
Conformational constraints imposed on a pan-neutralizing HIV1 antibody epitope result in increased...
15
Vaccine 23 (2005) 1559–1573 Conformational constraints imposed on a pan-neutralizing HIV-1 antibody epitope result in increased antigenicity but not neutralizing response Jason Ho a , Robert A. Uger b,1 , Michael B. Zwick c , Mark A. Luscher d,e , Brian H. Barber a,1 , Kelly S. MacDonald a,d,f,∗ a Department of Immunology, University of Toronto, Toronto, Ont., Canada M5S 1A8 b Immunology Department, Aventis Pasteur, 2075 Bayview Avenue, Toronto, Ont., Canada M4N 3M5 c Department of Immunology, IMM-2, Scripps Research Institute, La Jolla, CA 92037, USA d Department of Medicine, University of Toronto, Toronto, Ont., Canada M5S 1A8 e Department of Microbiology, University Health Network, Toronto, Ont., Canada M5G 2C4 f Department of Microbiology, Mt. Sinai Hospital Room 1484, 600 University Avenue, Toronto, Ont., Canada M5G 1X5 Received 25 May 2004; received in revised form 21 September 2004; accepted 24 September 2004 Available online 11 November 2004 Abstract 2F5 is one of the few broadly neutralizing monoclonal antibodies against type 1 Human Immunodeficiency Virus (HIV-1). It recognizes the amino acid sequence ELDKWAS in gp41. We have previously identified a number of immunotargeting 2F5-reactive candidate immunogens. Three of them (designated H-BT1–3) have the ELDKWAS sequence constrained at -turn sites within the immunoglobulin heavy chain. Two others (L-CT and L-CTx3) have the sequence attached at the C-terminus of the immunoglobulin light chain with minimal conformational constraints. In the present investigation, the H-BTs were found to bind 2F5 with up to 10-fold higher affinities than their unconstrained coun- terpart. When used as immunogens, immunogen-specific antibodies were induced with or without adjuvant, confirming the immunotargeting potential of these immunogen constructs. While HIV-1 gp160 cross-reactive antibodies were induced, virus neutralization was not detected. Thus, factors other than 2F5 binding affinity may have a critical role to play in the design of a 2F5-based vaccine. © 2004 Elsevier Ltd. All rights reserved. Keywords: Antibody engineering; Immunotargeting; HIV-1; gp41 1. Introduction In the global effort to develop a vaccine against HIV-1, strategies aimed at inducing mainly T cell mediated immune responses in the macaque model have been shown to con- fer partial protection against infection by the hybrid Simian- Human Immunodeficiency Virus (SHIV) through reducing ∗ Corresponding author. Tel.: +1 416 586 8879; fax: +1 416 586 8746. E-mail address: [email protected] (K.S. MacDonald). 1 Robert A. Uger and Brian H. Barber are no longer affiliated with the institutions indicated. set point plasma viral load and delaying disease progression (reviewed in [1]). Yet, subsequent emergence of viral escape mutants [2] suggests that protection against the establishment of infection may be transient. The goal of inducing steriliz- ing protective immunity thus remains the ultimate endpoint in the search for an HIV-1 vaccine. A number of recent reports have demonstrated that passive transfer of broadly neutraliz- ing monoclonal antibodies against HIV-1 could completely protect macaques against infection with SHIV (reviewed in [3]). One of these broadly neutralizing monoclonal antibodies is the 2F5 antibody [4]. Developing an immunogen capable of inducing antibodies with a similar neutralizing capacity 0264-410X/$ – see front matter © 2004 Elsevier Ltd. All rights reserved. doi:10.1016/j.vaccine.2004.09.037
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Conformational constraints imposed on a pan-neutralizing HIV1 antibody epitope result in increased...

Vaccine 23 (2005) 1559–1573
Conformational constraints imposed on a pan-neutralizing HIV-1antibody epitope result in increased antigenicity but not
neutralizing response
Jason Hoa, Robert A. Ugerb,1, Michael B. Zwickc, Mark A. Luscherd,e,Brian H. Barbera,1, Kelly S. MacDonalda,d,f,∗
a Department of Immunology, University of Toronto, Toronto, Ont., Canada M5S 1A8b Immunology Department, Aventis Pasteur, 2075 Bayview Avenue, Toronto, Ont., Canada M4N 3M5
c Department of Immunology, IMM-2, Scripps Research Institute, La Jolla, CA 92037, USAd Department of Medicine, University of Toronto, Toronto, Ont., Canada M5S 1A8
e Department of Microbiology, University Health Network, Toronto, Ont., Canada M5G 2C4f Department of Microbiology, Mt. Sinai Hospital Room 1484, 600 University Avenue,
Toronto, Ont., Canada M5G 1X5
Received 25 May 2004; received in revised form 21 September 2004; accepted 24 September 2004Available online 11 November 2004
A
nizes thea unogens.T woo rmationalc ned coun-t otargetingp t detected.T©
K
1
srfH
i
ssionape
mentiliz-nt inortsraliz-telyd indiesblecity
0d
bstract
2F5 is one of the few broadly neutralizing monoclonal antibodies against type 1 Human Immunodeficiency Virus (HIV-1). It recogmino acid sequence ELDKWAS in gp41. We have previously identified a number of immunotargeting 2F5-reactive candidate immhree of them (designated H-BT1–3) have the ELDKWAS sequence constrained at�-turn sites within the immunoglobulin heavy chain. Tthers (L-CT and L-CTx3) have the sequence attached at the C-terminus of the immunoglobulin light chain with minimal confoonstraints. In the present investigation, the H-BTs were found to bind 2F5 with up to 10-fold higher affinities than their unconstraierpart. When used as immunogens, immunogen-specific antibodies were induced with or without adjuvant, confirming the immunotential of these immunogen constructs. While HIV-1 gp160 cross-reactive antibodies were induced, virus neutralization was nohus, factors other than 2F5 binding affinity may have a critical role to play in the design of a 2F5-based vaccine.2004 Elsevier Ltd. All rights reserved.
eywords:Antibody engineering; Immunotargeting; HIV-1; gp41
. Introduction
In the global effort to develop a vaccine against HIV-1,trategies aimed at inducing mainly T cell mediated immuneesponses in the macaque model have been shown to con-er partial protection against infection by the hybrid Simian-uman Immunodeficiency Virus (SHIV) through reducing
∗ Corresponding author. Tel.: +1 416 586 8879; fax: +1 416 586 8746.E-mail address:[email protected] (K.S. MacDonald).
1 Robert A. Uger and Brian H. Barber are no longer affiliated with thenstitutions indicated.
set point plasma viral load and delaying disease progre(reviewed in[1]). Yet, subsequent emergence of viral escmutants[2] suggests that protection against the establishof infection may be transient. The goal of inducing stering protective immunity thus remains the ultimate endpoithe search for an HIV-1 vaccine. A number of recent rephave demonstrated that passive transfer of broadly neuting monoclonal antibodies against HIV-1 could compleprotect macaques against infection with SHIV (reviewe[3]). One of these broadly neutralizing monoclonal antibois the 2F5 antibody[4]. Developing an immunogen capaof inducing antibodies with a similar neutralizing capa
264-410X/$ – see front matter © 2004 Elsevier Ltd. All rights reserved.oi:10.1016/j.vaccine.2004.09.037

1560 J. Ho et al. / Vaccine 23 (2005) 1559–1573
comparable to 2F5 would, therefore, be a very useful com-ponent of an effective HIV-1 vaccine.
2F5 recognizes the seven amino acid sequence ELD-KWAS in gp41 [4–6]. Several laboratories have attemptedto induce neutralizing antibodies by incorporating the EL-DKWA sequence into various immunogenic molecules in-cluding the Hepatitis B surface antigen[7], HIV-1 gp120[8],and theEscherichia coliderived MalE protein[9], or by con-straining an ELDKWAS containing peptide[10,11], then im-munizing animals with these recombinant proteins. However,despite the induction of ELDKWA-specific antibodies in eachof those instances, virus neutralization was not observed inmost cases. These findings suggest that the conformation inwhich the ELDKWAS sequence was incorporated in theseimmunogens may not mimic the conformation of the native2F5 epitope, resulting in the induction of antibodies that haddifferent binding characteristics from those of 2F5. Under-standing the structure of the native 2F5 epitope is thereforeimportant if an effective ELDKWAS-based immunogen is tobe made.
We reported earlier the construction of a series of immuno-gens that incorporate the ELDKWAS sequence in the frame-work of an anti-HLA-DR immunoglobulin molecule[12].This approach provided two significant advantages. First, byselectively incorporating the ELDKWAS sequence in vari-o les,a ughtt then classI an-t Thiss ducea ad-ja sub-s vantl icu-l . Weh le int ASs H-B andt in-c ents( con-s sitesw ive2 n.I allr for-e uctsw ns.
-CTs rthers ncesi in-
duce ELDKWAS-specific antibody responses in vivo. Usingsurface plasmon resonance (SPR) technologies, it was foundthat the constructs containing the�-turn-constrained ELD-KWAS sequence were able to bind 2F5 with 5- to 10-foldhigher affinities than the unconstrained counterpart. Whenused as immunogens, these constructs were found to be ableto induce gp160 cross-reactive antibodies even without theaddition of extraneous adjuvants. This induction of antibodyresponses was found to be dependent on the class II MHCspecificity that was built into the immunogens. These find-ings demonstrate that immunotargeting is a useful strategy forvaccine designs that are aimed at inducing epitope-specificantibody responses. These studies also provide important in-sights with respect to the development of an HIV neutralizingantibody response based on the 2F5 epitope.
