
Fiber projections from trigeminal nucleus caudalis in primate (squirrel mon-key and baboon) J. comp....
15
Fiber Projections from Trigeminal Nucleus Caudalis in Primate (Squirrel Monkey and Baboon) RAJENDRA K. TIWAR12 AND ROBERT B. KING Department of Neurosurgery, State University of New York, Upstate Medical Center, Syracuse, New York 1321 0 ABSTRACT Small radiofrequency lesions were produced in the trigeminal nucleus caudalis and adjacent lateral reticular formation (LRF) in seven squirrel monkeys and four baboons. Frozen brain sections were stained for cellular morphology and for axonal and terminal degeneration. The lesions in pars marginalis and gelatinosa produced terminal degeneration in pars magnocellu- laris and LRF. Pars magnocellularis lesions produced degeneration into the ascending trigeminal intranuclear tract and the adjacent lateral reticular for- mation. Ascending thalamic and reticular projections emanated from LRF lesions. These observations suggest that elements of the LRF adjacent to the nucleus caudalis are an integral part of the trigeminal somesthetic fiber pro- jection system and may subserve the perception of noxious stimuli. The trigeminal nucleus caudalis has been described as a rostra1 continuation of the dorsal horn of the spinal cord (01- szewski, '50; Olszewski and Baxter, '54; Smith et al., '73). In Nissl stained trans- verse sections of the medulla, the trigemi- nal nucleus caudalis has been divided into three parts: pars marginalis, gelatin- osa, and magnocellularis (Olszewski, '50). Recent electrophysiological studies (Nord and Kyler, '68; Nord and Ross, '73) have suggested that the lateral reticular forma- tion, medial to pars magnocellularis, may be an important component in the relay of noxious somesthetic trigeminal stimuli. Although fiber projections to the thala- mus from trigeminal nucleus caudalis have been reported, the findings in such studies have not been uniform. Wallenberg (1896) was not able to demonstrate thalamic de- generation from lesions of the trigeminal nucleus caudalis in the rabbit using the Marchi technique. Van Gehuchten ('01) using the same technique and animal, did report degeneration into the thalamus. Similar discrepancies have persisted in more recent studies. Dunn and Matzke ('68) and Roberts and Matzke ('71) could find only sparse degeneration in the thal- amic nuclei, while other investigators have reported dense trigeminothalamic projec- tion from the nucleus caudalis (Walker, '39; Stewart and King, '63). Each of these investigators, however, reported results J. COMP.NEUR., 158: 191-206. from lesions which were not necessarily limited exclusively in the classic confines of the nucleus caudalis. The terminal pro- jection fields of trigeminothalamic neurons have also not been uniformly described. Trigeminal projections and the fields of termination secondary to small lesions confined to morphologic subdivisions of nucleus caudalis or the adjacent lateral reticular formation, therefore, warranted further investigation and form the basis of this report. Since both nucleus caudalis and the thalamic nuclei are further de- veloped in primates (Walker, '66) than in carnivores, we undertook this study in squirrel monkeys and baboons. METHODS small lesions were produced in the dor- solateral medulla of 28 primates using an LM3 radiofrequency generator (Grass In- strument Co.) and a stainless steel elec- trode with a 50 p uninsulated tip. Under pentobarbital anesthesia, the dorsolateral surface of the medulla was exposed through a midline suboccipital incision. The tuber trigeminum and obex were identified. Le- sions were made at various depths from the surface of the medulla, using a variety 1 This study was supported in part by Training Grant N05-T01-NS-05605 from the National Institute of Neu- rological Diseases and Stroke. 2 Spinalcord Injury Service, VA Hospital, LongBeach, California 90801. 191
Transcript of Fiber projections from trigeminal nucleus caudalis in primate (squirrel mon-key and baboon) J. comp....

Fiber Projections from Trigeminal Nucleus Caudalis in Primate (Squirrel Monkey and Baboon)
RAJENDRA K . TIWAR12 AND ROBERT B. KING Department of Neurosurgery, State University of New York, Upstate Medical Center, Syracuse, N e w York 1321 0
ABSTRACT Small radiofrequency lesions were produced in the trigeminal nucleus caudalis and adjacent lateral reticular formation (LRF) in seven squirrel monkeys and four baboons. Frozen brain sections were stained for cellular morphology and for axonal and terminal degeneration. The lesions in pars marginalis and gelatinosa produced terminal degeneration in pars magnocellu- laris and LRF. Pars magnocellularis lesions produced degeneration into the ascending trigeminal intranuclear tract and the adjacent lateral reticular for- mation. Ascending thalamic and reticular projections emanated from LRF lesions. These observations suggest that elements of the LRF adjacent to the nucleus caudalis are an integral part of the trigeminal somesthetic fiber pro- jection system and may subserve the perception of noxious stimuli.
The trigeminal nucleus caudalis has been described as a rostra1 continuation of the dorsal horn of the spinal cord (01- szewski, '50; Olszewski and Baxter, '54; Smith et al., '73). In Nissl stained trans- verse sections of the medulla, the trigemi- nal nucleus caudalis has been divided into three parts: pars marginalis, gelatin- osa, and magnocellularis (Olszewski, '50). Recent electrophysiological studies (Nord and Kyler, '68; Nord and Ross, '73) have suggested that the lateral reticular forma- tion, medial to pars magnocellularis, may be an important component in the relay of noxious somesthetic trigeminal stimuli.
