
Conducting polymer based electrochemical sensor for - OSTI ...
Upload
independentCategory
view
0download
0
Functional Interfacing of Rhodospirillum rubrum Chromatophores toa Conducting Support for Capture and Conversion of Solar EnergyJohn W. Harrold, Jr.,† Kamil Woronowicz,‡ Joana L. Lamptey,‡ John Awong,§ James Baird,§
Amir Moshar,¶ Michele Vittadello,§ Paul G. Falkowski,†,⊥ and Robert A. Niederman*,‡
†Department of Chemistry and Chemical Biology, Rutgers, The State University of New Jersey, 610 Taylor Road, Piscataway,New Jersey 08854, United States‡Department of Molecular Biology and Biochemistry, Rutgers, The State University of New Jersey, 604 Allison Road, Piscataway,New Jersey 08854-8082, United States§Energy Nanotechnology and Materials Chemistry Lab, Medgar Evers College of the City University of New York,1638 Bedford Avenue, Brooklyn, New York 11225, United States¶Asylum Research, 6310 Hollister Avenue, Santa Barbara, California 93117, United States⊥Institute for Marine and Coastal Sciences, Rutgers, The State University of New Jersey, 71 Dudley Road, New Brunswick,New Jersey 08901, United States
*S Supporting Information
ABSTRACT: Owing to the considerable current interest in replacing fossil fuelswith solar radiation as a clean, renewable, and secure energy source, light-drivenelectron transport in natural photosynthetic systems offers a valuable blueprint forconversion of sunlight to useful energy forms. In particular, intracytoplasmicmembrane vesicles (chromatophores) from the purple bacterium Rhodospirillumrubrum provide a fully functional and robust photosynthetic apparatus, ideal forbiophysical investigations of energy transduction and incorporation into biohybridphotoelectrochemical devices. These vesicular organelles, which arise by invagina-tion of the cytoplasmic membrane, are the sites of the photochemical reactioncenters and the light harvesting 1 (LH1) complex. The LH1 protein is responsiblefor collecting visible and near-IR radiant energy and funneling these excitationsto the reaction center for conversion into a transmembrane charge separation.Here, we have investigated the morphology, fluorescence kinetics and photocurrentgeneration of chromatophores from Rsp. rubrum deposited directly onto gold surfaces in the absence of chemical surfacemodifications. Atomic force microscopy showed a significant coverage of the gold electrode surface by Rsp. rubrum chro-matophores. By in situ fluorescence induction/relaxation measurements, a high retention of the quantum yield of photochemistrywas demonstrated in the photoactive films. Chronoamperometric measurements showed that the assembled bioelectrodes werecapable of generating sustained photocurrent under white light illumination at 220 mW/cm2 with a maximum current of 1.5 μA/cm2,which slowly declines in about 1 week. This study demonstrates the possibility of photoelectrochemical control of robustchromatophore preparations from Rsp. rubrum that paves the way for future incorporation into functional solar cells.
1. INTRODUCTION
The light reactions of oxygenic photosynthesis are initiated by aphoton-driven charge separation process catalyzed by photo-chemical reaction centers (RCs), resulting in the generation ofchemical bonds in the form of reduced pyridine nucleotides andATP.1 In the anoxygenic chlorophototrophic purple nonsulfurbacteria, such as Rhodospirillum rubrum, light energy isconverted to chemical potential by way of a type-II RC,organized within the intracytoplasmic membrane (ICM) as acore structure completely surrounded by the light harvesting1 (LH1) antenna complex.2,3 In addition to absorbing blue−green light via the carotenoid spirilloxanthin, the LH1 protein iscapable of collecting near-IR photons (maximally at 880 nm)and funneling their excitation energy to the RC bacterio-chlorophyll (BChl) special pair for transduction into a
transmembrane charge separation with a quantum yield nearunity.4 Figure 1 shows how this initiates a cycle of electron-transfer reactions between the primary iron−quinone acceptor(QA), the ubiquinol−cytochrome c2 oxidoreductase (cytochromebc1 complex), cytochrome c2, and the photooxidized RC−BChlspecial pair, resulting in the formation of an electrochemicalproton gradient, coupled to the synthesis of ATP by an F1FO-ATP synthase. Together with reduced organic compounds as anelectron-donor source, this cyclic electron flow process allows the
Special Issue: Rienk van Grondelle Festschrift
Received: February 28, 2013Revised: June 19, 2013Published: June 21, 2013
Article
pubs.acs.org/JPCB
© 2013 American Chemical Society 11249 dx.doi.org/10.1021/jp402108s | J. Phys. Chem. B 2013, 117, 11249−11259

RC to move electrons and undergo rereduction without thenecessity of an oxygen-evolving complex oxidizing water, as inthe PSII complex, which houses the type-II RC of oxygenicphototrophs.1
The biologically mediated electron-transfer events in purplebacteria were demonstrated to be capable of generating a cur-rent in a biohybrid photovoltaic device by Jantzen and Seibert,5
who attached Rhodobacter sphaeroides RCs directly to a SnO2electrode. In the studies of Katz and associates,6,7 a photo-induced charge separation was demonstrated with UQ-depletedRhodobacter sphaeroides RCs immobilized on a PtO electrodesurface modified with covalently bound quinonoid in the pres-ence of reduced cytochrome c as the electron donor.6 Currentgeneration was enhanced when UQ10 was added as adiffusionally mobile mediator of electron flow between theRC surface and the electrode, driven by 860 nm illuminationabsorbed by the RC−BChl special pair. In addition, RCmonolayers were covalently immobilized on a carbonelectrode surface via the activation of a thiol-bindingbifunctional reagent, resulting in oriented RC attachment.7
The limited separation between RC quinone sites and theelectrode surface provided efficient nondiffusional electronflow, eliminating the need for soluble electron-transfermediators, thereby demonstrating the importance of spatialand diffusional considerations in the design of efficientbiohybrid photoelectrochemical devices.Subsequent work has further established the bacterial RC as
an ideal model system for improvements in the design of bio-hybrid photoelectrochemical devices, and a number of RC immo-bilization strategies for enhanced photocurrent generation havebeen developed.8 These have included the orientated binding ofHis-tagged RCs on gold using Ni-nitrilotriacetic (Ni-NTA) self-assembled monolayers.9−11 With the primary donor orientedtoward the substrate, a unidirectional, light-induced cathodicphotocurrent was generated in the presence of UQ10, withelectron transfer between the gold electrode and the RCoccurring by an apparent tunneling mechanism.9 Photocurrentgeneration was improved in the presence of cytochrome cthrough formation of a RC−cytochrome c complex.10 Theefficiency of such biohybrid photoelectrochemical devices wasimproved by encapsulating stabilized RCs inside of electrodesconsisting of single-walled carbon nanotube arrays.12,13 As anadditional strategy for increasing the adsorptive surface area ofsuch biohybrid photoelectrochemical devices,14 RCs from a
thermophilic purple bacterium were adsorbed to folded surfaceof mesoporous silica. The associated RCs were shown to bestable, specifically bound within the silica pores, and capable ofgenerating a photocurrent on an ITO electrode.While RCs have provided a highly useful energy conversion