2. Methods
2.1. Cells and media
Unless otherwise stated, BTI-TN-5B1-4, also known asHigh Five
TMcells (Invitrogen, Carlsbad, CA), were cultured
using supplemented Express Five serum free medium (SFM)(Invitrogen). For each litre of SFM, the following supple-m )i cin(N vit-r le-m II.B n at2
2
lightc idatevI ructs,r vari-o pro-d sesw lightc entlyrB putu werep ALp nicp tD rus( ALc lightc trans-p n pu-
us conformational contexts in immunoglobulin molecund comparing their capacity to bind 2F5, the strategy so
o identify the structural context required for mimicry ofative 2F5 epitope. Second, by targeting the antigen to
I MHC expressing cells, it was designed to augmentibody responses in an adjuvant-independent manner.trategy, called immunotargeting, has been shown to inntibody responses without the addition of extraneous
uvants that are normally required (reviewed in[13]). Thebility to induce adjuvant-independent responses hastantial practical significance since alum, the only adjuicensed for use in human in North America, is not partarly effective in inducing immunogen-specific responsesave shown previously that 2F5 reactivity was detectab
hree of the recombinant constructs bearing the ELDKWequence at�-turn constraining sites (designated H-BT1,T2, and H-BT3), as well as in two others that had one
hree copies, respectively, of the ELDKWAS sequenceorporated as minimally constrained C-terminal attachmdesignated L-CT and L-CTx3). In contrast, none of thetructs incorporating the ELDKWAS sequence at helicalere recognized by 2F5[12]. This suggests that the natF5 epitope is more�-turn like than helical in conformatio
n addition, the HLA-DR binding capacity was retained inecombinant constructs despite the incorporation of theign ELDKWAS sequence, indicating that these constrould still constitute viable immunotargeting immunogeIn order to assess the potential of the H-BT and the L
eries of constructs as immunogens against HIV-1, futudies were carried out to further investigate the differen their 2F5 binding characteristics, and their abilities to
ent were added: (1) 90 ml of 20 mMl-glutamine (Sigman 0.89% NaCl, and (2) 10 ml of penicillin and streptomy10 000 units/ml and 10 000�g/ml, respectively) in 0.89%aCl (Invitrogen). Unless otherwise stated, Sf9 cells (In
ogen) was cultured in Sf-900II SFM (Invitrogen) suppented with in 0.89% NaCl (Invitrogen) per litre of Sf900oth High Five and Sf9 cells were grown in suspensio8◦C on an orbitalplatform shaking at 140 rpm.
.2. Expression and purification of protein constructs
Construction of the genes encoding the heavy and thehains of the immunoglobulin molecules used as candaccines in pIZ plasmids have been described elsewhere[12].n order to scale up production of these candidate constecombinant baculoviruses expressing the genes of theus heavy and light chains were constructed for proteinuction in High Five cells. These recombinant baculoviruere created by transferring the various heavy chain andhain genes into pFastBacDUAL plasmid, and subsequecombined into the baculovirus genome using theBac-To-ac® system (Invitrogen). The light chain genes werender the p10 promoter, and the heavy chain genesut under the polyhedrin promoter in the pFastBacDUlasmid. The procedure for the making of the bi-cistroFastBacDUAL constructs is shown inFig. 1. CompetenH10BacE. colicells containing the genome of baculovi
bacmid) were transformed with the various pFastBacDUonstructs. The segment containing the heavy and thehain encoding genes was integrated through specificoson sites. The resulting recombinant bacmid was the

J. Ho et al. / Vaccine 23 (2005) 1559–1573 1561
Fig. 1. Schematic diagram on the construction of bi-cistronic plasmids used in the making of recombinant baculoviruses for producing the various candidatevaccines.
rified and used in transfection of Sf9 insect cells (Invitrogen)to yield recombinant baculoviruses. For each construct, oneclone was selected by picking a plaque, and subsequently test-ing for the expression of antibodies. The titres of the plaquepurified virus stocks were amplified by several rounds of in-fection of Sf9 cells at a multiplicity of infection (MOI) ofaround 0.1.
To produce and purify the various antibody constructs,High Five cells were infected with the amplified recombinantbaculovirus stock at a higher MOI of 10. Supernatants wereharvested after three days for purification of antibodyconstructs. Supernatants were clarified by centrifugation,first at 500×g to remove the cells, then at 13 000×g toremove residual debris. Recombinant antibodies were thenpurified from the supernatants by immobilized Protein Aor Protein G (Pierce, Rockford, IL). Columns were washedextensively with Immunopure IgG Binding Buffer (Pierce).Bound antibodies were eluted in 1 ml fractions using 50 mMglycine, 150 mM NaCl, pH 2.8, and immediately neutralizedby 1/10 volume of 1 M Tris pH 9.5. Antibody positive frac-
tions, as judged by SDS–PAGE, were pooled, concentrated,and exchanged into clinical grade saline using MilliporeCentricon-Plus 20 concentrator (Millipore, Billerica, MA)with a molecular weight cut-off 10 000. The concentratedpreparations were then sterilized by filtering through 0.22�mlow protein binding Millex GV syringe filters (Millipore). Re-sulting protein samples were analyzed using 10% acrylamideBis-Tris SDS–PAGE, which was visualized by staining withSYPRO orange (Molecular Probes, Eugene, OR) accordingto manufacturer’s instructions. Protein concentration was de-termined using Bichinoninic Acid Assay (Pierce) as directedby manufacturer. Samples were stored in 4◦C until use.
2.3. Antibodies and peptides
The Fab′ fragment of 2F5[14], obtained by partial re-duction of the 2F5 F(ab′)2 [15], was generously provided byDr. Emil Pai at the University of Toronto. Antibody 33D-1[16] was generously provided by Aventis Pasteur (Toronto,Canada). Free ELDKWAS peptide was purified by high per-
1562 J. Ho et al. / Vaccine 23 (2005) 1559–1573
formance liquid chromatography and was also provided byAventis Pasteur.
2.4. Preparation of surfaces and regeneration
Surface plasmon resonance experiments were performedusing a Biacore 3000 instrument (Uppsala, Sweden). To ana-lyze binding between 2F5 and free ELDKWAS peptide, 2000response unit (RU) of 2F5 Fab′ was coated on the specific flowcell, while the same amount of an irrelevant antibody (33D-1)was coated on the control flow cell. To immobilize these lig-ands, carboxyl groups on CM5 chips (Biacore) were activatedusing 1-ethyl-3-(3-dimethylaminopropyl)carbodiimide hy-
drochloride (EDC) andN-hydroxysuccinimide (NHS)(Biacore) at a flow rate of 5�l/min for 7 min. Immunoglob-ulins were diluted to 30�g/ml in 10 mM sodium acetate. For2F5 Fab′, a pH of 5.5, determined empirically, was used; for33D-1, a pH of 4.0. In each case, the ligand was injected until1500 RU was immobilized. Unreacted carboxyl groups wereblocked with a 7 min injection of 1 M ethanolamine pH 8.5.Varying amount of free ELDKWAS peptide was then passedover these surfaces at 20�l/min for 3 min. Surfaces weresubsequently regenerated with a 30 s pulse of 1 mM NaOH.
To analyze the binding between 2F5 and the various candi-date immunogens, the constructs containing the ELDKWASsequence were coated on the specific flow cell, and an equal
FEIthtcTo
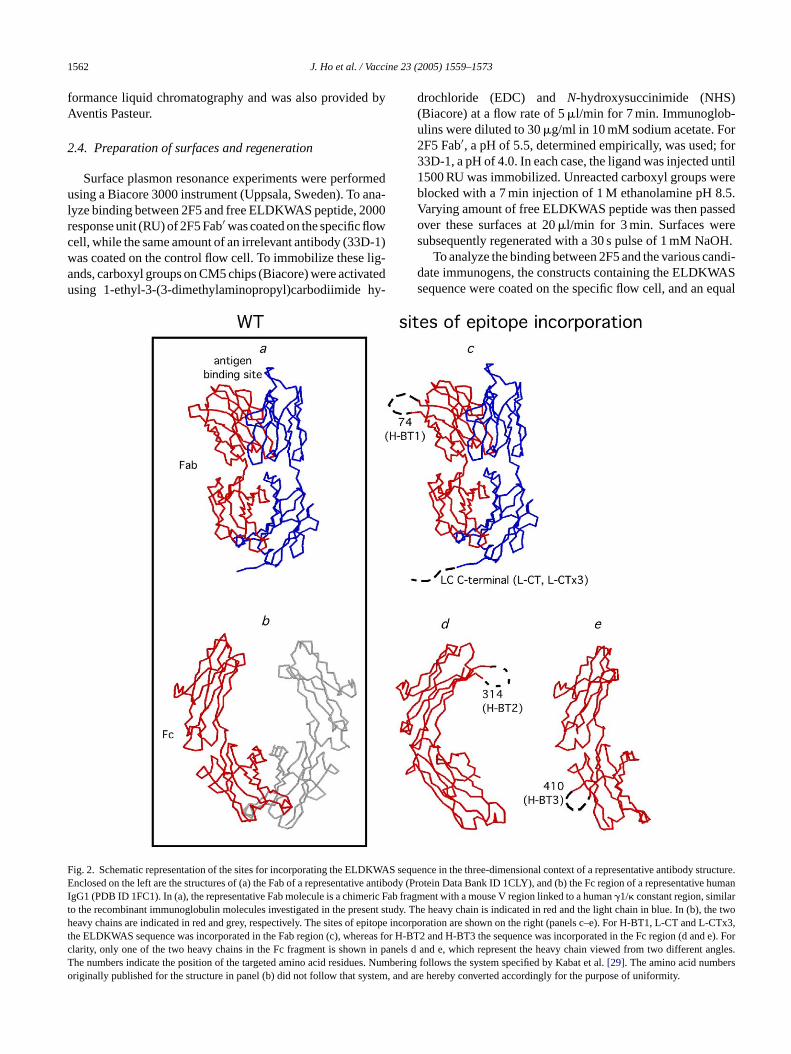
ig. 2. Schematic representation of the sites for incorporating the ELDKWASnclosed on the left are the structures of (a) the Fab of a representative antib
gG1 (PDB ID 1FC1). In (a), the representative Fab molecule is a chimeric Fao the recombinant immunoglobulin molecules investigated in the present stueavy chains are indicated in red and grey, respectively. The sites of epitope
he ELDKWAS sequence was incorporated in the Fab region (c), whereas folarity, only one of the two heavy chains in the Fc fragment is shown in panhe numbers indicate the position of the targeted amino acid residues. Numriginally published for the structure in panel (b) did not follow that system, a
sequence in the three-dimensional context of a representative antibodystructure.ody (Protein Data Bank ID 1CLY), and (b) the Fc region of a representativehumanb fragment with a mouse V region linked to a human�1/� constant region, similardy. The heavy chain is indicated in red and the light chain in blue. In (b),the twoincorporation are shown on the right (panels c–e). For H-BT1, L-CT and L-CTx3,
r H-BT2 and H-BT3 the sequence was incorporated in the Fc region (d and e). Forels d and e, which represent the heavy chain viewed from two different angles.bering follows the system specified by Kabat et al.[29]. The amino acid numbersnd are hereby converted accordingly for the purpose of uniformity.

