration into host chromosomes was not observed ineither cell line. The rate of chromosome loss per daywas calculated from the percentages of cells retainingHAC after 30 days under non-selective conditions. Thevalues were 0.2% and 0.5% for HT/GCH5-18 andHT/GCH2-10, respectively. These results indicated that anactive centromere/kinetochore structure was formed onthe HACs, and that the HACs were stably maintainedthrough mitosis.
DNA structure of HACs
To determine whether the HACs in HT/GCH2-10and HT/GCH5-18 were circular or linear, FISH wasperformed using a telomere sequence and BAC vectoras probes. No telomere signal was detected on the HACswhen the HACs were stained using a BAC vector probe(data not shown). In contrast, the ends of the chromo-somes from the host cell, HT1080, were hybridizedas clear speckles. As expected, BAC-derived HACs arelikely to be circular in form.
The DNA organization of the HACs was analysed byrestriction digestion of DNA isolated from HT/GCH2-10, HT/GCH5-18 and non-transfected HT1080 cells.The DNA samples were digested with
Bam
HI or
Stu
I,followed by conventional gel electrophoresis and hy-bridized with a probe consisting of either the upstream
Figure 2 FISH analysis for GCH1 signals on HAC. The HT/GCH2-10 cell line, generated by co-transfection of CMV/α100BAC and GCH1-BAC, and the cell line HT/GCH5-18generated by co-transfection of SV/α50 BAC and GCH1-BAC, were hybridized with probes for GCH1 exon 1 (green)and BAC vector (red) (Left) or with probes for GCH1 exon 4–6(green) and GCH1 exon 1 (red) (Right). Arrowheads indicateHACs.
GTC_580.fm Page 1024 Monday, September 9, 2002 7:09 PM

Human artificial chromosomes carrying GCH1
© Blackwell Science Limited
Genes to Cells (2002)
7
, 1021–1032
1025
region of GCH1 (US) or GCH1 exon 6. The size of the
Bam
HI fragments detected by the US probe were 5.0 kbfrom the endogenous GCH1 gene and 3.5 kb fromGCH1-BAC. The 5.0 and 3.5 kb fragments weredetected with DNA from HT/GCH2-10 and HT/GCH5-18 at almost the same signal intensity (Fig. 3A).The size of the
Stu
I fragments detected by the exon 6probe were 24.5 kb from the endogenous GCH1 geneand 14.4 kb from GCH1-BAC. The 24.5 and 14.4 kbfragments were detected with DNA from HT/GCH5-
18 at almost the same signal intensity, while three frag-ments heterogeneous in size were detected in additionto the endogenous fragment with the DNA from HT/GCH2-10 (Fig. 3B). The results indicated that GCH1-containing HACs in HT/GCH5-18 were established bythe assembly of about three copies of transfected GCH1-BAC DNA, since the karyo-type of HT1080 cells usedin this study is 3n, while HT/GCH2-10 was accompa-nied by some rearrangements of the terminal regionof GCH1 exon 6, but it may also contain three copies of
Figure 3 Structural analysis of GCH1-HAC. (A, B) Restriction analysis of GCH1genes in HACs. Genomic DNAs fromHT/GCH2-10, HT/GCH5-18 and non-transfected HT1080 were digested withBamHI (A) or StuI (B) and fractionated byconventional gel electrophoresis. Theexpected size of the BamHI and StuIfragments detected by the US probe(A) and the exon 6 probe (B), respectively,using the endogenous GCH1 locus andGCH1-HAC, are shown at the top. (C)Estimation of the copy number of GCH1-BAC and alphoid BAC in the HACs bydot hybridization. Left: The intensity valueobtained with the GCH1 exon 6 probe.Input DNA of GCH1-BAC (0.4, 0.2,0.1 ng) and genomic DNA (1.0, 0.5 µg)from HT1080, HT/GCH2-10 andHT/GCH5-18 were hybridized with theGCH1 exon 6 probe. The value obtainedwith 0.1 ng GCH1-BAC DNA was usedas a standard. Right: The intensity valueobtained with BAC vector probe. GCH1-BAC (0.5, 0.1, 0.05 ng) and genomic DNA(0.5, 0.25 µg) from HT1080, HT/GCH2-10 and HT/GCH5-18 were hybridizedwith the BAC vector probe. The signalintensity obtained with each probe wasdetermined using a Fuji image-analyserBAS1000.
GTC_580.fm Page 1025 Monday, September 9, 2002 7:09 PM

M Ikeno
et al.
Genes to Cells (2002)
7
, 1021–1032
© Blackwell Science Limited
1026
GCH1-BAC as judged by the density of the US band.The internal rearrangements of GCH1 genes wereconfirmed by the RT-PCR analyses of GCH1 transcriptsin HT/GCH2-10, which revealed the synthesis of abnor-mal transcripts (data not shown).
