
Salsolinol and dopamine in rat medial basal hypothalamus after chronic ethanol exposure
Upload
independentCategory
view
9download
0
BRAIN RESEARCH
ELSEVIER Brain Research 736 (1996) 227-236
Research report
FOS expression induced by interleukin-1 or acute morphine treatment in the rat hypothalamus is attenuated by chronic exposure to morphine
Sulie L. Chang a, *, Nilesh A. Patel a, Alejandro A. Romero a, James Thompson b, James E. Zadina c
a Department of Biology, Seton Hall University, South Orange, NJ 07079, USA b Department of Microbiology and Immunology, Louisiana State University Medical Center, New Orleans, LA 70119, USA
c VA Medical Center and Tulane Medical School, New Orleans, LA 70146, USA
Accepted 11 June 1996
Abstract
Interleukin-1 (IL-1) is a cytokine involved in a variety of biological activities. It has been hypothesized that the immunomodulatory effects of IL-1 are the result of both direct action on immune cells and indirect action on a regulatory cascade mediated through the hypothalamus. Chronic exposure to substances of abuse, such as morphine, appears to modulate immunoresponsiveness by mechanisms not yet defined. The expression of FOS, the protein product of the c-fos proto-oncogene, has been widely used as an anatomical marker for monitoring neuronal activity. We have previously shown that acute treatment with either morphine or IL-1 induces FOS immunoreactivity in the rat brain, including the paraventricular (PVN) and supraoptic (SON) nuclei of the hypothalamus. In this study, using immunocytochemical staining of FOS, we demonstrate that chronic exposure to morphine attenuates the cellular responsiveness to IL- 1 and to morphine in the PVN and SON, whereas pretreatment with naloxone, an opiate receptor antagonist, does not reverse the effect of IL-1 on FOS expression. The results not only confirm that the PVN and SON are neuroanatomical sites where the actions of both morphine and IL-1 converge, but also indicate that chronic exposure to morphine may desensitize the cellular response involved in hypothalamic functions through an IL-l-dependent pathway.
Keywords: FOS immunoreactivity; Paraventricular nucleus; Supraoptic nucleus; Caudate putamen; Desensitization
1. Introduct ion
Interleukin-1 ( IL-1) i s a 17-kDa polypeptide immunocy- tokine which exists in two forms, IL-lc~ and IL-I[3. These two forms are separate gene products, and have different physical properties, but share 2 2 - 3 0 % amino acid se- quence homology [22]. IL-1 was originally characterized as a product of macrophages [40], secreted in response to an antigen challenge. Considered as a key mediator of the inflammatory response, IL-1 is involved in neutrophilia, inflammatory cell chemotaxis, the proliferation of T-cells, and the augmentation of B-cell and natural kil ler cell function [19-21,49]. All of these actions contribute to a functional immune system which is necessary for the elimination of microbial invaders and neoplastic cells. Recently, IL-1 has been found in many other cell types including those of the central nervous system (CNS)
* Corresponding author. Fax: + 1 (201) 761-9772; E-mail: changsul @ lanmail.shu.edu
[29,48]. Within the CNS, IL-1 acts as a growth factor for glial cells [27], induces analgesia [42], and affects several hypothalamic functions, including fever production, en- hancement of slow wave sleep, reduction of food intake and insulin production, and alterations in neuroendocrine activities [24,30,51]. The most extensively characterized effect of IL-1 on the endocrine system involves the stimu- lation of the hypotha lamic-p i tu i t a ry-adrena l (HPA) axis [3,5,9,23,36,44,50,54]. IL-1 increases corticotropin-releas- ing factor (CRF) and adrenocorticotropic hormone (ACTH) release, which in turn elevates glucocorticoid concentra- tions. These steroids are involved not only with endocrine negative feedback to reduce CRF and ACTH release, but also with the inhibition of IL-1 production [3], and may be a mechanism underlying the functional role of IL-1 in the interactive endocrine and immune regulatory loops [5,44,50,54]. IL-1 has also been reported to affect other components of the neuroendocrine system, including the release of vasopressin and oxytocin [58], activation of oxytocin-producing neurons [12], and inhibition of the
0006-8993/96/$15.00 Copyright © 1996 Elsevier Science B.V. All rights reserved. PH S0006-8993(96)00698- 1

228 S.L. Chang et al./Brain Research 736 (1996) 227 236
hypothalamic-pituitary-gonadal axis [46]. The weight of current evidence indicates that these effects of IL-1 are mediated via the hypothalamus, particularly in the paraven- tricular (PVN) and supraoptic (SON) nuclei.
Opiates are best known for their analgesic effects and their potential for abuse. In the past two decades, however, considerable evidence has accumulated indicating that opi- ates may have detrimental immunomodulatory effects. Morphine treatment in vitro causes a decrease in prolifera- tive responses to mitogens, inhibition of E-rosette forma- tion, and a decrease in macrophage respiratory burst activ- ity [43]. In vivo, exposure to morphine has been shown to attenuate the activities of natural killer cells [53] and the lymphocyte proliferative responses to mitogens [7] to de- crease macrophage number, and to suppress antibody pro- duction [10,55]. Chronic use of morphine appears to de- crease the volume of primary lymphoid tissues [37], and to attenuate leukocyte-endothelial interaction which is a crit- ical indicator of a functional immune system [15].
Opiate-binding sites and receptors have been reported in a variety of immune cells, including thymocytes and T- lymphocytes [4,16,35,52]. Morphine, therefore, can exert its effects on immune responses via direct binding to various immune cells. There is also evidence that mor- phine modulates the immune system indirectly through the CNS [56]. Opiate alkaloids and peptides, for example, are important regulators of endocrine function, acting primar- ily at the level of the hypothalamus to affect most pituitary and adrenal hormones [34] which modulate immune func- tion, as discussed above. Through their common effects on hormones regulated via the hypothalamus, it is possible that interactions between opiates and IL-1 in the CNS can modulate peripheral systems, such as the immune system.
Recently, the expression of FOS, the protein product of the immediate early proto-oncogene c-[os, has been widely used as a neuroanatomical marker in monitoring neuronal activity [41]. At the basal level, FOS expression is usually low or absent. However, FOS expression can be selec- tively induced under a number of circumstances [1 1-13]. FOS is a nuclear protein, as shown by immunocytochemi- cal staining; and positive staining for FOS can be used to locate individual stimulated neurons, regardless of the sources or mechanisms of stimulation. In our previous studies, the induction of FOS immunoreactivity (FOSir) in specific brain nuclei, especially in the hypothalamic PVN and SON, following acute treatment with either morphine or IL-1 suggests that not only are these nuclei activated by each of these two stimuli, but that these nuclei may also mediate the involvement of both morphine and IL-1 with the neuroendocrine-immune axis. We, therefore, hypothe- sized that chronic exposure to morphine may modulate immune functions by causing the attenuation of the cellu- lar responsiveness to IL-1 in the CNS. To test this hypoth- esis, we examined both morphine-induced and IL-l-in- duced expression of FOS in the hypothalamic PVN and SON following chronic exposure to morphine.
