
Diversity anomalies and spatial climate heterogeneity
12
RESEARCH PAPER Diversity anomalies and spatial climate heterogeneity Iván Jiménez 1 * and Robert E. Ricklefs 2 1 Center for Conservation and Sustainable Development, Missouri Botanical Garden, PO 17 Box 299, St Louis, MO 63166-0299, USA, 2 Department of Biology, University of Missouri at St. Louis, One University Boulevard, St Louis, MO 63121-4499, USA ABSTRACT Aim Diversity anomalies are differences in species richness between areas that belong to different regions but have similar environments. Some hypotheses addressing the origin of well-known anomalies in plant diversity propose that regions with higher environmental spatial heterogeneity have higher diversity because heterogeneity fosters diversification or coexistence. Arguments supporting these hypotheses emphasize inter-regional comparisons of diversity and assume that spatial environmental heterogeneity is higher in: (1) eastern Asia (EA) than in eastern North America (ENA), (2) western North America (WNA) than in ENA, and (3) the Cape Floristic Region in southern Africa (CFR) than in the Southwest Australian Floristic Region (SWA). Here, we evaluate these assumptions by meas- uring two kinds of environmental heterogeneity – spatially implicit and explicit – each thought to affect diversity via different mechanisms. The former refers to environmental variation among sites within a region, regardless of site location. The latter refers to the spatial pattern of environmental variation across a region (e.g., monotonic or undulating). Location EA, ENA, WNA, CFR and SWA. Methods Multivariate and univariate analyses of spatially implicit and explicit heterogeneity in 17 climatic variables describing central tendency, variation and extremes of temperature and precipitation. Results Multivariate (spatially implicit and explicit) climate heterogeneity is higher in: (1) EA than in ENA, (2) WNA than in ENA, and (3) CFR than in SWA. However, univariate analysis revealed that the regions thought to be most homo- geneous (ENA and SWA) were actually most heterogeneous in three or four cli- matic variables, including precipitation during the driest (ENA) or wettest (SWA) seasons. Main conclusions The overall inter-regional pattern of spatially implicit and explicit heterogeneity in climate supports the three assumptions listed in the Aim. However, particular climate variables deviate from this overall pattern, implying that hypotheses linking diversity to regional heterogeneity can yield more precise predictions, and thus can be more stringently tested, than previously recognized. Keywords Climate, diversity anomalies, environmental heterogeneity, regional effects, spatial heterogeneity, spatially explicit, spatially implicit. *Correspondence: Iván Jiménez, Center for Conservation and Sustainable Development, Missouri Botanical Garden, PO 17 Box 299, St Louis, MO 63166-0299, USA. E-mail: [email protected] INTRODUCTION Explaining the broad-scale geographic variation in species rich- ness has presented a persistent challenge to ecologists and bio- geographers (Ricklefs, 2004; Mittelbach et al., 2007). Insights have come from attempts to understand the origin of relation- ships between species richness and the physical environment (Currie et al., 2004; Ricklefs, 2006), as well as regional deviations from those relationships known as ‘diversity anomalies’ (Ricklefs, 2004). Examples of diversity anomalies are well Global Ecology and Biogeography, (Global Ecol. Biogeogr.) (2014) © 2014 John Wiley & Sons Ltd DOI: 10.1111/geb.12181 http://wileyonlinelibrary.com/journal/geb 1
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Diversity anomalies and spatial climate heterogeneity

RESEARCHPAPER
Diversity anomalies and spatial climateheterogeneityIván Jiménez1* and Robert E. Ricklefs2
1Center for Conservation and Sustainable
Development, Missouri Botanical Garden, PO
17 Box 299, St Louis, MO 63166-0299, USA,2Department of Biology, University of Missouri
at St. Louis, One University Boulevard, St
Louis, MO 63121-4499, USA
ABSTRACT
Aim Diversity anomalies are differences in species richness between areas thatbelong to different regions but have similar environments. Some hypothesesaddressing the origin of well-known anomalies in plant diversity propose thatregions with higher environmental spatial heterogeneity have higher diversitybecause heterogeneity fosters diversification or coexistence. Arguments supportingthese hypotheses emphasize inter-regional comparisons of diversity and assumethat spatial environmental heterogeneity is higher in: (1) eastern Asia (EA) than ineastern North America (ENA), (2) western North America (WNA) than in ENA,and (3) the Cape Floristic Region in southern Africa (CFR) than in the SouthwestAustralian Floristic Region (SWA). Here, we evaluate these assumptions by meas-uring two kinds of environmental heterogeneity – spatially implicit and explicit –each thought to affect diversity via different mechanisms. The former refers toenvironmental variation among sites within a region, regardless of site location.The latter refers to the spatial pattern of environmental variation across a region(e.g., monotonic or undulating).
Location EA, ENA, WNA, CFR and SWA.
Methods Multivariate and univariate analyses of spatially implicit and explicitheterogeneity in 17 climatic variables describing central tendency, variation andextremes of temperature and precipitation.
Results Multivariate (spatially implicit and explicit) climate heterogeneity ishigher in: (1) EA than in ENA, (2) WNA than in ENA, and (3) CFR than in SWA.However, univariate analysis revealed that the regions thought to be most homo-geneous (ENA and SWA) were actually most heterogeneous in three or four cli-matic variables, including precipitation during the driest (ENA) or wettest (SWA)seasons.
Main conclusions The overall inter-regional pattern of spatially implicit andexplicit heterogeneity in climate supports the three assumptions listed in the Aim.However, particular climate variables deviate from this overall pattern, implyingthat hypotheses linking diversity to regional heterogeneity can yield more precisepredictions, and thus can be more stringently tested, than previously recognized.
KeywordsClimate, diversity anomalies, environmental heterogeneity, regional effects,spatial heterogeneity, spatially explicit, spatially implicit.
*Correspondence: Iván Jiménez, Center forConservation and Sustainable Development,Missouri Botanical Garden, PO 17 Box 299, StLouis, MO 63166-0299, USA.E-mail: [email protected]
INTRODUCTION
Explaining the broad-scale geographic variation in species rich-
ness has presented a persistent challenge to ecologists and bio-
geographers (Ricklefs, 2004; Mittelbach et al., 2007). Insights
have come from attempts to understand the origin of relation-
ships between species richness and the physical environment
(Currie et al., 2004; Ricklefs, 2006), as well as regional deviations
from those relationships known as ‘diversity anomalies’
(Ricklefs, 2004). Examples of diversity anomalies are well
bs_bs_banner
Global Ecology and Biogeography, (Global Ecol. Biogeogr.) (2014)
© 2014 John Wiley & Sons Ltd DOI: 10.1111/geb.12181http://wileyonlinelibrary.com/journal/geb 1

known (Schluter & Ricklefs, 1993; Qian & Ricklefs, 2000;
Ricklefs, 2004; Kreft & Jetz, 2007) but their causes remain elusive
(Harrison & Cornell, 2008). Hypotheses invoking ‘regional
effects’ suggest that differences among regions in geographic
configuration and history result in regional differences in rates
of speciation, immigration and extinction that underlie diver-
sity anomalies (Schluter & Ricklefs, 1993; Ricklefs, 2004). Here
we focus on hypotheses about how regional differences in the
geographic configuration of environmental variation may
underlie diversity anomalies. These hypotheses rely on ‘back-
ground assumptions’ (sensu Turner, 2005) about regional differ-
ences in one of two distinct aspects of environmental spatial
heterogeneity, each thought to affect diversity via different
mechanisms.