The copy numbers of the GCH1-BAC and thealphoid-BAC in GCH2-10 and GCH5-18 HACs weredetermined by dot hybridization using GCH1 exon 6and BAC vector, respectively, as probes. Relative copynumbers of each BAC in the HACs were estimatedfrom the hybridization signal-intensity values, which weredetermined using each DNA probe and standardizedusing the values obtained with 0.1 ng GCH1-BACDNA (Fig. 3C). In the case where GCH1 exon 6 was usedas the probe, the same hybridization intensity values tothose obtained with 0.1 ng GCH1-BAC DNA wereobtained with 0.5
µ
g DNA from both HT/GCH2-10and HT/GCH5-18, and with 1
µ
g DNA from HT1080(Fig. 3C, left). Since the HT1080 karyo-type used inthis study is 3n, three copies of GCH1 genes occuron its chromosomes, and given that HT/GCH2-10 andHT/GCH5-18 resulted in the same signal values withhalf the amount DNA as that from HT1080, they mustcontain six copies of GCH1 genes; three on the chro-mosomes and three on the HAC. The total copy numberof BACs was estimated from the intensity values obtainedwith the BAC vector probe. The same hybridizationintensity values to that obtained with 0.1 ng GCH1-BAC were obtained with 0.33
µ
g DNA from HT/GCH2-10 and HT/GCH5-18, while HT1080 showedno signal, as expected (Fig. 3C, Right). Therefore, bothHACs have roughly threefold more copies of the BACvector than copies of the GCH1 gene. Thus, copy numbersof the total BAC vectors must be approximately nine percell; three copies of GCH1 genes are in the form of GCH1-
BAC and the remaining six copies of BACs must exist inthe form of alphoid-BAC in both HACs.
Transfer of HAC-containing cell lines to mouse A9 cells
De novo
HAC formation using cloned alphoid DNAhas been successful in the human fibrocarcinoma cell lineHT1080. To determine the natural expression of theGCH1 gene in the neural cell line, the HAC that hasbeen established in the HT1080 cell line needs to betransferred into a neural cell line.
Fusion of the HAC-containing cell lines and mouseA9 cell line was performed using PEG which allowsmicrocell mediated chromosome transfer (MMCT)(Fournier & Ruddle 1977). BS- and Ouabain-resistantcell lines were analysed by FISH. Metaphase spreadswere hybridized with a BAC vector probe and Alu repeatprobe to identify the HACs and human chromosomes,respectively (Fig. 4A). One of the fusion cell lines,F/GCH5-18, contained one or two copies of HACtogether with 8 to 10 human chromosomes.
The HACs in the fusion cells were maintained stablyduring mitotic growth under non-selective conditionswith a loss of
≈
1% of the mitotic chromosomes perday (data not shown). The mitotic stability of humanchromosomes in mouse cell lines was sometimes caused bythe acquisition of minor satellite DNA from the mousewhich was localized at the centromere of the mousechromosomes and may serve as functional centromeresequences (Shen
et al
. 1997). Therefore the presence ofmouse minor satellite DNA on HAC was examined byFISH. Signals of minor satellite DNA were not detectedon HAC, while strong signals were detected at thecentromeres of mouse chromosomes (Fig. 4B). The
Figure 4 Cell fusion of HAC-containingcell line with mouse A9 cells. HT/GCH5-18 cell lines were fused with A9 cellsmediated by PEG. BS- and Ouabain-resistant cell lines were analysed by FISH.Metaphase spreads were hybridized withthe BAC vector probe (red) and an Alurepeat probe (green) (A) or hybridizedwith the BAC vector probe (green) and amouse minor satellite probe (red) (B). Arrowsindicate HACs.
GTC_580.fm Page 1026 Monday, September 9, 2002 7:09 PM

Human artificial chromosomes carrying GCH1
© Blackwell Science Limited Genes to Cells (2002) 7, 1021–1032 1027
fusion cell lines were able to form microcells under col-cemid treatment conditions (data not shown). Therefore,the HACs could easily be transferred to neural cell lines.
GCH1 expression from HAC
Naturally regulated gene expression was expected fromthe transgenes in the large genome segments carried byHACs. The GCH1-BAC used in the generation ofHACs contained over 100 kb of genomic sequence fromthe 5′ upstream region of the GCH1 exon 1. Therefore,we have measured GTP cyclohydrolase I (GCH1) activ-ities in HT1080 and the HAC-containing derivativesthat were developed from it. It would have beenexpected from the previous report that the activity ofGCH1 would have been hardly detectable in fibroblastcell lines, but up-regulated by induction of IFN-γ(Werner et al. 1990). GCH1 activity in HT1080, HT/GCH2-10 and HT/GCH5-18 were analysed in thepresence and absence of IFN-γ induction (Table 2).
