
Global Changes in the Rat Heart Proteome Induced by Prolonged Morphine Treatment and Withdrawal

Available online at www.sciencedirect.com
Drug and Alcohol Dependence 97 (2008) 21–32
Morphine and methadone pre-exposures differently modify brain regionalFos protein expression and locomotor activity responses
to morphine challenge in the rat
Ewa Taracha a,∗, Stanisław J. Chrapusta b, Małgorzata Lehner a, Anna Skorzewska a,Piotr Maciejak a,c, Janusz Szyndler c, Adam Płaznik a,c
a Department of Neurochemistry, Institute of Psychiatry and Neurology, 9 Sobieskiego Street, 02-957 Warsaw, Polandb Department of Experimental Pharmacology, Polish Academy of Sciences Medical Research Center, 5 Pawinskiego Street, 02-106 Warsaw, Polandc Department of Experimental and Clinical Pharmacology, Medical Academy of Warsaw, 26/28 Krakowskie Przedmiescie, 00-927 Warsaw, Poland
Received 4 November 2007; received in revised form 5 March 2008; accepted 8 March 2008Available online 15 May 2008
Abstract
Methadone is commonly used in substitution therapy of heroin addicts; hence, its potential for modifying reactions to opiates is of clinicalimportance. We compared the effects of repeated daily and every-other-day pre-exposure of rats to s.c. morphine and methadone on locomotoractivity and CNS neuronal activation (as assessed by Fos immunohistochemistry) responses to s.c. morphine challenge given 2 weeks after thecompletion of the pretreatment. The challenge revealed behavioral sensitization after daily morphine pretreatment only. Dorsomedial striatumand basolateral amygdaloid nucleus showed robust morphine-induced Fos protein induction that was unaffected by the pretreatments tested.Centrolateral striatum, shell and core of the nucleus accumbens, paraventricular thalamic nucleus and some layers of motor and somatosensorycortices showed but negligible Fos protein induction in drug-naive rats; this response was markedly enhanced by morphine pretreatment only,which effect might be related to the emergence of opiate addiction. Minor Fos responses to morphine were also found in layers IV and VI of thesomatosensory cortex and layer VI of the insular cortex of the drug-naıve rats; these responses were significantly enhanced both by morphineand methadone pretreatment. The similarity of methadone and morphine pretreatments’ effects in the latter cortical regions might be relevant to
the ability of methadone to alleviate signs of abstinence syndrome and craving in heroin addicts. In summary, this study revealed differing andrelatively long-lasting effects of prolonged administration of morphine and methadone on the profile of behavioral and CNS neuronal activationresponses to morphine challenge in the rat.© 2008 Elsevier Ireland Ltd. All rights reserved.K edia
1
imcmeip
a2beDsb
0d
eywords: Morphine; Methadone; Sensitization; Cortex; Striatal complex; Imm
. Introduction
Repeated administration of various psychoactive substances,ncluding opiates, may cause, even after an abstinence period, a
ore robust effect than that of the first dose. This phenomenon,alled sensitization, may play an important role in the develop-ent of drug addiction (Stewart and Badiani, 1993; De Vries
t al., 1998). Notably, � opioid receptor (MOR) agonists cannduce mutual cross-sensitization with other commonly abusedsychoactive substances (Nestby et al., 1997; Vanderschuren et
∗ Corresponding author. Tel.: +48 22 458 2619; fax: +48 22 458 2771.E-mail address: [email protected] (E. Taracha).
BMc1ism
376-8716/$ – see front matter © 2008 Elsevier Ireland Ltd. All rights reserved.oi:10.1016/j.drugalcdep.2008.03.013
te early genes
l., 1997; Cadoni et al., 2001; Ward et al., 2006; He and Grasing,007). There are many reports showing, in experimental animals,ehavioral sensitization from opiates administered with a vari-ty of regimens (e.g. see Vanderschuren et al., 1997; Cadoni andi Chiara, 1999; Trujillo et al., 2004). In particular, it has been
hown that the development of the sensitization is facilitatedy environmental novelty and intermittent dosage (reviewed inadiani and Robinson, 2004; see also Paolone et al., 2007).orphine-induced behavioral sensitization is long-lasting and
an resurface even after months of abstinence (Babbini et al.,
975; Pollock and Kornetsky, 1996). Results of those behav-oral studies are supported by neuronal activation data fromtudies using c-fos mRNA or Fos protein expression in rats andice (Curran et al., 1996; Frankel et al., 1999; D’Este et al.,
2 cohol
2a
aimitmiditalbeirt
itCttFr
2
2
cwihgibdaa
2
(PIsDsbaA(m
2
iin2(otdA4TtLattrdi
oaruitbhtLaop
2
lBlwscwitpi
2
pdtsc(m
2 E. Taracha et al. / Drug and Al
002; Pontieri et al., 1997; Kornetsky, 2004; Ammon-Treibernd Hollt, 2005).
Chronic nature and high probability of relapses of opi-te addiction are common knowledge. Sensitization, whichncreases susceptibility to the relapses, is considered detri-ental with regard to addiction therapy. Thus, one might ask
f methadone, a MOR agonist commonly used in substitu-ion therapy of heroin addicts, induces sensitization, and if
ethadone-aided therapy modifies heroin addicts’ reaction dur-ng relapses. Whereas the clinical utility of methadone is wellocumented (reviewed in Kreek, 2000; Kreek and Vocci, 2002),ts molecular bases have not been fully elucidated. In particular,he potential of methadone for cross-sensitization to morphinend for modifying the sensitization caused by opiate treatment inaboratory rodents has not been thoroughly studied. Comparingehavioral and molecular effects of these drugs is of both sci-ntific interest and potential importance for the development ofmproved therapies. In particular, it would be interesting to see ifepeated methadone treatment can modify subject’s sensitivityo eventual sporadic morphine intake.
The aims of this study were: (i) to set up a regimen for behav-oral sensitization to morphine of Sprague-Dawley rats fromhe stock of the Polish Academy of Sciences Medical Researchenter; (ii) to test the ability of methadone for cross-sensitizing
he rats to morphine; and (iii) to see if pre-exposing the ratso morphine and methadone will differently affect patterns ofos protein expression responses to morphine in selected brainegions.
. Experimental procedures
.1. Animals
Male Sprague-Dawley rats from the Polish Academy of Sciences Medi-al Research Center, Warsaw, initial body weight 176 ± 15 g (mean ± S.D.),ere housed four per cage in opaque plastic cages (56 cm × 35 cm floor size)
n a temperature- and humidity-controlled room (21 ± 2 ◦C, 60–70% relativeumidity) with a 12/12-h light/dark cycle (lights on at 7:00 a.m.), and wereiven standard rodent chow and tap water ad libitum. The rats were acclimatedn the animal house of the Institute of Psychiatry and Neurology for 1 weekefore the experiment (Obernier and Baldwin, 2006) and were subject to 1 minaily handling during this period. All animal use procedures were in compli-nce with the NIH Guide for the Care and Use of Laboratory Animals and wereccepted by the Bioethical Committee of the Medical Academy of Warsaw.
.2. Drugs and chemicals
(R,S)-Methadone hydrochloride was donated by Molteni FarmaceuticiScandicci, Italy), and morphine sulfate was purchased from Polfa (Warsaw,oland). Rabbit polyclonal anti-c-Fos antibody (anti-c-Fos (Ab-2) (4–17) rabbitgG) was from Calbiochem (San Diego, CA, USA); its specificity was verified asuggested by the manufacturer (Calbiochem Data Sheet PP10 Rev. 15-April-04CH). Normal donkey serum (NDS) was purchased from Jackson ImmunoRe-
earch Labs (West Grove, PA, USA), avidin–biotin–peroxidase complex and
iotinylated goat anti-rabbit IgGs – from Vector Labs (Burlingame, CA, USA),nd DePeX mounting medium – from VWR International Ltd., Poole, England.ll other reagents were of analytical-grade and were purchased from ChemPurPiekary Slaskie, Poland), POCh (Gliwice, Poland), Serva (Heidelberg, Ger-any), and Merck (Darmstadt, Germany).
p0fdm
Dependence 97 (2008) 21–32
.3. Experimental design
Rats were divided between seven experimental groups and were given s.c.njections of saline, morphine and methadone according to the schedules shownn Table 1. Since the development of sensitization is facilitated by environmentalovelty (Ferguson et al., 2004; Badiani and Robinson, 2004; Paolone et al.,007), the rats were transferred (in their home cages) to locomotor activityLA) testing room 30 min before each injection. The term ‘novelty’ is used inperational sense here to designate unique environment that is different fromhat of the home cage and does not imply cognitive novelty lasting for theuration of the experiment (see also Ferguson et al., 2004; Paolone et al., 2007).fter the injections, the rats were placed in another set of clear plastic cages0 cm × 25 cm floor size (post-injection or ‘novelty’ cages, four rats to a cage).he rats were either left undisturbed in those cages for 2 h, or were transferred
o LA testing chambers (see Section 2.4) 30 min later, subjected to a 55-minA test (after the 1st, 4th and 7th pretreatment dose if injected every-other-day,fter the 1st, 7th and 14th dose – if injected for 6 days a week (‘daily’), and afterhe challenge – irrespective of the pretreatment schedule) and then returned tohe post-injection cages for additional 35 min. Two hours after the injections allats were returned to their home cages in the animal house, except for the lastay of the experiment when they were sacrificed for Fos immunohistochemistrynstead.
Morphine doses for the daily pretreatment and challenge were chosen basedn earlier reports (Vanderschuren et al., 2001; Kalinichev et al., 2004; Taracha etl., 2006). Equivalent doses of methadone were estimated based on data on self-eported daily intake of ‘street’ heroin preparations and oral methadone dosessed for heroin addicts’ therapy in the Institute of Psychiatry and Neurologyn Warsaw (Taracha et al., 2005). When making the estimates we consideredhe uncertainty regarding the make-up of the heroin concoctions, poor relia-ility of the self-reports, oral methadone’s bioavailability, relative potencies oferoin and morphine, and, last but not least, differing biological half-lives ofhe drugs in rodents and humans (Aderjan and Skopp, 1998; Eap et al., 2002;ing et al., 1981). Because of the differences, we tested both the effects of dailynd every-other-day methadone treatment; the effects of morphine given every-ther-day were studied for the sake of comparison. The drugs were dissolved inhysiological saline to give injection volumes of 1 ml/kg.
