
Eyeblink conditioning anomalies in bipolar disorder suggest cerebellar dysfunction
14
Original Article Eyeblink conditioning anomalies in bipolar disorder suggest cerebellar dysfunction Despite the extraordinary personal and societal burden of bipolar disorder (BD), relatively little is known about neural mechanisms associated with the unusual swings and shifts in affective and cognitive function that characterize the disorder. Accumulating evidence suggests that structures (e.g., the cerebellum and basal ganglia) and neu- rotransmitter systems (e.g., dopamine and gluta- mate) associated with neural timing processes are abnormal in mood disorders (1–5). The overarch- ing theoretical model under investigation in the present study posits that classic symptoms of BD may arise from poor cerebellar-mediated temporal coordination of networks distributing and integrating cognitive and affective information. Although conventional conceptualizations of the cerebellum have been confined to motor coordina- tion, a growing body of empirical and theoretical evidence indicates that the cerebellum plays a significant role in psychological functions as well, including modulation of perceptual, cognitive, and affective functions (6–12). This evidence is broad- ening the traditional notion of the cerebellum and illuminating new avenues of research into the mechanisms of psychiatric illness (13, 14). Bolbecker AR, Mehta C, Johannesen JK, Edwards CR, OÕDonnell BF, Shekhar A, Nurnberger JI, Steinmetz JE, Hetrick WP. Eyeblink conditioning anomalies in bipolar disorder suggest cerebellar dysfunction. Bipolar Disord 2009: 11: 19–32. ª 2009 The Authors Journal compilation ª 2009 Blackwell Munksgaard Objectives: Accumulating research implicates the cerebellum in non- motor psychological processes and psychiatric diseases, including bipolar disorder (BD). Despite recent evidence that cerebellar lesions have been documented to trigger bipolar-like symptoms, few studies have directly examined the functional integrity of the cerebellum in those afflicted with BD. Methods: Using a single-cue delay eyeblink conditioning procedure, the functional integrity of the cerebellum was examined in 28 individuals with BD (9 manic, 8 mixed, and 11 euthymic) and 28 age-matched healthy controls. Results: Analysis of the bipolar group as a whole indicated a conditioned response acquisition and timing deficit compared to controls. However, when the bipolar group was categorized according to mood state (mixed, manic, euthymic), individuals tested during mixed episodes were strikingly impaired, performing significantly worse than all other groups on both the acquisition and timing of conditioned responses. Conclusions: These findings extend prior research implicating cerebellar functional abnormalities in BD and suggest that cerebellar dysfunction may be associated with mood state and course of illness. Amanda R Bolbecker a,b , Crystal Mehta a , Jason K Johannesen c , Chad R Edwards a , Brian F OÕDonnell a,b,d , Anantha Shekhar b,d , John I Nurnberger d , Joseph E Steinmetz e and William P Hetrick a,b,d a Department of Psychological and Brain Sciences, Indiana University, Bloomington, b Larue D. Carter Memorial Hospital, c Department of Psychology, Indiana University-Purdue University, d Department of Psychiatry, Indiana University School of Medicine, Indianapolis, IN, e Departments of Psychology and Molecular Biosciences, University of Kansas, Lawrence, KS, USA Key words: bipolar disorder – cerebellum – depression – eyeblink conditioning – mania – mixed – temporal – timing Received 22 April 2007, revised and accepted for publication 1 February 2008 Corresponding author: William P. Hetrick, PhD, Department of Psychological and Brain Sciences, Indiana University, 1101 E. Tenth Street, Bloomington, IN 47405, USA. Fax: 812 856 4544; e-mail: [email protected] The authors of this paper do not have any commercial associations that might pose a conflict of interest in connection with this manu- script. Bipolar Disorders 2009: 11: 19–32 ª 2009 The Authors Journal compilation ª 2009 Blackwell Munksgaard BIPOLAR DISORDERS 19
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Eyeblink conditioning anomalies in bipolar disorder suggest cerebellar dysfunction

Original Article
Eyeblink conditioning anomalies in bipolardisorder suggest cerebellar dysfunction
Despite the extraordinary personal and societalburden of bipolar disorder (BD), relatively little isknown about neural mechanisms associated withthe unusual swings and shifts in affective andcognitive function that characterize the disorder.Accumulating evidence suggests that structures(e.g., the cerebellum and basal ganglia) and neu-rotransmitter systems (e.g., dopamine and gluta-mate) associated with neural timing processes areabnormal in mood disorders (1–5). The overarch-
ing theoretical model under investigation in thepresent study posits that classic symptoms of BDmay arise from poor cerebellar-mediated temporalcoordination of networks distributing andintegrating cognitive and affective information.Although conventional conceptualizations of thecerebellum have been confined to motor coordina-tion, a growing body of empirical and theoreticalevidence indicates that the cerebellum plays asignificant role in psychological functions as well,including modulation of perceptual, cognitive, andaffective functions (6–12). This evidence is broad-ening the traditional notion of the cerebellum andilluminating new avenues of research into themechanisms of psychiatric illness (13, 14).
Bolbecker AR, Mehta C, Johannesen JK, Edwards CR, O�Donnell BF,Shekhar A, Nurnberger JI, Steinmetz JE, Hetrick WP. Eyeblinkconditioning anomalies in bipolar disorder suggest cerebellardysfunction.Bipolar Disord 2009: 11: 19–32. ª 2009 The Authors Journalcompilation ª 2009 Blackwell Munksgaard
Objectives: Accumulating research implicates the cerebellum in non-motor psychological processes and psychiatric diseases, including bipolardisorder (BD). Despite recent evidence that cerebellar lesions have beendocumented to trigger bipolar-like symptoms, few studies have directlyexamined the functional integrity of the cerebellum in those afflictedwith BD.
Methods: Using a single-cue delay eyeblink conditioning procedure,the functional integrity of the cerebellum was examined in 28 individualswith BD (9 manic, 8 mixed, and 11 euthymic) and 28 age-matchedhealthy controls.
Results: Analysis of the bipolar group as a whole indicated aconditioned response acquisition and timing deficit compared tocontrols. However, when the bipolar group was categorized according tomood state (mixed, manic, euthymic), individuals tested during mixedepisodes were strikingly impaired, performing significantly worse than allother groups on both the acquisition and timing of conditionedresponses.
Conclusions: These findings extend prior research implicatingcerebellar functional abnormalities in BD and suggest that cerebellardysfunction may be associated with mood state and course of illness.
Amanda R Bolbeckera,b, CrystalMehtaa, Jason K Johannesenc,Chad R Edwardsa, Brian FO�Donnella,b,d, Anantha Shekharb,d,John I Nurnbergerd, Joseph ESteinmetze and William PHetricka,b,d
aDepartment of Psychological and Brain Sciences,
Indiana University, Bloomington, bLarue D. Carter
Memorial Hospital, cDepartment of Psychology,
Indiana University-Purdue University, dDepartment
of Psychiatry, Indiana University School of
Medicine, Indianapolis, IN, eDepartments of
Psychology and Molecular Biosciences, University
of Kansas, Lawrence, KS, USA
Key words: bipolar disorder – cerebellum –
depression – eyeblink conditioning – mania –
mixed – temporal – timing
Received 22 April 2007, revised and accepted for
publication 1 February 2008
Corresponding author: William P. Hetrick, PhD,
Department of Psychological and Brain Sciences,
Indiana University, 1101 E. Tenth Street,
Bloomington, IN 47405, USA.
Fax: 812 856 4544;
e-mail: [email protected]
The authors of this paper do not have any commercial associations
that might pose a conflict of interest in connection with this manu-
script.
