
Frequency tuning, latencies, and responses to frequency-modulated sweeps in the inferior colliculus...
15
ORIGINAL PAPER M. J. Ferragamo Æ T. Haresign Æ J. A. Simmons Frequency tuning, latencies, and responses to frequency-modulated sweeps in the inferior colliculus of the echolocating bat, Eptesicus fuscus Accepted: 21 June 1997 Abstract Neurons in the inferior colliculus (IC) of the awake big brown bat, Eptesicus fuscus, were examined for joint frequency and latency response properties which could register the timing of the bat’s frequency- modulated (FM) biosonar echoes. Best frequencies (BFs) range from 10 kHz to 100 kHz with 50% tuning widths mostly from 1 kHz to 8 kHz. Neurons respond with one discharge per 2-ms tone burst or FM stimulus at a characteristic latency in the range of 3–45 ms, with latency variability (SD) of 50 ls to 4–6 ms or more. BF distribution is related to biosonar signal structure. As observed previously, on a linear frequency scale BFs appear biased to lower frequencies, with 20–40 kHz overrepresented. However, on a hyperbolic frequency (linear period) scale BFs appear more uniformly dis- tributed, with little overrepresentation. The cumulative proportion of BFs in FM 1 and FM 2 bands reconstructs a scaled version of the spectrogram of FM broadcasts. Correcting FM latencies for absolute BF latencies and BF time-in-sweep reveals a subset of IC cells which re- spond dynamically to the timing of their BFs in FM sweeps. Behaviorally, Eptesicus perceives echo delay and phase with microsecond or even submicrosecond accu- racy and resolution, but even with use of phase-locked FM and tone-burst stimuli the cell-by-cell precision of IC time-frequency registration seems inadequate by itself to account for the temporal acuity exhibited by the bat. Key words Bats Æ Biosonar Æ Echolocation Æ Target ranging Æ Inferior colliculus Abbreviations AM amplitude modulation Æ BF best frequency Æ BP best period (1/BF) Æ EF excitatory fre- quency Æ FM n frequency modulated (nth harmonic) Æ IC inferior colliculus Æ PSTH peristimulus time histo-gra- m Æ SD standard deviation Introduction Echolocating big brown bats (Eptesicus fuscus) perceive the distance to objects, or target range, from the delay of frequency-modulated (FM) biosonar echoes (Moss and Schnitzler 1995; Simmons et al. 1995). These bats can detect insect-sized targets at distances of about 3–5 m (Kick 1982). During pursuit of prey, the bat typically tracks the target continuously until it has approached to within 10–15 cm, when the target is seized. Eptesicus shortens the duration of its sounds progressively from 8 to 15 ms or more at the start of the approach to 2–5 ms when the bat has closed to distances of 0.5–1 m, and finally to as little as 0.5 ms at the moment of capture (Grin 1958; Kalko and Schnitzler 1993; Simmons 1989). Acoustic tracking of delay demonstrates that the bat perceives and progressively updates its estimates of target range throughout interception. The bat’s neural mechanisms for target ranging thus must accommodate a continuous span of echo delays from 25 to 30 ms down to 1 ms or less. General features of the neural responses associated with representing echo delay have been identified physiologically in four species of bats, including Epte- sicus (Berkowitz and Suga 1989; Dear et al. 1993a; Haplea et al. 1994; Kuwabara and Suga 1993; O’Neill 1995; Pollak et al. 1977a–c; Suga et al. 1995). Coding of echo delay involves systematic dispersal of response la- tencies to create delay lines that register the time-of- occurrence of dierent frequencies in the FM sweeps of sonar emissions and echoes in the auditory brainstem J Comp Physiol A (1998) 182: 65–79 Ó Springer-Verlag 1998 M.J. Ferragamo (&) 1 Æ T. Haresign 2 Æ J.A. Simmons Department of Neuroscience, Brown University, Providence, RI 02912, USA Tel.: +1-401 863-1542; Fax: +1-401 863-1074 1 Present addresses: Department of Neurophysiology, University of Wisconsin Medical Center, Madison, WI 53706, USA 2 NAMS, Stockton College, Pomona, NJ 08240, USA
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Frequency tuning, latencies, and responses to frequency-modulated sweeps in the inferior colliculus...

ORIGINAL PAPER
M. J. Ferragamo á T. Haresign á J. A. Simmons
Frequency tuning, latencies, and responses to frequency-modulatedsweeps in the inferior colliculus of the echolocating bat, Eptesicus fuscus
Accepted: 21 June 1997
Abstract Neurons in the inferior colliculus (IC) of theawake big brown bat, Eptesicus fuscus, were examinedfor joint frequency and latency response propertieswhich could register the timing of the bat's frequency-modulated (FM) biosonar echoes. Best frequencies(BFs) range from 10 kHz to 100 kHz with 50% tuningwidths mostly from 1 kHz to 8 kHz. Neurons respondwith one discharge per 2-ms tone burst or FM stimulusat a characteristic latency in the range of 3±45 ms, withlatency variability (SD) of 50 ls to 4±6 ms or more. BFdistribution is related to biosonar signal structure. Asobserved previously, on a linear frequency scale BFsappear biased to lower frequencies, with 20±40 kHzoverrepresented. However, on a hyperbolic frequency(linear period) scale BFs appear more uniformly dis-tributed, with little overrepresentation. The cumulativeproportion of BFs in FM1 and FM2 bands reconstructsa scaled version of the spectrogram of FM broadcasts.Correcting FM latencies for absolute BF latencies andBF time-in-sweep reveals a subset of IC cells which re-spond dynamically to the timing of their BFs in FMsweeps. Behaviorally, Eptesicus perceives echo delay andphase with microsecond or even submicrosecond accu-racy and resolution, but even with use of phase-lockedFM and tone-burst stimuli the cell-by-cell precision ofIC time-frequency registration seems inadequate by itselfto account for the temporal acuity exhibited by the bat.
Key words Bats á Biosonar á Echolocation áTarget ranging á Inferior colliculus
Abbreviations AM amplitude modulation á BF bestfrequency á BP best period (1/BF) á EF excitatory fre-quency á FMn frequency modulated(nth harmonic) á ICinferior colliculus á PSTH peristimulus time histo-gra-m á SD standard deviation
Introduction
Echolocating big brown bats (Eptesicus fuscus) perceivethe distance to objects, or target range, from the delay offrequency-modulated (FM) biosonar echoes (Moss andSchnitzler 1995; Simmons et al. 1995). These bats candetect insect-sized targets at distances of about 3±5 m(Kick 1982). During pursuit of prey, the bat typicallytracks the target continuously until it has approached towithin 10±15 cm, when the target is seized. Eptesicusshortens the duration of its sounds progressively from 8to 15 ms or more at the start of the approach to 2±5 mswhen the bat has closed to distances of 0.5±1 m, and®nally to as little as 0.5 ms at the moment of capture(Gri�n 1958; Kalko and Schnitzler 1993; Simmons1989). Acoustic tracking of delay demonstrates that thebat perceives and progressively updates its estimates oftarget range throughout interception. The bat's neuralmechanisms for target ranging thus must accommodatea continuous span of echo delays from 25 to 30 ms downto 1 ms or less.
General features of the neural responses associatedwith representing echo delay have been identi®edphysiologically in four species of bats, including Epte-sicus (Berkowitz and Suga 1989; Dear et al. 1993a;Haplea et al. 1994; Kuwabara and Suga 1993; O'Neill1995; Pollak et al. 1977a±c; Suga et al. 1995). Coding ofecho delay involves systematic dispersal of response la-tencies to create delay lines that register the time-of-occurrence of di�erent frequencies in the FM sweeps ofsonar emissions and echoes in the auditory brainstem
J Comp Physiol A (1998) 182: 65±79 Ó Springer-Verlag 1998
M.J. Ferragamo (&)1 á T. Haresign2 á J.A. SimmonsDepartment of Neuroscience, Brown University,Providence, RI 02912, USATel.: +1-401 863-1542; Fax: +1-401 863-1074
1Present addresses:Department of Neurophysiology,University of Wisconsin Medical Center,Madison, WI 53706, USA
2NAMS, Stockton College, Pomona,NJ 08240, USA

and inferior colliculus (IC) (Casseday and Covey 1995;Sullivan 1982b), followed by delay-selective coincidencemechanisms for creating echo delay-tuned neurons inthe IC, superior colliculus, thalamus and cortex (Dearet al. 1993a, b; Dear and Suga 1995; Feng et al. 1978;O'Neill 1995; Simmons 1980; Suga et al. 1995; Sullivan1982a, b; Valentine and Moss 1997; Wong and Shannon1988). Relatively short physiological delays of severalmilliseconds probably are a consequence of neuronalspike travel times from nucleus to nucleus up the audi-tory pathway, but longer delays seem to be created byinhibition, which retards responses for up to tens ofmilliseconds (Casseday et al. 1994; Saitoh and Suga1995). To create delay lines that match the full range ofbiologically relevant echo delays, dispersion of latenciesin the IC of Eptesicus densely covers a span of at least25±30 ms from shortest to longest (Haplea et al. 1994;Jen and Schlegel 1982; Poon et al. 1990), and read-out ofdelay lines by delay tuning is present for delays from25 ms to 30 ms down to ca. 1 ms (Dear et al. 1993a, b).
The neural mechanisms for delay coding are pre-sumed to support the bat's acoustic images (Moss andSchnitzler 1995; Simmons et al. 1995, 1996), but there isa large discrepancy between behavioral and physiologi-cal measures of the bat's acuity for determining delay. Insome experiments, big brown bats perceive echo delaywith submicrosecond accuracy (Menne et al. 1989; Mossand Schnitzler 1989; Simmons 1979; Simmons et al.1990a) and microsecond two-point resolution (Saillantet al. 1993), and bats also are sensitive to echo phaseshifts as correspondingly small shifts in apparent delay(Menne et al. 1989; Moss and Simmons 1993; Simmonset al. 1990a). In contrast, most interpretations of phys-iological data to estimate acuity for delay yield estimatesof roughly 50±100 ls at best (Pollak et al. 1977c;Schnitzler et al. 1985). This discrepancy has attractedconsiderable attention because understanding where itarises may be an important test of our knowledge ofbrain function (Carr 1993; Rieke et al. 1997). Althoughit is possible that the discrepancy merely re¯ects unno-ticed artifacts that unintentionally magnify behavioralmeasures of delay acuity (Menne et al. 1989; Pollak1993; Schnitzler et al. 1985), a variety of control proce-dures so far have failed to trace the results to any speci®cartifact (Simmons et al. 1990ab; Simmons 1993) andindeed have identi®ed new evidence that is consistentwith the results actually re¯ecting ®ne delay perception(Simmons 1996; Simmons et al. 1995).
