
Four Irritating Odorants Target the Trigeminal Chemoreceptor TRPA1
10
Four Irritating Odorants Target the Trigeminal Chemoreceptor TRPA1 Paige M. Richards & Erik C. Johnson & Wayne L. Silver Received: 8 June 2010 / Accepted: 9 September 2010 / Published online: 28 September 2010 # Springer Science+Business Media, LLC 2010 Abstract The trigeminal nerve responds to a variety of nociceptive stimuli, including many chemicals that activate the olfactory system at lower concentrations. However, the mechanisms by which specific odorants activate the trigeminal nerve are largely undetermined. We used an integrative approach to determine whether TRPA1 channels were the target of eight olfactory stimuli known to activate the trigeminal nerve. In a mammalian cell line stably expressing the human TRPA1 channel, we observed significant increases in intracellular calcium levels upon application of α-terpineol, amyl acetate, benzaldehyde, and toluene. Notably, these responses were greatly reduced when these chemicals were applied in conjunction with the TRPA1 inhibitor HC-030031. To determine whether TRPA1 is required for detecting these odorants as irritants in vivo, we evaluated physiological and behavioral responses to these stimuli in mice lacking the TRPA1 channel. A decline in respiration was seen in wild-type but not TRPA1 knockout mice upon exposure to α-terpineol, benzaldehyde, and toluene. Furthermore, we observed either attenuated or absent avoidance behaviors in TRPA1 knockout mice compared to wild-type mice when exposed to α-terpineol, amyl acetate, and benzaldehyde. Our results show that TRPA1 is a molecular target for four different odorants—α-terpineol, amyl acetate, benzaldehyde, and toluene. Additionally, this study suggests that TRPA1 is required for detection of α-terpineol, benzaldehyde, and toluene as trigeminal irritants. However, TRPA1 is likely only one of multiple trigeminal receptors detecting amyl acetate. Keywords Behavioral aversion . Intracellular calcium . Irritant . Respiratory pause . Trigeminal nerve . TRPA1 Introduction Humans detect chemical stimuli in the environment through three distinct sensory systems—olfaction, gustation, and chemethesis. Chemesthesis, the sense of irritation due to chemical stimuli, is mediated by a subset of pain, mechanosensory and temperature-sensitive nerve fibers (Bryant and Silver 2000; Bhattacharya et al. 2008). In mammals, the trigeminal nerve—which innervates the mouth, nose, and eyes—is a major anatomical site of chemesthesis. The small-diameter afferent fibers of the trigeminal nerve, both myelinated (Aδ) and unmyelinated (c), detect noxious chemical stimuli (Julius and Basbaum 2001). Activating trigeminal nerve fibers triggers a variety of protective reflexes such as sneezing, bronchoconstric- tion, increased secretions (e.g., nasal mucus and saliva), and decreased respiration rate (Alarie 1973). Many odorants stimulate trigeminal receptors in the nasal cavity (Tucker 1971), yet olfactory studies do not always consider possible trigeminal contributions to odor perception. Trigeminal thresholds are generally higher than olfactory thresholds (Cometto-Muñiz and Cain 1990), so it is often assumed that testing odorants at low concentrations will eliminate possible trigeminal input. However, the trigeminal and olfactory systems are heavily interrelated (Brand 2006). For example, previous trigeminal stimulation can decrease olfactory thresholds (Jacquot et al. 2004a) and loss of olfactory function reduces trigeminal sensitivity (Hummel et al. 1996; Frasnelli and Hummel 2007). These interactions justify efforts to understand the mechanisms by which olfactory stimuli are involved in trigeminal nerve stimulation in an effort to strengthen future studies of olfactory P. M. Richards (*) : E. C. Johnson : W. L. Silver Department of Biology, Wake Forest University, Winston-Salem, NC 27109, USA e-mail: [email protected] Chem. Percept. (2010) 3:190–199 DOI 10.1007/s12078-010-9081-1
-
Upload
catawbaschools -
Category
Documents
-
view
4 -
download
0
Transcript of Four Irritating Odorants Target the Trigeminal Chemoreceptor TRPA1

Four Irritating Odorants Target the TrigeminalChemoreceptor TRPA1
Paige M. Richards & Erik C. Johnson & Wayne L. Silver
Received: 8 June 2010 /Accepted: 9 September 2010 /Published online: 28 September 2010# Springer Science+Business Media, LLC 2010
Abstract The trigeminal nerve responds to a variety ofnociceptive stimuli, including many chemicals that activatethe olfactory system at lower concentrations. However, themechanisms by which specific odorants activate thetrigeminal nerve are largely undetermined. We used anintegrative approach to determine whether TRPA1 channelswere the target of eight olfactory stimuli known to activatethe trigeminal nerve. In a mammalian cell line stablyexpressing the human TRPA1 channel, we observedsignificant increases in intracellular calcium levels uponapplication of α-terpineol, amyl acetate, benzaldehyde, andtoluene. Notably, these responses were greatly reducedwhen these chemicals were applied in conjunction with theTRPA1 inhibitor HC-030031. To determine whetherTRPA1 is required for detecting these odorants as irritantsin vivo, we evaluated physiological and behavioralresponses to these stimuli in mice lacking the TRPA1channel. A decline in respiration was seen in wild-type butnot TRPA1 knockout mice upon exposure to α-terpineol,benzaldehyde, and toluene. Furthermore, we observedeither attenuated or absent avoidance behaviors in TRPA1knockout mice compared to wild-type mice when exposedto α-terpineol, amyl acetate, and benzaldehyde. Our resultsshow that TRPA1 is a molecular target for four differentodorants—α-terpineol, amyl acetate, benzaldehyde, andtoluene. Additionally, this study suggests that TRPA1 isrequired for detection of α-terpineol, benzaldehyde, andtoluene as trigeminal irritants. However, TRPA1 is likelyonly one of multiple trigeminal receptors detecting amylacetate.
Keywords Behavioral aversion . Intracellular calcium .