Bipolar Disorders 2009: 11: 19–32ª 2009 The Authors
Journal compilation ª 2009 Blackwell Munksgaard
BIPOLAR DISORDERS
19

Concordant with this more contemporary con-ceptualization, recent evidence suggests that cere-bellar abnormalities may play a role in mooddisorders. For example, structural imaging studiesindicate cerebellar abnormalities, especially cere-bellar atrophy, in BD (15–22) and unipolar depres-sion (23, 24), as well as neurochemical markersof cerebellar abnormalities in mood-disorderedchildren with a BD parent (25).Findings of cerebellar abnormalities in BD are
especially intriguing given evidence linking thecerebellum to emotional and behavioral regulation.For example, marked changes in affect and behav-ior have been reported following focal lesions tothe cerebellum, including emotional dysregulation,behavioral lability, contextually inappropriatebehavior, mania, depression, and rapid cycling(8–12, 26), all of which bear a striking resemblanceto hallmark symptoms of BD. However, it shouldbe noted that the psychological effects of cerebellarlesions in adults may subside within a year of lesionoccurrence (27).Further evidence for cerebellar influence on
affective states comes from experiments in whichcerebellar stimulation has resulted in modulationof emotion in humans (28, 29). In addition, brainimaging studies have documented increased acti-vation in the cerebellum during the induction ofsadness (30, 31) and happiness (32). Interestingly,in patients with cerebellar lesions, the processingof emotional material appears to be mediatedby different neural circuits compared to a healthycontrol group (33).The anatomical connections of the cerebellum
are also consistent with functional indications of itsrole in emotional modulation. For example, thecerebellum shares projections with cortical andsubcortical areas critical for affect regulation,including feedback loops with the prefrontal cortex(PFC) (34, 35) and limbic system (36–38). Inaddition, the cerebellum projects to brainstemnuclei (39) and cerebellar stimulation has beenshown to influence catecholamine turnover inforebrain areas critical to emotional processes,presumably by influencing norepinephrine anddopamine brainstem nuclei (40).Converging sources of evidence implicate the
cerebellum in timing of neural processes associatedwith not only motor, but also perceptual andcognitive functioning (6, 7, 41). Patients withcerebellar lesions have deficits in both time per-ception and production tasks (41, 42). Further-more, increased regional cerebral blood flow in thecerebellum has been demonstrated using positronemission tomography during an auditory intervaldiscrimination task (42). Because of the striking
uniformity of cerebellar architecture at the micro-anatomical level, it has been suggested that thecerebellum performs similar operations on infor-mation from different functional domains (e.g., 6,12). Therefore, given evidence of a cerebellar rolein both neural timing and affective functioning, aswell as reported structural abnormalities in BD,characteristic symptoms of BD may be related todeficient neural timing processes mediated by thecerebellum.While structural abnormalities in the cerebellum
have been reported in BD, less is known about theirfunctional consequences. Therefore, a delay eye-blink conditioning procedure was used to investi-gate the functional integrity of the cerebellumand itsassociated timing properties in BD. The neuralcircuitry associated with different forms of condi-tioning is distinct and well characterized, andincludes structures and pathways implicated in BD(e.g., 36, 37). The cerebellum is particularly criticalfor learning in the delay form of single-cue, classicaleyeblink conditioning (43, 44). While brain struc-tures other than the cerebellum are implicated intiming (e.g., basal ganglia), the cerebellum is thecritical brain structure for generating and timingconditioned eyeblink responses, most especiallywhen the time interval is short and the conditionedstimulus (CS) (e.g., a tone) and unconditionedstimulus (US) (e.g., an air puff) co-terminate, as isthe case in the delay eyeblink conditioning proce-dure. Furthermore, the magnitudes of the responsesduring conditioning are related to the morphologyand volume of the cerebellum in humans (45, 46),and thus could be altered by cerebellar atrophyobserved in BD. However, despite increasingevidence for cerebellar involvement in mood disor-ders, this well-established cerebellar-dependentmethodology has not been studied in BD.While the preponderance of evidence, especially
from rodent studies, indicates that the cerebellumand brainstem circuits are necessary and sufficientfor eyeblink conditioning, the involvement of otherareas of the brain cannot be definitively ruled out.An important issue in the present study is whethereyeblink conditioning can be influenced by abnor-mal function of the PFC, a brain region that hasbeen consistently implicated in BD (e.g., 47–49).Evidence in the animal literature suggests that thePFC does not play a role in the acquisition ofconditioned responses (CRs) in the delay eyeblinkconditioning procedure. For example, decerebraterodents can acquire and appropriately time re-sponses in the delay form of eyeblink conditioning,suggesting that only the brainstem and cerebellumare necessary for conditioning to occur (50). Inaddition, numerous studies have shown that PFC
Bolbecker et al.
20

lesions, especially to medial aspects of the PFC, canaffect trace eyeblink conditioning, but that delayconditioning is unimpaired by such lesions (51–53).However, it has been shown that the PFC, while notinvolved in acquisition, may be involved in retrievalof CRs, but not the amplitude or timing of theretrieved CRs (54). Nevertheless, because the PFCis not the same in human and non-human animals,it is difficult to generalize from these findings.Therefore, since the PFC has been implicated in BDand shares projections with the cerebellum, thepossibility that abnormal activation of the humanPFC may in turn influence the cerebellum duringconditioning cannot be ruled out. Even so, theavailable evidence, albeit primarily from non-human studies, argues against PFC involvementin the acquisition of delay eyeblink conditioning.In the delay eyeblink conditioning procedure,
repeated pairings of a tone (CS) with a forthcomingaversive corneal air puff (US) result in the occur-rence of an anticipatory eyelid closure just prior tothe onset of the air puff. This learned eyeblinkresponse is the CR. It was hypothesized that BDwould be associated with impaired acquisition (i.e.,fewer CRs) and impaired timing (i.e., short CRlatencies) of conditioned eyeblink responses. Fur-thermore, it was predicted that clinical status wouldbe associated with eyeblink conditioning, withsymptomatic BD patients, and to a lesser degreeeuthymic patients, showing fewer CRs and shorterlatencies compared to healthy controls (HCs). Wepredicted shorter latencies in BD because previousmorphological and volumetric studies of the cere-bellum in BD from both case histories and con-trolled comparison studies have pointed to wholecerebellum abnormalities (15, 20, 22), vermal atro-phy (16–18), or both cerebellar cortical and vermianatrophy (21). Thus, it was reasonable to predictabnormalities in both components of the cerebellarcircuitry underlying eyeblink conditioning: the deepcerebellar nuclei, which are involved in the acquisi-tion of the CRs, and the inhibitory and excitatorycortical system, which may contribute to acquisi-tion, but also to the precise timing and gain of theCRs (43). Since cortical lesions tend to result in shortlatency conditioned blink responses (55), and wepostulated a generalized cerebellar deficit in BD dueto a paucity of research suggesting localized deficits,we predicted shorter CR latencies in BD.
Methods
Participants
Participants were 28 individuals (12 men and 16women) with DSM-IV BD and 28 age- and sex-
matched non-psychiatric HCs (14 men and 14women). Diagnostic status was determined usingthe Structured Clinical Interview for DSM-IV(SCID) Axis I Disorders sections for mood disor-ders, psychotic disorders, and substance abusedisorders, and chart review. HCs were recruitedthrough newspaper advertisements and fliers, anddid not meet SCID criteria for any Axis I or Axis IIdisorder. Any participant who met criteria forsubstance dependency within three months prior totesting was excluded from the study. Diagnosticinterviews and clinical ratings were performed bytrained research personnel. Kappa inter-rater reli-ability in this laboratory setting has been 0.95 formood disorders versus schizophrenia or otherdiagnoses. The study procedures were approvedby the Indiana University-Purdue University Indi-anapolis Internal Review Board and the study wasconducted in accordance with the Declaration ofHelsinki (Edinburgh amendments). Written in-formed consent was obtained from all participants.The BD group included individuals tested during
manic (n = 9), mixed (n = 8), and euthymic(n = 11) episodes. Acute symptom severity duringthe preceding week was assessed using the YoungMania Rating Scale (56) and Montgomery-AsbergDepression Rating Scale (57) (see Table 1). Noconditioning variables were significantly correlatedwith these measures.Healthy controls were recruited and included in
the study if their age fell within two years of abipolar participant�s in order to ensure that thegroups were age matched. The mean age in years ofBD participants (41.39, SD = 11.89) did not differfrom controls (41.79, SD = 12.02), nor did themean age of the BD subtypes. Inclusion criteriawere completion of grade-school level education,normal or corrected to normal hearing and vision,no history of cardiovascular or neurological dis-ease, and no history of head injury that resulted inloss of consciousness. The BD group did not differfrom the HC group on educational level attained[v2(6) = 7.503, p = 0.277]. In both groups, themajority of participants had some college educa-tion (BD = 11, HC = 14) or had at least abachelor�s degree (BD = 9, HC = 10).
Table 1. Means and standard deviations of Young Mania Rating Scale(YMRS) and Montgomery-Asberg Depression Rating Scale (MADRS)scores for bipolar disorder subtypes
Bipolar subtypeYMRS sum scoreMean (SD)
MADRS sum scoreMean (SD)
Manic (n = 9) 24.8 (7.2) 7.5 (3.7)Euthymic (n = 11) 2.5 (2.6) 4.5 (2.8)Mixed (n = 8) 28.3 (3.9) 19.4 (7.9)
Eyeblink conditioning in bipolar disorder
21
Seven individuals with BD were unmedicatedat the time of testing and two were enrolled in aclinical study in which medication status has yetto be unblinded. Of the remaining 19 BDparticipants, 11 were on antipsychotics (10atypical, 1 typical), 11 were on mood stabilizers,and 6 were on other selective serotonin reuptakeinhibitors (SSRIs) or tricyclic antidepressants.
Eyeblink conditioning procedure
Participants completed a single-cue tone delayeyeblink conditioning task. A previous publica-tion by Brown et al. (58) describes this procedurein detail. The CS was a 400 ms, 1000 Hz (80 dBSPL) tone, which, on paired trials, co-terminatedwith a 50 ms air puff, the US. Subjects werepresented with eight US-alone trials with anintertrial interval (ITI) of 15 sec, followed by 10blocks of conditioning trials (mean ITI = 15 sec;range = 10 to 20 sec). The eight US-alone trialswere included to determine whether groups dif-fered in the performance of unconditioned blinkresponses before the conditioning phase began.Each of the 10 conditioning trial blocks consistedof nine CS-US paired trials and one CS-alonetrial. In order to maintain the participants�attention throughout the experiment, neutralphotographs selected from the InternationalAffective Picture System (59) were presented(2 sec duration) between each trial, and partici-pants rated the pleasantness of the images on aresponse pad. In addition, participants wereobserved via a closed-circuit monitor to ensurethat their eyes remained open. The experimentwas briefly suspended if signs of fatigue wereobserved so that the examiner could interact withthe participant.