.4. Locomotor activity testing
LA response was tested at standardized lighting and white laboratory noiseevels, using a model TSE Fear Conditioning System (TSE Systems GmbH,ad Homburg, Germany) equipped with rectangular Perspex chambers (35 cm
ength × 20 cm width × 19.3 cm height). The rat’s position inside the chamberas monitored with the help of two arrays (arranged in a horizontal plane at a
traight angle one to another) of 15 infra-red light barriers each. The barriersonsisted of one infra-red beam transmitter and one receiving sensor each andere located 23.6 mm (long axis) and 13 mm (short axis) apart. The arrays were
nterfaced with a computer that collected beam breaks and calculated the distanceraveled by the rat over pre-set consecutive 5-min time intervals 31–85 minost-injection. The chambers were thoroughly washed and dried prior to eachndividual session.
.5. Fos immunohistochemistry
Locations of the brain fields studied are shown in Fig. 1. All Fos stainingrocedures were done at room temperature unless indicated otherwise. Afterecapitation (with no perfusion, see Lehner et al., 2006; Taracha et al., 2006),he brains were instantly removed, snap-frozen in cold (−80 ◦C) isopentane andtored at −70 ◦C until processed further. For each region studied, two adjacentoronal 18 �m cryoslices were cut from each brain based on the rat brain atlasPaxinos and Watson, 1998), mounted on silanized glass slides, fixed for 5 min inethanol at −20 ◦C, and washed twice with 10 mM phosphate-buffered saline
H 7.4 (PBS; 2 × 15 min). The slices were then incubated for 30 min with a.3% H2O2 solution in methanol, washed with PBS (2 × 15 min) and incubatedor 2 h with a 5% NDS solution in PBS. Next, the slices were incubated for 3ays at 4–8 ◦C with diluted (1:2000) anti-c-Fos IgG solution in PBS supple-ented with 3% NDS, washed with PBS (3 × 15 min), incubated for 2 h with

E. Taracha et al. / Drug and Alcohol Dependence 97 (2008) 21–32 23
Table 1Rat treatment schedules
Group (number of rats) Pretreatment Withdrawal Challenge
Sal/Sal (N = 10) Physiological saline, s.c., 1 ml/kg, once dailyfor 6 days/week (the 2nd and 8thpretreatment days were drug-free), 14 doses
Two weeks of no injections/LA testing Physiological saline, s.c., 1 ml/kg
Sal/Morph5 (N = 16) Physiological saline, s.c., 1 ml/kg, once daily,6 days/week (break days as above), 14 doses
Two weeks of no injections/LA testing Morphine sulfate, s.c., 5 mg/kg
Morph5/Morph5 (N = 16) Morphine sulfate, s.c., 5 mg/kg, once daily, 6days/week (break days as above), 14 doses
Two weeks of no injections/LA testing Morphine sulfate, s.c., 5 mg/kg
Morph10/Morph5 (N = 10) Morphine sulfate, s.c., 10 mg/kg, once daily,6 days/week (break days as above), 14 doses
Two weeks of no injections/LA testing Morphine sulfate, s.c., 5 mg/kg
Morph10-2d/Morph5 (N = 8) Morphine sulfate, s.c., 10 mg/kg,every-other-day, 7 doses
Two weeks of no injections/LA testing Morphine sulfate, s.c., 5 mg/kg
Meth1/Morph5 (N = 12) Methadone hydrochloride, s.c., 1 mg/kg,once daily, 6 days/week (the 2nd and 8thpretreatment days were drug-free), 14 doses
Two weeks of no injections/LA testing Morphine sulfate, s.c., 5 mg/kg
Meth3-2d/Morph5 (N = 10)a Methadone hydrochloride, s.c.,every-other-day; 5 mg/kg on Day 1, followed
a
Two weeks of no injections/LA testing Morphine sulfate, s.c., 5 mg/kg
se of 5i alyse
bPabwTisqansMhfj
2
matelcAr
FTiptt
by 6 doses of 3 mg/kg
a At first there were 12 rats in this group, but the initially used methadone donjections, and behavioral data for the two rats were excluded from statistical an
iotinylated goat anti-rabbit IgGs supplemented with 3% NDS, washed withBS (2 × 15 min), incubated for 1 h with avidin–biotin–peroxidase complexccording to manufacturer’s instructions, and washed with 50 mM Tris–HCluffer pH 7.4 (2 × 15 min). Subsequently, the slices were incubated for 7 minith a 0.03% diaminobenzidine hydrochloride/0.003% H2O2 solution in 50 mMris–HCl, pH 7.4; the reaction was stopped by washing the slices with deion-
zed water. Finally, the slices were dehydrated using standard ethanol dilutioneries, dipped in xylene and coverslipped with DePeX. Fos immunostaining wasuantified at ×40 magnification by a person blinded to slices’ identity, usingn Olympus BX-51 microscope and a DP-70 Olympus camera. Fos-positiveuclei were counted bilaterally in both slices using a computerized analysis
ystem (Cell* Imaging Software for Life Sciences, Soft Imaging System Ltd.,unster, Germany), and the counts were averaged. Cortex layer I that showedardly any neuronal somata or Fos staining in any region studied was excludedrom the study, and Fos-positive nuclei in cortex layers II and III were countedointly.
v(cAS
ig. 1. Drawing of brain cross-sections (adapted from Paxinos and Watson, 1998, wihe cingulate cortex (CG), prelimbic cortex (PrL), motor cortex (M1/M2), primary so
nsular cortex (AI), basolateral amygdaloid nucleus (anterior part, BLA), lateral amygdart, CeC) and medial amygdaloid nucleus (Me) fields were of 0.1 mm2 each; the nuhe caudate-putamen (CPu) cross-sections examined were as shown in the middle pahe dentate gyrus (DG) field was of 1 mm2 area; the thalamic periventricular nucleus
mg/kg appeared lethal to two rats; hence the dosage was reduced for the nexts.
.6. Data analysis
LA data for the pretreatment period were first analyzed by three-way repeatedeasures’ analysis of variance (ANOVA) with drug regimen as the main factor
nd pretreatment day and post-injection time as the repeated measures’ fac-ors. Significance of within-group changes in LA response was then tested forach drug regimen separately by two-way repeated measures’ ANOVA fol-owed, when appropriate, by the t-test for dependent variables. Between-groupomparisons for individual days were done by two-way repeated measures’NOVA with drug regimen as the main factor and post-injection time as the
epeated measures’ factor, followed by the Bonferroni t-test for independent
ariables. Fos-positive cell counts were analyzed using the Kruskal–WallisK–W) ANOVA by ranks followed by the test of multiple comparisons versusontrol when appropriate. In all cases, a p < 0.05 was considered significant.ll the analyses were performed using the Statistica software v.7.1 (Stat-oft Inc., Tulsa, Oklahoma, USA), except that the multiple comparisons’th permission) and locations of fields analyzed by Fos immunohistochemistry.matosensory cortex (S1), granular/dysgranular cortex (GI/DI) cortex, agranularaloid nucleus (ventromedial part, LaVM), central amygdaloid nucleus (capsularcleus accumbens’ fields (core – AcbC, shell – AcbSh) were of 0.5 mm2 each;nel; the CA1, CA3 and IBl hippocampal fields studied were of 0.3 mm2 each;(PVP) and medial amygdaloid nucleus (Me) fields were of 0.2 mm2 each.
2 cohol Dependence 97 (2008) 21–32
t1
3
3
itdiautwLoo
ftwspp(ia(pLgict
31(itrwDtoir
3prtoFn
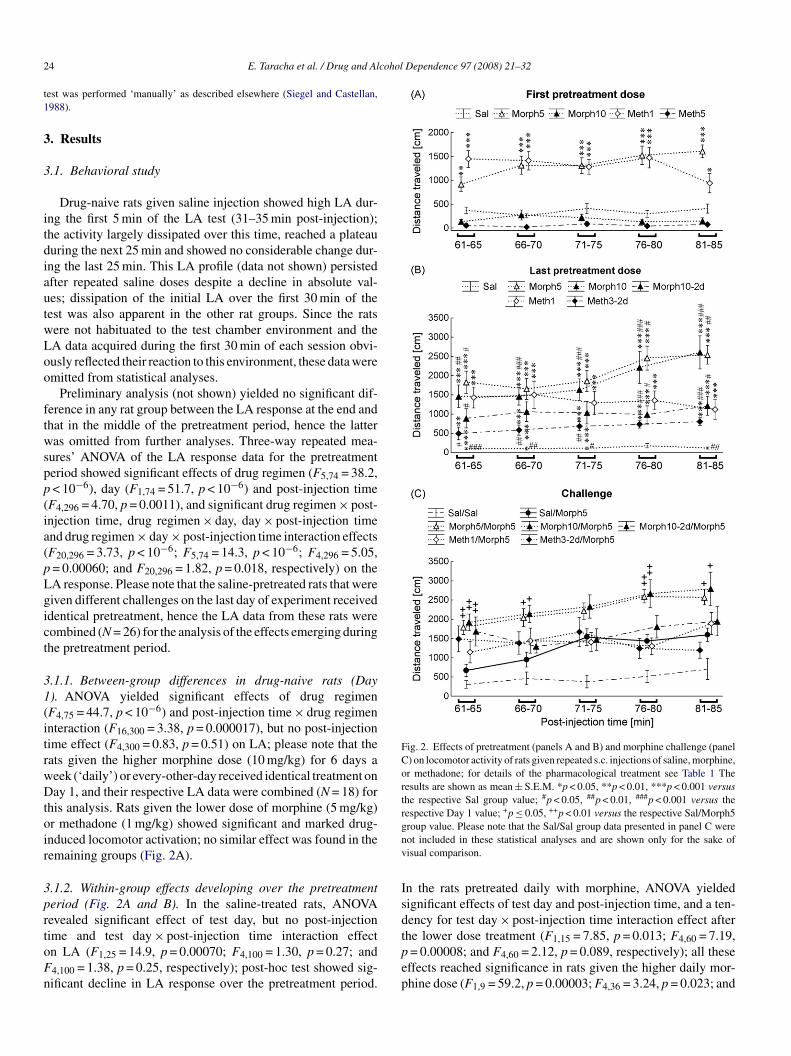
Fig. 2. Effects of pretreatment (panels A and B) and morphine challenge (panelC) on locomotor activity of rats given repeated s.c. injections of saline, morphine,or methadone; for details of the pharmacological treatment see Table 1 Theresults are shown as mean ± S.E.M. *p < 0.05, **p < 0.01, ***p < 0.001 versusthe respective Sal group value; #p < 0.05, ##p < 0.01, ###p < 0.001 versus therespective Day 1 value; +p ≤ 0.05, ++p < 0.01 versus the respective Sal/Morph5gnv
Isd
4 E. Taracha et al. / Drug and Al
est was performed ‘manually’ as described elsewhere (Siegel and Castellan,988).