system when serving as the biologically based component ofbiohybid photoelectrochemical devices, increased solar con-version efficiencies are obtained with RC−LH1 core particle,in which the LH1 antenna complex functions at the interfaceof light harvesting and electron cycling together with providinga markedly increased functional absorption cross section (σ).Nango and collaborators15,16 showed that RC−LH1 core com-plexes from purple bacteria could be stably self-assembled asmonolayers on appropriately modified electrodes surfaces,which, upon illumination at the LH1 absorption maximum(880 nm), gave rise to efficient energy transfer and photo-current responses. More recently, His-tagged RC−LH1 corecomplexes, engineered in Rba. sphaeroides with definedorientations, were immobilized onto a solid Ni-NTA substrateand shown to gave rise to orientation-dependent photo-currents.17 Such orientation-specific photocurrents were alsodemonstrated with Rba. sphaeroides LH proteins in which Cysresidues were inserted by site-directed mutagenesis on eachmembrane face and attached covalently to conducting goldsurfaces.18 LH functions were improved in comparison to self-assembled monolayers adsorbed onto gold, and superior LH1photostability was also confirmed.As a means of enhancing photocurrent generation, RCs were
interfaced directly to a bare gold electrode,19 and the effectsplayed by molecular relays and the inclusion of LH complexeswere investigated. Sizeable photocurrents were obtained incomparison with monolayers that were self-assembled with RCsspecifically modified to contain orienting linkers. This wasapparently a consequence of the lack of a layer insulating themetal surface from the RC pigments and the minimization ofelectron tunneling distances. Higher photocurrents were alsogenerated with RC−LH1 core complexes, but this was notattributed to the improved photoabsorption but rather toan enhanced surface coverage of core complexes that wereuniformly oriented and contained a tetraheme cytochrome,thought to serve as a connecting wire. Added cytochromec functioned as a molecular relay by intercalating betweenthe adhered RC pigments and the electrode to improve cur-rent production. Alterations in the order in which the photo-cell components were introduced resulted in a dynamicrearrangement of the cytochrome and RCs at the electrodesurface.Magis et al.20 have described the fabrication of a chromatophore-
based biohybid photoelectrochemical device in which ICM vesicles(chromatophores) from Rba. sphaeroides were immobilized onto abare gold electrode surface in the absence of surface functionaliza-tion. Confocal fluorescence spectroscopy and light-inducedelectrochemistry measurements demonstrated that the adsorbedmembranes maintain their energy and electron-transferringcapabilities. By applying an external potential within the range ofthe open-circuit potential of the RC and appropriate charge carriers(cytochrome c and the quinone Q0, which lacks an isoprenoid tail),light-induced currents of 10 μA/cm2 were generated for severalminutes under aerobic conditions and maintained for nearly3 days under continuous illumination. These electrochemicalmeasurements showed that a current of 300 nA was generatedat peak output, dropping off to a minimum of 50 nA over thefurther duration of the measurements. The LH1 complex
Figure 1. Schematic depiction of radiant energy transformation in theICM of Rsp. rubrum. Photon absorption and excitation energy transferby the LH1 antenna complex (black undulating arrow) drives RCcharge separation across the ICM and subsequent electron transport(blue arrows), coupled to proton movement (red arrows), whichresults in formation of an electrochemical proton gradient providingthe energy for the synthesis of ATP by an F1FO-ATPase.
The Journal of Physical Chemistry B Article
dx.doi.org/10.1021/jp402108s | J. Phys. Chem. B 2013, 117, 11249−1125911250

showed substantial durability, overcoming the main obstacle forusing biological-membrane materials based in biohybrid photo-electrochemical cells or in other technological applications.In contrast to the robustness demonstrated for the LH1
complex, the Rba. sphaeroides LH2 peripheral antenna proteinshowed a light-intensity-dependent decoupling from photo-conversion, with extensive photobleaching under high illumi-nation.20 Because of this lack of long-term LH2 stability, wehave initiated further photoelectrochemical studies withchromatophores from Rsp. rubrum, which forms a robustLH1−RC core structure while lacking LH2. Further impetusfor the choice of the Rsp. rubrum chromatophore prepara-tions was provided by the differences in the functionalabsorption cross section, obtained from fluorescenceinduction/relaxation measurements between these twoorganisms.21 Values of 16 Å2 were obtained for Rsp. rubrumcells,21 as compared to ∼50 and ∼120 Å2, respectively, forRba. sphaeroides cells grown at high light intensity and thoseacclimating to 48 h of weak illumination in which theLH2/LH1 molar ratio increased by ∼1.8-fold.22 Although asmaller antenna size results in decreases in the amount oflight harvested by individual cells, they have an enhancedactivity for the charge separation process on a photon basis,owing to decreases in nonproductive light acquisition and anattendant loss of captured excitations as heat or fluorescence.As noted above for Rba. sphaeroides chromatophoresadsorbed to gold,20 little advantage was encountered whenthe functional absorption cross section was expanded by thepresence of the LH2 complex.Consequently, we have adsorbed Rsp. rubrum chromato-
phores to gold electrode surfaces lacking functionalization andobtained views of the surface coverage of the immobilizedstructures by atomic force microscopy while assessing theretention of functional activities through fluorescence in-duction/relaxation and chronoamperometric measurements.These functional biohybrid photoelectrochemical cells werecompared to devices assembled using Rba. sphaeroideschromatophores with increasing LH2 levels. The adsorbedstructures showed a continuous surface coverage comprised offlattened Rsp. rubrum chromatophore vesicle patches that had asignificant residual quantum yield of photochemistry and thecapability of generating sustained photocurrents under bothwhite light and intense 850 nm illumination for a 1 weekperiod.
2. EXPERIMENTAL SECTION
2.1. Bacterial Growth and Chromatophore Isolation.Rba. sphaeroides was grown at a 30 °C in Cohen-Bazire medium23
under high light conditions (1100 W/m2) to a maximum OD680of 0.2 to limit cellular self-shading. The cells were switched to alower light level (100 W/m2) and incubated for another 24 h,reaching an OD680 of 0.7. Because the illumination level isrelated inversely to the levels of the peripheral LH2 antenna,24
the cells harvested at 4 and 24 h yielded increased LH2/LH1molar ratios when compared to the 0 h cells (Table 1). Rsp. rubrumwas grown at 30 °C in Ormerod medium,25,26 modified asdescribed in ref 16, at low light conditions (100 W/m2) to anOD680 of ∼1.0.Bacterial cells were harvested by centrifugation at 14 000g
for 10 min at 4 °C, washed at 18 000g in 1 mM Tris buffer(pH 7.5), and resuspended to 1.5 mg of wet weight in bufferby homogenization. The Rba. sphaeroides chromatophores wereisolated as described previously.27 For isolation of chromato-phores from Rsp. rubrum, harvested cells were treated for30 min at 37 °C with lysozyme (1.0 mg/mL), a few crystalsof DNAase I and protease inhibitors (Roche) were added, andthe cells were disrupted by three passages through a Frenchpressure cell, chilled to 4 °C, at 16 000 lb/in.;2 all subsequentprocedures were also performed at 4 °C. Unbroken cells andcellular debris were removed by centrifugation at 18 000g for10 min, and the supernatant was layered onto a 5−40% (w/w)sucrose gradient prepared in Tris buffer over a 60% sucrosecushion. After centrifugation for 3 h at 141 000g in an SW 28rotor, the chromatophore band in the bottom third of thegradient was removed with a Pasteur pipet and placed in anAmicon 100 kDa filter concentrator. During the concentrationprocess, the buffer was exchanged to 20 mM sodium phosphate(pH 7.5).