One such aspect of environmental spatial heterogeneity is
variation among different sampling points within a region,
regardless of the location of those points within the region. This
aspect of environmental spatial heterogeneity is referred to as
‘spatially implicit heterogeneity’ (Wiens, 2000) and can be
operationally defined as the variance in environmental condi-
tions across a region (Fig. 1). Spatially implicit heterogeneity has
been hypothesized to determine the number of species that may
co-occur within a region through ecological sorting (i.e. the
differential success of species in contrasting environments;
Chesson, 2000). It is also thought to partly determine the range
of environmental conditions potentially experienced by a
species within a region (Fig. 1) and, therefore, the selective pres-
sures experienced by different populations across a region and
the potential contribution of ecological speciation to regional
species richness (Schluter, 2000).
The second aspect of environmental spatial heterogeneity is
the geographic distribution of sample points with different
characteristics across a region, referred to as ‘spatially explicit
heterogeneity’ (Wiens, 2000). This is independent of the abso-
lute variation among localities, and can be operationally defined
as the proportion of variance in environmental variables across
a region due to different kinds of spatial structure (sensu
Legendre & Legendre, 2012; Fig. 1). In linear or other monot-
0 5 10 15 20
Spatial dimension
−1.
0−
0.5
0.0
0.5
1.0
Env
ironm
enta
l var
iabl
eS
uita
ble
habi
tat
B
A
0 5 10 15 20
Spatial dimension
−1.
0−
0.5
0.0
0.5
1.0
B
A
0 5 10 15 20
Spatial dimension
−1.
0−
0.5
0.0
0.5
1.0
Env
ironm
enta
l var
iabl
eS
uita
ble
habi
tat
B
A
0 5 10 15 20
Spatial dimension
−1.
0−
0.5
0.0
0.5
1.0
B
A
a b
c d
Figure 1 Spatially implicit environmental heterogeneity (i.e. variation among locations), spatially explicit environmental heterogeneity (i.e.arrangement of locations with different characteristics) and the distribution of species suitable habitat. Each panel shows environmentalvariation along a spatial dimension of an idealized region. Regions (a) and (b) are equal in spatially implicit variation (standarddeviation = 0.71) but differ in spatially explicit variation: a sinusoidal spatial structure dominates (a) and a monotonic structure dominates(b). Regions illustrated in (c) and (d) have lower spatially implicit variation (standard deviation = 0.28) than regions in (a) and (b).Nonetheless, spatially explicit variation does not differ between (a) and (c) nor between (b) and (d). The limits of the niche of species Aand B are shown as black and gray dotted lines, respectively. The niche is defined as the set of environmental conditions in which intrinsicgrowth rate is non-negative. The distribution of suitable habitat for species A and B is shown at the bottom of each panel as black and greylines, respectively. Suitable habitat for a species is defined as the set of areas having environments within the niche of the species.
I. Jiménez and R. E. Ricklefs
Global Ecology and Biogeography, © 2014 John Wiley & Sons Ltd2

onic spatial structures, patches of suitable habitat for particular
species are less isolated than expected by chance (Fig. 1 and
Appendix S1 and Fig. S1 in Supporting Information). In con-
trast, in sinusoidal (undulating) spatial structures patches of
suitable habitat for particular species can be more isolated than
expected by chance (Figs. 1 & S1, Appendix S1). These patterns
of isolation of suitable habitat are thought to determine how
regional species richness is influenced by metacommunity
dynamics (Holyoak et al., 2005) and opportunities for allopatric
speciation (Allmon, 1992; Barraclough, 2006). Based on the
effects of different spatial structures on the isolation of suitable
habitat (Appendix S1, Fig. S1), we consider spatially explicit
heterogeneity to be negatively related to the proportion of envi-
ronmental variance explained by linear or other monotonic
structures, and positively related to the proportion of environ-
mental variance explained by sinusoidal spatial structures
(Fig. 1).
Hypotheses about how regional differences in geographic
configuration may cause diversity anomalies can be scrutinized
by analysing their background assumptions, by separately quan-
tifying regional differences in spatially implicit and explicit het-
erogeneity. To the best of our knowledge, such comparisons have
not been made before. Previous regional comparisons of spatial
heterogeneity as a potential cause of diversity anomalies have
contrasted climate and vegetation maps for different regions
with little input from formal quantitative analyses of environ-
mental spatial variation (e.g. Hobbs et al., 1995; Cowling et al.,
1996; Qian & Ricklefs, 2000), and they have not distinguished
spatially implicit and explicit heterogeneity. Here, we evaluate
the background assumptions of hypotheses proposing mecha-
nisms by which differences in geographic configuration may
have caused well-known diversity anomalies. In particular, we
empirically examine the assumptions that spatially implicit and
explicit environmental heterogeneity are higher in: (1) moist
environments in eastern Asia (EA) than in eastern North
America (ENA), (2) western North America (WNA) than in
eastern North America (ENA), and (3) the Cape Floristic Region
(CFR) in southern Africa than in the Southwest Australian Flo-
ristic Region (SWA).
METHODS
Study system
We focused on salient examples of diversity anomalies. One is
the diversity anomaly between temperate disjunct floras of EA
and ENA (Wen, 1999). Disjunct plant genera occupy similar
moist environments in EA and ENA (Qian & Ricklefs, 2004) and
have, on average, twice as many species in EA as in ENA (Qian &
Ricklefs, 1999). Extinctions during the Neogene period of
cooling climate and glaciation are thought to explain the low
diversity in European moist temperate environments relative to
EA and ENA (Svenning, 2003), but not the contrast between EA
and ENA (Latham & Ricklefs, 1993). Detailed phylogenetic
studies of 10 plant genera indicate that differences in species
numbers are unlikely to be explained by the time available for
diversification on each continent (Xiang et al., 2004). Rather,
diversification rates (the net outcome of speciation and extinc-
tion rates) of clades restricted to EA are thought to be higher
than those of clades restricted to ENA (Qian & Ricklefs, 2000;
Xiang et al., 2004). A hypothesis advanced to explain these pre-
sumed differences in diversification rates suggests that global
temporal variation in climate and sea level during the late Ter-
tiary would have resulted in more opportunities for allopatric
speciation (Qian & Ricklefs, 2000), and also less extinction
(Xiang et al., 2004), in EA than in ENA due to higher physi-
ographic heterogeneity and geographic complexity in EA.
Furthermore, disjunct EA–North American plant genera that
are restricted to ENA within North America tend to have fewer
species than those extending their ranges into WNA, again, pur-
portedly due to differences in diversification rates caused by
regional differences in physiographic heterogeneity between
ENA and WNA (Qian & Ricklefs, 2000).
Another diversity anomaly involves comparisons between the
CFR (Born et al., 2007) and the SWA (Hopper & Gioia, 2004),
both with similar Mediterranean climates (Milewski, 1979) and
well known for their high plant diversity and endemism
(Cowling et al., 1996; Crisp et al., 2001; Goldblatt & Manning,
2002; Linder, 2003) despite reputedly little spatial environmen-
tal heterogeneity (Rosenzweig, 1995). High diversity in CFR and
SWA relative to other Mediterranean-climate regions has been
explained as the result of a combination of key features promot-
ing diversification, including high fire frequency, nutrient-poor
soils and a relatively mild Quaternary climate (Cowling et al.,
1996). In spite of these similarities, CFR has higher species rich-
ness than comparable environments in SWA (Cowling et al.,
1996), largely due to parallel differences in species richness in
several clades (e.g. Restionaceae, Linder et al., 2003; Proteaceae,
Sauquet et al., 2009). This diversity anomaly has been hypoth-
esized to result partly from relatively high physiographic hetero-
geneity in CFR, which might have increased the opportunities
for geographic isolation and ecological speciation and reduced
extinction risk across the region (Goldblatt & Manning, 2002;
Linder, 2003; Linder, 2005; Cowling et al., 2009; van der Niet &
Johnson, 2009; Verboom et al., 2009), perhaps conditional on
rainfall regimes (Cowling & Lombard, 2002; Forest et al., 2007).