J. Ho et al. / Vaccine 23 (2005) 1559–1573 1563
amount of WT construct was coated on the control flow cell.The same immobilization procedure was used as above, withthe coating step performed at pH 5.5. The 2F5 Fab′ fragmentwas injected as analyte at varying concentrations at 20�l/minfor 7 min, and the surface was regenerated by a 30 s pulse of1 mM NaOH.
2.5. Data collection and analysis
Surface plasmon data were collected at 1 s intervals andanalyzed using Biaevaluation version 3.0 (Biacore). For bind-ing of peptide to 2F5, the relative response level was aver-aged over a 10 s period at steady state to obtainReq. Thevalues were then plotted against concentration. The theoret-ical response correlating to saturation,Rmax, was estimatedby Quasi-Newton curve fit using MacCurveFit version 1.5.5(Kevin Raner Software, Mt. Waverley, Australia).Kd wasthen determined according to the equation:
Req = CRmax
C + Kd
whereC is the concentration of the peptide, andRmax is thetheoretical response at saturation. To model binding with twodifferent sites, the above model was modified as follow toi
R
w ndR if-f dbd
-s ligandm s tot ratec
K
w on-s riumc equi-l y fit-t he eno delw tion:
R
wt
2.6. Animals and immunization
Class II MHC directed immunogens were prepared as de-scribed above. A purified human antibody of the same iso-type (�1/�) but without class II MHC specificity (catalogue#I-5154, Sigma, St. Louis, MO) was used as a non-targetingnegative control. All immunogens were sterilized by filteringthrough 0.22�m filters. The immunotargeting antibody con-structs were designed ultimately for use in human, but sincethey also cross-reacts with rabbit class II MHC[17], a rabbitmodel was used to evaluate the in vivo immunotargeting po-tential of these immungens. Thus, female New Zealand White(NZW), rabbits 2.5–3 kg each (11–14 weeks old), were pur-chased from Charles River Laboratories (Wilmington, MA).All animals were rested for at least one week after arrivalbefore immunization schedule is started.
Rabbits were immunized with 200�g of the recombinantantibody molecules, boosted 4 weeks later with 200�g ofthe same protein, then once more another 4 weeks later with100�g of the same protein. The immunization schedule isschematically illustrated inFig. 3. Immunogens were pre-pared in 0.5 ml doses and administered subcutaneously. Therabbits were divided into two major immunization groups.In one, immunogens were administered in the priming pro-cedure as an emulsification with equal volume of CompleteFreund’s Adjuvant (CFA). In the other, it was administeredi vens priort indi-c co-au eateda ing.
2
xedw fer( atedc teinG s ofI pHe ndi pH9 usingC cu-l hateb n ster-iGs
2e
an-t Y)
ncorporate twoRmax values:
eq = CRmax 1
C + Kd1+ CRmax 2
C + Kd2
hereRmax1 andKd1 pertain to one set of binding sites, amax2 andKd2 pertain to a second of binding sites of d
erent characteristics. The twoRmax values were estimatey Quasi-Newton curve fit as above.Kd1 andKd2 were thenerived from above.
For binding of 2F5 Fab′ fragment to immobilized contructs, data were analyzed using the heterogeneousodel in the Biaevaluation software (where ligand refer
he molecules bound to the CM5 chip) to derive the ononstant (kon) and the off rate constant (koff ). The equation
d = koff
kon
herekon and koff represent the on rate and off rate ctants, respectively, was then used to derive the equilibonstants of dissociation. As an alternative method, theibrium constants of dissociation were also estimated bing the quasi-steady state response levels reached at tf the injection phase to a simple Langmuir binding moith one class of binding sites, as described by the equa
eq = CRmax
C + Kd
hereC is the concentration of the 2F5 Fab′, andRmax is theheoretical response at saturation.
d
n saline. Within each group, rabbits were divided into seubgroups, with three animals in each. Blood was drawno immunization, before each boost, and at week 12 asated inFig. 3. Collected blood samples were allowed togulate at 4◦C. Sera were then collected and stored at−70◦Cntil assayed. For neutralization assays, the sera were ht 56◦C for 30 min to inactivate complements prior to test
.7. Purification of IgG from rabbit sera
To purify total rabbit IgG from sera, samples were miith equal volume of ImmunoPure (G) Binding Buf
Pierce). The mixture was then loaded onto equilibrolumns containing ImmunoPure Plus Immobilized Pro
(Pierce). Columns were then washed with 25 volumemmunoPure (G) Binding Buffer, and eluted with a lowlution buffer (50 mM glycine, 150 mM NaCl, pH 2.8), a
mmediately neutralized with 1/10 volume of 1 M Tris,.5. Purified antibody samples were then concentratedentricon-Plus 20 concentrator (Millipore) with a mole
ar weight cut-off 10 000 Da, and exchanged into phospuffered saline. The concentrated preparations were the
lized by filtering through 0.22�m low protein binding MillexV syringe filters (Millipore), and stored at 4◦C until as-ayed.
.8. Determination of immunogen-specific antibodynd-point titres
To determine end-point titres of immunogen-specificibody in sera, Costar ELISA plates (Corning, Corning, N

1564 J. Ho et al. / Vaccine 23 (2005) 1559–1573
Fig. 3. Immunization groups and schedule. (a) 14 Groups of rabbits, three rabbits per group, were divided into two arms. In one arm, animals were primedwith immunogen in Complete Freund’s Adjuvant (CFA), and subsequently boosted twice with the same immunogen in saline. In the other arm, animals wereprimed and boosted with the same immunogens in saline. Within each arm, animals were immunized either with an immunotargeting immunogen containingconformationally constrained ELDKWAS sequence (H-BT1, 2, and 3 groups), an immunotargeting immunogen containing unconstrained ELDKWAS sequence(L-CT and L-CTx3 groups), an immunotargeting immunogen with no ELDKWAS sequence (WT group), or a non-targeting isotype control immunoglobulin(irrelevant human IgG/� group). (b) All animals were primed with 0.2 mg of the protein respective immunogen, then boosted 4 weeks later with 0.2 mg of thesame immunogen, then boosted once more another 4 weeks later with 0.1 mg of the same immunogen. (Abbreviations:Irr hIgG/� = irrelevant human IgG/�,MHC = Major Histocompatibility Complex, CFA = Complete Freund’s Adjuvant).
were coated with 100�l of the respective immunogen at1�g/ml at 4◦C overnight. Plates were then blocked with0.5% BSA in PBS with 0.05% Tween-20 and 1% nor-mal goat serum for 1 h at 37◦C. After washing the platesthree times with PBS with 0.05% Tween-20, 100�l rabbitsera, 4-fold serially diluted in blocking buffer, were dis-pensed into the wells. After 1 h of incubation at 37◦C,the plates were washed and the captured rabbit antibod-ies were detected by alkaline phosphatase conjugated goatanti-rabbit IgG (catalogue #L42008, Caltag Laboratories,Burlingame, CA). After washing the wells three times,the ELISA was developed usingp-nitrophenylphosphate
(Sigma) at 1 mg/ml in 10% diethanolamine, pH 9.8.Absorbance at 405 nm was determined using THERMO-max microplate reader (Molecular Devices, Sunnyvale, CA).Samples were tested in duplicates, and the values wereaveraged. End-point titres were defined as the highest di-lution that gives an absorbance reading above twice the av-eraged absorbance of four wells assayed without serum onthe same plate. For determination of gp160 cross-reactiveantibody titres, the same procedure was followed as above,except that the plates were coated with oligomeric HIV-1 gp160 (catalogue #2000LAV, Protein Sciences, Meriden,CT).