2.2. Chromatophore Immobilization. For surfacecharacterization, chromatophores were immobilized on a goldsubstrates consisting of 10 × 10 mm silicon chips with a 30 nmchromium adhesion layer covered by a 100 nm layer of gold(supplied by Structure Probes, Inc.). The substrate was preparedby hydrogen flaming to form a uniform and nearly atomically flatsurface. The chromatophores (0.1 mg of bacteriochlorophyll a(BChl)/ml) were incubated on the substrate surface for 1 h in thedark at 0−4 °C.For chronoampereometric measurements, chromatophores
were immobilized by incubation on a polished and rinsed 3 mm
Table 1. Chromatophores in Suspension with Cytochrome ca
sample LH1/LH2 F0 Fm Fv/Fm σRC (Å2/q) p τi (ms)
Rba. sphaeroides 0h 3.25:1.00 566 1715 0.67 86.2 0.10 τ1 =1.15 ± 0.09 α1 = 0.50τ2 = 15.3 ± 1.5 α2 = 0.32τ3 = 242 ± 30. α3 = 0.18
Rba. sphaeroides 4h chromatophores 2.71:1.00 1007 1552 0.35 87.7 0.02 τ1 =1.62 ± 0.26 α1 = 0.25τ2 = 47.2 ± 7.4 α2 = 0.30τ3 = 504 ± 69 α3 = 0.45
Rba. sphaeroides 24h chromatophores 1.00:1.00 1214 1737 0.30 157.4 0.04 τ1 =1.25 ± 0.19 α1 = 0.32τ2 = 34.9 ± 4.4 α2 = 0.39τ3 = 408 ± 75 α3 = 0.29
Rsp. rubrum chromatophores n/a 808 1699 0.52 24.0 0.42 τ1 = 1.11 ± 0.13 α1 = 0.24τ2 = 37.2 ± 3.3 α2 = 0.37τ3 = 280 ± 22 α3 = 0.39
aThe parameters refer to the fluorescence yield profiles shown in Figure 6a. F0 is the minimum of fluorescence, Fm is the maximum, Fv = Fm − F0,Fv/Fm is the quantum yield of photochemistry, σCC is the functional absorption cross section, I is the irradiance, τi refers to the equation f II(t) = ∑1
3
Ai(exp(−t/τi) used in the fitting of the fluorescence in region II, and αi corresponds to the ratio αi = Ai/(∑in Ai).
The Journal of Physical Chemistry B Article
dx.doi.org/10.1021/jp402108s | J. Phys. Chem. B 2013, 117, 11249−1125911251

round gold electrode (Bio-Logic) at the BChl concentrationspecified above for 1 h in the dark at 0−4 °C.2.3. Visible and Near-IR Absorption Spectroscopy.
Absorbance was measured between 450 and 950 nm at aresolution of 1 nm using a Beckman DU-640 spectropho-tometer. For Rba. sphaeroides chromatophores, LH1/LH2molar ratios and BChl concentrations were determined bydecomposition of near-IR absorption spectra.28
2.4. Atomic Force Microscopy. Electrode surfaces withadsorbed chromatophores were visualized using a MFP-3D-BioAFM with a Nikon microscope (Asylum Research, SantaBarbara, CA 93117). The measurements were made using thetapping mode in the liquid after the surfaces were dried with agentle stream of argon. BL-AC40TS cantilevers (Bio LeverMini), 0.25 N/m, were used in the liquid measurements.2.5. Fluorescence Induction/Relaxation (FIRe) Mea-
surements. A Satlantic FIRe system was used for fluorescenceinduction/relaxation measurements in which a quartz cuvetteand an 880 nm emission filter were used for bulk measurementson appropriately diluted chromatophore samples. Surface mea-surements were performed by immobilizing a gold substratebearing the chromatophores on a Delrin insert secured to thecuvette holder. The substrate was oriented at a 45° angle withrespect to the vertical axis of the cuvette holder in order todirectly face the excitation source. A single turnover flash (STF)of 300 μs was used to saturate the RCs of the Rba. sphaeroidesand Rsp. rubrum preparations. The excitation source consistedof a blue LED (450 nm, 30 nm bandwidth), which inRba. sphaeroides excites LH1- and LH2-bound spheroidene andspheroidenone to the S2(1Bu
+) state, from which excitationenergy is transferred to both the QX and QY absorption bandsof the LH1 and LH2 BChls.29 As shown by the excitationspectrum in Figure 2, for Rsp. rubrum, the 450 nm excitation
results in an S2(1Bu+) excited state in spirilloxanthin for
transfer to LH1 BChl with an efficiency of ∼36%. The excita-tion source is controlled by an LED circuit driver capable ofgenerating pulses of 1 μs to 50 ms duration. The fluorescence
emission is detected by a sensitive avalanche photodiodemodule, and the digitized fluorescence kinetic transients ob-tained at 880 nm are processed by computer-assisted analysis,which translates the measured signal into several physiologicalparameters, following the charge separation elicited betweenthe RC−BChl special pair and the primary QA. These param-eters include the functional absorption cross section (σ), thequantum yield of the primary charge separation (Fv/Fm), andthe electron-transfer turnover rates of the RC (τ1, τ2, and τ3),extracted from the multiphasic decay during the fluorescencerelaxation. Fv/Fm and σRC are obtained from the initial rapidinduction phase, which is related to the rate of increase in thefluorescence yield.
2.6. Electrochemical Measurements. Chronoamperom-etry was performed with a Bio-Logic SP-50 and VSPpotentiostat-galvanostats, with EC-Lab version 10 softwaresupplied by the manufacturer. Chromatophores were immobi-lized on a 3 mm working electrode that was placed into acommercially available cell, with a 23 cm coiled platinumcounter electrode (see Figure 3 for a schematic representation).
The cell was filled with 15 mL of a 20 mM sodium phosphatebuffer at pH 7.0 containing 100 mM Q0 and 20 mM equineheart cytochrome c. The system was set to a potential of−0.100 V versus the saturated calomel electrode (SCE).20 Theelectrochemical cell temperature was maintained at 25 °C andallowed to come to equilibrium in the dark for 45 min followedby long-term illumination of the immobilized chromatophoresamples either under white light or with an 850 nm LED lightsource at the light intensities indicted in the text.
3. RESULTS AND DISCUSSION3.1. Absorbance Spectra of Chromatopohore Prep-
arations. The electronic absorbance spectra of the Rba. sphaeroideschromatophore preparations isolated at various stages after atransfer from high (1100 W/m2) to low (100 W/m2) lightintensity (Figure 4) showed a marked increase in LH2 levels
Figure 2. Efficiency of excitation energy transfer from carotenoid to BChlin the Rsp. rubrum LH1 complex. Fractional absorption (1 − T) spectraand fluorescence excitation spectra obtained at 77 K were normalized atthe QY absorption maximum (897 nm). Efficiencies for excitation energytransfer from spirilloxanthin to both the QX and QY bands were ∼35−40% as compared to ∼36% in chromatophores. Spectra were obtained byQinghui Hu as described previously30 using a modified a JohnsonResearch Foundation DBS-3 spectrophotometer and a DN1704 liquidnitrogen cryostat (Oxford Instruments, Osney Mead, U.K.).
Figure 3. Schematic depiction of the chronoamperometry measure-ments. A gold working electrode (WE) with adsorbed chromatophoreson the surface was polarized at −0.1 V versus Ag/AgCl, with 3 MNaCl as the reference electrode (RE), at 25.00 ± 0.01 °C. Illuminationoccurred under white light conditions or at 850 nm. The electrontransfer from the WE to the RC in immediate contact with the surfaceoccurs directly. Indirect electron transfer is possible for RCs furtheraway from the surface via horse heart cytochrome c (Cyt c). QuinoneQ0 serves as a redox mediator toward the platinum counter electrode(CE). ECE indicates the potential difference between the CE and theRE, while EWE is the potential difference between the WE and RE.I indicates the electrical current between the WE and the CE. Themeasurement were conducted in aqueous buffer.