Beyond spatial heterogeneity, relatively high fire frequency and
climatic stability during the Pleistocene may have driven high
plant diversity in CFR, in addition to intrinsic factors such as
short dispersal distances and specialized pollination relation-
ships (Cowling & Lombard, 2002; Goldblatt & Manning, 2002;
Linder, 2003, 2005; Barraclough, 2006).
Sampling regional environments
We defined limits of regions of disjunct floras of EA, ENA and
WNA based on the distribution of distinct assemblages of
species (Qian & Ricklefs, 2000; Fig. 2a–c), and those of CFR and
SWA following previous floristic work (Hopper & Gioia, 2004;
Born et al., 2007; Fig. 2e–h). We described environmental het-
erogeneity in terms of 17 climate variables thought to influence
distribution of suitable habitat for plant species. Specifically, we
Diversity anomalies and climate heterogeneity
Global Ecology and Biogeography, © 2014 John Wiley & Sons Ltd 3

used data at a resolution of 30 arcsec available from WorldClim
(Hijmans et al., 2005; http://www.worldclim.org/) on nine tem-
perature and eight precipitation variables describing the central
tendency, variation and extremes of climatic conditions (Fig. 2,
Tables S1 & S2).
We obtained systematic samples of WorldClim maps of each
region by arranging sample points in regular grids of
50 km × 50 km across EA, ENA and WNA, and in regular grids
of 10 km × 10 km across the smaller areas of CFR and SWA.
Although denser grids would be desirable to capture variation at
smaller distances, we used the densest grids that did not exceed
computational limits for estimating spatially explicit measures
of heterogeneity. We relied on systematic sampling because it is
thought to outperform random sampling in terms of statistical
efficiency under realistic assumptions (Haining, 2003). None-
theless, we also conducted analyses of samples obtained by ran-
domly locating sample points across WorldClim maps of each
region. These latter analyses yielded the same conclusions as
those based on systematic samples, so we do not discuss them
further.
Testing regional differences in spatially implicit andexplicit heterogeneity
We first compared regions in terms of spatially implicit climatic
heterogeneity, measured as variance in climate across a region
(Fig. 1). In multivariate space we measured variance as the
determinant of the variance–covariance matrix for the 17
climate variables (i.e. the dispersion matrix), and tested regional
differences using Bartlett’s modified likelihood ratio test statis-
tic, Bm, and critical values based on a bootstrap procedure
appropriate for distributions that depart from multivariate nor-
mality (Zhang & Boos, 1992; Goodnight & Schwartz, 1997;
Fig. 3a). This procedure might potentially perform poorly
Figure 2 Equal-area projection maps ofclimate variables across eastern Asia (EA)(a, c), western North America (WNA)and eastern North Aamerica (ENA) (b,d), the Cape Floristic Region (CFR) (e,g), and south-west Australia (SWA) (f, h).The limits of these regions, shown asthick black lines, are based on thedistribution of distinct assemblages ofspecies (Qian & Ricklefs, 2000; Hopper &Gioia, 2004; Born et al., 2007). Annualmean temperature (a, b, e, f),precipitation of the driest month (c, d),and precipitation of the wettest month(g, h). Panels in the same row share asingle scale, shown in the left panel ofeach row.
I. Jiménez and R. E. Ricklefs
Global Ecology and Biogeography, © 2014 John Wiley & Sons Ltd4

because bootstrap samples do not attempt to preserve spatial
dependence in the data (Davison & Hinkley, 1997), so we used
an additional bootstrap approach that preserved spatial struc-
ture in the data (details in Appendix S2).
Next we compared regions in terms of spatially explicit het-
erogeneity in multivariate climate, measured as the proportion
of variance due to two kinds of spatial structure: linear and
sinusoidal (Fig. 1). We used redundancy analysis (RDA;
Legendre & Legendre, 2012) to regress multivariate climate data
on two kinds of explanatory variables: geographic coordinates
and spatial eigenvector maps (Borcard & Legendre, 2002; Dray
et al., 2006; Fig. 3b). Geographic coordinates accounted for
linear spatial structures in climate, while spatial eigenvector
maps accounted for sinusoidal spatial structures. We obtained
spatial eigenvector maps from a truncated matrix of geographic
distances among sampling sites, with a truncation distance equal
to the maximum distance of the minimum spanning tree across
sampling sites. Higher truncation distances are thought to
decrease the power to detect spatial structures at small scales
(Borcard & Legendre, 2002). When derived from regularly
spaced localities, as in our systematic samples of WorldClim
maps, spatial eigenvector maps are sinusoidal variables whose
wavelengths are associated with the respective eigenvalues
(Borcard et al., 2011; Figs S2 & S3). Because positive spatial
autocorrelation at short distances predominates in climatic vari-
ables, the RDA included only spatial eigenvector maps associ-
ated with positive eigenvalues, which represent positive spatial
autocorrelation at short distances (Dray et al., 2006).
We quantified the proportion of variation in climate due to
linear and sinusoidal spatial structures with the ‘adjusted redun-
dancy statistic’ (Peres-Neto et al., 2006; Fig. 3b) for geographic
coordinates and spatial eigenvector maps, respectively. We
examined regional differences in these proportions using a test
based on bootstrap samples constructed by resampling residuals
of RDA models (Peres-Neto et al., 2006). We did not perform
any forward or backward selection of spatial eigenvector maps,
because inclusion of null predictors in RDA models does not
bias the adjusted redundancy statistic (Peres-Neto et al., 2006).
Spatial dependence in residuals of RDA models, however, might
result in bootstrap samples that do not preserve spatial depend-
ence in the data and, consequently, the test might perform
poorly (Davison & Hinkley, 1997). To determine whether this
was a concern, we tested for spatial dependence in residuals of
RDA models using multivariate Mantel correlograms (Legendre
& Legendre, 2012).
The analyses above addressed regional differences in implicit
and explicit measures of multivariate climate heterogeneity that
might arise from proportional differences in all terms (variances
and covariances) of the dispersion matrices or from non-
proportional differences known as differences in matrix shape.
We examined regional differences in the shape of dispersion
matrices using a bootstrap procedure to test the null hypothesis
that both matrices have the same shape (Goodnight & Schwartz,
1997; Fig. 3c). The null hypothesis in this test is that the Mantel
correlation coefficient, rM, equals 1. To ensure that our conclu-
sions were unaffected by bootstrap samples that do not preserve
spatial dependence, we used an additional approach that pre-
served spatial structure in the data (Appendix S2).
Whenever we found significant regional differences in the
shape of dispersion matrices, we also examined whether single
climate variables showed patterns that were opposite to the
overall multivariate pattern. In particular, we compared regions
in terms of a spatially implicit estimate of univariate climatic
variance using a two-sample F-test statistic and critical values
based on a bootstrap procedure appropriate for non-normal
Raw data A
VCV matrix
A
Raw data B
VCV matrix
B
Bartlett’s modified
test
VCV matrix
A
VCV matrix
B
Manteltest
Raw data A
Explanatory variables
RDA Radj2 linear (± bootstrap CI)
Radj2 sinusoidal (± bootstrap CI)
rM statistic and P-value for the null hypothesis that the shape of VCV matrices does not differ between regions (i.e., H0: rM = 1).