Irritant . Respiratory pause . Trigeminal nerve . TRPA1
Introduction
Humans detect chemical stimuli in the environment throughthree distinct sensory systems—olfaction, gustation, andchemethesis. Chemesthesis, the sense of irritation due tochemical stimuli, is mediated by a subset of pain,mechanosensory and temperature-sensitive nerve fibers(Bryant and Silver 2000; Bhattacharya et al. 2008). Inmammals, the trigeminal nerve—which innervates themouth, nose, and eyes—is a major anatomical site ofchemesthesis. The small-diameter afferent fibers of thetrigeminal nerve, both myelinated (Aδ) and unmyelinated(c), detect noxious chemical stimuli (Julius and Basbaum2001). Activating trigeminal nerve fibers triggers a varietyof protective reflexes such as sneezing, bronchoconstric-tion, increased secretions (e.g., nasal mucus and saliva), anddecreased respiration rate (Alarie 1973).
Many odorants stimulate trigeminal receptors in the nasalcavity (Tucker 1971), yet olfactory studies do not alwaysconsider possible trigeminal contributions to odor perception.Trigeminal thresholds are generally higher than olfactorythresholds (Cometto-Muñiz and Cain 1990), so it is oftenassumed that testing odorants at low concentrations willeliminate possible trigeminal input. However, the trigeminaland olfactory systems are heavily interrelated (Brand 2006).For example, previous trigeminal stimulation can decreaseolfactory thresholds (Jacquot et al. 2004a) and loss ofolfactory function reduces trigeminal sensitivity (Hummelet al. 1996; Frasnelli and Hummel 2007). These interactionsjustify efforts to understand the mechanisms by whicholfactory stimuli are involved in trigeminal nerve stimulationin an effort to strengthen future studies of olfactory
P. M. Richards (*) : E. C. Johnson :W. L. SilverDepartment of Biology, Wake Forest University,Winston-Salem, NC 27109, USAe-mail: [email protected]
Chem. Percept. (2010) 3:190–199DOI 10.1007/s12078-010-9081-1

perception. The purpose of this study was to examine themechanism by which various olfactory stimuli activate thetrigeminal nerve. Understanding how these odorants stimu-late the trigeminal nerve may aid in determining possibletrigeminal contributions to the perception of these odors.
It is largely unknown which trigeminal receptors areactivated by specific olfactory stimuli. Noxious chemicalstimuli can activate the trigeminal nerve through a varietyof ligand-gated receptors including acid-sensing ion channels(ASIC; Ichikawa and Sugimoto 2002), P2X (Spehr et al.2004), nACh (Alimohammadi and Silver 2000), two-poreK+ (Bautista et al. 2008), and TRP channels, a family ofnonselective cation channels (Caterina et al. 1997; Jordt et al.2004). Additionally, trigeminal irritants can activate recep-tors on solitary chemoreceptor cells (SCCs) which areinnervated by trigeminal nerve fibers (Finger et al. 2003;Tizzano et al. 2010).
One candidate molecule for chemical trigeminal stimula-tion by odorants is the TRPA1 channel. This protein isexpressed in trigeminal sensory neurons (Story et al. 2003;Jordt et al. 2004) and is the primary mediator of trigeminalactivation for allyl-isothiocyanate (AITC; Jordt et al. 2004)and cinnamaldehyde (Bandell et al. 2004). Other stimuli likeeugenol activate the trigeminal nerve by stimulating TRPA1in addition to other trigeminal receptors (e.g., TRPV1; Yanget al. 2003; Bandell et al. 2004). Chemical stimuli are knownto activate TRPA1 by covalently modifying cytosoliccysteine or lysine residues (Hinman et al. 2006; MacPhersonet al. 2007); however, some stimuli may activate TRPA1 byinteracting with a more traditional binding pocket (Peterlinet al. 2007; Matta et al. 2008; Xiao et al. 2008). A largenumber of chemical stimuli with varying structures andchemical properties act as TRPA1 agonists.
Eight olfactory stimuli (Table 1) were examined for theirability to activate TRPA1. It was previously demonstratedthat all of these stimuli activate the trigeminal nerve (Bryantand Silver 2000; Silver et al. 2006). Overall, we identifiedfour previously unidentified TRPA1 agonists: α-terpineol,amyl acetate, benzaldehyde, and toluene. Application ofthese compounds to HEK cells expressing the TRPA1channel leads to increases in intracellular calcium levels.Additionally, behavioral aversion and/or physiologicalresponses to these compounds are attenuated or absent inmice lacking the TRPA1 gene.
Materials and Methods
Chemicals
AITC (Acros) and eight experimental stimuli [α-terpineol(IGN), acetic acid (Sigma), amyl acetate (Fisher), benzal-dehyde (Aldrich), cyclohexanone (Acros), D-limonene
(Kodak), (-)-nicotine (Alfa-Aesar), and toluene (Aldrich)]were tested in this study.
Animals
Sprague–Dawley rats (250–400 g; male) were used toobtain ethmoid nerve recordings (Harlan, Indianapolis, IN).For the behavioral aversion assay and respiration monitoring,eight C57Bl/6J (wild-type) and four B6;129P-Trpa1tm1Kykw/J(TRPA1 KO) mice were used. All mice were female.CJ57Bl/6 mice were obtained from Charles River Laborato-ries (Raleigh, NC), whereas TRPA1 knockout mice wereobtained from Jackson Laboratories (Bar Harbor, ME).Housing and procedures were approved by the Wake ForestUniversity Animal Care and Use Committee.
Ethmoid Nerve Recording
The procedure for recording from the rat ethmoid nerve hasbeen described previously (Silver et al. 2006). Briefly,animals were anesthetized with urethane (IP, ethyl carbamate:1.0 g/kg), and two tubes were placed into the trachea. Onetube was inserted toward the lungs to allow the rat to breathroom air. A second tube was inserted up through the tracheainto the nasopharynx. Ringer’s solution (5.4 mM KCl, 5 mMHEPES, 135 mMNaCl, 1.8 mMCaCl2, pH 7.2) was pumpedthrough the nasopharyngeal tube and allowed to drip fromthe nose. The rat was secured in a head holder, and theethmoid nerve was exposed in the orbit of the eye and placedon two platinum–iridium wire hook electrodes. The cavitywas filled with halocarbon oil (Sigma) to prevent the nervefrom drying out and to provide electrical insulation.
Multiunit neural activity was amplified (Grass P-511) andmonitored (Grass AM-8 audio monitor). The neural outputfrom the ethmoid nerve bundle was integrated using a short-time averaging circuit with a time constant of 0.5 s. Both theneural output and integrated responses were digitized (MP100,Biopac Systems) and recorded using the AcqKnowledge(v3.7.1) data acquisition program (Biopac Systems).