2. Materials and methods
2.1. Cannulation surgery
Adult male rats, 225-249 g, were obtained from Harlan Sprague-Dawley (Indianapolis, IN), and housed in a tem- perature-controlled environment with a 12-h light/dark cycle for at least a week prior to any further treatment. The animals were anesthetized with ketamine (80 mg/kg)/xylazine (8 mg/kg) for implantation of a 22- gauge guide cannula into the right lateral ventricle at 0.3 mm posterior to the bregma, 1.3 mm lateral to midline, and 4.5 mm below the skull surface. The cannula was anchored to the skull with two stainless steel screws and cranioplas- tic cement. A dummy cannula (26 gauge) was inserted into the guide cannula, and the animals were allowed to recover from surgery for 7-10 days. During the postoperative period, the dummy cannula was removed and re-inserted once a day to condition the animals to the injection.
2.2. Treatment with morphine and IL-1
At the end of the recovery period, the animals were randomly assigned to receive either two pellets of mor- phine sulfate (75 mg/pellet, National Institute on Drug Abuse, Rockville, MD) or placebo on the first day, and four pellets on the second day, subcutaneously. The two- plus-four regimen was shown to result in a high degree of opiate tolerance and dependence [47]. During the morphine pelleting procedure, the conditioning regimen described above was continued. On the fifth day, the animals were assigned to receive one of the following over a period of 30 s through an internal cannula (26 gauge) connected to a PE-10 tube inserted into the guide cannula: 200 ng of recombinant human interleukin-lc~ (rhIL-lc~) or inter- leukin-l[3 (rhIL-l[3) (Immunex, Seattle, WA), 10 txg of morphine sulfate (National Institute on Drug Abuse, Rockville, MD), or the vehicle, 5 Ixl of 100 mM Tris-C1, pH 7.6, with 1% BSA. After infusion, the internal cannula was left in position for 5 min before re-insertion of the dummy cannula. The length of treatment was 3 h.
2.3. Treatment with naloxone and IL-1
For naloxone pretreatment, after recovery from the can- nulation procedure, the animals were injected subcuta- neously either with naloxone (4 mg/kg, Sigma, St Louis, MO) or saline 5 min prior to the i.c.v, injection of IL-l~x, IL-113, morphine, or BSA control, as described above.
2.4. Animal perfusion and brain preparation
At the end of treatment, the rats were deeply anes- thetized with pentobarbital (0.5 ml, i.p.), and perfused transcardially with 250 ml of isotonic saline, followed by 500 ml of 4% paraformaldehyde in 0.1 M sodium phos-
S.L. Chang et al. / Brain Research 736 (1996) 227-236 229
pha t e bu f f e r (PB, p H 7.4). B r a i n s were r e m o v e d , pos t f ixed
for an add i t iona l 2 h at 4°C and c ryop ro t ec t ed in 3 0 %
sucrose in PB at 4°C unt i l the b ra ins sank. The b ra ins were
t hen f rozen on p o w d e r e d dry ice and s tored at - 8 0 ° C .
2.5. F O S i m m u n o c y t o c h e m i s t r y
C orona l sec t ions (50 Ixm) were cut on a f r eez ing mic ro -
t ome and soaked in PB for at leas t 1 h to r e m o v e the
f ixat ive. T h e t issues were then t rea ted wi th 0 .2% Tr i ton
X - 1 0 0 in PB for 30 rain at r o o m tempera tu re , f o l l owed by
two w a s h e s wi th PB for 15 rain. T h e sec t ions were then
s tored in PB at 4°C unt i l i m m u n o c y t o c h e m i c a l s t a in ing
wi th FO S a n t i s e r u m was pe r fo rmed .
T h e sec t ions were i n c u b a t e d wi th an t i -FOS a n t i s e r u m
( A B - 2 , lot n u m b e r 3922202 , O n c o g e n e Science , U n i o n -
dale, NY), d i lu ted 1:200 in n o r m a l goa t se rum, at 4°C for
40 h wi th gen t le shak ing on an orb i ta l shaker . T he sec t ions
i
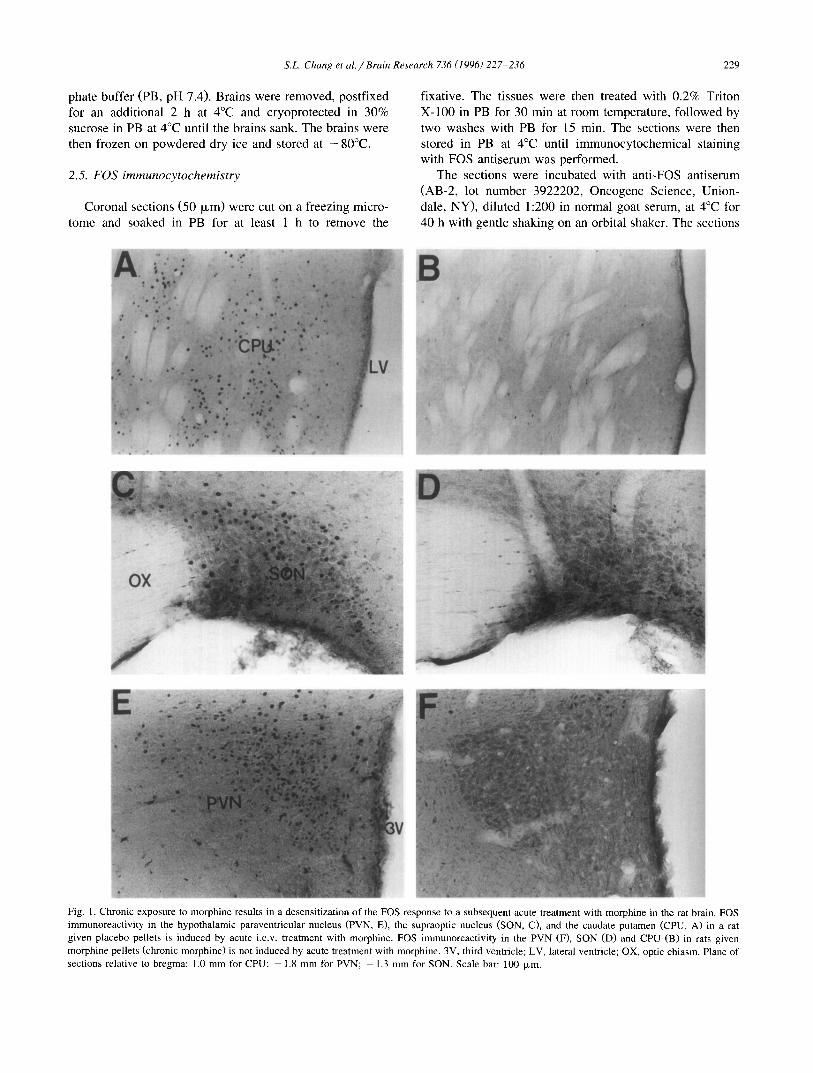
Fig. 1. Chronic exposure to morphine results in a desensitization of the FOS response to a subsequent acute treatment with morphine in the rat brain. FOS immunoreactivity in the hypothalamic paraventricular nucleus (PVN, E), the supraoptic nucleus (SON, C), and the caudate putamen (CPU, A) in a rat given placebo pellets is induced by acute i.c.v, treatment with morphine. FOS immunoreactivity in the PVN (F), SON (D) and CPU (B) in rats given morphine pellets (chronic morphine) is not induced by acute treatment with morphine. 3V, third ventricle; LV, lateral ventricle; OX, optic chiasm. Plane of sections relative to bregma: 1.0 mm for CPU; - 1.8 mm for PVN; - 1.3 mm for SON. Scale bar: 100 txm.
