
Expression of neuronal nitric oxide synthase during embryonic development of the rat cerebral cortex
18
Ž . Developmental Brain Research 111 1998 205–222 Research report Expression of neuronal nitric oxide synthase during embryonic development of the rat cerebral cortex M. Santacana ) , L.O. Uttenthal, M.L. Bentura, A.P. Fernandez, J. Serrano, ´ J. Martınez de Velasco, D. Alonso, R. Martınez-Murillo, J. Rodrigo ´ ´ Instituto Cajal, C.S.I.C., AÕenida del Dr. Arce, 37, E-28002 Madrid, Spain Accepted 15 September 1998 Abstract Ž . Ž. The expression of neuronal nitric oxide synthase nNOS during the development of the rat cerebral cortex from embryonic day E 13 Ž. to postnatal day P 0 was analyzed by immunocytochemical procedures using a specific antibody against rat brain nNOS. Expression of nNOS was first seen on E14 in cells of Cajal–Retzius morphology located in the marginal zone. Neuronal NOS immunoreactivity persisted in this layer throughout the embryonic period and only began to decrease on E20, when neuronal migration is coming to an end. From E17 onwards, migrating neurons expressing nNOS were observed in the intermediate zone with their leading processes directed towards the cortical plate. At the same time, efferent nNOS-immunoreactive axons originating from cortical plate cells entered the intermediate zone. From E19 onwards, cells expressing nNOS and with the morphological characteristics of migrating cells were observed in and near the subventricular zone. Confocal analysis of double immunostaining for nNOS and glial fibrillary acidic protein or nestin showed no coexpression of nNOS and glial markers in these cells, suggesting that nNOS-positive cells leaving the subventricular zone were not glial cells. Commissural, callosal and fimbrial fibers were seen to express nNOS on E18 and E19. This expression decreased from E20 and was very weak on E21 and P0. The observations suggest that nitric oxide is synthesized during embryonic life in relation to maturational processes such as the organization of cerebral lamination, and is involved in controlling migrational processes and fiber ingrowth. q 1998 Elsevier Science B.V. All rights reserved. Keywords: Neuronal nitric oxide synthase; nNOS; Embryo; Development; Rat 1. Introduction Ž . The biological significance of nitric oxide NO , a diffusible gas with free radical properties, first became evident when it was identified as the endothelium-derived Ž . relaxing factor EDRF , of major importance in vasodilata- w x tion 27,51 . NO is produced in tissues by the enzymatic oxidation of a guanidine nitrogen of arginine to form NO w x Ž . and citrulline 10 . Nitric oxide synthase NOS , the en- Abbreviations: CC: corpus callosum; CP: cortical plate; E: embryonic day; EDRF: endothelium-derived relaxing factor; eNOS: endothelial ni- tric oxide synthase; GFAP: glial fibrillary acidic protein; iNOS: inducible nitric oxide synthase; IZ: intermediate zone; LTD: long-term depression; LTP: long-term potentiation; MZ: marginal zone; NADPH: nicotinamide adenine dinucleotide phosphate; nNOS: neuronal nitric oxide synthase; NO: nitric oxide; NOS: nitric oxide synthase; P: postnatal day; SVZ: subventricular zone; VZ: ventricular zone; WM: white matter ) Corresponding author. Fax: q34-1-585-4754 zyme concerned, occurs in three main isoforms, encoded w x Ž . by different genes 10,15 : neuronal nNOS , endothelial Ž . Ž . eNOS and inducible iNOS , chiefly expressed in neu- rons, endothelial cells and macrophages, respectively. Both nNOS and eNOS are activated by Ca 2q and calmodulin w x 2q 46,71 , whereas the activity of iNOS is Ca -independent. NO has several different functions in the nervous sys- tem. It acts as a neurotransmitter andror neuromodulator w x 15,33,58 . It is involved in neuronal plasticity and is w x expressed in spinal neurons after injury 68 . It plays a role Ž . w x in hippocampal long-term potentiation LTP 2,22,23,29 Ž . w x and cerebellar long-term depression LTD 8,36 . All three isoforms of NOS are greatly increased after hy- w x poxicrischemic insult during postnatal 25 and adult life w x 47,55,72 . In these pathological situations, NO may act either as a protective or as a destructive agent depending on the stage of evolution of the ischemia and on the w x cellular source of NO 26 . 0165-3806r98r$ - see front matter q 1998 Elsevier Science B.V. All rights reserved. Ž . PII: S0165-3806 98 00140-0
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Expression of neuronal nitric oxide synthase during embryonic development of the rat cerebral cortex

Ž .Developmental Brain Research 111 1998 205–222
Research report
Expression of neuronal nitric oxide synthase during embryonicdevelopment of the rat cerebral cortex
M. Santacana ), L.O. Uttenthal, M.L. Bentura, A.P. Fernandez, J. Serrano,´J. Martınez de Velasco, D. Alonso, R. Martınez-Murillo, J. Rodrigo´ ´
Instituto Cajal, C.S.I.C., AÕenida del Dr. Arce, 37, E-28002 Madrid, Spain
Accepted 15 September 1998
Abstract
Ž . Ž .The expression of neuronal nitric oxide synthase nNOS during the development of the rat cerebral cortex from embryonic day E 13Ž .to postnatal day P 0 was analyzed by immunocytochemical procedures using a specific antibody against rat brain nNOS. Expression of
nNOS was first seen on E14 in cells of Cajal–Retzius morphology located in the marginal zone. Neuronal NOS immunoreactivitypersisted in this layer throughout the embryonic period and only began to decrease on E20, when neuronal migration is coming to an end.From E17 onwards, migrating neurons expressing nNOS were observed in the intermediate zone with their leading processes directedtowards the cortical plate. At the same time, efferent nNOS-immunoreactive axons originating from cortical plate cells entered theintermediate zone. From E19 onwards, cells expressing nNOS and with the morphological characteristics of migrating cells wereobserved in and near the subventricular zone. Confocal analysis of double immunostaining for nNOS and glial fibrillary acidic protein ornestin showed no coexpression of nNOS and glial markers in these cells, suggesting that nNOS-positive cells leaving the subventricularzone were not glial cells. Commissural, callosal and fimbrial fibers were seen to express nNOS on E18 and E19. This expressiondecreased from E20 and was very weak on E21 and P0. The observations suggest that nitric oxide is synthesized during embryonic life inrelation to maturational processes such as the organization of cerebral lamination, and is involved in controlling migrational processes andfiber ingrowth. q 1998 Elsevier Science B.V. All rights reserved.
Keywords: Neuronal nitric oxide synthase; nNOS; Embryo; Development; Rat
1. Introduction
Ž .The biological significance of nitric oxide NO , adiffusible gas with free radical properties, first becameevident when it was identified as the endothelium-derived
Ž .relaxing factor EDRF , of major importance in vasodilata-w xtion 27,51 . NO is produced in tissues by the enzymatic
oxidation of a guanidine nitrogen of arginine to form NOw x Ž .and citrulline 10 . Nitric oxide synthase NOS , the en-
Abbreviations: CC: corpus callosum; CP: cortical plate; E: embryonicday; EDRF: endothelium-derived relaxing factor; eNOS: endothelial ni-tric oxide synthase; GFAP: glial fibrillary acidic protein; iNOS: induciblenitric oxide synthase; IZ: intermediate zone; LTD: long-term depression;LTP: long-term potentiation; MZ: marginal zone; NADPH: nicotinamideadenine dinucleotide phosphate; nNOS: neuronal nitric oxide synthase;NO: nitric oxide; NOS: nitric oxide synthase; P: postnatal day; SVZ:subventricular zone; VZ: ventricular zone; WM: white matter
) Corresponding author. Fax: q34-1-585-4754
zyme concerned, occurs in three main isoforms, encodedw x Ž .by different genes 10,15 : neuronal nNOS , endothelial
Ž . Ž .eNOS and inducible iNOS , chiefly expressed in neu-rons, endothelial cells and macrophages, respectively. BothnNOS and eNOS are activated by Ca2q and calmodulinw x 2q46,71 , whereas the activity of iNOS is Ca -independent.
NO has several different functions in the nervous sys-tem. It acts as a neurotransmitter andror neuromodulatorw x15,33,58 . It is involved in neuronal plasticity and is
w xexpressed in spinal neurons after injury 68 . It plays a roleŽ . w xin hippocampal long-term potentiation LTP 2,22,23,29
Ž . w xand cerebellar long-term depression LTD 8,36 . All threeisoforms of NOS are greatly increased after hy-
w xpoxicrischemic insult during postnatal 25 and adult lifew x47,55,72 . In these pathological situations, NO may acteither as a protective or as a destructive agent dependingon the stage of evolution of the ischemia and on the
w xcellular source of NO 26 .