Although fiber projections to the thala- mus from trigeminal nucleus caudalis have been reported, the findings in such studies have not been uniform. Wallenberg (1896) was not able to demonstrate thalamic de- generation from lesions of the trigeminal nucleus caudalis in the rabbit using the Marchi technique. Van Gehuchten ('01) using the same technique and animal, did report degeneration into the thalamus. Similar discrepancies have persisted in more recent studies. Dunn and Matzke ('68) and Roberts and Matzke ('71) could find only sparse degeneration in the thal- amic nuclei, while other investigators have reported dense trigeminothalamic projec- tion from the nucleus caudalis (Walker, '39; Stewart and King, '63). Each of these investigators, however, reported results
J . COMP. NEUR., 158: 191-206.
from lesions which were not necessarily limited exclusively in the classic confines of the nucleus caudalis. The terminal pro- jection fields of trigeminothalamic neurons have also not been uniformly described.
Trigeminal projections and the fields of termination secondary to small lesions confined to morphologic subdivisions of nucleus caudalis or the adjacent lateral reticular formation, therefore, warranted further investigation and form the basis of this report. Since both nucleus caudalis and the thalamic nuclei are further de- veloped in primates (Walker, '66) than in carnivores, we undertook this study in squirrel monkeys and baboons.
METHODS
small lesions were produced in the dor- solateral medulla of 28 primates using an LM3 radiofrequency generator (Grass In- strument Co.) and a stainless steel elec- trode with a 50 p uninsulated tip. Under pentobarbital anesthesia, the dorsolateral surface of the medulla was exposed through a midline suboccipital incision. The tuber trigeminum and obex were identified. Le- sions were made at various depths from the surface of the medulla, using a variety
1 This study was supported in part by Training Grant N05-T01-NS-05605 from the National Institute of Neu- rological Diseases and Stroke.
2 Spinalcord Injury Service, VA Hospital, LongBeach, California 90801.
191

192 RAJENDRA K. TIWARI
of trajectories through the descending spinal tract below the level of the obex.
Animals were again anesthetized with Nembutal 12-18 days later and fixation initiated by perfusion with normal saline and 10% buffered formalin solution through the thoracic aorta. The calvarium, upper cervical and dorsal laminae were removed and the dura incised. The brain and spinal cord in situ were suspended for two days in 10% formalin. The brain and upper spinal cord were then carefully removed and cut in 8-10 mm slices in a coronal plane which was selected to ac- centuate longitudinally oriented tracts. These blocks were kept in 10% formalin for 2 4 weeks prior to sectioning, marked for lateralization (Tiwari and King, '72) and stored in a solution made up of 2% formalin, 6% glycol and normal saline. Every fifth section was prepared with Nissl (Fernstrom, '58), Fink and Heimer ('67) and Nauta Gygax ('54) stains. Two sec- tions were kept for confirmatory staining when necessary. Degenerating axon ter- minals and preterminals were traced mi- croscopically through these serial sections. Criteria used for the identifkation of de- generating axons and their terminals were those defined by Nauta ('54) and Fink and Heimer ('67). Photomicrographs of repre- sentative fields were obtained and the dis- tribution of degenerating axons and ter- minals plotted in sequential diagrams derived from appropriate sections of stereo- taxic atlases of the brain of squirrel mon- key (Gergen and MacLean, '62) and ba- boon (Davis and Huffman, '69).
OBSERVATIONS
Lesion characteristics Eleven animals (7 squirrel monkeys and
4 baboons) with small lesions limited to subdivisions of nucleus caudalis or the adjacent reticular formation were selected for this study.
Group A: Pars marginalis and substantia gelatinosa
In two squirrel monkeys lesions involved portions of pars marginalis and substantia gelatinosa. The trajectory of the electrodes passed through dorsal elements of the tri- geminal tract terminating at lesions (200- 300 p diameter) in the ventral portion of pars marginalis and pars substantia gel- atinosa 2.5 mm caudal to the obex (fig. la).
AND ROBERT B. KING
Group B : Pars magnocellularis In three squirrel monkeys and three ba-
boons lesions were limited to pars magno- cellularis (fig. lb). In the squirrel monkeys the lesions (200-400 p diameter) were located in the ventral and middle portions of pars magnocellularis and 2 mm below the obex. In the baboons the rostra1 ex- tent of the three lesions (0.1 mm to 1.0 mm diameter) was 1.5 mm caudal to the obex. These lesions in the baboons were slightly more medially positioned in pars magnocellularis than the lesions in the squirrel monkeys. The trajectories of the electrode tracts passed through the de- scending spinal trigeminal tract and varied in their dorsoventral position.