Physiological accuracy of delay coding depends onthe variability of response latencies to biosonar emis-sions and echoes in the brainstem and IC (Haplea et al.1994; Suga 1970; Pollak et al. 1977b, c), and the sharp-ness of delay-tuning curves in the auditory midbrain andcortex (Dear et al. 1993a; Suzuki and Suga 1990).However, these two neural response parameters do notyield the same estimates of physiological delay acuity. InEptesicus, latency variability in the brainstem is about20±50 ls, while in the IC it increases to perhaps 50±100 ls at best, and in most cells is hundreds of micro-
seconds or worse (Casseday and Covey 1995). Whenlatency variability and amplitude/latency trading areconsidered together, physiological delay acuity in theFM bat's IC is estimated to be no better than 50±100 ls(Pollak et al. 1977c). Delay-tuning curves typically havewidths of several milliseconds (Dear et al. 1993a, b; Dearand Suga 1995; Valentine and Moss 1997), but thesesame delay-tuned responses can have latency variabili-ties as small as 100±200 ls (Feng et al. 1978; Dear et al.1993a, b; Dear and Suga 1995). Such discrepancieswithin the physiological data reveal the di�culty ofpredicting the behavioral acuity of the bat, and hencethe scope of acceptable behavioral observations (Pollak1993), without knowing beforehand how the bat's brainreads its own internal codes for delay.
Most physiological experiments on which predictionsof delay acuity could be based have lacked one or moreof the following critical features: use of phase-lockedshort-duration FM stimuli, collection of both frequencyand temporal response parameters from the same neu-rons at the same time in the same experiment, andconsideration of data-display dimensions other thanlinear frequency and peristimulus time histogram(PSTH) time (raw latency) which might reveal subtle ordynamic properties of responses. For example, somestudies have reported only frequency tuning and tuningwidth, or latency and its variability, but not both to-gether for the same cells, and usually only for tonebursts rather than wideband FM sounds that mimicecholocation signals. It is critical that FM stimuli sim-ulating sonar emissions and echoes be phase lockedbecause bats can perceive echo phase shifts as changes inapparent echo delay (Menne et al. 1989; Moss andSimmons 1993; Simmons et al. 1990a). However, analogswept-frequency generators ordinarily cannot deliver therequisite phase stability across multiple stimulus pre-sentations. Here, we measured the ability of single unitsin the IC of Eptesicus to signal the time-of-occurrence oftheir best frequencies (BFs) in phase-locked FM sweepsas the physiological basis for echo delay perception. Weused a variety of alternative data-display strategies toexplore the time-frequency and dynamic organization ofthese responses. IC neurons specialized for respondingselectively to FM sweeps are assumed to be incorporatedinto neuronal circuits which bring together inhibitoryand excitatory inputs from adjacent frequencies at spe-ci®c times (manifested as inhibitory and excitatory re-gions of tuning curves; Casseday et al. 1994, 1997; Parkand Pollak 1993; Suga 1965, 1969, 1972; Suga andSchlegel 1973). Excitation for FM-selective responses toparticular frequencies in FM sweeps should be precededin time by a transient interval of very little or no re-sponse due to inhibitory sidebands around the excitatoryfrequency, and we examined ``dechirped'' single unitresponses to brief FM sweeps for evidence that thisdynamic e�ect occurs near each cell's BF. A related re-verse correlation strategy has been used e�ectively toidentify IC neurons that respond selectively to transientamplitude-modulation (AM) glints such as would be
66

produced in echoes by insect wing-beats (Lesser et al.1990). In this report we are concerned particularly withneural responses evoked by FM sounds with durationsof about 2 ms because this is approximately the durationof the signals Eptesicus uses in laboratory behavioralstudies of the acuity for echo-delay perception. Our in-tention was to search for dynamic features of responsesto FM sweeps and determine whether single unit re-sponses can account for the bat's behavioral delay acuitywhen the stimuli more closely mimic sounds receivedduring echolocation.
Materials and methods
Surgical procedure
Big brown bats, E. fuscus (Kurta and Baker 1990) were maintainedon a diet of mealworms in a colony room at a temperature of about22±25 °C. Individual bats were prepared for neurophysiologicalrecordings by a simple surgical procedure to expose the skull overthe IC and to attach a post to the anterodorsal surface of the skull.One day prior to this surgery, a healthy animal (15±25 g) was se-lected and the hair from the top of the head and from around theshoulders was removed with depilatory (``Nair''). On the day ofthe surgery, after the animal became settled under anesthesia[dosage of 0.004 ml g)1 of 15% (100 mgml)1) ketamine, 15%(100 mg ml)1) xylazine, 70% (0.09%) saline], an incision was madeat the midline of the skull. The overlying temporal muscles werepartially removed to expose the dorsal surface of the skull. The ¯athead of a small metal screw was mounted on the skull with cyan-oacrylate adhesive at the midline anterior to the site of the IC. Thebat was then allowed to recover for at least 48 h prior to the ®rstneurophysiological recording experiment.
On the day of an experiment, the bat was placed in a Plexiglasholder with its head, which protruded from the holder, ®xed byfastening the screw into a specially designed clamp. This apparatusenabled limited movement of the animal's torso, while also stabi-lizing the skull for extracellular recording. A small hole to admitthe recording electrode was hand-drilled into the skull overlying theIC with a sharpened no. 9 sewing needle. A single recording sessionwas limited to 8 h or less, with at least 1 day of rest between ses-sions. This report includes data collected from ten bats. Each batwas studied in multiple recording sessions for periods up to 4weeks.
Acoustic stimulation
The bat and the acoustic transducers for delivering stimuli wereplaced in an Industrial Acoustics sound-proof and vibration-iso-lated chamber lined with convoluted acoustic foam which served toreduce ultrasonic reverberations. Acoustic stimuli could be deliv-ered dichotically (independently to each ear) through a systemconsisting of two Bruel and Kjaer model 4135 (1/4 inch) micro-phones driven as earphones. However, the particular experimentsreported below used identical stimulation at the two ears, or dioticstimulation. The sound sources were inserted into the external earsnear the ear canals at a position above the tragus within the pinnaof each ear (Wenstrup et al. 1988). Digitally synthesized signalwaveforms were generated by computer programs which also si-multaneously supervised the collection of neural responses to thesesounds. These programs were developed to coordinate the opera-tion of both the A/D data-acquisition and D/A waveform-gener-ating boards (ISC-16 and 200 series manufactured by RCElectronics) within one IBM PC-AT computer. Stimulus wave-forms (16-bit accuracy, 1-MHz sampling rate) were generatedthrough two D/A channels, one channel leading to the sound-producing microphone in each ear. The computer programs
allowed on-line control of the frequency or FM sweep structure ofsounds, as well as the amplitude, time-of-arrival, and phase of thesignals independently at the two ears.
In the present study we presented digitally synthesized, phase-stable, tone bursts and FM bursts which were from 2 ms to 5 ms induration and which had an envelope shaped by a Hanning windowthat provided a rise-fall time of about 0.5 ms. All stimuli werepresented at a 4-Hz repetition rate. The phase and ®ne waveformstructure were identical from one repetition of the stimulus to thenext. By using a D-to-A sampling rate of 1 MHz, many timeshigher than the maximum Nyquist frequency of 200 kHz for our10- to 100-kHz FM sweeps and tone bursts, we avoided producingsignals whose spectra contained notches caused by incompletetracing of the peaks of individual cycles at certain frequencies. Eachdigitally synthesized signal was 5 kHz high-pass ®ltered (Rocklandmodel 442 ®lters) and then ampli®ed and added to a 200-V DC biassignal before being delivered to the sound sources. A combinationof digital and analog attenuation controlled the amplitude ofstimuli presented through the left and right ear channels.
The acoustic delivery system was calibrated by placing a 4138(1/8 inch) Bruel and Kjaer condenser microphone at the position ofthe tympanum in the head of a dead bat mounted in the apparatus[for details see Wotton et al. (1995)]. The peak-to-peak amplitudeof pure tone bursts was measured for each sound source at fre-quencies spanning 10±100 kHz in 5-kHz steps. The resulting fre-quency response shown in Fig. 1 is quite similar for the soundsource at each ear. Acoustic cross-talk between the left and theright ears was attenuated by more than 30 dB at frequencies be-tween 10 kHz and 30 kHz, and by more than 35 dB at frequenciesabove 30 kHz.
Physiological recording techniques
Action potentials generated by single and multiple units were re-corded with either glass micropipettes ®lled with 3 mol l)1 NaCland having impedances of 5±15 MW, or glass/carbon-®ber elec-trodes having impedances of about 0.25±1 MW. The recordingelectrode was advanced in approximately 2- to 3-lm steps by aremote hydraulic microdrive (Stolting). The depth was recordedfrom the microdrive scale during the experiment; it was later cor-rected to the brain surface contact point by listening to the onset ofhigh impedance noise in the recording upon electrode withdrawal.
The physiological signal was ampli®ed with a WPI DAM-05preampli®er, band-pass ®ltered (Rockland model 442 adjustable®lter) from 300 Hz to 3 kHz, and monitored audiovisuallythroughout the experiment as a means of observing the conditionof the physiological preparation. The occurrence of a neural actionpotential was detected with a custom-built window discriminatorwhich generated a TTL pulse that was delivered to the A/D board
Fig. 1 Calibration curves for left and right earphones based onfrequency response measurements of the stimulus delivered to theeardrum of a bat external ear preparation (see Wotton et al. 1995)
67

(RC Electronics model ISC-16 data acquisition system). The onsetof this digital signal was stored as representing the time-of-occur-rence of the neuronal spike. On-line, real-time, computer terminalraster displays of digitally registered spike times were used tomonitor the acquisition of data and as an aid for identifyingstimulus conditions for subsequent stages of experiments.
During advance of the recording electrode, the search stimulusfor isolating neurons to be studied was a 2-ms FM burst (hyper-bolic frequency sweep, or linear period sweep) that mimics the bat'sown sonar sounds (FM from 50 kHz to 25 kHz in the ®rst har-monic and from 100 kHz to 50 kHz in the second harmonic) at apeak-to-peak amplitude of 80±90 dB SPL (RMS amplitude71±81 dB SPL). If a unit was isolated and its spike appeared to beat an acceptable signal-to-noise ratio (peak-to-peak spike ampli-tude above background noise approximately 4:1 or better), a seriesof automated stimulus presentation sequences were run to gather asmuch data as possible about frequency tuning, dynamic range,latency to FM bursts, and various specialized aspects of responsesin the time the cell remained in good recording condition.