Electromyographic recording procedure
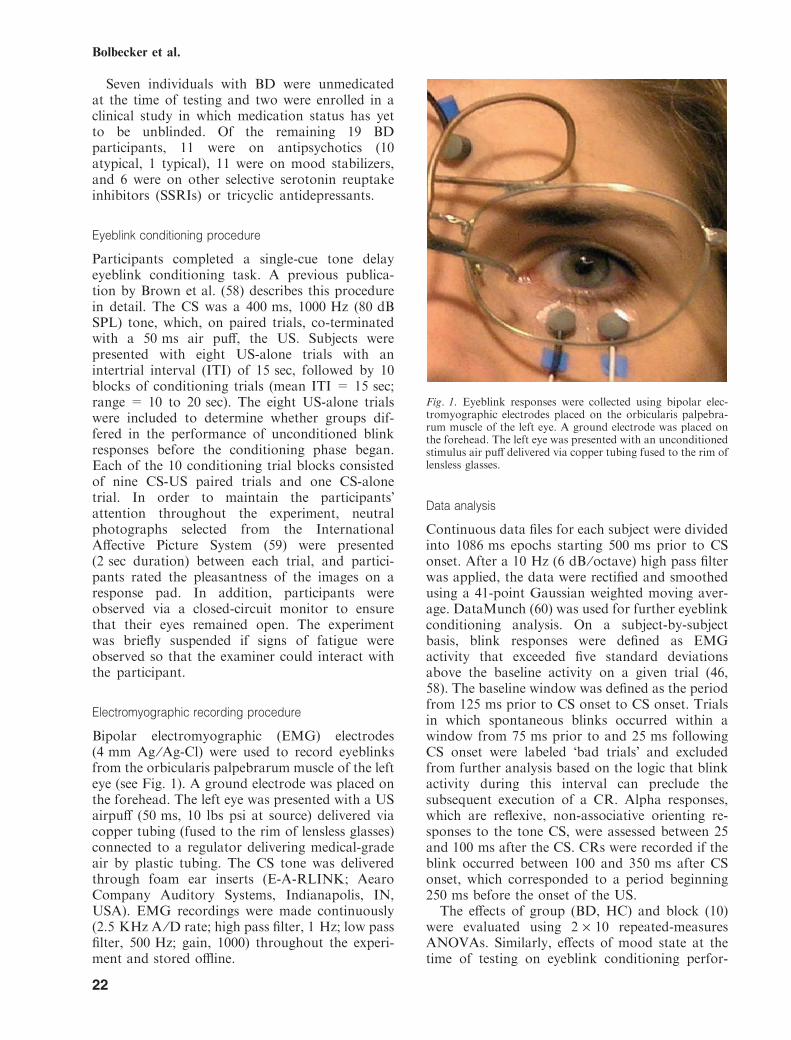
Bipolar electromyographic (EMG) electrodes(4 mm Ag ⁄Ag-Cl) were used to record eyeblinksfrom the orbicularis palpebrarum muscle of the lefteye (see Fig. 1). A ground electrode was placed onthe forehead. The left eye was presented with a USairpuff (50 ms, 10 lbs psi at source) delivered viacopper tubing (fused to the rim of lensless glasses)connected to a regulator delivering medical-gradeair by plastic tubing. The CS tone was deliveredthrough foam ear inserts (E-A-RLINK; AearoCompany Auditory Systems, Indianapolis, IN,USA). EMG recordings were made continuously(2.5 KHz A ⁄D rate; high pass filter, 1 Hz; low passfilter, 500 Hz; gain, 1000) throughout the experi-ment and stored offline.
Data analysis
Continuous data files for each subject were dividedinto 1086 ms epochs starting 500 ms prior to CSonset. After a 10 Hz (6 dB ⁄octave) high pass filterwas applied, the data were rectified and smoothedusing a 41-point Gaussian weighted moving aver-age. DataMunch (60) was used for further eyeblinkconditioning analysis. On a subject-by-subjectbasis, blink responses were defined as EMGactivity that exceeded five standard deviationsabove the baseline activity on a given trial (46,58). The baseline window was defined as the periodfrom 125 ms prior to CS onset to CS onset. Trialsin which spontaneous blinks occurred within awindow from 75 ms prior to and 25 ms followingCS onset were labeled �bad trials� and excludedfrom further analysis based on the logic that blinkactivity during this interval can preclude thesubsequent execution of a CR. Alpha responses,which are reflexive, non-associative orienting re-sponses to the tone CS, were assessed between 25and 100 ms after the CS. CRs were recorded if theblink occurred between 100 and 350 ms after CSonset, which corresponded to a period beginning250 ms before the onset of the US.The effects of group (BD, HC) and block (10)
were evaluated using 2 · 10 repeated-measuresANOVAs. Similarly, effects of mood state at thetime of testing on eyeblink conditioning perfor-
Fig. 1. Eyeblink responses were collected using bipolar elec-tromyographic electrodes placed on the orbicularis palpebra-rum muscle of the left eye. A ground electrode was placed onthe forehead. The left eye was presented with an unconditionedstimulus air puff delivered via copper tubing fused to the rim oflensless glasses.
Bolbecker et al.
22
mance were assessed by comparing bipolar sub-types (manic, mixed, euthymic, HC) in 4 · 10repeated-measures ANOVAs. In cases where therewas a significant departure from the assumption ofsphericity, the Huyn-Feldt corrected values wereused. Post hoc analyses were calculated to evaluatethe relationship between mood state and eyeblinkconditioning variables.Results of themajor depend-ent variables are reported with their correspond-ing effect sizes in the form of partial eta2 (gP
2). Arough estimate of effect size was provided byCohen (61): small effect sizes are less than 0.06;moderate effect sizes range from 0.06 to 0.14; largeeffect sizes are greater than 0.14. Alpha was set atp < 0.05.
Results
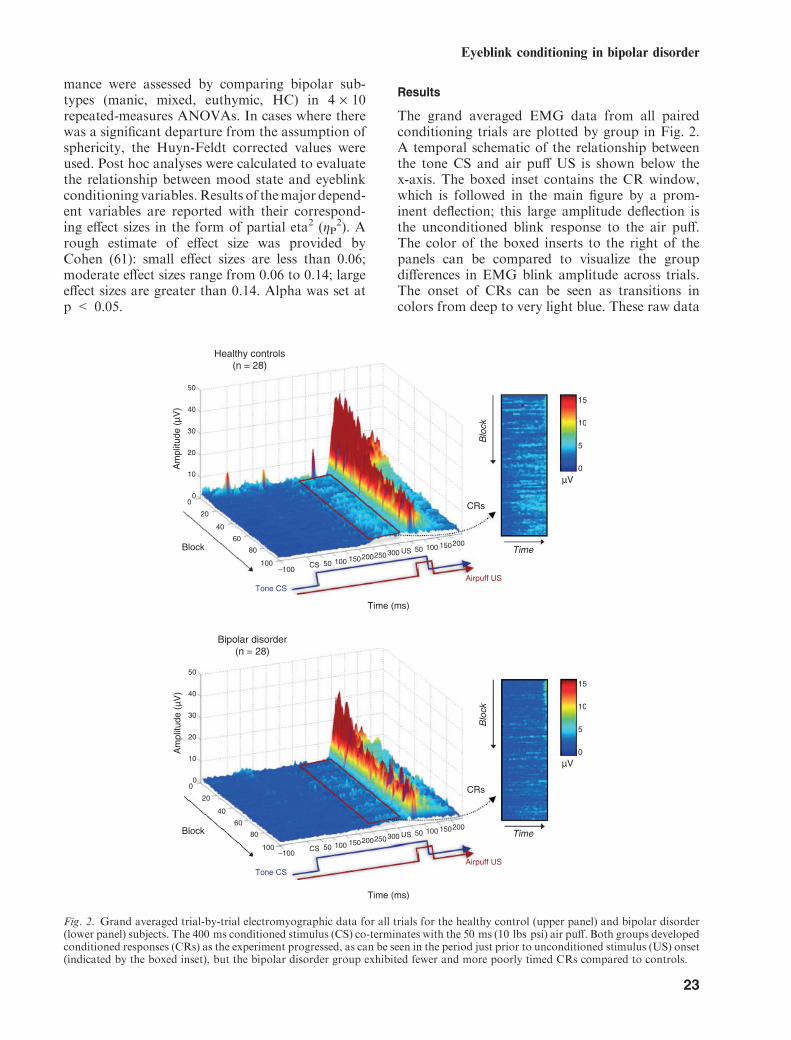
The grand averaged EMG data from all pairedconditioning trials are plotted by group in Fig. 2.A temporal schematic of the relationship betweenthe tone CS and air puff US is shown below thex-axis. The boxed inset contains the CR window,which is followed in the main figure by a prom-inent deflection; this large amplitude deflection isthe unconditioned blink response to the air puff.The color of the boxed inserts to the right of thepanels can be compared to visualize the groupdifferences in EMG blink amplitude across trials.The onset of CRs can be seen as transitions incolors from deep to very light blue. These raw data
Fig. 2. Grand averaged trial-by-trial electromyographic data for all trials for the healthy control (upper panel) and bipolar disorder(lower panel) subjects. The 400 ms conditioned stimulus (CS) co-terminates with the 50 ms (10 lbs psi) air puff. Both groups developedconditioned responses (CRs) as the experiment progressed, as can be seen in the period just prior to unconditioned stimulus (US) onset(indicated by the boxed inset), but the bipolar disorder group exhibited fewer and more poorly timed CRs compared to controls.