Group C : Lateral reticular formation Lesions in the lateral reticular forma-
tion (fig. lc) of two squirrel monkeys and one baboon varied slightly at the level of the medulla. The density of degeneration
Abbreviations
(-), Degenerating axons (x), Degenerating boutons CM, Centromedian nucleus of thalamus DM, Medial dorsal nucleus of thalamus FN, Facial nucleus IC, Inferior coliculus IL, Intralaminar nuclei of thalamus IOL, Inferior olivary nucleus LRF, Lateral reticular formation MG, Medial geniculate ML, Medial lemniscus MV, Trigeminal motor nucleus N 111, Oculomotor nucleus N VI, Abducens nucleus NA, Ambiguus nucleus NAC, Accessory nucleus NC, Cuneate nucleus NG, Gracile nucleus NH, Hypoglossal nucleus NRL, Lateral reticular nucleus NS, Solitarius nucleus and tract NV, Trigeminal sensory nucleus NVST, Vestibular nucleus PER, Peripeduncular dorsal nucleus PF, Parafascicular nucleus of thalamus PGL, Pars gelatinosa PMG, Pars magnocdlularis PMR, Pars marginalis PYR, Pyramidal tract RN, Red nucleus SC, Superior colliculus SCP, Superior cerebellar peduncle SN, Substantia nigra TV, Spinal trigeminal tract VH, Ventral horn of the spinal cord VPL, Ventral posterolateral nucleus of thalamus VPM, Ventral posteromedial nucleus of thalamus
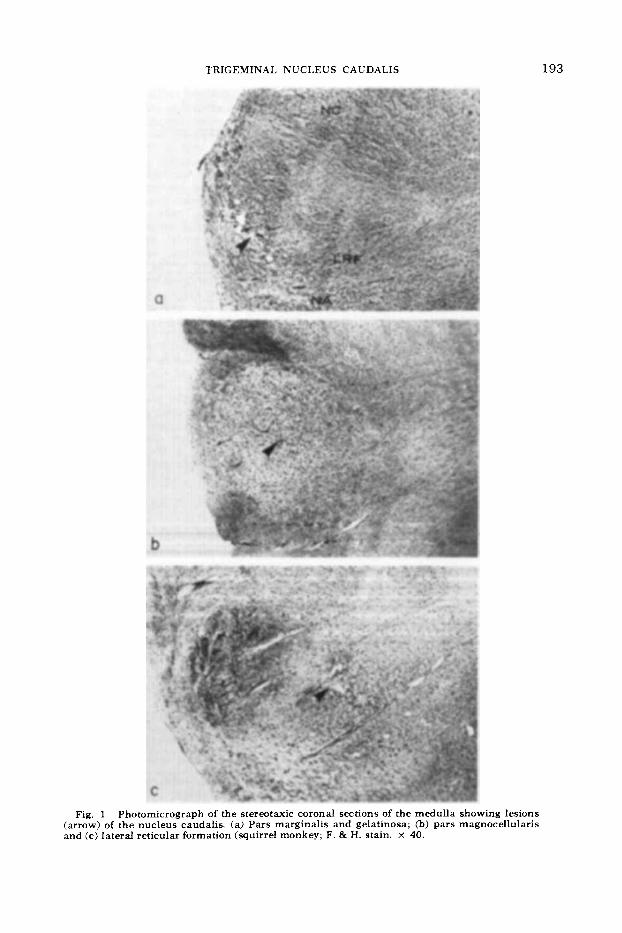
TRIGEMINAL NUCLEUS CAUDALIS
Fig. 1 Photomicrograph of the stereotaxic coronal sections of the medulla showing lesions (arrow) of the nucleus caudalis. (a) Pars marginalis and gelatinosa; (b) pars magnocellularis and (c) lateral reticular formation (squirrel monkey; F. & H. stain. X 40.
193

194 RAJENDRA K. TIWARI
in certain tracts varied somewhat between lesions.:%
Distribution of axonal and terminal degeneration
Group A lesions From lesions in pars marginalis and
substantia gelatinosa (fig. 2A) degenerat- ing axons were traced medially and cau- dally for a short distance in the medulla oblongata. They terminated around the cells of the various laminae of the nucleus caudalis, cuneate and accessory nuclei, and the ventral horn cells of the upper cervical cord ipsilaterally. The axons des- tined to terminate in the accessory nucleus and ventral horn cells passed caudally and ventromedially between the fascicles of the crossed pyramidal tract. The degener- ation in reticular formation and nucleus solitarious was bilateral. At the level of the lesion, the deeper cells of pars magno- cellularis and lateral reticular formation also showed degenerating terminals. The degeneration in the substantia gelatinosa was localized in the neighborhood of the lesion adjacent to the concave surface of the descending tract. The relative density of degeneration was marked in ipsilateral reticular formation, particularly in an area ventromedial to nucleus caudalis (fig. 4b). A thin bundle of degenerating axons crossed the midline ventral to the hypo- glossal nuclei and terminated around cells of contralateral medial reticular forma- tion. Ascending degeneration from these lesions extended only a short distance in the trigeminal spinal tract and nucleus to end in terminal degeneration around neighboring neurons of the pars marginalis and gelatinosa.
Group B lesions Degeneration from lesions in pars mag-
nocellularis of nucleus caudalis (fig. 2B) terminated primarily in the ventromedial part of the medullary reticular formation. The density of degenerating boutons in this area was striking. A number of de- generating axons also coursed medially, crossed the midline through the dorsal commissure, and terminated in the contra- lateral lateral reticular formation medial to trigeminal nucleus caudalis. Degener- ating fibers were also traced (with in- creased terminal degeneration compared
AND ROBERT B. KING
to Group A lesions) bilaterally, at solitary nuclei and scattered neurons in the dorsal commissural gray. Terminal degeneration was noted bilaterally in the hypoglossal nuclei. On the side of the lesions, some axons passed ventrally and medially to terminate in the nucleus ambiguus and the lateral reticular nucleus.
Rostra1 to the lesion degenerating axons, within the ventral part of the ipsilateral trigeminal tract and nucleus, ascended to the level of the facial nucleus. Here the fibers became diffuse; some turned ven- tromedially to end in the ipsilateral facial nucleus, while others terminated in the trigeminal main sensory nucleus, trigemi- nal motor nucleus and adjacent reticular formation. As this tract ascended, degen- erating terminals were also seen at neu- rons of the intervening trigeminal spinal nuclei (caudalis, interpolaris and oralis) but were not seen at the trigeminal mes- encephalic nucleus. The ipsilateral retic- ular formation of the medulla and pons also received degenerating terminals sub- jacent to spinal trigeminal nuclei.
Neither axonal nor terminal degenera- tion were evident in the mesencephalon or diencephalon from group B lesions.
Group C lesions The distribution of degenerating axons
3B6. The lesion in the ventral part of subtrigeminal reticular gray dorsal to nucleus retroambigualis was 1 mm in transverse diameter and 2 mm in its rostro- caudal extension. The electxode entered the medulla through the descending spinal trigeminal tract 3 mm below the obex with a traectory at the level of the lesion directed ventromedi ally.
