
Neutralizing DNA Aptamers against Swine Influenza H3N2 Viruses
Upload
independentCategory
view
1download
0
Journal of General Virology (1992), 73, 2913-2921. Printed in Great Britain 2913
Assembly of conformation-dependent neutralizing domains on glycoprotein B of human cytomegalovirus
Ishtiaq Qadri, David Navarro,t Pedro Paz and Lenore Pereira*
Division o f Oral Biology, School o f Dentistry, University o f California San Francisco, San Francisco, California 94143-0512, U.S.A.
We analysed the antigenic properties of human cytomegalovirus (CMV) glycoprotein B (gB) by con- structing a set of deletion derivatives lacking different portions of the carboxy terminus and reacting them with a panel of monoclonal antibodies with neutral- izing activity. We found that two novel antigenic domains that bind neutralizing antibodies were assembled on truncated forms ofgB, one in the amino- terminal half and one that spans the midregion of the molecule. Assembly of the conformation-dependent epitopes occurred independently of residues in the carboxy-terminal half of the molecule and did not depend on proteolytic cleavage of the molecule between amino acids 460 and 461. Ten antibodies recognized a derivative with 447 amino-terminal residues; their
failure to recognize a derivative 411 residues long suggested that the amino acids required for assembly of these epitopes either were incorrectly folded, or had been totally or partially deleted in this derivative. Epitopes for three antibodies with complement- independent neutralizing activity were assembled when amino acids from the midregion of gB between residues 447 and 476 were present. Two other antigenic domains were formed by the addition of residues 476 to 618 and 619 to 645 from the carboxy-terminal half of gB. Our results underscore the importance of confor- mation in the antigenic structure and functional properties of both the amino- and carboxy-terminal portions of gB.
Introduction
Human cytomegalovirus (CMV) is a pathogen that causes significant morbidity and mortality in immuno- compromised patients, including organ transplant re- cipients (Meyers et al., 1986; Singh et al., 1988) and congenitally infected newborns (Pass et al., 1980; Stagno et al., 1983). Among those at risk from life-threatening CMV pneumonia and CMV chorioretinitis are patients with AIDS (Drew, 1988; Jacobson & MiUs, 1988; Pepose, 1989). CMV infections elicit neutralizing anti- bodies that react with the viral glycoproteins in infected cells, particularly with a major virion envelope glyco- protein (Britt & Vugler, 1990; Pereira et al., 1982a, 1983, 1992; Rasmussen et al., 1991) that has been designated glycoprotein B (gB) on the basis of sequence similarity with herpes simplex virus 1 (HSV-1) glycoprotein B (Chee et al., 1990; Cranage et al., 1986; Pellett et al., 1985).
CMV gB is a major component of the virion envelope (Farrar & Greenaway, 1986). It is highly immunogenic,
t Present address: University of Valencia, Faculty of Medicine, Valencia, Spain.
inducing neutralizing antibodies in infected patients and in animals immunized with the glycoprotein (Britt et al., 1988; Gonczol et al., 1986; Rasmussen et al., 1985). CMV gB is made as a single polypeptide chain of approximately 120K which undergoes glycosylation and cleavage between residues 460 and 461 during transport through the exocytic pathway (Pereira et aL, 1984; Radsak et al., 1990; Rasmussen et al., 1988, Spaete et al., 1988). Cleavage of gB occurs in the Golgi compartment, and the specificity of the calcium-dependent protease for the target site is dependent on basic amino acids at or near this site (Spaete et al., 1990). The gB heterodimer is composed of the amino- and carboxy-terminal frag- ments, which remain disulphide-bonded (Britt, 1984) and are anchored in the membranes of infected cells by the hydrophobic transmembrane sequence (Spaete et al., 1988). The amino-terminal portion, having 15 N-glyco- sylation sites, is heavily modified and is approximately 110K; the carboxy-terminal portion, having five glyco- sylation sites, is approximately 58.5K (Meyer et al., 1990; Pereira et al., 1984; Rasmussen et al., 1988). Nucleotide sequence analyses ofgB genes from CMV strains AD169 and Towne show sequence variation at the extreme amino terminus between residues 28 and 67 (Cranage et
0001-1092 © 1992 SGM

2914 I. Qadri and others
al., 1986; Spaete et al., 1988). Analysis of several cl inical s trains also showed sequence var iabi l i ty in other regions of the molecule, inc luding residues 440 to 460 (Chou, 1992; Chou & Denni son , 1991).
Our studies on C M V gB have focused on m a p p i n g its ant igenic and funct ional regions using a panel of monoc lona l an t ibodies (MAbs). Analys is of the epitopes in the extreme a m i n o te rminus of gB revealed a region of cont inuous amino acids tha t b inds several ant ibodies , inc luding one with complemen t -dependen t neutra l iz ing act ivi ty (Basgoz et al., 1992; Meyer et al., 1992; Pereira et
al., 1991). In the present study, we made a set of delet ion constructs in the C M V (AD169) gB gene encoding derivat ives that lacked carboxy- terminal sequences of various lengths. Epi topes assembled on these derivat ives were identified by t rans ient ly expressing the products in COS-1 cells and detect ing them by immunofluorescence, and in some instances by immunoprec ip i t a t ion , using neutra l iz ing ant ibodies to gB. We found four confor- ma t ion -dependen t regions on gB that b ind neut ra l iz ing ant ibodies and identified the amino acids required for assembly of d iscont inuous epitopes in these regions. Our studies show that the amino- t e rmina l half of gB be tween residues 411 and 447 is required to assemble an ant igenic region recognized by ant ibodies with complement - independen t and -dependent neutra l iz ing activity. Assembly of the epitopes in this region is i ndependen t of the carboxy- terminal half of the molecule. We also identified the amino acids required to assemble epitopes in the midregion of gB that span the proteolytic cleavage site. Lastly, we more precisely defined the amino acids
required to assemble epitopes in the carboxy- terminal half of gB that had been identified in an earlier study (Banks et al., 1989).