Bm statistic and P-value for the null hypothesis that (multivariate) variance does not differ between regions
Radj2 linear (± bootstrap CI)
Radj2 sinusoidal (± bootstrap CI)
a) Regional differences in spatially implicit heterogeneity
b) Regional differences in spatially explicit heterogeneity
c) Regional differences in the shape of VCV matrices
Raw data B
Explanatory variables
RDA
Figure 3 Outline of multivariate statistical methods used toexamine regional differences in spatial climate heterogeneity. (a)The raw data for each region (A and B) consist of a table in whicheach row represents a sample point and each column represents aclimate variable. From these raw data the variance–covariancematrix (VCV matrix), also known as the dispersion matrix, iscalculated. Regional differences in spatially implicit heterogeneity,measured as the determinant of the VCV matrix, are examinedwith Bartlett’s modified likelihood ratio test statistic, Bm. (b) Theraw data are the response variables in a redundancy analysis(RDA) with two sets of explanatory variables: geographiccoordinates to account for linear spatial structures and spatialeigenvector maps to account for sinusoidal spatial structures.Spatially explicit heterogeneity is measured as the proportion ofvariation in climate due to linear and sinusoidal spatial structureswith the adjusted redundancy statistic, Radj
2. Regional differencesin these proportions are examined with a bootstrap procedure. (c)Regional differences in the structure of VCV matrices, known asdifferences in matrix shape, are examined using a Mantel test withthe null hypothesis that the Mantel’s correlation coefficient, rM,equals 1.
Diversity anomalies and climate heterogeneity
Global Ecology and Biogeography, © 2014 John Wiley & Sons Ltd 5

distributions (Boos & Brownie, 1989; Fig. 4a). We used multiple
regression to measure the proportion of variation in univariate
climate explained by linear and sinusoidal spatial structures
(Fig. 4b). Similar to multivariate analysis, we regressed the single
climate variable of interest on geographic coordinates and
spatial eigenvector maps. We compared regions in terms of vari-
ation in single climate variables explained by linear and
sinusoidal spatial structures using a test based on bootstrap
samples constructed by resampling residuals of regression
models (Peres-Neto et al., 2006). To ensure that our conclusions
were not affected by a failure to account for spatial dependence,
we used univariate versions of procedures described above for
multivariate tests (Appendix S2). All statistical procedures were
performed in the R environment (R Development Core Team,
2009).
RESULTS
Spatially implicit and explicit measures of multivariate climate
heterogeneity supported the three postulated contrasts between
regions. Spatially implicit measures showed that ENA was less
heterogeneous than EA (Bm = 10,732, P = 0.001; Fig. 5a) and
WNA (BM = 10,673, P = 0.001, Fig. 5a) and that SWA was less
heterogeneous than CFR (Bm = 29293, P = 0.001, Fig. 6a). Tests
preserving spatial structure in the data yielded the same conclu-
sions (Table S3). Spatially explicit measures showed that a higher
proportion of variation in multivariate climate was due to linear
structures in ENA than in EA and WNA (Fig. 5b) and in SWA
than in CFR (Fig. 6b). Conversely, a lower proportion of vari-
ation in multivariate climate was due to sinusoidal structures in
ENA than in EA and WNA (Fig. 5c) and in SWA than in CFR
(Fig. 6c). Statistically significant autocorrelations in residuals of
RDAs were small in all cases, suggesting a negligible impact on
results (Fig. S4).
Regional differences in multivariate climate heterogeneity did
not arise from proportional differences across components
(variances and covariances) of dispersion matrices. The shape of
the dispersion matrix for climate of ENA differed from that of
EA (rM = 0.57, P = 0.001, Fig. 7a,b) and WNA (rM = 0.65,
P = 0.001, Fig. 5c,d). The shape of the dispersion matrix of SWA
also differed from that of CFR (rM = 0.54, P = 0.001, Fig. 7e,f).
Tests preserving spatial structure in the data yielded the same
conclusions (Table S3). Importantly, these results indicate that
empirical support for the three postulated regional contrasts
might vary depending on the set of climatic variables included
in the analysis.
Indeed, regional differences in univariate implicit measures of
heterogeneity deviated from the respective multivariate pattern
(Figs 5a & 6a). Contrary to the three postulated regional con-
trasts, spatially implicit measures showed that ENA was more
heterogeneous than EA in precipitation for the driest month
(F = 1.58, P = 0.001, Fig. 2c,d), the driest quarter (F = 1.38,
P = 0.001) and the coldest quarter (F = 1.75, P = 0.001). ENA
was also more heterogeneous than WNA in precipitation for the
driest month (F = 2.66, P = 0.001, Fig. 2d) and the driest quarter
(F = 2.33, P = 0.001). Furthermore, SWA was more hetero-
geneous than CFR in mean temperature of the warmest quarter
(F = 1.49, P = 0.001), precipitation for the wettest month
(F = 2.67, P = 0.001, Fig. 2g,h), the wettest quarter (F = 2.20,
P = 0.001) and the coldest quarter (F = 2.01, P = 0.001). Tests
preserving spatial structure in the data yielded the same conclu-
sions (Table S3).
Univariate spatially implicit measures that deviated from the
multivariate pattern of regional differences were mirrored by the
respective univariate spatially explicit measures in only some
cases. In particular, the proportion of variation in precipitation
of the driest month, the driest quarter and the coldest quarter
due to sinusoidal structures was larger in ENA than in EA
(Fig. 5c), and thus deviated from the multivariate pattern of
regional differences in the same way as the univariate implicit
measures. Respective differences in the proportion of variation
due to linear structures were less marked (Fig. 5b), but also
tended to mirror univariate implicit measures. In all other cases,
implicit and explicit measures did not show similar deviations
from the multivariate pattern of regional differences. The pro-
portion of variation in precipitation of the driest month and the
Raw data A
Raw data B
F test
Raw data A
Explanatory variables
OLS Radj2 linear (± bootstrap CI)
Radj2 sinusoidal (± bootstrap CI)
F statistic and p-value for the null hypothesis that (univariate) variance does not differ between regions
Radj2 linear (± bootstrap CI)
Radj2 sinusoidal (± bootstrap CI)
a) Regional differences in spatially implicit heterogeneity
b) Regional differences in spatially explicit heterogeneity
Raw data B
Explanatory variables
OLS
Figure 4 Outline of univariate statistical methods used toexamine regional differences in spatial climate heterogeneity.Whenever regional differences in shape of variance–covariancematrices are significant (Fig. 3c), single climate variables areexamined to determine if they deviated from the overallmultivariate pattern. (a) Regional differences in spatially implicitheterogeneity of single climate variables (one shown as a blackrectangle in the raw data table) are examined using the F-test.(b) To measure spatially explicit heterogeneity, a single climatevariable (shown as a black rectangle in the raw data table) is theresponse variable in ordinary least square regression (OLS) withtwo sets of explanatory variables: geographic coordinates toaccount for linear spatial structures and spatial eigenvector mapsto account for sinusoidal spatial structures. The proportion ofvariation in a climate variable due to linear and sinusoidal spatialstructures is measured as the adjusted redundancy statistic, Radj
2.Regional differences in these proportions are then examined witha bootstrap procedure.