The Journal of Physical Chemistry B Article
dx.doi.org/10.1021/jp402108s | J. Phys. Chem. B 2013, 117, 11249−1125911252

relative to those of the RC−LH1 core structures, con-firming the inverse relation between ambient light intensityand LH2 formation in the ICM.24 By 4 h after the switch tolow light intensity, the dimeric LH2−B850 BChl QY-absorptionband at 850 nm overtook the 875 nm LH1 QY-band, which wasthe major band under the high-light conditions (LH2/LH1molar ratios of 0.31 and 0.37 at 0 and 4 h, respectively),ultimately reaching a ratio of 1.00 at 24 h. This overall 3.2-foldincrease in LH2 levels resulted in the expansion of the functionalabsorption cross section to accommodate growth at thereduced light levels. The resulting supramoleclar arrangement31
assured that excitation energy captured by LH2 was transferredwith high efficiency to LH1, ultimately reaching open RCphototraps. Figure 4 also shows that Rsp. rubrum contains aLH1 complex (absorption maximum at 880 nm) while lackingthe LH2 antenna. The RC, identified by the absorbance peakat ∼804 nm arising from the monomeric RC−BChl, exists in
association with the LH1 antenna to form the photosynthetic unitcore,32 shown below to be capable of remaining viable and robustwhen exposed to high light intensities.
3.2. Atomic Force Microscopy of ChromatophorePreparations Adsorbed to Gold Surfaces. The coverage ofchromatophores on the gold substrate surfaces was assessed byatomic force microscopy (AFM). Figure 5A shows the extent ofcoverage of the gold surface with Rsp. rubrum chromatophores.The diameter of chromatophores associated with the goldsurfaces observed here ranged between ∼68.0 and 73.6 nm,with a height of ∼46 nm. In contrast, AFM topographs ofLH1-enriched Rba. sphaeroides preparations showed a morecontinuous coverage (Figure 5B), consisting of chromatophorevesicles of similar diameter and height of ∼50 nm. Thus, inboth instances, our chromatophores, which represent com-plete photosynthetic units, have successfully adsorbed to a goldelectrode surface that requires only minor conditioning and nochemical modification and have assumed heights consistentwith those obtained with Rba. sphaeroides chromatophoreswhen deposited onto mica surfaces.20 These results demon-strate that contact mode AFM can be successfully applied tothe characterization of ICM vesicles adsorbed to gold surfaces.Our data indicate that these preparations have apparentlyretained their vesicular structure rather than giving rise tosingle-membrane bilayers when adsorbed to gold, as observedby Magis et al.20
Because our results are consistent with the retention of avesicular structure for both Rsp. rubrum and LH1-enrichedRba. sphaeroides chromatophores upon adsorption to gold, it ismore surprising that these preparations, as shown below, haveproven to be sufficient for permitting exogenous quinone andcytochrome c to conduct the flow of electrons from thechromatophores to and from the electrodes. While the con-ditions used here for chromatophore immobilization haveattracted the cytoplasmic surface of the chromatophoremembrane vesicles to the gold surface and are of sufficientstrength to survive multiple washings, they do not appear tohave disrupted vesicles into open bilayers, as observed by Magiset al.20 These authors hypothesized that this disruption arisesfrom strong attractive forces between the gold and the vesicularmembranes, sufficient to overcome native forces holding thevesicles together. Nevertheless, such forces may result in partialmembrane disruption, allowing redox carriers to functionaccording to the scheme shown in Figure 3. This would ulti-mately favor the action of cytochrome c in shuttling electronsfrom the working electrode to the RC. In turn, Q0 can transferelectrons from RC to the counter electrode surface, once alight-driven charge separation has taken place (Figure 3).
3.3. Fluorescence Measurements. The variable fluo-rescence arising from the initial chromatophore suspensionsand the chromatophore-modified electrodes was assessed fromfluorescence kinetics studies. The results (Figures 6 and 7)showed that partially functional RCs have been retained in theadsorbed chromatophore preparations and after immobilizationon gold surfaces (Tables 1−4). These measurements wereconducted in the presence of horse heart cytochrome c alone orcytochrome c and Q0. Cytochrome c is expected to play the roleof native cytochrome c2, while Q0 is a surrogate for nativeubiquinone Q (Figure 1). The fluorescence induction rates forthe initial Rba. sphaeroides chromatophore preparations in thepresence of cytochrome c gave values for the quantum yieldof the primary charge separation (Fv/Fm) of 0.67 for the 0 hsample and respective values of 0.35 and 0.30 for those
Figure 4. Visible and near-IR absorbance spectra of the isolatedchromatophores obtained at room temperature. For the Rba.sphaeroides chromatophores, the spectral crossover corrections andmolar extinction coefficients of Sturgis et al.18 were used to calculatethe molar levels of the LH1 and LH2 complexes in the ICMundergoing remodeling during a down-shift in light intensity. Culturegrowth conditions were 1100 W/cm2 for 24 h, shifted to lower light(100 W/cm2), and chromatophores were isolated at 0, 4, and 24 h.Note the accelerated appearance of increased LH2 levels during low-light growth. The left dotted vertical line shows the QY absorption ofLH2 at 850 nm, while the dotted vertical line to the right shows the QYabsorption of LH1 at 875 nm.14 In Rsp. rubrum, note the red shift ofthe LH1 QY absorption (continuous vertical line) band to 880 nm andthe absence of an LH2 antenna.
The Journal of Physical Chemistry B Article
dx.doi.org/10.1021/jp402108s | J. Phys. Chem. B 2013, 117, 11249−1125911253

obtained at 4 and 24 h after the light intensity downshift(Table 1). These values compare favorably to the value of 0.70obtained previously for the chromatophore preparationcharacterized in ref 21. When both cytochrome c and Q0 arepresent the values of Fv/Fm are 0.66, 0.42, and 0.42 for the Rba.sphaeroides chromatophores harvested after 0, 4, and 24 h,respectively (Table 2). With both redox mediators present, theRsp. rubrum chromatophores have a Fv/Fm value of 0.63. Underthese circumstances, the chromatophores in suspension show,in most cases, a higher activity of primary charge separation.The functional absorption cross section (σ) values were 86.2,
87.7, and 157.4 Å2/q for 0, 4, and 24 h chromatophores,respectively, in the presence of cytochrome c only. The samesamples resulted in the following values of σ: 80.9, 91.1, and
149.5 Å2/q, respectively, with cytochrome c and Q0. (Table 2).Consistent with previous observations,21 the values of σ in-crease in the Rba. sphaeroides chromatophores with the lowestLH1/LH2 ratio, in which a higher packing density of LH2 ringsresults in higher values for the functional absorption crosssection. Rsp. rubrum chromatophores exhibit σ values of24.0 and 21.0 Å2/q with cytochrome c only and cytochromec with Q0, respectively (Tables 1 and 2). A small cross section isconsistent with the exclusive presence of the LH1 complex asthe light-harvesting antenna. The higher values of theconnectivity parameter p (0.42 and 0.11 with cytochrome conly and cytochrome c with Q0, respectively) reflect the closerproximity of the RCs to LH1, the sole LH complex in Rsp.rubrum chromatophores. In the case of Rba. sphaeroides
Figure 5. AFM of chromatophores on gold supports. (A) Tapping mode AFM topography image of Rsp. rubrum chromatophores. Thechromatophores were adsorbed to a Au−Si electrode surface glued to a glass slide with epoxy resin, and measurements were made in a liquidmedium. Chromatophores (5 μL) were injected into 100 μL of 0.02 mM sodium phosphate buffer, and the surface was dried with a gentle stream ofargon. The surface was rinsed with 5 mL of deionized water before applying 1 mL of phosphate buffer, followed by 100 μL of 0.01 M Tris/0.15 MKCl buffer. A 1.0 × 1.0 μm scanned area is shown with the panel at the top representing a magnified cytoplasmic surface view showing RC−LH1core particles with a projecting globular surface of the RC subunit H at the center. Rsp. rubrum chromatophore vesicles appear as ellipsoidalstructures with an average size of ∼68.0−73.6 nm and a height averaging ∼50 nm. Vertical false-color scale bars show the tapping mode AFM heightand amplitude images. This signal is determined by the error in the AFM feedback loop of the cantilever’s amplitude and shows fine changes in thefeatures of the topography. The bottom panel shows the results of an analysis of the particle height distribution. (B) Tapping mode AFM topographyimage of a 1.0 × 1.0 μm scanned area containing LH1-enriched Rba. sphaeroides chromatophores isolated from cells grown at high light intensity(1100 W/cm2). The chromatophores were adsorbed to a Au−Si electrode surface as described in panel (a) for imaging in a liquid medlum. In themiddle and bottom panels, three vesicles are magnified to show a higher-resolution view of the scanned surface. Vertical false-color scale bars as inpanel (a).