J. Ho et al. / Vaccine 23 (2005) 1559–1573 1565
2.9. Neutralization assays
The neutralization assay, using R2 pseudotyped virus forsingle round infection, has been described elsewhere[18].Briefly, virus competent for a single round of replicationwas produced by transfecting 293 cells with two plasmids:one containing theenv-deleted molecular clone of NL4-3(pNL4-3.luc.R-E-), and another containing theenvgene fromR2 isolate. The plasmid pNL4-3.luc.R-E- contains a reporterfirefly luciferase gene inserted into thenefgene, and is ren-deredenv− andvpr− by two separate frameshifts. Since theenvgene is supplied in trans, the pseudovirions carrying theenv-deleted genome can effect only one round of replication.The incorporatedluc gene leads to expression of luciferaseupon infection. For target cells, cell line U87.CD4.CCR5,which expresses CD4 and CCR5 co-receptor, was used.2× 104 target cells were pre-incubated in 100�l of medium(DMEM containing 15% FBS, 1�g of puromycin/ml, 300�gof G418/ml, glutamine, and penicillin–streptomycin) in eachmicrowell for 24 h at 37◦C in 5% CO2 100�l of mediumcontaining an amount of virus, previously determined to yield∼100 000 luciferase-producing infectious units, was mixedwith various amounts of sera or purified IgG samples, incu-bated for 1 h at 37◦C, added to the target cells, and incubatedfor a further 3 days. The wells were aspirated and washed oncew nt( dt1 quea ea-s gent(
3
3i
werea eci-fi le.T oss-rs ew s( ana on-s demr n oft var-i thep eins,h pt ivei s ex-
pressing the respective heavy and light chains for each con-struct. The secreted recombinant immunoglobulin moleculeswere then purified by affinity chromatography. The purity ofthe protein samples was confirmed by SDS–PAGE analysis,which showed only the bands corresponding to the molecularmass of the heavy and the light chains of an immunoglobulin(Fig. 4).
3.2. Binding of the 2F5 Fab′ fragment to free ELDKWASpeptide
In order to gain a better understanding of how theconformation of the ELDKWAS sequence may affect thebinding of the sequence to 2F5, a series of surface plasmonresonance (SPR) experiments were carried out. In the firstset of experiment, an attempt was made to assess the affinityof free ELDKWAS peptide to the 2F5 antibody. Towardsthat end, the Fab′ fragment of the 2F5 antibody was immo-bilized on a CM5 chip, and a similar mass of an irrelevantantibody was immobilized on the surface of the controlflow cell. Free ELDKWAS peptide at various concentrationswas injected as analyte. Using the Langmuir model, theequilibrium constant of dissociation (Kd) of ELDKWASpeptide binding to the 2F5 Fab′ fragment was found to be1.62�M.
3cu
nt n-s tivet T,f ea-s tructs
F s andp rpo-r e usedt ructss roteinG lamideS underu
ith PBS, and 60�l of luciferase cell culture lysis reagePromega, Madison, WI) was added. 50�l were transferreo a round-bottom plate and centrifuged at 1800×g for0 min. Twenty microlitres were transferred to an opassay plate (Corning), and the luciferase activity was mured on a luminometer using luciferase assay reaPromega).
. Results
.1. Expression and purification of the candidatemmunogens
The series of candidate immunogens being studiedll recombinant immunoglobulin molecules bearing spcity for a framework determinant on the HLA-DR molecuhe specificity of these antibodies is also known to creact with the class II MHC molecules in rabbit[17]. Aschematically illustrated inFig. 2, the ELDKWAS sequencas incorporated as an insertion at�-turn constraining site
as in the constructs H-BT1, H-BT2, and H-BT3), or asttachment to the C-terminus of the light chain (as in ctructs L-CT and L-CTx3, the latter bearing three tanepeats of the ELDKWAS sequence). The constructiohe genes encoding the heavy and the light chains of theous recombinant immunoglobulin molecules, as well asreliminary characterization of these recombinant protave been previous documented[12]. In an effort to scale u
he production of these constructs for purification, High Fnsect cells were infected with recombinant baculoviruse
.3. ELDKWAS sequence constrained inβ-turnonformations have higher affinity towards 2F5 thannconstrained counterpart
To determine whether the�-turn constraints imposed ohe ELDKWAS sequence in the H-BT1, H-BT2, H-BT3 cotructs had resulted in differences in 2F5 binding relao the minimally constrained ELDKWAS sequence in L-Curther SPR experiments were carried out. In order to mure monovalent interactions between the various cons
ig. 4. Candidate immunogens were expressed from High Five cellurified by affinity chromatography. Recombinant baculoviruses incoating the respective heavy chain and light chain encoding genes wero infect High Five insect cells. Recombinant immunoglobulin constecreted into the supernatants were then purified using Protein A or P. Eluates were subsequently concentrated and analyzed by 10% acryDS–PAGE. The gel was stained with SYPRO Orange and visualizedltraviolet light.
1566 J. Ho et al. / Vaccine 23 (2005) 1559–1573
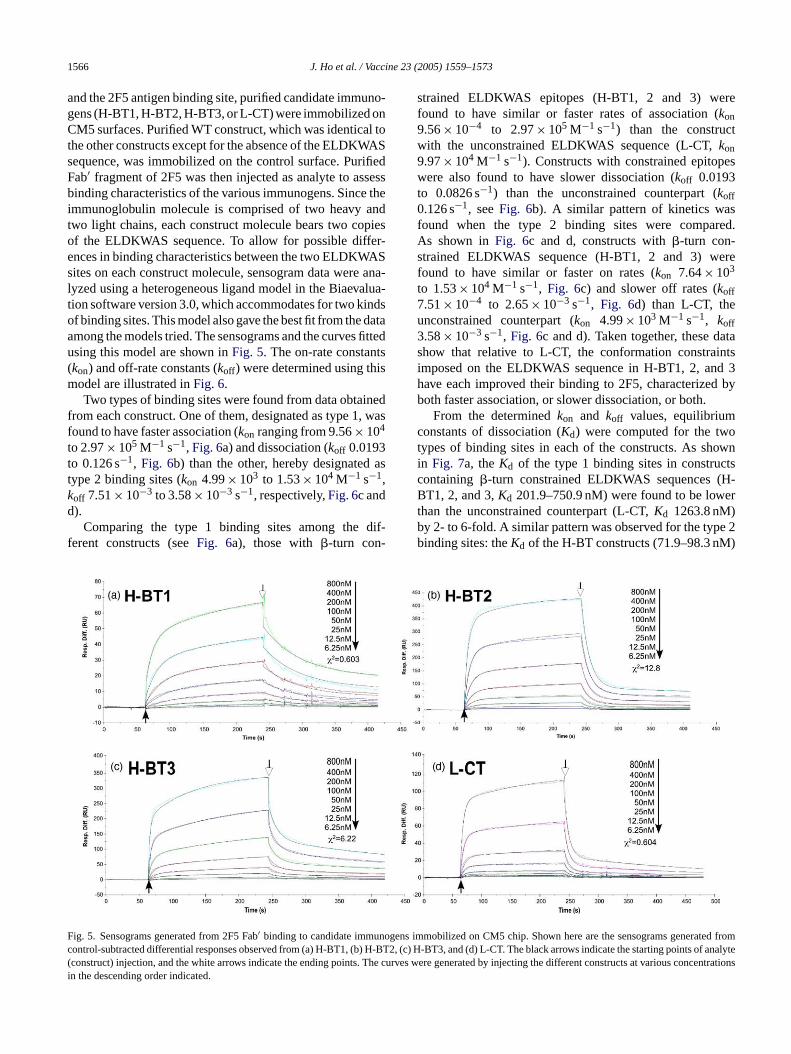
and the 2F5 antigen binding site, purified candidate immuno-gens (H-BT1, H-BT2, H-BT3, or L-CT) were immobilized onCM5 surfaces. Purified WT construct, which was identical tothe other constructs except for the absence of the ELDKWASsequence, was immobilized on the control surface. PurifiedFab′ fragment of 2F5 was then injected as analyte to assessbinding characteristics of the various immunogens. Since theimmunoglobulin molecule is comprised of two heavy andtwo light chains, each construct molecule bears two copiesof the ELDKWAS sequence. To allow for possible differ-ences in binding characteristics between the two ELDKWASsites on each construct molecule, sensogram data were ana-lyzed using a heterogeneous ligand model in the Biaevalua-tion software version 3.0, which accommodates for two kindsof binding sites. This model also gave the best fit from the dataamong the models tried. The sensograms and the curves fittedusing this model are shown inFig. 5. The on-rate constants(kon) and off-rate constants (koff ) were determined using thismodel are illustrated inFig. 6.