Eyeblink conditioning in bipolar disorder
23
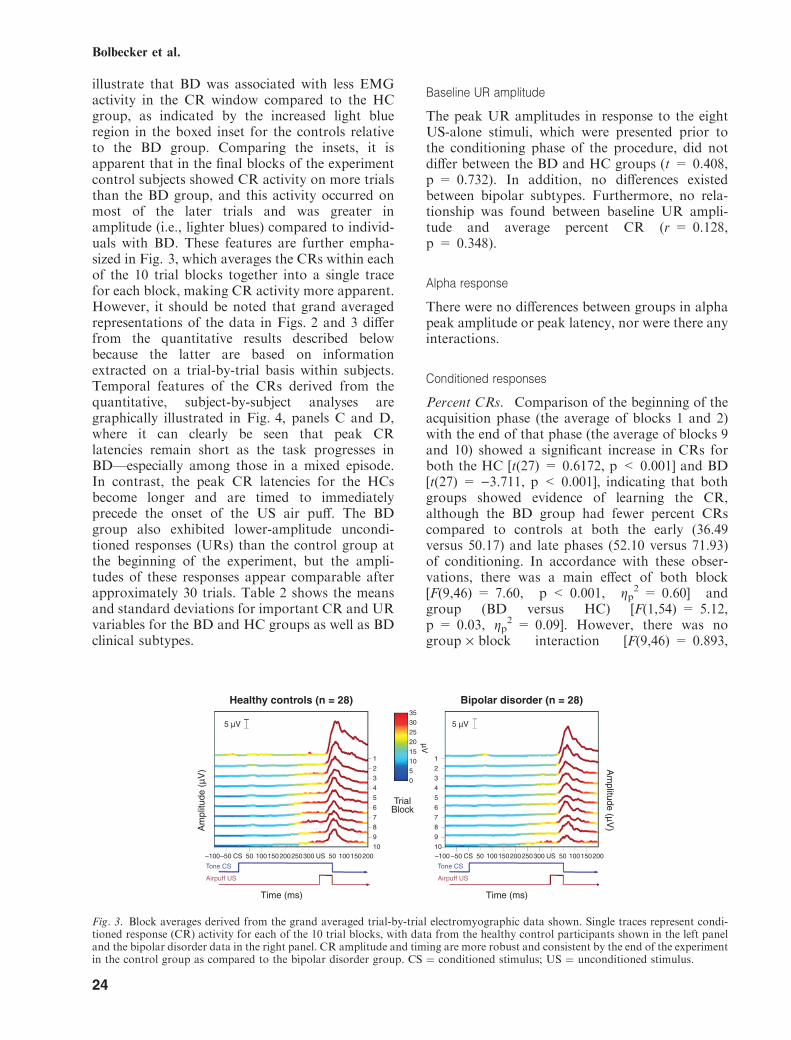
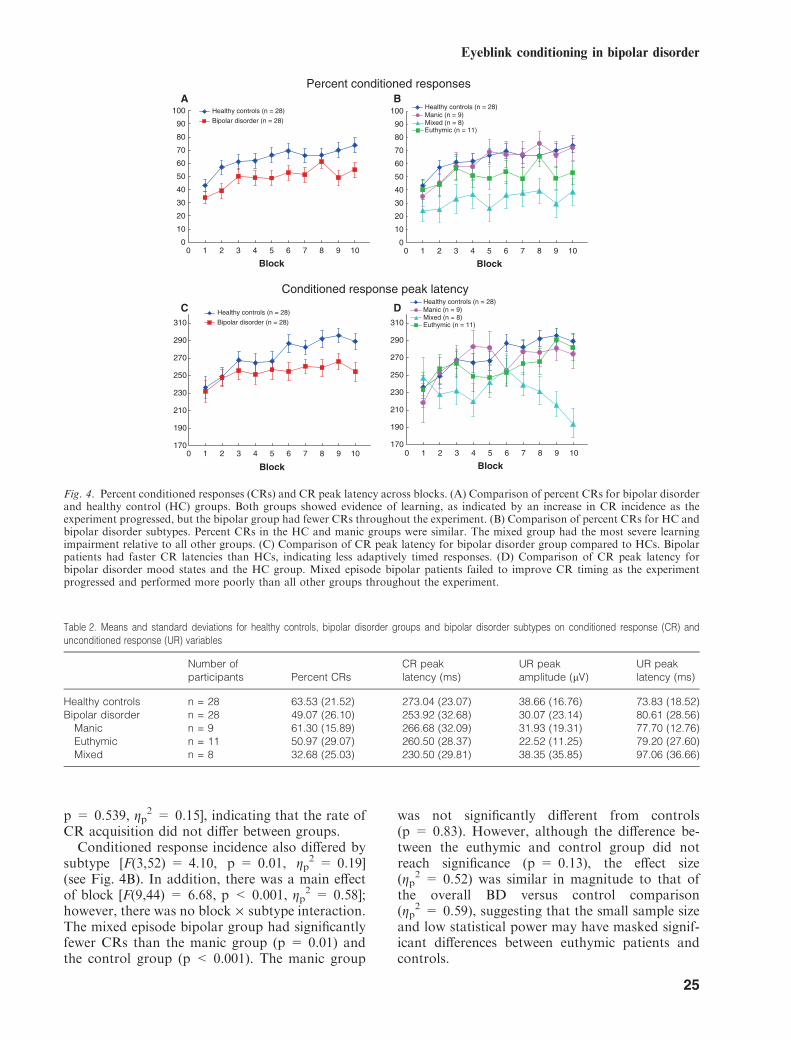
illustrate that BD was associated with less EMGactivity in the CR window compared to the HCgroup, as indicated by the increased light blueregion in the boxed inset for the controls relativeto the BD group. Comparing the insets, it isapparent that in the final blocks of the experimentcontrol subjects showed CR activity on more trialsthan the BD group, and this activity occurred onmost of the later trials and was greater inamplitude (i.e., lighter blues) compared to individ-uals with BD. These features are further empha-sized in Fig. 3, which averages the CRs within eachof the 10 trial blocks together into a single tracefor each block, making CR activity more apparent.However, it should be noted that grand averagedrepresentations of the data in Figs. 2 and 3 differfrom the quantitative results described belowbecause the latter are based on informationextracted on a trial-by-trial basis within subjects.Temporal features of the CRs derived from thequantitative, subject-by-subject analyses aregraphically illustrated in Fig. 4, panels C and D,where it can clearly be seen that peak CRlatencies remain short as the task progresses inBD—especially among those in a mixed episode.In contrast, the peak CR latencies for the HCsbecome longer and are timed to immediatelyprecede the onset of the US air puff. The BDgroup also exhibited lower-amplitude uncondi-tioned responses (URs) than the control group atthe beginning of the experiment, but the ampli-tudes of these responses appear comparable afterapproximately 30 trials. Table 2 shows the meansand standard deviations for important CR and URvariables for the BD and HC groups as well as BDclinical subtypes.
Baseline UR amplitude
The peak UR amplitudes in response to the eightUS-alone stimuli, which were presented prior tothe conditioning phase of the procedure, did notdiffer between the BD and HC groups (t = 0.408,p = 0.732). In addition, no differences existedbetween bipolar subtypes. Furthermore, no rela-tionship was found between baseline UR ampli-tude and average percent CR (r = 0.128,p = 0.348).
Alpha response
There were no differences between groups in alphapeak amplitude or peak latency, nor were there anyinteractions.
Conditioned responses
Percent CRs. Comparison of the beginning of theacquisition phase (the average of blocks 1 and 2)with the end of that phase (the average of blocks 9and 10) showed a significant increase in CRs forboth the HC [t(27) = 0.6172, p < 0.001] and BD[t(27) = )3.711, p < 0.001], indicating that bothgroups showed evidence of learning the CR,although the BD group had fewer percent CRscompared to controls at both the early (36.49versus 50.17) and late phases (52.10 versus 71.93)of conditioning. In accordance with these obser-vations, there was a main effect of both block[F(9,46) = 7.60, p < 0.001, gp
2 = 0.60] andgroup (BD versus HC) [F(1,54) = 5.12,p = 0.03, gp
2 = 0.09]. However, there was nogroup · block interaction [F(9,46) = 0.893,
Fig. 3. Block averages derived from the grand averaged trial-by-trial electromyographic data shown. Single traces represent condi-tioned response (CR) activity for each of the 10 trial blocks, with data from the healthy control participants shown in the left paneland the bipolar disorder data in the right panel. CR amplitude and timing are more robust and consistent by the end of the experimentin the control group as compared to the bipolar disorder group. CS ¼ conditioned stimulus; US ¼ unconditioned stimulus.
Bolbecker et al.
24
p = 0.539, gp2 = 0.15], indicating that the rate of
CR acquisition did not differ between groups.Conditioned response incidence also differed by
subtype [F(3,52) = 4.10, p = 0.01, gp2 = 0.19]
(see Fig. 4B). In addition, there was a main effectof block [F(9,44) = 6.68, p < 0.001, gp
2 = 0.58];however, there was no block · subtype interaction.The mixed episode bipolar group had significantlyfewer CRs than the manic group (p = 0.01) andthe control group (p < 0.001). The manic group
was not significantly different from controls(p = 0.83). However, although the difference be-tween the euthymic and control group did notreach significance (p = 0.13), the effect size(gp
2 = 0.52) was similar in magnitude to that ofthe overall BD versus control comparison(gp
2 = 0.59), suggesting that the small sample sizeand low statistical power may have masked signif-icant differences between euthymic patients andcontrols.
Fig. 4. Percent conditioned responses (CRs) and CR peak latency across blocks. (A) Comparison of percent CRs for bipolar disorderand healthy control (HC) groups. Both groups showed evidence of learning, as indicated by an increase in CR incidence as theexperiment progressed, but the bipolar group had fewer CRs throughout the experiment. (B) Comparison of percent CRs for HC andbipolar disorder subtypes. Percent CRs in the HC and manic groups were similar. The mixed group had the most severe learningimpairment relative to all other groups. (C) Comparison of CR peak latency for bipolar disorder group compared to HCs. Bipolarpatients had faster CR latencies than HCs, indicating less adaptively timed responses. (D) Comparison of CR peak latency forbipolar disorder mood states and the HC group. Mixed episode bipolar patients failed to improve CR timing as the experimentprogressed and performed more poorly than all other groups throughout the experiment.