Methods
Construction of deletion derivatives in CMV (AD169) gB. A panel of deletion derivatives of CMV (AD169) gB was constructed. BamHI fragment T containing the gB gene ofCMV (AD169) was excised from cosmid 7091 (Fleckenstein et al., 1982) and cloned into the BamHI site of pUC18. The EagI fragment (3125 bp) encoding gB was cloned into the SmaI site of the eukaryotic expression vector pMT2 (Wong et al., 1985) and designated pMTgB. A derivative, pMT3, was constructed by inserting a synthetic linker, SpeI, with stop codons in all three frames at the blunt-ended EcoRI site. Subfragments expressing gB of various lengths were then cloned into this plasmid at the appropriate restriction sites. The deletion constructs were obtained by using convenient restriction endonuclease sites within the gB gene. Plasmids p258, p411, p447 and p476 were obtained by excising the BgllI and blunt-ended BssHII, NdeI, ClaI and ApaLI fragments, respectively, from pMTgB and then cloning them into the BgllI and Sinai restriction sites of pMT3. Plasmids p618, p645 and p687 were obtained by excising the BglII-BgllI, SalI-SalI and EcoRI-EcoRI fragments, respectively, from pMTgB and cloning them into the compatible sites of pMT3. To construct plasmid p760, a 907 bp EcoRI subfragment of pMTgB was
clv
I I I CO,H CMV (ADI69) gB NH21 1 1 1 1 ~ I I [ ] 1 100 200 300 400 500 600 700 800 906 "
. . . . . . . . . . . . . CMV gB-(1-258) 1 258
1 411- . . . . . . . . . . CMV gB-(1-411) . . . . . . . . . CMV gB-(1-447)
1 447
1 476 . . . . . . . . CMV gB-(1-476) 1 618 . . . . . . CMV gB-(1-618) i 645- . . . . . CMV gB-(1-645) 1 687-- . . . . CMV gB-(1-687) 1 76ff - - - CMV gB-(l-760)
Fig. 1. Deletion constructs ofCMV (ADI69) gB. Designations for the products of the deletion constructs of the gB gene are shown on the right. The dashed region represents the deleted portion of the glycoprotein. The number of amino acids contained in the mutated forms ofgB is shown at the junction between the solid and dashed lines. The cleavage site in the intact gB molecule is indicated by an arrow (clv) (Spaete et al., 1988). The box indicates the transmembrane region.
first cloned into the EcoRI site of pUC18 and a SpeI linker containing stop codons in all three frames was added to the Bali site of the gB sequence. This subfragment, which contained a stop codon at residue 760, was cloned into the EcoRI site of the EagI fragment in the correct orientation.
Cells and medium. COS-1 cells were obtained from the American Type Culture Collection and were grown in Dulbecco's modified Eagle's minimum essential medium, supplemented with 10~ foetal bovine serum.
MAbs. The properties of the panel of MAbs to CMV (AD169) gB produced in this laboratory and used in this study have been reported (Banks et al., 1989; Pereira et al., 1982b, 1984). The mechanism of neutralization by these antibodies will be described elsewhere (D. Navarro et at., unpublished data). Cells were fixed in acetone and stained with MAbs after 48 h as previously described (Pereira et al., 1989). The pool of MAbs used for immunofluorescence reactions consisted of CH45-1, CH86-3, CH408-1 and CH386-3 which recognize continuous epitopes mapping in domain DClv (Table 2).
DNA transfections. Plasmid DNAs (10 to 20 ktg/106 cells) were precipitated on COS-I cells using calcium phosphate. Cells were exposed to DNA precipitates in medium containing chloroquine (100 ~tM) for 5 h, then rinsed and fresh medium was added.
Radiolabelling, immunoprecipitation and PAGE. Plasmid DNAs were precipitated on COS-1 ceils as described above and were labelled with [35S]methionine (5 ~tCi/ml; DuPont New England Nuclear) from 36 to 48 h post-transfection. The cells and culture medium were extracted with 1 ~ sodium deoxycholate, 1 ~ Nonidet P-40 and 0.1 ~ SDS. The extracts were reacted with the antibodies and the precipitates formed were eluted in SDS, boiled and electrophoresed in a denaturing polyacrylamide gel (9~ polyacrylamide cross-linked with diallyl- tartardiamide).
Results
Transient expression o f deletion constructs o f C M V g B
The structure of the delet ion der ivat ives in the gB gene of C M V (AD169) is shown in Fig. 1. Der ivat ives that

Antigenic domains assembled on C M V gB 2915
i i i i i
i
2 0 0 K - iii i i i i i , i ¸ ¸ ....
..... ~' :5 . ~ : :
Fig. 2. Immunofluorescence reactions of a pool of MAbs to CMV gB with products of deletion constructs o f the CMV gB gene. Cells were fixed in acetone at 48 h post-transfection and stained with a pool of MAbs to continuous epitopes mapping in domain DClv (Table 2). (a) Wild-type CMV gB, (b) gB-(1-258), (c) gB-(1-411), (d) gB-(1-447), (e) gB-(1-645), (f) gB-(1-760).