Two types of binding sites were found from data obtainedfrom each construct. One of them, designated as type 1, wasfound to have faster association (kon ranging from 9.56× 104
to 2.97× 105 M−1 s−1, Fig. 6a) and dissociation (koff 0.0193to 0.126 s−1, Fig. 6b) than the other, hereby designated astype 2 binding sites (kon 4.99× 103 to 1.53× 104 M−1 s−1,k −3 −3 −1
ddif-
f
strained ELDKWAS epitopes (H-BT1, 2 and 3) werefound to have similar or faster rates of association (kon9.56× 10−4 to 2.97× 105 M−1 s−1) than the constructwith the unconstrained ELDKWAS sequence (L-CT,kon9.97× 104 M−1 s−1). Constructs with constrained epitopeswere also found to have slower dissociation (koff 0.0193to 0.0826 s−1) than the unconstrained counterpart (koff0.126 s−1, seeFig. 6b). A similar pattern of kinetics wasfound when the type 2 binding sites were compared.As shown inFig. 6c and d, constructs with�-turn con-strained ELDKWAS sequence (H-BT1, 2 and 3) werefound to have similar or faster on rates (kon 7.64× 103
to 1.53× 104 M−1 s−1, Fig. 6c) and slower off rates (koff7.51× 10−4 to 2.65× 10−3 s−1, Fig. 6d) than L-CT, theunconstrained counterpart (kon 4.99× 103 M−1 s−1, koff3.58× 10−3 s−1, Fig. 6c and d). Taken together, these datashow that relative to L-CT, the conformation constraintsimposed on the ELDKWAS sequence in H-BT1, 2, and 3have each improved their binding to 2F5, characterized byboth faster association, or slower dissociation, or both.
From the determinedkon and koff values, equilibriumconstants of dissociation (Kd) were computed for the twotypes of binding sites in each of the constructs. As shownin Fig. 7a, theKd of the type 1 binding sites in constructscontaining �-turn constrained ELDKWAS sequences (H-B ertb e 2b )
F nogens ted fromc H-BT2(i
off 7.51× 10 to 3.58× 10 s , respectively,Fig. 6c and).
Comparing the type 1 binding sites among theerent constructs (seeFig. 6a), those with�-turn con-
ig. 5. Sensograms generated from 2F5 Fab′ binding to candidate immuontrol-subtracted differential responses observed from (a) H-BT1, (b)
construct) injection, and the white arrows indicate the ending points. The cun the descending order indicated.T1, 2, and 3,Kd 201.9–750.9 nM) were found to be lowhan the unconstrained counterpart (L-CT,Kd 1263.8 nM)y 2- to 6-fold. A similar pattern was observed for the typinding sites: theKd of the H-BT constructs (71.9–98.3 nM
immobilized on CM5 chip. Shown here are the sensograms genera, (c) H-BT3, and (d) L-CT. The black arrows indicate the starting points of analyte
rves were generated by injecting the different constructs at various concentrations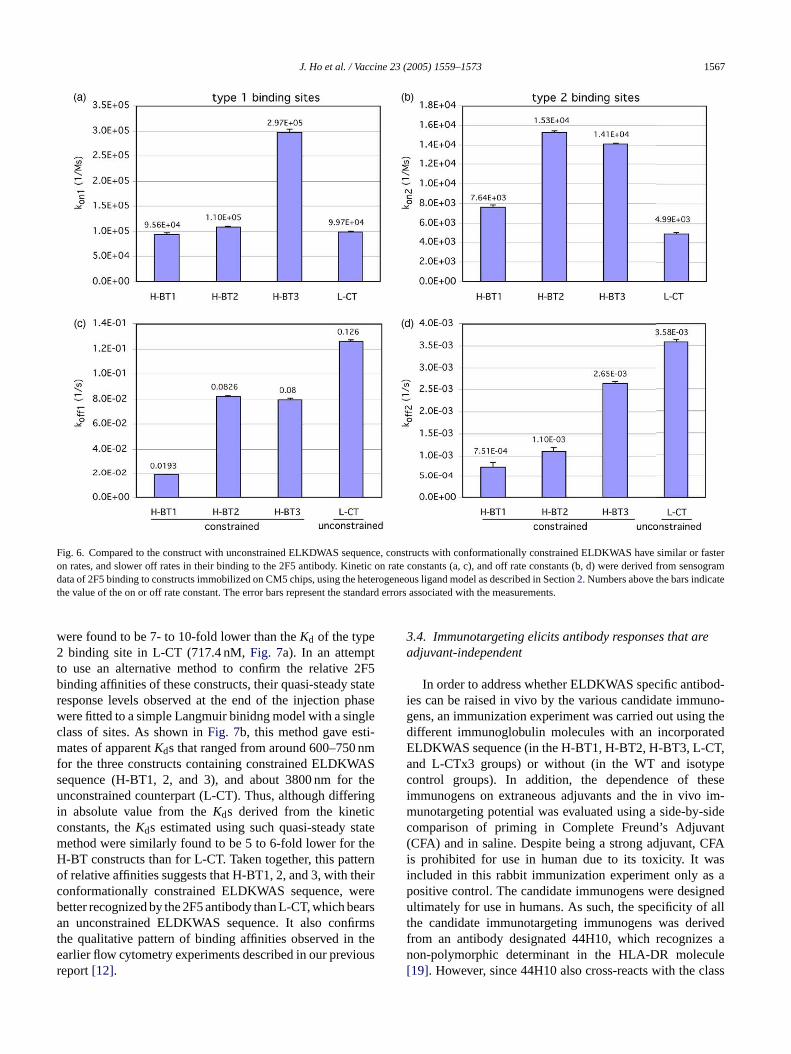
J. Ho et al. / Vaccine 23 (2005) 1559–1573 1567
Fig. 6. Compared to the construct with unconstrained ELKDWAS sequence, constructs with conformationally constrained ELDKWAS have similar or fasteron rates, and slower off rates in their binding to the 2F5 antibody. Kinetic on rate constants (a, c), and off rate constants (b, d) were derived from sensogramdata of 2F5 binding to constructs immobilized on CM5 chips, using the heterogeneous ligand model as described in Section2. Numbers above the bars indicatethe value of the on or off rate constant. The error bars represent the standard errors associated with the measurements.
were found to be 7- to 10-fold lower than theKd of the type2 binding site in L-CT (717.4 nM,Fig. 7a). In an attemptto use an alternative method to confirm the relative 2F5binding affinities of these constructs, their quasi-steady stateresponse levels observed at the end of the injection phasewere fitted to a simple Langmuir binidng model with a singleclass of sites. As shown inFig. 7b, this method gave esti-mates of apparentKds that ranged from around 600–750 nmfor the three constructs containing constrained ELDKWASsequence (H-BT1, 2, and 3), and about 3800 nm for theunconstrained counterpart (L-CT). Thus, although differingin absolute value from theKds derived from the kineticconstants, theKds estimated using such quasi-steady statemethod were similarly found to be 5 to 6-fold lower for theH-BT constructs than for L-CT. Taken together, this patternof relative affinities suggests that H-BT1, 2, and 3, with theirconformationally constrained ELDKWAS sequence, werebetter recognized by the 2F5 antibody than L-CT, which bearsan unconstrained ELDKWAS sequence. It also confirmsthe qualitative pattern of binding affinities observed in theearlier flow cytometry experiments described in our previousreport[12].
3.4. Immunotargeting elicits antibody responses that areadjuvant-independent
In order to address whether ELDKWAS specific antibod-ies can be raised in vivo by the various candidate immuno-gens, an immunization experiment was carried out using thedifferent immunoglobulin molecules with an incorporatedELDKWAS sequence (in the H-BT1, H-BT2, H-BT3, L-CT,and L-CTx3 groups) or without (in the WT and isotypecontrol groups). In addition, the dependence of theseimmunogens on extraneous adjuvants and the in vivo im-munotargeting potential was evaluated using a side-by-sidecomparison of priming in Complete Freund’s Adjuvant(CFA) and in saline. Despite being a strong adjuvant, CFAis prohibited for use in human due to its toxicity. It wasincluded in this rabbit immunization experiment only as apositive control. The candidate immunogens were designedultimately for use in humans. As such, the specificity of allthe candidate immunotargeting immunogens was derivedfrom an antibody designated 44H10, which recognizes anon-polymorphic determinant in the HLA-DR molecule[19]. However, since 44H10 also cross-reacts with the class
1568 J. Ho et al. / Vaccine 23 (2005) 1559–1573
Fig. 7. ELDKWAS sequence grafted into�-turns have higher affinity towards 2F5 than unconstrained counterpart. (a) Equilibrium constants of dissociation(Kd) for the two types of binding sites in the recombinant ELDKWAS-containing antibody constructs, estimated using the heterogeneous ligand model.Kdswere derived from the kinetic rates of association and dissociation by the formulaKd = koff /kon. The numbers above the bars indicate the calculatedKd value ofeach type of binding site in the construct. The error bars, representing the estimated standard errors, are included but too small to be visible in thisgraph. (b)Equilibrium constants of dissociation (Kd) for the various constructs estimated by fitting quasi-steady state binding levels into a Langmuir binding model. Theerror bars represent the standard errors of the estimation procedure.
II MHC molecule on rabbit PBMC[17], it was possible toevaluate the immunotargeting potential of these immunogensin this small animal model.
As shown inFig. 8, all rabbits that received immunogenstargeting class II MHC (panels a–f) developed IgG antibodyresponse whether they were primed with immunogens in CFAor saline alone. In contrast, without such specificity, the iso-type control immunogen could induce antibodies only whenit was administered in CFA but not in saline (Fig. 8g). Thesedata suggest that the candidate immunogens were able to in-duce antibody response without extraneous adjuvants, andthat such induction was mediated by the targeting of theseimmunogens to class II MHC molecules.
In terms of the magnitude of antibody response,immunogen-specific antibody titres plateau at about 106 ingroups primed with CFA. In comparison, animals primedwith immunotargeting immunogens in saline developedimmunogen-specific titres that plateaued at the somewhatlower level of about 105 (seeFig. 8).