D
Fig.
2.
Chr
onic
exp
osur
e to
mor
phin
e at
tenu
ates
the
acti
vati
on o
f F
OSi
r by
IL
-I i
n th
e hy
poth
alam
ic p
arav
entr
icul
ar nu
cleu
s (P
VN
). F
OS
imm
unor
eact
ivit
y in
the
PV
N i
n ra
ts g
iven
pla
cebo
pel
lets
(A
,B,C
) is
in
duce
d by
an
i.c.v
, in
ject
ion
of I
L- 1
~ (B
) or
IL
- 1 ]3
(C
) as
com
pare
d to
tha
t in
rats
giv
en a
n i.c
.v,
inje
ctio
n of
con
trol
veh
icle
(A
). F
OS
imm
unor
eact
ivit
y in
the
PV
N o
f ra
ts g
iven
mor
phin
e pe
llet
s (D
,E,F
) is
no
t in
duce
d by
an
i.c.v
, in
ject
ion
of I
L-1
e~ (
E)
or I
L-1
[3 (
F)
as c
ompa
red
to t
hat
in r
ats
rece
ivin
g an
i.c
.v,
inje
ctio
n w
ith
cont
rol v
ehic
le (
D).
Abb
revi
atio
ns, p
lane
of
sect
ions
rel
ativ
e to
bre
gma,
and
sca
le b
ar
are
the
sam
e as
tho
se i
n F
ig.
1.
~z
2 e~
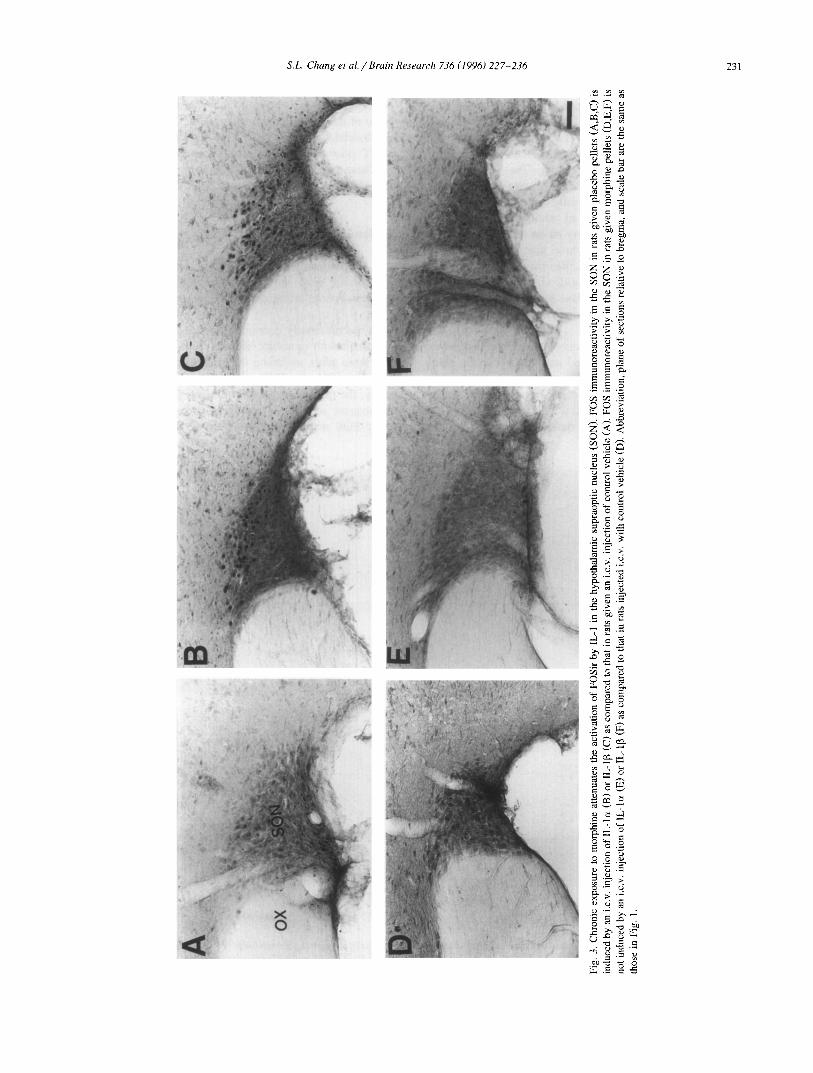
Fig.
3,
Chr
onic
exp
osur
e to
mor
phin
e at
tenu
ates
the
act
ivat
ion
of F
OSi
r by
IL
-1 i
n th
e hy
poth
alam
ic s
upra
opti
c nu
cleu
s (S
ON
). F
OS
imm
unor
eact
ivit
y in
the
SO
N i
n ra
ts g
iven
pla
cebo
pel
lets
(A
,B,C
) is
in
duce
d by
an
i.c.v
, in
ject
ion
of I
L- 1
c~ (
B)
or I
L- 1
13 (C
) as
com
pare
d to
tha
t in
rat
s gi
ven
an i
.c.v
, in
ject
ion
of c
ontr
ol v
ehic
le (
A).