0165-3806r98r$ - see front matter q 1998 Elsevier Science B.V. All rights reserved.Ž .PII: S0165-3806 98 00140-0

( )M. Santacana et al.rDeÕelopmental Brain Research 111 1998 205–222206
Neuronal NOS is expressed in the brain during embryo-nic development and is considered to be an importantfactor in maturational processes. It has been shown by
w xBredt and Snyder 9 that nNOS is expressed in corticalplate neurons at E15, and decreases postnatally. Other
w xauthors 11,21,39 , using NADPH-diaphorase histochem-istry or in situ hybridization, suggested the involvement ofNO in development during late embryonic stages or in theneonatal animal. In the developing brain, NO produced bythe action nNOS appears to perform several functions. Itplays a leading role in refining axonal connectivityw x14,19,24,54,65,66,69 and may also be involved in cellu-
w xlar migration 1,17,60 . Another suggested role is to inducew xthe apoptosis 9 that is a ubiquitous feature of develop-
w xment. Palluy and Rigaud 50 have shown that NO inducesapoptosis in cortical neuronal cell cultures.
An extensive description of the expression of nNOSduring embryonic development of the cortical anlage isstill lacking. The purpose of our study was to determinethe expression of nNOS during the development of thetelencephalic vesicle, in correlation with the well-describedchanges that occur during embryonic maturation. As cau-dorostral and ventrodorsal gradients are seen in cortical
w xdevelopment 6 , we have focused our study on the so-matosensory level of the telencephalic vesicle to avoidinterference from these gradients. Neuronal NOS-immuno-reactivity was identified by immunocytochemistry using apolyclonal antibody against rat brain nNOS. Here wedescribe the expression of nNOS from embryonic day 13Ž . Ž .E13 to postnatal day 0 P0 in the different layers of theembryonic telencephalon.
The last embryonic layer to appear is the cell-producingŽ .subventricular zone SVZ . This produces astrocytes and
oligodendrocytes as well as neurons. Although nNOS isexpressed in neurons, recent findings have indicated that
w xglial cells may also express nNOS 3,35,61 . We havetherefore used fluorescent double immunostaining for glial
Ž .markers GFAP and nestinrRAT 401 and nNOS to iden-tify the nature of cells generated in the SVZ in lateembryonic stages and just after birth.
2. Materials and methods
2.1. Animals
Surgical operations and perfusion of embryos wereperformed under appropriate anesthesia. Dams were deeply
Ž .anesthetized with Equithesin Janssen, 3 mlrkg . Embryosfrom E18 onwards were anesthetized by hypothermia.Embryos were collected from timed pregnant dams byCesarean operation. The day of conception was determinedby examining vaginal smears from female rats afterovernight mating. The day of sperm-positive smears was
Ž .considered as embryonic day zero E0 . Postnatal day zero
Table 1Number of animals examined at each stage
E13 E14 E15 E16 E17 E18 E19 E20 E21 P0
12 10 7 6 6 6 10 6 4 4
Ž .P0 was the day of birth. The number of animals exam-ined at each stage is given in Table 1.
2.2. Antibodies
A polyclonal antibody to purified rat brain nNOS wasw xraised in rabbits as described elsewhere 59 and used at a
dilution of 1:3000 in phosphate-buffered saline, pH 7.4Ž .PBS . The antiserum thus obtained has been shown toreact in Western blots with nNOS from different mam-malian species and to react immunocytochemically with
w xcertain neurons of these species 53,59 .ŽThe anti-GFAP antibody a gift from Dr. J.M. Polak,
.Imperial College School of Medicine, London was a ratmonoclonal antibody that recognizes adult astrocytes andwas used at a dilution of 1:100 in PBS.
The anti-nestin antibody was the mouse monoclonalantibody RAT 401 from the Developmental Studies Hy-
Ž .bridoma Bank University of Iowa, USA . This antibodyrecognizes neuroepithelial precursor cells in both the pe-ripheral and central nervous system at very early stages ofdevelopment, as well as radial glia. It was used undiluted.
2.3. Immunocytochemistry
Immunocytochemistry was performed on brains from71 rats, fixed with 4% paraformaldehyde in 0.1 M phos-
Ž .phate buffer, pH 7.4 PB . Small embryos, from E13 toE15, were fixed entire by immersion in the fixative solu-tion. From E16 to P0 animals were perfused through theleft ventricle. Brains were removed and postfixed in freshfixative solution for 3 h at 48C. They were then cryopro-tected by soaking overnight in 30% sucrose. Vibratomesections 80 mm thick, or frozen sections 50 mm thick,were cut in frontal or sagittal planes. Free-floating sectionswere first incubated in PBS containing 3% hydrogen per-oxide for 25 min to quench endogenous peroxidase activ-ity. After washing three times in PBS for 5 min, sectionswere incubated overnight with nNOS antiserum. No Tritonwas used and sections were processed by the avidin–biotin
Ž .complex ABC method. After three washes in PBS, sec-tions were incubated with biotinylated anti-rabbit second
Ž .antibody Vector Laboratories at 1:100 for 60 min, washedŽ .in PBS and incubated in ABC Vector for 90 min. After
three washes in PBS they were washed at least three timesin sodium acetate buffer, pH 6.0. The reaction product wasvisualized by incubating the sections in 0.05% 3,3X-di-
Ž .aminobenzidine in acetate buffer containing 2% nickel IIŽ .sulfamate Merck . After incubation for 10 min, 0.03%
hydrogen peroxide was added for an additional 3 min.
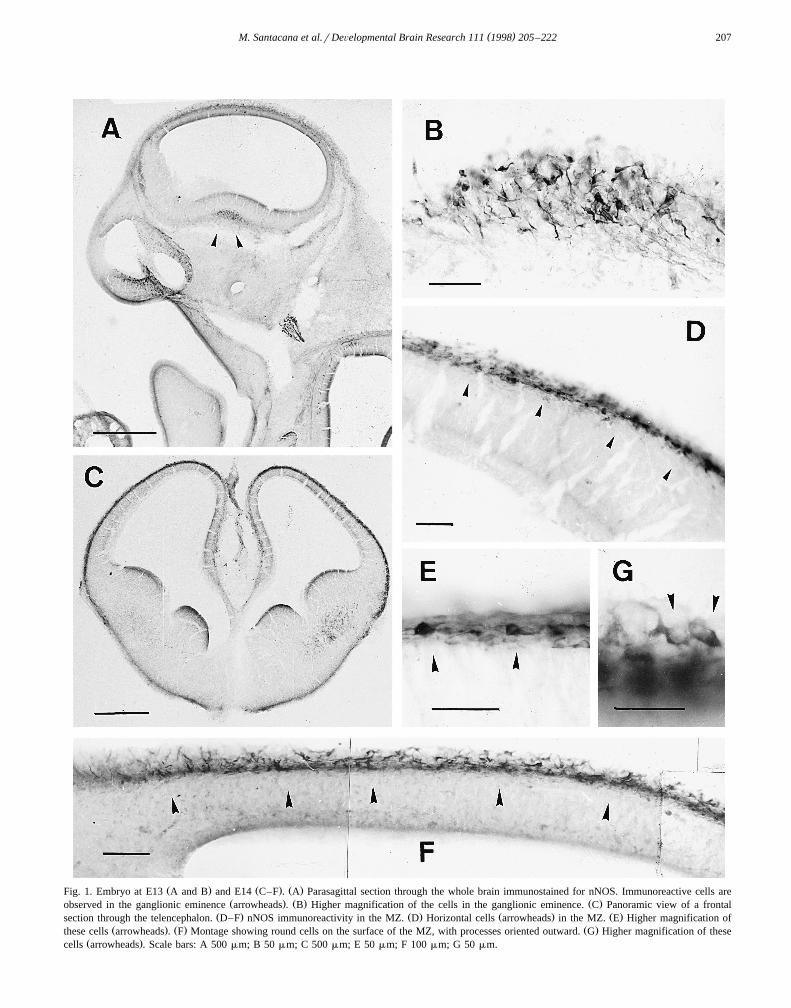
( )M. Santacana et al.rDeÕelopmental Brain Research 111 1998 205–222 207
Ž . Ž . Ž .Fig. 1. Embryo at E13 A and B and E14 C–F . A Parasagittal section through the whole brain immunostained for nNOS. Immunoreactive cells areŽ . Ž . Ž .observed in the ganglionic eminence arrowheads . B Higher magnification of the cells in the ganglionic eminence. C Panoramic view of a frontal
Ž . Ž . Ž . Ž .section through the telencephalon. D–F nNOS immunoreactivity in the MZ. D Horizontal cells arrowheads in the MZ. E Higher magnification ofŽ . Ž . Ž .these cells arrowheads . F Montage showing round cells on the surface of the MZ, with processes oriented outward. G Higher magnification of these
Ž .cells arrowheads . Scale bars: A 500 mm; B 50 mm; C 500 mm; E 50 mm; F 100 mm; G 50 mm.