Fig. 3. Immunoprecipitates of selected truncated forms of CMV gB transiently expressed in COS-1 cells. Aliquots of radiolabelled cell extracts were reacted with the pool of MAbs to continuous epitopes mapping in domain DClv (Table 2). Lane M, Mr markers, gB bands contained in the precipitates are indicated by dots.
spanned the amino-terminal half of gB expressed the residues from positions I to 258, 411 and 447. The longer derivatives expressed the residues from positions 1 to 476, 618, 645, 687 and 760 in the carboxy-terminal half beyond the cleavage site between amino acids 460 and 461. In the first series of experiments, the derivatives of gB were expressed by transfecting COS-1 cells with plasmid DNAs containing the deletion constructs. Expression of the truncated glycoproteins was first monitored by immunofluorescence with a pool of MAbs to continuous epitopes mapping at the extreme amino terminus within residues 28 to 67 (Basgoz et al., 1992; Pereira et al., 1991). Each construct expressed a protein reactive with the antibody pool at levels approximately equivalent to that detected in cells transfected with the intact gB gene (Fig. 2). In general, the staining pattern exhibited by these mutated proteins did not differ significantly from the pattern observed in cells express- ing wild-type gB. The exception was gB-(1-411) (Fig. 2c), which showed a reticular distribution when compared with gB and the other derivatives.
To determine whether the actual sizes of the proteins agreed with estimates of their size based on the number
of residues contained in the derivatives, immunoprecipi- tation reactions were performed with the pool of antibodies to the extreme amino terminus of gB. Transfected COS-1 cells were radiolabelled and precipi- tated as described in Methods. Fig. 3 compares the electrophoretic mobilities of the mutated forms with that of intact gB. Analysis of the precipitates formed from the cell extracts showed that the truncated products mi- grated more slowly than the predicted size and appeared to be modified by the addition of sugars. The shift in mobility for each product depended on the number of N-glycosylation sites retained after truncation of the molecule, based on the predicted Asn-X-Ser/Thr consen- sus sequences in the AD169 gB gene (Cranage et al., 1986). The apparent Mr of the derivatives was as follows: gB-(1-258), 46K; gB-(1-411), 72K; gB-(1-447), 82K; gB- (1-476), 91K; gB-(1-618), 101K; gB-(1-645), 106K; gB- (1-687), 110K; gB-(1-760), 116K. Analysis of precipitates formed after the reaction of antibodies with the culture medium indicated that the truncated forms of gB were poorly released from cells (data not shown).
We had noted previously that trace amounts of cleaved gB were generated in transient expression of the CMV

2916 L Qadri and others
Table 1. Grouping of neutralizing MAbs to CMV (AD169) gB by immunofluorescence and immunoprecipitation reactions with truncated derivatives
Reaction with truncated derivatives Antigenic
region MAb 258* 411 447 476 618 645 687 760
DClv CH408-1 + + + + + + + + D1 CH177-3, CH253-1, CH358-5, -- - + + + + + +
CH382-2, CH388-4, CH395-1, CH87-1t, CH92-1 t, CH105-7~, CH112-1~
D2a CH244-4, CH130-9, CH143-13 - - - + + + + + D2b CH424-1, CH432-1, CH434-1, - - - - + + + +
CH436-1, CH442-1, CH446-2 D3 CH51-4, CHl14-5, CH409-2 . . . . . + + +
* Numbers indicate the length (no. of amino acids) of CMV (AD169) amino-terminal derivatives. t Immunofluorescence reactions were weak but reproducible with CMV gB-(1-447).
strain Towne gB gene (Banks et al., 1989). In the present study, the carboxy-terminal cleavage fragments expected to be generated from intact gB were again barely detectable. Cleavage of the longer derivatives gB- (1-645), gB-(1-687) and gB-(1-760) was not detected even though the size of these fragments would have been within the separation limits of this gel and others of higher acrylamide concentration. Insofar as intact gB and the longer derivatives are not cleaved in COS-1 cells (Fig. 3), it appears that this cell type does not contain significant amounts of the calcium-dependent protease found in CHO cells stably expressing a truncated form of the gB gene (Spaete et al., 1990). We have reported that the panel of MAbs to CMV gB immunoprecipitates both the intact form of gB and the faster migrating cleavage products (Pereira & Hoffman, 1986; Pereira et al., 1984); thus cleavage does not affect the conformation of these epitopes. In order to confirm that uncleaved gB molecules do not differ significantly in conformation from cleaved forms, a CHO cell line expressing the mutant CMV gB pXgB24clv4, which contains three altered residues near the cleavage site (Spaete et al., 1990), was reacted with a subset of MAbs to domains D1, D2a, D2b and D3 (Table 2) spanning the length of the molecule (D. Spaete, unpublished observations). It was found that the antibodies recognized the uncleaved derivative ofgB, indicating that lack of cleavage does not affect these epitopes.
