
Towards measurement of the time-resolved heat release of protein conformation dynamics

Plant Physiol. (1975) 55, 393-400
The Effect of Adenosine 5'-Triphosphate on the Shibata Shiftand on Associated Structural Changes in the Conformationof the Prolamellar Body in Isolated Maize Etioplasts'
Received for publication June 25, 1974 and in revised form September 23, 1974
PETER HORTON2 AND RACHEL M. LEECHDepartment of Biology, University of York, Heslington, York, England
ABSTRACT
Isolated maize (Zea mays var. kelvedon glory) etioplasts havebeen used to investigate the relationships between the spectralshifts and ultrastructural changes which occur during light-in-duced chloroplast development. After primary photoconversion,the Shibata shift was observed as a change from 680 to 670 nmin the chlorophyllide absorption maximum. When 1.5 nM ATPwas added to the incubation medium the maximum was 675 nmeven after 3.5 hours of illumination. Difference spectra forthis effect indicate ATP inhibition of the Shibata shift. Twobands with maxima at 682 and 669 nm can be used to fitspectra of both ATP-treated and control etioplasts, the esti-mated proportions of chlorophyllide 682 being 36% and 6%,respectively. Quantitative analysis of electron micrographs ofthe etioplasts showed that the frequency of untransformedprolameliar bodies was also higher in the presence of ATP(73% untransformed compared to 22% in the absence ofATP). A similar correlation was observed when transforma-tion was measured for two etioplast fractions which show theshift to different extents. These results imply that the Shibatashift and prolamellar body transformation are related events,both being inhibited by the presence of ATP. ATP may there-fore have an important role in regulating the early stages ofplastid development.
The early stages of the light-induced transformation ofgreening etioplasts into chloroplasts in leaves involve severalshifts in the position of the chlorophyll(ide) absorption maxi-mum (1, 8, 11, 18, 28, 30-32). The first slow shift was char-acterized by Shibata (28) and involves changes in maximumfrom 680 to 684 nm to 670 to 673 nm and occurs in etiolatedgreening leaves within 10 to 40 min following the formation ofChle.3 It has been shown that the shift can be induced in whole
1 This research was supported by Science Research Council GrantB/SR/8692 to R. M. L. and a studentship to P. H.
2 Present address: Department of Biological Sciences, PurdueUniversity, West Lafayette, Ind. 47907.
3Abbreviations: Chle: chlorophyllide; PChle: protochlorophyllide;PChl: protochlorophyll; PChl/e and Chl/e refer to mixtures ofphytylated and unphytylated pigments; PLB: prolamellar body. Foridentification purposes several spectral forms of protochlorophylland Chl are referred to as follows: PChl/e 630, absorption maxi-mum 628 to 630 nm; PChle 650, maximum at 650 nm; Chl/e 670,
leaves exposed to freezing and thawing (8, 12), suggesting thatthe changes in association between different pigment moleculesor between pigment molecules and other membrane com-ponents is the cause of the shift (18). This hypothesis is sup-ported by the demonstration that the shift also occurs inisolated Chle-holochrome suspensions (3, 14, 26), and otherrecent evidence indicates that a change in holochrome con-formation occurs during the shift (14, 22, 27). An alternativeview is that phytylation is the cause of the Shibata shift (29)but there is little direct experimental support for this view.The rate of the Shibata shift is variable and appears to de-
pend on several parameters. Electron microscopical studieshave indicated that the shift is itself correlated with specificmembrane changes occurring in the developing plastid; ac-cording to Von Wettstein's group, prolamellar body dispersalcan occur simultaneously with the Shibata shift in barley butthey add that in other species the relationship between thespectral and ultrastructural changes is not always as clearlyseen (13, 43).
Studies using whole plants in which attempts have beenmade to examine the control of the Shibata shift and its rela-tionship to membrane changes have been hindered by the com-plexity of the metabolism of the intact tissue and by the diffi-culty in inducing alteration of specific processes from outsidethe whole organism. Recently systems of isolated etioplastshave been described by Wellburn and Wellburn (40, 41, 42) andby Horton and Leech (15, 16) capable of being used to studylimited plastid development in vitro. Using isolated maizeetioplasts, we have reported a marked effect of ATP on theproportions of PChle forms having maxima at 650 and 630nm (16) and these etioplast suspensions also provide an idealsystem in which to study the changes associated with theShibata shift. This paper reports experiments showing thatATP also affects both the spectral changes associated withthe Shibata shift and the transformation of the prolamellarbody. The ATP effect was found to be a valuable probe infurther investigating the nature of the association betweenthe spectral and ultrastructural changes.
MATERIALS AND METHODS
Isolation of Etioplasts. Suspensions of etioplasts were pre-pared from seedlings of dark-grown maize (Zea mays var. kelve-don glory) as described previously (20). "Etioplasts" refer tothose prepared from the initial high speed centrifugation, andthe suspensions contain approximately 60% envelope-bound
maximum at 669 to 674 nm; Chle 680, maximum at 680 to 685nm. The ranges of maxima refer to previously published values.
393

HORTON AND LEECH
plastids. Washed etioplast suspensions were prepared by furthercentrifugation through 0.6M sucrose. These preparations con-tained 90% envelope-bound etioplasts. After resuspension in amedium containing 0.5M sucrose, 0.4% BSA (Cohn, fraction V,Koch Light) and 2.5% Ficoll, 5 mm cysteine,1 mm MgCl2, 1 mMEDTA, 5 mm KHCO3, 0.5 mm sodium acetate, 1mM L-glu-tamic acid, in 67 mm Na2HPO,-KH2PO, buffer (pH 7.5), theetioplasts were incubated either unstirred in a 1X 1X 4.5cm cuvette (dark experiments) orunder continuous rotation inthe light. The Shibata shift was recorded for etioplast suspen-sions incubated either in darkness or during rotation (16, 25)under continuous low intensity illumination (1.6X 103 ergcm-2s-') provided by a 25-w tungsten filament lamp. In both casesetioplasts were initially illuminated to photoconvert the PChle650 into Chle 680 as described previously(16). ATP was addedwhere indicated as the disodium salt (Sigma) to give a con-centration of 1.5 mm. Absorption spectra were recorded atroom temperature in a 3-ml volume containing between 1.2and 2.3 nmoles PChl/e- ml using a Shimadzu MP550L multi-purpose spectrophotometer. A Dupont 310 curve resolver wasused to analyze the Chl bands into component curves.