S16. The trJectory of the electrode started 2 mm b e low the obex into the trigeminal nucleus caudalis through which it extended ventromedially into the lesion in the lateral reticular formation. The lesion was 1 mm in transverse and rostral caudal diameters. I t was centered in the ventral part of the lateral reticular formation, dorsolateral to nucleus ambiguus.
S17. The lesion in the ventral part of the lateral r e ticular formation at the level of the obex and dorsal to nucleus ambiguus. It was 1 mm in all diameters, and was centered slightly more medially in the reticu- lar formation and extended slightly more rostrally than the lesion in S16.
Fig. 2 Diagrams of the brain stem (stereotaxic coronal plane) showing axonal (-) and terminal (x) degenerations. (A) Lesions of pars marginalis and substantia gelatinosa. Degeneration largely confined to the medulla. Note the course of the degenerating axons through the crossing pyra- midal tracts. (B) Lesions of pars magnocellularis. Extension of degeneration into ascending intra- trigeminal tract terminating at main sensory nucleus and trigeminal motor and facial nuclei (see text).

TRIGEMINAL NUCLEUS CAUDALIS 195
B Figure 2

196 RAJENDRA K. TIWARI
and terminals, associated with the lesions of the reticular formation, extended into contralateral thalamic nuclei, mesenceph- alon and pontomedullary reticular forma- tion (fig. 3C). It was in this respect dis- tinctly different from that which occurred incident to lesions in more superficial ele- ments of nucleus caudalis. Unlike the pars marginalis, gelatinosa and magnocellu- laris lesions dense degenerating axons were noted in the trigeminothalamic tract (fig. 5). Most of the degeneration in the trigeminothalamic tract was in the form of discrete fiber bundles which traversed ventromedially towards the lower third of the ipsilateral inferior olivary nucleus. These fascicles passed through inferior olivary nucleus and then crossed the mid- line in the interolivary region to lie in the ventral part of the contralateral medial lemniscus. In the midbrain, some of these fibers passed through the superior cere- bellar peduncle. In the thalamus, trigem- inothalamic fibers passed through the cau- dolateral part of the centromedianum (CM) into internal medullary lamina to termi- nate in the ventral posteromedial (VPM) and intralaminar (IL) nuclei.
In animal S17, where the lesion ex- tended more medially into the medullary reticular gray, the terminal degeneration extended beyond VPM into the medial portion of ventral posterolateral nucleus (VPL). In the mesencephalon, degenerat- ing axons branched out from the ascend- ing crossed trigeminothalamic tract to end in terminal degeneration in the adjacent mesencephalic reticular nuclei and ventro- lateral part of the periaqueductal gray. Occasionally some scattered terminal de- generation was noted in the region of the tectum of midbrain.
The ascending intratrigeminal nuclear tract was again evident and was even more prominent medially than in Group B le- sions because of the addition of ascending degeneration in reticular formation. It extended with terminal degeneration at all levels of the spinal trigeminal nuclei and main sensory nucleus rostrally into the mesencephalon to end in the ipsilat- era1 mesencephalic tegmental reticular nuclei and ventrolateral part of the peri- aqueductal gray.
Degeneration from the lesions of lateral reticular formation was observed in the
AND ROBERT B. KING
contralateral trigeminal nuclei and was most pronounced at the level of the lesion. It also extended into the dorsal column nuclei bilaterally and ipsilateral inferior vestibular nucleus.
The distribution of degeneration in med- ullary reticular formation was diffuse and bilateral. A distinct bundle of degenerat- ing fibers was observed in animals B6 and S16 extending dorsomedially towards the ipsilateral hypoglossal nucleus. Ventral to the hypoglossal nucleus, these fibers turned medially towards the midline where they broke into several thinner fascicles. These fascicles then turned ventrolateral and rostral, crossed the midline and terminated into con;ralateral medullary reticular formation.
DISCUSSION
The small lesions which we have studied, confined to the subdivisions of the tri- geminal nucleus caudalis, have made pos- sible an analysis of patterns of degenera- tion from these subdivisions. Rexed ('52) on morphological basis divided the dorsal horn of the spinal cord into six laminae. Trigeminal nucleus caudalis is morpho- logically similar to elements of the dorsal horn of the spinal cord. Classically (01- szewski, '50), however, only three subdivi- sions were made: pars marginalis, gelatin- osa, and magnocellularis. Pars marginalis and gelatinosa of nucleus caudalis resem- ble lamina I and I1 (of the dorsal horn of the spinal cord respectively). Pars magno- cellularis has not shown further neuronal segregation but contained an admixture of neurons of various sizes (10-30 r ) . In degeneration studies, following trigeminal rhizotomy, however, the portion of the pars magnocellularis adjacent to pars gel- atinosa has shown a heavy concentration of degenerating terminals (Kerr, '70). Thus it resembles lamina I11 of the dorsal horn of the spinal cord, which showed similar distribution of degenerating terminals fol- lowing dorsal root section (Heimer and Wall, '68).
The deeper elements of pars magnocel- lularis have more recently been considered
~ ~~~
Fig. 3 Diagrams of brainstem and thalamus showing degeneration from lesions of LRF subja- cent to pars magnocellularis. Projections extended to midbrain and contralateral thalamic nuclei (see text).

TRIGEMINAL NUCLEUS CAIJDALIS 197

198 RAJENDRA K. TIWARI AND ROBERT B. KING
analagous to lamina IV. Further subdivi- sion of the trigeminal nucleus caudalis and adjacent reticular formation in the medulla oblongata has largely been de- pendent on electrophysiological studies (Mosso and Kruger, '73; Nord and Ross, '73). The present study, which defines pro- jections of degenerating fibers from the lateral reticular formation (LRF) to the midbrain and thalamus, implies that ele- ments of nucleus caudalis (which are analagous to Rexeds lamina V and VI of the spinal cord) lie within the LRF sub- jacent to pars magnocellularis.