MAbs recognize epitopes assembled by truncated derivatives of gB
In the next series of experiments, we located the amino acids required to assemble the conformation-dependent neutralizing epitopes on gB by reacting each truncated glycoprotein in immunofluorescence tests with the
individual MAbs. Results of these experiments (Table 1) are summarized as follows. (i) The shortest glycopro- teins, gB-(1-258) and gB-(1-411), did not assemble any of the discontinuous epitopes detected by the antibody panel. However, gB-(1-258) contained the continuous neutralizing epitope recognized by antibody CH408-1, which had been shown previously to map in domain DC 1 v in the extreme amino terminus (Basgoz et al., 1992; Meyer et al., 1992; Pereira et al., 1991). (ii) In contrast to the results obtained with gB-(1-411), analysis of the next longer form gB-(1-447), comprising almost the entire amino-terminal half of gB, showed that 10 conformation- dependent epitopes were present. Of these epitopes, five (CH177-3, CH253-1, CH358-5, CH382-2 and CH388-4) had complement-independent neutralizing activity and five had complement-dependent activity (Table 2) (D. Navarro et al., unpublished data). Four antibodies (CH87-1, CH92-1, CH105-7 and CHII2-1) reacted weakly but reproducibly with gB-(1-447), indicating that their epitopes, grouped here in domain D1, may be only partially assembled on this form of the molecule. The neutralizing domain assembled when amino acids 411 to 447 are present, designated domain D1, is the first discontinuous antigenic region to be identified in the amino-terminal half of CMV gB. (iii) Three additional discontinuous epitopes, recognized by neutralizing anti- bodies CH244-4, CH130-9 and CH143-13, were as- sembled on gB-(1-476). These epitopes were grouped into domain D2a and are assembled when residues 447 to 476 are present. (iv) Six more discontinuous epitopes, recognized by antibodies CH424-1, CH432-1, CH434-1, CH436-1, CH442-1 and CH446-2, were expressed on gB- (1-618). These epitopes were grouped into domain D2b, which is assembled by the addition of residues 476 to 618 in the carboxy-terminal half of gB. (v) gB-(1-645) contained epitopes for three neutralizing antibodies,

Antigenic domains assembled on C M V gB 2917
Table 2. Grouping o f 36 epitopes into antigenic domains on C M V (AD169) gB
Antigenic domain/ Neutralizing Epitopes expressed on gB deletion derivatives, epitope location MAb activity* fusion proteins and synthetic peptides
DCI~, amino terminus: 28 to 67 Present on AD169 gB-(I-258) and on fl-galactosidase N-1 CH408-1 - fusion protein, amino acids 27 to 84t N-2 CH45-1 N-3 CH86-3 N-4 CH386-3 N-5 CH396-3 N-6 CH404-4 N-7 CH412-2
DI, midregion: 411 to 447 M-1 CH177-3 + M-2 CH253-1 + M-3 CH358-5 + M-4 CH382-2 + M-5 CH388-4 + M-6 CH395-I - M-7 CH87-1 - M-8 CH92-1 - M-9 CHI05-7 - M-10 CHl12-1 -
D2a, midregion: 447 to 476 M-11 CH244-4 + M-12 CH130-9 + M-13 CH143-13 +
D2b, carboxy-terminal half: 476 to 618 C- 1 CH424-1 - C-2 CH432-1 + C-3 CH434-1 - C-4 CH442-1 + C-5 CH446-2 + C-6 CH436-1 +
D3, carboxy-terminal half: 618 to 645 C-7 CH51-4 - C-8 CH114-5 - C-9 CH409-2 -
DC2, intracellular: 716 to 906 I-1 CH158-2 I-2 CH216-2 I-3 CH380-4 1-4 CH381-1 1-5 CH385-3 1-6 CH402-5 1-7 CH410-3
DC3, intraceUular: 833 to 898 I-8 CH405-1 1-9 CH421-5 1-10 CH28-2
Assembled on gB-(1-447) but not gB-(1-411)
Assembled on gB-(1-476) but not gB-(1-447)
Assembled on gB-(1-618) but not gB-(1-476) (antigenic region identified by others:~)
Assembled on gB-(1-645) but not gB-(l-618) (antigenic region identified by otbers:~)
Present on gB-(716-906)t but not gB-(1-832)
Present on gB-(716-906)t but not gB-(1-832) Peptide (833 to 852) Peptide (833 to 852) Peptide (878 to 898)
* Complement-independent (D. Navarro et al., unpublished data). t (Basgoz et al., 1992; Meyer et al., 1992; Pereira et al., 1991). :~ (Kniess et al., 1991; Liu et al., 1991; Utz et al., 1989; Spaete et al., 1988).
CH51-4, CHl14-5 and CH409-2, shown in an earlier
study to be assembled when residues 619 to 680 are present (Banks et al., 1989). This result indica ted that the residues required for assembly of these d iscont inuous epitopes are located be tween amino acids 619 and 645 in the carboxy te rminus , in a d o m a i n which we have designated D3. (vi) gB-(1-687) and gB-(1-760) expressed all of the d iscont inuous epitopes con ta ined in gB-(1-645), but failed to express the cont inuous epitopes in domains
D C 2 and DC3, which had been previously m a p p e d in the intracel lular carboxy t e rminus (Basgoz et al., 1992;
Pereira et al., 1991).
Discontinuous epitopes assembled by residues in the amino-terminal hal f and midregion o f gB
Some of the epitopes found in the present study to assemble in domains D1 and D2 were reported to require
2918 I. Qadri and others
CMV gB-(1-447) CMV gB-(1-411)
7 t-~
97K-
69K--
46K--
on eq "7,
: i ̧̧ ' i ̧̧~:~: :iJ
~,~, , ............