( )M. Santacana et al.rDeÕelopmental Brain Research 111 1998 205–222208
Sections were washed with 0.1 M acetate buffer, then inPBS, mounted on gelatin-coated slides and coverslippedwith DPX.
2.4. Fluorescent double immunostaining
To study whether or not the cells expressing nNOSimmunoreactivity within or near the SVZ on E21 and P0were glial cells, double immunostaining for nNOS and
Ž .glial fibrillary acidic protein GFAP , or nNOS and RAT401, was carried out. The nNOS antiserum was used at adilution of 1:1000 in PBS. Two additional E21 embryosand four additional P0 animals were processed as de-
w xscribed 56 . Free-floating sections were incubated with2% BSA in PBS for 30 min and then with the primaryantibody against nNOS, overnight at 48C. After washes inPBS, sections were incubated for 3 h at room temperaturewith FluroLink-CY-2-labeled goat anti-rabbit immuno-
Ž .globulin Amersham, UK diluted 1:500. All subsequentsteps were carried out in the dark. After washes in PBS,sections were incubated overnight at 48C with either anti-GFAP or RAT 401 at the stated dilutions. After severalwashes in PBS, sections were incubated for 60 min withbiotinylated antibodies against rat or mouse immuno-globulin, diluted 1:100 in PBS. Sections were washed andincubated for 3 h at room temperature with FluroLink-CY-
Ž .5-avidin Amersham diluted 1:500 in PBS. They werethen washed again and mounted on slides. CY-2 gives agreen color with absorbance and fluorescence maxima at489 nm and 506 nm, respectively. CY-5 gives a red colorwith absorbance and fluorescence maxima at 650 nm and667 nm, respectively. The double-stained sections were
Ž .analyzed in a confocal scanner Leika TCS 4D equippedwith a low-noise ArrKr multiline laser.
2.5. Controls
Control sections were processed routinely, omitting onlythe primary antibodies. No staining was observed in thesesections.
3. Results
The nomenclature of the embryonic layers is that pro-w xposed by the Boulder Committee 7 . Brief descriptions of
the stages of embryonic development based on Nissl-stained preparations are used to place the observed distri-bution of nNOS immunoreactivity in context.
At E13, the telencephalic vesicle is composed of onlyŽ .two layers: the ventricular zone VZ , where all neurons
Ž .are generated, and the marginal zone MZ , where thefirst-generated neurons are settling.
Neuronal NOS immunoreactivity was not observed inthe telencephalic vesicle at this stage in either transverse orlongitudinal sections, but nNOS-expressing cells were
Ž .found in the ganglionic eminence Fig. 1A . These cellsshowed varying morphological features: some were bipolarwhile others were multipolar, but they had well-developed
Ž .processes Fig. 1B . No fibers originating from these cellswere seen to enter the neighboring telencephalic vesicle.Cells and fibers expressing nNOS were observed in theposterior part of the mesencephalic vesicle, cranial nervesand ganglia, and in the spinal cord, as well as in theolfactory pit. No immunoreactivity was observed in the VZof either the telencephalic or the mesencephalic vesicles.
At E14, the whole telencephalic vesicle has greatlyenlarged and increased in thickness, mainly due to growthof the VZ. The MZ is also thicker. An incipient intermedi-
Ž .ate zone IZ between the VZ and the MZ is observed. Acharacteristic feature at this stage is the appearance in theganglionic eminence of a ventricular fold, which corre-sponds to the intermediate subpallial sulcus separating thestriatal from the pallial neuroepithelium.
The difference in nNOS immunoreactivity between E13and E14 embryos was striking. In contrast to the situationat E13, the whole MZ of the telencephalic vesicle was
Ž .populated by nNOS-positive cells Fig. 1C–G . Therewere two chief types of nNOS-immunoreactive cells: largecells in the middle part of the MZ with long tangentiallyoriented processes, which showed the morphological char-
Ž .acteristics of Cajal–Retzius cells Fig. 1D,E , and exter-nally situated round cells with processes oriented upwardlyŽ .Fig. 1F,G . The latter type of cell could be observed up toE15 but was not seen thereafter. Fibers containing nNOSimmunoreactivity were observed running tangentially inthe lower part of the MZ. No nNOS immunostaining wasseen in the VZ.
As the embryo develops, more layers appear in thetelencephalic vesicle. E15 is characterized by the presenceof three clearly visible layers: the VZ, IZ and MZ. Thetelencephalic vesicle is thicker because of the growth ofthe IZ and, to a lesser extent, because of the expansion of
Ž .the VZ. The cortical plate CP is beginning to appear inthe lateral aspects of the telencephalic vesicle.
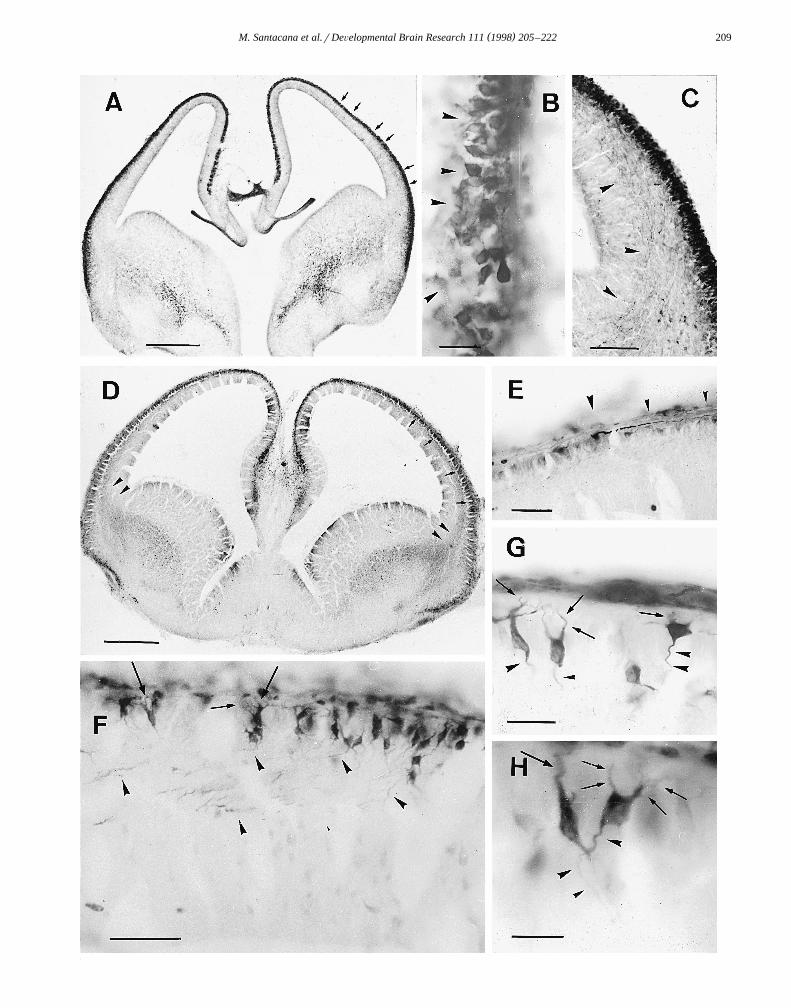
At this stage, the whole extent of the MZ was populatedŽ .by nNOS-positive cells Fig. 2A . These cells displayed
Ž . Ž . Ž . Ž . Ž .Fig. 2. Embryo at E15 A–C and E16 D–H . A Frontal section trough the telencephalon; the MZ is intensely immunostained arrows . B LateralŽ . Ž . Ž . Ž .aspect of the telencephalic vesicle showing immunoreactive CP cells arrowheads . C Immunostained fibers entering the IZ arrowheads . D Panoramic
Ž . Ž .view of a frontal section through the telencephalon; the CP shows nNOS immunoreactivity arrows and immunoreactive fibers enter the IZ arrowheads .Ž . Ž . Ž .E Example of an immunoreactive cell in the MZ with a long process arrowheads . F Immunoreactive cells in the CP; dendritic processes are directed to
Ž . Ž . Ž .the MZ arrows and axonal processes enter the IZ arrowheads . G and H Higher magnification of CP cells showing their dendritic arborizationsŽ . Ž .arrows and axonal processes arrowheads . Scale bars: A 500 mm; B 50 mm; C 100 mm; D 500 mm; E and F 100 mm; G and H 50 mm.