In Golgi studies of the trigeminal nu- cleus caudalis, Cajal ('09) observed axons of marginal cells which projected into trigeminothalamic tract but final termina- tion of these neurons were not determined. Similar studies by Scheibel and Scheibel ('68) on the dorsal horn of the spinal cord revealed axons of the marginal cells run- ning in the ipsilateral dorsolateral white matter rather than the contralateral an- terolateral ascending tracts. This is in contrast to the findings of Kuru ('49) on retrograde cell degeneration studies where chromatolysis of the marginal cells of the dorsal horn resulted following sectioning of the contralateral anterolateral column (spinothalamic) of the spinal cord in man. Kuru's anatomical findings are supported by electrophysiological studies of Trevino et al. ('73) who found antidromic responses from neurons in the pericoronal region (lamina I and 11) of the dorsal horn follow- ing stimulation of the contralateral an- terolateral column of the spinal cord in cat.
A further physiologic analogy exists be- tween the periphery of the dorsal horn of the spinal cord and the trigeminal nucleus caudalis in response to noxious stimula- tion. Christensen and Per1 ('70) found concentration of neuronal units respon- sive to noxious stimulation in the mar- ginal cell layer of the dorsal horn of the spinal cord of cat. Mosso and Kruger ('73) found noxious units in the pars marginalis of the trigeminal nucleus caudalis in the cat, although the density of such neurons was sparse.
It is possible that sparsely distributed marginal neurons of the nucleus caudalis (Kerr, '70) may have escaped destruction
in Group A lesions which were ventrally located. However, marginal neurons or their process must have been involved in the electrode trajectory or in the lesions of the Group B, yet no degeneration in the thalamus was observed in conjunction with any of these lesions. It appears then that the marginal cells of the nucleus caudalis do not directly project to the mid- brain or thalamus. Axons of marginal cells and elements of substantia gelatinosa pro- ject into deeper layers of trigeminal nu- cleus caudalis (pars magnocellular) and the adjacent LRF, and a short distance into the spinal trigeminal tract. This is in accordance with the Golgi study of the dorsal horn of the spinal cord in which the axons or axon collaterals of the gel- atinosa neurons are spread between the lamina I1 and V (Matsushita, '69). Ter- minal degeneration in LRF from group A lesions was denser than that which follow trigeminal rhizotomy (Kerr, 'Sl), suggest- ing that the marginal and perhaps gela- tinosa neurons primarily project to synaptic terminations at LRF neurons.
The ascending degeneration in the tri- geminal spinal tract observed adjacent to these superficial lesions extended only a few sections rostrally reflecting axons or axon collaterals either from neurons of pars gelatinosa and marginal cells or deeper neurons which are known to con- tribute into Lissaurs' tract in the dorsal horn of the spinal cord (Scheibel and Schei- bel, '68; Matsushita, '69). These fibers may represent the short trigeminal intra- nuclear projections observed by Roberts and Matzke ('71).
Cajal ('09) in Golgi preparations of brain stem, described axons from substantia gelatinosa of the trigeminal spinal nucleus ascending to the main sensory nucleus. Stewart and King ('63) found this tract to arise from lesions of the trigeminal nu- cleus caudalis extending rostrally up to the midbrain. This tract was further con- firmed by degeneration studies to have neurons of origin in the trigeminal nu- cleus caudalis (Dunn and Matzke, '68; Roberts and Matzke, '71). Dunn and Matzke ('68) proposed short and long in- tranuclear projection systems as the de- generation at the level of the trigeminal main sensory nucleus became sparse. Later

TRIGEMINAL NUCLEUS CAUDALIS 199
Roberts and Matzke ('71) observed and found no decrease in degeneration ros- trally.
Our observations confirm the existence of the ascending trigeminal intranuclear projection system emanating from the nu- cleus caudalis. The long ascending degen- eration in the tract and nucleus were largely confined in the ventral part of the trigeminal complex. This location of as- cending degeneration was also observed by Dunn and Matzke ('68) in marmoset monkey and by Roberts and Matzke ('71) in sheep, although their lesions were larg- er than ours and also involved the dorsal part of the nucleus caudalis. It appears that the ascending intranuclear fibers segregate in the ventral part of the tri- geminal complex. Matsushita ('69) in Golgi preparations found neurons in lamina 111 and IV of spinal dorsal horn with axons projecting into the dorsal part of the lat- eral funiculus (ventral to the dorsal horn of the spinal cord) analogous to ascending projections from pars magnocellularis in nucleus caudalis. In this respect, also, pars magnocellularis is similar to lamina 111 and IV of the dorsal horn.
Following trigeminal tractotomy, an al- teration in the response of the rostral trigeminal nuclei (interpolaris, oralis and main sensory) on peripheral stimulation has been observed (Scibetta and King, '69; Young and King, '72). Because of the close proximity to the spinal tract, the ascending intranuclear fibers must have been involved as a result of tractotomy in the above experiments. The anatomic sub- strate for these observations may well be represented in the ascending trigeminal intranuclear projection systems originat- ing in pars magnocellularis.