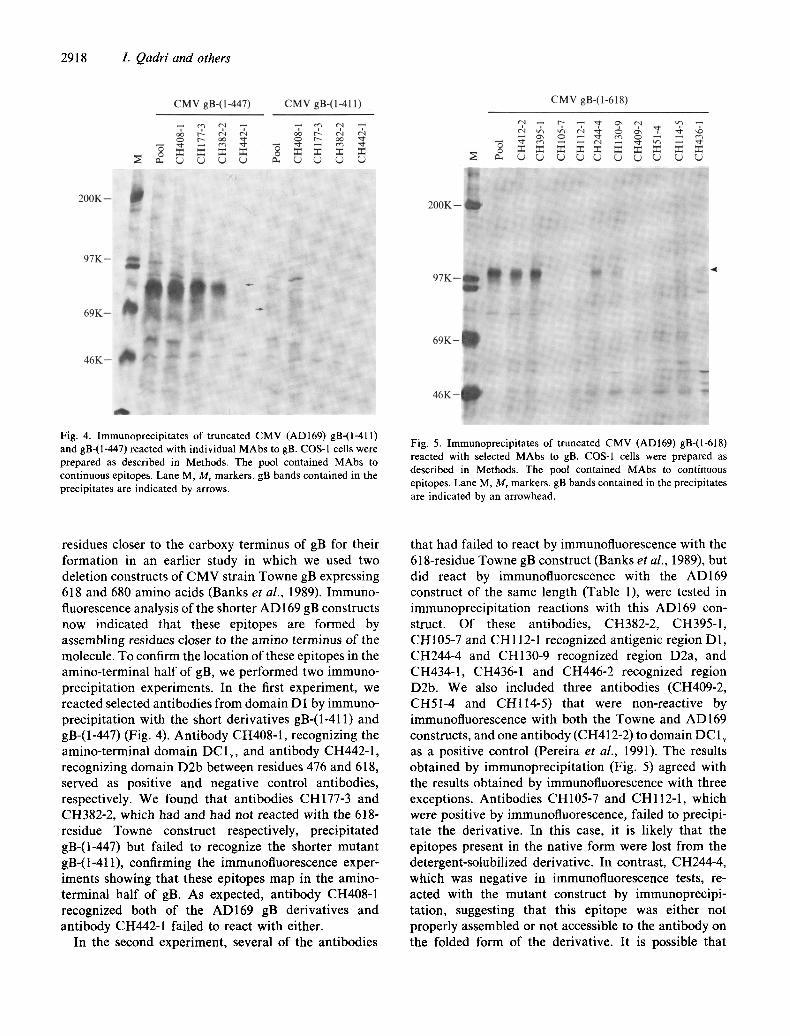
Fig. 4. Immunoprecipitates of truncated CMV (AD169) gB-(l-411) and gB-(1-447) reacted with individual MAbs to gB. COS-1 cells were prepared as described in Methods. The pool contained MAbs to continuous epitopes. Lane M, Mr markers, gB bands contained in the precipitates are indicated by arrows.
CMV gB-(1-618)
Fig. 5. Immunoprecipitates of truncated CMV (AD169) gB-(1-618) reacted with selected MAbs to gB. COS-I cells were prepared as described in Methods. The pool contained MAbs to continuous epitopes. Lane M, Mr markers, gB bands contained in the precipitates are indicated by an arrowhead.
residues closer to the carboxy terminus of gB for their formation in an earlier study in which we used two deletion constructs of CMV strain Towne gB expressing 618 and 680 amino acids (Banks et al., 1989). Immuno- fluorescence analysis of the shorter AD 169 gB constructs now indicated that these epitopes are formed by assembling residues closer to the amino terminus of the molecule. To confirm the location of these epitopes in the amino-terminal half of gB, we performed two immuno- precipitation experiments. In the first experiment, we reacted selected antibodies from domain D 1 by immuno- precipitation with the short derivatives gB-(1-411) and gB-(1-447) (Fig. 4). Antibody CH408-1, recognizing the amino-terminal domain DClv, and antibody CH442-1, recognizing domain D2b between residues 476 and 618, served as positive and negative control antibodies, respectively. We found that antibodies CH177-3 and CH382-2, which had and had not reacted with the 618- residue Towne construct respectively, precipitated gB-(1-447) but failed to recognize the shorter mutant gB-(1-41t), confirming the immunofluorescence exper- iments showing that these epitopes map in the amino- terminal half of gB. As expected, antibody CH408-1 recognized both of the AD169 gB derivatives and antibody CH442-1 failed to react with either.
In the second experiment, several of the antibodies
that had failed to react by immunofluorescence with the 618-residue Towne gB construct (Banks et al., 1989), but did react by immunofluorescence with the AD169 construct of the same length (Table 1), were tested in immunoprecipitation reactions with this AD169 con- struct. Of these antibodies, CH382-2, CH395-1, CH105-7 and CH112-1 recognized antigenic region D1, CH244-4 and CH130-9 recognized region D2a, and CH434-1, CH436-1 and CH446-2 recognized region D2b. We also included three antibodies (CH409-2, CH51-4 and CHl14-5) that were non-reactive by immunofluorescence with both the Towne and AD169 constructs, and one antibody (CH412-2) to domain DC l v as a positive control (Pereira et al., 1991). The results obtained by immunoprecipitation (Fig. 5) agreed with the results obtained by immunofluorescence with three exceptions. Antibodies CH105-7 and CHll2-1, which were positive by immunofluorescence, failed to precipi- tate the derivative. In this case, it is likely that the epitopes present in the native form were lost from the detergent-solubilized derivative. In contrast, CH244-4, which was negative in immunofluorescence tests, re- acted with the mutant construct by immunoprecipi- ration, suggesting that this epitope was either not properly assembled or not accessible to the antibody on the folded form of the derivative. It is possible that

Ant igen i c d o m a i n s a s s e m b l e d on C M V g B 2919
N M C 100 200 300 400 500 600
D1, ~)2 and D2b ~J II Major functional region: ~ [ promotes vlnon entry, I I cell-to-cen spread of I [ infection and fusion J [
I
700 800 906 1
N H 2 1 ~ CO2H I DC2 DC3 ~Continuous epitopes
Neutralizing domain lmmunodominant Neutralizing domain neutralizing Diseontinous epitopes Var ab e ep topes domain
Fig. 6. Topographic map of the antigenic and functional domains mapped on CMV (AD169) gB. Antigenic domains are shown as ellipses. Domains DCIv, DC2 and DC3 containing continuous epitopes were mapped previously (Basgoz et al., 1992; Pereira et aL, 1991). Boxes show our designations for the antigenic regions of gB and list the functional properties that have been identified for certain neutralizing domains (D. Navarro et al., unpublished data). Four antigenic regions of the gB molecule are indicated: N (amino terminus; positions 28 to 67), M (midregion; 411 to 476), C (carboxy terminus; 476 to 645) and I (intracellular; 719 to 906). The cleavage site is indicated by an arrow (clv) (Spaete et al., 1988). The shaded box designates the hydrophobic transmembrane domain (Cranage et aL, 1986). Immunodominant neutralizing domain D2b and neutralizing domain D3 contain antigenic sites mapped by other laboratories (Kniess et al., 1991 ; Liu et al., 1991 ; Spaete et aL, 1988; U tz et al., 1989).