HT1080 without GCH1-HAC exhibited barelydetectable levels of GCH1 activity in the absence ofIFN-γ induction, while the activity was increased15-fold following the addition of IFN-γ. In the HT/GCH2-10 cell line in the absence of IFN-γ induction,the GCH1 level was three times the value of HT1080without a HAC. After the IFN-γ induction, a nearly30-fold up-regulation was observed. In contrast, GCH1activity in HT/GCH5-18 was elevated 70-fold in theabsence of IFN-γ, and the addition of IFN-γ further up-regulated the activity fivefold. In both HT/GCH-HACcell lines, the GCH1 activities were elevated but differin degree, possibly reflecting the difference in chromatinstructure and/or DNA rearrangements in HACs. Theyare still susceptible to IFN-γ induction, just like theresponse of the expression of the GCH1 gene from theauthentic chromosome.
DiscussionThe generation of HAC containing large DNA fragments
We obtained HACs containing large DNA fragmentswith the GCH1 gene (GCH1-HAC) by simple co-transfection methods using alphoid-BAC and GCH1-BAC at a DNA ratio of 1 : 1. The GCH1-HAC wasmaintained at one copy after 30 or more rounds of gen-eration under non-selective conditions, in spite of beingcircular in form without telomeres, indicating that HACreplicates once in each cell cycle and is segregatedprecisely into daughter cells. Therefore, the circular HACsin this study did not cause a topological problem, whichmay result in the abnormal segregation of the circularchromosomes, since the catenated form arose fromDNA replication. The HACs were cytologically mega-bases in size and approximately 10-fold larger than thetransfected BAC DNA.
The DNA structure of the HAC was examined tounderstand the properties and mechanism of de novogeneration of HAC. Restriction analysis of the wholearea of the GCH1 gene was difficult because almostall rare-cutting enzyme sites in the BAC constructs weresubjected to methylation and the cell lines contain theendogenous GCH1 locus. Therefore, we applied restric-tion analysis to the region corresponding to the junctionof the BAC vector and the GCH1 locus. The resultshowed that GCH1-HACs in the two cell lines con-tained three copies of the GCH1-BAC and six copiesof the alphoid-BAC as components (Fig. 3). Although theexact mechanism of the formation of the HAC remainsunknown, GCH1-HAC was composed of a multimer ofthe input DNA, which was similar to the HAC gener-ated by alphoid-YAC (Ikeno et al. 1998) or alphoid-BACalone (data not shown), as an input element. The factthat the formation of HAC was accompanied by anassembly of the distinct BACs indicated that the mul-timerization of BAC molecules might be mediated by anonspecific recombination of input BAC in addition tothe amplification of BAC DNA itself.
Generation of the HAC containing large humangenomic DNA was previously reported using a 140 kbor 162 kb HPRT locus (Grimes et al. 2001; Mejia et al.2001). They obtained the HAC containing HPRT genein the HPRT-deficient HT1080 cell lines in HAT mediumdepending on complementation. The feasibility of suchan approach for genes with tissue and stage specificexpression (i.e. not house-keeping genes) will be low inHT1080 cells. In this study, we found that as little as50 kb of α21-I alphoid DNA in BAC was able to gen-erate HACs (centromere/kinetochore) and that the BACs
Table 2 GCH1 activity in HAC-containing cell lines
GCH1 activity (pmol/h/mg protein)
Ratio of increaseCell lines –IFN +IFN
HT1080 0.05 ± 0.02 0.76 ± 0.03 15HT/GCH2-10 0.16 ± 0.02 5.0 ± 0.2 31HT/GCH5-18 3.4 ± 0.2 16.5 ± 0.6 4.8
GCH1 activity of HT/GCH2-10, HT/GCH5-18 and HT1080cells was measured in the presence or absence of IFN-γ induction.Data are mean ± SD values from three independent experiments.
GTC_580.fm Page 1027 Monday, September 9, 2002 7:09 PM

M Ikeno et al.
Genes to Cells (2002) 7, 1021–1032 © Blackwell Science Limited1028
containing a large transgene without selection markerscould be incorporated efficiently into the HAC, since50% of the HACs included the transgene. Thus, HACscontaining any large genomic region of interest could begenerated using alphoid-BAC containing 50 kb ofalphoid DNA and a readily available BAC library with-out any modification. The completeness of the incorpo-rated transgenes may be checked after HAC generation.
Selection of transformants with the CMV promoter-driven Bsd gene increases the number of BS-resistantcells, but the FISH signals for HACs or integrated loci onthe host chromosomes were not found in the majority oftransformants (Table 1). Southern hybridization analysesusing alphoid and BAC vector sequences as probes indi-cated that these cell lines only have Bsd genes integratedin the chromosomes. Thus, the selection marker, drivenby a high expression promoter, was not suitable for thescreening of HAC-containing cell lines.