Stimulus regime for characterizing neuronal responses
The BF and response threshold of single units and small multiunitrecords were initially approximated by audiovisually assessing thefrequency which evoked the greatest spike probability (few neuronsdischarged more than one to two spikes per stimulus, so responserate is not a good term for what is measured). To quantify BF by auniform and objective paradigm, a series of 2-ms tone bursts(10±110 kHz) were presented in small frequency steps within aspeci®ed frequency range about 10 dB above the audiovisually es-timated threshold, and usually also at several other amplitude levelsin the range up to about 40 dB above threshold. These were pre-sented at a selected amplitude, then repeated at a lower amplitudeuntil a threshold was obtained. We then generated monaural am-plitude response curves (input-output curves) in 5-dB steps for theipsi- and contralateral ear with the tone bursts set at the cell's BF(these data to be published separately). After this, we recorded theunit's responses to 2- to 5-ms FM bursts either having a 20- to30-kHz bandwidth centered at BF or else having two harmonicsrepresenting the full range of echolocation frequencies ± FM1 from50 kHz to 25 kHz and FM2 from 100 kHz to 50 kHz (hyperbolicfrequency sweeps). To characterize FM latencies, we used ``stan-dard'' 2-ms hyperbolic downward FM sweeps with sweepwidths of20±30 kHz around BF and with envelopes shaped by a Hanningwindow (see Fig. 4A for starting and ending frequencies). Eachsuch sweep represented a segment clipped from the wider FM1 orFM2 sweeps used by the bat. These FM stimuli were presented atan amplitude approximately 10 dB above BF tone-burst threshold,and usually also at higher amplitudes as well. In some cases, FMsounds were presented with both upward and downward frequencysweeps.
In all these protocols, 40 or 48 repetitions of each stimulus weregiven while the corresponding responses were collected, along withthe spontaneous activity occurring before and after each sequenceof di�erent test stimuli. To control for changes in neural responsesover time due to changes in the state of the awake animal, stimuliassessing response sensitivity to any given acoustic parameter werepresented in blocks of four sequences which interleaved di�erentvalues of the parameter being studied.
Most of the response parameters we studied could be deter-mined by straightforwardly plotting response probability or re-sponse strength against magnitudes of di�erent stimulusparameters. Response latencies, however, presented special prob-lems for objective quanti®cation that had to be addressed speci®-cally rather than falling back on audiovisual methods. Responselatencies of all cells were quanti®ed by two independent methods,both performed automatically by computer software. The ®rstmethod calculated the latency of the ®rst spike (with 50-ls reso-lution) for each stimulus presentation, along with the mean andstandard deviation of the resulting latency distribution. The secondmethod for determining latency examined the cumulative PSTH to
search for the onset of a positive change in slope beyond a speci®ccriterion value.
Results
A total of 102 single units and 128 small multiunitclusters were recorded and subjected to analysis aftertheir spikes were discriminated by the amplitude windowtechnique (digital spike timing). At some recording sitesnot all stimulus conditions were run due to inability to``hold'' the cell long enough to run every protocol. Re-cording sites were located at depths of 100±2100 lm inthe IC, and at numerous positions along the rostral-caudal and mediolateral extent of the IC. Responses ofsingle units were identi®ed as such by consistency ofspike amplitude and waveshape, especially across mul-tiple presentations of stimuli having di�erent ampli-tudes, and by the all-or-none character of the spikes (onsome stimulus presentations the response failed to occurat all rather than exhibiting a graded amplitude). Thosemultiunit responses that we analyzed by the window-discriminator technique contained spikes from only afew cells and yielded unambiguous frequency-tuningestimates and latencies from the cumulative PSTHmethod. Most frequency domain response properties ±including the distribution of BFs, response thresholds atBF, topographic organization of BFs in the IC ± weregenerally similar to ®ndings in previous neurophysio-logical studies of the IC of anesthetized and unanes-thetized E. fuscus (Casseday and Covey 1992; Jen andSchlegel 1982; Poon et al. 1990), but latencies of re-sponses covered a somewhat wider span of times thanhas previously been reported. Our search stimuli wereFM sweeps, but we found no cells that responded to FMsweeps that could not also be driven by tone bursts andcharacterized as having a well-de®ned BF.
Frequency tuning of responses
Best frequencies
All single units and multiunit clusters whose spikes weredetected by the window discriminator and treated asdigital events responded most strongly at a single,readily characterized, and unambiguous BF. They dif-fered only in the speci®c value of BF and in the sharp-ness of tuning around BF. Figure 2 shows the relationbetween BF and various other parameters of the singleunit responses. BFs were measured at values from 14 to101 kHz, which spans the entire range of frequenciesemitted in the bat's biosonar signals (Fig. 2A showsdistribution of BFs in 5-kHz bins; Fig. 2B±G shows BFvalues to 1 kHz). Of these BFs, 71.3% (n � 164=230) fellbetween 20 and 45 kHz, in the range of the ®rst har-monic of the bat's sounds. Only 16.5% (n � 38=230) ofthe cells had BFs above 45 kHz. This uneven BF dis-tribution has been observed previously in Eptesicus and
68

is described as overrepresentation of frequencies from 20to 40 kHz (Casseday and Covey 1992; Jen and Schlegel1982; Poon et al. 1990). No cells we recorded from theIC had dual or multi-peaked excitatory tuning curves,although multi-peaked tuning curves are found exten-sively in the auditory cortex of Eptesicus (Dear et al.1993a).
BFs at di�erent recording depths
Single-unit BFs generally increased as a function of re-cording depth along the dorsal-ventral axis of the IC(Fig. 2B). At depths of 0±1 mm, there is a somewhattighter clustering and obvious sloping relation covering20±45 kHz (frequencies of FM1 in the bat's sonarsounds), but there is considerably more scatter above45 kHz (mostly FM2 frequencies) at depths of 1±2 mm.Even in the frequency range where a relation betweenBF and recording depth is evident, however, there still isconsiderable BF scatter. In e�ect, neurons with BFs in aspan 20 kHz wide around a particular site's nominaltonotopic frequency can be found at depths down to0.75±1.0 mm, while to a lesser degree neurons with
almost any BF from 20 to 80 kHz can be found atdepths from 0.25 mm to 2 mm.
Thresholds for responses at di�erent frequencies
The response threshold was de®ned as the stimulusamplitude at BF for which there was a response justabove the spontaneous ®ring rate. This was relativelyeasy to determine since 75.7% (n � 174=230) of units inthe IC of Eptesicus were not at all spontaneously active,and nearly all cells responded with no more than onespike per stimulus (see below). Figure 2C shows therange of response thresholds at di�erent BFs. The solidcurve in this graph traces the pure-tone behavioral au-diogram of E. fuscus (Dalland 1965; Koay et al. 1997).At BF the thresholds for single units in the IC weredistributed upward from the bat's minimum hearingsensitivity at each frequency throughout the whole rangeof ultrasonic hearing of the animal.
Response thresholds associated with speci®c BFs alsowere measured for FM sounds sweeping through theseBFs. In general, single units responded at a higherprobability if the FM sweep's center frequency, whichwas the frequency of greatest energy within the envelope,matched the BF determined from tone bursts. However,thresholds to FM signals were usually 5±20 dB higherthan those to tone bursts at BF. Since these FM signalswere of the same duration (2 ms) as the tone bursts,higher thresholds to the FM sounds may be attributed tolower e�ective stimulus energy due to the short dwelltime of the sweep in the neighborhood of BF. That is tosay, the BF is present within the FM sweep for only ashort segment of the sweep's 2-ms duration compared toa tone burst, which dwells at a single frequency for thefull 2-ms duration.
Sharpness of tuning
The frequency selectivity of single units in the IC wasdetermined by plotting the strength of responses (typi-cally number of spikes accumulated in PSTH for 40stimulus presentations) evoked by di�erent frequenciespresented at an amplitude 10 dB above the threshold atBF and then measuring the width of the resulting re-sponse area at 50% of its greatest height (maximumresponse strength at BF). The value for this ``50% tun-ing width'' (usually rounded to nearest kHz) is an ab-solute measure for sharpness of tuning which we preferto the inverted relative index, Q10dB. Figure 2D showsthe 50% tuning widths for di�erent units at di�erentBFs. The majority of cells had relatively narrow fre-quency-tuning curves (50% tuning width less than halfthe value of BF). Ninety-six percent (n � 170=177) oftested units had 50% tuning widths of 10 kHz or less(mostly at BFs of 20±50 kHz), and 65.3% of those cells(n � 111=170) had 50% tuning widths of 5 kHz or less.At each BF, the distribution of tuning widths is con-
Fig. 2A±G Graphs showing density distribution of best frequencies(BFs) for single units in the inferior colliculus (IC) ofEptesicus (A) atdi�erent recording depths (B), together with BF dependence ofthreshold (C), sharpness of tuning at BF (D), on-response latency (E),and latency variability (standard deviation, SD) (F, G)
69
tinuous and dense from about 1 kHz to 8 kHz, with ascattering of broader tuning widths up to 14±16 kHz.Most of the tuning widths in Fig. 2D are in the range of2±8 kHz at BFs of 20±50 kHz, which are approximatelyequivalent to Q10dB values of 2.5±25, and the narrowesttuning widths are equivalent to Q10dB values as high as40±45 (see Casseday and Covey 1992).
Amplitude e�ects
The shapes of amplitude response curves or input-out-put functions for IC units were usually nonmonotonic(90.3%; n � 102=113). Most often the discharge proba-bility of each unit would increase linearly with increasingstimulus amplitude plotted in decibels until it saturatedat high amplitudes and thereafter remained ®xed(49.6%; n � 56=113). The dynamic range from thresholdto saturation was usually from 15 dB to 30 dB, butcould be as high as 50 dB in some cells. Approximately19% (n � 18=93) of cells tested for this particular featurehad nonmonotonic input-output curves which peaked ata particular amplitude and then gradually declined to-ward baseline with increasing amplitude beyond thispeak. Fully unsaturated monotonic input-output curves,in which discharge rate increased smoothly and nearlylinearly over the total range of stimulus amplitudes, werefound for only a few cells (9.7%; n � 11=113). In dif-ferent cells the range of ``linearity'' in these responsesvaried from 30 dB to 80 dB. It is likely that responses offully monotonic cells would saturate at a higher ampli-tude than the maximum output of our acoustic deliverysystem (approximately 95 dB SPL peak to peak), butsounds of this strength do not usually stimulate the bat'sears during echolocation (Henson 1970; Kick and Sim-mons 1984).