ElectronMicroscopy. Etioplast samples were prepared forelectron microscopy according to a procedure similar to thatdescribed by Wellburn and Wellburn (40).
Samples of etioplasts were fixed in an equal volume of 5%glutaraldehyde in 34 mm Na2HPO-KH2PO4 buffer (pH 7.5)containing 0.5M sucrose. Fixation was at 0 C for 2 hr in com-plete darkness and the suspension was then centrifuged to forma pellet at the base of the centrifuge tube. The supernatant wasremoved, and the pellet was diluted successively with bufferscontaining 0.5M, 0.35M, and 0.2M sucrose. The pellet waspostfixed with 2% (w/v) osmium in phosphate buffer contain-ing 0.1 M sucrose, for 2 hr at 0 C in darkness. After removalof the osmium, the pellet was dehydrated and embedded asdescribed by Mackender and Leech (21) except that the 1 mlof 30% acetone which began the dehydration series contained0.1M sucrose. The sucrose concentration was gradually dilutedto10 mM. First 1 ml, then 2 ml, and finally 4 ml of acetonewere added to increase the acetone concentration to 65, 83,and 88 %. respectively. Finally, the acetone was removed anddehydration was completed by adding 1 ml of acetone andleaving overnight. All manipulations until the end of dehydra-tion were carried out under a dim green safelight (Ilford 905X),and the samples, whenever possible, were kept in completedarkness.The pellet was embedded in Spurr's resin (31) after care-
fully dissecting it to locate the top, middle, and bottom of thepellet. This was necessary, because profiles from differentregions of the pellet were found to differ markedly in theircontents. Sections were cut on an LKB ultramicrotome using adiamond knife, picked up on uncoated copper grids, andstained in uranyl acetate followed by lead citrate (24). Analternative staining procedure, modified from one describedby Sutton (33), was also used. This procedure involved treat-ing the sections with 2% KMnO4 for 5 min. rinsing in H20,and bleaching with a mixture of 1.2% sodium sulphate (3drops), 1% oxalic acid (3 drops), and water (10 ml), beforestaining for 30 to 60 sec in lead citrate. KMnO4 appears tomake the tissue components more receptive to lead, givingmore contrast (34), and is especially useful when employingSpurr's resin. Sections were viewed with an AEI EM6B elec-tron microscope at 60 kv. For quantitative assessment of theamount of prolamellar body membranes occurring in etioplastsuspensions, two methods were used to analyze the electronmicrographs.
1. Prolamellar bodies were assigned quantitatively to three
crystalline (tight), completely transformed (loose), and con-
taining areas in both the loose and tight configurations (mixed).Examples of these three categories are shown in FigureThis method does not give a precise indication of the aMOnntof prolamellar body material in each configuration. In par-
ticular the mixed class varies from almost totally tight tototally loose.
2. An alternative method of quantification was to measure
the total area of PLB showing the tight and the loose configura-tion. To estimate the relative proportions of tight and looseprolamellar bodies, areas containing tight or loose regions were
cut out from photocopies of etioplast profiles and weighed.Weight was found to be consistently proportional to the area
of the photocopy paper. For mixed prolamellar bodies, the area
was divided into loose and tight areas before weighing. Thetotal weight, after correction for different magnification. was
calculated and the ratio of tight and loose proiamellar bodiesdetermined.
Both methods ofmeasurement require random selection ofetioplast profiles. Randomness was ensured by examiningphotographs whichwere taken at 4000 to 8000 times magnifi-cation on the microscope and which therefore always con-
tained several plastids. Moreover, we were not involved in theprocess of selecting grid areas for photography, and the iden-tity of samples was concealed until after the analysis of theprints was complete.
Analysis of Chlorophyllide and Chlorophyll. The phytylated(Chl) and unphytylated Chle were separated by a partition be-
tween petroleum ether and aqueous acetone using the methodof Treffry (35). Petroleum ether (1.8 ml) was added to5 ml ofan 80% acetone etioplast extract. The pigment concentrationsin the petroleum and the aqueous acetone phases were deter-mined spectrophotometrically as described by Anderson nndBoardman (2).