different results obtained with the Towne construct in the previous study (Banks et al. , 1989) were due to small differences in the amino acid sequence between the strains; however, this seems unlikely since the antibodies that failed to react with the Towne derivative neverthe- less recognized epitopes shared by the intact gB of both strains AD169 and Towne. The most likely explanation is that the differences in reactivities with the Towne construct may have resulted from a small spontaneous in- frame internal deletion generated during the construc- tion of this mutant gB. Particularly relevant to the folding of the truncated forms of gB are four cysteine residues mapping between amino acids 411 and 618. A mutation that altered a cysteine residue might have led to mispairing of the cysteines, causing formation of mismatched disulphide bonds, partial malfolding of the molecule and loss of selected epitopes. The detailed analysis of the set of eight deletion derivatives in the AD169 gB gene that we have now completed enables us to locate correctly the amino acids required to assemble the conformation-dependent epitopes in the various antigenic regions of the molecule.
Discussion
In this study, we generated from the CMV gB gene a set of deletion constructs that lacked different lengths of the sequences specifying the carboxy terminus, in order to
locate residues that confer reactivity with our panel of MAbs. Table 2 summarizes our findings to date regarding the antigenic regions of CMV (AD169) gB, which have been analysed by identifying amino acids required to assemble the epitopes recognized by neutral- izing antibodies. The locations of these regions are summarized in a schematic diagram (Fig. 6). We have divided the gB molecule into four antigenic regions: the amino (N) terminus residues from 28 to 67, the midregion (M) residues from 411 to 476, the carboxy (C) terminus residues from 476 to 645, and the intracellular (I) residues from 716 to 906. Epitopes of antibodies recognizing each antigenic region have been grouped in Table 2 by their location in the gB molecule, e.g. N-I, M-l, etc.
We have identified three regions of continuous epitopes: DClv, a neutralizing domain in the amino terminus region which contains strain-specific epitopes, and DC2 and DC3, located in the intracellular region of the molecule (Basgoz et al. , 1992; Pereira et al. , 1991). Our work and that of others (Meyer et al. , 1990) has shown that nucleotide sequence differences in the extreme amino terminus of gB specified by strains AD169 and Towne (Spaete et al. , 1988) affect their antigenic properties. Both conserved and variable antigenic sites are contained in the region encompass- ing domain DClv (Meyer et al. , 1992). Other investi- gators have identified continuous neutralizing epitopes mapping between residues 589 and 645 (Kniess et al. ,
1991) and between 608 and 625 (Utz et al., 1989). Discon- tinuous neutralizing epitopes are assembled in four domains, D1, D2a and D2b, and D3. Domain D1, one of two novel conformation-dependent domains identified in this study, is assembled in part by the folding of residues 411 to 447 in the amino-terminal half of gB. Domain D2a, the second novel domain, is assembled in the midregion by residues 447 to 476, spanning the cleavage site. Domain D2b, also in the midregion, is assembled by amino acids 476 to 618, and domain D3 by amino acids 619 to 645 in the carboxy-terminal half of the molecule. The antigenic region designated AD-1 by others (Kniess et al. , 1991), which contains a continuous epitope mapping between residues 589 and 645, partially overlaps domains D2 and D3. In locating the recognition sites of a large number of neutralizing MAbs to the region of gB between residues 476 and 645, our studies and those of others suggest that this region is immuno- dominant (Banks et al . , 1989; Kniess et al. , 1991 ; Liu et
al . , 1991; Spaete et al . , 1988; Utz e t al. , 1989). Since most of the neutralizing epitopes in our panel of
antibodies are dependent on the conformation of gB, the formation of the antibody recognition sites may require the juxtaposition not only of discontinuous amino acids composing those sites but also of other discontinuous

2920 I. Qadri and others
residues in the amino-terminal half of gB that promote the proper folding of the antigenic domains identified in the present study. We are in the process of constructing mutant gB molecules with small deletions spanning the antigenic domains of the molecule to distinguish more precisely the amino acids that affect the epitope folding from those which are assembled into the antibody recognition site (I. Qadri & L. Pereira, unpublished data). Preliminary analysis of the antigenic properties of these mutated forms showed that deletion of residues 170 to 411 and of residues 411 to 447 precludes assembly of domain D1 epitopes. This finding shows that amino acids 170 to 411 also play a role in assembling these epitopes. In addition, domain D3 was perturbed by deleting residues 411 to 447, which supports our finding that folding of the carboxy-terminal half of gB depends on certain residues in the amino-terminal half of the molecule. Epitopes in domain D3 are lost when residues 548 to 618 are deleted, suggesting that adjacent residues participate in the antigenic structure of this region. No antigenic changes were induced by deleting residues 645 to 702, suggesting that this region is not directly involved in assembling epitopes in the extracellular region of gB, as indicated in the present study. We recently carried out a detailed analysis of the neutralizing activities of complement-independent antibodies to CMV gB, which revealed that they prevent virion entry by blocking fusion of the viral envelope with the cell membrane, the spread of infection from cell to cell, and the formation of syncytia by infected glioblastoma cells (D. Navarro et al., unpublished data). The detailed mutational analysis of CMV gB now underway will enable us to formulate a model for the folding of the molecule that is consistent with its antigenic structure and its function in virion infectivity and the cell-to-cell transmission of infection.
We thank Richard Spaete for analysing the CHO cell line expressing the CMV gB mutant pXgB24clv4 with the panel of MAbs and Scott Frank for assisting in the analysis of neutralizing epitopes. These studies were supported by Public Health Service grants A123592 and AI30873 from the National Institute of Allergy and Infectious Diseases and by grant HL33713 from the National Heart and Lung Institute to Stanford University. D.N. was supported by a fellowship from the Spanish Ministry of Education and Science.