Temporal organization of responses
Temporal response patterns
The overwhelming proportion of neurons we recordedfrom Eptesicus responded with an average of just onespike for each presentation of a tone burst at BF or adownward FM sweep that encompassed BF. Ninety-onepercent (n � 209=230) of IC units responded only onceper stimulus presentation for 2-ms FM bursts. Whenstimulated by 2-ms tone bursts, only 7% (n � 16=230)responded with more than one spike ± usually in achopper-like pattern, discharging several spikes in aregular periodic sequence to each stimulus presentation.The interspike intervals of the oscillatory dischargesmaking up chopper-like responses were from 1 ms to2 ms. These usually consisted of about two to four dis-charges. Even these chopper units nevertheless re-sponded with only one to two spikes per stimulus for2-ms FM bursts rather than with the chopper patternitself. Consequently, for the majority of single units,
basic properties of the responses can be subsumed bystatistics associated with measurement of BF (shown inFig. 2) and response latency (shown in Fig. 3) since allcells had a unique BF (Fig. 2A, D) and nearly always aunique latency to each presentation of the stimulus.
Response latencies
Latencies were determined either from the slope of thecumulative PSTH (latency distribution in Fig. 3A), ordetermined directly from the mean time-of-occurrence ofthe ®rst spike (latency distribution in Fig. 3B). Latenciesfor on-responses of IC single units extended from theminimum of 3±4 ms required to ``insert'' inputs into theIC from lower auditory centers (Haplea et al. 1994; Suga1969) to a maximum of almost 50 ms. The PSTH slope-based distribution in Fig. 3A has a range of latenciesfrom 3.6 ms to 47.6 ms (n � 217), with a mean of16.4 ms and a standard deviation of 7.5 ms. The spike-time distribution in Fig. 3B has a range of latencies from3.6 ms to 46.4 ms (n � 181), with a mean of 18.9 ms and
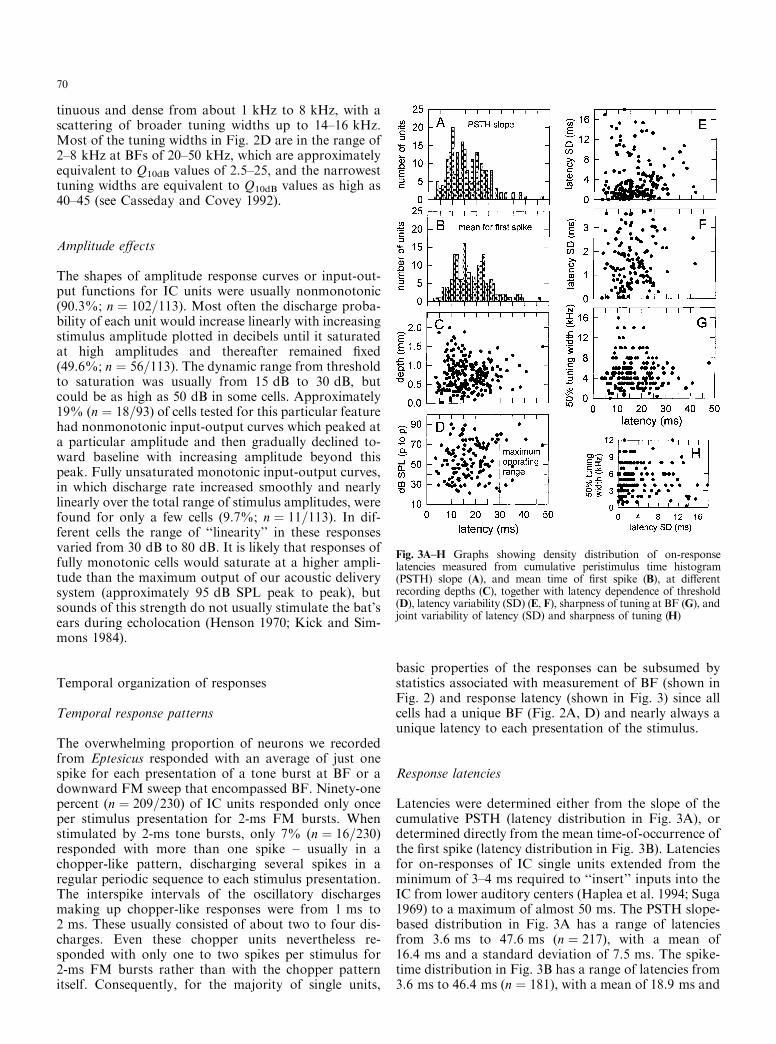
Fig. 3A±H Graphs showing density distribution of on-responselatencies measured from cumulative peristimulus time histogram(PSTH) slope (A), and mean time of ®rst spike (B), at di�erentrecording depths (C), together with latency dependence of threshold(D), latency variability (SD) (E, F), sharpness of tuning at BF (G), andjoint variability of latency (SD) and sharpness of tuning (H)
70

a standard deviation of 7.7 ms. These two di�erent cri-teria for specifying the latency of on-responses yieldeddistributions whose means are signi®cantly di�erent(Student's t-test, t � 3:216, P < 0:05). All latency valuesfor the small multiunit clusters were determined with theslope-detection algorithm (Fig. 3A). For single units thismethod yields virtually the same latency as the mean-spike-time method (Fig. 3B), but for multiunits thisprocedure reports only the shortest-latency unit in thecluster and thus should be biased to report shorter val-ues on the whole, which in fact it did.
Latencies at di�erent frequencies
Figure 2E shows the spread of on-response latencies atdi�erent BFs from 10 kHz to 100 kHz (latencies deter-mined by the slope-detection method). Cells with iden-tical BFs have a wide range of latencies as well as a widerange of tuning widths (Fig. 2D). At BFs correspondingto ®rst harmonic frequencies of 20±45 kHz, the latenciesin Fig. 2E extend densely from 4±5 ms to 25±30 ms,with a scattering of longer latencies out to 40±50 ms. Athigher frequencies of 50±90 kHz latencies are mostlyfrom 5 ms to 10±15 ms, with a few as long as 20±30 ms.Aside from a broader span of latencies past 25±30 ms inour data, the observation that on-response latencies aredispersed in general, and dispersed more widely at lowerfrequencies, is similar to what has been reported earlierfor Eptesicus (Haplea et al. 1994; Jen and Schlegel 1982).
Latencies at di�erent recording depths
Figure 3 shows relations between response latency andother parameters of the single unit responses. Figure 3Cshows the distribution of response latencies at di�erentrecording depths in the IC. In general, latencies from 6or 7 ms to over 25±30 ms were encountered at all depthsfrom 0 mm to 1.5 mm, with a truncation of the range oflatencies to shorter maximum latencies of about 15 msat the deepest locations of 1.5 mm to 2 mm, and ex-tension of the range to slightly shorter latencies of 4±5 ms at shallower sites of 0±0.6 mm. The distribution ofresponse latencies in Fig. 3C is wider in the dorsallaminae of the IC, the locations of cells tuned to lowBFs, than in the most ventral laminae, which containcells with the highest BFs, but the breadth of the latencydistribution nevertheless remains great over intermediatedepths in the IC, too. Beyond this crudest of e�ects,there appears to be no topographic organization ofsingle unit latencies by depth in the IC; just about alllatencies can be found at any depth. BF is related torecording depth by the tonotopic organization of the IC(Fig. 2B), and the distribution of latencies depends uponBF (Fig. 2E). However, the distribution of BF withdepth has appreciable scatter, with BFs from 20 kHz to80 kHz likely to be encountered at depths anywherefrom 0.25 mm to 2 mm (Fig. 2B). Given the breadth of
the distributions for both BF with depth and BF withlatency, it is not surprising that the distribution oflatency with depth is so broad.
Thresholds for responses at di�erent latencies
Figure 3D shows the threshold measured at each unit'sBF (from Fig. 2C) and the response latency obtained ata level 10 dB above this threshold. The dense distribu-tion of latencies from 5 ms to over 25 ms (Fig. 3A, B)uniformly covers all threshold levels from 20 dB SPL to80 dB SPL in Fig. 3D. These thresholds correspond tothe range of sound pressures either for the broadcastsound as it reaches the bat's ears directly through the airfrom the mouth or for echoes which stimulate the bat'sears during echolocation (Dear et al. 1993a; Henson1970; Kick and Simmons 1984; Suga and Schlegel 1972).Responses either to emitted sounds at their directly re-ceived amplitudes or to echoes at any realistic strengthoccur in the IC at all times from about 4±5 ms to25±30 ms after reception of the emission or the echo atthe ear. The vertical dashed line in Fig. 3D shows thetime after the stimulus when echoes arrive from targetsat a distance of 5 m, which is approximately the maxi-mum operating range of echolocation in Eptesicus (Kick1982). This line marks the end of the period of timewhen responses occur frequently at all thresholds.
Variability of response latency
Figure 3E shows the mean absolute latency and the la-tency variability (standard deviation, SD) of the ®rstspike in the PSTHs for di�erent single units (meanSD � 4:31 ms, n � 189; latencies measured with bin-widths of 50 ls.) Latency SD is an index of the temporaljitter inherent in the registration of the time-of-occur-rence of the stimulus by each cell (see Pollak et al.1977c). Figure 3F magni®es the broad, continuousspread of variabilities in the densest part of the distri-bution in Fig. 3E. Latency variabilities as small as50±100 ls are found at all absolute latencies from 4 msto over 25 ms, and a fairly dense distribution of di�erentvariabilities from 50 ls to 100 ls to 2±4 ms is found atall these absolute latencies. A wide scatter of even highervariabilities is also found over this range of absolutelatencies. Approximately 46% (n � 88) of combinedsingle and multiple units had an SD below 2 ms, and18% (n � 16) of those units had an SD below 500 ls.The most surprising result is that the size of the latencyvariability is not related to absolute latency; in particu-lar, narrow variability is not restricted to short latenciesbut is prevalent at all latencies of biological relevance fortarget ranging by Eptesicus (out to about 25±30 ms).Remarkably, there are responses at latencies of20±35 ms (approximating echo delays near the bat'smaximum sonar operating range) that vary in time by only50±100 ls from one stimulus presentation to the next.
71

Response latencies for tone bursts and FM sweeps
A total of 63 single units were tested both with 2-ms tonebursts to determine their BFs and characteristic latencies(mean ®rst spike latency) and then tested with 2-ms FMsweeps (hyperbolic downward FM sweeps over range of20 kHz containing the BFs of individual cells) to de-termine FM latency. Figure 4A shows the frequencyrange for each 20-kHz wide FM stimulus used in thisparticular series of experiments in relation to the BF ofeach unit. The upper and lower ®lled circles mark thestarting and ending frequencies of individual FMsweeps, while each open circle marks the BF value forthe unit associated with that particular sweep. Sweeptime thus moves from top to bottom in Fig. 4A. Thesweep frequencies shown in Fig. 4A were chosen quicklywhile recording was in progress to place the BFs ofdi�erent units at a variety of locations along the dura-tion (2 ms) of the FM sweep.