Centromere/kinetochore structure of HAC
Gene expression was affected by chromosome structure.The insertion of a transgene into a chromosome oftenresults in stably inherited gene silencing in a clonalsubpopulation of the cells, a phenomenon which iscommonly known as position effect variegation (PEV)(Karpen 1994). Recent molecular analysis showed thatthe methylation of histone H3 on lysine 9 contributes tothe targeting of HP1 to the chromatin and results in het-erochromatinization and the silencing of gene expression(Platero et al. 1995; Bannister et al. 2001; Lachner et al.2001). Gene silencing at or near the centromere/kine-tochore in yeast and fly was also reported (Karpen &Allshire 1997), and was expected to occur in mammaliancells. We have recently demonstrated that in the alphoidarray of a HAC, once the centomere/kinetochore struc-ture has formed, the expression of the short markergenes inserted into the HACs were repressed strongly,even if they were driven by strong promoters (T. Abe,K. Yamamoto & T. Okazaki, submitted). Thus, to getexpression of transgenes in HACs, we will need to solvethe topological problem of the genes in relation to thecentromere/kinetochore structure.
The GCH1 gene expression from HAC might becorrelated with the chromatin structure at or near theGCH1 locus. We addressed whether the centromere/kinetochore structure was formed on only the alphoidarray or whether it spread into the GCH1 locus. SinceCENP-A is an essential protein for a functional cen-tromere/kinetochore and constitutes the histone com-ponent for centromere specific nucleosomes (Palmer et al.1991; Howman et al. 2000), we analysed the chromatin
structure on HAC by ChIP using an anti-CENP-A anti-body (Ando et al. 2002). In such analyses using HeLa andHT1080 cells, the alphoid array was enriched to 60–80%in total immunoprecipitated DNA. The alphoid array inthe GCH1-HAC was also enriched by anti-CENP-Aantibody, while the GCH1 region was not. In contrast,the BAC vector sequence about 3 kb away from thealphoid sequence was also immunoprecipitated, indi-cating that the centromere/kinetochore structure wasformed on the alphoid array and spread to the flankingnonalphoid region (data not shown). Invasion of theGCH1 locus in the HAC by centromere/kinetochorestructure was prevented by an as yet unknown protectionmechanism that probably resides in the upstream regula-tory sequence.
Authentic gene expression from HAC
GCH1 encodes the first and rate-limiting enzymefor the biosynthetic pathway of tetrahydrobiopterin(Nichol et al. 1985), the co-factor of aromatic aminoacid hydroxylase (PAH, TH, TPH), as well as nitric oxidesynthase (NOS), and is present in higher organisms(Kaufman 1993). The GCH1 gene is a causative genefor dopamine deficiency in dopa responsive dystonia(DRD/Segawa’s disease) (Ichinose et al. 1994). Defi-ciency of GCH1 in conjunction with a mutation in theTH gene results in severe early onset dystonia/parkin-sonism (Ichinose et al. 1999).
Although only limited analyses have been performedon the upstream regulatory sequence of the humanand mouse GCH1 gene, it has been reported that theCCAAT and TATA boxes are conserved (Ichinoseet al. 1995; Hubbard et al. 2002). It was established thatGCH1 gene expression could be induced by IFN-γ invarious rodent and human cells (Werner et al. 1990).However, the exact mechanism involved in IFN-γ signaltransduction is as yet unknown.