Functionally, the LRF adjacent to tri- geminal nucleus caudalis appears anal- agous to the basal laminae (V and VI) of the dorsal horn of the spinal cord. This supposition is consistent with the report of neurons in LRF responsive to noxious stimulation of the face in cat (Gordon et al., '61; Burton, '68), in rat (Nord and Kyler, '68) and monkey (Nord and Ross, '73). The anatomical findings of degener- ating axons into contralateral trigemino- thalamic tract from LRF lesions resemble the physiological studies in the basal lam-
inae of the dorsal horn and its projection into the spinothalamic tract of the spinal cord (Trevino et al., '73). Neurons were found in the laminae IV and V of the dorsal horn antidromically responsive on stimulation of the contralateral spinothal- amic tract and VPL. Matsushita ('69) in Golgi studies traced the axon from the neurons at lamina V to project into con- tralateral funiculus of the spinal cord in cat. The LRF lesions in this study produce axons and terminal degeneration into trigeminothalamic and reticular system. It is probable, therefore, that there is an admixture of somesthetic and reticular neurons in the LRF subjacent to pars mag- nocellularis.
Wallenberg (1896) did not find thalamic degeneration following lesions in nucleus caudalis in rabbit using Marchi techniques. This he attributed to the superficial loca- tion of the lesion in the nucleus caudalis. Dunn and Matzke ('68) in sheep and Roberts and Matzke ('71) in marmoset monkey, using Nauta technique, found sparse thalamic degeneration from the lesions of the nucleus caudalis and thought that the nuclei which produce trigemino- thalamic projections may be located in the rostral part of the spinal trigeminal nucleus. The lesions described by others in which thalamic projections from nu- cleus caudalis were noted (Walker, '39; Stewart and King, '63) were large enough to extend into LRF and the medulla.
Our observations concerning the dis- tribution of trigeminal projections to VPM are in accord with Walker ('39) and Stew- art and King ('63). The question then arises whether this termination into VPM belongs to the lemniscal system or spino- thalamic system for the face. It is clear that the spinal trigeminal nucleus which functionally must belong to the spinothal- amic system also possesses some charac- teristics of lemniscal neurons (Kruger and Michael, '62). This duality of somethetic representation has been further stressed by the studies of Mosso and Kruger ('73) who found neurons responsive to noxious and non-noxious facial stimuli in nucleus caudalis in cat, and by Nord and Ross ('73) in the adjacent LRF in monkey. The VPM may have dual functional projection, i.e., lemniscal and spinothalamic system. Such

200 RAJENDRA K . TIWARI AND ROBERT B. KING
Fig. 4 Terminal degeneration from pars marginalis and gelatinosa lesions; (a) sparse degeneration in pars gelatinosa and magnocellularis and (b) dense degeneration around the cells of the lateral reticular formation (squirrel monkey; F. & H . x 480).

TRIGEMINAL NUCLEUS CAUDALIS
Fig. 5 Representative section of the medulla rostra1 to lesions (a) of pars magnocellularis showing paucity of degenerating axons in the trigeminothalamic tracts and; (b) of LRF with an abundance of degenerating axons (squirrel monkey; Nauta. X 400).
20 I

202 RAJENDRA K. TIWARI AND ROBERT B. KING
a conclusion does not necessarily indicate that VPM is a thalamic relay for trigemi- nal pain. The latter may be diffuse as suggested by Poggio and Mountcastle (‘60). Trigeminal projection into the centrome- dianum and intralaminar nuclei was ob- served from the nucleus caudalis by Stew- art and King (’63). However, whether these were degenerating axons “en pas- sage’’ or terminals could not be confirmed on the basis of Nauta stain alone. The use of the technique to selectively enhance the staining of the terminal degeneration (Fink and Heimer, ’67) confirms the dense trigeminal projection into the intralaminar nuclei from the Group C lesions. No cer- tain terminal degeneration in the centro- medianum was noted. In the lateral part of centromedianum axonal degeneration and scattered terminal degeneration (fig. 6a) were evident adjacent to the internal medullary lamina. It appears that these neurons are perhaps part of trigeminoin- tralaminar projections in primate. Unlike Stewart and King’s (’63) finding, no degen- eration was noted in the medial geniculate nuclei.
Terminal degeneration in VPL was lo- calized into the area just rostrolateral to VPM. This part of the VPL will represent somatotopically thalamic projection of the cervical spinal segments. The dorsal horn of the spinal cord blends with nucleus caudalis and upper dorsal root and de- scending spinal tract overlap in their pro- jection in the rostral cervical spinal cord (Kerr, ’61). The fibers representing upper cervical segments may pass through LRF and may be involved in the lesion as they ascend to thalamus. An increase in VPL degeneration from S17 supports cervical spinal origin of these fibers.
Following trigeminal rhizotomy, preter- minal degeneration is sparse in the retic- ular formation of the medulla oblongata (Kerr, ’61). These primary afferent ter- minations are localized to ipsilateral retic- ular formation adjacent to the trigeminal nucleus. Progressive increase in the ax- onal and terminal degeneration in the reticular formation with extension of nu- cleus caudalis lesions medially into the medulla suggests cumulative origin of these projections from neurons of nucleus caudalis and the adjacent LRF.
The projection from the trigeminal spin- al nucleus to the medullary reticular for-
mation ha- been described in both Golgi studies (Cajal, ’09) and silver degenera- tion studies (Nauta and Kuyper, ’58; Car- penter and Hanna, ’61; Stewart and King, ’63; Dunn and Matzke, ’68). Nauta and Kuyper (’58) reported bilateral projections into reticular formation. Predominantly ipsilateral projection to medullary reticu- lar formation was observed by Stewart and King (’63) and Dunn and Matzke (‘68). Cajal (’09) reported predominantly contra- lateral trigemino-reticular projection. This was substantiated by Carpenter and Hanna (‘61) from lesions in the rostral part of the spin a1 trigeminal nucleus.