Cell Culture
We utilized HEK-293 cells, kindly donated by Dr. DavidJulius, which stably expressed the TRPA1 channel underthe control of an inducible tetracycline promoter. NaiveHEK-293 and hTRPA1-HEK 293 cells (McNamara et al.2007) were cultured using Dulbecco’s modified eaglemedium (DMEM; Hyclone) supplemented with 10% fetalbovine serum (FBS; PAA) and 1% antibiotic/antimycoticsolution. Cells were kept in an incubator at 37 °C with 5%CO2 and split 3:1 at ~80% confluency using trypsin(CellGro). Before experiments, cells were seeded in apolylysine coated, black-walled, 96-well plate (Corning).
Chem. Percept. (2010) 3:190–199 191

Table 1 Collected information for eight tested odorants
10 sec
Stimulus Structure
Representative Ethmoid Nerve Recordinga
Human Olfactory Threshold (ppm)b
Threshold for activating Ethmoid Nerve (ppm)c
Trigeminal Receptor Targets
α-Terpineol
Acetic Acid
ASICNot
TRPV1d-e
Amyl Acetate f
Benzaldehyde f
Cyclohexanone f
Limonene f
Nicotine Not
Determined 5 nACHg, TRPA1h
Toluene
0.04 19 N/D
0.14 82
0.03 252 Not TRPV1
0.04 125 Not TRPV1
0.71 112 TRPV1
0.44 497 Not TRPV1
1.55 2404 Not TRPV1f
This chart contains the structure of each stimulus tested and representative ethmoid nerve recordings obtained using the stimulus. Additionally, thehuman olfactory threshold, the activation threshold of the ethmoid branch of the trigeminal nerve in rats, and what is known about the trigeminalreceptor targets of each stimulus is reporteda The current studyb Devos et al. 1990c Bryant and Silver 2000d Symanowicz et al. 2004e Kobayashi et al. 2009f Silver et al. 2006g Alimohammadi and Silver 2000h Talavera et al. 2009
192 Chem. Percept. (2010) 3:190–199

Tetracycline (1 μg/ml; Sigma) was added to wells containinghTRPA1-HEK cells to induce expression of TRPA1.
Intracellular Calcium Levels Determination
Intracellular calcium levels were assessed before and afterstimulus addition like previously described (Johnson et al.2003; Bandell et al. 2004; Birse et al. 2006). Briefly, changesin calcium levels of naive HEK and hTRPA1-HEK cellswere detected in an end-point fashion using the calcium-sensitive dye, FLUO-3AM (Molecular Probes), and a multi-label plate reader. Twenty-four hours after plating, the mediawas replaced with HBSS (Sigma) containing 5 μg/mlFLUO-3AM dissolved in 20% Pluronic F-127 (Sigma).Cells were then incubated for 30 min before replacing mediawith HBSS that did not contain FLUO-3AM. Cells werewashed three times with Tyrode’s solution (Silver et al.2006) before adding Tyrode’s solution plus 0.5% dimethylsulfoxide (DMSO; MP Biologicals). For inhibition experi-ments using HC-030031, a TRPA1 inhibitor (McNamara etal. 2007), the inhibitor was dissolved in DMSO and tested ata final concentration of 100 μM in Tyrode’s solution.
After measuring baseline fluorescence with a VictorWallac III multilabel plate reader (Perkin-Elmer), a stimuluswas added to replicate wells. Fluorescence measurementswere then repeated. Due to the nature of the plate reader used,these measurements were end-point measurements. There-fore, kinetic data was not gathered. All stimuli, except D-limonene, were prepared in Tyrode’s solution. D-limonenewas prepared as a 120 mM stock solution in Tyrode’splus 0.466% DMSO and then diluted to the appropriateconcentration for testing. The concentration ranges testedwere determined based on previous physiological testingand an examination of the literature. AITC was tested at afinal stimulus concentration of 0.001–1 mM. Acetic acid,amyl acetate benzaldehyde, cyclohexanone, D-limonene,and nicotine were tested at a final concentration of 0.001–40 mM. Two of the stimuli, α-terpineol and toluene weretested as suspensions in Tyrode’s solution (ranges of 0–6.17and 0–3.69 g/l, respectively). All solutions were pH 7–7.5.
AITC and a calcium ionophore (A23187, Fisher) weretested on each plate to ensure proper dye incorporation andhTRPA1 expression. Each stimulus was tested on threeindependent replicates of three wells (n=9) for each of thefour experimental situations (naive HEK, hTRPA1-HEK,naive HEK plus inhibitor, and hTRPA1-HEK plus inhibitor).
Dose–response curves were developed for tested stimulifrom wells not receiving the TRPA1 inhibitor (for allcompounds except α-terpineol and toluene). These data arereported as a percentage of the maximum responseobtained. EC50 values were calculated from concentrationresponse curves using computerized nonlinear curve fitting(PRISM 3.0; GraphPad, San Diego, CA, USA). Change in
fluorescence of each well was calculated by dividing thefinal fluorescence level (after stimulus addition) by theinitial fluorescence level (prior to chemical addition).Changes in fluorescence upon stimulus addition were thennormalized to vehicle addition. A one-tailed independent ttest was conducted between fluorescence ratios of naiveHEK 293 cells and hTRPA1-HEK 293 cells exposed to thesame irritant concentration to determine statistical signifi-cance (n=9; p<0.05).
Inhibition was examined by normalizing changes influorescence first to vehicle addition and then to fluorescentchanges in naive HEK-293 cells. A one-tailed independent ttest was conducted between fluorescence ratios of hTRPA1-HEK 293 cells with and without addition of HC-030031 ateach concentration tested to determine statistical signifi-cance (n=9; p<0.05).