Table 2. Means and standard deviations for healthy controls, bipolar disorder groups and bipolar disorder subtypes on conditioned response (CR) andunconditioned response (UR) variables
Number ofparticipants Percent CRs
CR peaklatency (ms)
UR peakamplitude (lV)
UR peaklatency (ms)
Healthy controls n = 28 63.53 (21.52) 273.04 (23.07) 38.66 (16.76) 73.83 (18.52)Bipolar disorder n = 28 49.07 (26.10) 253.92 (32.68) 30.07 (23.14) 80.61 (28.56)
Manic n = 9 61.30 (15.89) 266.68 (32.09) 31.93 (19.31) 77.70 (12.76)Euthymic n = 11 50.97 (29.07) 260.50 (28.37) 22.52 (11.25) 79.20 (27.60)Mixed n = 8 32.68 (25.03) 230.50 (29.81) 38.35 (35.85) 97.06 (36.66)
Eyeblink conditioning in bipolar disorder
25
CR peak timing. As learning occurs across theconditioning phase, the timing of the CR shouldshift to more precisely approximate the onset of theair puff; hence, CRs which occur shortly after CSonset are less adaptive compared to CRs occurringlater and immediately prior to US onset. Aspredicted, CR peak latency increased acrossblocks for both groups [F(7.27,392.31) = 4.59,p < 0.001, gP
2 = 0.08], but the BD group�s laten-cies were consistently shorter than the controlgroup, yielding a main effect of group[F(1,54) = 6.40, p = 0.01, gP
2 = 0.11] (seeFig. 4C). There was no significant block · groupinteraction. Differences between groups on theaverage peak latency for blocks 1–5 and for blocks6–10 were then compared. While there was nosignificant difference between groups during thefirst half of the experiment (t = 0.897, p = 0.374),the bipolar group had significantly shorter latenciesthan the control group (t = 3.498, p < 0.001)during the second half of the conditioning phase.These results suggest that the controls learned to
more accurately time CRs relative to BD patientsduring the final blocks on conditioning.Conditioned response peak latency also differed
by bipolar subtype [F(3,52) = 5.38, p < 0.01,gP
2 = 0.24]. The mixed group exhibited shorterlatency CRs relative to all other subtypes andcontrols: controls (p < 0.001), manic (p < 0.01),and euthymic (p = 0.02) (see Fig. 4D). There wasno block · subtype interaction.
Unconditioned responses
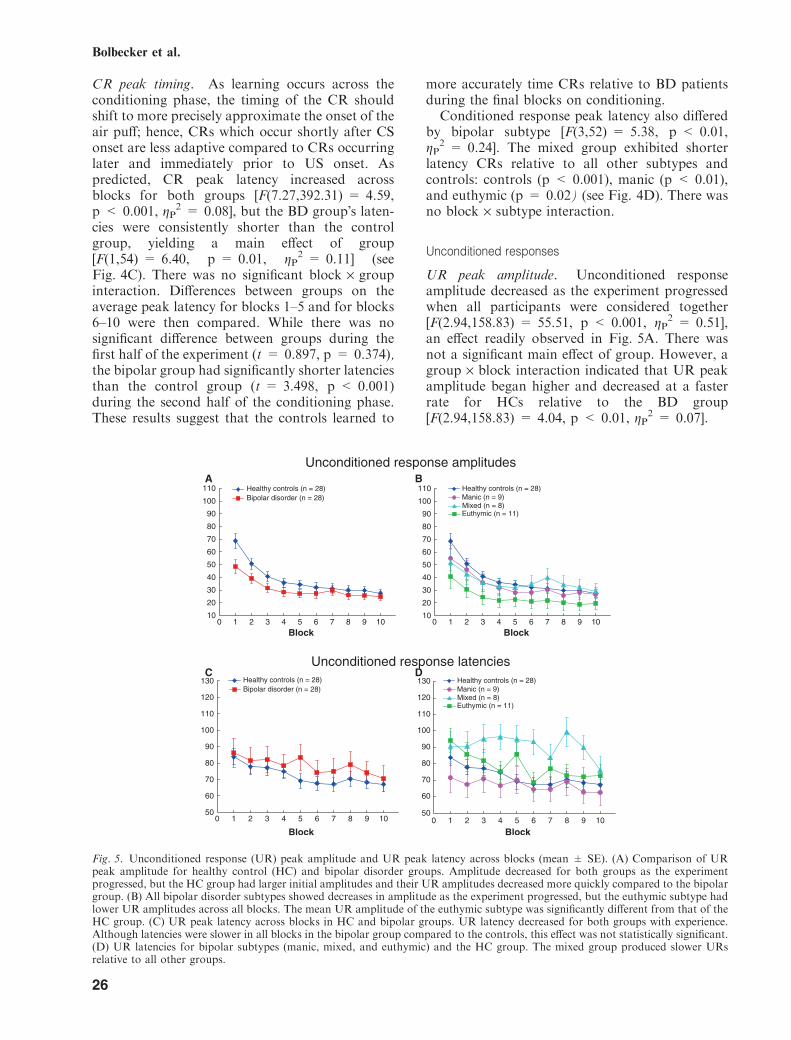
UR peak amplitude. Unconditioned responseamplitude decreased as the experiment progressedwhen all participants were considered together[F(2.94,158.83) = 55.51, p < 0.001, gP
2 = 0.51],an effect readily observed in Fig. 5A. There wasnot a significant main effect of group. However, agroup · block interaction indicated that UR peakamplitude began higher and decreased at a fasterrate for HCs relative to the BD group[F(2.94,158.83) = 4.04, p < 0.01, gP
2 = 0.07].
Fig. 5. Unconditioned response (UR) peak amplitude and UR peak latency across blocks (mean ± SE). (A) Comparison of URpeak amplitude for healthy control (HC) and bipolar disorder groups. Amplitude decreased for both groups as the experimentprogressed, but the HC group had larger initial amplitudes and their UR amplitudes decreased more quickly compared to the bipolargroup. (B) All bipolar disorder subtypes showed decreases in amplitude as the experiment progressed, but the euthymic subtype hadlower UR amplitudes across all blocks. The mean UR amplitude of the euthymic subtype was significantly different from that of theHC group. (C) UR peak latency across blocks in HC and bipolar groups. UR latency decreased for both groups with experience.Although latencies were slower in all blocks in the bipolar group compared to the controls, this effect was not statistically significant.(D) UR latencies for bipolar subtypes (manic, mixed, and euthymic) and the HC group. The mixed group produced slower URsrelative to all other groups.
Bolbecker et al.
26

While there was not a significant main effect ofsubtype [F(3,52) = 1.630, p = 0.20], exploratorypost hoc analyses indicated a significant differencebetween the HC group and the euthymic group(p = 0.03). The control group had consistentlylarger amplitudes than any of the BD groups andthe euthymic group had the lowest mean ampli-tudes of all groups in every block (Fig. 5B).
UR peak latency. Unconditioned response peaklatency decreased with experience when partici-pants were collapsed across groups (seeFig. 5C), resulting in a main effect of block[F(7.26,392.94) = 6.20, p < 0.001, gP
2 = 0.10].There was no main effect of group or ablock · group interaction.While there was not a main effect of subtype,
exploratory post hoc analyses indicated that thepeak UR latency of the mixed group was signifi-cantly longer than the HC group (p = 0.02) andthe manic group (p = 0.01) (see Fig. 5D). Therewas no block · subtype interaction.
Bad trials analyses. The bad trial window exists toexclude trials where EMG activity is increasedimmediately before ()75 ms) and shortly after(+25 ms) CS onset. If a subject exhibits EMGblink activity during this interval, it is questionablewhether a CR can be emitted immediately there-after. Stated differently, spontaneous blinks occur-ring immediately prior to and following CS onset(i.e., in the bad trial window) may interfere withthe subsequent execution of a CR. It should benoted that blinks recorded in the bad trial windoware considered spontaneous blinks because theyoccur too early in reference to CS onset to beconsidered either tone related or conditioningrelated. Accordingly, the number of bad trialscan be used as a rough index of spontaneous blinkrate.The average number of bad trials rejected from
analysis in the bipolar group was 18.8 (SD =10.3), whereas for the HC group it was 14.2(SD = 6.5), which was significantly different(p = 0.05). Importantly, group differences re-mained unchanged when the number of bad trialswas used as a covariate for the percent CRs[F(1,53) = 6.20, p = 0.02, gP
2 = 0.11] and peaklatency analyses [F(1,53) = 5.40, p = 0.02,gP
2 = 0.09].