3.5. Antibodies cross-reactive with intact oligomergp160 protein were raised by ELDKWAS-containingimmunogens
In an attempt to investigate whether the candidate im-munogens could induce any antibodies against the incorpo-r in as as at ld bed S-c u-n d-p andL ngt odiesw tion
groups, although the level of induction was lower (Fig. 9lower panels). Taken together, the findings show that EL-DKWAS specific antibodies can be raised by the variousELDKWAS-containing immunogens, and that the inducedantibodies can recognize the intact gp160 protein.
3.6. In vitro neutralization of HIV-1
To assess the neutralizing capacity of the induced antibod-ies, whole sera taken from animals with the highest gp160cross-reactive titres were tested for neutralization of R2 pseu-dotyped virus in vitro using a method previously described[20]. No significant difference was detected between the
Table 1Results of neutralization assay using R2 pseudotyped virus and total IgGpurified from the antisera at week 12 of animals in selected CFA primedgroupsa
Animalb [Antibody] (�g/ml)
3000 (%) 1500 (%) 750 (%) 330 (%) 190 (%) 95 (%)
A1 122 99 155 154 152 160A13 132 188 112 135 119 139A15 124 165 105 117 125 149A17 160 140 108 107 168 121A18 155 136 151 107 153 120A1 157 144 151 144 136 197
)
2
izedw -c undredp cted inv
15w T(
ated ELDKWAS sequence, rabbit antisera were testederies of ELISA experiments using intact gp160 proteinarget. As expected, gp160 cross-reactive antibodies couetected in antisera of animals immunized with ELDKWAontaining immunogens in CFA, but not in animals immized with the WT control (Fig. 9 upper panels). The enoint titres of gp160 cross-reactive antibodies in H-BT1-CTx3 groups reached 104 after the second boost. Amo
he saline primed animals, gp160 cross-reactive antibere also detected in the BT-1 and L-CTx3 immuniza
[2F5] (�g/ml)25 (%) 12 (%) 6 (%) 3 (%) 1.5 (%) 0.75 (%
F5 (control) 19 37 63 53 59 100a IgG was purified from sera taken at week 12 from animals immunith H-BT1 (A1–A3) or L-CTx3 (A13–A15) with CFA in the priming proedure, and tested for neutralization against R2 pseudotyped virus. Hercent detectable infection was defined as luciferase counts deteirus culture without any antibody.b Animals A1 and A3 were immunized with H-BT1; A13 and Aere immunized with L-CTx3; A17 and A18 were immunized with W
CFA = Complete Freund’s Adjuvant).
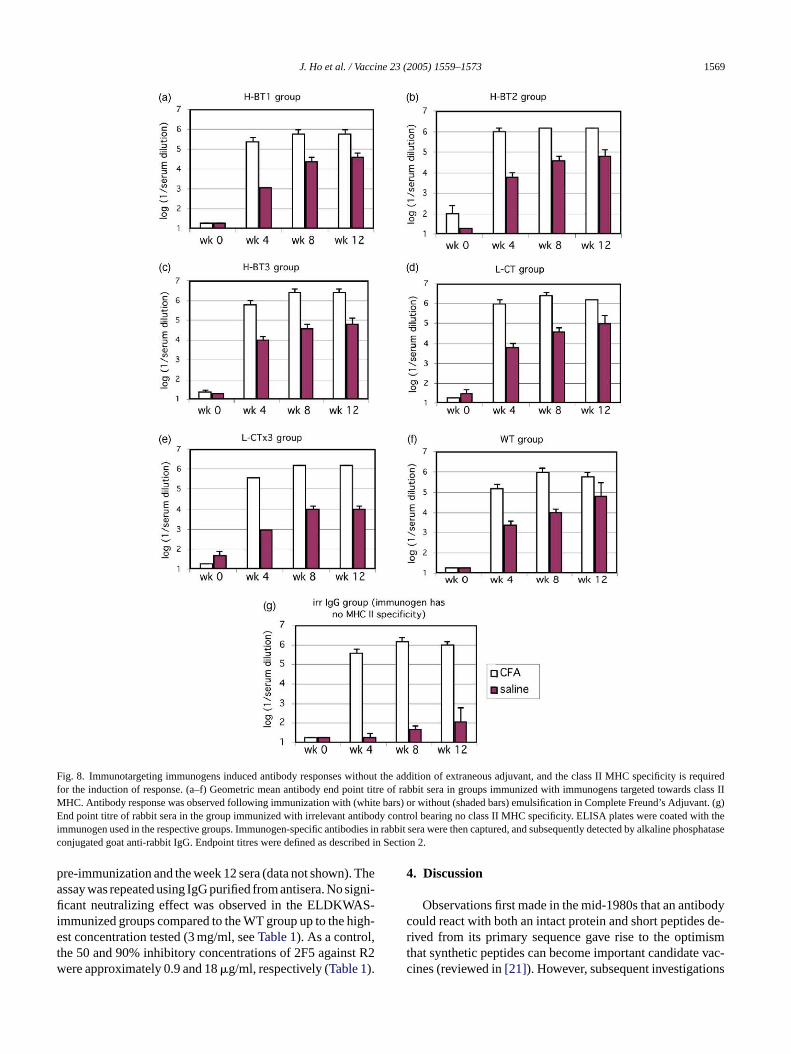
J. Ho et al. / Vaccine 23 (2005) 1559–1573 1569
Fig. 8. Immunotargeting immunogens induced antibody responses without the addition of extraneous adjuvant, and the class II MHC specificity is requiredfor the induction of response. (a–f) Geometric mean antibody end point titre of rabbit sera in groups immunized with immunogens targeted towards class IIMHC. Antibody response was observed following immunization with (white bars) or without (shaded bars) emulsification in Complete Freund’s Adjuvant. (g)End point titre of rabbit sera in the group immunized with irrelevant antibody control bearing no class II MHC specificity. ELISA plates were coated with theimmunogen used in the respective groups. Immunogen-specific antibodies in rabbit sera were then captured, and subsequently detected by alkaline phosphataseconjugated goat anti-rabbit IgG. Endpoint titres were defined as described in Section2.
pre-immunization and the week 12 sera (data not shown). Theassay was repeated using IgG purified from antisera. No signi-ficant neutralizing effect was observed in the ELDKWAS-immunized groups compared to the WT group up to the high-est concentration tested (3 mg/ml, seeTable 1). As a control,the 50 and 90% inhibitory concentrations of 2F5 against R2were approximately 0.9 and 18�g/ml, respectively (Table 1).
4. Discussion
Observations first made in the mid-1980s that an antibodycould react with both an intact protein and short peptides de-rived from its primary sequence gave rise to the optimismthat synthetic peptides can become important candidate vac-cines (reviewed in[21]). However, subsequent investigations
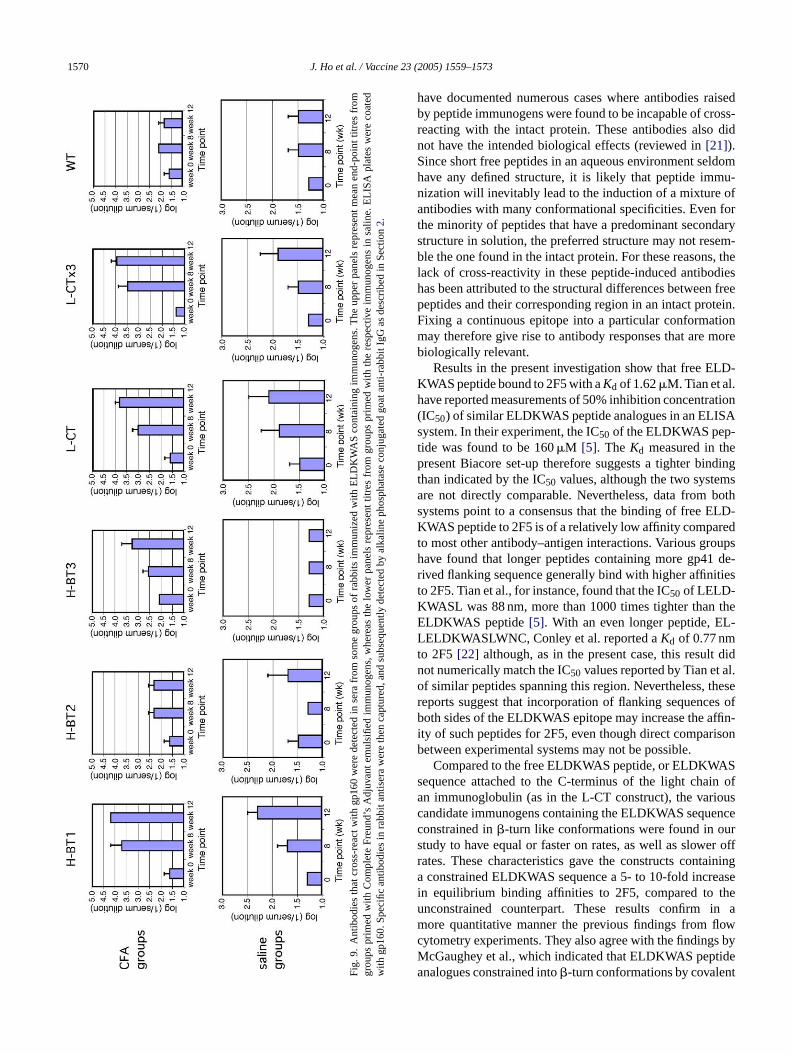
1570 J. Ho et al. / Vaccine 23 (2005) 1559–1573
Fig
.9.