( )M. Santacana et al.rDeÕelopmental Brain Research 111 1998 205–222 209

( )M. Santacana et al.rDeÕelopmental Brain Research 111 1998 205–222210
the morphological characteristics of Cajal–Retzius cells asdescribed above. In the lateral aspect of the telencephalicvesicle, some nNOS-positive cells belonging to the incipi-
Ž .ent CP had appeared Fig. 2B . Some of their immunoreac-tive processes were observed to extend radially into the IZ.In addition, immunoreactive axonal processes were also
Ž .seen to enter the lateroventral aspect of the IZ Fig. 2C .These processes originated from the ganglionic eminence.They were more numerous in the lateral part of the IZ anddecreased in number in the dorsal and medial aspects,where they were found under the lower border of the MZ.As in previous stages, no labeling was observed in the VZ.
At E16, the telencephalic vesicle has four layers: theVZ, which occupies roughly one-third of the thickness, theIZ, with easily visible fibers coursing along it, the CP, andthe MZ. The CP consists of a few rows of cells and ismore prominent in the lateral than in the dorsal aspect ofthe telencephalic vesicle. It does not reach the dorsalmostpart of the telencephalic vesicle at the level of the so-matosensorial primordium.
Neuronal NOS immunoreactivity was found in the MZin tangentially oriented cells resembling Cajal–Retziuscells. These cells had processes coursing tangentially within
Ž .the MZ Fig. 2E . Many nNOS-immunoreactive cells wereŽ .observed in the CP Fig. 2D,F–H . These cells were
oriented radially, with dendritic processes running to theMZ and arborizing in it, and an axonal process running
Ž .obliquely into the IZ Fig. 2F . Apart from these axonalprocesses, many immunoreactive fibers were observedcrossing between the ganglionic eminence and the telen-
Žcephalic vesicle to enter the IZ at its lateral aspect Fig..2D .Less well-developed embryos of the same litter showed
many Cajal–Retzius cells in the MZ and fewer immunos-tained cells in the CP. As in previous stages, no immuno-staining was observed in the VZ.
At E17, the structure of the telencephalic vesicle hasbecome more complex. The CP is well-developed between
Ž .the MZ and the IZ. Below the CP the subplate SPappears as a pale band, but is not completely deficient incells and fibers. The SVZ begins to develop between theIZ and the VZ, while the latter shows approximately thesame thickness as at E16.
At this stage, nNOS immunoreactivity was foundthroughout the telencephalic vesicle. At low magnificationit appeared as three layers corresponding to the MZ, the
Ž .CP and the IZ Fig. 3A . The SVZ was not clearlydelimited from the IZ and both layers appeared as a singleband. At higher magnification it was observed that theheavy immunostaining of the MZ was due to the dendritic
processes of the immunoreactive CP cells and to im-munoreactive intrinsic cells showing the morphological
Ž .features of Cajal–Retzius cells Fig. 3B . These cells weredistributed along the whole extent of the MZ and weremore abundant near the anlage of the piriform cortex. Inthe CP, nNOS-positive cells were bipolar and radiallyoriented, with dendritic processes extending upward to theMZ. They were densely packed and formed a dense pal-
Ž .isade of cells and fibers in some parts of the CP Fig. 3A .Many nNOS-immunoreactive fibers were seen in the
IZ. Some were axonal processes from the internal andexternal capsules, while others originated from cells of the
Ž .cortical plate Fig. 3A and C . The most striking featurewas that many migrating cells were observed crossing thewhole extent of the IZ. They were radially oriented, with
Ž .their leading processes directed toward the CP Fig. 3C .Some of these cells were arranged horizontally, suggestinga tangential migration. No immunostaining was found inthe VZ.
From E18 onwards, important developmental featuresinclude the consolidation of incipient commissural connec-tions and the development of the SVZ. A significantfeature is the shrinking of the lateral ventricles due to thethickening of the telencephalic vesicle, and the develop-ment of the basal ganglia, septal and thalamic structures.The intermediate subpallial sulcus of the lateral ventricle isno longer present. The VZ begins to decrease, while theSVZ, IZ and CP continue to increase in thickness. The CPhas enlarged and now consists of numerous rows of denselypacked cells. The SP is observed as a pale band of cellsbeneath the CP. The IZ and the SVZ are clearly distin-guishable in Nissl-stained preparations. The VZ appears asa heavily stained band surrounding the ventricle.
At E18, nNOS immunoreactivity observed at low mag-Ž .nification showed five clearly defined layers Fig. 4A,B .
The MZ was densely labeled, and the underlying CPshowed a band of immunostained cells in which nearlyevery cell was stained, so that it was difficult to identifysingle cells. The SP appeared as a faintly stained layer. Incontrast with the Nissl stained preparations, the IZ andSVZ appeared as a single heavily stained band withoutdistinction between the two layers. The VZ was seen as apale band devoid of staining. At higher magnification thelabeling in the MZ was seen to be due to both the intrinsicneurons of the MZ and the dendritic arborizations of CP
Ž .neurons Fig. 4C,D , as at the previous stage. Fibers werealso observed coursing horizontally in the MZ. The CPwas seen as a palisade of homogeneously stained cells,with processes extending radially to the MZ. The SPshowed less immunoreactivity than the CP and appeared as
Ž . Ž .Fig. 3. Embryo at E17. A Frontal section through the telencephalon; note the intense immunostaining of the CP arrows and immunoreactive fibersŽ . Ž . Ž .entering the IZ arrowheads . B High magnification view of an immunostained horizontal cell in the MZ. C Montage showing the telencephalic vesicle
Ž . Ž .at higher magnification. Note the elongated immunostained CP cells long arrows , immunostained fibers coursing horizontally along the IZ short arrowsŽ .and many immunoreactive migrating cells traversing the IZ arrowheads . Scale bars: A 500 mm; B and C 50 mm.

( )M. Santacana et al.rDeÕelopmental Brain Research 111 1998 205–222 211

( )M. Santacana et al.rDeÕelopmental Brain Research 111 1998 205–222212

( )M. Santacana et al.rDeÕelopmental Brain Research 111 1998 205–222 213
a pale band beneath it, crossed by many radially orientedfibers. These fibers might be processes of neurons duringtheir migration to the CP. The IZ was similar to that ofE17 embryos: many immunoreactive fibers ran in a tan-gential direction while some showed a radial orientation,and numerous immunoreactive migrating cells were ob-served.
At this stage, the immunoreactive fibers destined toform the corpus callosum began to approach the midlineand showed strong nNOS immunoreactivity. The fimbria–
Ž .fornix system was also heavily stained Fig. 4A . Nolabeling was observed in the VZ.
At E19, the whole brain has greatly increased in vol-ume. Commissural connections such as the corpus callo-sum and anterior commissure, or long-distance connectionssuch as the fornix, are well-developed and close to reach-ing their destinations. The CP is well-developed and ex-tends along the whole telencephalic vesicle, accounting for30% of the total thickness. The SP appears as a band ofcells separated from the CP by a thin band of fibers. TheIZ is rich in fibers and the white matter primordium hasappeared in its upper part. The SVZ is well-developed andis clearly delimited between the IZ and the VZ, whereasthe latter is now notably reduced.