Our preparations supported the obser- vation which suggests predominantly ipsi- lateral trigemino-reticular projections from the lesions localized to nucleus caudalis. However, the pattern of reticular projec- tion changed when the lesion extended to the LRF. In this circumstance there was a distinct increase in the projections to the reticular formation in the contralat- era1 side, especially above the level of the obex. It resembled projections from the rostral trigeminal lesion described by Nauta and Kuypers (’58) and Carpenter and Hanna (’61). A t the level of the pyra- midal decussation a distinct band of de- generation crossed to the opposite medul- lary reticular formation from the LRF. These fibers resembled the beginning of the dorsal trigeminothalamic tract from nucleus caudalis described by Wallenberg (1896) and Van Gehuchten (’Ol), but they clearly ended in synaptic terminations around cells of the contralateral medullary reticular formation in this study. The pres- ence of these degenerating fibers bridging between the reticular formation of the two halves of the medulla, only at the level of the pyramidal decussation, suggests that these may by “commissural” fibers which have been displaced dorsally and aggre- gated in the form of compact bundles over- lying the decussating pyramidal fibers.
The projection into mesencephalic retic- ular formation (Stewart and King, ’63) and to the vestibular nuclei (Dunn and
Fig. 6 Degenerations in contralateral thalamic nuclei form the lateral reticular formation lesions; (a) sparse terminal degeneration (arrow) in CM; (b) dense terminal degeneration in the intralam- inar nuclei. Axonal degeneration is also seen as fiber en passage to terminate into terminal degen- eration in the (c) VPM (squirrel monkey, F. & H. X 480).
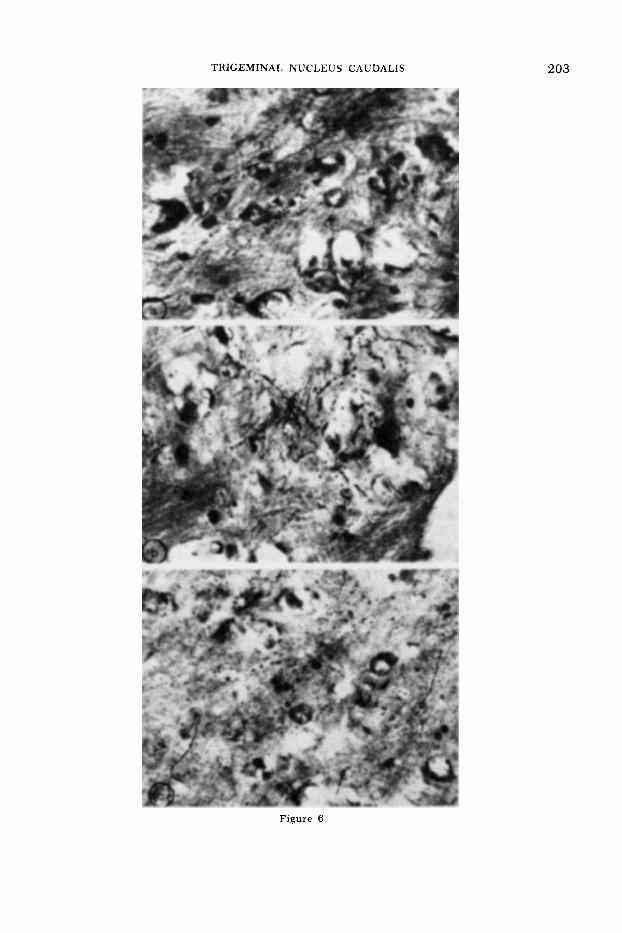
TRIGEMINAL NUCLEUS CAUDALIS 203
Figure 6

204 RAJENDRA K. TIWARI
Fig. 7 Schematic drawing of a transverse s e e tion of the dorsolateral medulla to show the pro- posed homology between the laminae ( I to VI) of the dorsal horn of the spinal cord and the trigem- inal nucleus caudalis and aaacen t lateral reticular form a tion.
Matzke, ’68) were not associated with ob- served degeneration in the trigeminal in- tranuclear ascending tract until degen- eration involved the LRF (Group C). I t appears that medial to the intratrigeminal intranuclear tract, a “reticular” compo- nent is added, when the lesion extends deep into medullary reticular formation with more diffuse projections.
Spinal trigeminal projections terminat- ing in the mesencephalon have been re- ported by several authors (Carpenter and Hanna, ’61; Nauta and Kuyper, ’68; Stew- art and King, ’63; Dunn and Matzke, ’68). Our study indicates that the neurons of origin for these projections are elements of the LRF subjacent to pars magnocellu- laris. Degenerating fibers ascended medial to the ipsilateral ascending trigeminal in- tranuclear tract. We cannot exclude the possibility on the basis of our material that some of this degeneration may be related to ventromedial location of our LRF le- sions resulting in interruption of medial fibers of the ascending spinotectal tracts, which project to the ipsilateral midbrain.
LITERATURE CITED
Burton, H. 1968 Somatic sensory properties of caudal bulbar reticular neurons in the cat (felis domestica). Brain Res., 1 I : 357-372.
AND ROBERT B. KING
Cajal, S. R. Y . 1909 Histologie du Systeme Nerveux de L’Homme et des Vertebres. Paris Maloine. Reprinted 1952. Ceniejo Superior d e Investigaciones Centificias, Institute Ramon Y Cajal.
Carpenter, M. D., and G. R. Hanna 1961 Fiber projections from the spinal trigeminal nucleus in the cat. J. Comp. Neur., 1 17: 117-125.
Christensen, B. N., and E. R. Per1 1970 Spinal neurons specifically excited by noxious or ther- mal stimuli. Marginal zone of the dorsal horn. J. Neurophysiol., 33: 293-307.
Davis, R., and R. Huffman 1969 A stereotaxic atlas of the brain of the baboon. University of Texas Press. Austin - London.
Dunn, J . D., and H. A. Matzke 1968 Efferent fiber connections of the marmoset (Oedipomidas oedipus) trigeminal nucleus caudalis. J . Comp. Neur., 133: 429438 .