. Results
.1. Behavioral study
Drug-naive rats given saline injection showed high LA dur-ng the first 5 min of the LA test (31–35 min post-injection);he activity largely dissipated over this time, reached a plateauuring the next 25 min and showed no considerable change dur-ng the last 25 min. This LA profile (data not shown) persistedfter repeated saline doses despite a decline in absolute val-es; dissipation of the initial LA over the first 30 min of theest was also apparent in the other rat groups. Since the ratsere not habituated to the test chamber environment and theA data acquired during the first 30 min of each session obvi-usly reflected their reaction to this environment, these data weremitted from statistical analyses.
Preliminary analysis (not shown) yielded no significant dif-erence in any rat group between the LA response at the end andhat in the middle of the pretreatment period, hence the latteras omitted from further analyses. Three-way repeated mea-
ures’ ANOVA of the LA response data for the pretreatmenteriod showed significant effects of drug regimen (F5,74 = 38.2,< 10−6), day (F1,74 = 51.7, p < 10−6) and post-injection time
F4,296 = 4.70, p = 0.0011), and significant drug regimen × post-njection time, drug regimen × day, day × post-injection timend drug regimen × day × post-injection time interaction effectsF20,296 = 3.73, p < 10−6; F5,74 = 14.3, p < 10−6; F4,296 = 5.05,= 0.00060; and F20,296 = 1.82, p = 0.018, respectively) on theA response. Please note that the saline-pretreated rats that wereiven different challenges on the last day of experiment receiveddentical pretreatment, hence the LA data from these rats wereombined (N = 26) for the analysis of the effects emerging duringhe pretreatment period.
.1.1. Between-group differences in drug-naive rats (Day). ANOVA yielded significant effects of drug regimenF4,75 = 44.7, p < 10−6) and post-injection time × drug regimennteraction (F16,300 = 3.38, p = 0.000017), but no post-injectionime effect (F4,300 = 0.83, p = 0.51) on LA; please note that theats given the higher morphine dose (10 mg/kg) for 6 days aeek (‘daily’) or every-other-day received identical treatment onay 1, and their respective LA data were combined (N = 18) for
his analysis. Rats given the lower dose of morphine (5 mg/kg)r methadone (1 mg/kg) showed significant and marked drug-nduced locomotor activation; no similar effect was found in theemaining groups (Fig. 2A).
.1.2. Within-group effects developing over the pretreatmenteriod (Fig. 2A and B). In the saline-treated rats, ANOVAevealed significant effect of test day, but no post-injection
ime and test day × post-injection time interaction effectn LA (F1,25 = 14.9, p = 0.00070; F4,100 = 1.30, p = 0.27; and4,100 = 1.38, p = 0.25, respectively); post-hoc test showed sig-ificant decline in LA response over the pretreatment period.tpep
roup value. Please note that the Sal/Sal group data presented in panel C wereot included in these statistical analyses and are shown only for the sake ofisual comparison.
n the rats pretreated daily with morphine, ANOVA yieldedignificant effects of test day and post-injection time, and a ten-ency for test day × post-injection time interaction effect after
he lower dose treatment (F1,15 = 7.85, p = 0.013; F4,60 = 7.19,= 0.00008; and F4,60 = 2.12, p = 0.089, respectively); all theseffects reached significance in rats given the higher daily mor-hine dose (F1,9 = 59.2, p = 0.00003; F4,36 = 3.24, p = 0.023; and
cohol
FadicdmpcdepFdmFatwt
3mr(tOs
3cd(i(ew
3Lwm(oyoptC
3
3Acs
3ilpmFiwnStct
3ApHlpbcstpcFMIhramer
3ilpptbaMtIeodSp
E. Taracha et al. / Drug and Al
4,36 = 3.60, p = 0.014, respectively). Post-hoc analysis showedrobust increase in LA response in the rats given the higher
aily morphine dose pretreatment, and a smaller but significantncrease in their low dose-pretreated counterparts. A signifi-ant effect of LA test day, but not of post-injection time or testay × post-injection time interaction was found in the rats givenorphine every-other-day (F1,7 = 6.24, p = 0.041; F4,28 = 1.39,= 0.26; and F4,28 = 1.14, p = 0.36, respectively); post-hoc testonfirmed increased LA response to morphine. No effect of testay, post-injection time or interaction between the two factors’ffects on LA was found in the daily low dose methadone-retreated rats (F1,11 = 0.01, p = 0.92; F4,44 = 1.90, p = 0.13; and4,44 = 0.31, p = 0.87, respectively). Significant effect of testay only was found in the rats given the higher dose ofethadone (3 mg/kg) every-other-day (F1,7 = 68.5, p = 0.00007;4,28 = 1.33, p = 0.28; and F4,28 = 0.82, p = 0.52, respectively),nd post-hoc analysis showed an increase in LA response overhe pretreatment period. However, this result should be takenith caution because of the forced decrease in drug dose after
he first day (see Table 1).
.1.3. Between-group differences after the last pretreat-ent dose. ANOVA yielded significant effects of drug
egimen (F5,74 = 24.0, p < 10−6), post-injection timeF4,296 = 6.90, p = 0.000025) and drug regimen × post-injectionime (F20,296 = 2.94, p = 0.000038) on the LA response.pioid-treated rats showed significantly higher LA than the
aline-treated controls (Fig. 2B).
.1.4. Pretreatment effects on the LA response to morphinehallenge. ANOVA yielded significant effects of pretreatmentrug regimen (F5,62 = 5.24, p = 0.00045), post-injection timeF4,248 = 12.2, p < 10−6) and drug regimen × post-injection timenteraction effect on the response to the morphine challengeF20,248 = 2.28, p = 0.0019). Post-hoc test showed significantnhancement of the LA response only in the rats pretreated dailyith morphine (Fig. 2C).
.1.5. LA response to saline challenge in saline-pretreated rats.A response data for the saline-pretreated saline-challenged ratsere analyzed separately to verify if the pretreatment lastinglyodified rats’ reactivity. Two-way repeated measures’ ANOVA
with repeated measures on LA test day and post-injection time)f the respective Day 1 and challenge day LA response dataielded no significant effect of test day, post-injection time,r interaction between the two factors’ effects (F1,9 = 0.014,= 0.91; F4,36 = 1.78, p = 0.15; and F4,36 = 1.33, p = 0.28, respec-
ively; see also the respective LA response curves in Fig. 2A and).
.2. Fos immunohistochemistry
.2.1. Cingulate cortex (CG) and prelimbic cortex (PrL). K–WNOVA showed no between-group difference in Fos-positive
ell counts in any CG or PrL layer studied (p ≥ 0.13; data nothown).
Ip
3s
Dependence 97 (2008) 21–32 25
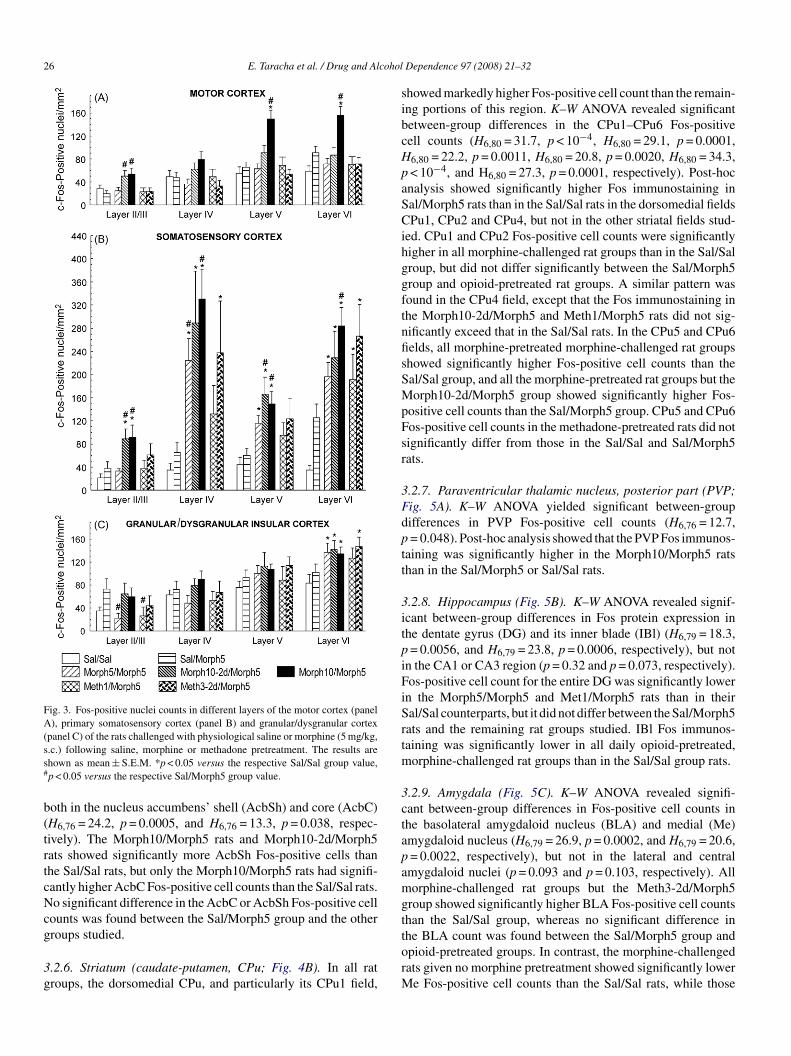
.2.2. Motor cortex (Fig. 3A). K–W ANOVA revealed signif-cant between-group differences in Fos protein expression inayers II/III, V and VI (H6,79 = 15.9, p = 0.0141, H6,79 = 23.5,= 0.0006, and H6,79 = 20.5, p = 0.0023, respectively). Post-hocultiple comparisons’ test showed no significant difference inos immunostaining between the Sal/Sal and Sal/Morph5 rats
n any motor cortex layer. Morphine-challenged rats pretreatedith high morphine dose daily or every-other-day showed sig-ificantly higher layer II/III Fos-positive cell counts than theal/Morph5 rats, but not the Sal/Sal rats. Only the rats given
he daily high morphine dose pretreatment showed signifi-antly higher layers’ V and VI Fos-positive cell counts thanhe Sal/Morph5 or the Sal/Sal rats.