Neuronal NOS immunoreactivity could be observed inevery layer except the VZ, showing well-defined bands of
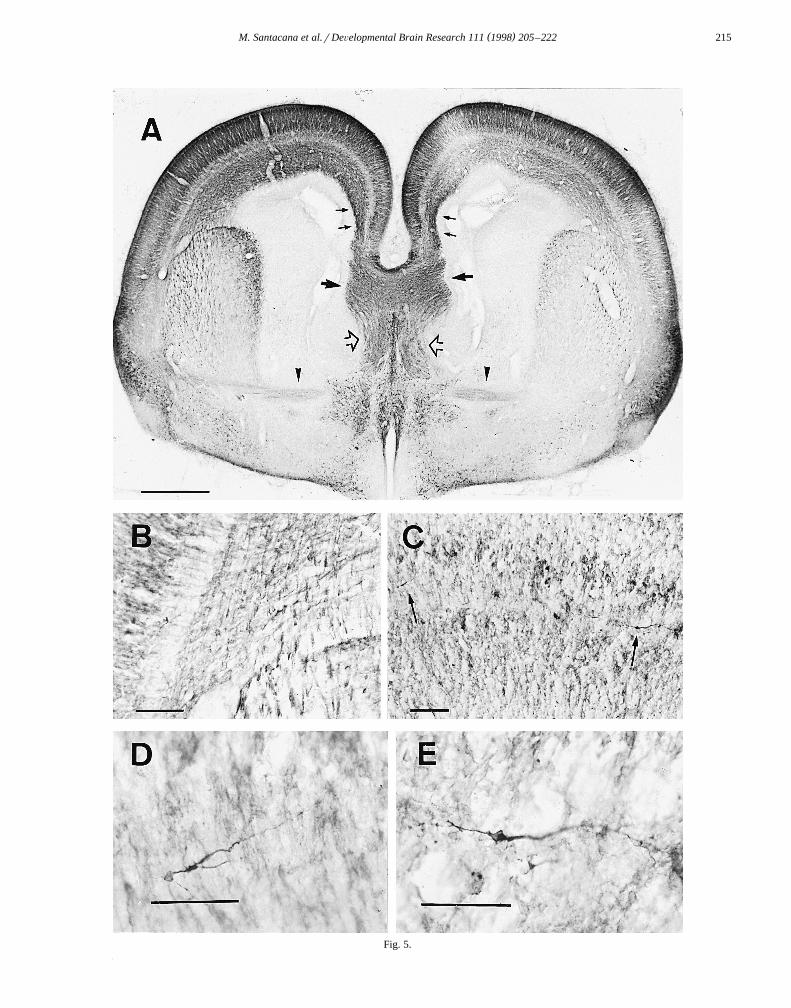
Žintense immunoreactivity and bands of paler staining Fig..5A . The most intense immunoreactivity was seen in the
ŽMZ, the deeper layers of the CP prospective cortical.layers V and VI and the IZ. A notable feature was the
intense staining of discrete regions of the SVZ. Thisstaining was most prominent in the SVZ below the cingu-late gyrus. In the SP, horizontally oriented immunostainedneurons were observed. The morphology and location ofthese neurons suggested that they were migrating neurons.Their horizontal disposition suggested a tangential migra-
Ž .tion Fig. 5C–E . As in previous stages, fibers from theinternal capsule were observed crossing the boundary of
Ž .the striatum to the IZ Fig. 5B .Intensely immunostained callosal axons were seen to
cross the midline at this stage. They were densely im-munoreactive, as were fibers of the fimbria–fornix and
Ž .those of the anterior commissure Fig. 5A .At E20, the CP is thicker and with the SP accounts for
half the thickness of the telencephalic tissue. The appear-ance of the SP is similar to that of the preceding stage. TheIZ has also increased in thickness and the SVZ is clearlydelimited. The VZ is heavily stained in Nissl preparations,
surrounding the walls of the ventricles but accounting foronly 6% of the total neocortical volume.
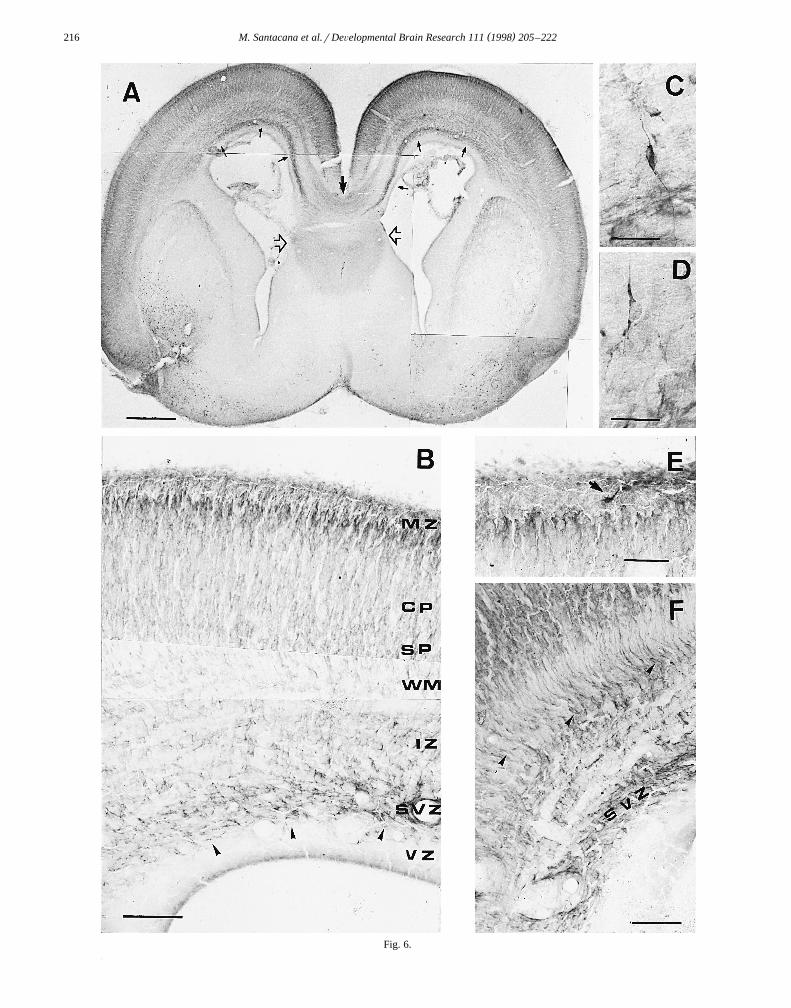
At this stage, the whole telencephalic vesicle was heav-ily stained for nNOS immunoreactivity but the MZ had
Ž .begun to lose its labeling Fig. 6A,B . Immunostained cellsand fibers could nevertheless be observed in this layerŽ .Fig. 6E . As in the previous stage, different bands oflabeling were observed throughout the telencephalic vesi-
Ž .cle Fig. 6A . In the CP, the most intense immunostainingwas observed in the prospective layers IIrIII, just below
Ž .the MZ Fig. 6B . Labeling was also seen in the deeperlayers of the CP, while less intense immunoreactivityoccurred in the SP, which was crossed by many perpendic-ular immunostained fibers. The IZ was occupied by im-munoreactive fibers corresponding to the internal capsule,the external capsule and to axons originating from CPneurons. The SVZ was heavily stained, mainly at the level
Ž .of the cingulate gyrus Fig. 6F . Many immunoreactivecells were observed in and near the SVZ, directing theirleading processes to the cingulate gyrus and the neighbor-
Ž .ing cortex Fig. 6C,D . No staining was seen in the VZ.The heavy immunostaining of the corpus callosum ob-
served at E19 was weaker at E20, by which time almost allthe commissural fibers have crossed the midline. Staining
Žof the fimbria–fornix complex had also decreased Fig..6A .At E21, the telencephalon largely resembles that of
newborn rats. The ventricles have shrunk and many struc-tures can easily be recognized as similar to those of moremature rats. The VZ is greatly decreased and is reduced toa thin band lining the ventricular wall. The SVZ is veryprominent in Nissl-stained preparations, bordering the lat-eral and dorsal aspects of the lateral ventricles. The IZ hasgreatly increased in thickness and can be subdivided intolower and upper parts. The SP is clearly visible and similarto the previous stage. The CP has increased in thicknessbecause of the arrival of new neurons and the developmentof layers V and VI.