The Journal of Physical Chemistry B Article
dx.doi.org/10.1021/jp402108s | J. Phys. Chem. B 2013, 117, 11249−1125911254

chromatophores, the connectivity parameters vary in theintervals of 0.02−0.10 with cytochrome c and 0.04−0.05 withcytochrome c and Q0, consistent with a lack of sigmoidicity inthe fluorescence rise kinetics.The fluorescence transients reported in Figure 6 are
sufficiently resolved to allow a successful fitting of the relaxationprofiles after the 300 μs STF with three exponential terms withboth types of chromatophores (Figure 8, Table S1 (SupportingInformation)). To the best of our knowledge, this is the firsttime that all three τ values for Rba. sphaeroides and Rsp. rubrumchromatophores are reported. The τ1 time constant (1 ms) is
attributed to the double reduction of mobile native ubiquinoneQB (or the substitute Q0) by the fixed ubiquinone QA. The τ2time constant (tens of ms) is ascribed to the reductionof ubiquinone QB (or Q0) entering the pocket from theexternal quinone pool, while τ3 (hundreds of ms) wasattributed to inactive and damaged RC. This assignmentreflects the similarity of the RC to PSII core complexes.33
Clearly, when both cytochrome c and Q0 are present, thechromatophores exhibit a higher degree of functionalintegrity. This observation is reflected by the lower valuesof τ3 when cytochrome c and Q0 are available (Table 2) as
Figure 6. Fluorescence kinetics studies of Rba. sphaeroides and Rsp. rubrum chromatophores in suspension, obtained from fluorescence induction andrelaxation measurements. The STF−τ transients (STF: single turnover flash, phase I; τ: fluorescence relaxation, phase II) are shown; (STF, 300 μs; τ,1451 ms), the latter represents the width of the relaxation region. All measurements are the average of 50 iterations. (A) Performed in the presenceof horse heart cytochrome c. (B) Performed in the presence of cytochrome c and quinone Q0.
Table 2. Chromatophores in Suspension with Cytochrome c and Q0a
sample LH1/LH2 F0 Fm Fv/Fm σRC (Å2/q) p τi (ms)
Rba. sphaeroides 0h chromatophores 3.25:1.00 482 1416 0.66 80.9 0.05 τ1 = 1.17 ± 0.09 α1 = 0.39τ2 = 14.2 ± 0.8 α2 = 0.47τ3 = 247 ± 30. α3 = 0.14
Rba. sphaeroides 4h chromatophores 2.71:1.00 1020 1754 0.42 91.1 0.04 τ1 = 1.51 ± 0.11 α1 = 0.44τ2 = 20.5 ± 2.1 α2 = 0.30τ3 = 268 ± 23 α3 = 0.26
Rba. sphaeroides 24h chromatophores 1.00:1.00 1071 1847 0.42 149.5 0.04 τ1 = 1.11 ± 0.12 α1 = 0.40τ2 = 21.3 ± 2.5 α2 = 0.33τ3 = 259 ± 29. α3 = 0.27
Rsp. rubrum chromatophores n\a 635 1733 0.63 21.0 0.11 τ1 = 1.31 ± 0.09 α1 = 0.48τ2 = 25.9 ± 3.2 α2 = 0.28τ3 = 282 ± 35 α3 = 0.23
aThe parameters refer to the fluorescence yield profiles shown in Figure 6b. F0 is the minimum of fluorescence, Fm is the maximum, Fv = Fm − F0,Fv/Fm is the quantum yield of photochemistry, σCC is the functional absorption cross section, I is the irradiance, τi refers to the equation f II(t) = ∑1
3
Ai(exp(−t/τi) used in the fitting of the fluorescence in region II, and αi corresponds to the ratio αi = Ai/(∑in Ai).
The Journal of Physical Chemistry B Article
dx.doi.org/10.1021/jp402108s | J. Phys. Chem. B 2013, 117, 11249−1125911255

opposed to when only cytochrome c is provided (Table 1).A smaller portion of inactive and damaged RCs is measuredwith cytochrome c and Q0 in comparison cytochrome conly, as shown by lower values of α3.Following adsorption to the gold electrode surface, in the
presence of cytochrome c, the Fv/Fm values decreased to 0.51,0.26, and 0.21 for the respective Rba. sphaeroides 0, 4, and 24 hsamples, representing a ∼70% retention of photosyntheticactivity (Table 3). The Rsp. rubrum chromatophore preparationgave an Fv/Fm value of 0.39, as compared the 0.74 observed inwhole cells.21 This Fv/Fm value for the Rsp. rubrum chro-matophores on gold with cytochrome c represents a ∼90%retention of photosynthetic activity. Respective values of 0.38,0.17, and 0.36 were obtained for the 0, 4, and 24 h for Rba.sphaeroides when Q0 was added. Thus, for chromatophores ongold surfaces, the addition of cytochrome c and Q0 showed noobvious effect on the quantum yield of the primary chargeseparation. We attribute this apparent inconsistency with what
was observed in suspension to the substantially differentbehavior of the chromatophores on the surface and insuspension. While Q0 can easily access the QB pockets of theRCs in the chromatophores in suspension, the same cannot beexpected for the chromatophores on the gold surfaces. Theadhesion characteristics of the chromatophores on the surfaceand the partial disruption of the vesicles caused by interactionwith gold will determine their ultimate photosynthetic activity.The lack of active mixing of the film of liquid on thechromatophores is also a factor negatively affecting activity.Despite this, the value of τavg of the Rba. sphaeroideschromatophores on gold appears to be smaller when bothcytochrome c and Q0 are present, while no similar effect wasobserved for Rsp. rubrum chromatophores on gold. In the caseof the Rba. sphaeroides chromatophores, it seems that thefunctional integrity is enhanced by the presence of Q0, but thisdoes not consistently result in a higher photosynthetic activity(as reflected by the Fv/Fm).
Figure 7. Fluorescence kinetics studies of Rba. sphaeroides and Rsp. rubrunm chromatophores on gold-coated silicon substrates, obtained fromfluorescence induction and relaxation measurements. The STF−τ transients are described in the Figure 6 legend. All measurements are the averageof 50 iterations. (A) Performed in the presence of horse heart cytochrome c. (B) Performed in presence of cytochrome c and quinone Q0.