Some gene expression, such as that for human β-globin, is regulated by the locus control regions (LCRs)responsible for initiating and maintaining a stable tissue-specific open chromatin structure (Festentein et al.1996; Milot et al. 1996). The GCH1-HACs used in thisstudy carry a 180 kb genomic fragment containing theGCH1 gene, and may therefore contain the regulatorysequences required for tissue specific expression and forprevention of the silencing effect of the flanking centro-mere. The expression of the GCH1 gene from HACwas measured by GTP cyclohydrolase I activity in thepresence and absence of IFN-γ (Table 2). Activity in theHAC-containing cell line, HT/GCH2-10, was slightlyhigher than the activity obtained with HT1080 in
GTC_580.fm Page 1028 Monday, September 9, 2002 7:09 PM

Human artificial chromosomes carrying GCH1
© Blackwell Science Limited Genes to Cells (2002) 7, 1021–1032 1029
the absence of IFN-γ. Addition of IFN-γ increasedthe GCH1 activity approximately 30-fold. In anothercell line, HT/GCH5-18, which also carries twice thenumber of GCH1 genes as HT1080, showed a 70-foldhigher enzyme activity than TH1080 in the absence ofIFN-γ and the activity was further increased fivefold byIFN-γ induction. The small difference in values betweenHT/GCH2-10 and HT1080 may correspond to thesmall copy numbers of the intact GCH1 genes, since itseems that some copies of GCH1 genes on GCH2-10HAC have the structural abnormality described in theResults section. It will require further study to deter-mine the exact reason why the basal level of the GCH1activity in HT/GCH5-18 is higher. One possibility,however, might be the elevated copy numbers of GCH1loci on the HAC, since there is a greater likelihood thatchromatin assembly on some of the copies might beimprecise. These results also indicated that, although thegene expression of GCH1 may be affected by the differ-ence of chromatin structure assembled on the GCH1locus in the HAC, the genes still responded to IFN-γ.Considering that the value of the GCH1 activity ofIFN-γ induced human macrophage was 240 pmol/h/mg (Werner et al. 1990) and that of LPS inducedmurine neuroblastoma cell line N1E was more than300 pmol/h/mg (Ota et al. 1996), the final levels ofGCH1 activity after IFN-γ induction in the cell lines werestill repressed and kept to a similar order of magnitude.This might indicate the presence of complex cel-lular regulation systems to keep GCH1 activity in theproper range. The GCH1-HAC should prove to be asuitable system for understanding the complex regula-tory mechanisms of GCH1 expression in vivo. This isthe first report in which gene expression on the HACwas quantitatively analysed and the results positively sup-ported the HAC system.
Utility of HAC as a gene transfer vector for mammalian cells
The adeno-associated virus (AAV) vector has often beenused for gene therapy in a helper virus-dependent man-ner for productive infection. The AAV vector has a lim-ited cloning capacity that usually carries cDNA withoutthe original regulatory sequence for gene expression(Dong et al. 1996). GCH1 is necessary for efficientdopamine production together with tyrosine hydro-xylase (TH) and aromatic-l-amino acid decarboxylase(AADC). The expression of these three enzymes fromthe AAV vector in the striatum resulted in relativelylong-term behavioural recovery in a primate model ofParkinson’s disease (Muramatsu et al. 2002).
Recently, an Epstein-Barr virus (EBV)-based episomalvector was reported that was capable of transferringan HPRT gene (115 kb) to some mammalian cell lines,in which the expression was not silenced (Wade-Martinset al. 2000). However, EBV-based vectors are lost morerapidly than HAC in the absence of selection and theirreplication is reliant on the presence of the viral trans-activator, EBNA1. Safety in clinical gene therapy withEBV vectors requires further investigation. HACs mayovercome the above problems as gene transfer vectorsand have further advantages in term of safety. HACs car-rying a long genomic locus in this study were maintainedextrachromosomally, and expressed regulated levels ofgenes for long periods. Therefore, the HAC containingTH, AADC and GCH1 may offer a potential therapeuticstrategy for Parkinson’s disease.
However, the low efficiency of the de novo generationof HACs from BACs, the requirement of the limited cellline for generation of HACs and the large size of HACspresents a difficulty in the delivery of HACs to cells ortissues at required sites. To utilize the HAC as a genetransfer vector, the HAC that has been established inHT1080 needs to be transferred into suitable cell lines.The HAC could be transferred by MMCT using mouseA9 cells, which enable the formation of microcells(Fournier & Ruddle 1977). We have established mouseA9 cell lines, which maintained HACs stably in mitoticgrowth under non-selective conditions without detect-able structural changes in the HAC. The HACs would beeasily transferrable from A9 to other cell lines.
In this study we have demonstrated the generation ofa HAC containing the GCH1 gene, together with itsoriginal regulatory region, from a GCH1-BAC by co-transfection with the alphoid-BAC. The GCH1-HACsexpressed GCH1 genes in a regulated manner and thusproved to be a good system for study of the regulatorymechanism of the GCH1 gene in vivo. Further study onthe GCH1-HACs will reveal the minimum numberof alphoid arrays used to assemble centromere/kineto-chore, the structure of the upstream region required forthe regulated expression of the GCH1 gene and thesites of action of transcription factors on the regulatoryregion. The results we have obtained also indicated thatthe GCH1-HAC may also serve as a gene delivery toolin animal models or future therapeutic trials.
Experimental proceduresConstruction of alphoid-BAC
pBAC-TAN was created by the insertion of a MluI-SfiI-SacIIlinker into the XhoI site of pBeloBAC11. pBAC-CMV and
GTC_580.fm Page 1029 Monday, September 9, 2002 7:09 PM

M Ikeno et al.