FO
S im
mun
orea
ctiv
ity
in t
he S
ON
in
rats
giv
en m
orph
ine
pelle
ts (
D,E
.F)
is
not
indu
ced
by a
n i,c
.w i
njec
tion
of
IL-I
c~ (E
) or
IL-
1[3
(F)
as c
ompa
red
to t
hat
in r
ats
inje
cted
i.c
.v,
wit
h co
ntro
l ve
hicl
e (D
). A
bbre
viat
ion,
pla
ne o
f se
ctio
ns r
elat
ive
to b
regm
a, a
nd s
cale
bar
are
the
sam
e as
th
ose
in F
ig.
I.
2"
232 S.L. Chang et al. / Brain Research 736 (1996) 227-236
were washed with PB three times for 15 min each, with gentle shaking, at room temperature. The sections were then incubated with biotinylated goat anti-rabbit IgG (Vectastain ABC kit, Vector Laboratories, Burlingame, CA), diluted 1:300 in normal goat serum, for 2 h on an orbital shaker at room temperature. The sections were washed with PB three times for 15 min each at room temperature, then incubated with the av id in -b io t i n -pe r - oxidase complex in PB for an additional 2 h at room temperature with gentle shaking on an orbital shaker. The sections were again washed three times with PB for 15 min each at room temperature, then incubated with 3,3'-di- aminobenzidine tetrahydrochloride (DAB) as the substrate for the av id in -b io t in -pe rox idase reaction for 5 min, fol- lowed by the addition of hydrogen peroxide solution to develop the brown stain.
After FOS staining, the sections were mounted on the slides, air dried, counter-stained with Methyl green, dehy- drated through graded alcohols into Hemo-de (Fisher Sci- entific), and coversl ipped with Permount (Fisher Scien- tific). The FOS-stained cells were counted against the Methyl green-stained cells for quantitation.
Two controls for specificity of immunostaining were performed: (1) incubation in non-immune rabbit serum in place of the anti-FOS anti-sera; and (2) anti-FOS anti-sera preabsorbed with the synthetic M peptide (10 -3 M) for 8 h at 4°C prior to use in the primary incubation.
These procedures were repeated at least 5 times with reproducible results.
3. Results
3.1. Chronic exposure to morphine results in desensitiza-
tion of the FOS immunoreactivi~ (FOSir) response to
acute morphine in the rat brain
It has previously been reported that acute subcutaneous treatment with morphine induces FOS expression in sev- eral brain areas, including the caudate putamen (CPU) [11] and hypothalamic PVN and SON [14]. In the present study, the rats implanted with placebo pellets, followed by subsequent acute i.c.v, treatment with morphine, showed clear FOS induction in the brain. FOSir induced in the
C
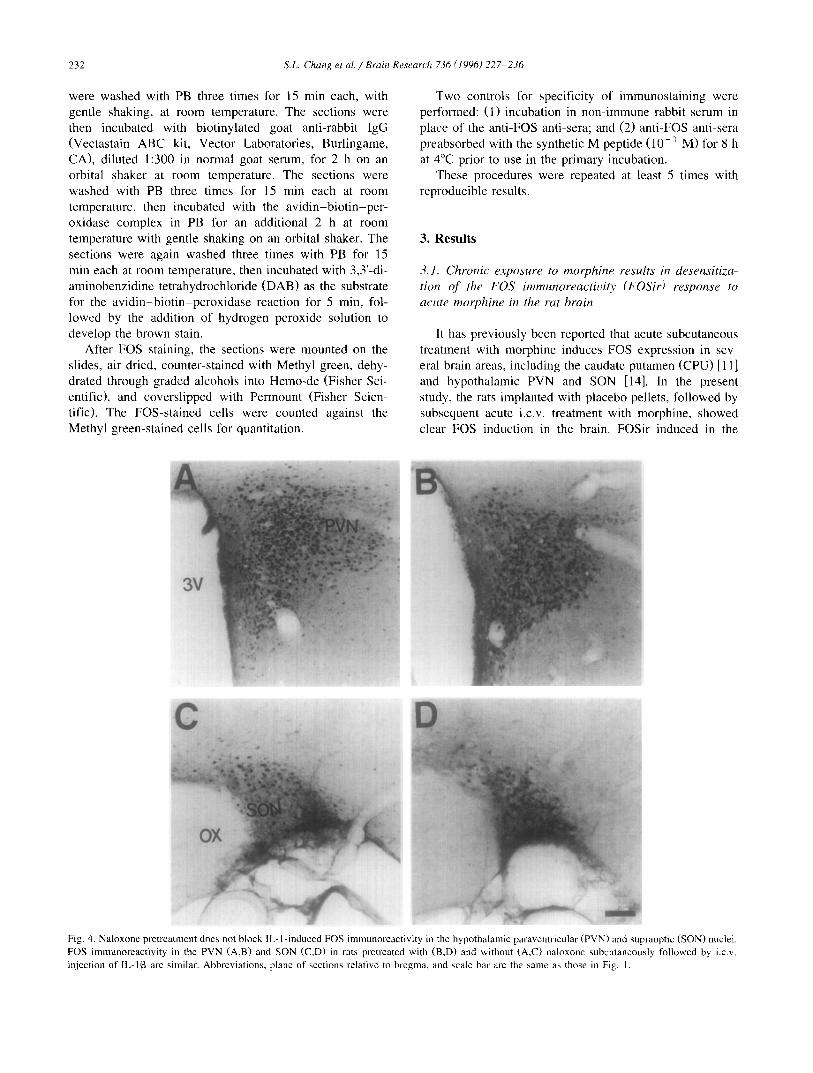
Fig. 4. Naloxone pretreatment does not block IL- l-induced FOS immunoreactivity in the hypothalamic paraventricular (PVN) and supraoptic (SON) nuclei. FOS immunoreactivity in the PVN (A,B) and SON (C,D) in rats pretreated with (B,D) and without (A,C) naloxone subcutaneously followed by i.c.v. injection of IL-113 are similar. Abbreviations, plane of sections relative to bregma, and scale bar are the same as those in Fig. I.

S.L. Chang et a l . / Brain Research 736 (1996) 227-236 233
CPU, PVN and SON (Fig. 1A,C,E) is consistent with previous results [11,14]. Following chronic exposure to morphine, subsequent acute i.c.v, treatment with morphine failed to induce FOSir in these nuclei (Fig. 1B,D,F), indicating that chronic exposure to morphine results in a loss of FOS responsiveness (desensitization) in specific brain areas that are normally responsive to acute morphine treatment.