Table 3. Chromatophores on a Gold Surface with Cytochrome ca
sample LH1/LH2 F0 Fm Fv/Fm % (Fv/Fm)b σRC (Å2/q) τi (ms)
Rba. sphaeroides 0h chromatophores on Au−glass 3.25:1.00 450 918 0.51 76 27.2 τavg = 47.3 ± 7.9Rba. sphaeroides 4h chromatophores on Au−glass 2.71:1.00 926 1245 0.26 73 38.7 τavg = 49.5 ± 9.0Rba. sphaeroides 24h chromatophores on Au−glass 1.00:1.00 915 1160 0.21 70. 42.7 τavg = 39.0 ± 4.7Rsp. rubrum chromatophores on Au−glass n/a 1010 1643 0.39 93 16.0 τavg = 52.8 ± 6.3
aThe parameters refer to the fluorescence yield profiles shown in Figure 7a. F0 is the minimum of fluorescence, Fm is the maximum, Fv = Fm − F0,Fv/Fm is the quantum yield of photochemistry, σCC is the functional absorption cross section, I is the irradiance, τi refers to the equation f II(t) = ∑1
3
Ai(exp(−t/τi) used in the fitting of the fluorescence in region II, and αi corresponds to the ratio αi = Ai/(∑in Ai).
bPercentage retention of thequantum yield of primary charge separation (Fv/Fm) with respect to the corresponding chromatophore suspensions in Table 1.
The Journal of Physical Chemistry B Article
dx.doi.org/10.1021/jp402108s | J. Phys. Chem. B 2013, 117, 11249−1125911256
It is possible to conclude that some loss of photosyntheticactivity of chromatophores on the gold electrode surface hasarisen from limitations in the supply of accessible chromato-phores following adsorption to the gold surface. In chromato-phore suspensions, inactive chromatophores can be replacedwith active ones by simple dispersion while the immobilizedchromatophores are stationary and cannot replace lower-activityvesicles with more active ones. This ensemble effect, which isabsent in the immobilized structures, favors the chromatophoresthat have the largest and most active antenna among those insuspension. Nevertheless, it is shown below that sufficientantenna and RC activity was maintained for production of asignificant current at the chromatophore electrode surface.
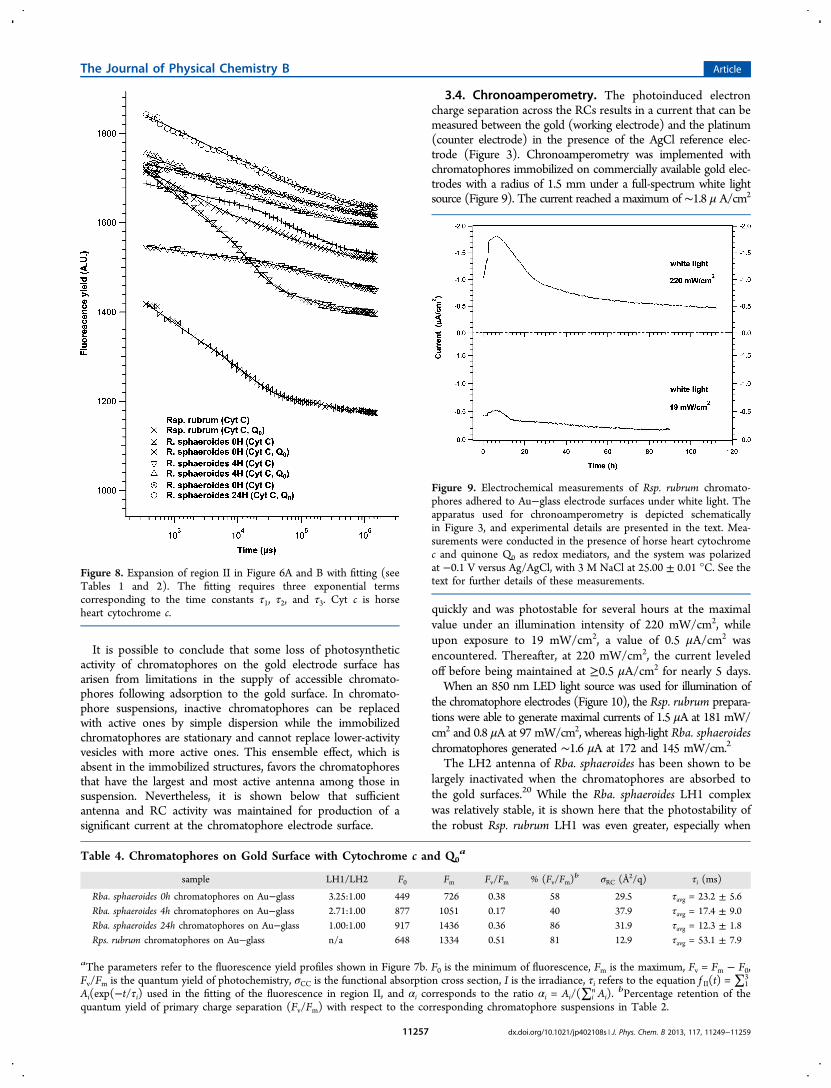
3.4. Chronoamperometry. The photoinduced electroncharge separation across the RCs results in a current that can bemeasured between the gold (working electrode) and the platinum(counter electrode) in the presence of the AgCl reference elec-trode (Figure 3). Chronoamperometry was implemented withchromatophores immobilized on commercially available gold elec-trodes with a radius of 1.5 mm under a full-spectrum white lightsource (Figure 9). The current reached a maximum of ∼1.8 μ A/cm2
quickly and was photostable for several hours at the maximalvalue under an illumination intensity of 220 mW/cm2, whileupon exposure to 19 mW/cm2, a value of 0.5 μA/cm2 wasencountered. Thereafter, at 220 mW/cm2, the current leveledoff before being maintained at ≥0.5 μA/cm2 for nearly 5 days.When an 850 nm LED light source was used for illumination of
the chromatophore electrodes (Figure 10), the Rsp. rubrum prepara-tions were able to generate maximal currents of 1.5 μA at 181 mW/cm2 and 0.8 μA at 97 mW/cm2, whereas high-light Rba. sphaeroideschromatophores generated ∼1.6 μA at 172 and 145 mW/cm.2
The LH2 antenna of Rba. sphaeroides has been shown to belargely inactivated when the chromatophores are absorbed tothe gold surfaces.20 While the Rba. sphaeroides LH1 complexwas relatively stable, it is shown here that the photostability ofthe robust Rsp. rubrum LH1 was even greater, especially when
Figure 8. Expansion of region II in Figure 6A and B with fitting (seeTables 1 and 2). The fitting requires three exponential termscorresponding to the time constants τ1, τ2, and τ3. Cyt c is horseheart cytochrome c.
Table 4. Chromatophores on Gold Surface with Cytochrome c and Q0a
sample LH1/LH2 F0 Fm Fv/Fm % (Fv/Fm)b σRC (Å2/q) τi (ms)
Rba. sphaeroides 0h chromatophores on Au−glass 3.25:1.00 449 726 0.38 58 29.5 τavg = 23.2 ± 5.6Rba. sphaeroides 4h chromatophores on Au−glass 2.71:1.00 877 1051 0.17 40 37.9 τavg = 17.4 ± 9.0Rba. sphaeroides 24h chromatophores on Au−glass 1.00:1.00 917 1436 0.36 86 31.9 τavg = 12.3 ± 1.8Rps. rubrum chromatophores on Au−glass n/a 648 1334 0.51 81 12.9 τavg = 53.1 ± 7.9
aThe parameters refer to the fluorescence yield profiles shown in Figure 7b. F0 is the minimum of fluorescence, Fm is the maximum, Fv = Fm − F0,Fv/Fm is the quantum yield of photochemistry, σCC is the functional absorption cross section, I is the irradiance, τi refers to the equation f II(t) = ∑1
3
Ai(exp(−t/τi) used in the fitting of the fluorescence in region II, and αi corresponds to the ratio αi = Ai/(∑in Ai).
bPercentage retention of thequantum yield of primary charge separation (Fv/Fm) with respect to the corresponding chromatophore suspensions in Table 2.