RESULTS
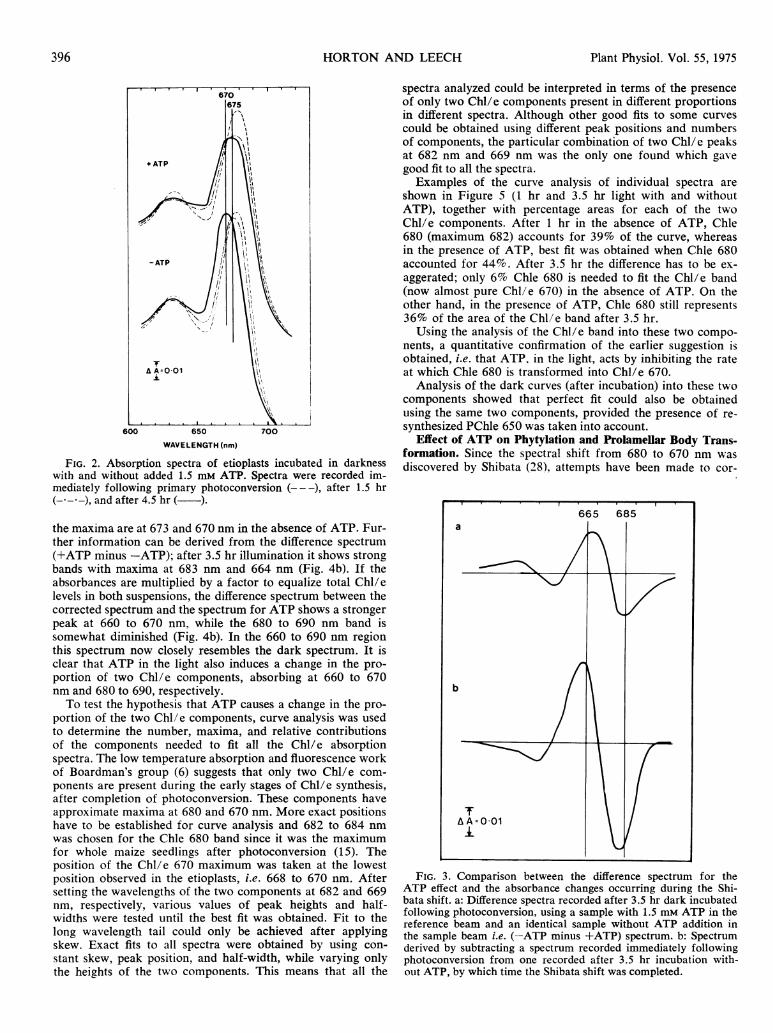
Effect of ATP on Shibata Shift. The Shibata shift was firstexamined in darkness so that problems due to additional light-induced synthesis of Chl were eliminated. Figure 2 shows theabsorption spectra of etioplast samples immediately followingillumination, after 1.5 hr in darkness and after 4.5 hr. A shiftfrom near 680 nm to 675 nm after 1.5 hr and to 670 nm after4.5 hr is observed. Incubation in the presence of ATP markedlyinhibited the Shibata shift. The effect of ATP on the maximumwas already recorded after 1.5 hr but the greatest differencein the spectra with and without ATP occurred after 4.5 hrwhen the maxima were at 674 and 670 nm, respectively. Thepresence of isobestic points as observed by Shibata (28) is
seen at 674 nm. indicaiting direct conversion of one componentinto another. Thus ATP did not appear to inhibit the initiallyrapid shift from over 680 to about 678 nm but only the later,slower stage from 676 to 670 nm.
Difference spectra were next recorded using the sample withadded ATP as the reference. Because of overlap of Chle bandsit is possible to obtain only approximate estimates of individualpeak positions from difference spectra but the spectra in Fig-ure 3a clearly show the type of change caused by the additionof ATP. Minima are seen at 650 nm and at 684 to 685 nm.
Maxima are visible at 630 and 666 to 667 nm. The positiveabsorbance in the 670 region and the negative absorbance illthe 680 region indicate an ATP-induced change in the pro-
portions of two Chl components, one with a maximum in theregion of 670 nm, the other in the region of 680 nm. Mlore-over, as can be seen by comparison with Figure 3b. the(-ATP minus +ATP) difference spectrum resembles closely
categories by observation of etioplast profiles: completely
394 Plant Physiol. Vol. 55, 1975
the difference wectrum for the completed Shibata shift, (3.5 to

SHIBATA SHIFT AND PROLAMELLAR BODY
a
A B
U. *.u -.- 9-
..e\i . ',.;
.i 4 * w FFe4s S < S _ ''ft.
* .. @;ff _<. , .^ . v. ..<A.sOL
4 2;\ V < ~~~.rl
4t-..->t. .s > ;.¢< t
A*g@>~9 6 s,x,>* i6.'ty w
C r'sP
¾.;
A.~~'W3 ~ 0*t
A~~~~~~ ~ ~ ~ ~~~~~~~~~~~~~/
FIG. 1. a: Classification of the degree of PLB transformation. A: Tight PLB (X 9,000); B: mixed PLB (X 14,000); and C: loose PLB (X
17,000). Profiles are of chloroplasts incubated for 3.5 hr under continuous illumination with 1.5 mMt ATP (A) and without ATP (B and C).
b: Electron micrographs of chloroplasts showing the effect of the presence of ATP on PLB transformation after 3.5 hr illumination ill vitro
(X 6,500). A: Incubated with ATP (note all the PLBS are tight); B: incubated without ATP. All three types of PLB are present. In one (arrow),
the PLB is dispersing into a mass of perforated lamellae.
-0 hr). The shape of the difference spectra in the 630 to 650 nmregion confirms our previous work in which the 630 and 650 nmpeaks were shown to be due to ATP-induced resynthesis ofPChle 650 from endogenous PChle 630 (15, 16). The resultsare consistent with the hypothesis that ATP is inhibiting thetransformation of Chle 680 into Chle 670, rather than that itis creating an entirely new Chi/e form at 673 to 675 nm.