Figure 4B shows latencies for responses of these 63single units to the two di�erent stimuli; 2-ms tonebursts at BF and 10 dB above threshold (®lled circles),and 2-ms downward 20-kHz wide hyperbolic FM sweeps(open circles). BF latencies and FM latencies are highlycorrelated (r � 0:91). Both in terms of BFs and in termsof latencies at BF; the cells in Fig. 4B are representativeof our data set as a whole (see Fig. 2E). The units shownin Fig. 4B have BFs from 15 kHz to 85 kHz (seeFig. 2A), with response latencies distributed denselyfrom 5 ms to 25 ms, and scatter out to 30±45 ms (seeFig. 3A±B). The latencies of responses to 2-ms FMsweeps are dispersed over the same range as latencies to
tone bursts at BF, and they retain the same pattern ofwider spread of latencies at lower BFs.
Figure 4C (repeated with a somewhat expanded timescale in Fig. 4D) is a histogram showing di�erences be-tween BF and FM latency for these units, or the latencyof responses to the FM sound relative to its onset time at0 ms, with the various absolute latencies associated withstimulation at BF removed (mean FM-BF latency dif-ference � 1.22 ms, SD � 3.40 ms, n � 63). This sub-traction make use of the absolute latency of responses totone bursts at BF to ``calibrate'' the FM latencies, re-moving the large component of ``brain'' latency attrib-utable to various kinds of neuronal propagation delaysand other circuit delays involved in creating the latencydispersion that characterizes IC responses generally (seeDiscussion). The histogram in Fig. 4C and D shows thatmost neurons respond within a time window about2±3 ms wide (relative to FM sweep onset), which re-sembles the envelope of the FM signal. However, inFig. 4C and D no regard is taken for where each BF islocated in the sweep. When the location of each BFalong the sweep is taken into account by ``dechirping''the sweep to provide a BF sweep-time reference for la-tency instead of sweep onset, the response distributionchanges in shape. Figure 4E shows these same FM-BFlatency di�erences corrected for the small 0±2 ms timedelay for each cell's BF in the sweep. (Fig. 4E aligns thetime-of-occurrence of responses with the time-of-occur-rence of individual BFs in the FM sweeps.) The broad,smooth histogram resembling the envelope of the 2-msFM sounds in Fig. 4C and D is transformed into ahistogram which contains a sharply de®ned, narrowpeak located at about 0.2 ms in Fig. 4E. This narrowpeak is caused by a subset of the neurons (n � 15=63 or24%) discharging very close to the relative time-of-occurrence of their BFs in the FM sweeps. In associationwith this peak, no responses at all occur in a narrowwindow of time about 0.2 ms prior to the occurrence ofBF in the sweep. Moreover, the shape of the histogramin Fig. 4E is not otherwise smooth or especially simple ±the large peak at 0.2 ms is surrounded by a structure ofmultiple peaks.
Discussion
Complexity of frequency-time representationin IC responses
Single units in our data from the IC of Eptesicus areunambiguously tuned to a speci®c frequency region (BF;Fig. 2A) with di�erent degrees of sharpness of tuning(50% tuning width; Fig. 2D). The majority of cells alsorespond at a speci®c, unambiguous latency (Fig. 3A, B)with di�erent degrees of variability, or temporal preci-sion (Fig. 3E, F). Cells thus register the time-of-occur-rence of their BFs, but they di�er in the time at whichthey express this response, collectively spreading re-sponses over a span of approximately 25±30 ms after the
Fig. 4 A Starting and ending frequencies in 2-ms FM sweeps (®lledcircles) used as stimuli to compare FM latencies to latencies for tonebursts at BF (open circles). (B) On-response latencies of 63 single unitsfor tone bursts at BF (®lled circles) and for FM sweeps in A (opencircles). C, D Histogram of latency di�erences referred to sweep onsettime = 0 ms (FM±BF latencies from B). E Histogram of latencydi�erences after correction for time-of-BF in sweep, or ``dechirping,''referred to BF time-in-sweep = 0 ms
72

stimulus takes place at the ear (Figs. 2E, 4B). Theseresults are entirely consistent with results from previousstudies of the IC in Eptesicus (Haplea et al. 1994; Jenand Schlegel 1982; Poon et al. 1990), and they also aresimilar to results from other species of FM bats(Bodenhamer and Pollak 1981; Pollak et al. 1977a, b, c;Suga 1969; Vater and Schlegel 1979), but we observed asomewhat broader span of latencies overall.
The predominance of phasic on-responses, eachtuned to a speci®c BF at a speci®c latency, means thatthe primary mode of representation for short-durationwideband sounds in the IC of Eptesicus mixes the timedomain and the frequency domain together (Boden-hamer and Pollak 1981; Casseday and Covey 1995; Suga1972, 1973). This is equivalent to a spectrogram-likerepresentation made up of ``points'' in time-frequencyspace de®ned by individual neural discharges. The fre-quency dimension of these ``spectrograms'' is encoded interms of which neurons produce their discharges to eachfrequency, but the time dimension of these spectrogramsremains time itself ± when the discharges occur. It is truethat responses of IC neurons are selective for di�erenttemporal features or patterns in sounds or trains ofsounds, e.g., duration, repetition rate, AM or FMmodulation rate, intersound delay (Casseday et al. 1994,1997; Condon et al. 1994; Friend et al. 1966; Grinnell1963; Jen et al. 1993; Moriyama et al. 1994; Pinheiro etal. 1991; Pollak et al. 1977a; Saitoh and Suga 1995; Suga1965, 1969, 1972, 1973; Suga and Schlegel 1973; We-nstrup 1995), as well as being selective to di�erent BFs;however, this temporal selectivity is coarse and the time-to-place transformation it represents is not complete. Inother words, rate-code tuning curves for such temporalfeatures as repetition rate, duration, or intersound in-terval are relatively broad, while the latency variabilityof spikes evoked by these temporal patterns still can benarrow. This means that the timing of responses retainsinformation about the ®ne temporal structure ofbroadband waveforms even though individual cells arerate-tuned to di�erent temporal patterns. In Eptesicus,tuned values of temporal features correspond rather wellwith biologically relevant modulation rates of 5±170 Hz(Casseday et al. 1994, 1997), and segregation of re-sponses into di�erent cells ± a ``place'' code according tothese features ± probably facilitates control of motorresponses (Casseday and Covey 1995, 1996), but it doesnot follow that bats perceive such acoustic features asrepetition rate, duration, or delay exclusively because aspeci®c subset of temporal-tuned cells is activated. (It isquestionable why a bat might need to hear repetitionrate or duration in the context of echolocation because itnecessarily has already determined what these parame-ters are to be in the course of producing its sonarsounds.) Instead, the presence of temporal pattern tun-ing may serve to segregate responses into di�erent sub-populations of neurons to facilitate motor control andpossibly to minimize masking and interference ratherthan speci®cally to encode these features for perception.We raise this point because, although temporal patterns
in sounds determine whether a given neuron will re-spond to sounds of a speci®c duration or at a speci®cmodulation rate, the precise timing of the cell's BF [orbetter, excitatory frequency (EF) as de®ned by Bode-nhamer and Pollak (1981)] in a complex sound such asan FM sweep still will be represented by the precisetiming of the on-discharge, not just by which cells re-spond. Temporal pattern tuning in the IC does not ap-pear to completely transpose registration of BF time orEF time in complex signals entirely into a response rateor ``place'' code; it just ®lters information about thetiming of EFs for di�erent temporal patterns into dif-ferent neurons according to gross temporal features suchas duration, repetition rate, or frequency modulation.BF time or EF time itself still appears to be coded bylatency.
FM sweep shape in the BF distribution
The scaling of frequency in the auditory systems ofecholocating bats in relation to the frequency structureof the sonar sounds has proved to be important infor-mation for assessing the underlying mechanisms of echoprocessing (Neuweiler 1990; Pollak and Casseday 1989;Schnitzler and Henson 1980). Di�erent experimentshave now yielded about 840 published values for singleunit BFs in the IC of Eptesicus (Casseday and Covey1992; Jen and Schlegel 1982; Poon et al. 1990; Fig. 2Aabove), a su�cient number of cells to examine frequencyscaling in relation to the detailed frequency compositionof echolocation sounds in this species. With regard tothe bat's sounds, during the bat's approach to a targetFM1 sweeps from about 55±60 kHz down to 22±28 kHzand FM2 sweeps from 95±100 kHz down to 45±55 kHz(Gri�n 1958; Hartley 1992; Kick and Simmons 1984;Simmons 1989). During the terminal stage the frequen-cies slide noticeably lower, with FM1 sweeping from 30or 35 kHz down to 15 kHz and FM2 sweeping from70 kHz down to 30±35 kHz. Except for the terminalstage of interception, FM1 thus consists of frequenciesfrom about 55±60 kHz down to 22±28 kHz and FM2
consists of frequencies from 95±100 kHz down to about45±55 kHz. Speci®c aspects of this frequency and har-monic organization in the bat's sounds appear in the BFvalues for neurons.
The distribution of BFs across the frequency bandused for echolocation (Fig. 2A) shows a strong emphasison frequencies of 25±30 kHz, with progressive loss ofemphasis on higher frequencies from 30 kHz to 80 kHz.Overall, frequencies from 20 kHz to 40 kHz dominatethis distribution. The strong prevalence of BFs in the25±30 kHz region has been termed an overrepresenta-tion of these frequencies, suggesting some special func-tion or need for emphasizing information in this bandcompared to other bands (Casseday and Covey 1992).There is good evidence that a proportion of the cellstuned to 25±30 kHz indeed do serve a special function inecholocation apart from other neurons tuned to these
73

frequencies. When searching for prey in open spaces,Eptesicus broadcasts very shallow FM sounds sweepingfrom 28 kHz to 25 kHz in FM1 (see Simmons 1989),and about 24% of IC units (called ``®lter units'') tunedto frequencies of 22±30 kHz are more sharply tunedthan units found at higher or lower frequencies, pre-sumably to receive echoes of these shallow sweeping``search mode'' signals (Casseday and Covey 1992).However, even if the histogram bars for 20±30 kHz inFig. 2A were made lower by 24% to approximatelyremove the contribution of these demonstrably special®lter units from the distribution of BFs, there still wouldremain an apparent overrepresentation of frequenciesfrom 20 kHz to 40 kHz. This emphasis on some fre-quencies over others may be related to the FM sweeps inthe signals.
Figure 5A illustrates the cumulative proportion ofBFs for a large population of single units pooled fromFig. 2A as well as from three previous studies of Epte-sicus (Casseday and Covey 1992; Jen and Schlegel 1982;Poon et al. 1990). Cells are plotted linearly in descendingorder of BF by assigning each unit a number in thesequence from highest BF to lowest BF (horizontal ``cell
number'' axis in Fig. 5A). The unit with the highest BFin the entire data set is numbered as 1, with the nexthighest numbered as 2, and so forth. This plot showsBFs for over 800 single units tuned to frequencies from20 kHz to 90 kHz. In Fig. 5A, the alignment of datapoints manifestly is not linear with frequency but cur-vilinear, with progressively more cells at each frequencyas frequency decreases over the echolocation band from90 kHz to 22 kHz.