Figure 9. Electrochemical measurements of Rsp. rubrum chromato-phores adhered to Au−glass electrode surfaces under white light. Theapparatus used for chronoamperometry is depicted schematicallyin Figure 3, and experimental details are presented in the text. Mea-surements were conducted in the presence of horse heart cytochromec and quinone Q0 as redox mediators, and the system was polarizedat −0.1 V versus Ag/AgCl, with 3 M NaCl at 25.00 ± 0.01 °C. See thetext for further details of these measurements.
The Journal of Physical Chemistry B Article
dx.doi.org/10.1021/jp402108s | J. Phys. Chem. B 2013, 117, 11249−1125911257

illuminated under white light. This can be explained by thepresence of LH2 in the Rba. sphaeroides chromatophores, whichwill not have as strong of a contribution to the current as theLH1, which Rsp. rubrum chromatophores utilize as the only light-harvesting antenna. The lack of LH2 would allow more surfacearea to be occupied by RCs that have a fixed ratio to LH1.32
4. CONCLUSIONS
This study demonstrates that chromatophores isolated fromRsp. rubrum can be adsorbed onto gold surfaces. The adsorbedchromatophores were shown to generate a photodriven currentwhen exposed to both white light and 850 nm radiation. Whencompared to chromatophores isolated from Rba. sphaeroides,these preparations showed more robustness when exposed to highlight. Under high 850 nm illumination, the total charge generatedby the electrode is greater with the Rsp. rubrum chromatophoreseven though the peak current is slightly lower. This makeschromatophores isolated from Rsp. rubrum more suitable forincorporation into photovoltaic devices. In this connection, wehave recently fabricated a biohybrid near-IR sensing dye-sensitizedsolar cell in which the light-harvesting dye was replaced withrobust Rsp. rubrum chromatophores and the electrolytes with Q0and cytochrome c.34 In the present study, we demonstrate that inthe presence of residual cytochrome c and Q0, it is possible toresolve three time constants in the fluorescence relaxation kineticsof both types of chromatophores in suspension. Moreover, thefluorescence studies reported here have facilitated determinationof the survivng photosynthetic activity of the chromatophoreswhen adsorbed to gold electrode surfaces.
■ ASSOCIATED CONTENT
*S Supporting InformationTable S1, Assessment of Validity of Reaction Center ElectronTransfer Turnover Transient Curve Fitting. This material isavailable free of charge via the Internet at http://pubs.acs.org.
■ AUTHOR INFORMATIONCorresponding Author*E-mail: [email protected]. Fax: (732) 445-4213.
NotesThe authors declare no competing financial interest.
■ ACKNOWLEDGMENTSSupported by a grant from the U.S. Air Force Office ofScientific Research (M.V., P.G.F.) and the U.S. Department ofEnergy Grant No. DE-FG02-08ER15957 from the ChemicalSciences, Geosciences and Biosciences Division, Office of BasicEnergy Sciences, Office of Science (R.A.N.). We thank ArpitPatel for assistance in these studies, Maxim Gorbunov for usefuldiscussions on fluorescence induction/relaxation measurement,and Daniel T. Mastrogiovanni for obtaining the initial atomicforce microscopy topographs.
■ REFERENCES(1) Blankenship, R. E. Molecular Mechanisms of Photosynthesis;Blackwell Science Inc.: London, 2002.(2) Parson, W. W.; Warshel, A. Mechanism of Charge Separation inPurple Bacterial Reaction Centers. In The Purple Phototrophic Bacteria;Hunter, C. N., Daldal, F., Thurnauer, M. C., Beatty, J. T., Eds.;Springer: Dordrecht, The Netherlands, 2009; pp 355−377.(3) Karrasch, S.; Bullough, P. A.; Ghosh, R. The 8.5 Å ProjectionMap of the Light-Harvesting Complex I from Rhodospirillum rubrumReveals a Ring Composed of 16 Subunits. EMBO J. 1995, 14, 631−638.(4) Fotiadis, D.; Qian, P.; Philippsen, A.; Bullough, P. A.; Engel, A.;Hunter, C. N. Structural Analysis of the Reaction Center Light-Harvesting Complex I Photosynthetic Core Complex of Rhodospir-illum rubrum Using Atomic Force Microscopy. J. Biol. Chem. 2004,279, 2063−2068.(5) Janzen, A. F.; Seibert, M. Photoelectrochemical ConversionUsing Reaction-Center Electrodes. Nature 1980, 286, 584−585.(6) Katz, E. Y.; Shkuropatov, A. Y.; Vagabova, O. I.; Shuvalov, V. A.Coupling of Photoinduced Charge Separation in Reaction Centers of
Figure 10. Electrochemical measurements of (A) Rsp. rubrum and (B) high-light Rba. sphaeroides chromatophores adhered to Au−glass electrodesurfaces under near-IR light (850 nm ± 10 nm bandwidth filter). The apparatus and experimental details are described in the text and in the Figure 9legend.
The Journal of Physical Chemistry B Article
dx.doi.org/10.1021/jp402108s | J. Phys. Chem. B 2013, 117, 11249−1125911258

Photosynthetic Bacteria with Electron Transfer to a ChemicallyModified Electrode. Biochim. Biophys. Acta 1989, 976, 121−128.(7) Katz, E. Application of Bifunctional Reagents for Immobilizationof Proteins on a Carbon Electrode Surface: Oriented Immobilizationof Photosynthetic Reaction Centers. J. Electroanal. Chem. 1994, 365,157−164.(8) Lu, Y.; Xu, J.; Liu, B.; Kong, J. Photosynthetic Reaction CenterFunctionalized Nanocomposite Films: Effective Strategies for Probingand Exploiting the Photo-Induced Electron Transfer of PhotosensitiveMembrane Protein. Biosens. Bioelectron. 2007, 22, 1173−1185.(9) Trammell, S. A.; Wang, L.; Zullo, J. M.; Shashidhar, R.; Lebedev,N. Orientated Binding of Photosynthetic Reaction Centers on GoldUsing Ni-NTA Self-Assembled Monolayers. Biosens. Bioelectron. 2004,19, 1649−1655.(10) Trammell, S. A.; Spano, A.; Price, R.; Lebedev, N. Effect ofProtein Orientation on Electron Transfer Between PhotosyntheticReaction Centers and Carbon Electrodes. Biosens. Bioelectron. 2006,21, 1023−1028.(11) Lebedev, N.; Trammell, S. A.; Spano, A.; Lukashev, E.; Griva, I.;Schnur, J. J. Conductive Wiring of Immobilized PhotosyntheticReaction Center to Electrode by Cytochrome c. J. Am. Chem. Soc.2006, 128, 12044−12045.(12) Dorogi, M.; Balint, Z.; Miko, C.; Vileno, B.; Milas, M.; Hernadi,K.; Forro, L.; Varo, G.; Nagy, L. Stabilization Effect of Single-WalledCarbon Nanotubes on the Functioning of Photosynthetic ReactionCenters. J. Phys .Chem. B 2006, 110, 21473−21479.