A time course for the Shibata shift in etioplast suspensionsmaintained under continuous illumination with and withoutATP was next determined in order to investigate whether a
similar mechanism operated in the light and in the dark.Spectra recorded for washed etioplasts are shown in Figure4a. After 2 hr and 3.5 hr in the presence of ATP, broad bandswith maxima at 678 and 675 nm, respectively, are observed;
a
395Plant Physiol. Vol. 55, 1975
Plant Physiol. Vol. 55, 1975
650
WAVELENGTH (nm)
FIG. 2. Absorption spectra of etioplasts incubated in darknesswith and without added 1.5 mM ATP. Spectra were recorded im-mediately following primary photoconversion (---), after 1.5 hr(- - -), and after 4.5 hr ( ).
the maxima are at 673 and 670 nm in the absence of ATP. Fur-ther information can be derived from the difference spectrum(+ATP minus -ATP); after 3.5 hr illumination it shows strongbands with maxima at 683 nm and 664 nm (Fig. 4b). If theabsorbances are multiplied by a factor to equalize total Chl/elevels in both suspensions, the difference spectrum between thecorrected spectrum and the spectrum for ATP shows a strongerpeak at 660 to 670 nm. while the 680 to 690 nm band issomewhat diminished (Fig. 4b). In the 660 to 690 nm regionthis spectrum now closely resembles the dark spectrum. It isclear that ATP in the light also induces a change in the pro-portion of two Chl/e components, absorbing at 660 to 670nm and 680 to 690, respectively.To test the hypothesis that ATP causes a change in the pro-
portion of the two Chl/e components, curve analysis was usedto determine the number, maxima, and relative contributionsof the components needed to fit all the Chl/e absorptionspectra. The low temperature absorption and fluorescence workof Boardman's group (6) suggests that only two Chl/e com-ponents are present during the early stages of Chl/e synthesis,after completion of photoconversion. These components haveapproximate maxima at 680 and 670 nm. More exact positionshave to be established for curve analysis and 682 to 684 nmwas chosen for the Chle 680 band since it was the maximumfor whole maize seedlings after photoconversion (15). Theposition of the Chl/e 670 maximum was taken at the lowestposition observed in the etioplasts, i.e. 668 to 670 nm. Aftersetting the wavelengths of the two components at 682 and 669nm, respectively, various values of peak heights and half-widths were tested until the best fit was obtained. Fit to thelong wavelength tail could only be achieved after applyingskew. Exact fits to all spectra were obtained by using con-stant skew, peak position, and half-width, while varying onlythe heights of the two components. This means that all the
spectra analyzed could be interpreted in terms of the presenceof only two Chl/e components present in different proportionsin different spectra. Although other good fits to some curvescould be obtained using different peak positions and numbersof components, the particular combination of two Chl/e peaksat 682 nm and 669 nm was the only one found which gavegood fit to all the spectra.
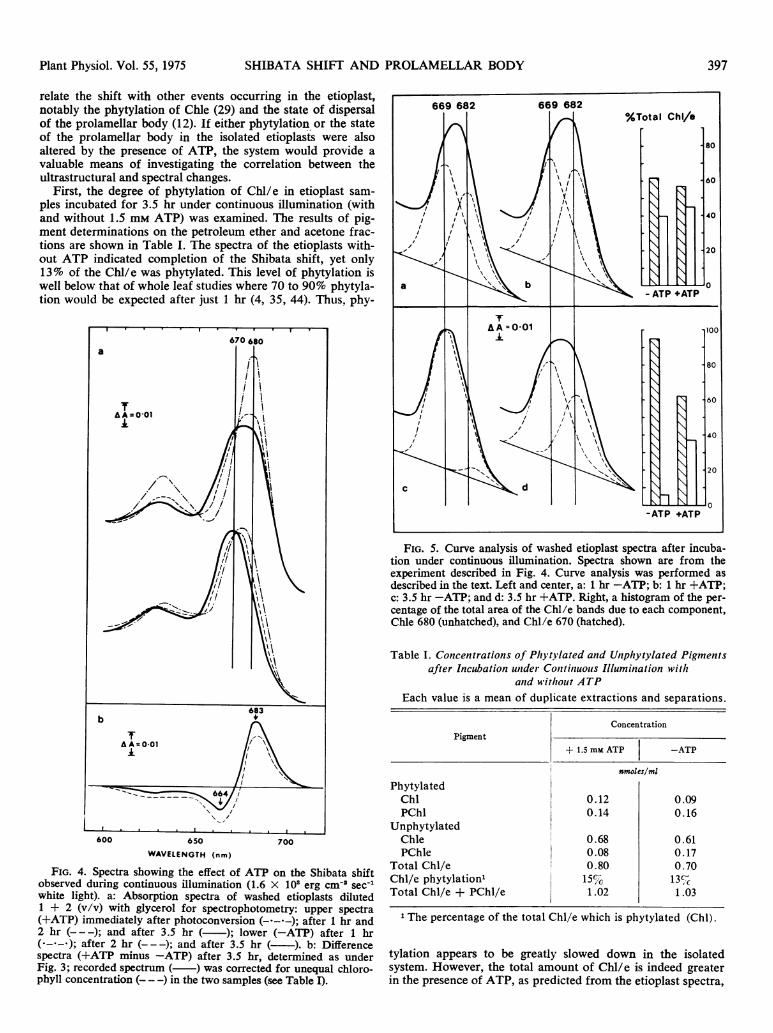
Examples of the curve analysis of individual spectra areshown in Figure 5 (1 hr and 3.5 hr light with and withoutATP), together with percentage areas for each of the twoChl/e components. After 1 hr in the absence of ATP, Chle680 (maximum 682) accounts for 39% of the curve, whereasin the presence of ATP, best fit was obtained when Chle 680accounted for 44%. After 3.5 hr the difference has to be ex-aggerated; only 6% Chle 680 is needed to fit the Chl/e band(now almost pure Chl/e 670) in the absence of ATP. On theother hand, in the presence of ATP, Chle 680 still represents36% of the area of the Chl/e band after 3.5 hr.
Using the analysis of the Chl/e band into these two compo-nents, a quantitative confirmation of the earlier suggestion isobtained, i.e. that ATP. in the light, acts by inhibiting the rateat which Chle 680 is transformed into Chl/e 670.
Analysis of the dark curves (after incubation) into these twocomponents showed that perfect fit could also be obtainedusing the same two components, provided the presence of re-synthesized PChle 650 was taken into account.