.2.3. Primary somatosensory cortex (S1; Fig. 3B). K–WNOVA revealed significant between-group differences in Fosrotein expression in all S1 layers examined (layers II/III:6,N=80 = 21.2, p = 0.0017; layer IV: H6,N=80 = 33.6, p < 10−4;
ayer V: H6,N=80 = 24.3, p = 0.0005; layer VI: H6,N=80 = 35.8,< 10−4). Post-hoc analysis showed no significant differenceetween Sal/Morph5 and Sal/Sal rats in Fos-positive cellounts in any S1 layer. In all S1 layers, Morph10/Morph5 ratshowed significantly stronger Fos immunostaining than eitherhe Sal/Morph5 rats or the Sal/Sal rats. Rats pretreated with mor-hine every-other-day showed significantly more Fos-positiveells in layers II–VI than the Sal/Sal rats and significantly moreos-positive cells in layers II/III and V than the Sal/Morph5 rats.orph5/Morph5 rats showed significantly stronger S1 layers’
V–V Fos immunostaining than the Sal/Sal rats, and significantlyigher layer IV Fos protein expression than the Sal/Morph5ats. No significant difference between methadone-pretreatednd saline-pretreated rats was found in any S1 layer after theorphine challenge, whereas Fos-positive cell counts in S1 lay-
rs IV and VI were significantly higher in Meth3-2d/Morph5ats than in the Sal/Sal rats.
.2.4. Insular cortex (Fig. 3C). K–W ANOVA showed signif-cant between-group differences in Fos-positive cell counts inayers II/III and VI (H6,80 = 17.6, p = 0.0075, and H6,80 = 16.5,= 0.0114, respectively), but not IV and V (p = 0.069 and= 0.16, respectively) of the granular/dysgranular insular cor-
ex field examined (GI/DI). There was no significant differenceetween Sal/Sal and Sal/Morph5 rats in Fos immunostaining inny GI/DI layer. In layers II/III, both the Morph5/Morph5 andeth1/Morph5 rats showed significantly weaker Fos immunos-
aining than the Sal/Morph5 rats, but not the Sal/Sal controls.n layer VI, there was no significant difference in Fos proteinxpression between the Sal/Morph5 and the other rats, but allpioid-pretreated groups except that given the low methadoneose showed significantly higher Fos protein expression than theal/Sal group. No significant between-group difference in Fos-ositive cell counts was found in the agranular cortex (layersI/III: p = 0.068; layer IV: p = 0.37; layer V: p = 0.97; layer VI:
= 0.89; data not shown)..2.5. Nucleus accumbens (Fig. 4A). K–W ANOVA revealedignificant between-group differences in Fos immunostaining
26 E. Taracha et al. / Drug and Alcohol
Fig. 3. Fos-positive nuclei counts in different layers of the motor cortex (panelA), primary somatosensory cortex (panel B) and granular/dysgranular cortex(panel C) of the rats challenged with physiological saline or morphine (5 mg/kg,ss#
b(trtcNcg
3g
sibcHpaSCihggftnfisSMpFsr
3Fdptt
3itpiFiSrtm
3ctapamgt
.c.) following saline, morphine or methadone pretreatment. The results arehown as mean ± S.E.M. *p < 0.05 versus the respective Sal/Sal group value,p < 0.05 versus the respective Sal/Morph5 group value.
oth in the nucleus accumbens’ shell (AcbSh) and core (AcbC)H6,76 = 24.2, p = 0.0005, and H6,76 = 13.3, p = 0.038, respec-ively). The Morph10/Morph5 rats and Morph10-2d/Morph5ats showed significantly more AcbSh Fos-positive cells thanhe Sal/Sal rats, but only the Morph10/Morph5 rats had signifi-antly higher AcbC Fos-positive cell counts than the Sal/Sal rats.o significant difference in the AcbC or AcbSh Fos-positive cell
ounts was found between the Sal/Morph5 group and the other
roups studied..2.6. Striatum (caudate-putamen, CPu; Fig. 4B). In all ratroups, the dorsomedial CPu, and particularly its CPu1 field,
torM
Dependence 97 (2008) 21–32
howed markedly higher Fos-positive cell count than the remain-ng portions of this region. K–W ANOVA revealed significantetween-group differences in the CPu1–CPu6 Fos-positiveell counts (H6,80 = 31.7, p < 10−4, H6,80 = 29.1, p = 0.0001,6,80 = 22.2, p = 0.0011, H6,80 = 20.8, p = 0.0020, H6,80 = 34.3,< 10−4, and H6,80 = 27.3, p = 0.0001, respectively). Post-hocnalysis showed significantly higher Fos immunostaining inal/Morph5 rats than in the Sal/Sal rats in the dorsomedial fieldsPu1, CPu2 and CPu4, but not in the other striatal fields stud-
ed. CPu1 and CPu2 Fos-positive cell counts were significantlyigher in all morphine-challenged rat groups than in the Sal/Salroup, but did not differ significantly between the Sal/Morph5roup and opioid-pretreated rat groups. A similar pattern wasound in the CPu4 field, except that the Fos immunostaining inhe Morph10-2d/Morph5 and Meth1/Morph5 rats did not sig-ificantly exceed that in the Sal/Sal rats. In the CPu5 and CPu6elds, all morphine-pretreated morphine-challenged rat groupshowed significantly higher Fos-positive cell counts than theal/Sal group, and all the morphine-pretreated rat groups but theorph10-2d/Morph5 group showed significantly higher Fos-
ositive cell counts than the Sal/Morph5 group. CPu5 and CPu6os-positive cell counts in the methadone-pretreated rats did notignificantly differ from those in the Sal/Sal and Sal/Morph5ats.
.2.7. Paraventricular thalamic nucleus, posterior part (PVP;ig. 5A). K–W ANOVA yielded significant between-groupifferences in PVP Fos-positive cell counts (H6,76 = 12.7,= 0.048). Post-hoc analysis showed that the PVP Fos immunos-
aining was significantly higher in the Morph10/Morph5 ratshan in the Sal/Morph5 or Sal/Sal rats.
.2.8. Hippocampus (Fig. 5B). K–W ANOVA revealed signif-cant between-group differences in Fos protein expression inhe dentate gyrus (DG) and its inner blade (IBl) (H6,79 = 18.3,= 0.0056, and H6,79 = 23.8, p = 0.0006, respectively), but not
n the CA1 or CA3 region (p = 0.32 and p = 0.073, respectively).os-positive cell count for the entire DG was significantly lower
n the Morph5/Morph5 and Met1/Morph5 rats than in theiral/Sal counterparts, but it did not differ between the Sal/Morph5ats and the remaining rat groups studied. IBl Fos immunos-aining was significantly lower in all daily opioid-pretreated,
orphine-challenged rat groups than in the Sal/Sal group rats.