Respiration Monitoring
Respiration was monitored in wild-type and TRPA1 KOmice similar to previously described (Tizzano et al. 2010).This method requires anesthetization of the animals, but itallows testing of both volatile and nonvolatile compoundsas well as easy determination of the irritant concentrationreaching the nasal cavity. Six stimuli (AITC, α-terpineol,amyl acetate, benzaldehyde, cyclohexanone, and toluene)were tested to determine their effect on trigeminal respira-tory responses in these animals. Stimuli were prepared asfollows: AITC (0.99 g/l); α-terpineol (1.54 g/l), amylacetate (1.30 g/l), benzaldehyde (1.06 g/l), cyclohexanone(9.8 g/l), and toluene (9.2 g/l) in Ringer’s solution. Theselevels were chosen as to be above reported RD50 values(Schaper 1993). A tracheotomy was performed afteranesthesizing mice with urethane (1.0 g/kg) and polyethylenecannulas were placed into both ends of the trachea—oneallowing the mouse to breathe room air and the other leadingto the nasopharynx. A thermistor wire connected to adifferential amplifier (DAM 50, WPI) was placed in thebreathing cannula. Ringer’s solution was perfused continuallythrough the nasopharyngeal cannula (flow rate=10 ml/min).Stimuli (~1 cc) were injected into the cannula at 3–5 minintervals. Acknowledge 3.7.3 (Biopac Systems) was used toacquire and analyze recorded data. The duration of a breathwas divided by the mean duration of breath events (inhalationfollowed by an exhalation) before stimulation (n>10).Relative breath duration of wild-type and TRPA1-KO micewas compared using a two-tailed, independent t test (p<0.05).
Behavioral Aversion Assay
We evaluated behavioral responses of wild-type andTRPA1 KO mice to eight experimental stimuli (Alimo-hammadi 2004). Mice were placed individually into opaque
Chem. Percept. (2010) 3:190–199 193

cages with a 1 cm2 side opening and given 30 min toacclimate before experimentation. Stimuli were presentedby inserting a stimulus soaked cotton-tipped applicator(Puritan) through the cage opening. The behavioral re-sponse of the mouse was then evaluated (Table 2) on adiscrete scale based on distinctive behavioral actions.Stimulus presentation was alternated between trigeminalstimuli and neutral (H2O) or appetitive stimuli. Theobserver was blind to the particular stimulus tested. Micewere rested at least 7 min between subsequent stimuluspresentations.
A behavioral aversion score was determined for eachmouse from the average score of three stimulus pre-sentations. Behavioral aversion scores were comparedbetween wild-type and TRPA1 KO mice using a two-wayanalysis of variance (ANOVA; p<0.05; GraphPad).Additionally, a correlation analysis was completed tomeasure test–retest reliability. AITC (Acros) was used asa positive control as it is known to stimulate TRPA1 (Jordtet al. 2004). All stimulus mixtures were prepared indistilled water to prevent vehicle interference and mixedwell before each presentation. Stimuli were prepared asfollows: α-terpineol (15.4 g/l), acetic acid (60.1 g/l), AITC(9.9 g/l), amyl acetate (13.0 g/l), benzaldehyde (10.6 g/l),cyclohexanone (9.8 g/l), D-limonene (13.6 g/l), nicotine(16.2 g/l), and toluene (9.2 g/l). These irritant levels werechosen as they elicited behavioral responses from wild-type mice but did not cause learned aversion to stimuluspresentation. Appetitive stimulus solutions were preparedby mixing whole wheat powder (King Arthur) or choco-late powder (Hershey’s, Nestle’s, Nesquik) in deionizedwater.
Results
Stimulation of Trigeminal Nerve by Tested Odorants
All of the tested odorants activated the trigeminal nerve asdemonstrated by ethmoid nerve recordings (see Table 1). Therepresentative recordings shown were obtained using thefollowing stimulus preparations in Ringer’s solution:α-terpineol (15.4 g/l), acetic acid (6.0 g/l), amyl acetate(13.0 g/l), benzaldehyde (10.6 g/l), cyclohexanone (9.8 g/l),D-limonene (13.6 g/l), nicotine (1.6 g/l), and toluene (9.2 g/l).
Activation of TRPA1 by Four Odorants
To determine whether TRPA1 is activated by the testedodorants, we monitored changes in calcium levels ofHEK cells upon presentation of these chemicals in anend-point fashion. Following induction of hTRPA1-HEKcells with tetracycline, we observed dramatic increases inintracellular calcium with the presentation of AITC, aknown TRPA1 agonist (Jordt et al. 2004). Notably,neither naive HEK cells nor cells lacking tetracyclineinduction (data not shown) showed this level of activation.Four of the experimental stimuli (α-terpineol, amylacetate, benzaldehyde, and toluene) elicited significantcalcium influxes in hTRPA1-HEK cells but not naive HEKcells (p<0.05, Fig. 1). In contrast, acetic acid, cyclohex-anone, D-limonene, and (-)-nicotine, did not cause highercalcium influxes in hTRPA1-HEK than naive HEK cells.Since cyclohexanone and (-)-nicotine elicited calciuminfluxes in naive HEK cells at high concentrations (∂≥40and ≥10 mM, respectively), the fluorescence ratio belowthose levels are reported.
We then evaluated the sensitivities of the responseselicited by the agonistic stimuli by assessing TRPA1activation under a range of concentrations. Specifically,we found the order of sensitivity from highest to lowest tobe: AITC (EC50=5.8 μM), benzaldehyde (EC50=53 μM,amyl acetate (EC50=1,900 μM) (Fig. 2). Increased intra-cellular calcium levels were seen only when α-terpineol andtoluene were added as a suspension (≥6.17 and ≥0.92 g/l,respectively. Therefore, no dose–response curve was con-structed for α-terpineol and toluene.
To ensure elicited responses to these odorants weredue to selective TRPA1 activation and not unspecificcellular responses, we employed a TRPA1 inhibitor HC-030031 (McNamara et al. 2007). Co-application of HC-030031 with agents that elicited responses from hTRPA1-HEK cells caused a significant reduction in calciuminfluxes (Fig. 3). Specifically, HC-030031 eliminated theresponses to amyl acetate, α-terpineol, and toluene andsignificantly reduced the amplitude of the responses toAITC and benzaldehyde. It should be noted that one study
Table 2 Scoring system for the behavioral aversion assay
Behavioralscore
Category Behavioral description
–2 Trigeminalreflex
Rapid, reflexive withdrawalmovement away from stimulus source
−1 Aversiveresponse
Rejection of stimulus markedby a head turn movement
0 Neutralresponse
No response or lack of interest instimulus presentation
1 Positiveresponse
Investigation of stimulus source,marked by an increase in inspiratorysniffing
2 Feedingattempt
Attempted manipulation of stimulussource or attempted feeding behavior
Mice were scored on a discrete scale from −2 to +2 based on thedistinctive behaviors described when presented with a stimulus-saturated cotton swab. The overall behavioral aversion score wasdetermined for each mouse from the average score of three stimuluspresentations
194 Chem. Percept. (2010) 3:190–199

does indicate that HC-030031 inhibits TRPV1 in additionto TRPA1 (Iwasaki et al. 2009); however, this is notrelevant in the current study as TRPV1 was not present inthe HEK 293 cells tested.