Genes to Cells (2002) 7, 1021–1032 © Blackwell Science Limited1030
pBAC-SV were created by the insertion of a 1.3 kb NotI-HindIIIfragment from pCMV/Bsd (Invitrogen) or a 2.6 kb PvuII-EcoRIfragment from pSV2bsr (Kakenseiyaku), both containing a Blasti-cidin S resistance gene, into the NotI-HindIII sites of pBAC-TAN.The 25 kb α 21-I alphoid fragment (α25) was isolated fromthe cosmid clone, Q25F12, obtained from the LL21NC02 library(Lawrence Livermore Laboratory) by SfiI digestion and clonedinto the SfiI site of pBAC-TAN. The resulting alphoid-BACs,which contain either 50 kb or 100 kb of tandem alphoid insert,were digested with MluI and SacII, and the alphoid fragments wereinserted into the MluI-SacII sites of pBAC-CMV or pBAC-SV,respectively.
Transfection
BAC-DNAs were purified by CsCl banding using a gradient. Forthe generation of HAC, 0.5 µg of alphoid-BACs and 1.0 µg ofGCH1-BAC (186L09, Genomesystems) were co-transfected intoHT1080 cells (5 × 105) using lipofectamine (Gibco BRL) accord-ing to the manufacturer’s instructions. The cells were selectedwith 4 µg/mL Blasticidin S (BS, Kakenseiyaku) and colonies werepicked after 10 days.
FISH
Metaphase chromosome spreads were prepared on glass slidesafter methanol/acetate (3 : 1) fixation and FISH was carried outaccording to conventional procedures. For the detection of HAC,biotin-labelled α 21-I alphoid DNA (11–4) (Ikeno et al. 1994) anddigoxigenin-labelled pBeloBAC11 were used as probes. GCH1genes were detected with a biotin-labelled 13 kb fragment con-taining exon 1 and with a digoxigenin-labelled 8 kb fragmentcontaining GCH1 exons 4, 5 and 6. In dual FISH, biotin-labelledDNA was visualized with FITC conjugated avidin (Vector),and digoxigenin-labelled DNA was visualized with TRITCconjugated anti-digoxigenin (Boehringer Mannheim). Photo-graphs were taken using a CCD camera (Princeton instruments)mounted on a Zeiss microscope. Images were processed usingIPLab and Adobe Photoshop 6.0.
Indirect immunofluorescence
Swollen and 1% paraformaldehyde fixed cells were incubated withanti-CENP-A (Ando et al. 2002) or anti-CENP-E (Santa Cruz)antibodies. Antibody localization was visualized with FITC-conjugated anti-mouse IgG. For the subsequent FISH analysis, thecells were fixed again with 1% paraformaldehyde and then withmethanol/acetate (3 : 1).
Restriction-enzyme digestion and hybridization
Genomic DNA (5 µg) from cell lines were digested for 4 h andfractionated by conventional gel electrophoresis. The DNA in thegel was transferred to a nylon membrane and hybridized with 32Plabelled DNA probe prepared from GCH1 exon 6 (2.1 kb) and
the upstream region of GCH1 (1.4 kb, position 595–1959 inGCH1-BAC).
Mitotic stability
The mitotic stability of GCH1-HAC was measured by culturingthe cells for approximately 30 days under non-selective growthconditions. With each of the cell lines after 10, 20 and 30 days culturein the absence of BS selection, spreads of metaphase chromosomeswere prepared, and the presence of HACs was analysed by FISH.The percentage of metaphase cells with HAC, from a total of 50spreads, was measured. The rate of HAC loss (R) was calculated bythe following formula: Nn = N0 × (1 − R)n, where N0 is the numberof the metaphase spreads containing a HAC under selective con-ditions, and Nn is the number of metaphase spreads containing aHAC after n days of culture under non-selective conditions.
Cell fusion
Cell lines containing a HAC (5 × 105) and mouse A9 cells(5 × 105) were co-cultivated and fused in PEG/DMSO solution(Sigma). BS- and Ouabain-resistant cells were selected with2.5 µg/mL BS and 3 µm Ouabain.
Assay of GCH1 activity
Cells were grown in the absence or presence of human IFN-γat 250 U/mL in culture medium for 48 h. Trypsinized cells werewashed in phosphate-buffered saline (PBS), and then lysedin 0.1 m Tris-HCl (pH 8.0), 0.3 m KCl, 2.5 mm EDTA and 10%glycerol. GCH1 activity was measured as previously described(Hibiya et al. 2000).
Chromatin immunoprecipitation (ChIP)
The nuclei of HT/GCH5-18 cells (5 × 107) were isolated anddissolved in WB (20 mm HEPES (pH 8.0), 20 mm KCl, 0.5 mmEDTA, 0.5 mm dithiothreitol, 0.05 mm phenylmethylsulphonylfluoride). After digestion with MNase, solubilized chromatin wasimmunoprecipitated using anti-CENP-A antibody as previouslydescribed (Ando et al. 2002).