At low magnification nNOS immunoreactivity was ob-served to be distributed in different bands of heavy and
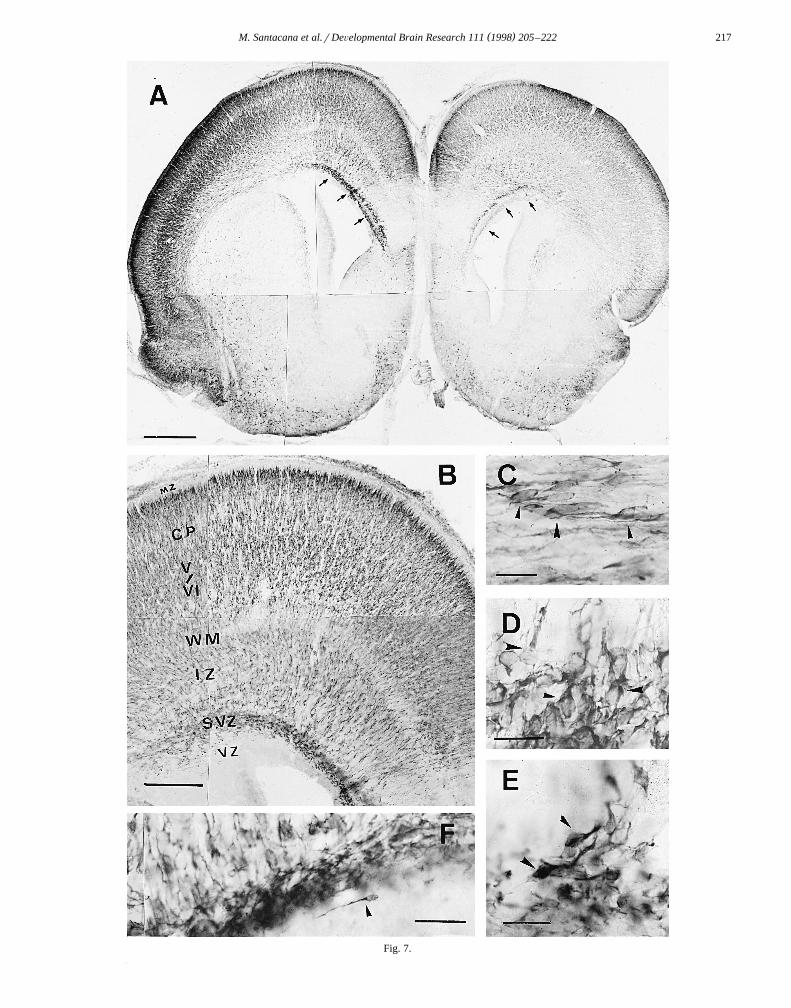
Ž .weak staining Fig. 7A . The most notable feature was adecrease in immunoreactivity of the MZ. At low magnifi-
Ž .cation this layer appeared as a clear band Fig. 7B similarto that observed in Nissl-stained preparations. Higher mag-nification revealed the presence of some immunoreactivecells retaining the morphology of Cajal–Retzius cells.Horizontally coursing nNOS-immunoreactive fibers couldalso be observed in the MZ. The most heavily stained band
Ž .Fig. 4. Embryo at E18. A Frontal section through the telencephalon showing nNOS immunoreactivity. Immunostaining is distributed in bandscorresponding to the different embryonic layers, the MZ being the most heavily stained, while the CP and IZ are also well-stained. Note the
Ž . Ž .immunostaining of the corpus callosum and fornix arrowheads . B Montage of a sagittal section at higher magnification showing the distribution ofimmunoreactivity in the different embryonic layers; a group of highly immunoreactive cells is seen at the rostral pole of the telencephalic vesicleŽ . Ž . Ž . Ž . Ž .arrowheads . C Higher magnification view of immunoreactive cells in the MZ arrowheads . D The MZ shows arborization of CP cells arrowheads ,
Ž .together with intrinsic neurons arrows . Scale bars: A 500 mm; B 200 mm; C 25 mm; D 50 mm.

( )M. Santacana et al.rDeÕelopmental Brain Research 111 1998 205–222214
corresponded to that destined to form layers IIrIII at laterstages of development. Cells of this layer were elongated,with dendrites directed towards layer I, but the immuno-staining of these processes disappeared when they entered
Ž .the MZ Fig. 7B . Layers V and VI also showed stronglabeling of the pyramidal neurons. Layer IV, although notcompletely devoid of immunoreactivity, showed neuronsthat were pale in comparison with those of layers IIrIIIand layers V and VI. The SP was less stained than layersV and VI, but contained some immunoreactive cells. TheIZ showed two types of immunostained fibers: in thelateral part, horizontally coursing fibers had the sameappearance as in previous stages, migrating cells being
Ž .observed among them Fig. 7C , while in the medial part,just below the cingulate gyrus, many radially orientedfibers and numerous cells arising from the SVZ were
Ž .observed Fig. 7B,D–F . The latter cells also had themorphological features of migrating cells and their leadingprocesses were oriented toward the cingulate gyrus and theneighboring neocortex. A very few cells with the morphol-ogy of migrating cells were observed just below the SVZŽ .Fig. 7F . Only weak immunoreactivity was observed inthe corpus callosum, whereas the external capsule and theinternal capsule were strongly immunoreactive.
Ž .At the day of birth P0 , the general appearance of thebrain is quite similar to that of older animals, althoughdevelopment is far from complete. The cortical neurons areimmature and their dendritic arborizations very poorlydeveloped. The layers characteristic of the mature cortexare still not well-defined and the callosal and thalamicaxons do not reach their final destinations until the P3–5.The SVZ is very prominent at this stage, specially at thelateral border of the lateral ventricles, and the VZ isreduced to a thin layer lining the ventricles.
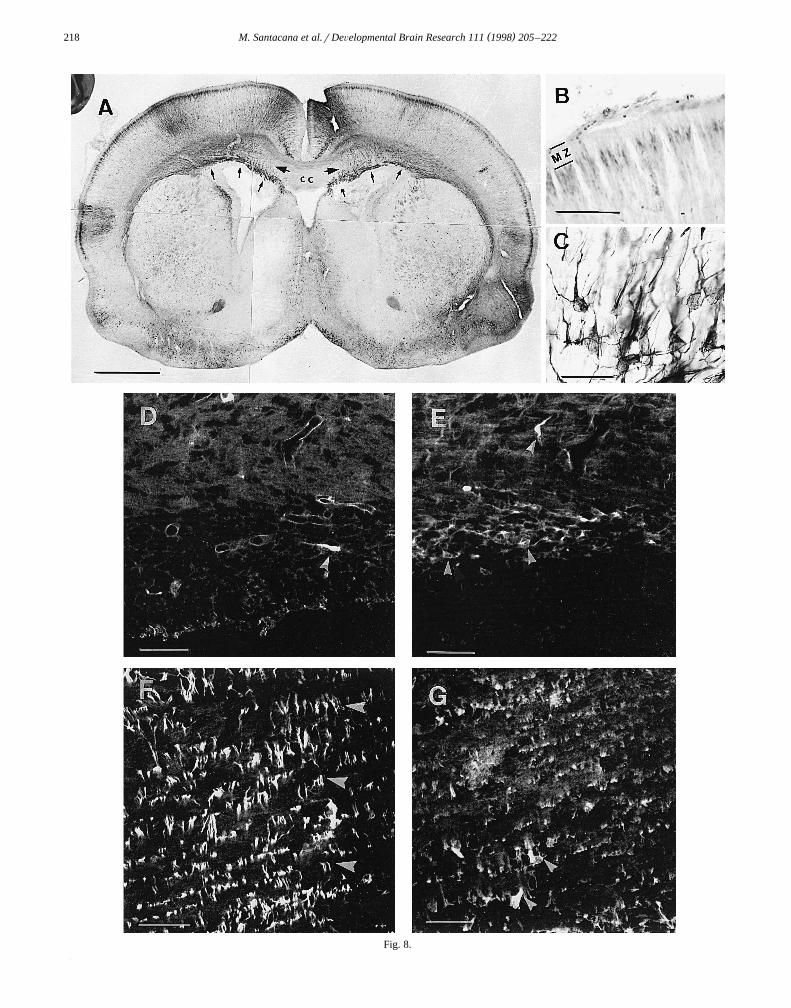
At P0, the MZ was almost devoid of nNOS immuno-reactivity and appeared as a clear band. Nevertheless,
Žsome cells and fibers could be observed in this layer Fig..8A,B . In the CP, most immunostaining was observed in
prospective layers IIrIII. As in the previous stage, theprocesses that reached layer I lost their labeling when they
Ž .entered it Fig. 8B . The CP was populated by manyimmunoreactive cells and radially oriented fibers. Most ofthe nNOS-immunoreactive neurons were pyramidal cellsof layers V and VI. Layer IV showed less staining than therest of the CP. Apparently, all fibers populating the IZ,
which is now forming the white matter and the corpuscallosum, were nNOS immunoreactive. The SVZ washeavily stained, with many cells arising from it and orient-
Žing their leading processes toward the cingulate gyrus Fig..8C .The SVZ is considered a source of glial cells, but it also
produces neurons. To define the nature of the immunoreac-tive cells seen in and near the SVZ, double immunofluo-rescence staining for nNOS and GFAP, and nNOS andRAT 401, was performed. As expected in animals of thisage, little GFAP labeling was observed. It was neverthe-less sufficient to show the absence of double immuno-staining within the same cells; elements expressing GFAPdid not coexpress nNOS and those expressing nNOS did
Ž .not coexpress GFAP Fig. 8D and E . RAT 401 labelingoccurred throughout the telencephalic vesicle and no dou-ble immunostaining with nNOS and RAT 401 antibodies
Ž .was observed Fig. 8F,G .