Figure 5B shows a reorganization of the BF data inFig. 5A by plotting the reciprocals of BF as best periodsor BPs (BP�1/BF) for the same units in ascending orderfrom 10 ls to 50 ls. In contrast to BFs in Fig. 5A, thearrangement of data points along the BP scale in Fig. 5Bis more linear. The overall curvilinear relation in Fig. 5Ais greatly attenuated. In particular, the apparent over-representation of lower ultrasonic frequencies around20±40 kHz in Fig. 5A is transformed into a nearly uni-form representation of all ultrasonic periods in Fig. 5B.The approximately linear relation in Fig. 5B arguesagainst overrepresentation of low frequencies being acomplete explanation for the peaked appearance ofhistograms of BFs (as in Fig. 2A). Instead, the BFdensity bias towards lower frequencies could well just bea consequence of the use of a linear period representa-tion for auditory computations in echolocation (Saillantet al. 1993; Simmons et al. 1996). [Note that BF densityis not necessarily the same as tonotopic representation;we make no inference about the cochlear frequency axisbeyond assuming in a general sense that it is probablynot linear with frequency ± see KoÈ ssl and Vater (1995).]The previously identi®ed sharply tuned ``®lter'' unitsat 22±30 kHz (Casseday and Covey 1992) still are anoverrepresentation, however. When their contribution of24% to the BFs at frequencies of 22±30 kHz is removedfrom the BP plot (narrow horizontal bars in Fig. 5B),the adjusted BP plot is even more linear with period.
The break in the slope for the BF curve in Fig. 5A at60±62 kHz is suspiciously close to the frequency boun-dary between FM1 and FM2 in the bat's sonar sounds.The frequencies from 20 kHz to 60 kHz and the fre-quencies from 60 kHz to 100 kHz can be treated sepa-rately in the BF data by numbering the cells from 1 to133 for the total of 133 units tuned above 60 kHz andrenumbering cells from 1 to 657 for the total of 657 unitstuned below 60 kHz (equivalent to subtracting 133 fromeach cell number for cells tuned below 60 kHz). Fig-ure 5C shows the proportion of cells tuned to di�erentBFs above 60 kHz (proportion of 133 for each cellnumber) or below 60 kHz (proportion of 657 for eachcell number) rather than the absolute serial number ofeach cell as in Fig. 5A. When the FM1 and FM2 fre-quency bands are separated as in Fig. 5C, the resultingBF curve appears remarkably like a scaled picture of thespectrogram for the ®rst two harmonics in the bat'ssounds ± a spectrogram derived entirely from physio-logical measurements of BFs in the IC. It thus appearsthat Eptesicus may place an emphasis on sampling dif-ferent ultrasonic frequencies by scaling frequency with a
Fig. 5 A Plot of the serial descending order of BFs in over 800 ICneurons (data pooled from Fig. 2A and from Casseday and Covey(1992); Jen and Schlegel (1982); Poon et al. (1990). B Plot of serialascending order of best periods (BPs) (BP=1/BF) for the same ICneurons as shown in A. Thick horizontal bars show data fromall neurons; thin bars show data with 24 % of neurons tuned to22±30 kHz (``®lter'' cells; Casseday and Covey 1992) removed. Linearregression shown as thin solid lines in A, B. C Plot of serial order in theproportion of neurons tuned to BFs above 60 kHz and neurons tunedto BFs below 60 kHz
74

density according to the relative rate at which the bat'ssonar sounds sweep through these frequencies in eachharmonic. A related model has previously been pro-posed for the relation between biosonar spectrogramsand sharpness of tuning in the bat's auditory-nerve®bers (Menne 1988).
Latency dispersion and delay lines for target ranging
The prevalence of phasic on-responses is a particularlyprominent feature of neural responses in the bat's IC thathas been observed repeatedly (Bodenhamer and Pollak1981; Haplea et al. 1994; Jen and Schlegel 1982; Park andPollak 1993; Pollak et al. 1977b, c; Poon et al. 1990; Suga1964, 1965, 1969, 1970; Vater and Schlegel 1979). Withrespect to earlier studies, the surprising ®nding in thepresent study is that, in the IC of awake Eptesicus, vir-tually all responses to 2-ms downward FM sweeps arephasic; on average one spike occurs at a speci®c latencyin each neuron. The wide dispersion of absolute latenciesis another previously observed, prominent feature ofneural responses in the bat's IC. Figure 6A illustrateslatencies of IC responses in Eptesicus [present data plusthose of Haplea et al. (1994)]. Latency dispersion inFig. 6A is thought to create delay lines for retarding theregistration of the broadcast sonar sound until responsesto echoes can commence (e.g., Berkowitz and Suga 1989;Haplea et al. 1994; Kuwabara and Suga 1993; O'Neill1995; Park and Pollak 1993; Pollak and Casseday 1989;
Saitoh and Suga 1995; Simmons 1980; Simmons et al.1995; Suga 1984, 1990; Suga et al. 1990, 1995; Sullivan1982b). If responses to the outgoing sound are still inprogress when responses to the echo start to occur, ac-tive comparisons between the broadcast and its echoesare made possible. This ``comparison'' is achievedthrough creation of delay-tuned responses in neuronswhose inputs come from both the emitted sound and thereturning echo. Figure 6B illustrates the range of delaytuning or delay sensitivity observed in the auditorycortex of Eptesicus (Dear et al. 1993a; Fritz 1995). De-lay-tuned cells are driven by speci®c combinations ofinput latencies for the emitted sound and for the echo[latency di�erence for strongest response = best delay;e.g., Carr (1993); O'Neill (1995); Sullivan (1982b)]. Thelatencies of on-responses in the IC provide delay-line``taps'' adequate to determine echo delays from a mini-mum of 3±4 ms out to a maximum of 30 ms or more(Fig. 3D), which are equivalent to target ranges from atleast 50 cm out to about 5 m. The broader dispersal oflatencies at lower frequencies results in longer delay linesat these frequencies, which return at still audible am-plitudes from targets at longer ranges due to smalleramounts of atmospheric absorption at lower frequenciesthan higher frequencies (Gri�n 1958; Lawrence andSimmons 1982). In Eptesicus, delay-tuned neurons rep-resenting ``read-out'' of the IC's neuronal delay lineshave been found in the midbrain (Dear and Suga 1995;Feng et al. 1978; Valentine and Moss 1997) and auditorycortex (Dear et al. 1993a,b). This delay line read-out isre¯ected in the distribution of best delays for the pop-ulation of cortical delay-selective cells (Fig. 6B), whichmirrors the span of dispersed latencies which the ICdelivers as inputs (Fig. 6A).
Responses to FM sounds
One question we wish to address with our data setconsisting of BFs and BF and FM latencies is whetherthe value of each cell's latency to a tone burst at BF canbe used to estimate the physiological or ``brain'' latencycaused by neuronal propagation and circuitry. If so,removal of this ``brain'' component of latency from re-sponses to FM stimuli should leave behind the compo-nent of latency due to the timing of BFs in FM sweeps.The underlying assumption is that the excitatory inputsto some cells arrive over the same neuronal path fortone-bursts at BF and for FM sweeps. That is, inhibitorysidebands needed to create FM selectivity (see Suga1972; Casseday and Covey 1992) might prevent re-sponses to adjacent frequencies at speci®c times withoutmaterially perturbing the excitatory latency itself(Bodenhamer and Pollak 1981), but this can only betested by experiment. Figure 4 illustrates this treatmentof response latencies for 63 single units from the IC ofEptesicus. Here, the absolute latency values obtainedfrom on-responses to tone bursts at BF (®lled circles inFig. 4B) are assumed to represent the time required for
Fig. 6 A Latency density distribution from this study and fromHaplea et al. 1994. A horizontal solid line representing most of thelatency dispersion is plotted below the densest portion of thecombined data distribution. B Histogram of best delays (4 ms bins)of delay-tuned cells recorded in the auditory cortex ofEptesicus fuscus(Dear et al. 1993a; Fritz 1995; this study). The solid line from A isreplotted above the delay-tuning histogram
75

responses to propagate through neuronal circuits toreach and excite individual cells in the IC. This physio-logical delay includes both strict travel times and alsoany contribution from neuronal circuits involving in-hibitory and excitatory inputs converging on target cellsat di�erent times (Casseday et al. 1994). Next, latenciesto 2-ms FM sweeps (open circles in Fig. 4B) containingeach cell's BF are assumed to contain both neuronalpropagation delays and also those delays attributable tothe timing of speci®c frequencies within the 2-ms FMwaveform. The di�erence between FM and BF latencies(FM latency ± BF latency from Fig. 4B) should containchie¯y the contribution of the FM structure itself tolatency, shorn of the contribution from the physiologicaldelays associated with each cell's excitatory input at BF.Figure 4C and D shows FM latencies relative to BFlatencies time-locked just to the envelope of the FMstimuli (time zero is onset of sweep in Fig. 4C, D). Thedistribution of latencies in Fig. 4C and D resembles theenvelope of a 2-ms sound, with some additional laggingdispersion chie¯y over a span of an additional millisec-ond or so. When the latencies relative to the onset of theFM sound (FM ± BF latency) in Fig. 4D are addition-ally ``dechirped'' or corrected for the speci®c time atwhich each cell's BF occurred in the FM sweep, thedistribution changes its shape in Fig. 4E. The dechirpedhistogram has a sharp peak near the time of BF (zerotime in Fig. 4E) that is preceded by a narrow intervalwith no responses in any cells. The dechirped histogramas a whole is spread over a span of about 3.5 ms inFig. 4E, but the presence of a sharp discontinuity at itscenter (around zero time) re¯ects transient responsive-ness to BF in the sweep preceded by transient unre-sponsiveness. This is consistent with FM selectivitybeing created by inhibition to frequencies just above BFfollowed by excitation at BF (Suga 1972). The adjust-ment of histogram zero-time for dechirping of FM la-tencies is crudely analogous to reverse correlation ofresponses to speci®c frequencies in FM sweeps, which isrelated to a more sophisticated analysis previously givenfor responses to transient AM events in the IC of adi�erent species of bat (Lesser et al. 1990).