Effect of ATP on Phytylation and Prolameliar Body Trans-formation. Since the spectral shift from 680 to 670 nm wasdiscovered by Shibata (28), attempts have been made to cor-
FIG. 3. Comparison between the difference spectrum for theATP effect and the absorbance changes occurring during the Shi-bata shift. a: Difference spectra recorded after 3.5 hr dark incubatedfollowing photoconversion, using a sample with 1.5 mM ATP in thereference beam and an identical sample without ATP addition inthe sample beam i.e. (-ATP minus +ATP) spectrum. b: Spectrumderived by subtracting a spectrum recorded immediately followingphotoconversion from one recorded after 3.5 hr incubation with-out ATP, by which time the Shibata shift was completed.
396 HORTON AND LEECH
SHIBATA SHIFT AND PROLAMELLAR BODY
relate the shift with other events occurring in the etioplast,notably the phytylation of Chle (29) and the state of dispersalof the prolamellar body (12). If either phytylation or the stateof the prolamellar body in the isolated etioplasts were alsoaltered by the presence of ATP, the system would provide avaluable means of investigating the correlation between theultrastructural and spectral changes.
First, the degree of phytylation of Chl/e in etioplast sam-ples incubated for 3.5 hr under continuous illumination (withand without 1.5 m-m ATP) was examined. The results of pig-ment determinations on the petroleum ether and acetone frac-tions are shown in Table I. The spectra of the etioplasts with-out ATP indicated completion of the Shibata shift, yet only13% of the Chl/e was phytylated. This level of phytylation iswell below that of whole leaf studies where 70 to 90% phytyla-tion would be expected after just 1 hr (4, 35, 44). Thus, phy-
600 650 700
WAVELENGTH (nm)
FIG. 4. Spectra showing the effect of ATP on the Shibata shiftobserved during continuous illumination (1.6 x 101 erg cm-' sec-'white light). a: Absorption spectra of washed etioplasts diluted1 + 2 (v/v) with glycerol for spectrophotometry: upper spectra(+ATP) immediately after photoconversion (- - -); after 1 hr and2 hr (---); and after 3.5 hr ( ); lower (-ATP) after 1 hr(-----); after 2 hr (---); and after 3.5 hr ( ). b: Differencespectra (+ATP minus -ATP) after 3.5 hr, determined as underFig. 3; recorded spectrum ( ) was corrected for unequal chloro-phyll concentration (- -) in the two samples (see Table I).
FIG. 5. Curve analysis of washed etioplast spectra after incuba-tion under continuous illumination. Spectra shown are from theexperiment described in Fig. 4. Curve analysis was performed asdescribed in the text. Left and center, a: 1 hr -ATP; b: 1 hr +ATP;C: 3.5 hr -ATP; and d: 3.5 hr +ATP. Right, a histogram of the per-centage of the total area of the Chl/e bands due to each component,Chle 680 (unhatched), and Chl/e 670 (hatched).
Table I. Concentrations of Phytylated and Unphytylated Pigmentsafter Incubation untder Conttinzuous Illumination with
anld without ATPEach value is a mean of duplicate extractions and separations.
ConcentrationPigment
+ 1.5 mm ATP -ATP
nmoles/ml
PhytylatedChl 0.12 0.09PChl 0.14 0.16
UnphytylatedChle 0.68 0.61PChle 0.08 0.17
Total Chl/e 0.80 0.70Chl/e phytylation' 15%SS 13%cTotal Chl/e + PChl/e 1.02 1.03
1 The percentage of the total Chl/e which is phytylated (Chl).
tylation appears to be greatly slowed down in the isolatedsystem. However, the total amount of Chl/e is indeed greaterin the presence of ATP, as predicted from the etioplast spectra,
Plant Physiol. Vol. 55, 1975 397

Plant Physiol. Vol. 55, 1975
Table II. Effect of ATP oni PLB Transfiormzation:Qlualntitative Assessmnetit
Washed etioplasts were incubated under continuous illumina-tion for 3.5 hr with and without ATP. A: Distribution of intactetioplast profiles according to the state of the PLB as exemplifiedin Fig. 1; B: estimates of total PLB areas in the tight and looseconfiguration from approximately 80 etioplast profiles. Areas areexpressed as weight (g) of cut out paper.
iAmount of PLB TransformationConfiguration of PLB
+1.5 mm ATP -ATP
A. Tight 44 12Mixed 14 45Loose 25 27
B. Tight 4.4 1.1Loose 1.6 3 .9
Tight 73/ '2'.
Table Ill. Quianztified Estimates of the Amoanlt oJ PLBTransJormationz ini Convenitionial and Washed
Etioplast SaispenisionsEtioplasts and washed etioplasts prepared in a single experi-
ment were incubated in the presence of 1.5 mm ATP under con-tinuous illumination for 5 hr (white light 1.6 X 103 erg cm- sec-').PLB transformation was quantified as for Table II, with A and Brepresenting etioplast distribution and total areas respectively.Approximately 100 etioplasts were analyzed.
Amount of PLB TransformationConfiguration
Etioplasts W'ashed etioplasts
A. Tight 21 43Mixed 10 22Loose 37 21
B. Tight 1.1 5.1Loose 3.1 2.7
Tight 26% 66%'
but whether ATP is added to the suspension or not, the pro-portion of phytylated Chle remains the same.