I. Jiménez and R. E. Ricklefs
Global Ecology and Biogeography, © 2014 John Wiley & Sons Ltd6
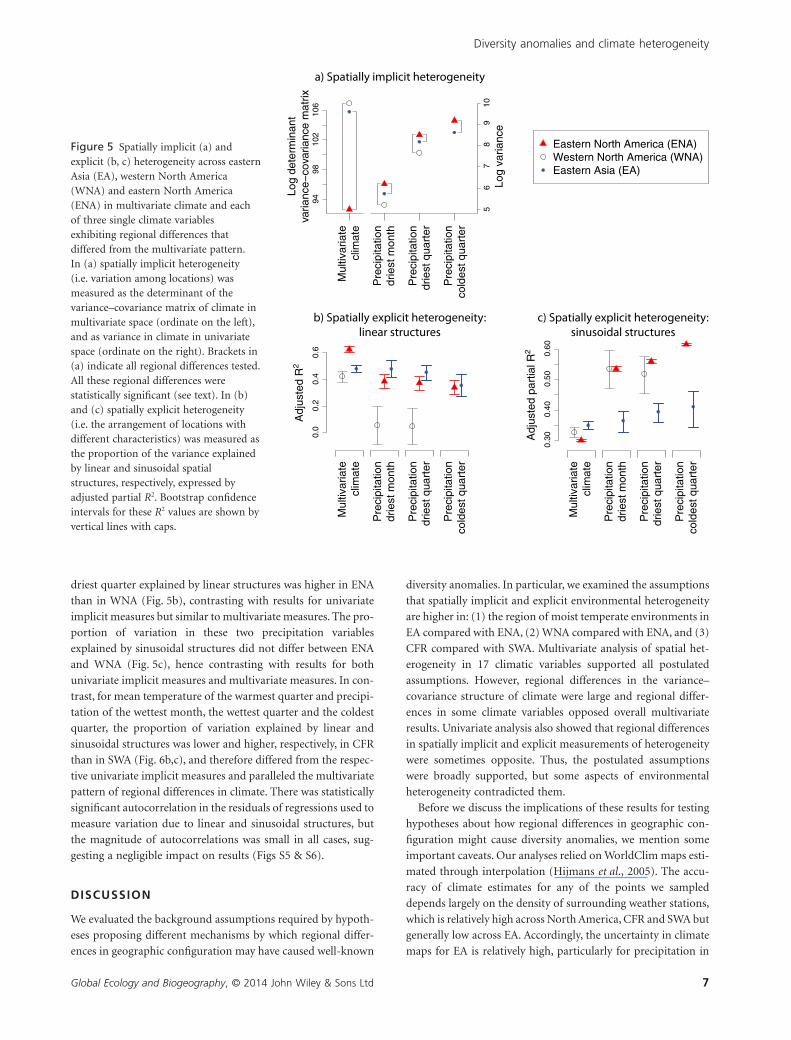
driest quarter explained by linear structures was higher in ENA
than in WNA (Fig. 5b), contrasting with results for univariate
implicit measures but similar to multivariate measures. The pro-
portion of variation in these two precipitation variables
explained by sinusoidal structures did not differ between ENA
and WNA (Fig. 5c), hence contrasting with results for both
univariate implicit measures and multivariate measures. In con-
trast, for mean temperature of the warmest quarter and precipi-
tation of the wettest month, the wettest quarter and the coldest
quarter, the proportion of variation explained by linear and
sinusoidal structures was lower and higher, respectively, in CFR
than in SWA (Fig. 6b,c), and therefore differed from the respec-
tive univariate implicit measures and paralleled the multivariate
pattern of regional differences in climate. There was statistically
significant autocorrelation in the residuals of regressions used to
measure variation due to linear and sinusoidal structures, but
the magnitude of autocorrelations was small in all cases, sug-
gesting a negligible impact on results (Figs S5 & S6).
DISCUSSION
We evaluated the background assumptions required by hypoth-
eses proposing different mechanisms by which regional differ-
ences in geographic configuration may have caused well-known
diversity anomalies. In particular, we examined the assumptions
that spatially implicit and explicit environmental heterogeneity
are higher in: (1) the region of moist temperate environments in
EA compared with ENA, (2) WNA compared with ENA, and (3)
CFR compared with SWA. Multivariate analysis of spatial het-
erogeneity in 17 climatic variables supported all postulated
assumptions. However, regional differences in the variance–
covariance structure of climate were large and regional differ-
ences in some climate variables opposed overall multivariate
results. Univariate analysis also showed that regional differences
in spatially implicit and explicit measurements of heterogeneity
were sometimes opposite. Thus, the postulated assumptions
were broadly supported, but some aspects of environmental
heterogeneity contradicted them.
Before we discuss the implications of these results for testing
hypotheses about how regional differences in geographic con-
figuration might cause diversity anomalies, we mention some
important caveats. Our analyses relied on WorldClim maps esti-
mated through interpolation (Hijmans et al., 2005). The accu-
racy of climate estimates for any of the points we sampled
depends largely on the density of surrounding weather stations,
which is relatively high across North America, CFR and SWA but
generally low across EA. Accordingly, the uncertainty in climate
maps for EA is relatively high, particularly for precipitation in
9498
102
106
Log
dete
rmin
ant
varia
nce−
cova
rianc
e m
atrix
56
78
910
Log
varia
nce
0.0
0.2
0.4
0.6
Adj
uste
d R
2
Mul
tivar
iate
clim
ate
Pre
cipi
tatio
ndr
iest
mon
th
Pre
cipi
tatio
ndr
iest
qua
rter
Pre
cipi
tatio
nco
ldes
t qua
rter
Mul
tivar
iate
clim
ate
Pre
cipi
tatio
ndr
iest
mon
th
Pre
cipi
tatio
ndr
iest
qua
rter
Pre
cipi
tatio
nco
ldes
t qua
rter
Mul
tivar
iate
clim
ate
Pre
cipi
tatio
ndr
iest
mon
th
Pre
cipi
tatio
ndr
iest
qua
rter
Pre
cipi
tatio
nco
ldes
t qua
rter
0.30
0.40
0.50
0.60
Adj
uste
d pa
rtia
l R2
Eastern North America (ENA)Western North America (WNA)Eastern Asia (EA)
a) Spatially implicit heterogeneity
b) Spatially explicit heterogeneity:linear structures
c) Spatially explicit heterogeneity:sinusoidal structures
Figure 5 Spatially implicit (a) andexplicit (b, c) heterogeneity across easternAsia (EA), western North America(WNA) and eastern North America(ENA) in multivariate climate and eachof three single climate variablesexhibiting regional differences thatdiffered from the multivariate pattern.In (a) spatially implicit heterogeneity(i.e. variation among locations) wasmeasured as the determinant of thevariance–covariance matrix of climate inmultivariate space (ordinate on the left),and as variance in climate in univariatespace (ordinate on the right). Brackets in(a) indicate all regional differences tested.All these regional differences werestatistically significant (see text). In (b)and (c) spatially explicit heterogeneity(i.e. the arrangement of locations withdifferent characteristics) was measured asthe proportion of the variance explainedby linear and sinusoidal spatialstructures, respectively, expressed byadjusted partial R2. Bootstrap confidenceintervals for these R2 values are shown byvertical lines with caps.
Diversity anomalies and climate heterogeneity
Global Ecology and Biogeography, © 2014 John Wiley & Sons Ltd 7

mountainous areas with local effects such as rain shadows
(Hijmans et al., 2005). Nonetheless, we believe that WorldClim
maps are accurate enough to portray broad-scale spatial hetero-
geneity across each of the five regions. Further analyses would be
useful when improved climate maps become available. We also
stress that our analyses are restricted to climate and do not
address other dimensions of physiography that may be impor-
tant for explaining diversity anomalies (e.g. soil properties).
The scale along which environment is measured would ideally
be meaningful in terms species niches (Fig. 1), but that may be
difficult to accomplish in practice and results may depend on
scales of measurement. For example, on a linear scale, spatially
implicit heterogeneity in precipitation of the driest month and
quarter was higher in ENA than WNA (Fig. 5a). Precipitation of
the driest month and quarter are generally low in WNA and
variable in ENA (Fig. 2c,d, Table ST1). If variation at low values
of precipitation is ecologically more important than variation at
higher values, then a logarithmic scale might be more meaning-
ful than a linear scale. On a logarithmic scale, ENA was not more
heterogeneous than WNA in precipitation of the driest month
(F = 0.34, P = 1) or the driest quarter (F = 0.31, P = 1). These
results emphasize the difficulty in testing hypotheses about envi-
ronmental heterogeneity as a determinant of diversity anomalies
if there is uncertainty regarding the appropriate scales of envi-
ronmental measurement.
The regional differences in climate we describe here are rel-
evant to hypotheses about how regional differences in geo-
graphic configuration may cause diversity anomalies via
processes that may have operated over relatively recent times.