Medication effects
Complete medication information was availablefor 26 of 28 BD participants. The remaining twoBD patients were in a blinded clinical trial at the
time of participation and were either unmedicatedor on licarbamezapine monotherapy. Seven partic-ipants with BD were unmedicated at the time oftesting, having discontinued their medicationsprior to admission to hospital. Of the remaining19 BD participants, 11 were on antipsychotics (10atypical, 1 typical), 11 were on mood stabilizers,and 6 were on other SSRIs or tricyclic antidepres-sants.The seven unmedicated BD participants were
compared with their seven age-matched HCs on thepercentage of CRs and CR peak latencies, theprimary dependent variables of interest, in a 2(group: HCs, unmedicated BD) · 10 (block) re-peated-measuresANOVA. In this analysis, themaineffect of group on percent CRs yielded a large effectsize (gP
2 = 0.25), indicating decreased CRs in BD,but this difference was significant at a trend level dueto the small sample sizes and decreased statisticalpower [F(1,12) = 4.01, p = 0.07]. There was also amain effect of block [F(5.93,71.14) = 5.40,p < 0.001, gP
2 = 0.31], but no block by groupinteraction [F(5.93,71.14) = 1.32, p = 0.26,gP
2 = 0.10], as was the case when the entire bipolargroup was analyzed. The seven unmedicated BDparticipants also exhibited significantly shorter CRonset latencies compared to their age-matched HCs[F(1,12) = 6.63, p = 0.02].Again, the effect size forthis analysis was larger (gP
2 = 0.36) than wasobserved when all HC and bipolar participantswere analyzed (gP
2 = 0.11). There was no maineffect of block [F(5.69,68.21) = 1.48, p = 0.20,gP
2 = 0.11] and no interaction [F(5.69,68.21)= 0.80, p = 0.57, gP
2 = 0.06]. The fact that theeffect sizes for these groupdifferences in percentCRsand CR peak latencies were larger when only theunmedicated BD participants were compared totheir age-matched HCs suggests that medicationsmay in fact attenuate eyeblink conditioning deficitsrather than be the source of the effects in bipolarparticipants.In another analysis, chlorpromazine equivalent
dosage was calculated for the 10 patients onatypical antipsychotics (reliable dosage informa-tion was unavailable for the one patient who hadreceived the typical antipsychotic haloperidol priorto testing). Chlorpromazine dose was not signifi-cantly correlated with average percent CRs(r = 0.26, p = 0.89) or average CR peak latency(r = 0.06, p = 0.87). Similarly, valproate dosage(n = 7) was not significantly correlated withpercent CRs (r = )0.06, p = 0.89) or CR peaklatency (r = )0.385, p = 0.39). Because onlythree participants were taking lithium, its effectswere not analyzed. Reliable analyses of medicationeffects comparable to those reported above were
Eyeblink conditioning in bipolar disorder
27

not feasible for the bipolar subtypes due to theeven smaller sample sizes.
Discussion
Individuals with BD demonstrated performanceand timing deficits in a delay eyeblink conditioningtask that depends critically on the cerebellum andrelated brain stem nuclei. Interestingly, perfor-mance on this task covaried with emotional state,with the most pronounced conditioning deficitsseen in participants with the greatest mood lability(i.e., current mixed mood episode). These findingsare consistent with models in which cerebellardysfunction may contribute to poor temporalcoordination of information processing and emo-tional regulation in BD (14, 33), and warrantfurther investigation as to their source and clinicalcorrelates.Evidence for impairment in the functional
integrity of the cerebellum and related brain stemcircuits in BD was indicated by two primaryfindings. First, the bipolar group produced signif-icantly fewer CRs overall. Second, when CRs wereproduced, they were timed unusually early com-pared with US air puff onset. Moreover, timing ofthe CRs of the control and BD groups were similarduring the first half of the experiment, but becamesignificantly different during the last half of theexperiment, suggesting that the control grouplearned to produce more adaptively timed CRsrelative to the bipolar group. These latency effectsin the bipolar group could not be attributed to ageneral slowing of neural processes because CRlatencies were faster in the bipolar group than thecontrol group. Importantly, in terms of support forthe cerebellar-mediated dysregulation hypothesis,the observed deficits in the performance and timingof CRs were most pronounced in the subgroup ofindividuals experiencing a mixed mood episode,which was the most emotionally labile bipolarsubgroup.Several potential statistical and interpretive
confounds were examined to further substantiatethese conclusions. First, the bipolar group exhib-ited significantly fewer conditioned blink responsesacross the acquisition phase, an effect that wasespecially evident in the mixed episode subtype;however, the absence of a group by block statisticalinteraction raised the possibility that the bipolargroup may have had a pre-existing deficit in theperformance of blinks rather than a cerebellar-mediated eyeblink conditioning deficit per se. Anumber of observations run counter to this inter-pretation. Neither the bipolar group as a whole norany of the subtypes differed from the HCs in the
performance of unconditioned blink responses tothe solitary air puffs presented before the condi-tioning phase, indicating comparable motorresponsiveness between groups. In addition, bipo-lar participants showed a small but statisticallysignificant increase in blink activity on an index ofspontaneous blinks which, although uncorrelatedwith conditioning, would tend to inflate thepercentage of conditioned eyeblink responsesrather than lead to the observed decrease. Takentogether, these findings argue against a precondi-tioning blink performance deficit in BD that couldhave accounted for the observed conditioningabnormalities and strengthen the interpretationthat the observed findings suggest cerebellar dys-function in the disorder. Nevertheless, as discussedfurther below, more research will be necessary torule out the possibility of a non-cerebellar-medi-ated cause of the observed deficits in BD, includingabnormalities in brainstem nuclei receiving outputfrom the cerebellum (e.g., red nucleus).A second potential confound concerned possible
effects of psychotropic medication treatment oneyeblink conditioning, specifically the possibilitythat medication use may have impaired condition-ing. This possibility appears unlikely, given thatthe statistical effect size of the decrease in thepercentage of CRs was larger for the medication-withdrawn bipolar compared to the bipolar groupas a whole when contrasted with the age-matchedHC participants. In addition, antipsychotic andvalproate medication dosage did not covary withthe conditioning variables. Nevertheless, furtherstudy of these potential confounds is warranted inlarger patient samples—especially never-medicatedand currently medication-withdrawn patients,given that the bipolar participants were on multiplemedications and there is no reliable methodto account for possible interactions betweenmedications.Both symptomatic and euthymic individuals
with BD were studied, making it possible toaddress the question of whether impaired eyeblinkconditioning performance in this disorder is a stateor a trait effect. The euthymic subgroup performedsimilarly to the symptomatic subgroups as a whole(i.e., mixed and manic subtypes combined) on CRacquisition, but they were not significantly differentfrom controls. However, similar effect size magni-tudes were observed when the symptomatic bipolargroup was compared with controls (gP
2 = 0.65)and when the euthymic subgroup was compared tocontrols (gP
2 = 0.52). Although these comparisonsare consistent with a trait-like effect, moreresearch, with large sample sizes and greatervariability in mood state will be necessary to
Bolbecker et al.
28

properly disentangle state and trait factors. Inaddition, the potential contribution of depressivesymptoms to the conditioning deficits exhibited bythe mixed group requires further study, especiallygiven recent evidence of eyeblink conditioningdeficits (62) and cerebellar atrophy (23, 24) inunipolar depression.On the basis of an extensive animal and human
literature, the present findings implicate the cere-bellar cortex, the interpositus nucleus, and ⁄orbrainstem nuclei receiving cerebellar output in thedeficits observed in BD. Animal studies haveshown that the critical CS-US association occursin the anterior interpositus nucleus (63–65), one ofthe deep cerebellar nuclei. The cortex of thecerebellum is thought to control the expression ofCRs and modulate both their timing and ampli-tude (54, 66, 67). The anterior lobe of the cerebel-lum seems to be specifically involved in responsetiming. For example, lesions to the anterior lobe inrabbits caused previously acquired responses tooccur at a fixed, short latency (i.e., non-adaptive),suggesting that this region plays a role in delayingthe onset of CRs until just prior to the US onset(55, 68). Findings from these animal studies aresupported by the human literature (69). Togetherwith the present results, these previous studiessuggest possible abnormalities in the anteriorinterpositus nuclei and cerebellar cortex, both ofwhich could underlie the observed CR acquisitiondeficits. Possible abnormalities in the anterior lobeof the cerebellar cortex in the BD group are morespecifically suggested by a failure to adaptivelytime CRs. The CR peak latencies did not differbetween bipolar patients and controls during theinitial half of the conditioning phase; however,during the final half, controls produced longer CRlatencies, which more closely preceded the onset ofthe air puff. In other words, the bipolar group wasunable to adaptively delay their conditioned blinkresponses to coincide with the air puff, which isconsistent with anterior lobe dysfunction, asdescribed above. Finally, while the observeddeficits in eyeblink conditioning may be relatedto a failure in patients with BD to acquire acerebellar-mediated CR, it is important to notealternative explanations that require further study.For example, it could be that the cerebellarcircuitry was in fact intact in BD but that a criticalnucleus downstream from the cerebellum did notpropagate the learning signal, thus leading to afailure to produce a CR. The interpositus nucleusof the cerebellum projects to the red nucleus, whoseinactivation abolishes expression of CRs (70). Asshown in the UR analysis, BD patients did produceblinks, but it is still possible that the CR message