Altered Behavioral and Physiological Responses to TRPA1Stimuli
Since these results demonstrate that TRPA1 is a moleculartarget of α-terpineol, amyl acetate, benzaldehyde, and toluene,we monitored respiration in wild-type and TRPA1 -/- mice toassess whether TRPA1 was required for trigeminal activationby these odorants (Fig. 4). Activation of the trigeminal nervecauses decreased respiratory frequency (Vijayaraghavan1993). Wild-type mice exhibited a short-term decrease inrespiration (measured as relative breath duration) whenexposed to all stimuli tested except the vehicle (Ringers).TRPA1 KO mice showed a similar decrease in respiration(p>0.05) when exposed to amyl acetate or cyclohexanone(data not shown); however, this response was absent inanimals exposed to AITC, α-terpineol, benzaldehyde, ortoluene.
We then used a behavioral aversion assay to furtherassess whether TRPA1 was necessary for trigeminallyinduced aversive behavioral responses. To evaluatewhether TRPA1 is necessary for aversive behavior, wetested mice lacking the TRPA1 gene (TRPA1 KO) withD-limonene, which was shown not to activate TRPA1. Wefound that the TRPA1 KO mice were indistinguishablefrom wild-type mice in the level that they avoidedD-limonene (p>0.05). Additionally, the two mouse strainswere indistinguishable (p>0.05) in their response to boththe vehicle (H2O) and toluene (Fig. 5). In contrast, TRPA1KOs displayed significantly lower aversion to AITC,α-terpineol, benzaldehyde, and amyl acetate (p<0.001)as compared to wild-type mice. When evaluated using atwo-way ANOVA, all three components (genotype, chem-ical, and interaction) were significant. Wild-type mice
Fig. 1 Four of the tested odorants activate TRPA1 in a heterologousexpression system. Relative fluorescence ratios (mean ± SE) aredisplayed for HEK 293 and hTRPA1-HEK 293 cells for the positivecontrol (AITC) and the eight experimental stimuli. Fluorescence wasmeasured on a multilabel plate reader, so only end-point data areavailable. Stimulus preparations represented are as follows: α-terpineol (6.17 g/l), acetic acid (2.4 g/l), AITC (0.1 g/l), amyl acetate(5.2 g/l), benzaldehyde (4.2 g/l), cyclohexanone (1.96 g/l), D-limonene(5.4 g/l), nicotine (0.16 g/l), and toluene (3.69 g/l). Exposure to thepositive control (AITC), α-terpineol, amyl acetate, benzaldehyde, andtoluene elicits larger intracellular calcium increases (mean ± SE) inhTRPA1-HEK 293 cells than in naive HEK 293 cells (one-tailed,Student’s t test: p<0.05). n=9
Fig. 2 Dose–response curves of hTRPA1-HEK 293 cells to stimulithat elicited intracellular calcium increases (mean ± SE) compared tonaive HEK cells. Curves are displayed as a percentage of themaximum response. Vehicle addition was 83.6%–88.8% of themaximum response (i.e., largest increase in intracellular calcium with
addition of stimulus) for each stimulus tested. The calculated EC50
values were as follows: AITC (5.8 μM), amyl acetate (1,900 μM), andbenzaldehyde (53 μM). A dose–response curve is not included for α-terpineol or toluene as these stimuli only elicited intracellular calciumincreases when tested as suspensions. n=9
Chem. Percept. (2010) 3:190–199 195
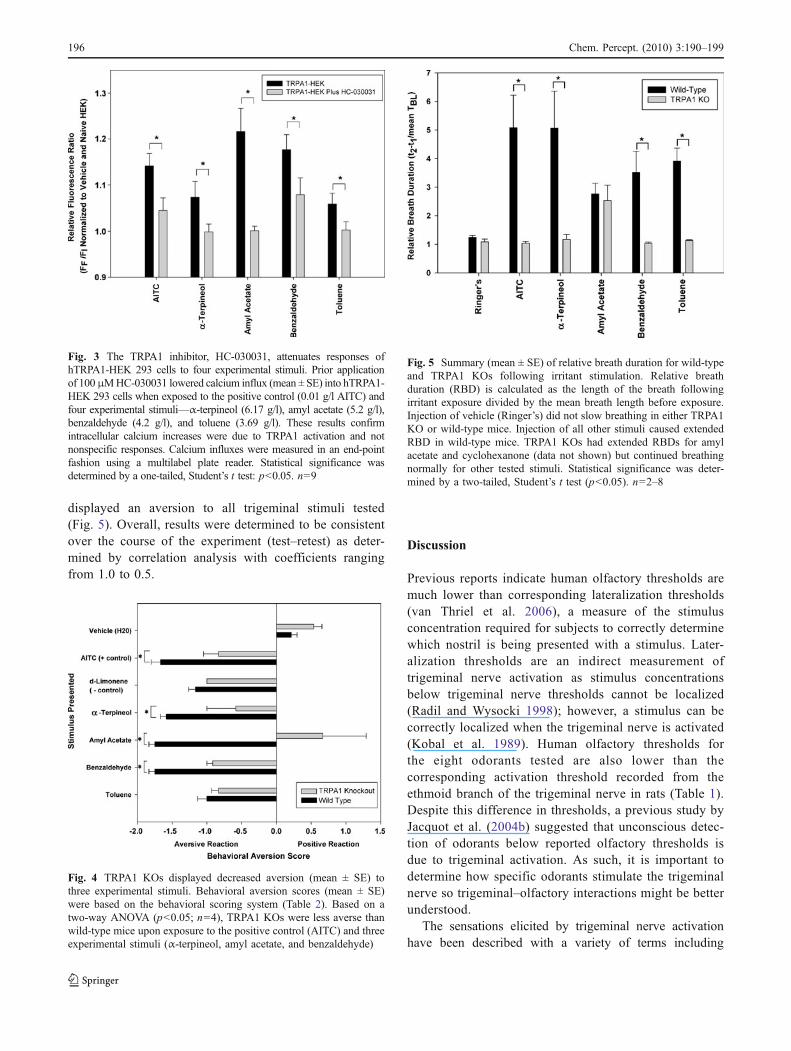
displayed an aversion to all trigeminal stimuli tested(Fig. 5). Overall, results were determined to be consistentover the course of the experiment (test–retest) as deter-mined by correlation analysis with coefficients rangingfrom 1.0 to 0.5.