Ant
ibod
ies
that
cros
s-re
actw
ithgp
160
wer
ede
tect
edin
sera
from
som
egr
oups
ofra
bbits
imm
uniz
edw
ithE
LDK
WA
Sco
ntai
ning
imm
unog
ens.
The
uppe
rpa
nels
repr
esen
tmea
nen
d-po
intt
itres
from
grou
pspr
imed
with
Com
plet
eF
reun
d’s
Adj
uvan
tem
ulsi
fied
imm
unog
ens,
whe
reas
the
low
erpa
nels
repr
esen
ttit
res
from
grou
pspr
imed
with
the
resp
ecti
veim
mun
ogen
sin
salin
e.E
LIS
Apl
ates
wer
eco
ated
with
gp16
0.S
peci
fican
tibod
ies
inra
bbit
antis
era
wer
eth
enca
ptur
ed,a
ndsu
bseq
uent
lyde
tect
edby
alka
line
phos
phat
ase
conj
ugat
edgo
atan
ti-ra
bbit
IgG
asde
scrib
edin
Sec
tion2.
have documented numerous cases where antibodies raisedby peptide immunogens were found to be incapable of cross-reacting with the intact protein. These antibodies also didnot have the intended biological effects (reviewed in[21]).Since short free peptides in an aqueous environment seldomhave any defined structure, it is likely that peptide immu-nization will inevitably lead to the induction of a mixture ofantibodies with many conformational specificities. Even forthe minority of peptides that have a predominant secondarystructure in solution, the preferred structure may not resem-ble the one found in the intact protein. For these reasons, thelack of cross-reactivity in these peptide-induced antibodieshas been attributed to the structural differences between freepeptides and their corresponding region in an intact protein.Fixing a continuous epitope into a particular conformationmay therefore give rise to antibody responses that are morebiologically relevant.
Results in the present investigation show that free ELD-KWAS peptide bound to 2F5 with aKd of 1.62�M. Tian et al.have reported measurements of 50% inhibition concentration(IC50) of similar ELDKWAS peptide analogues in an ELISAsystem. In their experiment, the IC50 of the ELDKWAS pep-tide was found to be 160�M [5]. TheKd measured in thepresent Biacore set-up therefore suggests a tighter bindingthan indicated by the IC50 values, although the two systemsa boths LD-K edt oupsh de-r itiestK theE L-Lt t didn al.o theser es ofb ffin-i isonb
Ss in ofa ousc encec urs r offr aininga easei theu in am flowc s byM tidea nt
re not directly comparable. Nevertheless, data fromystems point to a consensus that the binding of free EWAS peptide to 2F5 is of a relatively low affinity compar
o most other antibody–antigen interactions. Various grave found that longer peptides containing more gp41ived flanking sequence generally bind with higher affino 2F5. Tian et al., for instance, found that the IC50 of LELD-WASL was 88 nm, more than 1000 times tighter thanLDKWAS peptide[5]. With an even longer peptide, EELDKWASLWNC, Conley et al. reported aKd of 0.77 nm
o 2F5[22] although, as in the present case, this resulot numerically match the IC50 values reported by Tian etf similar peptides spanning this region. Nevertheless,eports suggest that incorporation of flanking sequencoth sides of the ELDKWAS epitope may increase the a
ty of such peptides for 2F5, even though direct comparetween experimental systems may not be possible.
Compared to the free ELDKWAS peptide, or ELDKWAequence attached to the C-terminus of the light chan immunoglobulin (as in the L-CT construct), the variandidate immunogens containing the ELDKWAS sequonstrained in�-turn like conformations were found in otudy to have equal or faster on rates, as well as sloweates. These characteristics gave the constructs contconstrained ELDKWAS sequence a 5- to 10-fold incr
n equilibrium binding affinities to 2F5, compared tonconstrained counterpart. These results confirmore quantitative manner the previous findings from
ytometry experiments. They also agree with the findingcGaughey et al., which indicated that ELDKWAS pepnalogues constrained into�-turn conformations by covale

J. Ho et al. / Vaccine 23 (2005) 1559–1573 1571
linking of the second and the sixth residues were foundto have higher affinities to 2F5 than the non-constrainedpeptide[11]. Collectively, these findings indicate that theantigenicity of ELDKWAS recognized by 2F5 can be sig-nificantly increased by constraining the sequence in�-turnlike sites in the scaffold of an immunoglobulin molecule.
The collective data generated from the previous in-vestigation[12] and the present one also suggest that the�-turn-constrained ELDKWAS sequence in these constructsrepresents a better mimic of the native epitope of 2F5 thanthe unconstrained sequence or the free peptide. Such a modelagrees with the X-ray crystallography study carried out byPai et al., where the ELDKWAS peptide was found to adopta�-turn conformation when bound to the 2F5 antibody[23].It also agrees with the data obtained from a recent NMRstudy carried out by Barbato et al., where the majority of theconformers of the ELLELDKWASLWN peptide in solutionwas found to adopt a� turn in the DKW region, which con-stitutes the most crucial residues in the interaction between2F5 and the ELDKWAS peptide[24]. The authors alsofound that the shorter ELDKWAS peptide was too flexiblein solution to have any detectable preferred conformation[24], which could explain why immunogens containing con-formationally unconstrained ELDKWAS sequence have allfailed to induce 2F5-like neutralizing antibodies in vivo. Thee al inc o sta-b dp edf sents
2F5e dings g isa roteini acem res,d n tob on ofe wedi re-s resb Ther theirc cea ne-o odyr ificityb gsf vanti milarc ,i eref dies.I thouta score
the usefulness of the antibody scaffold as a viable platformfor conformationally constrained epitope vaccine designs.
Unfortunately, despite the presence of gp160 cross-reactive antibodies, no dramatic neutralizing activity was de-tected in the antisera from the highest responding animals, orin the IgG purified from these antisera. A number of reasonsmay explain the inability of the candidate vaccines to inducedetectable neutralizing antibodies. One possibility is that thestructure of the ELDKWAS sequence in the immunogen maystill be inappropriate for the induction of neutralizing clonesof antibodies. Alternatively, neutralizing clones might havebeen induced, but that the lack of observed neutralizationwas a result of insufficient concentration of such antibodies,and/or the insufficient sensitivity in the in vitro assay systemused. A third possibility is that broadly neutralizing antibod-ies against HIV may require certain unusual characteristics.Of the broadly neutralizing antibodies against HIV, at leasttwo – 2F5[25], and b12[26] – have unusually long CDR3regions in their heavy chain. The 2G12 antibody furthermorehas been found to have a highly unusual heavy chain–heavychain association that gives rise to two secondary antigenbinding sites, in addition to the two usual primary sites formedby heavy chain–light chain association. This unorthodox con-figuration results in a tetravalent molecule[27]. It is possiblethat the ELDKWAS-recognizing antibodies generated by thec auset gion,w lizingm d es-s onala f thei nes.
n ofw ss int thea ypeo nt culars butp ues,m
blen od-i theyw lowt dis-c neu-t than1 of3 cedE no-c er of1 zinge liza-t rtiono uno-
pitope of the antibody may be more loop-like than heliconformation. Indeed, it was postulated that strategies tilize that region into a� turn-like conformation would yielromising vaccine candidates[24], the same concept deriv
rom our earlier investigation and tested further in the pretudy.
Aside from the structural issues surrounding thepitope, it is of equal importance that the chosen 2F5-bintructure be made immunogenic in vivo. Immunotargetinstrategy intended to augment antibody responses to p
mmunogens by directing them to particular cell surfarkers. Among a number of possible target structuirecting immunogens to class II MHC has been showe able to induce antibody responses without the additixtraneous adjuvants that are normally required (revie
n [13]). Thus, the possibility of augmenting antibodyponses to the identified 2F5-binding ELDKWAS structuy means of immunotargeting was also investigated.esults indicate that the candidate immunogens, withlass II MHC targeting specificity, were able to induntibody responses with or without the addition of extraus adjuvant. In the latter case, the induction of antibesponses was dependent on the class II MHC specuilt into the immunogens. This further confirms findinrom numerous previous reports that showed adjundependent induction of antibody responses using a silass II MHC targeting strategy[13]. More importantly
mmunogens containing the ELDKWAS sequence wound to be able to induce gp160 cross-reactive antibon some cases, such antibodies were elicited even widdition of extraneous adjuvant. These results under
andidate immunogens have failed to neutralize HIV bechey do not possess the unusual length in the CDR3 rehich seems to be characteristic of these broadly neutraonoclonal antibodies. If such characteristics are indee
ential for the neutralizing capacity of these rare monoclntibodies, the lack of neutralization could be a result o
nability of the candidate immunogens to induce such cloThe unusually long CDR3 of 2F5 raises the questio
hether it, like its b12 counterpart, is probing into a recehe envelope glycoprotein. In this regard, elucidation oftomic structure of 3H6, the antibody specific for the idiotf 2F5 [28], will certainly offer very useful information o
his issue. If that is indeed the case, a more complex moletructure, involving not only the ELDKWAS sequence,ossibly also discontinuous regions of amino acid residay be more suited for the recapitulation of its epitope.Another possibility that can explain the lack of detecta
eutralization in most of the antisera is that the antibes required for neutralization were induced, but thatere simply present at concentrations that were too
o be detected in the in vitro neutralization assays. Asussed earlier, the assays used could only detect 2F5ralization when it was present at concentrations higher�g/ml. Thus, by testing total IgG up to a concentrationmg/ml, neutralization could only be detected if the induLDKWAS-specific antibodies equivalent to the 2F5 molonal were present in frequencies higher than the ord:1000 in the serum. Otherwise, their potential neutraliffects would not be detectable in such a virus neutra
ion assay. The data also showed that a significant pof the immune response induced by the candidate imm

1572 J. Ho et al. / Vaccine 23 (2005) 1559–1573
gens in the rabbit model was directed against the human im-munoglobulin backbone. One might be able to increase theproportion of ELDKWAS specific response by exchangingthe human constant region in these constructs with a rabbitconstant region, resulting in a series of “rabbitized” immuno-gens that would further focus the response onto the ELD-KWAS sequence. Alternatively, since the immunogens beingtested were humanized in the constant region with the in-tention of being used as human immunogens, the backbonewould be seen as self by the immune system in human recip-ients. The diminished anti-backbone response in human mayrender the incorporated ELDKWAS epitope much more im-munogenic in relative terms than is possible in the presentrabbit model. Thus, problems associated with insufficientELDKWAS-specific antibody concentrations may be natu-rally overcome in the first human studies. Furthermore, thisELDKWAS-specific response could potentially be enhancedin human clinical studies by the addition of a licensed adju-vant like alum, analogous to the enhanced immune responsesobserved in the present rabbit experiments when CFA wasused.