3.2. Chronic exposure to morphine attenuates the activa- tion of FOSir by IL-1 in the PVN and SON
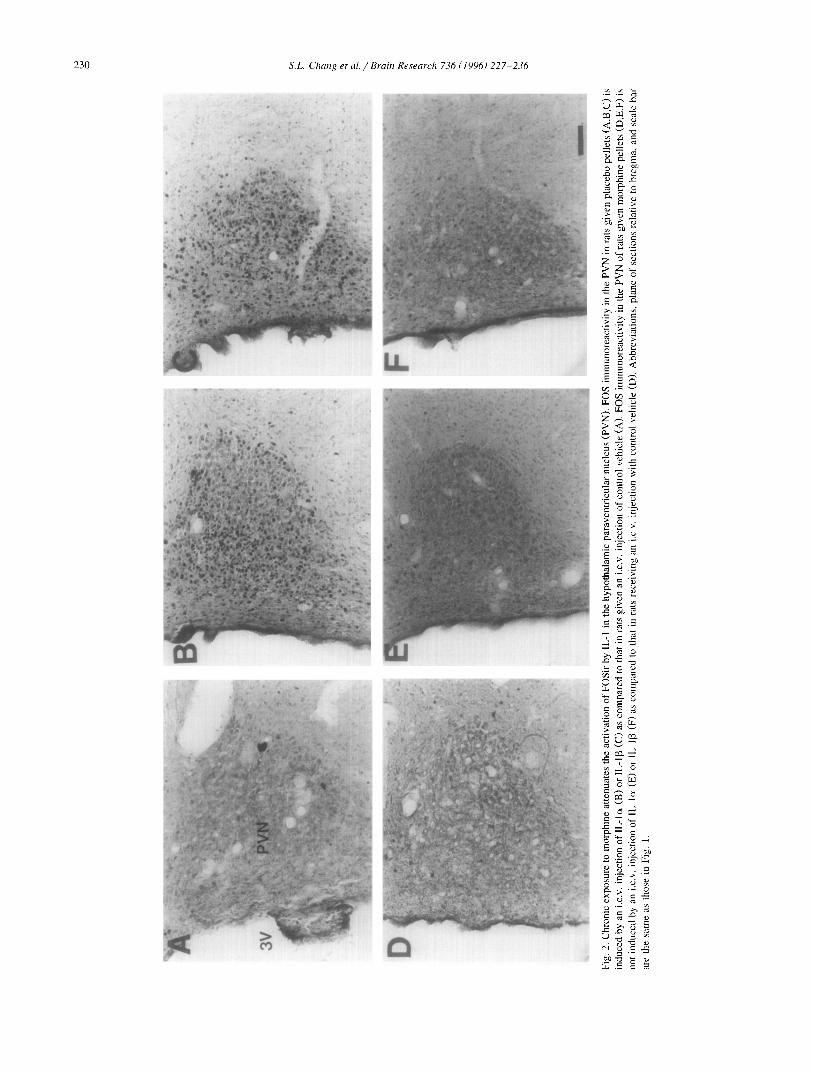
We and others have previously reported that IL-1 treat- ment induces FOS expression in the hypothalamic PVN [12,45] and SON [23] of the rat brain. We now find that following placebo implantation, FOSir was induced by a subsequent i.c.v, treatment of IL-1 (o~ and 13 as shown in Fig. 2B,C and Fig. 3B,C). These data are consistent with the results previously observed with animals given only an acute treatment with IL-1 [12]. In the animals implanted with morphine pellets (chronic treatment), i.c.v, injection of either IL-lcx or IL-I[3 failed to induce FOSir in either the PVN or SON (Fig. 2E,F and Fig. 3E,F). Chronic exposure to morphine appears to prevent the activation of FOSir induced by either acute treatment with morphine or IL-l , as noted above. Virtually no FOSir was seen in the PVN and SON of rats implanted with either morphine or placebo following treatment with control vehicle (Fig. 2A,D and Fig. 3A,D). These results are consistent with those presented in Fig. 1 illustrating the desensitization of FOS responsiveness following chronic exposure to mor- phine.
3.3. Naloxone pretreatment does not block IL-l-induced FOS activation in the PVN and SON
In order to determine whether activation of opiate re- ceptors is involved in the IL-l-induced FOS immuno- reactivity in the PVN and SON, naloxone pretreatment was performed prior to the i.c.v, infusion of IL-lot or IL-I[3. As shown in Fig. 4, FOSir induced by IL-113 was seen in the animals with and without subcutaneous pretreatment with naloxone. Similar results were seen with IL-lo~. By contrast, this regimen of naloxone pretreatment blocks induction of FOSir following i.c.v, injection of morphine (data not shown). Taken together, these data suggest that the IL-l-induced FOSir in the PVN and SON that was seen in this study, as well as in previous studies, is not due to activation of the opiate receptor.
4. Discussion
The c-fos proto-oncogene is one of the immediate early genes that encodes an inducible, nuclear protein that func- tions as a transcriptional regulator, c-fos mRNA expres-
sion, as measured by in situ hybridization, and FOS pro- tein levels, demonstrated by immunocytochemistry, have been widely used as anatomical markers to illustrate the loci of neuronal activity. The induction of FOSir in the hypothalamic PVN and SON following acute treatment with either morphine or IL-1, shown previously and in this study, suggests that not only are the PVN and SON activated by each of these two stimuli, but that these nuclei may also mediate the involvement of both morphine and IL-1 with the neuroendocrine-immune axis. The PVN and SON can, therefore, be considered as neuroanatomical sites where biological actions of morphine and IL-1 con- verge.
Studies have shown the ability of IL-1 to increase the release of CRF [58], to increase the plasma concentrations of ACTH [32], and to induce FOS in the PVN and SON [12,46]. Similarly, acute morphine treatment causes release of these hormones in rats [8,28] and FOS induction in the PVN and SON [14]. Chronic morphine treatment resulted in a desensitization of the response to morphine by FOS (this study), and by CRF and ACTH [8,28]. Taken to- gether, these data raise the possibility of a common path- way, at the molecular level, through which morphine and IL-1 regulate CRF and ACTH. FOS protein may play a critical role in this pathway at the level of gene expression because the promoter region of the precursors for CRF and ACTH (POMC) contain AP-1 sites, the sites to which the FOS-JUN transcription factor complex binds [41].