Discussion
Previous reports indicate human olfactory thresholds aremuch lower than corresponding lateralization thresholds(van Thriel et al. 2006), a measure of the stimulusconcentration required for subjects to correctly determinewhich nostril is being presented with a stimulus. Later-alization thresholds are an indirect measurement oftrigeminal nerve activation as stimulus concentrationsbelow trigeminal nerve thresholds cannot be localized(Radil and Wysocki 1998); however, a stimulus can becorrectly localized when the trigeminal nerve is activated(Kobal et al. 1989). Human olfactory thresholds forthe eight odorants tested are also lower than thecorresponding activation threshold recorded from theethmoid branch of the trigeminal nerve in rats (Table 1).Despite this difference in thresholds, a previous study byJacquot et al. (2004b) suggested that unconscious detec-tion of odorants below reported olfactory thresholds isdue to trigeminal activation. As such, it is important todetermine how specific odorants stimulate the trigeminalnerve so trigeminal–olfactory interactions might be betterunderstood.
The sensations elicited by trigeminal nerve activationhave been described with a variety of terms including
Fig. 5 Summary (mean ± SE) of relative breath duration for wild-typeand TRPA1 KOs following irritant stimulation. Relative breathduration (RBD) is calculated as the length of the breath followingirritant exposure divided by the mean breath length before exposure.Injection of vehicle (Ringer’s) did not slow breathing in either TRPA1KO or wild-type mice. Injection of all other stimuli caused extendedRBD in wild-type mice. TRPA1 KOs had extended RBDs for amylacetate and cyclohexanone (data not shown) but continued breathingnormally for other tested stimuli. Statistical significance was deter-mined by a two-tailed, Student’s t test (p<0.05). n=2–8
Fig. 3 The TRPA1 inhibitor, HC-030031, attenuates responses ofhTRPA1-HEK 293 cells to four experimental stimuli. Prior applicationof 100μMHC-030031 lowered calcium influx (mean ± SE) into hTRPA1-HEK 293 cells when exposed to the positive control (0.01 g/l AITC) andfour experimental stimuli—α-terpineol (6.17 g/l), amyl acetate (5.2 g/l),benzaldehyde (4.2 g/l), and toluene (3.69 g/l). These results confirmintracellular calcium increases were due to TRPA1 activation and notnonspecific responses. Calcium influxes were measured in an end-pointfashion using a multilabel plate reader. Statistical significance wasdetermined by a one-tailed, Student’s t test: p<0.05. n=9
Fig. 4 TRPA1 KOs displayed decreased aversion (mean ± SE) tothree experimental stimuli. Behavioral aversion scores (mean ± SE)were based on the behavioral scoring system (Table 2). Based on atwo-way ANOVA (p<0.05; n=4), TRPA1 KOs were less averse thanwild-type mice upon exposure to the positive control (AITC) and threeexperimental stimuli (α-terpineol, amyl acetate, and benzaldehyde)
196 Chem. Percept. (2010) 3:190–199

“tingling,” “pungent,” “burning,” or “cooling”. The specificsensations experienced often depend on the type ofchemoreceptors activated by the stimulus. For example,capsaicin, the “hot” component of chili peppers, activatesTRPV1 (Caterina et al. 1997), whereas menthol, the “cool”component in many gums or mouthwashes, activatesTRPM8 (McKemy et al. 2002; Peier et al. 2002). Manyodorants have been identified as trigeminal stimuli byrecording from the ethmoid branch of the trigeminal nerveor monitoring intracellular calcium increases in trigeminalneurons. It is not known which trigeminal chemoreceptorsmany specific olfactory stimuli target. In the current study,eight odorants, which act as trigeminal agonists, were testedto determine if they activate the trigeminal chemoreceptorTRPA1.
We provide multiple lines of evidence that activation ofthe trigeminal system by α-terpineol, amyl acetate, benzal-dehyde, and toluene is mediated via the TRPA1 channel.First, the introduction of the human TRPA1 channel toHEK-293 cells conferred sensitivity to these compounds asassessed by calcium levels. Notably, these calcium changeswere attenuated when a known TRPA1 inhibitor was co-applied. Furthermore, both the behavioral and physiologicalresponses elicited by trigeminal nerve activation arediminished in animals lacking the TRPA1 channel. Ingeneral, the sensitivity of TRPA1 to these chemicals (andAITC) correlates with previous estimates of trigeminalsensitivity. One notable difference in reported thresholdswas the calculated threshold for α-terpineol. In theintracellular calcium assay, α-terpineol only activated thehTRPA1-HEK cells when applied as a suspension (6.17 g/l),whereas reported nerve thresholds (Bryant and Silver 2000)and concentrations eliciting a respiratory pause in the wild-type mice (1.54 g/l) are much lower. This discrepancy inthreshold is likely due to inherent differences in the assaysthemselves or to species-specific activation patterns asspecies differences in TRPA1 have been demonstrated (Chenand Kym 2009).
Although the current study did not investigate howthese agonists stimulate TRPA1, the chemical reactivity ofthe compound likely determines the mechanism by whichthe stimulus activates TRPA1 (Hinman et al. 2006;MacPherson et al. 2007; Peterlin et al. 2007) if it bindsdirectly. These studies suggest that electrophilic compoundsmodify cysteine or lysine residues covalently to activate theTRPA1 channel. Although amyl acetate and benzaldehydeare not as strongly electrophilic as the compoundsreferenced in these studies, it is possible these mildelectrophiles activate the channel in a similar manner.Nonelectrophilic agonists like α-terpineol and toluenemight activate TRPA1 through a more traditional bindingpocket (Peterlin et al. 2007). However, it should be notedthat TRPA1 can be sensitized or activated by intracellular
messengers like calcium (Doerner et al. 2007) or byintracellular acidification (Wang et al. 2010). If this werethe case in the present study, intracellular calcium increaseswould have been noted in the naive HEK cells; however,downstream activation of TRPA1 due to a differentintracellular messenger or by intracellular acidificationcannot be eliminated.