4. Discussion
The first embryonic neurons to express nNOS in thetelencephalic vesicle were those situated in the MZ. Ac-
w xcording to the studies of Bayer and Altman 5 usingtritiated thymidine, the Cajal–Retzius cells that are charac-teristic of this layer are generated at E13 and E14. Theyare easy to recognize from their morphological featuresand not easily confused with other cells found in this layerw x40 . Cajal–Retzius cells play a significant role throughoutthe development of the cerebral cortex. According to
w xMarin-Padilla 41 , every cell generated during the devel-opment of the cerebral cortex establishes contact withCajal–Retzius cells of the MZ. As maturation proceeds,only pyramidal neurons retain and expand their originalconnections with the MZ, while other neuronal types losethem. Using organotypic cultures with ablation of Cajal–
w xRetzius cells, Del Rio et al. 16 have recently shown thatin the hippocampus, these cells direct the ingrowth ofentorhinal fibers to the appropriate target. Another impor-tant role of the Cajal–Retzius cells is to direct the migra-tion of every neuron generated to the appropriate position
w xin the CP. Ogawa et al. 49 have shown that Cajal–Re-tzius cells express the protein reelin. This protein, absentin reeler mice, is crucial for the correct migration of
Ž .Fig. 5. Embryo at E19. A Frontal section through the telencephalon. Immunostaining is distributed in different bands, as in the preceding stage. Note theŽ . Ž . Ž . Ž .immunostaining of the SVZ small arrows . The corpus callosum larger arrows , fornix clear arrows and anterior commissure arrowheads are
Ž . Ž . Ž . Žwell-labeled, as are axons of the internal capsule. B Internal capsule axons crossing to the IZ. C Immunoreactive migrating cells in the SP arrows . D.and E Higher magnification view of these cells. Scale bars: A 500 mm; B and C 100 mm; D and E 50 mm.
Ž .Fig. 6. Embryo at E20. A Frontal section through the telencephalon showing the distribution of nNOS immunostaining. Note the increasedŽ . Ž . Ž .immunoreactivity in the SVZ small arrows . The corpus callosum medium arrow and fimbria–fornix system clear arrows show nNOS immunoreactiv-
Ž .ity. B Higher magnification view of the telencephalic vesicle. The MZ is clearer than in preceding stages, while the border between the MZ and CPŽ .shows increased immunoreactivity. C and D Higher magnification views showing the fusiform morphology of migrating cells arising from the SVZ, with
Ž . Ž .their leading processes directed toward the cingulate gyrus. E The MZ showing a Cajal–Retzius-like cell. F Detail of the SVZ showing radial fibersŽ .arrowheads . Scale bars: A 500 mm; B and E 100 mm; C and D 25 mm; F 50 mm.
( )M. Santacana et al.rDeÕelopmental Brain Research 111 1998 205–222 215
Fig. 5.
( )M. Santacana et al.rDeÕelopmental Brain Research 111 1998 205–222216
Fig. 6.
( )M. Santacana et al.rDeÕelopmental Brain Research 111 1998 205–222 217
Fig. 7.
( )M. Santacana et al.rDeÕelopmental Brain Research 111 1998 205–222218
Fig. 8.

( )M. Santacana et al.rDeÕelopmental Brain Research 111 1998 205–222 219
cortical neurons and the establishment of appropriate corti-cal lamination.
Only a few authors have studied nNOS or NADPH-di-aphorase expression in Cajal–Retzius cells during embry-
w xonic development 9,44,69 . These cells are seldom seen inmature animals, but whether they die or are dispersed in amuch greater tissue volume during most of adult life is apoint of controversy among different authors. In the adult
w xrat, layer I cells are not stained for nNOS 53 , but we haverecently shown that nNOS-immunopositive neurons of Ca-jal–Retzius morphology can be found in layer I of aged
w xrats 62 . These results suggest that the lack of nNOSexpression by these cells of layer I is a prolonged, buttransient feature of adult life. In the present study, theintense immunostaining and morphological characteristicsof these cells persisted during the embryonic period andonly began to decrease at E20, when neuronal migration iscoming to an end.
The loss of nNOS expression by Cajal–Retzius cells isnot concomitant with their disappearance. In previous stud-ies on the development of the telencephalic vesicle using
w xtritiated thymidine 64 , we observed that cells populatingthe MZ were abundant from E20 to the first postnatalstages. Autoradiographic studies had shown that these cellswere generated between E12 and E14, as is the case for the
w xCajal–Retzius cells. Recently, Meyer et al. 43 have showna new class of reelin-expressing cells in the rodent MZ,with the same morphology and location as the roundneurons with outwardly oriented processes that we haveobserved at E14 in the external part of the MZ. Accordingto these authors this type of cell persists for a longerpostnatal period than the Cajal–Retzius cells. This sug-gests that the loss of nNOS expression by either type ofneuron is not due to the disappearance of the cells them-selves but may be due to a change in their function.Considering the role of these neurons during development,the massive expression of nNOS in the MZ elements andits coincidence with the duration of the migrational pro-cess, we suggest that NO may be involved in directing theingrowing axons and in the migrational process.
During the development of the telencephalic vesicle,numerous afferent fibers reach the cortex from subcortical
Ž .structures, but some of them may express trophic factor sbefore any expression of neurotransmitter. It is well-knownw x4 that at E15 afferent fibers from the nucleus basalismagnocellularis enter through the anterolateral edge of thecortex, and grow dorsally, medially and posteriorly tocover the entire cortex. These do not express cholineacetyltransferase until the second postnatal week, whereasNGF is expressed in the embryonic basal forebrain as early
w xas E13 34 . Another set of fibers arriving at the telen-cephalic vesicle at this stage are the dopamine fibersoriginating from the substantia nigra and the ventral
w xtegmental area 30 . Their progress to the telencephalicvesicle is similar to that of fibers from the basal forebrainw x6 . In the present study, many nNOS-positive fibers havebeen observed to arise from the ganglionic eminence and
w xenter the IZ, as noted by the above authors 4,30 . The pathfollowed by these fibers corresponds to that described forthe first fibers to reach the cortical primordium. This isalso in agreement with our observation of nNOS expres-sion in cells of the ganglionic eminence at E13. The timeof appearance of these cells corresponds to that described
w xby Koh and Loy 34 for the first cells to be generated inthe basal forebrain. In addition to these afferent fibers,
ŽnNOS-positive axons originating from cells in the CP E15.onwards enter the IZ and take an oblique direction within
it. This observation is interesting because it proves thatefferent as well as afferent fibers in the IZ express nNOS.While the present study does not permit us to identify theother neurotransmitters destined to be produced by thenNOS-positive afferent fibers reaching the telencephalicvesicle, these observations strongly suggest that NO isamong the first transmitters to be synthesized in the brainŽ w xe.g., contrasting the present data and that in Ref. 31 with
w x.Ref. 52 , and plays a role in situations in which axonalpathfinding is taking place, thus contributing to neuronaland cortical plasticity.
At E17, there is a parallel expression of nNOS in thecells of the CP and the fibers populating the IZ. Apart
Ž .Fig. 7. Embryo at E21. A Montage of a frontal section through the telencephalon showing the general distribution of nNOS immunoreactivity; increasedŽ . Ž .immunostaining of the SVZ is observed small arrows . B Montage at higher magnification of a frontal section of part of the telencephalic vesicle,
showing the now unlabeled MZ, increased staining of the prospective cortical layers IIrIII and the general distribution of nNOS immunoreactivity in otherŽ . Ž . Ž . Ž . Ž .layers. C Migrating cells in the lateral aspect of the SVZ arrowheads . D and E Cells arising from the SVZ arrowheads . F A migrating cell located
Ž .below the SVZ arrowhead . Scale bars: A 500 mm; B 200 mm; C–F 25 mm.
Ž . Ž .Fig. 8. Newborn P0 . A Montage showing the distribution of nNOS immunoreactivity in a frontal section of the telencephalon. The MZ is unlabeled,while prospective cortical layers IIrIII form a heavily stained band. Some immunostaining is observed in the dorsal part of the corpus callosum. The
Ž . Ž .heaviest staining is observed in the SVZ small arrows . from which radially oriented immunostained fibers medium arrows are directed toward theŽ . Ž .cingulate gyrus. B Photomicrograph showing the MZ in which no immunostaining is observed. C Magnification of some of the cells found arising from
Ž .the SVZ. D and E Confocal scanning reconstructions of double immunostaining for GFAPrnNOS in the SVZ. GFAP immunoreactivity was stained withŽ . Ž . Ž . Ž .the CY-5 fluorochrome D and nNOS immunoreactivity with the CY-2 fluorochrome E . The cell marked in D arrowhead expresses GFAP but does
Ž . Ž . Ž .not coexpress nNOS. The cells marked in E arrowheads contain nNOS but do not coexpress GFAP. F and G Confocal scanning reconstructions ofŽ .double immunostaining for RAT 401rnNOS in a region just above the SVZ. RAT 401 was stained with the CY-5 fluorochrome F and nNOS with the
Ž .CY-2 fluorochrome G . The numerous RAT 401-reactive fibers observed in F do not show nNOS immunoreactivity, while the nNOS-immunoreactiveŽ . Ž .cells arrowheads and fibers seen in G show no RAT-401 immunoreactivity. Scale bars: A 1000 mm; B 100 mm; C 30 mm; D–G 50 mm.