Of course, the neuronal circuits excited by tone burstsand FM sweeps can be di�erent enough to deliver ex-citation at di�erent times as well as at the same time.The wide dispersion of latencies in the IC can be cor-rected to show that some cells respond dynamically atthe time-of-BF in an FM sweep (Fig. 4E), but manyother cells respond to FM sweeps at di�erent times (seehistogram in Fig. 4E at locations away from zero time),and evaluating their responses requires using FM sweepsof di�erent durations (Bodenhamer and Pollak 1981),not comparison of BF and FM latencies. Neithermethod obviates use of the other because they producecomplementary kinds of data. Independent knowledgeof BF o�ers an estimate of the time at which inhibitionfollowed by excitation might be expected to occur withinan FM sweep, whereas data just for FM sweeps withdi�erent durations cannot be used to evaluate the pos-
sibility of inhibition followed by excitation so directly; inthat case, all of the interpretation rests upon responsesto FM stimuli, and responses necessarily cannot occurduring a window of time several hundred microsecondsprior to the occurrence of a spike.
Temporal acuity in neural responses
Previous physiological experiments with other species ofFM bats have led to the conclusion that the acuity forregistration of echo delay by single cells in the IC is inthe region of 50±100 ls at best, with more typical valuesin hundreds of microseconds (e.g., Pollak et al. 1977c).Other studies in which latency measurements were madein Eptesicus found that neurons in some brainstem sites,e.g., cochlear nucleus, nucleus of the lateral lemniscus(Covey and Casseday 1991; Haplea et al. 1994) havelatency variability as small as 20±50 ls, but not smaller.Latencies in the IC have been measured either for tonebursts, which yield latencies at di�erent BFs, or for FMsweeps of di�erent durations produced by an analogswept-frequency generator. With regard to temporalacuity, lack of control over the phase of FM stimuliproduced by an analog system makes it di�cult toconclude that the observed variability of FM latencyrepresents the intrinsic limit for coding of the time-of-occurrence of speci®c frequencies in an FM sweep. Be-haviorally, Eptesicus detects echo phase shifts (Menneet al. 1989; Moss and Simmons 1993) and perceives echophase shifts as corresponding shifts in echo delay(Simmons et al. 1990a). One possible means for encod-ing echo phase is in terms of latencies across di�erentfrequencies. Changes in stimulus phase over successivepresentations of FM sounds due to generator phase driftwould shift neural responses by 10±50 ls over one fullphase rotation, assuming that these responses really arelocked to instantaneous sweep frequency, as is indicatedfor one species of FM bat (Bodenhamer and Pollak1981). The size of neural response-latency shifts to beexpected from stimulus phase shifts is suspiciously closeto the usually quoted latency accuracy of about 50 ls.Thus, there is a real possibility the observed latencyvariability is due to stimulus phase drift during experi-ments rather than intrinsic neuronal timing accuracy.The present physiological experiments were carried outpartly to determine whether the intrinsic temporal acuityexhibited in neural responses to phase-stable FM signalswould be similar to previously reported physiologicalresults or similar to the ®ner acuity exhibited behavior-ally. We found that latency variability in the IC corre-sponds to earlier reports ± typically hundreds ofmicroseconds rather than tens of microseconds in mostneurons. Only a few cells in the IC showed latencyvariability as small as 50±100 ls, which still is roughly100 times worse than the most conservative estimates ofbehavioral acuity in the range of 0.5 ls (Moss andSchnitzler 1995) and another factor of ten or more worsethan the ®nest acuity so far measured behaviorally
76

(Simmons et al. 1990a). Assuming that latency vari-ability is independent in di�erent cells (no covariance oflatencies exists across cells), there still appears to be asigni®cant gap between the level of temporal acuityplausibly accounted for by neural responses in singleunits and the temporal acuity shown behaviorally.
It is not self-evident that the performance of an an-imal in a di�cult, near-threshold discrimination taskwith dynamically changing stimuli must be observablein responses of individual neurons taken one at a time; adistributed code with some sort of cross-populationcomputation might be more e�ective for explainingperformance (Suzuki and Suga 1990). This would implycovariance of latencies across di�erent neurons, whichpreliminary physiological data suggests may occur(Simmons et al. 1996). Eptesicus performs at very highlevels (high percentage correct responses) for echo delaychanges of only 0.05±0.5 ls (Simmons 1993; Simmonset al. 1990a; see also Moss and Schnitzler 1995). While itmight be unrealistic to expect that the smallest behav-iorally detected echo-delay changes of less than 0.05 lsshould be observable in responses of single neurons, itdoes not seem unrealistic to expect that changes in echodelay that the bat easily can detect might be manifestedin single-unit responses. As already mentioned, it ispossible that the bat's echo delay accuracy really ispoorer (e.g. 1±10 ls rather than 0.01 ls) because someartifact was present in the behavioral experiments, butno evidence has appeared to identify this artifact. At-tempts to implicate the most likely artifact ± a spectralchange in echoes due to interference ± have only servedto strengthen the case that the bat's temporal acuity is afraction of a microsecond (Simmons 1993). How can wego about determining the physiological substrate forsuch ®ne acuity? Under the assumption that observedlatency variability in di�erent cells is statistically inde-pendent, it does not appear likely that just making moremeasurements of BF and latency in di�erent neuronswill eventually reveal the basis for the bat's performancewhen echo delay changes by 1±10 ls, much less the bat'sstill ®ner echo-delay acuity of a fraction of a micro-second. It seems more fruitful to explore the possibilitythat temporal patterns of responses across di�erentneurons might contain the information used by the batto perceive small changes in delay. The observation thateach neuron only discharges once for each soundpractically requires groups of neurons to cooperate atspeci®c times if information about a sound is to bemade available for perception because each single spikecan only o�er its time-of-occurrence as a variable. Fu-ture physiological experiments might focus on somemeasure of responses in groups of neurons acting to-gether when stimuli change only in phase or small in-tervals of delay.
Acknowledgements These results were reported at Association forResearch in Otolaryngology Winter Research Meetings in 1993and 1994. We thank J.H. Casseday, E. Covey, S.P. Dear, A.M.Simmons, N. Suga, and numerous colleagues at ARO for sugges-
tions about this work. We also thank J.B. Fritz for allowing us topool some data from his Ph.D. thesis into Fig. 6B. The researchwas supported by ONR Grant No. N00014-89-J-3055, NIMHGrant No. MH00521 (RSDA), NSF Grant No. BCS-9216718,NIMH Training Grant No. MH19118, McDonnell-Pew Grant No.T89-01245-023, and DRF funds. Work conducted according toBrown University IACUC animal-care guidelines.
References
Berkowitz A, Suga N (1989) Neural mechanisms of ranging aredi�erent in two species of bats. Hear Res 41: 255±264
Bodenhamer RD, Pollak GD (1981) Time and frequency domainprocessing in the inferior colliculus of echolocating bats. HearRes 5: 317±355
Carr CE (1993) Processing of temporal information in the brain.Annu Rev Neurosci 16: 233±243
Casseday JH, Covey E (1992) Frequency tuning properties ofneurons in the inferior colliculus of an FM bat. J Comp Neurol319: 34±50
Casseday JH, Covey E (1995) mechanisms for analysis of auditorytemporal patterns in the brainstem of echolocating bats. In:Covey E, Hawkins HL, Port RF (eds) Neural representation oftemporal patterns. Plenum Press, New York, pp 25±52
Casseday JH, Covey E (1996) A neuroethological theory of theoperation of the inferior colliculus. Brain Behav Evol 47: 311±336
Casseday JH, Erhlich D, Covey E (1994) Neural tuning for soundduration: role of inhibitory mechanisms in the inferior col-liculus. Science 264: 847±850
Casseday JH, Covey E, Grothe B (1997) Neural selectivity andtuning for sinusoidal frequency modulations in the inferiorcolliculus of the big brown bat, Eptesicus fuscus. J Neurophysiol77: 1595±1605
Condon CJ, White KR, Feng AS (1994) Processing of amplitude-modulated signals that mimic echoes from ¯uttering targets inthe inferior colliculus of the little brown bat, Myotis luci®gus.J Neurophysiol 71: 768±784
Covey E, Casseday JH (1991) The monaural nuclei of the laterallemniscus in an echolocating bat: Parallel pathways for ana-lyzing temporal features of sound. J Neurosci 11: 3456±3470
Dalland J (1965) Hearing sensitivity in bats. Science 150: 1185±1186
Dear SP, Suga N (1995) Delay-tuned neurons in the midbrain ofthe big brown bat. J Neurophysiol 73: 1084±1100
Dear SP, Fritz J, Haresign T, Ferragamo M, Simmons JA (1993a)Tonotopic and functional organization in the auditory cortex ofthe big brown bat, Eptesicus fuscus. J Neurophysiol 70: 1988±2009
Dear SP, Simmons JA, Fritz J (1993b) A possible neuronal basisfor representation of acoustic scenes in auditory cortex of thebig brown bat. Nature 364: 620±623
Feng AS, Simmons JA, Kick SA (1978) Echo detection and target-ranging neurons in the auditory system of the bat, Eptesicusfuscus. Science 202: 645±648
Friend JH, Suga N, Suthers RA (1966) Neural responses in theinferior colliculus of echolocating bats to arti®cial orientationsounds and echoes. J Cell Physiol 67: 319±332
Fritz JB (1995) The neurobiology of echolocation: Insights fromneurophysiological studies of auditory cortex in the big brownbat, Eptesicus fuscus. PhD. Dissertation, Brown University
Gri�n DR (1958) Listening in the dark. Yale University Press,New Haven, Conn. (2nd edn., Cornell University Press, Ithaca,NY, 1986)
Grinnell AD (1963) The neurophysiology of audition in bats:temporal parameters. J Physiol (Lond) 167: 67±96
Haplea S, Covey E, Casseday JH (1994) Frequency tuning andresponse latencies at three levels in the brainstem of the echo-locating bat, Eptesicus fuscus. J Comp Physiol A 174: 671±683
77