References
BANKS, T., HUO, B., KOUSOULAS, K., SPAETE, R., PACHL, C. & PEREIRA, L. (1989). A major neutralizing domain maps within the carboxyl- terminal half of the cleaved cytomegalovirus B glycoprotein. Journal of General Virology 70, 979 985.
BAS6OZ, N., QADR1, I., NAVARRO, D., SEARS, A., LENNETTE, E., YOUNGBLOM, J. & PEREIRA, L. (1992). The amino terminus of human cytomegalovirus glycoprotein B contains epitopes that vary among strains. Journal of General Virology 73, 983-988.
BRITT, W. J. (1984). Neutralizing antibodies detect a disulfide-linked glycoprotein complex within the envelope of human cytomegalo- virus. Virology 135, 369-378.
BRITT, W. J. & VUGLER, L. G. (1990). Antiviral antibody responses in mothers and their newborn infants with clinical and subclinical congenital cytomegalovirus infections. Journal of Infectious Diseases 161, 214-219.
BR1Tlr, W. J., VUGLER, L. & STEPHENS, E. B. (1988). Induction of complement-dependent and -independent neutralizing antibodies by recombinant-derived human cytomegalovirus gp55-116 (gB). Journal of Virology 62, 3309-3318.
CHEE, i . S., BANKIER, A. T., BECK, S., BOHNI, R., BROWN, C. M., CERNY, R., HORSNELL, T., HUTCHISON, C. A., KOUZARIDES, T., MARTIGNETTI, J. A., PREDDIE, E., SATCHWELL, S. C., TOMLINSON, P., WESTON, K. M. & BARRELL, B. G. (1990). Analysis of the protein- coding content of the sequence of human cytomegalovirus strain AD169. Current Topics in Microbiology and Immunology 154, 125- 170.
CHou, S. (1992). Comparative analysis of sequence variation in gp116 and gp55 components of glycoprotein B of human cytomegalovirus. Virology 188, 388-390.
CHOU, S. W. & DENNISON, K. M. 0991). Analysis of interstrain variation in cytomegalovirus glycoprotein B sequences encoding neutralization-related epitopes. Journal of Infectious Diseases 163, 1229 1234.
CRANAGE, M. P., KOUZARIDES, T., BANKIER, A. T., SATCHWELL, S., WESTON, K., TOMLINSON, P., BARRELL, B., HART, H., BELL, S. E., MINSON, A. C. & SMITH, G. L. (1986). Identification of the human cytomegalovirus glycoprotein B gene and induction of neutralizing antibodies via its expression in recombinant vaccinia virus. EMBO Journal 5, 3057-3063.
DREW, L. (1988). Cytomegalovirus infection in patients with AIDS. Journal of Infectious Diseases 158, 449-456.
FARRAR, G. H. & GREENAWAY, P. J. (1986). Characterization of glycoprotein complexes present in human cytomegalovirus envelopes. Journal of General Virology 67, 1469-1473.
FLECKENSTEIN, B., MULLER, I. & COLLINS, J. (1982). Cloning of the complete human cytomegalovirus genome as cosmids. Gene 18, 3%46.
GONCZOL, E., HUDECZ, F., IANACONE, J., DIETZSCHOLD, B., STARR, S. & PLOTKIN, S. A. (1986). Immune responses to isolated human cytomegalovirus envelope proteins. Journal of Virology 58, 661- 664.
JACOBSON, M. A. • MILLS, J. (1988). Serious cytomegalovirus disease in the acquired immunodeficiency syndrome (AIDS). Annals of Internal Medicine 108, 585-594.
KNIESS, N., MACH, M., FAY, J. & BRITT, W. J. (1991). Distribution of linear antigenic sites on glycoprotein gp55 of human cytomegalo- virus. Journal of Virology 65, 138-146.
Ltu, Y.-N. C., KLAUS, A., KARl, B., STINSKI, M. F., ECKHARDT, K. & GEHRZ, R. C. (1991). The N-terminal 513 amino acids of the envelope glycoprotein gB of human cytomegalovirus stimulates both B- and T-cell immune responses in humans. Journal of Virology 65, 1644 1648.
MEYER, H., MASUHO, Y. & MACH, M. (1990). The gpl l6 of the gp58/116 complex of human cytomegalovirus represents the amino- terminal part of the precursor molecule and contains a neutralizing epitope. Journal of General Virology 71, 2443-2450.
MEYER, H., SUNDQVIST, V.-A., PEREIRA, L. & MACH, M. (i992). Glycoprotein gpl 16 of human cytomegalovirus contains epitopes for strain-common and strain-specific antibodies. Journal of General Virology 73, 2375-2383.
MEYERS, J. D., FLOURNOY, N. &THOMAS, E. D. (1986). Risk factors for cytomegalovirus infection after human marrow transplantation. Journal of lnfectious Diseases 153, 478-488.
PAss, R. F., STAGNO, S., MYERS, G. J. & ALFORD, C. A. (1980). Outcome of symptomatic congenital cytomegalovirus infection: results of long-term longitudinal follow-up. Pediatrics 66, 758-762.
PELLETr, P. E., KOUSOULAS, K. G., PEREIRA, L. & ROIZMAN, B. (1985). Anatomy of the herpes simplex virus 1 strain F glycoprotein B gene: primary sequence and predicted protein structure of the wild type and of monoclonal antibody-resistant mutants. Journal of Virology 53, 243-253.
PEPOSE, J. (1989). Cytomegalovirus infections of the retina. In Retina, pp. 589-596. Edited by S. J. Ryan. St Louis: C. V. Mosby Co.