Specifically, our multivariate results support a background
assumption required of the hypothesis that higher spatially
implicit climatic heterogeneity may have allowed more species
to coexist through ecological sorting (Chesson, 2000) in EA and
WNA than in ENA, and in CFR than in SWA. Under this
hypothesis, higher regional diversity is fostered by higher vari-
ance in the physical attributes of a region. Our results also
support a background assumption needed by a different
hypothesis, focused on metacommunity dynamics (Holyoak
et al., 2005), according to which higher spatially explicit climatic
heterogeneity would allow more species to coexist in EA and
WNA than in ENA, and in CFR than in SWA. Under the latter
hypothesis, isolation of patches of similar habitat fosters coex-
istence by moderating the impact of species interactions.
Whether our measurements of regional differences in spatial
heterogeneity are relevant to hypotheses emphasizing processes,
such as allopatric speciation, operating through geological
a) Spatially implicit heterogeneity
b) Spatially explicit heterogeneity:linear structures
c) Spatially explicit heterogeneity:sinusoidal structures
5660
6468
Log
dete
rmin
ant
varia
nce−
cova
rianc
e m
atrix
56
78
910
Log
varia
nce
Mul
tivar
iate
clim
ate
Mea
n te
mp.
war
mes
t qua
rter
Pre
cipi
tatio
nw
ette
st m
onth
Pre
cipi
tatio
nw
ette
st q
uart
er
Pre
cipi
tatio
nco
ldes
t qua
rter
Mul
tivar
iate
clim
ate
Mea
n te
mp.
war
mes
t qua
rter
Pre
cipi
tatio
nw
ette
st m
onth
Pre
cipi
tatio
nw
ette
st q
uart
er
Pre
cipi
tatio
nco
ldes
t qua
rter
Mul
tivar
iate
clim
ate
Mea
n te
mp.
war
mes
t qua
rter
Pre
cipi
tatio
nw
ette
st m
onth
Pre
cipi
tatio
nw
ette
st q
uart
er
Pre
cipi
tatio
nco
ldes
t qua
rter
0.0
0.2
0.4
0.6
0.8
1.0
Adj
uste
d R
2
0.0
0.2
0.4
0.6
0.8
Adj
uste
d pa
rtia
l R2
Southwest Australia (SWA)Cape Floristic Region (CFR)
Figure 6 Spatially implicit (a) andexplicit (b, c) heterogeneity across theCape Floristic Region (CFR) andsouth-west Australia (SWA) inmultivariate climate and each of foursingle climate variables exhibitingregional differences that differed fromthe multivariate pattern. In (a) spatiallyimplicit heterogeneity (i.e. variationamong locations) was measured as thedeterminant of the variance–covariancematrix in multivariate space (ordinate onthe left) and as variance in univariatespace (ordinate on the right). Brackets in(a) indicate all regional differences tested.All these regional differences werestatistically significant (see text). In (b)and (c) spatially explicit heterogeneity(i.e. the arrangement of locations withdifferent characteristics) was measured asthe proportion of the variance explainedby linear and sinusoidal spatialstructures, respectively, expressed byadjusted partial R2. Bootstrap confidenceintervals for these R2 values are shown byvertical lines with caps.
I. Jiménez and R. E. Ricklefs
Global Ecology and Biogeography, © 2014 John Wiley & Sons Ltd8

epochs depends on how the physiography of each region has
changed through time. An approximate reconstruction of the
physiographic heterogeneity of CFR since the Oligocene sug-
gests substantial changes before the end of the Pliocene
(Cowling et al., 2009). In contrast, the physiography of SWA is
thought to have been more stable (Hopper & Gioia, 2004;
Cowling et al., 2009). We note that physiographic change
through geological time need not imply that present-day relative
differences in spatial heterogeneity between regions do not
reflect past relative differences. For example, despite substantial
temporal change, relative differences in physiographic heteroge-
neity between the western and eastern portions of CFR are
estimated to have persisted since at least the late Oligocene
(Cowling et al., 2009). Nonetheless, reconstructions of regional
differences in physiography through geological time might be
hampered by ‘information-destroying’ processes linking the past
and present, so that competing historical hypotheses might be
more likely to be empirically equivalent than non-historical
hypotheses (Turner, 2005).
To the extent that current regional differences in spatial het-
erogeneity reflect past regional differences, our results would be
relevant to hypotheses focused on the impacts of speciation on
regional diversity. Our multivariate results would support a
background assumption needed by the hypothesis that higher
spatially implicit climatic heterogeneity fostered ecological spe-
ciation leading to higher regional diversity in EA and WNA than
in ENA, and in CFR than in SWA. This hypothesis is based on
the notion that variation in the physical attributes of a region
partly determines the number of peaks in the fitness function of
phenotypes of closely related species (Schluter, 2000). Our mul-
tivariate results would also support a background assumption
needed by the hypothesis that higher spatially explicit environ-
−1.
5−
0.5
0.5
1.0
1.5
Temperature
Precipitation
−2.
0−
1.0
0.0
1.0
2.0
−2
−1
01
23
Sta
ndar
dize
d va
rianc
e-co
vara
ince
a) Eastern North America b) Eastern Asia
c) Eastern North America d) Western North America
e) Southwest Australia f) Cape Floristic Region
Figure 7 Regional variance–covariancematrices for 17 climate variables. Eachrow of panels compares matricesstandardized to a single scale (shown inthe left of each row), followingGoodnight & Schwartz (1997; AppendixS3). The longest diagonal of each matrixshows the variances of nine temperaturevariables followed by the variances ofeight precipitation variables. Shorterdiagonals show the respectivecovariances. From bottom to top, climaticvariables along the longest diagonal are:annual mean temperature, mean diurnalrange, temperature seasonality (standarddeviation), maximum temperature of thewarmest month, minimum temperatureof the coldest month, mean temperatureof the wettest quarter, mean temperatureof the driest quarter, mean temperatureof the warmest quarter, meantemperature of the coldest quarter,annual precipitation, precipitation of thewettest month, precipitation of the driestmonth, precipitation seasonality(coefficient of variation), precipitation ofthe wettest quarter, precipitation of thedriest quarter, precipitation of thewarmest quarter and precipitation of thecoldest quarter.
Diversity anomalies and climate heterogeneity
Global Ecology and Biogeography, © 2014 John Wiley & Sons Ltd 9

mental heterogeneity resulted in more opportunities for
allopatric speciation (Allmon, 1992; Qian & Ricklefs, 2000;
Barraclough, 2006) and thus higher regional diversity in EA and
WNA than in ENA, and in CFR than in SWA.
Although multivariate analyses supported the background
assumptions required by hypotheses about how differences in
geographic configuration may have caused diversity anomalies,
regional differences in the variance–covariance structure of
climate were large and regional differences in some climate vari-
ables opposed overall multivariate results. These deviations
from multivariate patterns can be used to derive novel predic-
tions from the hypotheses of interest. Those predictions can be
tested using clades as replicates (Xiang et al., 2004; Barraclough,
2006; Ricklefs, 2006). As an example, we found that, opposite to
the multivariate pattern, spatially implicit and explicit heteroge-
neity in precipitation of the driest month, the driest quarter and
the coldest quarter were higher in ENA than in EA (Fig. 5). If
these variables are particularly important in defining suitable
habitat for some clades, then predictions about regional differ-
ences in diversification rates for those clades would be opposite
to what is expected from overall regional differences in multi-
variate climate. Simulation experiments illustrate this prediction
(Appendix S4, Fig. S7).
Novel predictions may also be derived by considering spatially
explicit heterogeneity at various scales. We used the overall pro-
portion of regional environmental variation due to sinusoidal
spatial structures as a measure of spatially explicit heterogeneity.