Fernstrom, R. 1958 A durable nissl stain for frozen and paraffin sections. Stain Tech., 33: 175-1 76.
Gergen, J . A, , and P. D. MacLean 1962 A ster- eotaxic atlas of the squirrel monkey’s brain (Saimir sciureus) USHEW, PHS Publication 933, Washington.
Gordon, G., S. Landgren and W. A. Seed 1961 The functional characteristics of single cells in the caudal part of the spinal nucleus of the trigeminal nerve of the cat. J . Physiol., 158. 544-588.
Fink, R., and L. Heimer 1967 Two methods for selective impregnation of degenerating axons and their synaptic endings in the central nerv- ous system. Brain Res., 4 : 369-374.
Heimer, L., and P. D. Wall 1968 The dorsal root distribution to the substantia gelatinosa of the rat with a note on the distribution in the cat. Exp. Brain Res., 6 . 8S90.
Kerr, F. W. L. 1961 Structural relation of the trigeminal spinal tract to upper cervical roots and the solitary nucleus in the cat. Exp. Neurol., 4 : 134-148.
1970 The fine structure of the subnu- cleus caudalis of the trigeminal nerve. Brain Res., 23: 129-145.
Kruger, L . . and F. Michel 1962 A morpholog- ical and somatotopic analysis of single unit ac- tivity in the trigeminal sensory complex of the cat. Exp. Neurol., 5: 139-156.
Matsushita, M. 1969 Some aspects of the inter- neuronal connections in cats’ spinal gray mat- ter. J. Comp. Neur., 136: 57-80.
Mosso, J . A., and L. Kruger 1973 Receptor categories represented in spinal trigeminal nu- cleus caudalis. J. Neurophysiol., 36: 472-488.
Nauta, W. J. H., and P. A. Gygax 1954 Silver impregnation of degenerated axons in the cen- tral nervous system. A modified technique. Stain Tech., 29: 91-93.
Nauta, W. J. H., and H. G. J. M. Kuypers 1958 Some ascending pathways in the brainstem reticular formation. Reticular formation of the brain. Little, Brown and Co., Boston, pp. 3-30.
A single unit analysis of trigeminal projections to bulbar re ticular nuclei of the rat. J. Comp. Neur., 134: 485495 .
Nord, S. G., and G. S. Ross 1973 Responses of
Nord, S. G., and H. J. Kyler 1968

TRIGEMINAL NUCLEUS CAUDALIS 205
trigeminal units in the monkey bulbar lateral reticular formation to noxious and non-noxious stimulation of the face: experimental and theo- retical considerations. Brain Res., 58: 385-399.
Olszewski, J. 1950 On the anatomical and func- tional organization of the spinal trigeminal nu- cleus. J. Comp. Neur., 92: 401413 .
Olszewski, J., and D. Baxter 1954 Cytoarchi- tecture of the human brain stem. S. Karger, New York, Basel.
Poggio, F. G . . and V. B. Mountcastle 1960 A study of the functional contribution of the lem- niscal and spinothalamic systems to somatic sensibility. Bull. Johns Hopkins Hosp., 106: 266-3 16.
Rexed, B. 1952 The cytoarchitectonic organiza- tion of the spinal cord in the cat. J. Comp. Neur., 96: 415496 .
Roberts, P. A, , and H. A . Matzke 1971 Projec- tions of the subnucleus caudalis of the trigem- inal nucleus in the sheep. J. Comp. Neur., 141: 273-282.
Scheibel, M. E., and A . B. Scheibel 1968 Ter- minal axonal patterns in cat spinal cord. 11. The dorsal horn. Brain Res., 9. 32-58.
Scibetta, C. J., and R. B. King 1969 Hyperpo- larizing influence of trigeminal nucleus caudalis on primary afferent preterminals in trigeminal nucleus oralis. J. Neurophysiol., 32: 22S238.
Smith, 0. A . , K. G. Kastella and D. C. Randall 1973 A stereotaxic atlas of the brainstem for
Macaca mulatta in the sitting position. J . Comp. Neur., 145: 1-24.
Stewart, W., and R. B. King 1963 Fiber pro- jections from the nucleus caudalis of the spinal trigeminal nucleus. J . Comp. Neur., 121 271- 286.
Trevino, D. L.. J. D. Coulter and W. D. Wills 1973 Location of cells of origin of spinothal- amic tract in Iumbar enlargement of the mon- key. J. Neurophysiol., 36; 750-761.
Tiwari, R. K., and R. B. King 1971 Needle- track reference marks of India ink for right-left orientation of brain sections. Stain Tech., 46: 321.
Van Geuhuchten, A. 1901 Recherches sur les voies sensitives centrales la voie central du trijemeau. Nevraxe, 3: 237-261.
Walker, A. E. 1939 The origin, course and ter- mination of the secondary pathways of the trigeminal nerve in primates. J. Comp. Neur., 7 1 . 59-89.
1966 Internal structure and afferent- efferent relations of the thalamus. In: The Thal- amus. D. Purpura and M. Yahr, eds., Columbia University Press, New York. pp. 1-12.
Wallenberg, A. 1896 Die secundare bahn des sensiblen trigeminus. Anat. Anz., 12: 95-100.
Young, R. F., and R. B. King 1972 Excitability changes in trigeminal primary afferent fibers in response to noxious and non-noxious stimuli. J. Neurophysiol., 35. 87-95.


![Mapping muscarinic receptors in human and baboon brain using [N-11C-methyl]-benztropine](https://static.fdokumen.com/doc/165x107/6344f35df474639c9b049d90/mapping-muscarinic-receptors-in-human-and-baboon-brain-using-n-11c-methyl-benztropine.jpg)

