.2.9. Amygdala (Fig. 5C). K–W ANOVA revealed signifi-ant between-group differences in Fos-positive cell counts inhe basolateral amygdaloid nucleus (BLA) and medial (Me)mygdaloid nucleus (H6,79 = 26.9, p = 0.0002, and H6,79 = 20.6,= 0.0022, respectively), but not in the lateral and centralmygdaloid nuclei (p = 0.093 and p = 0.103, respectively). Allorphine-challenged rat groups but the Meth3-2d/Morph5
roup showed significantly higher BLA Fos-positive cell countshan the Sal/Sal group, whereas no significant difference in
he BLA count was found between the Sal/Morph5 group andpioid-pretreated groups. In contrast, the morphine-challengedats given no morphine pretreatment showed significantly lowere Fos-positive cell counts than the Sal/Sal rats, while those
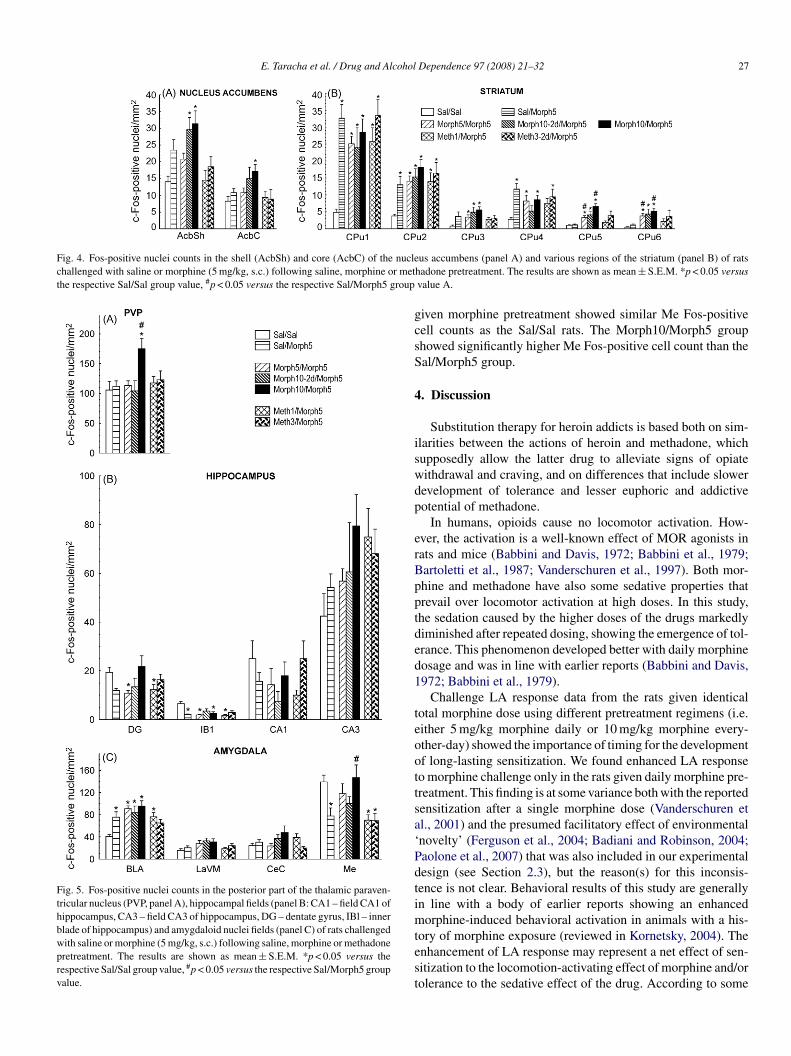
E. Taracha et al. / Drug and Alcohol Dependence 97 (2008) 21–32 27
Fig. 4. Fos-positive nuclei counts in the shell (AcbSh) and core (AcbC) of the nuclechallenged with saline or morphine (5 mg/kg, s.c.) following saline, morphine or metthe respective Sal/Sal group value, #p < 0.05 versus the respective Sal/Morph5 group
Fig. 5. Fos-positive nuclei counts in the posterior part of the thalamic paraven-tricular nucleus (PVP, panel A), hippocampal fields (panel B: CA1 – field CA1 ofhippocampus, CA3 – field CA3 of hippocampus, DG – dentate gyrus, IBl – innerblade of hippocampus) and amygdaloid nuclei fields (panel C) of rats challengedwith saline or morphine (5 mg/kg, s.c.) following saline, morphine or methadonepretreatment. The results are shown as mean ± S.E.M. *p < 0.05 versus therespective Sal/Sal group value, #p < 0.05 versus the respective Sal/Morph5 groupvalue.
gcsS
4
iswdp
erBpptded1
teoottsa‘Pdtimtest
us accumbens (panel A) and various regions of the striatum (panel B) of ratshadone pretreatment. The results are shown as mean ± S.E.M. *p < 0.05 versusvalue A.
iven morphine pretreatment showed similar Me Fos-positiveell counts as the Sal/Sal rats. The Morph10/Morph5 grouphowed significantly higher Me Fos-positive cell count than theal/Morph5 group.
. Discussion
Substitution therapy for heroin addicts is based both on sim-larities between the actions of heroin and methadone, whichupposedly allow the latter drug to alleviate signs of opiateithdrawal and craving, and on differences that include slowerevelopment of tolerance and lesser euphoric and addictiveotential of methadone.
In humans, opioids cause no locomotor activation. How-ver, the activation is a well-known effect of MOR agonists inats and mice (Babbini and Davis, 1972; Babbini et al., 1979;artoletti et al., 1987; Vanderschuren et al., 1997). Both mor-hine and methadone have also some sedative properties thatrevail over locomotor activation at high doses. In this study,he sedation caused by the higher doses of the drugs markedlyiminished after repeated dosing, showing the emergence of tol-rance. This phenomenon developed better with daily morphineosage and was in line with earlier reports (Babbini and Davis,972; Babbini et al., 1979).
Challenge LA response data from the rats given identicalotal morphine dose using different pretreatment regimens (i.e.ither 5 mg/kg morphine daily or 10 mg/kg morphine every-ther-day) showed the importance of timing for the developmentf long-lasting sensitization. We found enhanced LA responseo morphine challenge only in the rats given daily morphine pre-reatment. This finding is at some variance both with the reportedensitization after a single morphine dose (Vanderschuren etl., 2001) and the presumed facilitatory effect of environmentalnovelty’ (Ferguson et al., 2004; Badiani and Robinson, 2004;aolone et al., 2007) that was also included in our experimentalesign (see Section 2.3), but the reason(s) for this inconsis-ence is not clear. Behavioral results of this study are generallyn line with a body of earlier reports showing an enhanced
orphine-induced behavioral activation in animals with a his-
ory of morphine exposure (reviewed in Kornetsky, 2004). Thenhancement of LA response may represent a net effect of sen-itization to the locomotion-activating effect of morphine and/orolerance to the sedative effect of the drug. According to some
2 cohol
rano
earlotwbdTai
emGar1mswmtcftfd
artezeidbtCacse
occitfi
srestHpavoa
cIRbtsttlpcptirsevho
cpstttHccmiettioepdi
8 E. Taracha et al. / Drug and Al
esearchers (see Zernig et al., 2004) the tolerance alone beingmajor factor in drug dependence would be the most parsimo-ious explanation of the apparent behavioral sensitization effectf the pretreatment.
Despite the well-known cross-tolerance to the analgesicffects of morphine and methadone, and the similarity betweencute effects of these drugs on LA in the present study, the LAesponse of the methadone-pretreated rats to morphine chal-enge indicated no cross-tolerance or cross-sensitization to thepiate. No report from a rodent study is available to disputehis observation. However, this finding appears to correspondith a recent report on methadone dosage required to com-at withdrawal symptoms in heroin addicts who presented foretoxification upon multiple admissions (over up to 7 years).hose patients showed only slight tolerance during the repeatedbuse/detoxification cycles and no sensitization to the reinforc-ng effects of the opioids (Madlung et al., 2006).
In most brain regions, Fos expression is low at baseline (Sagart al., 1988), but can increase after a variety of stimuli, includingorphine (Chang et al., 1988; Erdtmann-Vourliotis et al., 1998;utstein et al., 1998). Morphine shows a pattern of brain c-fos
ctivation that differs between drug-naive and post-dependentats (Curran et al., 1996; Harlan and Garcia, 1998; Frankel et al.,999; Ferguson et al., 2004). We have shown earlier that singleethadone doses enhance Fos protein expression in rat dorsal
triatum and AcbC, but not in AcbSh (Taracha et al., 2006);e were also interested if repeated methadone treatment canodify brain’s sensitivity to morphine. As the challenge used in
his study was the same for all rat groups except for the Sal/Salontrols, the between-group differences found reflected the dif-erences in past exposure to opioids. These results revealed thathe effects of the pretreatment on Fos protein expression dif-ered among the brain regions examined with regard to both theosing regimen and identity of the pretreatment drug.
Morphine acts mostly via inhibitory MORs (Laugwitz etl., 1993); its direct inhibitory action, e.g. on striatal dopamineelease, can be seen after local administration. This effect showshe development of tolerance after repeated dosage (Piepponent al., 1999) and may have some relevance to behavioral sensiti-ation after repeated systemic morphine doses. The stimulatoryffect of systemic morphine on striatal and accumbal Fos, whichs dopamine-mediated, is indirect and results from predominantisinhibition of mesolimbic and nigrostriatal dopamine neuronsy the suppression of GABA interneurons in the ventral tegmen-al area and substantia nigra pars reticularis, respectively (Dihiara and North, 1992; Johnson and North, 1992; Bontempind Sharp, 1997; Steffensen et al., 2006). Brain GABA-ergicircuits appear to play an important role also in behavioral sen-itization to morphine (e.g. see Bartoletti et al., 2007; Zarrindastt al., 2007).
Since dopaminergic circuits play an important role in opi-id effects on locomotion and Fos expression, one mightonsider psychoactive drugs’ potential for interfering with spe-
ific dopamine reuptake as a factor in these phenomena. Its known that methadone inhibits monoamine reuptake andhat co-administration of monoamine uptake inhibitors modi-es some opiate effects. For instance, co-administration of theo
aw
Dependence 97 (2008) 21–32
elective serotonin reuptake inhibitor fluoxetine potentiates theewarding effect of morphine in a conditioned place prefer-nce paradigm (Subhan et al., 2000), whereas venlafaxine (aerotonin/noradrenaline reuptake inhibitor) attenuates acquisi-ion of heroin self-administration in rats (Magalas et al., 2005).owever, morphine showed considerably stronger sensitizationotential than methadone in our study despite having no knownffinity for specific monoamine reuptake systems. This obser-ation suggests that the differences between the effects of thesepioids in our study were not related to the efficacy of the drugst those sites.
In some brain regions, systemic morphine given in advancean suppress Fos protein expression caused by other stimuli.n our study, such stimulus likely was the novelty effect (seeyabinin et al., 1995; Ferguson et al., 2004) of the LA test cham-er, and possibly also of the post-injection cage. As reflected byhe waning LA response to repeated saline injections, our ratshowed some habituation to these environments during the pre-reatment period. However, this effect dissipated entirely overhe 2-week withdrawal period that followed. The morphine chal-enge preceding the final LA test partially counteracted theutative stimulatory effect of the post-injection cage/LA testinghamber novelty, resulting, in some brain regions, in a lower Fosrotein expression than that in the Sal/Sal controls. The extent ofhis suppression was related to the history of opioid exposure; fornstance, the suppression was evident in the Me provided the ratseceived no morphine pretreatment. This suggested that expo-ure to morphine caused lasting tolerance to the Fos inhibitoryffect of the drug; the tolerance showed some regimen-relatedariability, but could not be induced by methadone. On the otherand, we found no substantial difference between the abilitiesf these opioids for inducing the tolerance in DG or its IBl field.