However, this proportion can be partitioned into fractions
explained by sinusoidal structures having different wavelengths
and, thus, representing different spatial scales (Figs S2 & S3). For
example, although a lower overall proportion of spatial vari-
ation in multivariate climate was due to sinusoidal structures in
ENA than EA and WNA (Fig. 5c), and in SWA than in CFR
(Fig. 6c), these patterns are reversed at certain spatial scales (Fig.
S8). These scale-dependent regional differences can be used to
derive clade-specific predictions if clades differ in the scale at
which spatially explicit heterogeneity affects metacommunity
dynamics or opportunities for allopatric speciation.
Distinguishing spatially implicit and explicit environmental
heterogeneity can help derive exclusive predictions from
hypotheses that would otherwise appear empirically equivalent.
Where spatially implicit and explicit measures show opposite
patterns, hypotheses emphasizing different mechanisms yield
opposite predictions. Our results showed that regional differ-
ences in spatially explicit and implicit measures of heterogeneity
were consistent when measured in multivariate space, but dif-
ferent in some univariate analyses. In particular, spatially
implicit measures of mean temperature of the warmest quarter,
precipitation of the wettest month, the wettest quarter and the
coldest quarter were higher in SWA than in CFR, but the oppo-
site was true for spatially explicit measures (Fig. 6). Given that
these variables are important for some organisms, hypotheses
proposing that ecological sorting and ecological speciation are
fostered by spatially implicit heterogeneity would yield predic-
tions different from those derived from hypotheses suggesting
that coexistence through metacommunity dynamics and rates
of allopatric speciation are promoted by spatially explicit
heterogeneity.
Regional spatial environmental heterogeneity has been both
dismissed (Rosenzweig, 1995; Cowling et al., 1996) and empha-
sized (Qian & Ricklefs, 2000) as an explanation of salient exam-
ples of diversity anomalies. But it has not always been clear
whether spatially implicit or explicit heterogeneity was being
discussed, and there seems to be little previous empirical work
estimating regional differences in both kinds of heterogeneity.
We suggest it would be useful to distinguish and measure spa-
tially implicit and explicit heterogeneity when gauging empirical
support for different views of how the geographic configuration
of regions may be causally related to diversity anomalies. These
two kinds of spatial heterogeneity are not necessarily correlated
across regions and they are hypothesized to foster diversity
anomalies in distinct ways.
ACKNOWLEDGEMENTS
R.E.R. gratefully acknowledges support from the Curators of the
University of Missouri and from the Alexander von Humboldt
Foundation. Sebastián Tello provided helpful suggestions on
statistical analyses. Comments by David Currie, José Alexandre
Diniz-Filho, Bradford Hawkins and an anonymous referee sub-
stantially improved the manuscript.
REFERENCES
Allmon, D.J. (1992) A causal analysis of stages in allopatric
speciation. Oxford surveys in evolutionary biology (ed. by D.J.
Futuyma and J. Antonovics), pp. 219–257. Oxford University
Press, New York.
Barraclough, T.G. (2006) What can phylogenetics tell us about
speciation in the Cape flora? Diversity and Distributions, 12,
21–26.
Boos, D.D. & Brownie, C. (1989) Bootstrap methods for testing
homogeneity of variances. Technometrics, 31, 69–82.
Borcard, D. & Legendre, P. (2002) All-scale spatial analysis of
ecological data by means of principal coordinates of neigh-
bour matrices. Ecological Modelling, 153, 51–68.
Borcard, D., Gillet, F. & Legendre, P. (2011) Numerical ecology
with R. Springer, New York.
Born, J., Linder, H.P. & Desmet, P. (2007) The greater Cape
Floristic Region. Journal of Biogeography, 34, 147–162.
Chesson, P. (2000) Mechanisms of maintenance of species diver-
sity. Annual Review of Ecology and Systematics, 31, 343–366.
Cowling, R., Rundel, P.H, Lamont, B.B., Arroyo, M.K. &
Arianoutsou, M. (1996) Plant diversity in Mediterranean-
climate regions. Trends in Ecology and Evolution, 11, 362–366.
Cowling, R.M. & Lombard, A.T. (2002) Heterogeneity,
speciation/extinction history and climate: explaining regional
plant diversity patterns in the Cape Floristic Region. Diversity
and Distributions, 8, 163–179.
Cowling, R.M., Proches, S. & Partridge, T.C. (2009) Explaining
the uniqueness of the Cape flora: incorporating geomorphic
I. Jiménez and R. E. Ricklefs
Global Ecology and Biogeography, © 2014 John Wiley & Sons Ltd10

evolution as a factor for explaining its diversification. Molecu-
lar Phylogenetics and Evolution, 51, 64–74.
Crisp, M.D., Laffan, S., Linder, H.P. & Monro, A. (2001)
Endemism in the Australian flora. Journal of Biogeography, 28,
183–198.
Currie, D.J., Mittelbach, G.G., Cornell, H.V., Field, R., Guegan, J.,
Hawkins, B.A., Kaufman, D.M., Kerr, J.T., Oberdorff, T.,
O’Brien, E. & Turner, J.R.G. (2004) Predictions and tests of
climate-based hypotheses of broad-scale variation in taxo-
nomic richness. Ecology Letters, 7, 1121–1134.
Davison, A.C. & Hinkley, D.V. (1997) Bootstrap methods
and their application. Cambridge University Press, New
York.
Dray, S., Legendre, P. & Peres-Neto, P.R. (2006) Spatial model-
ling: a comprehensive framework for principal coordinate
analysis of neighbour matrices (PCNM). Ecological Modelling,
196, 483–493.
Forest, F., Grenyer, R., Rouget, M., Davies, T.J., Cowling, R.M.,
Faith, D.P., Balmford, A., Manning, J.C., Proches, S.,
van der Bank, M., Reeves, G., Hedderson, T.A.J. & Savolainen,
V. (2007) Preserving the evolutionary potential of floras in
biodiversity hotspots. Nature, 445, 757–760.
Goldblatt, P. & Manning, J. (2002) Plant diversity of the cape
region of South Africa. Annals of the Missouri Botanical
Garden, 89, 281–302.
Goodnight, C.J. & Schwartz, J.M. (1997) A bootstrap compari-
son of genetic covariance matrices. Biometrics, 53, 1026–1039.
Haining, R. (2003) Spatial data analysis: theory and practice.
Cambridge University Press, Cambridge.
Harrison, S. & Cornell, H. (2008) Toward a better understanding
of the regional causes of local community richness. Ecology
Letters, 11, 969–979.
Hijmans, R.J., Cameron, S.E., Parra, J.L., Jones, P.G. & Jarvis, A.
(2005) Very high resolution interpolated climate surfaces for
global land areas. International Journal of Climatology, 25,
1965–1978.
Hobbs, R.J., Richardson, D.M. & Davis, G.W. (1995) Mediter-
ranean-type ecosystems: opportunities and constraints for
studying the function of biodiversity. Mediterranean-type eco-
systems: the function of biodiversity (ed. by G.W. Davis and
D.M. Richardson), pp. 1–42. Springer-Verlag, Berlin.
Holyoak, M., Leibold, M.A. & Holt, R.D. (2005)
Metacommunities: spatial dynamics and ecological commu-
nities. University of Chicago Press, Chicago, IL.
Hopper, S.D. & Gioia, P. (2004) The Southwest Australian Flo-
ristic Region: evolution and conservation of a global hot spot
of biodiversity. Annual Review of Ecology, Evolution and Sys-
tematics, 35, 623–650.
Kreft, H. & Jetz, W. (2007) Global patterns and determinants of
vascular plant diversity. Proceedings of the National Academy of
Sciences USA, 104, 5925–5930.