IL-1 exerts its intracellular effects through specific cell surface receptors. The relative absence of IL-1 receptor expression in the rodent hypothalamus was demonstrated by receptor-ligand binding studies [26], in situ hybridiza- tion [18], and polymerase chain reaction [48]. However, these data are not consistent with the IL-l-induced FOS expression in the PVN and SON reported here and previ- ously by this and other laboratories [12,45]. Several expla- nations are possible for this discrepancy. One possibility is that the PVN may contain very low, but significant, quanti- ties of IL-1 receptors, or may contain as yet unidentified IL-I receptor subtypes. Another possibility is that the initial site of action for IL-1 is outside of the PVN, with afferents into the PVN from brain areas with greater concentrations of both IL-1 and opiate receptors providing critical input leading to the induction of FOS in the PVN.
Early studies reporting IL-l-binding to opiate receptors raised the possibility that morphine and 1L-1 could be interacting through IL-1 binding to the opiate receptor. Purified IL-1 was shown to reduce opioid binding in guinea pig brain [1]. When recombinant human IL-1 (rhIL- 1) was used, however, this observation could not he re- peated [57]. In the present study, pretreatment with the opiate receptor antagonist, naloxone, did not prevent the induction of FOS immunoreactivity by IL-1 in the PVN and SON (Fig. 4). Finally, like IL-I receptors, opiate receptors are sparse in the PVN [38]. Taken together, these results indicate that IL-1 is unlikely to act on opiate

234 S.L. Chang et al. / Brain Research 736 (1996) 227-236
receptors to induce FOS immunoreactivity in the PVN and SON. Down-regulation or desensitization of IL-1 receptors or elements of the IL-I signaling pathway are alternative mechanisms underlying the attenuation of IL-l- induced FOSir by chronic exposure to morphine.
Several laboratories have also shown that IL-1 induces FOS in the PVN after peripheral [6,25,45] administration, and its entry into the brain may be mediated via facilitated transport [2]. Two CNS regions have been proposed as candidates for relaying the stimulus from peripherally ad- ministered IL-1 to the PVN, the organum vasculosum laminae terminalis (OVLT) [33] and the catecholaminergic neurons of the nucleus tractus solitarius (NTS) and ventral medulla [6,25]. It has been suggested that the OVLT functions as a gate for the entry of blood-borne IL-113 into the brain, and a site where the initial biochemical response to IL-113 occurs. Discreet lesions of the ascending cate- cholamine projections from the NTS, a region of dense opiate receptors [39], to the PVN attenuate the FOS re- sponse to peripherally administered IL-1 [25]. Regions such as the NTS may be the initial site of interaction between morphine and IL-1, with manifestation of the interaction occurring at the PVN. It is possible that cen- trally administered IL-1 (as in this study) shares a pathway with peripherally administered IL- 1 for stimulation of FOS in the PVN.
The hippocampus is a brain area currently receiving considerable attention as a potential regulator of the HPA axis [31] because of its well-characterized efferent and afferent connections with the hypothalamus [25]. In addi- tion, the high density of IL-I receptors [18,26] and opiate receptors [39] within the hippocampus suggests that IL-1 and morphine may affect the hypothalamus by activating opiate- and IL-l-sensitive neurons within the hippocam- pus.
Chronic opiate usage has been shown to result in toler- ance and dependence at the biochemical, pharmacological, and behavioral levels [17]. After chronic exposure to opi- old receptor agonists, desensitization, or a loss of sig- nalling responsiveness to that agonist, is observed. Regard- less of the original site(s) of action, this study shows that chronic exposure to morphine culminates in a dramatic attenuation of the effects of IL-1 on FOS induction in areas of the hypothalamus (PVN, SON) critical to neuroen- docrine- immune interactions. In addition, we show that, after chronic exposure to morphine, subsequent acute treat- ment with morphine fails to induce FOSir in brain nuclei, including the PVN. These results indicate that chronic exposure to morphine results in a loss of EOS responsive- ness (desensitization) in specific brain areas that are nor- mally responsive to acute treatment with morphine and to IL-I[3. Chronic exposure to morphine results in both ho- mologous and heterologous desensitization of the FOS response. Loss of responsiveness to the agonist used for the chronic exposure, or to agonists acting at the same receptor is referred to as homologous desensitization. Thus,
the loss of FOS responsiveness to a challenge dose of morphine following chronic morphine treatment demon- strates homologous desensitization of the FOS response. By contrast, diminished responsiveness to agonists that act at different receptors, but whose signal transduction path- ways converge, is known as heterologous desensitization. This type of desensitization is demonstrated by the dra- matic reduction in the capacity of both morphine and IL-l[3 to induce FOSir following chronic exposure to morphine, possibly by an alteration in a common signaling pathway from morphine to FOS and from IL-I[3 to FOS. These results support the concept that the PVN and SON are convergent sites of action for morphine and IL-l[3 in the CNS, and that the interaction between morphine and IL-l[3 is mediated through the regulation of the transcrip- tional factor, FOS.
In summary, the hypothalamic paraventricular and supraoptic nuclei appear to be neuroanatomical sites where the actions of morphine and IL-I converge in the neuroen- docrine-immune axis. Chronic exposure to morphine then desensitizes the cellular response of the neurons within the PVN and SON, as shown by the loss of FOS expression in these nuclei. Since FOS protein functions as a transcrip- tional factor, the desensitization may extend to the level of gene regulation of the critical hypothalamic hormone pre- cursors, such as CRF and POMC. The impairment of the HPA axis following chronic treatment with morphine may be one of the mechanisms underlying the dysfunction of the immune system often seen in opiate addiction.
A c k n o w l e d g e m e n t s
The authors wish to thank Immunex Inc. (Seattle, WA) for the generous gift of recombinant human interleukin-1 o~ and -[3. This work was supported by grants awarded by the National Institute on Drug Abuse and Louisiana State University Neuroscience Center of Excellence, and a VA Merit Review award.
References
[1] Ahmed, M.S., LLanos-Q., J., Dinarello, C.A. and Blatteis, C.M., Interleukin-1 reduces opioid binding in guinea pig brain, Peptides, 6 (1985) 1149-1154.
[2] Banks, W.A., Ortiz, L., Plotkin, S.R. and Kastin, A.J., Human interleukin (IL) lot, murine IL-lc~ and murine 1L-113 are transported from blood to brain in the mouse by a shared saturable mechanism, J. Pharmacol. Exp. Ther., 259 (1991) 988-996.
[3] Bateman, A., Singh, A., Kral, T. and Solomon, S., The immune hy- pothalamic-pituitary-adrenal axis, Endocr. Rec., 10 (1989) 92-98.