In the remaining brain regions studied the effect of morphinehallenge on Fos protein expression varied depending on theretreatment regimen employed. In the BLA and dorsomedialtriatum there was no difference between rats with divergent his-ories of past opioid exposure. This would suggest no adaptationo repeated opioid treatment and might relate these regions tohe effects that show no change during long-term opiate use.owever, it has been shown by others that BLA undergoes
onsiderable neuroplastic changes in response to various psy-hoactive substances. For instance, repeated morphine injectionsodify the expression of many proteins in this region, includ-
ng NMDAR1 receptor (Turchan et al., 2003), and induce thexpression of the anti-adhesive glycoprotein SPARC that seemso be causally related to the emergence of behavioral sensitiza-ion (Ikemoto et al., 2000); BLA may also play a pivotal role (asndicated by altered FosB protein expression) in the expressionf negative symptoms after amphetamine withdrawal (Murphyt al., 2003). The fact that opioid pretreatment did not modify Fosrotein expression in the BLA and dorsomedial CPu in our studyoes not preclude a role of this protein as an important player innitiating expression of proteins and neuroplastic modification
f these structures.In the centrolateral part of CPu, Acb, PVP, layers II/III, Vnd VI of M1/M2 cortex and layers II/III and V of S1 cortex, ofhich regions CPu and Acb are considered essential for the
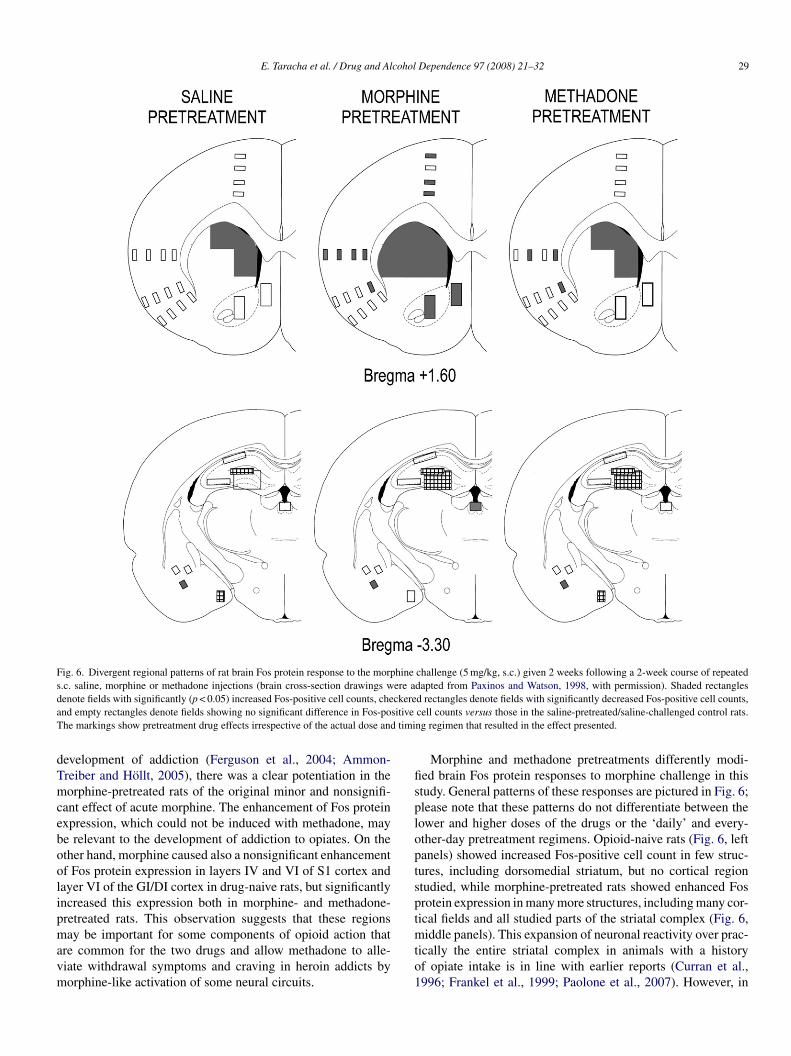
E. Taracha et al. / Drug and Alcohol Dependence 97 (2008) 21–32 29
Fig. 6. Divergent regional patterns of rat brain Fos protein response to the morphine challenge (5 mg/kg, s.c.) given 2 weeks following a 2-week course of repeateds.c. saline, morphine or methadone injections (brain cross-section drawings were adapted from Paxinos and Watson, 1998, with permission). Shaded rectanglesd ckerea sitiveT timi
dTmceboolipmavm
fisploptspt
enote fields with significantly (p < 0.05) increased Fos-positive cell counts, chend empty rectangles denote fields showing no significant difference in Fos-pohe markings show pretreatment drug effects irrespective of the actual dose and
evelopment of addiction (Ferguson et al., 2004; Ammon-reiber and Hollt, 2005), there was a clear potentiation in theorphine-pretreated rats of the original minor and nonsignifi-
ant effect of acute morphine. The enhancement of Fos proteinxpression, which could not be induced with methadone, maye relevant to the development of addiction to opiates. On thether hand, morphine caused also a nonsignificant enhancementf Fos protein expression in layers IV and VI of S1 cortex andayer VI of the GI/DI cortex in drug-naive rats, but significantlyncreased this expression both in morphine- and methadone-retreated rats. This observation suggests that these regions
ay be important for some components of opioid action thatre common for the two drugs and allow methadone to alle-iate withdrawal symptoms and craving in heroin addicts byorphine-like activation of some neural circuits.
mto1
d rectangles denote fields with significantly decreased Fos-positive cell counts,cell counts versus those in the saline-pretreated/saline-challenged control rats.ng regimen that resulted in the effect presented.
Morphine and methadone pretreatments differently modi-ed brain Fos protein responses to morphine challenge in thistudy. General patterns of these responses are pictured in Fig. 6;lease note that these patterns do not differentiate between theower and higher doses of the drugs or the ‘daily’ and every-ther-day pretreatment regimens. Opioid-naive rats (Fig. 6, leftanels) showed increased Fos-positive cell count in few struc-ures, including dorsomedial striatum, but no cortical regiontudied, while morphine-pretreated rats showed enhanced Fosrotein expression in many more structures, including many cor-ical fields and all studied parts of the striatal complex (Fig. 6,
iddle panels). This expansion of neuronal reactivity over prac-ically the entire striatal complex in animals with a historyf opiate intake is in line with earlier reports (Curran et al.,996; Frankel et al., 1999; Paolone et al., 2007). However, in

3 cohol
cehcdeOeomemtic(bsi
msmredaNbagaSit2Mcftrvmo
itofttNofimsb
sM
btdweiupaoitottAmiros
5
lcpptieNtaaaWpimmmtet
A
0 E. Taracha et al. / Drug and Al
ontrast to our data, heroin-induced suppression of Fos proteinxpression has been reported in the somatosensory cortex oferoin-sensitized rats (Paolone et al., 2007). This apparent dis-repancy may be explained by the use of small (1 mg/kg) heroinoses instead of relatively high morphine doses and studying theffect in another region of somatosensory cortex in that report.n the other hand, our data are in agreement with those of Curran
t al. (1996), who reported c-fos activation in layers IV and VIf the primary somatosensory cortex of rats given repeated i.p.orphine doses of 10 mg/kg. We found increased Fos protein
xpression also in other layers of the cortex after similar treat-ent, but this difference could be due to our using s.c. route
hat is known to be more effective. Morphine pretreatment mod-fied also the pattern of morphine challenge-evoked inhibitoryhanges in Fos protein expression. Methadone-pretreated ratsFig. 6, right panels) demonstrated an intermediate pattern ofoth stimulatory and inhibitory changes in Fos protein expres-ion, but showed more resemblance to the morphine-naive ratsn this respect.
It has been demonstrated that repeated administration oforphine as compared with that of methadone exerts much
tronger effects in the CNS. This observation is in line withuch higher potential of the natural opioid for inducing neu-
oplastic changes that result in tolerance and/or sensitization atither behavioral or molecular level. Some researchers relate thisifference to methadone’s, and particularly its S-enantiomer’sntagonistic properties at NMDA receptors (Inturrisi, 2005).otably, the induction of expression of c-fos in CPu and Acby morphine and at least some psychostimulants (cocaine,mphetamine), which should lead to long-term changes ofene expression, can be blocked with the NMDA receptorntagonist MK-801 (Liu et al., 1994; Sharp et al., 1995).timulation of these receptors supposedly plays a pivotal role
n the neuroplastic changes underlying the development ofolerance and sensitization (reviewed in Bisaga and Popik,000; see also Raith and Hochhaus, 2004; Popik et al., 2006;endez and Trujillo, 2008). Moreover, there is both biochemi-
al and behavioral evidence showing increased NMDA receptorunction in the nucleus accumbens, with respect to an interac-ion with the limbic dopamine system, in morphine-dependentats (Murray et al., 2007). These observations may be rele-ant to the differences between the effects of morphine andethadone pretreatments on Fos expression in this structure in
ur study.Yet another explanation of the markedly stronger tolerance-
nducing and addictive potential of morphine as compared tohat of methadone and other MOR agonists (e.g. opioid peptidesr fentanyl) may be the relatively low potential of morphineor inducing MOR internalization. This ‘deficiency’ may leado superactivation of the cAMP pathway and NMDA recep-ors (He and Whistler, 2005). S-Methadone is a relatively weakMDA receptor antagonist and its dose was relatively low inur study (0.5 mg/kg for the daily methadone pretreatment and
rst 2.5 and later 1.5 mg/kg for the every-other-day pretreat-ent) as compared to that used by others (e.g. 10 mg/kg in thetudy of Davis and Inturrisi, 1999). Considering these facts weelieve that the generally stronger effects of morphine in our
Ne
t
Dependence 97 (2008) 21–32
tudy should rather be ascribed to its low propensity for inducingOR internalization.The relation of the pretreatment regimen to the changes in
rain Fos protein expression is even more complex than thato the changes in LA response. For instance, no substantialifference in LA response was found between rats pretreatedith different daily morphine doses, while the higher doses
nhanced Fos protein expression in more brain regions. Still,n some areas, e.g. in the AcbSh, similar effects were obtainedsing either the daily or every-other-day high morphine doseretreatment. This suggests that the creation of the putative, opi-te use-related molecular adaptation required repeated dosingf certain minimum drug dose, whereas the timing character-stic of the pretreatment regimen was of less importance. Onhe other hand, every-other-day pretreatments with morphiner methadone were equally effective in inducing IBl Fos pro-ein ‘tolerance, in our rats, which indicated that, in this region,he timing was more important than the identity of the opioid.pparently, the effects of repeated treatment with morphine andethadone are regiospecific and their relations to dosing reg-
men (dose, timing, etc.) depend on functions related to theseegions. Regional specificity of the differences between the twopioids finds support also in the studies on the effects of theseubstances on MOR desensitization (Martin et al., 2007).