TRPA1 KO mice do not slow respiration when exposedto α-terpineol, benzaldehyde, and toluene, indicating thatTRPA1 is necessary for trigeminal nerve activation by theseodorants. Additionally, TRPA1 KO mice display decreasedbehavioral aversion to α-terpineol and benzaldehydefurther supporting this conclusion. Compared to wild-typemice, mice lacking the TRPA1 gene display equivalentbehavioral aversion to toluene. There are at least twopossible explanations for this discrepancy. It is possible thatthe behavioral aversion of TRPA1 KO mice to toluene maybe due to aversive olfactory responses as all mice had intactolfactory systems. One interpretation would be that aver-sive olfactory input overrode any differences in trigeminalsensitivity between the two genotypes. However, it ispossible that the final concentration of toluene reachingthe free nerve endings in the mouse was not high enough inthis assay to elicit behavioral reflexes due to trigeminalirritation in either strain. Based on this interpretation, theaversion seen in both strains is due solely to olfaction, andthe behavioral similarity of the two strains is due to the lackof trigeminal input. The threshold for detection of tolueneas an irritant has been reported to be much higher than theolfactory threshold (Cometto-Muñiz and Cain 1995;Cometto-Muñiz et al. 2002), suggesting that aversivebehavior due to olfaction could be seen at much lowerconcentrations than required for trigeminal mediatedresponses. As suspensions up to 9.2 g/l did not causedecreased respiration in TRPA1 KOs, we suspect thatTRPA1 is the primary trigeminal target for toluene.
In contrast, our results suggest that amyl acetate mayactivate other receptors in addition to TRPA1 at highconcentrations. TRPA1 KO and wild-type mice displayedthe same respiratory slowing upon exposure to amylacetate, suggesting that TRPA1 is not required for amylacetate detection. However, during the behavioral aversionassay, stark differences were observed between the behaviorof TRPA1 KOs and wild-type mice exposed to amylacetate. The discrepancy between the results of these twoassays may be a matter of concentration. We suspect thatthe concentration of amyl acetate actually detected by themice was lower in the behavioral aversion assay (due tovolatility and dilution by air) than the concentrationsdirectly applied when measuring respiratory responses.This study demonstrates that TRPA1 is a molecular targetof amyl acetate; however, multiple trigeminal receptors arelikely activated by this odorant. Future studies should
Chem. Percept. (2010) 3:190–199 197

consider other trigeminal nerve or SCC receptors, with theexception of TRPV1 (Silver et al. 2006), as potentialtrigeminal targets of this odorant.
Four of the eight trigeminal irritants tested did notactivate TRPA1. Talavera et al. (2009) reported thatnicotine activates TRPA1 within a limited concentrationrange, whereas our results did not demonstrate activation ofTRPA1 by nicotine application. However, the concentra-tions that we tested were not within the concentration rangeused by Talavera et al. (2009), so it is not surprising that wefound nicotine did not activate TRPA1 at those concen-trations. All of the tested odorants that did not elicitresponses from TRPA1, except D-limonene, have identifiedtrigeminal receptor targets (Table 1). As TRPV1 has alreadybeen eliminated as a target receptor for D-limonene (Silveret al. 2006) and the current study eliminates TRPA1, eitherdifferent TRP channels (e.g., TRPM8) or alternativereceptors could be the basis for trigeminal stimulation bythis odorant.
Conclusions
Overall, these experiments identified four new TRPA1agonists: α-terpineol, amyl acetate, benzaldehyde, andtoluene. Each of these odorants elicited intracellularcalcium increases in HEK-293 cells expressing hTRPA1,demonstrating that TRPA1 is a molecular target. Further-more, TRPA1 is required for detection of α-terpineol,benzaldehyde, and toluene, as TRPA1 knockout micelacked respiratory responses to these compounds. Behav-ioral aversion supports the hypothesis that TRPA1 mediatesthe trigeminal responses to these odorants. In contrast, amylacetate is likely to target multiple receptors, in addition toTRPA1. Future studies using these odorants should takeinto account not only that these stimuli activate trigeminalnerve receptors but also that the mechanism by whichthey activate the nerve as this understanding may clarifythe role of the trigeminal system in the perception of theseodorants.
Acknowledgments This work was supported by the Wake ForestUniversity Science Research Fund. We thank Dr. David Julius (UCSF)for providing the hTRPA1-HEK 293 stable line and Dr. T. MichaelAnderson (WFU) for his aid in statistical analysis.
References
Alarie YY (1973) Sensory irritation by airborne chemicals. CRC CritRev Toxicol 2:299–363
Alimohammadi H (2004) Trigeminal nerve-mediated nasal chemes-thesis: chemosensory mechanisms of the Vth cranial nerve.Dissertation, Wake Forest University
Alimohammadi H, Silver WL (2000) Evidence for nicotinic acetyl-choline receptors on nasal trigeminal nerve endings of the rat.Chem Senses 25:61–66
Bandell M, Story GM, Hwang SW et al (2004) Noxious cold ionchannel TRPA1 is activated by pungent compounds andbradykinin. Neuron 41:849–857
Bautista DM, Sigal YM, Milstein AD et al (2008) Pungent agentsfrom Szechuan peppers excite sensory neurons by inhibiting two-pore potassium channels. Nat Neurosci 7:772–779
Bhattacharya MR, Bautista DM, Wu K et al (2008) Radial stretchreveals distinct populations of mechanosensitive mammaliansomatosensory neurons. Proc Natl Acad Sci USA 105:20015–20020
Birse RT, Johnson EC, Taghert PH et al (2006) Widely distributedDrosophila G-protein coupled receptor (CG7887) is activated byendogenous tachykinin-related peptides. J Neurobiol 66:33–46
Brand G (2006) Olfactory/trigeminal interactions in nasal chemore-ception. Neurosci Biobehav Rev 30:908–917