was relayed incorrectly to the red nucleus. Hence,rather than a learning deficit, a production deficitcould cause the reduced incidence of CRs in BD.Another possible explanation for a reduction inCRs is that in the bipolar subjects fewer neuronswere recruited in the cerebellar circuitry responsi-ble for CR production, thus leading to smaller CRsthat did not rise to the threshold required to belabeled as such by our data analysis technique.Interestingly, recent evidence suggests that individ-uals with BD experience significant loss of cerebel-lar gray matter across time and that this loss ispositively correlated with increasing numbers ofmood episodes (15). Under these conceptualiza-tions, the results may point to a performancedeficit rather than an acquisition, or learning,deficit.Although the possibility exists that a deficit
exists outside of the cerebellum (in the red nucleusitself, for example), there is evidence that structuralimpairments in the cerebellum are associated withmood disorders. For example, patients with cere-bellar lesions sometimes show dramatic changes inmood regulation, exhibiting symptoms such asmania, depression, and contextually inappropriatebehavior (8–12, 26). In addition, structural abnor-malities have been reported in patients withunipolar depression (23, 24) and BD (15–22).Furthermore, cerebellar stimulation has beenreported to improve mood (28).Further research will be necessary to disentan-
gle the extent to which BD is associated withcerebellar-mediated CR acquisition versus pureproduction deficits. For example, eyeblink condi-tioning procedures that place a premium ontemporal components of this task would beinformative in further investigations of cerebellardysfunction in BD. One such task, the interstim-ulus interval (ISI) shift, is believed to dependmore on cerebellar cortex. Therefore, if cerebellardeficits are indeed responsible for the presentresults, BD patients should show a more pro-nounced ISI shift deficit—irrespective of basallevels of CR production—since the ability to shiftCRs to the new ISI depends on the cerebellarcortex. Hence, although the BD group�s basal rateof responding could be low in the ISI shift task, ifthere is a problem in the red nucleus, this taskwould have a higher probability of uncovering acerebellar deficit if it exists.The observed impairment in eyeblink condition-
ing is consistent with previous reports of neuro-chemical and structural cerebellar anomalies inBD. For example, BD has been linked to deficits incerebellar reelin (71, 72), which is an importantsecretory glycoprotein that guides neurons and glia
Eyeblink conditioning in bipolar disorder
29

during embryonic brain development, ensuringnormal brain lamination, and in the adult brain,may mediate neuroplasticity. With regard tostructural anomalies, BD has been associated withdecreased cerebellar volume (15–22), which iscorrelated with an increased number of moodepisodes (15–17), an index of illness severity. Thisis of particular interest because the most severely illBD participants in the present study, the mixedepisode subgroup, exhibited the most severe defi-cits in cerebellar-mediated eyeblink conditioning.Patients who experience mixed episodes generallyconstitute a more chronic and severe sample. Forexample, mixed episodes have been associated withrapid cycling, an increased number of moodepisodes, longer duration of illness, earlier age ofonset, and fewer symptom-free days per year (73–75). Therefore, the observed eyeblink conditioningimpairments in mixed-episode subjects may beassociated with a more severe course of illness andcorrespondingly severe cerebellar abnormalities.Theoretical and empirical findings suggest that
cerebellar abnormalities may result in temporalprocessing deficits in other functional domainsover which the cerebellum exerts modulatoryinfluence. For example, the striking invariance ofcerebellar circuitry suggests that it performs similaroperations on its neuronal inputs regardless oftheir source (6, 12, 14). Findings of topographicprojections organized in bidirectional informationstreams connecting the cerebellum with motor, pre-frontal, and posterior parietal cortex [Schmahmann(12) provides an excellent review] have led tosuggestions that the cerebellum integrates infor-mation from different functional domains (6, 12,14). This integration may provide the moment-by-moment context for behavioral output, bindingtogether perceptual, cognitive, emotional, andmotivational information from disparate brainregions (14). Given such a model, timing deficitsrevealed in one domain, such as motor timing ineyeblink conditioning, may indicate abnormalitiesin temporal coordination in other domains. It iswell known that cerebellar lesions can lead tomotor dysmetria, which is characterized by tem-poral incoordination of movement. Lauterbach(26) drew an analogy between the motor andaffective consequences he had observed in patientswith cerebellar lesions with particular referenceto BD:
�Loss of cerebellar input leads to … oscillationsproducing tremor, increased tonic muscleactivity, and asynchronous bursting in antago-nist musculature. Slower, more sustained anal-ogous phenomena in oscillating mood circuitsmight include environmentally-reactive labile
mood, exaggerated mood states of mania ordepression, failure to maintain the timing eu-thymia, or rapid cycling bipolar mood swings�(26, p. 728).
Hence, given the anatomical connectionsbetween the cerebellum and prefrontal, limbic,and brainstem regions, emotional effects that areanalogous to the motor effects of cerebellar lesionsmay result in aberrant temporal coordination ofemotional and motivational behavior in mooddisorders such as BD when cerebellar timing isdisrupted (14, 26). To further investigate the theorythat cerebellar-mediated temporal discoordinationmay contribute to symptoms of BD, futureexperiments should include additional cerebellar-dependent tasks that could provide furtherconvergent validity for the eyeblink conditioningtask employed in this research.
Acknowledgements
We are grateful to Misty Bodkins, Jennifer Boggs, ColleenMerrill, and Ashley Steffen for their assistance in collectingeyeblink conditioning data. We also thank the clinical researchteam at Larue D. Carter Memorial Hospital and the IndianaUniversity Neuroscience Clinical Research Center for theirsupport.
References
1. Konarski JZ, McIntyre RS, Grupp LA, Kennedy SH. Isthe cerebellum relevant in the circuitry of neuropsychiatricdisorders? J Psychiatry Neurosci 2005; 30: 178–186.
2. Nestler EJ, Carlezon WA Jr. The mesolimbic dopaminereward circuit in depression. Biol Psychiatry 2006; 59:1151–1159.
3. Kugaya A, Sanacora G. Beyond monoamines: glutama-tergic function in mood disorders. CNS Spectr 2005; 10:808–819.
4. Soares JC, Mann JJ. The functional neuroanatomy ofmood disorders. J Psychiatr Res 1997; 31: 393–432.
5. Carlson PJ, Singh JB, Zarate CA Jr, Drevets WC, ManjiHK. Neural circuitry and neuroplasticity in mood disor-ders: insights for novel therapeutic targets. NeuroRx 2006;3: 22–41.
6. Katz DB, Steinmetz JE. Psychological functions of thecerebellum. Behav Cogn Neurosci Rev 2002; 1: 229–241.
7. Leiner HC, Leiner AL, Dow RS. Cognitive and languagefunctions of the human cerebellum. Trends Neurosci 1993;16: 444–447.
8. Schmahmann JD, Sherman JC. Cerebellar cognitive affec-tive syndrome. Int Rev Neurobiol 1997; 41: 433–440.
9. Schmahmann JD, Sherman JC. The cerebellar cognitiveaffective syndrome. Brain 1998; 121: 561–579.
10. Schmahmann JD. The cerebrocerebellar system: anatomicsubstrates of the cerebellar contribution to cognition andemotion. Int Rev Psychiatry 2001; 13: 247–260.
11. Schmahmann JD. The cerebellar cognitive affective syn-drome: clinical correlations of the dysmetria of thoughthypothesis. Int Rev Psychiatry 2001; 13: 313–322.
12. Schmahmann JD. Disorders of the cerebellum: ataxia,dysmetria of thought, and the cerebellar cognitive affective
Bolbecker et al.
30

syndrome. J Neuropsychiatry Clin Neurosci 2004; 16: 367–378.
13. Andreasen NC. A unitary model of schizophrenia: Bleul-er�s ‘‘fragmented phrene’’ as schizencephaly. Arch GenPsychiatry 1999; 56: 781–787.
14. Schutter DJ, van Honk J. The cerebellum on the rise inhuman emotion. Cerebellum 2005; 4: 290–294.
15. Moorhead TW, McKirdy J, Sussman JE et al. Progressivegray matter loss in patients with bipolar disorder. BiolPsychiatry 2007; 62: 894–900.
16. Mills NP, Delbello MP, Adler CM, Strakowski SM. MRIanalysis of cerebellar vermal abnormalities in bipolardisorder. Am J Psychiatry 2005; 162: 1530–1532.
17. DelBello MP, Strakowski SM, Zimmerman ME, HawkinsJM, Sax KW. MRI analysis of the cerebellum in bipolardisorder: a pilot study. Neuropsychopharmacology 1999;21: 63–68.
18. Lippmann S, Manshadi M, Baldwin H, Drasin G, Rice J,Alrajeh S. Cerebellar vermis dimensions on computerizedtopographic scans of schizophrenic and bipolar patients.Am J Psychiatry 1982; 139: 667–668.
19. Nasrallah HA, Jacoby CG, McCalley-Whitters M. Cere-bellar atrophy in schizophrenia and mania. Lancet 1981;16: 1102.
20. Nasrallah HA, McCalley-Whitters M, Jacoby CG. Corti-cal atrophy in schizophrenia and mania: a comparative CTstudy. J Clin Psychiatry 1982; 43: 439–441.
21. Yadalam KG, Jain AK, Simpson GM. Mania in twosisters with similar cerebellar disturbance. Am J Psychiatry1985; 142: 1067–1069.
22. Cutting JC. Chronic mania in childhood: case report of apossible association with a radiological picture of cerebel-lar disease. Psychol Med 1976; 6: 635–642.
23. Escalona PR, Early B, McDonald WM et al. Reduction ofcerebellar volume in major depression: a controlled MRIstudy. Depression 1993; 1: 156–158.
24. Shah SA, Doraiswamy PM, Husain MM et al. Posteriorfossa abnormalities in major depression: a controlledmagnetic resonance imaging study. Acta Psychiatr Scand1992; 85: 474–479.
25. Cecil KM, DelBello MP, Sellars MC, Strakowski SM.Proton magnetic resonance spectroscopy of the frontallobe and cerebellar vermis in children with a mood disorderand a familial risk for bipolar disorders. J Child AdolescPsychopharmacol 2003; 13: 545–555.
26. Lauterbach EC. Bipolar disorders, dystonia, and compul-sion after dysfunction of the cerebellum, dentatorubrotha-lamic tract, and substantia nigra. Biol Psychiatry 1996; 40:726–730.
27. Neau JP, Arroyo-Anllo E, Bonnaud V, Ingrand P, Gil R.Neuropsychological disturbances in cerebellar infarcts.Acta Neurol Scand 2000; 102: 363–370.
28. Schutter DJ, van Honk J, d�Alfonso AA, Peper JS,Panksepp J. High frequency repetitive transcranial mag-netic over the medial cerebellum induces a shift in theprefrontal electroencephalography gamma spectrum: apilot study in humans. Neurosci Lett 2003; 336: 73–76.
29. Heath RG. Modulation of emotion with a brain pacem-amer. Treatment for intractable psychiatric illness. J NervMent Dis 1977; 165: 300–317.