. Concluding remarks
Two main findings of this study are (i) that pro-onged methadone administration causes no behavioralross-sensitization to morphine, and (ii) that the histories ofre-exposure to these opioids differently modify brain regionalatterns of neuronal activation by morphine in the rat. Forma-ion of lasting biochemical/molecular and behavioral ‘traces’s a peculiar attribute of addictive substances and suppos-dly plays a pivotal role in addiction relapses (reviewed inestler, 2004; Le Moal and Koob, 2007). Methadone substi-
ution is a well-known method of harm reduction in heroinddicts, but its addictive potential and cross-tolerance betweennalgesic effects of opiates and methadone bring doubtsbout the actual curative efficacy of the synthetic opioid.
hereas repeated methadone administration enhanced mor-hine challenge-induced activation of Fos protein expressionn few brain regions of unknown relevance to the develop-
ent of opioid addiction/cross-sensitization, this effect wasostly observed in rats given high methadone doses. Notably,ethadone did not show potential for a lasting enhancement of
he neuronal activation response to morphine in regions consid-red pivotal for the development of addiction to opiates, e.g. inhe nucleus accumbens.
cknowledgment
The authors thank Mrs. A. Biegaj of the Department of
eurochemistry, Institute of Psychiatry and Neurology, for herxcellent technical assistance.Role of funding source: The study was supported by the Insti-
ute of Psychiatry and Neurology statutory fund #501-03-07-64

cohol
agnidt
phiDtdabhdc
tt
R
A
A
B
B
B
B
B
B
B
B
C
C
C
C
D
D
D
D
E
E
F
F
G
H
H
H
I
I
J
K
K
K
K
L
L
L
E. Taracha et al. / Drug and Al
nd by the Polish Ministry of Science and Higher Educationrant #26/E-40/SN-023/2006 “Search for new sites of action ofeurotropic agents”. The funding institutions had no further rolen study design, in the collection, analysis and interpretation ofata, in the writing of the report, and in the decision to submithe paper for publication.
Contributors: Dr. Taracha designed the study, wrote therotocol, supervised the experiments, quantified Fos immuno-istochemistry, managed the literature searches and analyses,nterpreted the data and prepared the first draft of the manuscript.r. Chrapusta performed the statistical analyses, co-managed
he literature searches and analyses, and helped to interpret theata and prepare the first draft of the manuscript. Dr. Lehnernd M.Sc. Skorzewska ran the behavioral tests and collected therains for immunohistochemistry. Drs. Maciejak and Szyndlerelped to run Fos immunochemistry. Prof. Płaznik assisted inesigning the study and helped to interpret the data. All authorsontributed to and have approved the final manuscript.
Conflict of interest: All the authors/contributors declare thathey have no conflicts of interest related to the subject and con-ent of this report.
eferences
derjan, R.E., Skopp, C., 1998. Formation and clearance of active and inactivemetabolites of opiates in humans. Ther. Drug Monit. 20, 561–569.
mmon-Treiber, S., Hollt, V., 2005. Morphine-induced changes of gene expres-sion in the brain. Addict. Biol. 10, 81–89.
abbini, M., Davis, W.M., 1972. Time-dose relationships for locomotor activityeffects of morphine after acute or repeated treatment. Br. J. Pharmacol. 46,213–224.
abbini, M., Gaiardi, M., Bartoletti, M., 1975. Persistence of chronic mor-phine effects upon activity in rats 8 months after ceasing the treatment.Neuropharmacology 14, 611–614.
abbini, M., Gaiardi, M., Bartoletti, M., 1979. Dose-time motility effects ofmorphine and methadone in naive or morphinized rats. Pharmacol. Res.Commun. 11, 809–815.
adiani, A., Robinson, T.E., 2004. Drug-induced neurobehavioral plasticity:role of environmental context. Behav. Pharmacol. 15, 327–339.
artoletti, M., Gaiardi, M., Gebellini, C., Bacchi, A., Nannini, M., 1987. Pre-vious treatment with morphine and sensitization to the excitatory actions ofopiates: dose-effect relationship. Neuropharmacology 26, 115–119.
artoletti, M., Ricci, F., Gaiardi, M., 2007. A GABAB agonist reverses the behav-ioral sensitization to morphine in rats. Psychopharmacology 192, 79–85.
isaga, A., Popik, P., 2000. In search of a new pharmacological treatmentfor drug and alcohol addiction: N-methyl-d-aspartate (NMDA) antagonists.Drug Alcohol Depend. 59, 1–15.
ontempi, B., Sharp, F.R., 1997. Systemic morphine-induced Fos protein in therat striatum and nucleus accumbens is regulated by � opioid receptors in thesubstantia nigra and ventral tegmental area. J. Neurosci. 17, 8596–8612.
adoni, C., Di Chiara, G., 1999. Reciprocal changes in dopamine responsivenessin the nucleus accumbens shell and core and in the dorsal caudate-putamenin rats sensitized to morphine. Neuroscience 90, 447–455.
adoni, C., Pisanu, A., Solinas, M., Acquas, E., Di Chiara, G., 2001. Behaviouralsensitization after repeated exposure to �9-tetrahydrocannabinol and cross-sensitization with morphine. Psychopharmacology 158, 259–266.
hang, S.L., Squinto, S.P., Harlan, R.E., 1988. Morphine activation of c-Fos
expression in rat brain. Biochem. Biophys. Res. Commun. 157, 698–704.urran, E.J., Akil, H., Watson, S.J., 1996. Psychomotor stimulant and opiate-induced c-fos mRNA expression patterns in the rat forebrain: comparisonsbetween acute drug treatment and a drug challenge in sensitized animals.Neurochem. Res. 21, 1425–1435.
L
L
Dependence 97 (2008) 21–32 31
’Este, L., Scontrini, A., Casini, A., Pontieri, F.E., Renda, T.G., 2002. Heroinsensitization as mapped by c-Fos immunoreactivity in the rat striatum. BrainRes. 933, 144–149.
avis, A.M., Inturrisi, C.E., 1999. d-Methadone blocks morphine tolerance andN-methyl-d-aspartate-induced hyperalgesia. J. Pharmacol. Exp. Ther. 289,1048–1053.
e Vries, T.J., Schoffelmeer, A.N., Binnekade, R., Mulder, A.H., Van-derschuren, L.J., 1998. Drug-induced reinstatement of heroin- andcocaine-seeking behaviour following long-term extinction is associated withexpression of behavioural sensitization. Eur. J. Neurosci. 10, 3565–3571.
i Chiara, G., North, R.A., 1992. Neurobiology of opiate abuse. Trends Phar-macol. Sci. 13, 185–193.
ap, C.B., Buclin, T., Baumann, P., 2002. Interindividual variability of the clini-cal pharmacokinetics of methadone: implications for the treatment of opioiddependence. Clin. Pharmacokinet. 41, 1153–1193.
rdtmann-Vourliotis, M., Mayer, P., Riechert, U., Grecksch, G., Hollt, V., 1998.Identification of brain regions that are markedly activated by morphine intolerant but not in naive rats. Mol. Brain Res. 61, 51–61.
erguson, S.M., Thomas, M.J., Robinson, T.E., 2004. Morphine-induced c-fosmRNA expression in striatofugal circuits: modulation by dose, environmen-tal context, and drug history. Neuropsychopharmacology 29, 1664–1674.
rankel, P.S., Harlan, R.E., Garcia, M.M., 1999. Chronic administration ofmorphine alters immediate-early gene expression in the forebrain of post-dependent rats. Brain Res. 835, 204–212.
utstein, H.B., Thome, J.L., Fine, J.L., Watson, S.J., Akil, H., 1998. Patternof c-fos m-RNA induction in rat brain by acute morphine. Can. Physiol.Pharmacol. 76, 294–303.
arlan, R.E., Garcia, M.M., 1998. Drugs of abuse and immediate-early genesin the forebrain. Mol. Neurobiol. 16, 221–267.
e, S.T., Grasing, K., 2007. Chronic opiate treatment enhances both cocaine-reinforced and cocaine-seeking behaviors following opiate withdrawal. DrugAlcohol Depend. 75, 215–221.
e, L., Whistler, J.L., 2005. An opiate cocktail that reduces morphine toleranceand dependence. Curr. Biol. 15, 1028–1033.
kemoto, M., Takita, M., Imamura, T., Inoue, K., 2000. Increased sensitivity tothe stimulant effects of morphine conferred by anti-adhesive glycoproteinSPARC in amygdala. Nature Med. 6, 910–915.
nturrisi, C.E., 2005. Pharmacology of methadone and its isomers. MinervaAnestesiol. 71, 435–437.
ohnson, S.W., North, R.A., 1992. Opioids excite dopamine neurons by hyper-polarization of local interneurons. J. Neurosci. 12, 483–488.