[4] Belkowski, S.M., Zhu, J., Liu-Chen, L.-Y., Eisenstein, T.K., Adler, M.A. and Rogers, T.J., Sequence of K-opioid receptor cDNA in the R 1.1 thymoma cell line, J. Neuroimmunol., 62 (1995) 113-117.
[5] Berkenbosch, F., van-Oers, J., del-Rey, A.. Tilders, F. and Bese- dovsky, H., Corticotropin-releasing factor-producing neurons in the rat activated by interleukin-1, Science. 238 (1987) 524-526.

S.L. Chang et al . /Brain Research 736 (1996) 227-236 235
[6] Brady, L.S., Allison, B.L., Herkenham, M. and Gottesfeld, Z., Systemic interleukin-I induces early and late patterns of c-fos mRNA expression in brain, J. Neurosci., 14 (1994) 4951-4964.
[7] Bryant, H.U., Bernton, E.W. and Holaday, J.W., Immunosuppresive effects of chronic morphine treatment in mice, Life Sci., 41 (1987) 1731-1738.
[8] Buckingham, J.C. and Cooper, T.A., Differences in hypothalamic- pituitary-adrenocortical activity in the rat after acute and prolonged treatment with morphine, Neuroendocrinology, 38 (1984) 411-417.
[9] Busbridge, N,J. and Grossman, A.B., Stress and the single cytokine: interleukin modulation of the pituitary-adrenal axis, Mol. Cell Endocrinol., 82 (1991) C209-C214.
[10] Bussiere, J.L., Adler, M.W., Rogers, T.J. and Eisenstein, T.K., Cytokine reversal of morphine-induced suppression of the antibody response, J. Pharmacol. Exp. Ther., 264 (1993) 591-597.
[11] Chang, S.L., Squinto, S.P. and Harlan, R.E., Morphine activation of c-fos expression in the rat brain, Biochem. Biophys. Res. Commun., 157 (1988) 698-704.
[12] Chang, S.L., Ren, T. and Zadina, J.E., Interleukin-I activation of FOS proto-oncogene protein in the rat hypotbalamus, Brain Res., 617 (1993) 123-130.
[13] Chang, S.L., Patel, N.A. and Romero, A.A., Activation and desensi- tization of FOS immunoreactivity in the rat brain following ethanol administration, Brain Res., 679 (1995) 89-98.
[14] Chang, S.L., Kenigs, V., Moldow, R.L. and Zadina, J.E., Chronic treatment with morphine and ethanol, but not cocaine, attenuates IL-113 activation of Fos expression in the rat hypothalamic paraven- tricular nucleus, in B.M. Sharp, T.K. Eisentein, J.J. Madden and H. Friedman (Eds.), The Brain Immune Axis and Substance Abuse, Plenum Press, New York, 1995, pp. 201-208.
[15] Chang, S.L., Bersig, J.M., Wu, G.-D., Patel, N.A., Zadina, J.E. and House, S.D., Chronic exposure to morphine attenuates leukocyte- endothelial interactions in the rat mesentery, N1DA Research Monogr., 162 (1995) 189.
[16] Chuang, L.F., Chuang, T.K., Killam, K.F.J., Chuang, A.J., Kung, H., Yu, L. and Chuang, R.Y., Delta opioid receptor gene expression in lymphocytes, Biochem. Biophys. Res. Commun., 202 (1994) 1291-1299.
[17] Cox, B.M. and Werling, L.L., Opioid tolerance and dependence. In A. Pratt (Ed.), The Biological Bases of Drug Tolerance and Depen- dence, Academic Press, San Diego, 1991, pp. 199-229.
[18] Cunningham, E.T., Jr., Wada, E., Carter, D.B., Tracey, D.E., Battey, J.F. and DeSouza, E.G., In situ bistochemical localization of type I interleukin-1 receptor messenger RNA in the central nervous system, pituitary and adrenal gland of the mouse, J. Neurosci., 12 (1992) 1101-1114.
[19] Dinarello, C.A., Interleukin-I and its biologically related cytokines, Adv. lmmunol., 44 (1989) 153-205.
[20] Dinarello, C.A. and Savage, N., Interleukin-1 and its receptor, Crit. Rev. lmmunol., 9 (1989) 1-20.
[21] Dinarello, C.A., Interleukin-I and interleukin-1 antagonism, Blood, 77 (1991) 1627-1652.
[22] Dower, S.K., Bird, T.A. and Sims, J.E., lnterleukin-l, Adv, Neu- roimmunol., 12 (1992) 1-16.
[23] Dunn, A.J., Interleukin-I as a stimulator of hormone secretion, Prog. Neuroendocrinimmunol., 3 (1990)26-34.
[24] Eizirik, D.L., Welsh, M,, Strandell, E., Welsh, N. and Sandier, S., Interleukin-1 (13) depletes insulin messenger ribonucleic acid and increases the heat shock protein hsp70 in mouse pancreatic islets without impairing the glucose metabolism, Endocrinology, 127 (1990) 2290-2297.
[25] Ericsson, A., Krisztina, J. and Sawchenko, P.E., A functional anatomical analysis of central pathways subserving the effects of interleukin-1 on stress-related neuroendocrine neurons, J. Neurosci., 14 (1994) 897-913.
[26] Farrar, W.L., Kilian, P.L., Ruff, M.R., Hill, J.M. and Pert, C.B,,
Visualization and characterization of interleukin-1 receptors in brain, J. lmmunol., 139 (1987) 459-463.
[27] Friedman, W.J., Larkfors, L., Ayer-Lelievre, C., Ebendal, T., Olson, L. and Persson, H., Regulation of 13-nerve growth factor expression by inflammatory mediators in hippocampal cultures, J. Neurosci. Res., 27 (1990) 374-382.
[28] George, R. and Way, E.L, Studies on the mechanism of pituitary- adrenal activation by morphine, Br. J. Pharmacol. Chemother., 10 (1955) 260-264.
[29] Hopkins, S.J. and Rothwell, N.J., Cytokines and the nervous system I: expression and recognition, Trends Neurosci., 18 (1995) 83-88.
[30] Hughes, J.H., Colca, J.R., Easom, R.A., Turk, J. and McDaniel, M.L., Interleukin-I inhibits insulin secretion from isolated rat pan- creatic islets by a process that requires gene transcription and mRNA translation, J. Clin. Invest., 86 (1990) 856-863.
[31] Jacobson, L. and Sapolsky, R., The role of the hippocampus in feedback regulation of the hypothalamic-pituitary-adrenocortical axis, Endocr. Rev., 12 (1991) 118-134.