Bryant B, Silver WL (2000) Chemesthesis: the common chemicalsense. In: Finger TE, Silver WL, Restrepo D (eds) Theneurobiology of taste and smell. Wiley-Liss, New York
Caterina MJ, Schumacher MA, Tominaga M et al (1997) Thecapsaicin receptor: a heat-activated channel in the pain pathway.Nature 389:816–824
Chen J, Kym PR (2009) TRPA1: the species difference. J Gen Physiol133:623–625
Cometto-Muñiz JE, Cain WS (1990) Thresholds for odor and nasalpungency. Physiol Behav 48:719–725
Cometto-Muñiz JE, Cain WS (1995) Relative sensitivity of the oculartrigeminal, nasal trigeminal and olfactory systems to airbornechemicals. Chem Senses 20:191–198
Cometto-Muñiz JE, Cain WS, Abraham MH et al (2002) Psychomet-ric functions for the olfactory and trigeminal detectability ofbutyl acetate and toluene. J Appl Toxicol 22:25–30
Devos M, Patte F, Rouault J et al (1990) Standardized humanolfactory thresholds. Oxford University Press, New York
Doerner JF, Gisselmann G, Hatt H et al (2007) Transient receptorpotential channel A1 is directly gated by calcium ions. J BiolChem 282:13180–13189
Finger TE, Bottger B, Hansen A et al (2003) Solitary chemoreceptorcells in the nasal cavity serve as sentinels of respiration. ProcNatl Acad Sci USA 100:8981–8986
Frasnelli J, Hummel T (2007) Interactions between the chemicalsenses: trigeminal function in patients with olfactory loss. Int JPsychophysiol 65:177–181
Hinman A, Chuang H, Bautista DM et al (2006) TRP channelactivation by reversible covalent modification. Proc Natl AcadSci USA 103:19564–19568
Hummel T, Barz S, Lotsch J et al (1996) Loss of olfactory function leadsto a decrease of trigeminal sensitivity. Chem Senses 21:75–79
Ichikawa H, Sugimoto T (2002) The co-expression of ASIC3 withcalcitonin gene-related peptide and parvalbumin in the rattrigeminal ganglion. Brain Res 943:287–291
Iwasaki Y, Tanabe M, Kayama Y et al (2009) Miogadial andmiogatrial with alpha, beta-unsaturated 1, 4-dialdehyde moieties—novel and potent TRPA1 agonists. Life Sci 85:60–69
Jacquot L, Monnin J, Brand G (2004a) Influence of trigeminal stimulion olfactory sensitivity. C R Biol 327:305–311
Jacquot L, Monnin J, Brand G (2004b) Unconcious odor detectioncould not be due to odor itself. Brain Res 1002:51–54
Johnson EC, Garczynski SF, Park D et al (2003) Identification andcharacterization of a G protein-coupled receptor for the neuro-peptide proctolin in Drosophila melanogaster. Proc Natl AcadSci USA 100:6198–6203
Jordt SE, Bautista DM, Chuang HH et al (2004) Mustard oils andcannabinoids excite sensory nerve fibres through the TRPchannel ANKTM1. Nature 427:260–265
198 Chem. Percept. (2010) 3:190–199

Julius D, Basbaum AI (2001) Molecular mechanisms of nociception.Nature 413:203–210
Kobal G, van Toller S, Hummel T (1989) Is there directionalsmelling? Experientia 45:130–132
Kobayashi H, Yoshiyama M, Zakoji H et al (2009) Sex differences inthe expression profile of acid-sensing ion channels in the mouseurinary bladder: a possible involvement of irritative bladdersymptoms. BJU Int 104:1746–1751
MacPherson LJ, Dubin AE, Evans MJ et al (2007) Noxiouscompounds activate TRPA1 ion channels through covalentmodification of cysteines. Nature 445:541–545
Matta JA, Cornett PM, Miyares RL et al (2008) General anestheticsactivate a nociceptive ion channel to enhance pain andinflammation. Proc Natl Acad Sci USA 105:8784–8789
McKemy DD, Neuhausser WM, Julius D (2002) Identification of acold receptor reveals a general role for TRP channels inthermosensation. Nature 416:52–58
McNamara CR, Mandel-Brehm J, Bautista DM et al (2007) TRPA1mediates formalin-induced pain. Proc Natl Acad Sci USA104:13525–13530
Peier AM, Moqrich A, Hergarden AC et al (2002) ATRP channel thatsenses cold stimuli and menthol. Cell 108:705–715
Peterlin Z, Chesler A, Firestein S (2007) A painful TRP can be abonding experience. Neuron 53:635–638
Radil T, Wysocki CJ (1998) Spatiotemporal masking in pure olfaction.Ann NY Acad Sci 855:641–644
Schaper M (1993) Development of a database for sensory irritants andits use in establishing occupational exposure limits. Am Ind HygAssoc J 54:488–544
Silver WL, Clapp TR, Stone LM et al (2006) TRPV1 receptors andnasal trigeminal chemesthesis. Chem Senses 31:807–812
Spehr J, Spehr M, Hatt H et al (2004) Subunit-specific P2X-receptorexpression defines chemosensory properties of trigeminal neurons.Eur J Neurosci 19:2497–2510
Story GM, Peier AM, Reeve AJ et al (2003) ANKTM1, a TRP-likechannel expressed in nociceptive neurons, is activated by coldtemperatures. Cell 112:819–829
Symanowicz PT, Gianutsos G, Morris JB (2004) Lack of role forthe vanilloid receptor in response to several inspired irritantair pollutants in the C57Bl/6J mouse. Neurosci Lett 362:150–153
Talavera K, Maarten G, Karashima Y et al (2009) Nicotine activatesthe chemosensory cation channel TRPA1. Nat Neurosci12:1293–1299
Tizzano M, Gulbransen BD, Vandenbeuch AV et al (2010) Nasalchemosensory cells use bitter taste signaling to detect irritantsand bacterial signals. Proc Natl Acad Sci USA 107:3210–3215
Tucker D (1971) Nonolfactory responses from the nasal cavity:Jacobson’s organ and the trigeminal system. In: Beidler LM (ed)Handbook of sensory physiology, vol IV. Chemical senses.Springer-Verlag, Berlin
van Thriel C, Schaper M, Kiesswetter E et al (2006) From chemo-sensory thresholds to whole body exposures—experimentalapproaches evaluating chemosensory effects of chemicals. IntArch Occup Environ Health 79:308–321
Vijayaraghavan R (1993) Characteristic modifications of the breathingpattern of mice to evaluate the effects of airborne chemicals onthe respiratory tract. Arch Toxicol 67:478–490
Wang YY, Chang RB, Liman ER (2010) TRPA1 dependent detectionof CO2 by nociceptors: CO2 sensing by TRPA1. J Neurosci (inpress)
Xiao B, Dubin AE, Bursulaya B et al (2008) Identification oftransmembrane domain 5 as a critical molecular determinant ofmenthol sensitivity in mammalian TRPA1 channels. J Neurosci28:9640–9651
Yang BH, Piao ZG, Kim YB et al (2003) Activation of vanilloidreceptor 1 (VR1) by eugenol. J Dent Res 82:781–785
Chem. Percept. (2010) 3:190–199 199