Although neutralizing antibodies could not be detectedin the antisera from most of the animals in the presentstudy, the results nevertheless establish novel sites in theimmunoglobulin molecule where foreign sequences couldb thef hatB allyi fold,a theseB ousa udiess d byt theu gy toi
roms or ana hasr -m zinga ieso itsi hed ingv h av thec c.
R
de-
ac-ic T
[3] Ferrantelli F, Ruprecht RM. Neutralizing antibodies againstHIV—back in the major leagues? Curr Opin Immunol 2002;14(4):495–502.
[4] Muster T, Steindl F, Purtscher M, et al. A conserved neutralizingepitope on gp41 of human immunodeficiency virus type 1. J Virol1993;67(11):6642–7.
[5] Tian Y, Ramesh CV, Ma X, et al. Structure-affinity relationships inthe gp41 ELDKWA epitope for the HIV-1 neutralizing monoclonalantibody 2F5: effects of side-chain and backbone modifications andconformational constraints. J Pept Res 2002;59(6):264–76.
[6] Parker CE, Deterding LJ, Hager-Braun C, et al. Fine definition ofthe epitope on the gp41 glycoprotein of human immunodeficiencyvirus type 1 for the neutralizing monoclonal antibody 2F5. J Virol2001;75(22):10906–11.
[7] Eckhart L, Raffelsberger W, Ferko B, et al. Immunogenic presenta-tion of a conserved gp41 epitope of human immunodeficiency virustype 1 on recombinant surface antigen of hepatitis B virus. J GenVirol 1996;77(Pt 9):2001–8.
[8] Liang X, Munshi S, Shendure J, et al. Epitope insertion into variableloops of HIV-1 gp120 as a potential means to improve immunogenic-ity of viral envelope protein. Vaccine 1999;17(22):2862–72.
[9] Coeffier E, Clement JM, Cussac V, et al. Antigenicity and immuno-genicity of the HIV-1 gp41 epitope ELDKWA inserted into permis-sive sites of the MalE protein. Vaccine 2000;19(7–8):684–93.
[10] Joyce JG, Hurni WM, Bogusky MJ, et al. Enhancement of alpha-helicity in the HIV-1 inhibitory peptide DP178 leads to an increasedaffinity for human monoclonal antibody 2F5 but does not elicit neu-tralizing responses in vitro, implications for vaccine design. J BiolChem 2002;277(48):45811–20.
[11] McGaughey GB, Citron M, Danzeisen RC, et al. HIV-1 vaccine de-nding3.
[ bi-zingcine
[ ndent
[ hu-iency.
[ manllo-Fab
[ on-cad
[ oss-ules
[ ofock-98–
[ rfacesing
[ nti-V1,
f theirol
[ etic
[ di-mary
e incorporated without affecting the expression orunctional folding of the molecule. They also illustrate t
cell epitopes can be antigenically and immunogenicncorporated at these sites in the immunoglobulin scafnd that antibody response can be elicited against
cell epitopes even without the addition of extranedjuvants. These results confirm the results of earlier sthowing that antibody responses can be augmentehe immunotargeting approach, and further indicatesefulness of immunotargeting as a vaccination strate
nduce responses against protein immunogens.With the emergence of the encouraging results f
everal recent passive transfer studies, the search fntibody-mediated protective vaccine against HIV-1eceived renewed enthusiasm[3]. At the moment, inforation on the structures capable of inducing neutralintibodies reliably is still quite limited. Further studn how the epitopic structure of the 2F5 antibody and
mmunogenicity can be improved will be crucial for tevelopment of an effective, neutralizing antibody-inducaccine against HIV-1. The ultimate availability of sucaccine will certainly be a significant contribution toontrol of the currently unchecked global AIDS epidemi
eferences
[1] Barouch DH, Letvin NL. Viral evolution and challenges in thevelopment of HIV vaccines. Vaccine 2002;20(Suppl. 4):A66–8.
[2] Barouch DH, Kunstman J, Kuroda MJ, et al. Eventual AIDS vcine failure in a rhesus monkey by viral escape from cytotoxlymphocytes. Nature 2002;415(6869):335–9.
velopment: constrained peptide immunogens show improved bito the anti-HIV-1 gp41 MAb. Biochemistry 2003;42(11):3214–2
12] Ho J, MacDonald KS, Barber BH. Construction of recomnant targeting immunogens incorporating an HIV-1 neutraliepitope into sites of differing conformational constraint. Vac2002;20(7–8):1169–80.
13] Barber BH. The immunotargeting approach to adjuvant-indepesubunit vaccine design. Semin Immunol 1997;9(5):293–301.
14] Purtscher M, Trkola A, Gruber G, et al. A broadly neutralizingman monoclonal antibody against gp41 of human immunodeficvirus type 1. AIDS Res Hum Retroviruses 1994;10(12):1651–8
15] Bryson S, Cunningham A, Ho J, et al. Cross-neutralizing humonoclonal anti-HIV-1 antibody 2F5: preparation and crystagraphic analysis of the free and epitope-complexed forms of its′fragment. Protein Pept Lett 2001;8(5):413–8.
16] Nussenzweig MC, Steinman RM, Witmer MD, Gutchinov B. A moclonal antibody specific for mouse dendritic cells. Proc Natl ASci USA 1982;79(1):161–5.
17] Dubiski S, Cinader B, Chou CT, Charpentier L, Letarte M. Crreaction of a monoclonal antibody to human MHC class II molecwith rabbit B cells. Mol Immunol 1988;25(8):713–8.
18] Zwick MB, Wang M, Poignard P, et al. Neutralization synergyhuman immunodeficiency virus type 1 primary isolates by ctails of broadly neutralizing antibodies. J Virol 2001;75(24):121208.
19] Quackenbush EJ, Letarte M. Identification of several cell suproteins of non-T, non-B acute lymphoblastic leukemia by umonoclonal antibodies. J Immunol 1985;134(2):1276–85.
20] Zwick MB, Kelleher R, Jensen R, et al. A novel human abody against human immunodeficiency virus type 1 gp120 isV2, and V3 loop dependent and helps delimit the epitope obroadly neutralizing antibody immunoglobulin G1 b12. J V2003;77(12):6965–78.
21] Van Regenmortel MH. Antigenicity and immunogenicity of synthpeptides. Biologicals 2001;29(3–4):209–13.
22] Conley AJ, Kessler 2nd JA, Boots LJ, et al. Neutralization ofvergent human immunodeficiency virus type 1 variants and pri

J. Ho et al. / Vaccine 23 (2005) 1559–1573 1573
isolates by IAM-41-2F5, an anti-gp41 human monoclonal antibody.Proc Natl Acad Sci USA 1994;91(8):3348–52.
[23] Pai EF, Klein MH, Chong P, Pedyczak A. Fab′-Epitope Complexfrom the HIV1-Cross-Neutralizing Monoclonal Antibody 2F5. WorldIntellectual Property Organization 2000; WO-00/61618.
[24] Barbato G, Bianchi E, Ingallinella P, et al. Structural analysis of theepitope of the anti-HIV antibody 2F5 sheds light into its mechanismof neutralization and HIV fusion. J Mol Biol 2003;330(5):1101–15.
[25] Kunert R, Ruker F, Katinger H. Molecular characterization of fiveneutralizing anti-HIV type 1 antibodies: identification of nonconven-tional D segments in the human monoclonal antibodies 2G12 and2F5. AIDS Res Hum Retroviruses 1998;14(13):1115–28.
[26] Saphire EO, Parren PW, Pantophlet R, et al. Crystal structure ofa neutralizing human IGG against HIV-1: a template for vaccinedesign. Science 2001;293(5532):1155–9.
[27] Calarese DA, Scanlan CN, Zwick MB, et al. Antibody domain ex-change is an immunological solution to carbohydrate cluster recog-nition. Science 2003;300(5628):2065–71.
[28] Kunert RE, Weik R, Ferko B, Stiegler G, Katinger H. Anti-idiotypicantibody Ab2/3H6 mimics the epitope of the neutralizing anti-HIV-1monoclonal antibody 2F5. AIDS 2002;16(4):667–8.
[29] Kabat EA, Wu TT, Perry HM, Gottesman KS, Foeller C. Sequencesof proteins of immunological interest. Bethesda, MD: U.S. Depart-ment of Health and Human Services; 1991.




