[32] Katsuura, G., Gottschall, P.E., Dahl, R. and Arimura, A., Adrenocor- ticotropin release induced by intracerebroventricular injection of recombinant human interleukin-I in rats: possible involvement of prostaglandin, Endocrinology, 122 (1988) 1773-1779.
[33] Katsuura, G., Arimura, A., Koves, K. and Gottschall, P.E., Involve- ment of organum vasculosum of lamina terminalis and preoptic area in interleukin- 113-induced ACTH release, Am. Z Physiol., 258 (1990) E163-EI71.
[34] Kuhn, C.M. and Windh, R.T., Endocrine actions of opiates. In R.R. Watson, (Ed.), Biochemistry and Physiology of Substance Abuse, Vol. 111, CRC Press, Boca Raton, 1989, pp. 247-277.
[35] Lawrence, D,M. and Bidlack, J.M., Kappa opioid binding sites on the RI.I murine lymphoma cell line, J. Neuroimmunol., 41 (1992) 223-230.
[36] Lee, S. and Rivier, C., Hypophysiotropic role and hypothalamic gene expression of corticotropin-releasing factor and vasopressin in rats injected with interleukin-I beta systemically or into the brain ventricles, J. Neuroendocrinol., 6 (1994)217-223.
[37] LeVier, D.G., McCay, J.A., Stern, M.L., Harris, L.S., Page, D. and R.D. Brown, Immunotoxicological profile of morphine sulfate in B6C3FI female mice, Fundam. Appl. Toxicol., 22 (1994) 525-542.
[38] Mansour, A., Khachaturian, H., Lewis, M.E., Akil, H. and Watson, S.J., Autoradiographic differentiation of mu, delta, and kappa opioid receptors in the rat forebrain and midbrain, J. Neurosci., 7 (1987) 2445-2464.
[39] Mansour, A., Fox, C.A., Thompson, R.C., Akil, H. and Watson, S.J., Mu-Opioid receptors mRNA expression in the rat CNS: com- parison to mu-receptor binding, Brain Res., 643 (1994) 245-265.
[40] Martin, M. and Resch, K., Interleukin-l: more than a mediator between leukocytes, Trends Pharmacol. Sci., 9 (1988) 171-177.
[41] Morgan, J.I. and Curran, T., Stimulus-transcription coupling in the nervous system: involvement of the inducible proto-oncogenes los and jun, Annu. Rev. Neurosci., 14 (1991) 421-451.
[42] Nakamura, H., Nakanishi, K., Kita, A. and Kadokawa, T., Inter- leukin-1 induces analgesia in mice by a central action, Eur. J. Pharmacol., 149 (1988) 49-54.
[43] Peterson, P.K., Molitor, T.W., Chao, C.C. and Sharp, B., Opiates and cell-mediated immunity. In R.R. Watson, (Ed.), Drugs of Abuse and Immune Function, CRC Press, Boca Raton, 1990, pp. 2-12.
[44] Rivest, S. and Rivier, C., Influence of the paraventricular nucleus of the hypothalamus in the alteration of neuroendocrine functions induced by intermittent footshock or interleukin, Endocrinology, 129 (1991) 2049-2057.
[45] Rivest, S., Torres, G. and Rivier, C., Differential effects of central and peripheral injection of interleukin-113 on brain c-fos expression and neuroendocrine functions, Brain Res., 587 (1992) 13-23.
[46] Rivest, S. and Rivier, C., lnterleukin-113 inhibits the endogenous expression of the early gene c-los located within the nucleus of

236 S,L. Chang et al. / Brain Research 736 (1996) 227-236
LH-RH neurons and interferes with hypothalamic LH-RH release during proestrus in the rat, Brain Res., 613 (1993) 132-142.
[47] Rothman, R.B., Danks, J.A., Jacobson, A.E., Burke, Jr., T.R.. Tortella, T.E. and Holaday, J,W., Morphine tolerance increases Ix-noncompetitive K-binding sites, Eur. J. Pharmacol., 124 (1986) 113-119.
[48] Rothwell, N.J., Functions and mechanisms of interleukin-I in the brain, Trends Pharmaeol. Sci., 12 (1991) 430-436.
[49] Sauder, D.N. lnterleukin-l, Arch. Dermatol., 125 (1989) 679-682. [50] Sapolsky, R., Rivier, C., Yamamoto, G., Plotsky, P. and Vale, W.,
lnterleukin-1 stimulates the secretion of hypothalamic corticotropin- releasing factor, Science, 238 (1987) 522 524.
[51] Schettini, G., Interleukin-I in the neuroendocrine system: fi-om gene to function, Prog. Neuroendocrinol. Immunol., 3 (1990) 157-166.
[52] Sedqi, M., Roy, S., Ramakrishnan, S., Elde, R. and Lob, H.H., Complementary DNA cloning of a mu-opioid receptor from peri- toneal macrophages, Biochem. Biophys. Res. Commun., 209 (1995) 563-574.
[53] Shavit, Y., Terman, G.W., Lewis, J.W., Zane, C.J., Gale, R.P. and Liebeskind, J.C., Effects of footsbock stress and morphine on natural
killer lymphocytes in rats: studies of tolerance and cross-tolerance, Brain Res., 372 (1986) 382-385.
[54] Uehara, A., Gottschall, P.E., Dahl, R.R. and Arimura, A., Inter- leukin-I stimulates ACTH release by an indirect action which requires endogenous corticotropin releasing factor. Endocrinology, 121 (1987) 1580-1582.
[55] Weber, R.J., Ikejiri, B., Rice, K,C., Pert, A. and Hagan, A.A., Opiate receptor mediated regulation of the immune response in vivo, NlDA Research Monogr., 76 (1987) 341-348.
[56] Weber, R.J. and Pert, A., The periaqueductal gray matter (PAG) mediates opiate-induced immunosuppression, Science. 245 (1989) 188-190.
[57] Wiedermann, C.J., lnterleukin-1 interaction with neuroregulatory systems: selective enhancement by recombinant human and mouse interleukin-I of in vitro opioid peptide receptor binding in rat brain, J. Neurosci. Res., 22 (1988) 172-180.
[58] Yasin, S.A., Costa, A., Forsling, M.L. and Grossman. A., Inter- leukin-113 and interleukin-6 stimulate neurohypophyseal hormone release in vitro, .L Neuroendoerinol., 6 (1994) 179-184,
Copyright © 2022 FDOKUMEN




