Henson OW (1970) The ear and audition. In Wimsatt WA (ed)Biology of bats, vol II. Academic Press, New York, pp 181±263
Hartley DJ (1992) Stabilization of perceived echo amplitudes inecholocating bats. II. The acoustic behaviour of the big brownbat, Eptesicus fuscus, when tracking moving prey. J Acoust SocAm 91: 1133±1149
Jen PHS, Schlegel PA (1982) Auditory physiological properties ofneurons in the inferior colliculus of the big brown bat, Eptesicusfuscus. J Comp Physiol A 147: 351±363
Jen PHS, Hou T, Wu M (1993) Neurons in the inferior colliculus,auditory cortex and pontine nuclei of the FM bat, Eptesicusfuscus respond to pulse repetition rate di�erently. Brain Res613: 152±155
Kalko EKV, Schnitzler H-U (1993) Plasticity in echolocation sig-nals of European pipistrelle bats in search ¯ight: implicationsfor habitat use and prey detection. Behav Ecol Sociobiol 33:415±428
Kick SA (1982) Target detection by the echolocating bat, Eptesicusfuscus. J Comp Physiol A 145: 431±43
Kick SA, Simmons JA (1984) Automatic gain control in the bat'ssonar receiver and the neuroethology of echolocation. J Neu-rosci 4: 2725±2737
Koay G, He�ner HE, He�ner RS (1997) Audiogram of the bigbrown bat (Eptesicus fuscus). Hear Res 105: 202±210
KoÈ ssl M,Vater M (1995) Cochlear structure and function in bats.In: Popper AN, Fay RR (eds) Hearing by bats. Springerhandbook of auditory research. Springer, Berlin HeidelbergNew York, pp 191±234
Kurta A, Baker RH (1990) Eptesicus fuscus. Mamm Species 356: 1±10
Kuwabara N, Suga N (1993) Delay lines and amplitude selectivityare created in subthalamic auditory nuclei: the brachium of theinferior colliculus of the mustached bat. J Neurophysiol 69:1713±1724
Lawrence BD, Simmons JA (1982) measurements of atmosphericattenuation at ultrasonic frequencies and the signi®cance forecholocation by bats. J Acoust Soc Am 71:585±590
Lesser HD, O'Neill WE, Frisina RD, Emerson RC (1990) ON-OFF units in the mustached bat inferior colliculus are selectivefor transients resembling ``acoustic glint'' from ¯uttering insecttargets. Exp Brain Res 82: 137±148
Menne D (1988) Is the structure of bat echolocation calls an ad-aptation to the mammalian hearing system? J Acoust Soc Am83: 2447±2449
Menne D, Kaipf I, Wagner I, Ostwald J, Schnitzler H-U (1989)Range estimation by echolocation in the bat Eptesicus fuscus:trading of phase versus time cues. J Acoust Soc Am 85: 2642±2650
Moriyama T, Hou T, Wu M, Jen PHS (1994) Responses of inferiorcolliular neurons of the FM bat, Eptesicus fuscus, to pulse trainswith varied pulse amplitudes. Hear Res 79: 105±114
Moss CF, Schnitzler H-U (1989) Accuracy of target ranging inecholocating bats: acoustic information processing. J CompPhysiol A 165: 383±393
Moss CF, Schnitzler H-U (1995) Behavioral studies of auditoryinformation processing. In: Popper AN, Fay RR (eds) Hearingby bats. Springer handbook of auditory research. Springer,Berlin Heidelberg New York, pp 87±145
Moss CF, Simmons JA (1993) Acoustic image representation of apoint target in the bat, Eptesicus fuscus: evidence for sensitivityto echo phase in bat sonar. J Acoust Soc Am 93: 1553±1562
Neuweiler G (1990) Auditory adaptations for prey capture inecholocating bats. Physiol Rev 70: 615±641
O'Neill WE (1995) The bat auditory cortex. In: Popper AN, FayRR (eds) Hearing by bats. Springer handbook of auditory re-search. Springer, Berlin Heidelberg New York, pp 416±480
Park TJ, Pollak GD (1993) GABA shapes a topographic organi-zation of response latency in the mustache bat's inferior col-liculus. J Neurosci 13: 5172±5187
Pinheiro AD, Wu M, Jen PHS (1991) Encoding repetition rateand duration in the inferior colliculus of the big brown bat,Eptesicus fuscus. J Comp Physiol A 169: 69±85
Pollak GD (1993) Some comments on the proposed perception ofphase and nanosecond time disparities by echolocating bats.J Comp Physiol A 172: 523±531
Pollak GD, Casseday JH (1989) The neural basis of echolocation inbats. Springer, Berlin Heidelberg New York
Pollak GD, Bodenhamer R, Marsh DS, Souther A (1977a) Re-covery cycles of single neurons in the inferior colliculus of un-anesthetized bats obtained with frequency-modulated andconstant-frequency sounds. J Comp Physiol 120: 215±250
Pollak GD, Marsh D, Bodenhamer R, Souther A (1977b) Echo-detecting characteristics of neurons in inferior colliculus ofunanesthetized bats. Science 196: 675±678
Pollak GD, Marsh DS, Bodenhamer R, Souther A (1977c) Char-acteristics of phasic-on neurons in the inferior colliculus ofunanesthetized bats with observations related to mechanismsfor echo ranging. J Neurophysiol 40: 926±941
Poon PWF, Sun X, Kamada T, Jen PHS (1990) Frequency andspace representation in the inferior colliculus of the FM bat,Eptesicus fuscus. Exp Brain Res 79: 83±91
Rieke F, Warland D, Ruyter van Steveninck R de, Bialek W (1997)Spikes: exploring the neural code. MIT Press, Cambridge, Mass
Saillant PA, Simmons JA, Dear SP, McMullen TA (1993) Acomputational model of echo processing and acoustic imagingin frequency-modulated echolocating bats: the spectrogramcorrelation and transformation receiver. J Acoust Soc Am 94:2691±2712
Saitoh I, Suga N (1995) Long delay lines for ranging are created byinhibition in the inferior colliculus of the mustached bat.J Neurophysiol 74: 1±11
Schnitzler H-U, Henson OW (1980) Performance of airborne ani-mal sonar sytems: I. microchiroptera. In: Busnel RG, Fish JF(eds) Animal sonar systems. Plenum Press, New York, pp 109±181
Schnitzler H-U, Menne D, Hackbarth H (1985) Range deter-mination by measuring time delay in echolocating bats. In:Michelsen A (ed) Time resolution in auditory systems. Springer,Berlin Heidelberg New York, pp 180±204
Simmons JA (1979) Perception of echo phase information in batsonar. Science 207: 1336±1338
Simmons JA (1980) The processing of sonar echoes by bats. In:Busnel R-G, Fish JF (eds) Animal sonar systems. Plenum Press,New York, pp 695±714
Simmons JA (1989) A view of the world through the bat's ear: theformation of acoustic images in echolocation. Cognition 33:155±199
Simmons JA (1993) Evidence for perception of ®ne echo delay andphase by the FM bat, Eptesicus fuscus. J Comp Physiol A 172:533±547
Simmons JA (1996) Formation of perceptual ``objects'' from thetiming of neural responses: target-range images in bat sonar.In: Llina s R, Churchland PS (eds) The mind-brain continuum± sensory processes. MIT Press, Cambridge, Mass, pp 219±250
Simmons JA, Ferragamo M, Moss CF, Stevenson SB, Altes RA(1990a) Discrimination of jittered sonar echoes by the echolo-cating bat, Eptesicus fuscus: the shape of target images inecholocation. J Comp Physiol A 167: 589±616
Simmons JA, Moss CF, Ferragamo M (1990b) Convergence oftemporal and spectral information into acoustic images ofcomplex sonar targets perceived by the echolocating bat,Eptesicus fuscus. J Comp Physiol A 166: 449±470
Simmons JA, Ferragamo MJ, Saillant PA, Haresign T, WottonJM, Dear SP, Lee DN (1995) Auditory dimensions of acousticimages in echolocation. In: Popper AN, Fay RR (eds) Hearingby bats. Springer handbook of auditory research. Springer,Berlin Heidelberg New York, pp 146±190
Simmons JA, Saillant PA, Ferragamo MJ, Haresign T, Dear SP,Fritz J, McMullen TA (1996) Auditory computations for bio-sonar target imaging in bats. In: Hawkins HL, McMullen TA,Popper AN, Fay RR (eds) Auditory computation. Springerhandbook of auditory research. Springer, Berlin HeidelbergNew York, pp 401±468
78

Suga N (1964) Single unit activity in cochlear nucleus andinferior colliculus of echo-locating bats. J Physiol (Lond) 172:449±474
Suga N (1965) Analysis of frequency modulated sounds by audi-tory neurones of echolocating bats. J Physiol (Lond) 181: 26±53
Suga N (1969) Classi®cation of inferior collicular neurones of batsin terms of responses to pure tones, FM sounds, and noisebursts. J Physiol (Lond) 200: 555±574
Suga N (1970) Echo-ranging neurons in the inferior colliculus ofbats. Science 170: 449±452
Suga N (1972) Analysis of information-bearing elements in com-plex sounds by auditory neurons of bats. Audiology 11: 58±72
Suga N (1973) Feature extraction in the auditory system of bats. In:Moller AR (ed) Basic mechanisms in hearing. Academic Press,New York, pp 675±744
Suga N (1984) The extent to which biosonar information is rep-resented in the bat auditory cortex. In: Edelman GM, Gall WE,Cowan WM (eds) Dynamic aspects of neocortical function.Wiley, New York, pp 315±374
Suga N (1990) Cortical computational maps for auditory imaging.Neural Networks 3: 3±21
Suga N, Schlegel P (1973) Coding and processing in the auditorysystems of the FM-signal-producing bats. J Acoust Soc Am54:174±190
Suga N, Olsen JF, Butman JA (1990) Specialized subsystems forprocessing biologically important complex sounds: cross-cor-relation analysis for ranging in the bat's brain. Cold SpringHarbor Symp Quant Biol LV: 585±597
Suga N, Butman JA, Teng H, Yan J, Olsen JF (1995) Neuralprocessing of target-distance information in the mustached bat.
In: Flock A, Ottoson D, Ulfendahl M (eds) Active hearing.Pergamon Press, New York, pp 13±30
Sullivan WE (1982a) Neural representation of target distance inauditory cortex of the echolocating bat Myotis lucifugus.J Neurophysiol 48: 1011±1032
Sullivan WE (1982b) Possible neural mechanisms of target distancecoding in auditory system of the echolocating bat Myotislucifugus. J Neurophysiol 48: 1033±1047
Suzuki M, Suga N (1990) Acuity in ranging based upon neuralresponses in the FM-FM area of the mustached bat. SocNeurosci Abstr 16: 445
Valentine DE, Moss CF (1997) Spatially selective auditory re-sponses in the superior colliculus of the echolocating bat.J Neurosci 17: 1720±1733
Vater M, Schlegel P (1979) Comparative auditory neurophysiologyof the inferior colliculus of two molossid bats, Mollossus aterand Mollossus molossus. J Comp Physiol A 131: 147±160
Wenstrup JJ (1995) The auditory thalamus in bats. In: Popper AN,Fay RR (eds) Hearing by bats. Springer handbook of auditoryresearch. Springer, Berlin Heidelberg New York, pp 368±415
Wenstrup JJ, Fuzessery ZM, Pollak GD (1988) Binaural neurons inthe mustache bat's inferior colliculus. I. Responses of 60 kHzEI units to dichotic sound stimulation. J Neurophysiol 60:1369±1483
Wong D, Shannon SL (1988) Functional zones in the auditorycortex of the echolocating bat,Myotis lucifugus. Brain Res 453:349±352
Wotton JM, Haresign T, Simmons JA (1995) Spatially dependentacoustical cues generated by the external ear of the big brownbat, Eptesicus fuscus. J Acoust Soc Am 98: 1423±1445
79




