(13) Lebedev, N.; Trammell, S. A.; Tsoi, S.; Spano, A.; Kim, J. H.;Xu, J.; Twigg, M. E. .; Schnur, J. M. Increasing Efficiency ofPhotoelectronic Conversion by Encapsulation of PhotosyntheticReaction Center Proteins in Arrayed Carbon Nanotube Electrode.Langmuir 2008, 24, 8871−8876.(14) Oda, I.; Iwaki, M.; Fujita, D.; Tsutsui, Y.; Ishizaka, S.; Dewa, M.;Nango, M.; Kajino, T.; Fukushima, Y.; Itoh, S. Photosynthetic ElectronTransfer from Reaction Center Pigment−Protein Complex in SilicaNanopores. Langmuir 2010, 26, 13399−13406.(15) Suemori, Y.; Nagata, M.; Nakamura, Y.; Nakagawa, K.; Okuda,A.; Inagaki, J.-I.; Shinohara, K.; Ogawa, M.; Iida, K.; Dewa, T.;Yamashita, K.; Gardiner, A.; Cogdell, R. J.; Nango, M. Self-AssembledMonolayer of Light-Harvesting Core Complexes of PhotosyntheticBacteria on an Amino-Terminated ITO Electrode. Photosynth. Res.2006, 90, 17−21.(16) Kondo, M.; Nakamura, Y.; Fujii, K.; Nagata, M.; Suemori, Y.;Dewa, T.; Iida, K.; Gardiner, A. T.; Cogdell, R. J.; Nango, M. Self-Assembled Monolayer of Light-Harvesting Core Complexes fromPhotosynthetic Bacteria on a Gold Electrode Modified withAlkanethiols. Biomacromolecules 2007, 8, 2457−2463.(17) Kondo, M.; Iida, K.; Dewa, T.; Tanaka, H.; Ogawa, T.;Nagashima, S.; Nagashima, K. V.; Shimada, K.; Hashimoto, H.;Gardiner, A. T.; Cogdell, R. J.; Nango, M. Photocurrent and ElectronicActivities of Oriented-His-Tagged Photosynthetic Light-Harvesting/Reaction Center Core Complexes Assembled onto a Gold Electrode.Biomacromolecules 2012, 13, 432−438.(18) Magis, G. J.; Olsen, J. D.; Reynolds, N. P.; Leggett, G. J.;Hunter, C. N.; Aartsma, T. J.; Frese, R. N. Use of Engineered UniqueCysteine Residues to Facilitate Oriented Coupling of Proteins Directlyto a Gold Substrate. Photochem. Photobiol. 2011, 87, 1050−1057.(19) den Hollander, M. J.; Magis, J. G.; Fuchsenberger, P.; Aartsma,T. J; Jones, M. R.; Frese, R. N. Enhanced Photocurrent Generation byPhotosynthetic Bacterial Reaction Centers through Molecular Relays,Light-Harvesting Complexes, and Direct Protein−Gold Interactions.Langmuir 2011, 27, 10282−10294.(20) Magis, G.; den Hollander, M.-J.; Onderwaater, W. G.; Olsen, J.D.; Hunter, C. N.; Aartsma, T. J.; Frese, R. N. Light Harvesting,Energy Transfer and Electron Cycling of a Native PhotosyntheticMembrane Adsorbed onto a Gold Surface. Biochim. Biophys. Acta2010, 1798, 637−645.(21) Koblízek, M.; Shih, J. D.; Breitbart, S. I.; Ratcliffe, E. C.; Kolber,Z. S.; Hunter, C. N.; Niederman, R. A. Sequential Assembly ofPhotosynthetic Units in Rhodobacter sphaeroides as Revealed by Fast
Repetition Rate Analysis of Variable Bacteriochlorophyll a Fluo-rescence. Biochim. Biophys. Acta 2005, 1706, 220−231.(22) Woronowicz, K.; Sha, D.; Frese, R. N.; Sturgis, J. N.; Nanda, V.;Niederman, R. A. The Effects of Protein Crowding in BacterialPhotosynthetic Membranes on the Flow of Quinone Redox SpeciesBetween the Photochemical Reaction Center and the Ubiquinol−Cytochrome c2 Oxidoreductase. Metallomics 2011, 3, 765−774.(23) Cohen-Bazire, G.; Sistrom, W. R.; Stanier, R. Y. Kinetic Studiesof Pigment Synthesis by Non-Sulfur Purple Bacteria. J. Cell. Comp.Physiol. 1956, 49, 25−68.(24) Aagaard, J.; Sistrom, W. R. Control of Synthesis of ReactionCenter Bacteriochlorophyll in Photosynthetic Bacteria. Photochem.Photobiol. 1972, 15, 209−225.(25) Ormerod, J. G.; Ormerod, K. S.; Gest, H. Light-DependentUtilization of Organic Compounds and Photoproduction of MolecularHydrogen by Photosynthetic Bacteria; Relationships with NitrogenMetabolism. Arch. Biochem. Biophys. 1961, 94, 449−463.(26) Inamine, G. S.; Niederman, R. A. Development and Growth ofPhotosynthetic Membranes of Rhodospirillum rubrum. J. Bacteriol.1982, 150, 1145−1153.(27) Woronowicz, K.; Sha, D.; Frese, R. N.; Niederman, R. A. TheAccumulation of the Light-Harvesting 2 Complex during Remodelingof the Rhodobacter sphaeroides Intracytoplasmic Membrane Results in aSlowing of the Electron Transfer Turnover Rate of PhotochemicalReaction Centers. Biochemistry 2011, 50, 4819−4829.(28) Sturgis, J. N.; Hunter, C. N.; Niederman, R. A. Spectra andExtinction Coefficients of Near-Infrared Absorption Bands inMembranes of Rhodobacter sphaeroides Mutants Lacking Light-Harvesting and Reaction Center Complexes. Photochem. Photobiol.1988, 48, 243−247.(29) Papagiannakis, E.; Kennis, J. T. M.; van Stokkum, I. H. M.;Cogdell, R. J.; van Grondelle, R. An Alternative Carotenoid-to-Bacteriochlorophyll Energy Transfer Pathway in Photosynthetic LightHarvesting. Proc. Natl. Acad. Sci. U.S.A. 2002, 99, 6017−6022.(30) Westerhuis, W. H. J.; Sturgis, J. N.; Ratcliffe, E. C.; Hunter, C.N.; Niederman, R. A. Isolation, Size Estimates and SpectralHeterogeneity of an Oligomeric Series of Light-Harvesting 1Complexes from Rhodobacter sphaeroides. Biochemistry 2002, 41,8698−8707.(31) Bahatyrova, S.; Frese, R. N.; Siebert, C. A.; van der Werf, K. O.;van Grondelle, R.; Niederman, R. A.; Bullough, P. A.; Otto, C.; Olsen,J. D.; Hunter, C. N. The Native Architecture of a PhotosyntheticMembrane. Nature 2004, 430, 1058−1062.(32) Jamieson, S. J.; Wang, P.; Qian, P.; Kirkland, J. Y.; Conroy, M. J.;Hunter, C. N.; Bullough, P. A. Projection Structure of thePhotosynthetic Reaction Centre−Antenna Complex of Rhodospir-illum rubrum at 8.5 A Resolution. EMBO J. 2002, 21, 3927−3935.(33) Vittadello, M.; Gorbunov, M. Y.; Mastrogiovanni, D. T.;Wielunski, L. S.; Garfunkel, E. L.; Guerrero, F.; Kirilovsky, D.; Sugiura,M.; Rutherford, A. W.; Safari, A.; Falkowski, P. G. PhotoelectronGeneration by Photosystem II Core Complexes Tethered to GoldSurfaces. ChemSusChem 2010, 3, 471−475.(34) Woronowicz, K.; Ahmed, S.; Biradar, A. A.; Biradar, A. V.;Birnie, D. P.; Asefa, T.; Niederman, R. A. Near-IR Absorbing SolarCell Sensitized with Bacterial Photosynthetic Membranes. Photochem.Photobiol. 2012, 88, 1467−1472.
The Journal of Physical Chemistry B Article
dx.doi.org/10.1021/jp402108s | J. Phys. Chem. B 2013, 117, 11249−1125911259
Copyright © 2022 FDOKUMEN




