Using the known ATP effect on spectral changes as a
probe, prolamellar body transformation and its relationshipto spectral changes was next investigated. Figure lb showssome typical etioplast profiles from washed suspensions after3.5 hr illumination in the presence and absence of ATP. Thehigher proportion of tight PLBs induced by ATP is readilyapparent; in the absence of ATP loose PLBs predominate. Asderived from the results in Table II, part A, 53% of the pro-files showed tight PLBs in the presence of ATP, but withoutATP only 14% were tight. Area measurements confirm theeffect of ATP. The area of tight PLB configuration is 73% of
the total PLB area in the presence of ATP; when ATP is ab-sent only 22% have the tight configuration.Washed etioplasts were found to differ from those conven-
tionally prepared in the extent of the Shibata shift, havingChle maxima of 675 to 678 nm (instead of 673-674 nm) after
5 hr illumination in the presence of ATP (15). The two meth-ods of PLB transformation measurement were therefore also
applied to normal and washed etioplasts to see if structuralchanges were correlated quantitatively with the spectralchanges. The washed etioplasts contain 50% tight PLBs,whereas only 32% of the profiles of conventionally preparedetioplasts had completely tight PLBs. Similarly, area measure-ments indicate that 26% of the PLB membranes were tight inconventional etioplasts, compared to 66% for washed etio-plasts (Table III). In this experiment the maximum after wash-ing was at 678 nm, and 673 nm before washing. Therefore,etioplasts which are incubated after washing show a greaterproportion of crystalline PLBs and also a greater inhibition ofthe Shibata shift than etioplasts conventionally prepared, andthis correlation between ultrastructural and spectral charac-teristics parallels the similarly correlated response of etioplaststo ATP treatment.
DISCUSSION
The effects of ATP on plastid development have beenidentified using isolated maize etioplasts. Results from thetime course for changes in the absorption maxima, differencespectroscopy, and curve analysis into two Chl/e componentsall strongly suggest that one effect of ATP is to inhibit therate of conversion of Chle 680 into Chl/e both in the lightand in darkness. Quantitative analysis of etioplast profiles ob-tained by electron microscopy show that ATP also interruptsthe process of PLB transformation. The presence of the tightPLBs in the illuminated etioplast suspensions could not be cor-related with the presence of either PChle 630 or PChle 650.A reasonable correlation was found between the extent of PLBtransformation and the Shibata shift. In the presence of ATP,the estimated Chle 680 content was 36%, and 73% of thePLB area was tight. In the absence of ATP the Chle 680 con-tent was only 6% and the amount of tight PLB material hadfallen to 22% (Table IV). The correlation between inhibitionof the transformation of the prolamellar body and inhibitionof the Shibata shift was again clearly seen when the behaviorof washed and conventionally prepared etioplasts was com-pared.The question arises whether the effects of ATP on the Shi-
bata shift and on PLB transformation are independent orwhether they both result from a single event induced by ATP.The consistently correlated effects of ATP suggest that anintimate association exists between the two processes. Previ-ously it had been proposed that the shift is associated with themuch later membrane change of prolamellar body dispersal,
Table lV. Quantitative Comparisoni betwreeni Degree of ShibataShift an,d Tranisfomniationi of PLB
Data for PLB transformation are taken from Tables II and III.Curve analysis of spectra taken from the same samples as usedfor electron microscopy was used to estimate the proportion ofChle 680. A: Comparison in the presence and absence of ATPafter 3.5 hr illumination; B: compares chloroplasts and washedchloroplasts after 5 hr illumination.
Treatment Chle 680 Tight ProlamellarBodies
% total arei oftotalCIzl e prolamiellar body
A. Plus 1.5 m\t ATP 36 73Minus ATP 6 22
B. Washed etioplasts 55 66Crude etioplasts 30 26
398 HORTON AND LEECH

SHIBATA SHIFT AND
while prolamellar body transformation parallels the earlierchange of PChle photoconversion (12, 13, 43). However, inthe present experiments, untransformed prolamellar bodieswere repeatedly observed after complete photoconversion ofPChle, and a similar situation has been reported in wholeleaves (36, 39). In addition, it is known that the Shibata shiftcan take place without concomitant PLB dispersal (13). Toreconcile these observations, we propose, as a working hy-pothesis, that an indirect association between PLB transforma-tion and the shift exists as a two stage process. After photo-conversion of the PChl 650 into Chle 680, which itself causesno ultrastructural change, the Chle 680 changes into anotherpigment species, spectrally identical, but now having the po-tential to undergo the Shibata shift (stage I). The pigmentchange is manifested as an alteration in the prolamellar bodytubule interactions, resulting in transformation to the looseconfiguration. Subsequently, dispersal of the PLB membranesand the formation of Chle 670 occurs (stage II). The stage II
events must be only loosely coupled, since Chle 670 can beformed without dispersal occurring (13).On the basis of current knowledge of the nature of the
components referred to as Chle 680 and Chle 670, it seemsmost likely that the effect of ATP is on the conformation ofthe holochrome protein. Chle 680, a major constituent of theprolamellar body (5, 17), is a pigment species bound to aprotein holochrome (7, 14, 22, 23) and the Shibata shift to670 nm is thought to result from changes in interactions be-tween the porphyrin rings (18), perhaps as a result of a dis-sociation of pigment-protein dimers (22, 27). However, sincethe shift can also occur in an isolated low molecular weightpigment-protein complex containing no interacting pigmentmolecules, Henningsen et al. (14) have recently proposed thatthe shift results merely from a simple conformational relaxa-tion of the individual holochrome subunits. On either inter-pretation, since ATP inhibits the Shibata shift, it is presum-ably affecting the conformation of the holochrome andpreventing the dissociation responsible for inducing alterationin the absorption properties of bound Chle. This effect is thusanalogous to the effect of ATP in controlling the ratio ofPChle 630 to PChle 650 (15, 16) where ATP apparently in-creases the degree of association between PChle and holo-chrome. The apparent paradox that ATP stimulates Chle syn-thesis yet inhibits the subsequent shift is thus reconciled. Thenature of the response of the two processes to ATP adds furthersupport to the suggestion that since energy transfer can occurbetween the two pigments after partial photoconversion (37,38), PChle 650 and Chle 680 occupy the same site on theholochrome. PChle 650 and Chle 680 also show similar re-sponses to extreme heat or freeze thaw treatment of leaves (8,10, 12) and appear at the same time during the growth ofetiolated leaves (19). The extent to which Chle is removedfrom the vicinity of the holochrome appears to be correlatedwith the ultimate dispersal of the prolamellar body (6, 14), soonly under circumstances where PChle synthesis and PLB dis-persal are virtually absent, as in isolated etioplasts, will thetwo effects of ATP come into opposition.The present experiments illustrate the value of isolated etio-
plasts for the study of discrete developmental changes. Furtherinformation about the possible role of ATP in plastid develop-ment requires examination of intact leaves. The finding of DeGreef et al. (9) of high ATP levels when many tight prolamellarbodies are still present during the early stages of slow greeningof leaves under far red light provides an interesting startingpoint for further experimentation.