AcknowledgementsWe thank K. Yoda for the anti-CENP-A antibody and E. Soedafor the Q25F12 cosmid clone. This work was supported by aspecial grant for Core Research for Evolutional Science andTechnology (CREST) of Japan Science and Technology Corporation( JST), a Grant for Establishment of transgenic agro-biofarmsystems from the Ministry of Agriculture, Forestry and Fisheries ofJapan, a Grant-in-Aid for International Scientific Research ( JointResearch) from the Ministry of Education, Culture, Sports, Scienceand Technology, Special Coordination Funds for PromotingScience and Technology from the Science and Technology Agency,and the Human Frontier Science Program.
GTC_580.fm Page 1030 Monday, September 9, 2002 7:09 PM

Human artificial chromosomes carrying GCH1
© Blackwell Science Limited Genes to Cells (2002) 7, 1021–1032 1031
ReferencesAndo, S., Yang, H., Nozaki, N., Okazaki, T. & Yoda, K. (2002)
CENP-A, -B, and -C chromatin complex that contains theI-type alpha-satellite array constitutes the prekinetochore inHeLa cells. Mol. Cell. Biol. 22, 2229–2241.
Bannister, A.J., Zegerman, P., Partridge, J.F., et al. (2001) Selectiverecognition of methylated lysine 9 on histone H3 by the HP1chromo domain. Nature 410, 120–124.
Dong, J.Y., Fan, P.D. & Frizzell, R.A. (1996) Quantitative analysisof the packaging capacity of recombinant adeno-associatedvirus. Hum. Gene Ther. 7, 2101–2112.
Ebersole, T.A., Ross, A., Clark, E., et al. (2000) Mammalianartificial chromosome formation from circular alphoid inputDNA does not require telomere repeats. Hum. Mol. Genet. 9,1623–1631.
Festentein, R., Tolaini, M., Corbella, P., et al. (1996) Locus controlregion function and heterochromatin-induced position effectvariegation. Science 271, 1123–1125.
Fisher, K.J., Jooss, K., Alston, J., et al. (1997) Recombinant adeno-associated virus for muscle directed gene therapy. Nature Med.3, 306–312.
Fournier, R.E.K. & Ruddle, F.H. (1977) Microcell-mediatedtransfer of murine chromosomes into mouse, Chinese hamster,and human somatic cells. Proc. Natl. Acad. Sci. USA 74, 319–323.
Grimes, B.R., Schindelhauer, D., McGill, N.I., et al. (2001) Stablegene expression from a mammalian artificial chromosome.EMBO Rep. 2, 910–914.
Henning, K.A., Novotny, E.A., Compton, S.T., et al. (1999)Human artificial chromosomes generated by modification of ayeast artificial chromosome containing both human alphasatellite and single-copy DNA sequences. Proc. Natl. Acad. Sci.USA 96, 592–597.
Hibiya, M., Ichinose, H., Ozaki, N., et al. (2000) Normal valuesand age-dependent changes in GTP cyclohydrolase I activityin stimulated mononuclear blood cells measured by high-performance liquid chromatography. J. Chromatogr. B. Biomed.Sci. Appl. 740, 35–42.
Howman, E.V., Fowler, K.J., Newson, A.J., et al. (2000) Early dis-ruption of centromeric chromatin organization in centromereprotein A (Cenpa) null mice. Proc. Natl. Acad. Sci. USA 97,1148–1153.
Hubbard, T., Barker, D., Birney, E., et al. (2002) The Ensemblgenome database project. Nucl. Acids Res. 30, 38–41.
Ichinose, H., Ohye, T., Matsuda, Y., et al. (1995) Characterizationof mouse and human GTP cyclohydrolase I genes. Mutationsin patients with GTP cyclohydrolase I deficiency. J. Biol. Chem.270, 10062–10071.
Ichinose, H., Ohye, T., Takahashi, E., et al. (1994) Hereditaryprogressive dystonia with marked diurnal fluctuation caused bymutations in the GTP cyclohydrolase I gene. Nature Genet. 8,236–242.
Ichinose, H., Suzuki, T., Inagaki, H., Ohye, T. & Nagatsu, T.(1999) Molecular genetics of dopa-responsive dystonia. Biol.Chem. 380, 1355–1364.
Ikeno, M., Grimes, B., Okazaki, T., et al. (1998) Construction of
YAC-based mammalian artificial chromosomes. Nature Biotech-nol. 16, 431–439.
Ikeno, M., Masumoto, H. & Okazaki, T. (1994) Distribution ofCENP-B boxes reflected in CREST centromere antigenic siteson long-range alpha-satellite DNA arrays of human chromo-some 21. Hum. Mol. Genet. 3, 1245–1257.
Karpen, G.H. (1994) Position-effect variegation and the newbiology of heterochromatin. Curr. Opin. Genet. Dev. 4, 281–291.