30. Liotti M, Mayberg HS, Brannan SK, McGinnis S, JerabekP, Fox PT. Differential limbic–cortical correlates of sad-ness and anxiety in healthy subjects: implications foraffective disorders. Biol Psychiatry 2000; 48: 30–42.
31. Damasio AR, Grabowski TJ, Bechara A et al. Subcorticaland cortical brain activity during the feeling of self-generated emotions. Nat Neurosci 2000; 3: 1049–1056.
32. Habel U, Klein M, Kellermann T, Shah NJ, SchneiderF. Same or different? Neural correlates of happy andsad mood in healthy males. Neuroimage 2005; 26: 206–214.
33. Turner BM, Paradiso S, Marvel CL et al. The cerebellumand emotional experience. Neuropsychologia 2007; 45:1331–1341.
34. Kelly RM, Strick PL. Cerebellar loops with motor cortexand prefrontal cortex of a nonhuman primate. J Neurosci2003; 23: 8432–8444.
35. Middleton FA, Strick PL. Cerebellar projections to theprefrontal cortex of the primate. J Neurosci 2001; 21: 700–712.
36. Heath RG, Dempesy CW, Fontana CJ, Myers WA.Cerebellar stimulation: effects on septal region, hippocam-pus, and amygdala of cats and rats. Biol Psychiatry 1978;13: 501–529.
37. Heath RG, Dempesy CW, Fontana CJ, Fitzjarrell AT.Feedback loop between cerebellum and septal-hippocam-pal sites: its role in emotion and epilepsy. Biol Psychiatry1980; 15: 541–556.
38. Dietrichs E, Haines DE, Roste GK, Roste LS. Hypotha-lamocerebellar and cerebellohypothalamic projections –circuits for regulating nonsomatic cerebellar activity?Histol Histopathol 1994; 9: 603–614.
39. Voogd J, Glickstein M. The anatomy of the cerebellum.Trends Neurosci 1998; 21: 370–375.
40. Snider SR, Snider RS. Kainic acid: enduring alterations incerebellar morphology and in cerebral catecholamine andGABA concentrations after cerebellar injection in the rat.Neurosci Lett 1979; 12: 339–342.
41. Ivry RB, Keele SW. Timing of functions of the cerebellum.J Cogn Neurosci 1989; 1: 136–152.
42. Ivry RB, Keele SW, Diener HC. Dissociation of thelateral and medial cerebellum in movement timingand movement execution. Exp Brain Res 1988; 73: 167–180.
43. Steinmetz JE. Brain substrates of classical eyeblink condi-tioning: a highly localized but also distributed system.Behav Brain Res 2000; 110: 13–24.
44. Woodruff-Pak DS, Papka M, Ivry RB. Cerebellar involve-ment in eyeblink classical conditioning in humans. Neuro-psychology 1996; 10: 443–458.
45. Woodruff-Pak DS, Goldenberg G, Downey-Lamb MM,Boyko OB, Lemieux SK. Cerebellar volume in humansrelated to magnitude of classical conditioning. Neurore-port 2000; 11: 609–615.
46. Edwards CR, Newman S, Bismark A et al. Cerebellumvolume and eyeblink conditioning in schizophrenia. Psy-chiatry Res 2007; 162: 185–194.
47. Strakowski SM, Delbello MP, Adler CM. The functionalneuroanatomy of bipolar disorder: a review of neuroim-aging findings. Mol Psychiatry 2005; 10: 105–116.
48. Frazier JA, Ahn MS, DeJong S, Bent EK, Breeze JL,Giuliano AJ. Magnetic resonance imaging studies in early-onset bipolar disorder: a critical review. Harv Rev Psychi-atry 2005; 13: 125–140.
49. Haldane M, Frangou S. New insights help define thepathophysiology of bipolar affective disorder: neuroimag-ing and neuropathology findings. Prog Neuropsychophar-macol Biol Psychiatry 2004; 28: 943–960.
50. Strakowski SM, DelBello MP, Adler C, Cecil DM, SaxKW. Neuroimaging in bipolar disorder. Bipolar Disord2000; 2: 148–164.
51. Takehara-Nishiuchi K, Kawahara S, Kirino Y. NMDAreceptor-dependent processes in the medial prefrontalcortex are important for acquisition and the early stage
Eyeblink conditioning in bipolar disorder
31

of consolidation during trace, but not delay eyeblinkconditioning. Learn Mem 2005; 12: 606–614.
52. McLaughlin J, Skaggs H, Churchwell J, Powell DA.Medial prefrontal cortex and pavlovian conditioning: traceversus delay conditioning. Behav Neurosci 2002; 116: 37–47.
53. Oswald B, Knuckley B, Mahan K, Sanders C, Powell DA.Prefrontal control of trace versus delay eyeblink condi-tioning: role of the unconditioned stimulus in rabbits(Oryctolagus cuniculus). Behav Neurosci 2006; 120: 1033–1042.
54. Powell DA, Skaggs H, Churchwell J, McLaughlin J.Posttraining lesions of the medial prefrontal cortex impairperformance of Pavlovian eyeblink conditioning but haveno effect on concomitant heart rate changes in rabbits(Oryctolagus cuniculus). Behav Neurosci 2001; 115: 1029–1038.
55. Perrett SP, Ruiz BP, Mauk MD. Cerebellar cortex lesionsdisrupt learning-dependent timing of conditioned eyelidresponses. J Neurosci 1993; 13: 1708–1718.
56. Young RC, Biggs JT, Ziegler VE, Meyer DA. A ratingscale for mania: reliability, validity and sensitivity. Br JPsychiatry 1978; 133: 429–435.
57. Montgomery SA, Asberg M. A new depression scaledesigned to be sensitive to change. Br J Psychiatry 1979;134: 382–389.
58. Brown SM, Kieffaber PD, Carroll CA et al. Eyeblinkconditioning deficits indicate timing and cerebellarabnormalities in schizophrenia. Brain Cogn 2005; 58: 94–108.
59. Lang PJ, Greenwald MK. The International AffectivePicture System Standardization Procedure and InitialGroup Results for Affective Judgments: Technical Report1A. Gainesville, FL: The Center for Research in Psycho-physiology, University of Florida, 1988.
60. King DAT, Tracy J. DataMunch: A Matlab m-FileCollection for the Analysis of Trial-Based Spike andBehavioral Data. http://www.novl.indiana.edu/~dmunch/.1999.
61. Cohen J. Eta-squared and partial eta-squared in commu-nication science. Hum Commun Res 1973; 28: 473–490.
62. Greer TL, Trivedi MH, Thompson LT. Impaired delayand trace eyeblink conditioning performance in majordepressive disorder. J Affect Disord 2005; 86: 235–245.
63. Yeo CH, Hardiman MJ, Glickstein M. Classical condi-tioning of the nictitating membrane response of the rabbit.
II. Lesions of the cerebellar cortex. Exp Brain Res 1985; 60:99–113.
64. Steinmetz JE, LavondDG, IvkovichD, LoganCG, Thomp-son RF. Disruption of classical eyelid conditioning aftercerebellar lesions: damage to a memory trace system ora simple performance deficit? J Neurosci 1992; 12: 4403–4426.
65. Lavond DG. Role of the nuclei in eyeblink conditioning.Ann N Y Acad Sci 2002; 978: 93–105.
66. Garcia KS, Mauk MD. Pharmacological analysis ofcerebellar contributions to the timing and expression ofconditioned eyelid responses. Neuropharmacology 1998;37: 471–480.
67. Steinmetz JE, Kim J, Thompson RF. Biological models ofassociative learning. In: Gallagher M, Nelson R eds.Handbook of Psychology, Volume 3. Biological Psychol-ogy. New York: Wiley, 2002: 499–541.
68. Garcia KS, Steele PM, Mauk MD. Cerebellar cortexlesions prevent acquisition of conditioned eyelid responses.J Neurosci 1999; 19: 10940–10947.
69. Gerwig M, Hajjar K, Dimitrova A et al. Timing ofconditioned eyeblink responses is impaired in cerebellarpatients. J Neurosci 2005; 25: 3919–3931.
70. Chapman PF, Steinmetz JE, Sears LL, Thompson RF.Effects of lidocaine injection in the interpositus nucleus andred nucleus on conditioned behavioral and neuronalresponses. Brain Res 1990; 537: 149–156.
71. Fatemi SH, Stary JM, Earle JA, Araghi-Niknam M, EaganE. GABAergic dysfunction in schizophrenia and mooddisorders as reflected by decreased levels of glutamic aciddecarboxylase 65 and 67 kDa and Reelin proteins incerebellum. Schizophr Res 2005; 72: 109–122.
72. Guidotti A, Auta J, Davis JM et al. Decrease in reelin andglutamic acid decarboxylase67 (GAD67) expression inschizophrenia and bipolar disorder: a postmortem brainstudy. Arch Gen Psychiatry 2000; 57: 1061–1069.
73. Wolpert EA, Goldberg JF, Harrow M. Rapid cycling inunipolar and bipolar affective disorders. Am J Psychiatry1990; 147: 725–728.
74. Kupka RW, Luckenbaugh DA, Post RM et al. Compar-ison of rapid-cycling and non-rapid-cycling bipolar disor-der based on prospective mood ratings in 539 outpatients.Am J Psychiatry 2005; 162: 1273–1280.
75. Keller MB, Lavori PW, Coryell W, Endicott J, Mueller TI.Bipolar I: a five-year prospective follow-up. J Nerv MentDis 1993; 181: 238–245.
Bolbecker et al.
32




