A4cknouledgment-We thank 'Mrs. W. Crosby for assistance in preparing theelectron micrographs.
PROLAMELLAR BODY 399
LITERATURE CITED
1. AKOYC-NOGLOU, G. A. AND H. W. SIEGELMAN-. 1968. Protochlorophyllide re-
synthesis in dark-grown bean leaves. Plant Physiol. 43: 66-68.2. A.-NDERSON-, J. M. AND N. K. BOARDMANZ. 1964. Studies on the greening of
dark-grown bean plants. II. Development of photochenmical activity. Aust.J. Biol. Sci. 17: 93-101.
3. BOARD.MA-N, N. K. 1966. Protochlorophyll. In: L. P. V-ernon andl G. R. Seely,eds., The Chlorophylls. Academic Press, New York. pp. 437-479.
4. BOARDOMAN, N. K. 1967. Chloroplast structure and development. In: A. SanPietro, F. A. Greer, and T. J. Army, eds., Harvesting tlse Sun. AcademicPress, New York. pp. 211-230.
O. BOARD'MAN, N. K. AND J. M. ANDERSON. 1964. Studies oln the greening ofdark-grown bean plants. I. Formation of chloroplasts from proplastids.Aust. J. Biol. Sci. 17: 86-92.
6. BOARD'MAN, N. K., J. M. AN-DERSON, A. KAHN, S. W. THORNE, AND T. E.TREFFRY. 1970. Formation of photosynthetic membranes during clhlor-o-plast development. In: N. K. Boardman, A. W. Linnane, and R. M.
Smillie, ecls., Autonomy and Biogenesis of Mitochondria and Chloroplasts.-North Holland, Amsterdam. pp. 70-84.
7. BOGORAD, L., L. LABER, AND LM. L. GASSMIAN-. 1968. Aspects of chloroplastdevelopmient: transitory pigment-protein complexes and protoclhlorophyllideregeneration. In: K. Shibata, A. Takamiya, A. T. Jagendorf, and R. C.Fuller. eds., Comparative Biochemistry and Biophysics of Photosynthesis.University Park Press, State College, Pa., pp. 299-312.
8. BUTLER, W. L. AND W. R. BRIGGS. 1966. The relation betwN een structure andpigments during the first stages of proplastid greening. Biochiim. Biophys.Acta 112: 45-53.
9. DE GREEF, J., W. L. BUTLER, AND R. F. ROTH. 1971. Greening of etiolated beanleaves in far red light. Plant Physiol. 47: 457-464.
10. DUJARDIN, E. AN-D C. SIRONVAL. 1970. The reduction of protochlorophyllidleinto chlorophyllide. III. The phototransformability of the forms of theprotochlorophyllide-lipoprotein complex found in darkness. Photosyntlhetica,4: 129-138.
11. GASSIMAN_\, 'M. L. 1973. The conversion of photoinactiVe protochlorophyllide633to phototransformable protochlorophyllideeso in etiolated bean leavestreated with 6-aminolevulinic acid. Plant Physiol. 52: 590-594.
12. HEN-NIN-GSEN, K. W. 1970. 'Macromolecular physiology of plastids. VI. Clsangesin membrane structure associated with shifts in the absorption maxima ofthe chlorophyllous pigments. J. Cell Sci. 7: 587-621.
13. HENNINGSEN, K. W. AND J. E. BOYN-TON. 1969. 'Macromolecular physiologyof plastids. VII. The effect of a brief illumination on plastids of dark-grown barley leaves. J. Cell Sci. 5: 757-793.
14. HENNINGSEN, K. W., S. W. THORN-E, AND N. K. BOARDMAN. 1974. Propertiesof protochlorophyllide and chlorophyllide holochromes from etiolated andgreening leaves. Plant Physiol. 53: 419-425.
15. HORTON, P. 1973. Chlorophyll biosynthesis and membrane changes in isolatedetioplasts. D. Phil. thesis. University of York, England.
16. HORTON, P. AN-D R. 'M. LEECH. 1972. The effect of ATP on plsotoconversion ofprotochloroplsyllide into chlorophyllide in isolated etioplasts. FEBS Lett.26: 277-280.
17. KAHN-, A. 1968. Developmental physiology of bean leaf plastids. II. Negativecontrast electron microscopy of tubular membranes in prolanmellar bodies.Plant Plivsiol. 43: 1769-1780.
18. KIRK, J. T. 0. 1970. Biochemical aspects of chloroplast developinent. Annu.Rev. Plant Physiol. 21: 11-42.
19. KLEIN, S. AND SCHIFF, J. A. 1972. The correlated appearance of prolamellarbodies, protoclilorophyllide species, and the Shibata shift during develop-ment of bean etioplasts in the dark. Plant Physiol. 49: 619-626.