Latham, R.E. & Ricklefs, R.E. (1993) Continental comparisons
of temperate-zone tree species diversity. Species diversity in
ecological communities: historical and geographical perspectives
(ed. by R.E. Ricklefs and D. Schluter), pp. 294–314. University
of Chicago Press, Chicago, IL.
Legendre, P. & Legendre, L. (2012) Numerical ecology, 3rd edn.
Elsevier, Amsterdam.
Linder, H.P. (2003) The radiation of the Cape flora, southern
Africa. Biology Reviews, 78, 597–638.
Linder, H.P. (2005) Evolution of diversity: the Cape flora. Trends
in Plant Science, 10, 536–541.
Linder, H.P., Eldenas, P. & Briggs, B.G. (2003) Contrasting pat-
terns of radiation in African and Australian Restionaceae. Evo-
lution, 57, 2688–2702.
Milewski, A.V. (1979) A climatic basis for the study of conver-
gence of vegetation structure in Mediterranean Australia and
southern Africa. Journal of Biogeography, 6, 293–299.
Mittelbach, G.G., Schemske, D.W., Cornell, H.V. et al. (2007)
Evolution and the latitudinal diversity gradient: speciation,
extinction, and biogeography. Ecology Letters, 10, 315–331.
van der Niet, D. & Johnson, S.D. (2009) Patterns of plant spe-
ciation in the Cape floristic region. Molecular Phylogenetics
and Evolution, 51, 85–93.
Peres-Neto, P.R., Legendre, P., Dray, S. & Borcard, D.
(2006) Variation partitioning of species data matrices:
estimation and comparison of fractions. Ecology, 87, 2614–
2625.
Qian, H. & Ricklefs, R.E. (1999) A comparison of the taxonomic
richness of vascular plants in China and the United States. The
American Naturalist, 154, 160–181.
Qian, H. & Ricklefs, R.E. (2000) Large-scale processes and the
Asian bias in species diversity of temperate plants. Nature,
407, 180–182.
Qian, H. & Ricklefs, R.E. (2004) Geographical distribution and
ecological conservatism of disjunct genera of vascular plants
in eastern Asia and eastern North America. Journal of Ecology,
92, 253–265.
R Development Core Team (2009) R: a language and environ-
ment for statistical computing. R Foundation for Statistical
Computing, Vienna, Austria. Available at: http://www
.R-project.org (accessed July 2009).
Ricklefs, R.E. (2004) A comprehensive framework for global
patterns in biodiversity. Ecology Letters, 7, 1–15.
Ricklefs, R.E. (2006) Evolutionary diversification and the origin
of the diversity–environment relationship. Ecology, 87, 3–13.
Rosenzweig, M.L. (1995) Species diversity in space and time.
Cambridge University Press, Cambridge.
Sauquet, H., Weston, P.H., Anderson, C.L., Barker, N.P., Cantrill,
D.J., Mast, A.R. & Savolainen, V. (2009) Contrasted patterns of
hyperdiversification in Mediterranean hotspots. Proceedings of
the National Academy of Sciences USA, 106, 221–225.
Schluter, D. (2000) The ecology of adaptive radiation. Oxford
University Press, New York.
Schluter, D. & Ricklefs, R.E. (1993) Convergence and the
regional component of species diversity. Species diversity in
ecological communities: historical and geographical perspectives
(ed. by R.E. Ricklefs and D. Schluter), pp. 230–242. University
of Chicago Press, Chicago, IL.
Svenning, J.C. (2003) Deterministic Plio-Pleistocene extinctions
in the European cool-temperate tree flora. Ecology Letters, 6,
646–653.
Diversity anomalies and climate heterogeneity
Global Ecology and Biogeography, © 2014 John Wiley & Sons Ltd 11

Turner, D. (2005) Local underdetermination in historical
science. Philosophy of Science, 72, 209–230.
Verboom, G.A., Archibald, J.K., Bakker, F.T., Bellstedt, D.U.,
Conrad, F., Dreyer, L.L., Forest, F., Galley, C., Goldblatt, P.,
Henning, J.F., Mummenhoff, K., Linder, H.P., Muasya, A.M.,
Oberlander, K.C., Savolainen, V., Snijman, D.A., van der Niet,
T. & Nowell, T.L. (2009) Origin and diversification of the
Greater Cape flora: ancient species repository, hot-bed of
recent radiation, or both? Molecular Phylogenetics and Evolu-
tion, 51, 44–53.
Wen, J. (1999) Evolution of eastern Asian and eastern North
American disjunct distributions on flowering plants. Annual
Review of Ecology and Systematics, 30, 421–455.
Wiens, M.J. (2000) Ecological heterogeneity: an ontogeny of
concepts and approaches. The ecological consequences of envi-
ronmental heterogeneity: the 40th symposium of the British Eco-
logical Society, held at the University of Sussex, 23–25 March
1999 (ed. by M.J. Hutchings, E.A. John & A.J.A. Stewart), pp.
9–31. Cambridge University Press, Cambridge.
Xiang, Q., Zhang, W.H., Ricklefs, R.E., Qian, H., Chen, Z.D.,
Wen, J. & Li, J.H. (2004) Regional differences in rates of plant
speciation and molecular evolution: a comparison between
eastern Asia and eastern North America. Evolution, 58, 2175–
2184.
Zhang, J. & Boos, D.D. (1992) Bootstrap critical values for
testing homogeneity of covariance matrices. Journal of the
American Statistical Association, 87, 425–429.
SUPPORTING INFORMATION
Additional supporting information may be found in the online
version of this article at the publisher’s web-site.
Figure S1 Illustration of computer simulation experiments
testing the effects of linear and sinusoidal spatial structures on
isolation of species suitable habitat.
Figure S2 Examples of spatial eigenvector maps of each region.
Figure S3 Examples of correlograms for spatial eigenvectors
maps of each region.
Figure S4 Mantel correlograms for redundancy analysis
residuals.
Figure S5 Correlograms for regression residuals corresponding
to western North America, eastern North America and eastern
Asia.
Figure S6 Correlograms for regression residuals corresponding
to the Cape Floristic Region and south-west Australia.
Figure S7 Simulation results show contrasting effects of spatial
heterogeneity in precipitation of the driest month and annual
mean precipitation on isolation of species suitable habitat in
eastern North America and eastern Asia.
Figure S8 Proportion of variation in multivariate climate
explained by sinusoidal spatial structures at different spatial
scales across western North America, eastern North America,
eastern Asia, the Cape Floristic Region and south-west Australia.
Table S1 Central tendency and variation in 17 climatic variables
across western North America, eastern North America and
eastern Asia.
Table S2 Central tendency and variation in 17 climatic variables
across the Cape Floristic Region and south-west Australia.
Table S3 Results of tests for differences in climate heterogeneity
that preserve spatial structure.
Appendix S1 Computer simulation experiments testing the
effect of linear and sinusoidal spatial structures on isolation of
species suitable habitat.
Appendix S2 Testing for differences in climate heterogeneity
while preserving spatial structure.
Appendix S3 Standardization of dispersion matrices.
Appendix S4 Computer simulation experiments comparing the
effects of spatial heterogeneity in precipitation of the driest
month and annual mean precipitation on isolation of species
suitable habitat in eastern North America and eastern Asia.
BIOSKETCHES
Iván Jiménez is interested in testing theory that
attempts to explain the spatial patterns of diversity and
the size, structure, and dynamics of geographic ranges
of taxa.
Robert E. Ricklefs researches diversity in ecological
systems at several levels of organization and scales of
time and space; he has a long-standing interest in the
evolutionary diversification of avian life histories and
the historical development of ecological communities
and regional species richness.
Editor: José Alexandre Diniz-Filho
I. Jiménez and R. E. Ricklefs
Global Ecology and Biogeography, © 2014 John Wiley & Sons Ltd12




