Karpen, G.H. & Allshire, R.C. (1997) The case for epigeneticeffects on centromere identity and function. Trends Genet. 13,489–496.
Kaufman, S. (1993) New tetrahydrobiopterin-dependent systems.Annu. Rev. Nutr. 13, 261–286.
Lachner, M., O’Carroll, D., Rea, S., Mechtler, K. & Jenuwein, T.(2001) Methylation of histone H3 lysine 9 creates a binding sitefor HP1 proteins. Nature 410, 116–120.
Masumoto, H., Ikeno, M., Nakano, M., et al. (1998) Assay ofcentromere function using a human artificial chromosome.Chromosoma 107, 406–416.
Mejia, J.E., Willmott, A., Levy, E., Earnshaw, W.C. & Larin, Z.(2001) Functional complementation of a genetic deficiencywith human artificial chromosomes. Am. J. Hum. Genet. 69,315–326.
Milot, E., Strouboulis, J., Trimborn, T., et al. (1996) Heterochro-matin effects on the frequency and duration of LCR-mediatedgene transcription. Cell 87, 105–114.
Mineta, T., Rabkin, S.D., Yazaki, T., Hunter, W.D. & Martuza, R.L.(1995) Attenuated multi-mutated herpes simplex virus-1 forthe treatment of malignant gliomas. Nature Med. 1, 938–943.
Muramatsu, S., Fujimoto, K., Ikeguchi, K., et al. (2002) Behavioralrecovery in a primate model of Parkinson’s disease by tripletransduction of striatal cells with adeno-associated viral vectorsexpressing dopamine-synthesizing enzymes. Hum. Gene Ther.13, 345–354.
Nichol, C.A., Smith, G.K. & Duch, D.S. (1985) Biosynthesis andmetabolism of tetrahydrobiopterin and molybdopterin. Annu.Rev. Biochem. 54, 729–764.
Ota, A., Yoshida, S., Nomura, T., et al. (1996) Tetrahydrobio-pterin biosynthesis enhanced by lipopolysaccharide stimulationin murine neuroblastoma cell line N1E-115. J. Neurochem. 67,2540–2548.
Palmer, D.K., O’Day, K., Trong, H.L., Charbonneau, H. &Margolis, R.L. (1991) Purification of the centromere-specificprotein CENP-A and demonstration that it is a distinctivehistone. Proc. Natl. Acad. Sci. USA 88, 3734–3738.
Pfeifer, A. & Verma, I.M. (2001) Gene therapy: promises andproblems. Annu. Rev. Genomics Hum. Genet. 2, 177–211.
Platero, J.S., Hartnett, T. & Eissenberg, J.C. (1995) Functionalanalysis of the chromo domain of HP1. EMBO J. 14, 3977–3986.
Shen, M.H., Yang, J., Loupart, M.L., Smith, A. & Brown, W.(1997) Human mini-chromosomes in mouse embryonal stemcells. Hum. Mol. Genet. 6, 1375–1382.
Tanaka, K., Kaufman, S. & Milstein, S. (1989) Tetrahydrobio-pterin, the cofactor for aromatic amino acid hydroxylases, issynthesized by and regulates proliferation of erythroid cells.Proc. Natl. Acad. Sci. USA 86, 5864–5867.
GTC_580.fm Page 1031 Monday, September 9, 2002 7:09 PM

M Ikeno et al.
Genes to Cells (2002) 7, 1021–1032 © Blackwell Science Limited1032
Wade-Martins, R., White, R.E., Kimura, H., Cook, P.R. &James, M.R. (2000) Stable correction of a genetic deficiency inhuman cells by an episome carrying a 115 kb genomic trans-gene. Nature Biotechnol. 18, 1311–1314.
Werner, E.R., Werner-Felmayer, G., Fuchs, D., et al. (1990)Tetrahydrobiopterin biosynthetic activities in human macro-phages, fibroblasts, THP-1, and T 24 cells. GTP-cyclohydrolaseI is stimulated by interferon-γ, and 6-pyruvoyl tetrahydropterinsynthase and sepiapterin reductase are constitutively present.J. Biol. Chem. 265, 3189–3192.
Werner, E.R., Werner-Felmayer, G. & Wachter, H. (1993)Tetrahydrobiopterin and cytokines. Proc. Soc. Exp. Biol. Med.203, 1–12.
Yen, T.J., Compton, D.A., Wise, D., et al. (1991) CENP-E, anovel human centromere-associated protein required forprogression from metaphase to anaphase. EMBO J. 10,1245–1254.
Received: 6 June 2002 Accepted: 6 July 2002
GTC_580.fm Page 1032 Monday, September 9, 2002 7:09 PM




