20. LEESE, B. M., R. 'M. LEECH, AND W. W. THO'MSON. 1972. Isolation of plastidsfrom different regions of developing maize leaves. In: G. Forti, Al. Avronand A. 'Melandri, eds., Proceedings 2nd International Congress on Photo-synthetic Research, Vol. 2. Dr. W. Junk N.V., The Hague. pp. 1485-1494.
21. 'MACEENDER, R. 0. AND R. M. LEECH. 1970. Isolation of clsloroplast envelopemembranes. Nature 228: 1347-1349.
22. MATHIS, P. AN-D K. SAUER. 1972. Circular dichroism studies on the structureand the photochemistry of protochlorophyllide and chlorophyllide bolo-chrome. Biochim. Biophys. Acta 267: 498510.
23. MAIATHIS, P. AND K. SAUER. 1973. Chlorophyll formation in greening beanleaves during the early stages. Plant Physiol. 51: 115-119.
24. REY-NOLDS, E. S. 1963. The use of lead citrate at high pH as an electronopaque stain in electron microscopy. J. Cell Biol. 17: 208-212.
25. RIDLEY, S. M. AND R. M. LEECH. 1969. Chloroplast survival in vitro. Ill:H. IMetzner ed., Progress in Photosynthesis Research, Vol. 1. H. Laupp, Jr.
Tubingen. pp. 229-244.
26. SCHOPFER, P. AND H. W. SIEGELMIAN-. 1968. Purification of protochloroplix-Ilideholochrome. Plant Physiol. 43: 990-996.
27. SCHULTZ, A. AND K. SAUER. 1972. Circular dichroism and fluorescence changesaccompanying the protochlorophyllide to chlorophyllide transformation ingreenin1g leaves and holochrome preparations. Biochim. Biophsys. Acta 267:
320-340.28. SHIBATA, K. 1957. Spectroscopic studies on chlorophyll formationi in intact
leaves. J. Biochem. 44: 147-173.
Plant Physiol. Vol. 55, 1975

400 HORTON)
29. SIRONVAL, C., M. R. MICHEL-WOLWERTZ, AND A. MADSEN. 1965. On thenature and possible functions of the 673- and 684-m,A forms of in vivochlorophyll. Biochim. Biophys. Acta 94: 344-354.
30. SMITH, J. H. C. 1960. Protochlorophyll transformations. In: M. B. Allen, ed.,Comparative Biochemistry of Photoreactive Systems. Academic Press,New York. pp. 257-277.
31. SPERR, A. R. 1969. A low viscosity epoxy resin embedding medium forelectron microscopy. J. Ultrastruct. Res. 26: 31-43.
32. SUNDQVIST, C. 1973. The relationship between chlorophyllide accumulation,the amount of protochlorophyllide636 and protochlorophyllidee&o in dark-grown wheat leaves treated with 6-amninolevulinic acid. Physiol. Plant. 28:464W470.
33. SUTTON, J. S. 1968. Potassium permanganate staining of ultrathin sections forelectron microscopy. J. Ultrastruct. Res. 21: 424-429.
34. TILNEY, L. G. AND B. BYERS. 1969. Studies on the microtubules in Heliozoa.V. Organisation of microtubules in the axonemal pattern in Echinosphaeriumnucleofilum. J. Cell Biol. 43: 148-165.
35. TREFFRY, T. E. 1970. Phytylation of chlorophyllide and prolamellar bodytransformation in etiolated peas. Planta 91: 279-284.
36. TREFFRY, T. E. 1973. Chloroplast development in etiolated peas: reformationof prolamellar bodies in red light without accumulation of protochloro-phyllide. J. Exp. Bot. 24: 185-195.
N4D LEECH Plant Physiol. Vol. 55, 1975
37. THORNE, S. W. 1971. The greening of etiolated bean leaves. T. The initialphotoconversion process. Biochim. Biophys. Acta 226: 113-127.
38. THORNE, S. W. 1971. The greening of etiolated bean leaves. II. Secondaryand further photoconversion processes. Biochim. Biophys. Acta 226: 128-134.
39. WEIER, T. E., R. D. SJOLUND, AND D. L. BROWN. 1970. Changes induced bylow light intensities on the prolamellar body of 8-day, dark-grown seedlings.Amer. J. Bot. 57: 276-284.
40. WELLBURTN, A. R. AN-D F. A. M. WELL3URN. 1971. A new method for theisolation of etioplasts with intact envelopes. J. Exp. Bot. 22: 972-979.
41. WELLBURN, A. R. AND F. A. M. WELLBURN. 1973. Developmental changes inetioplasts in isolated suspensions and in situ. Ann. Bot. 37: 11-19.
42. 'WELLBURN, F. A. M. AND A. R. WELLBURN. 1971. Development changesoccurring in isolated intact etioplasts. J. Cell Sci. 9: 271-287.
43. VON WETTSTEIN, D., K. W. HENNINGSEN, J. E. BOYNTON, G. C. KAN-
NANTGARA, AND 0. F. NIELSEN. 1970. The genetic control of chloroplastdevelopment in barley. In: N. K. Boardman, A. W. Linnane, and R. M.Smillie, eds., Autonomy and Biogenesis of Mitochondria and Chloroplasts.North Holland, Amsterdam. pp. 205-223.
44. WVOLFF, J. B. AND L. PRICE. 1957. Terminal steps of chlorophyll or bio-synthesis in higher plants. Arch. Biochem. Biophys. 72: 293-301.
AT~
Copyright © 2022 FDOKUMEN














![Problems with a conformation assignment of aryl-substituted resorc[4]arenes](https://static.fdokumen.com/doc/165x107/6324d12685efe380f30661c8/problems-with-a-conformation-assignment-of-aryl-substituted-resorc4arenes.jpg)