alinichev, M., White, D.A., Holtzman, S.G., 2004. Individual differences inlocomotor reactivity to a novel environment and sensitivity to opioid drugsin the rat. I. Expression of morphine-induced locomotor sensitisation. Psy-chopharmacology 177, 61–67.
ornetsky, C., 2004. Brain–stimulation reward, morphine-induced oral stereo-typy, and sensitization: implications for abuse. Neurosci. Biobehav. Rev. 27,777–786.
reek, M.J., 2000. Methadone-related opioid agonist pharmacotherapy forheroin addiction. History, recent molecular and neurochemical research andfuture in mainstream medicine. Ann. N. Y. Acad. Sci. 909, 186–216.
reek, M.J., Vocci, F.J., 2002. History and current status of opioid mainte-nance treatments: blending conference session. J. Subst. Abuse Treat. 23, 93–105.
augwitz, K.L., Offermanns, S., Spicher, K., Schultz, G., 1993. � and � opioidreceptors differentially couple to G protein subtypes in membranes of humanneuroblastoma SH-SY5Y cells. Neuron 10, 233–242.
e Moal, M., Koob, G.F., 2007. Drug addiction: pathways to the disease andpathophysiological perspectives. Eur. Neuropsychopharmacol. 17, 377–393.
ehner, M., Taracha, E., Skorzewska, A., Maciejak, P., Wislowska-Stanek, A.,Zienowicz, M., Szyndler, J., Bidzinski, A., Plaznik, A., 2006. Behavioral,immunocytochemical and biochemical studies in rats differing in their sen-sitivity to pain. Behav. Brain Res. 171, 189–198.
ing, G.S., Umans, J.G., Inturrisi, C.E., 1981. Methadone: radioimmunoassayand pharmacokinetics in the rat. J. Pharmacol. Exp. Ther. 217, 147–151.
iu, J., Nickolenko, J., Sharp, F.R., 1994. Morphine induces c-fos and junB instriatum and nucleus accumbens via D1 and N-methyl-d-aspartate receptors.Proc. Natl. Acad. Sci. USA 91, 8537–8541.

3 cohol
M
M
M
M
M
M
N
N
O
P
P
P
P
P
P
R
R
S
S
S
S
S
S
T
T
T
T
V
V
W
Z
2 E. Taracha et al. / Drug and Al
adlung, E., Haring, C., Crespo, J.A., Saria, A., Grubinger, P., Zernig, G.,2006. Methadone doses upon multiple readmissions to inpatient detoxifica-tion: clinical evidence for very moderate opioid tolerance. Pharmacology78, 38–43.
agalas, Z., De Vry, J., Tzschentke, T.M., 2005. The serotonin/noradrenalinereuptake inhibitor venlafaxine attenuates acquisition, but not maintenance,of intravenous selfadministration of heroin in rats. Eur. J. Pharmacol. 528,103–109.
artin, T.J., Kahn, W.R., Xiao, R., Childers, S.R., 2007. Differential regionaleffects of methadone maintenance compared to heroin dependence on �-opioid receptor desensitization in rat brain. Synapse 61, 176–184.
endez, I.A., Trujillo, K.A., 2008. NMDA receptor antagonists inhibit opiateantinociceptive tolerance and locomotor sensitization in rats. Psychophar-macology 196, 497–509.
urphy, C.A., Russig, H., Pezze, M.A., Ferger, B., Feldon, J., 2003.Amphetamine withdrawal modulates FosB expression in mesolimbicdopaminergic target nuclei: effects of different schedules of administration.Neuropharmacology 44, 926–939.
urray, F., Harrison, N.J., Grimwood, S., Bristow, L.J., Hutson, P.H., 2007.Nucleus accumbens NMDA receptor subunit expression and function isenhanced in morphine-dependent rats. Eur. J. Pharmacol. 562, 191–197.
estby, P., Vanderschuren, L.J., De Vries, T.J., Hogenboom, F., Wardeh, G.,Mulder, A.H., Schoffelmeer, A.N., 1997. Ethanol, like psychostimulantsand morphine, causes long-lasting hyperreactivity of dopamine and acetyl-choline neurons of rat nucleus accumbens: possible role in behaviouralsensitization. Psychopharmacology 133, 69–76.
estler, E.J., 2004. Historical review: molecular and cellular mechanisms ofopiate and cocaine addiction. Trends Pharmacol. Sci. 25, 210–218.
bernier, J.A., Baldwin, R.L., 2006. Establishing an appropriate period ofacclimatization following transportation of laboratory animals. ILAR J. 47,364–369.
aolone, G., Conversi, D., Caprioli, D., Del Bianco, P., Nencini, P., Cabib, S.,Badiani, A., 2007. Modulatory effect of environmental context and drughistory on heroin-induced psychomotor activity and Fos protein expressionin the rat brain. Neuropsychopharmacology 32, 2611–2623.
axinos, G., Watson, C., 1998. The Rat Brain in Stereotaxic Coordinates, fourthed. Academic Press, San Diego.
iepponen, T.P., Mikkola, J.A.V., Ruotsalainen, M., Jonker, D., Ahtee, L.,1999. Characterization of the decrease of extracellular striatal dopamineinduced by intrastriatal morphine administration. Br. J. Pharmacol. 127, 268–274.
ollock, J., Kornetsky, C., 1996. Reexpression of morphine oral stereotypy sixmonths after last morphine sensitizing dose. Pharmacol. Biochem. Behav.53, 67–71.
ontieri, F.E., Calo, L., Di Grezia, R., Orzi, F., Passarelli, F., 1997. Functionalcorrelates of heroin sensitization in the rat brain. Eur. J. Pharmacol. 335,133–137.
opik, P., Wrobel, M., Bisaga, A., 2006. Reinstatement of morphine-conditionedreward is blocked by memantine. Neuropsychopharmacology 31, 160–170.
aith, K., Hochhaus, G., 2004. Drugs used in the tratment of opioid toleranceand physical dependence. A review. Int. J. Clin. Pharmacol. Ther. 42, 191–203.
Z
Dependence 97 (2008) 21–32
yabinin, A.E., Melia, K.R., Cole, M., Bloom, F.E., Wilson, M.C., 1995. Alcoholselectively attenuates stress-induced c-fos expression in rat hippocampus. J.Neurosci. 15, 721–730.
agar, S.M., Sharp, F.R., Curran, T., 1988. Expression of c-fos protein in brain:metabolic mapping at the cellular level. Science 240, 1328–1331.
harp, F.R., Liu, J., Nickolenko, J., Bontempi, B., 1995. NMDA and D1 receptorsmediate induction of c-fos and junB genes in striatum following morphineadministration: implications for studies of memory. Behav. Brain Res. 66,225–230.
iegel, S., Castellan Jr., N.J., 1988. Nonparametric Statistics for the BehavioralSciences, second ed. McGraw-Hill Inc., New York, pp. 213–215.
teffensen, S.C., Stobbs, S.H., Colago, E.E., Lee, R.S., Koob, G.F., Gallegos,R.A., 2006. Contingent and noncontingent effects of heroin on �-opioidreceptor-containing ventral tegmental area GABA neurons. Exp. Neurol.202, 139–151.
tewart, J., Badiani, A., 1993. Tolerance and sensitization to the behavioraleffects of drugs. Behav. Pharmacol. 4, 289–312.
ubhan, F., Pache, D.M., Sewell, R.D.E., 2000. Potentiation of opioid-inducedconditioned place preference by the selective serotonin reuptake inhibitorfluoxetine. Eur. J. Pharmacol. 390, 137–143.
aracha, E., Habrat, B., Chmielewska, K., Baran-Furga, H., 2005. Excretionprofile in dependent patients in relation to route of administration and typeof drug measured in urine with immunoassay. J. Anal. Toxicol. 29, 19–21.
aracha, E., Lehner, M., Wisłowska-Stanek, A., Zienowicz, M., Maciejak, P.,Bidzinski, A., Skorzewska, A., Turzynska, D., Płaznik, A., 2006. Effects ofmethadone and morphine on c-Fos expression in the rat brain: similaritiesand differences. Pharmacol. Rep. 58, 120–124.
rujillo, K.A., Kubota, K.S., Warmoth, K.P., 2004. Continuous administrationof opioids produces locomotor sensitization. Pharmacol. Biochem. Behav.79, 661–669.
urchan, J., Maj, M., Przewłocka, B., 2003. The effects of drugs of abuseon NMDAR1 receptor expression in the rat limbic systems. Drug AlcoholDepend. 72, 193–196.
anderschuren, L.J.M.J., Tjon, G.H.K., Nestby, P., Mulder, A.H., Schoffelmeer,A.N.M., De Vries, T.J., 1997. Morphine-induced long-term sensitization tothe locomotor effects of morphine and amphetamine depends on the temporalpattern of pretreatment regimen. Psychopharmacology 131, 115–122.
anderschuren, L.J.M.J., De Vries, T.J., Wardeh, G., Hogenboom, F.A.C.M.,Schoffelmeer, A.N.M., 2001. A single exposure to morphine induces long-lasting behavioural and neurochemical sensitization in rats. Eur. J. Neurosci.14, 1533–1538.
ard, S.J., Lack, C., Morgan, D., Roberts, D.C., 2006. Discrete-trials heroin self-administration produces sensitization to the reinforcing effects of cocaine inrats. Psychopharmacology 185, 150–159.
arrindast, M.R., Heidari-Darvishani, A., Rezayof, A., Fathi-Azarbaijani, F.,Jafari-Sabet, M., Hajizadeh-Moghaddam, A., 2007. Morphine-induced sen-sitization in mice: changes in locomotor activity by prior scheduled exposure
to GABAA receptor agents. Behav. Pharmacol. 18, 303–310.ernig, G., Wakonigg, G., Madlung, E., Haring, C., Saria, A., 2004. Do verticalshifts in dose response rate-relationships in operant conditioning proce-dures indicate “sensitization” to “drug wanting”? Psychopharmacology 171,349–351.
Copyright © 2022 FDOKUMEN




