Antigenic domains assembled on C M V gB 2921
PEREmA, L. & HOFFMAN, M. (1986). Immunology of human cyto- megalovirus glycoproteins. In Human Herpesvirus Infections: Patho- genesis, Diagnosis, and Treatment, pp. 69-92. Edited by C. Lopez & B. Roizman. New York: Raven Press.
PEREIRA, L., HOFFMAN, M. & CREMER, N. (1982a). Electrophoretic analysis of polypeptides immune precipitated from extracts of cytomegalovirus infected cells by human sera. Infection and Immunity 36, 933-942.
PEREIRA, L., HOFFMAN, M., GALLO, O. & CREMER, N. (1982b). Monoclonal antibodies to human cytomegalovirus. I. Three cell surface proteins with unique immunologic and electrophoretic properties specify cross-reactive determinants. Infection and Immunity 36, 924 932.
PEREIRA, L., STAGNO, S., HOFFMAN, M. & VOLANAKIS, J. (1983). Cytomegalovirus infected cell polypeptides immune precipitated by children with congenital and perinatal cytomegalovirus infections. Infection and Immunity 39, 100-108.
PEREIRA, L., HOFFMAN, M., TATSUNO, M. & DONDERO, D. (1984). Polymorphism of human cytomegalovirus glycoproteins character- ized by monoclonal antibodies. Virology 139, 73-86.
PERE1RA, L., ALl, M., KOUSOULAS, K., Huo, B. & BANKS, T. (1989). Domain structure of herpes simplex virus 1 glycoprotein B: neutralizing epitopes map in regions of continuous and discon- tinuous residues. Virology 172, 11 24.
PEREIRA, L., BASGOZ, N., NAVARRO, D., QADRI, I., YOUNGBLOM, J., SEARS, A. & LENNETI'E, E. (1991). Mapping conserved antigenic regions and strain-specific epitopes within the glycoprotein B gene of human cytomegalovirus. In Progress in Cytomegalovirus Research, pp. 195-198. Edited by M. P. Landini. Amsterdam: Elsevier.
PEREIRA, L., JAHN, G. & NAVARRO, n. (1992). Proteins of human cytomegalovirus that elicit humoral immunity. In Molecular Aspects of Human Cytomegalovirus Diseases. Edited by Y. Becker, G. Darai & E.-S. Huang. Heidelberg: Springer-Verlag (in press).
RADSAK, K., BRUCHER, K. H., BRITT, W., SHIOU, H., SCHNEIDER, D. & KOLLERT, A. (1990). Nuclear compartmentation of glycoprotein B of human cytomegalovirus. Virology 177, 515-522.
RASMUSSEN, L., MULLENAX, J., NELSON, M. & MERIGAN, T. C. (1985). Human cytomegalovirus polypeptides stimulate neutralizing anti- body in vivo. Virology 145, 18(~190.
RASMUSSEN, U, NELSON, M., NEFF, M. & MERIGAN, T. C. (1988). Characterization of two different human cytomegalovirus glycopro- reins which are targets for virus neutralizing antibody. Virology 163, 309-318.
RASMUSSEN, L., MATKIN, C., SPAETE, R., PACHL, C. & MERIGAN, T~ C. (1991). Antibody response to human cytomegalovirus glycoproteins gB and gH after natural infection in humans. Journal of Infectious Diseases 164, 835-842.
SINGH, N., DUMMER, J. S., KUSNE, S., BREINIG, M. K., ARMSTRONG, J. A., MAKOWKA, L., STARZL, T. E. & Ho, M. (1988). Infections with cytomegalovirus and other herpesviruses in 121 liver transplant recipients: transmission by donated organ and the effect of OKT3 antibodies. Journal of Infectious Diseases 158, 124-131.
SPAETE, R. R., THAYER, R. M., PROBERT, W. S., MASIARZ, F. R , CHAMBERLAIN, S. H., RASMUSSEN, L., MERIGAN, T. C. & PACHL, C. (1988). Human cytomegalovirus strain Towne glycoprotein B is processed by proteolytic cleavage. Virology 167, 207-225.
SPAETE, R. R., SAXENA, A., SCOTT, P. I., SONG, G. J., PROBERT, W. S., BRiar, W. J., GmSON, W., RASMUSSEN, L. & PACHL, C. (1990). Sequence requirements for proteolytic processing of glycoprotein B of human cytomegalovirus strain Towne. Journal of Virology, 64, 2922-2931.
STAGNO, S., PASS, R. F., DWORSKY, M. E. & ALFORD, C. A. (1983). Congenital and perinatal cytomegalovirus infections. Seminars in Perinatology 7, 31-42.
UTZ, U., BRITr, W., VUGLER, L. & MATH, M. (1989). Identification of a neutralizing epitope on glycoprotein gp58 of human cytomegalovirus. Journal of Virology 63, 1995-2001.
WONG, G. G., WITEK, J. S., TEMPLE, P. A., WILKENS, K. M., LEARY, A. C., LUXENBERG, D. P., JONES, S. S., BROWN, E. L., KAY, R. M., ORR, E. C., SHOEMAKER, C., GOLDE, D. W., KAUFMAN, R. J., HEWlCK, R. M., WANG, E. A. & CLARK, S. C. (1985). Human GM-CSF: molecular cloning of the complementary DNA and purification of the natural and recombinant proteins. Science 228, 810-812.
(Received 11 May 1992; Accepted 17 July 1992)
Copyright © 2022 FDOKUMEN
















![Problems with a conformation assignment of aryl-substituted resorc[4]arenes](https://static.fdokumen.com/doc/165x107/6324d12685efe380f30661c8/problems-with-a-conformation-assignment-of-aryl-substituted-resorc4arenes.jpg)