( )M. Santacana et al.rDeÕelopmental Brain Research 111 1998 205–222220
from the fibers from the basal forebrain and the dopamin-ergic fibers described above, fibers from the thalamic
w xnuclei also reach the telencephalic vesicle 12,37,67 . Theyfollow a path similar to that of the fibers that have arrivedearlier. In the present study, we have observed that suchfibers express nNOS and contribute to the strong labelingof the IZ at this stage. The coincident expression of nNOSin the cells of the CP and the axons arriving there suggestsan interaction between the target region and the ingrowingaxons mediated by NO. This situation is comparable withdevelopmental mechanisms in the visual system of the
w x w xpostnatal ferret 13,14 , the rat superior colliculus 45 andw xthe avian cerebellum 17 , although in these cases it is the
retrograde action of NO synthesized in the target regionthat is regarded as significant. In our opinion, a similarmechanism may act in the establishment of connectionsbetween the ingrowing axons from different subcorticalstructures and the nNOS-expressing cells of the rat CP.
Ž .Another important feature at this stage E17 is thepresence of migrating cells expressing nNOS in the IZ.The expression of nNOS or NADPH-diaphorase and therole of NO in cell migration are controversial issues. Some
w xauthors 57 suggest that, in the cerebellum, cells expressnNOS only when migration is completed. But studies in
w xslice culture of rat cerebellum 60 showed that granuleŽ .cell migration was inhibited by N-nitro-L-arginine L-NNA
and by hemoglobin, indicating that NO was involved incell migration and in the differentiation of granule cellsand Bergman glia. In our study, the presence of migratingcells expressing nNOS in the IZ of the telencephalicvesicle suggests that NO is involved in cell migration. It isconsistent with the marked expression of nNOS by theCajal–Retzius cells already described, and their function ofdirecting the migration of every cell in the telencephalicvesicle.
From E18 onwards, a new embryonic layer is clearlyobserved in Nissl-stained sections, the SVZ. As the mainsource of neurogenesis, the VZ, shrinks, many cells areproduced in the SVZ. This layer does not completelydisappear after birth but continues to generate cells. It hasbeen thought that the germinal cells in the SVZ produce
w xonly astrocytes and oligodendrocytes 6 . In vitro experi-ments have nevertheless shown that neurons are producedin the SVZ together with glial cells in the juvenile animalw x70 . There is no doubt that bulbar granular cells areproduced postnatally in the lateral horn of the lateralventricle and migrate to the olfactory bulb in the rostral
w xmigratory stream 18,32,38,42 . In the present study, wehave observed a notable nNOS immunostaining of theSVZ from E19 onwards. We have also seen nNOS-positivecells with the morphological characteristics of migratingcells in and near the SVZ. These results agree with those
w xof Gates et al. 20 , who used NADPH-diaphorase as aneuronal marker for cells observed around the subventricu-lar zone during early postnatal stages. The lack of coex-pression of nNOS and GFAP or nNOS and RAT 401 leads
us to conclude that the nNOS-positive cells leaving theSVZ are not glial cells. The coincidence of the cell-gener-ating activity of the SVZ and the migrating morphology ofthese nNOS-positive elements suggests, again, a possiblerole of NO in migratory activity.
Another point of interest is the expression of nNOSduring the establishment of the commissural callosal fibers.At E18, fibers destined to form the corpus callosum begin
w xto approach the midline 28,63 . At this stage, nNOS-im-munoreactive fibers were observed in the corpus callosumand the immunostaining was maximal at E19. From E20onwards, nNOS expression decreased and was very weakat E21 and at P0. These observations may imply a role ofNO in the establishment of callosal connections.
The present study describes the expression of nNOS inthe telencephalic vesicle during embryonic development.Neuronal NOS expression may be considered to be corre-lated with the production of NO and may thus provideindirect evidence for a role of NO in development. Duringthe embryonic stages, the role of NO is probably differentfrom that in the adult. This is suggested by the muchgreater expression of nNOS found in the cerebral cortexduring the embryonic stages than in later life. In thetelencephalic vesicle, the neuronal elements expressingnNOS are much more abundant in embryonic stages than
w xin neonates or adults 9,21,48 . Not only is the amount ofnNOS expressed in embryonic and adult life different, butthe distribution and general appearance of cells and fibers
w xexpressing nNOS are also different 53 . In the adultnervous system, the role of NO in physiological conditions
w xis that of a neurotransmitter or neuromodulator 15 andŽ . w xNO is involved in long-term potentiation LTP 2,22,29
Ž . w xand long-term depression LTD 8,36 . It also plays a rolew xin memory, especially spatial memory 66 . None of these
roles has been demonstrated in embryos. However, thelarge amount of nNOS in the embryonic telencephalicvesicle suggests an important function at that stage. Theresults presented here suggest that NO is expressed when-ever there is an activity related to maturational processes,i.e., the organization of cerebral lamination through itsinvolvement in migrational processes, and the regulation ofthe ingrowth of nerve fibers.
Acknowledgements
This study was supported by the Direccion General de´ŽInvestigacion Cientıfica y Tecnica grant DGICYT PM95-´ ´ ´
.0009-C02-01 .
References
w x1 S. Akbarian, W.E. Bunney Jr., S.G. Potkin, S.B. Wigal, J.O. Hag-man, C.A. Sandman, E.G. Jones, Altered distribution of nicotin-

( )M. Santacana et al.rDeÕelopmental Brain Research 111 1998 205–222 221
amide-adenine dinucleotide phosphate-diaphorase cells in frontallobe of schizophrenics implies disturbances of cortical development,
Ž .Arch. Gen. Psychiatry 50 1993 169–177.w x2 O. Arancio, M. Kiebler, C.J. Lee, V. Lev-Ram, R.Y. Tsien, E.R.
Kandel, R.D. Hawkins, Nitric oxide acts directly in the presynapticneuron to produce long-term potentiation in cultured hippocampal
Ž .neurons, Cell 87 1996 1025–1035.w x3 M.L. Arbones, J. Ribera, L. Agullo, M.A. Baltrons, A. Casanovas,
V. Riveros-Moreno, A. Garcia, Characteristics of nitric oxide syn-Ž .thase type I of rat cerebellar astrocytes, Glia 18 1996 224–232.
w x4 S. Bayer, Neurogenesis of the magnocellular basal telencephalicŽ .nuclei in the rat, Int. J. Dev. Neurosci. 5 1985 229–243.
w x5 S.A. Bayer, J. Altman, Development of layer I and the subplate inŽ .the rat neocortex, Exp. Neurol. 107 1990 48–62.
w x6 S. Bayer, J. Altman, Neocortical Development, Raven Press, NewYork, 1991, pp. 257–262.
w x7 Boulder Committee, Embryonic vertebrate central nervous system:Ž .revised terminology, Anat. Rec. 166 1970 257–262.
w x8 A.R. Boxal, J. Garthwaite, Long-term depression in rat cerebellumrequires both NO synthase and NO-sensitive guanylyl cyclase, Eur.
Ž .J. Neurosci. 8 1996 2209–2212.w x9 D.S. Bredt, S.H. Snyder, Transient nitric oxide synthase neurons in
embryonic cerebral cortical plate, sensory ganglia and olfactoryŽ .epithelium, Neuron 13 1994 301–313.
w x10 D.S. Bredt, S.H. Snyder, Nitric oxide: a physiologic messengerŽ .molecule, Annu. Rev. Biochem. 63 1994 175–195.
w x11 G. Bruning, NADPH-diaphorase histochemistry in the postnatalmouse cerebellum suggests specific developmental functions for
Ž .nitric oxide, J. Neurosci. Res. 36 1993 580–587.w x12 S.M. Catalano, R.T. Robertson, H.P. Killackey, Early ingrowth of
thalamocortical afferents to the neocortex of the prenatal rat, Proc.Ž .Natl. Acad. Sci. USA 88 1991 2999–3003.
w x13 K.S. Cramer, M. Sur, Activity-dependent remodeling of connectionsŽ .in the mammalian visual system, Curr. Opin. Neurobiol. 5 1995
106–111.w x14 K.S. Cramer, A. Angelucci, J.-O. Hahm, M.B. Bogdanov, M.A. Sur,
Role for nitric oxide in the development of the ferret retinogenicu-Ž .late projection, J. Neurosci. 16 1996 7995–8004.
w x15 T.M. Dawson, V.L. Dawson, Nitric oxide synthase: role as a neuro-transmitterrmediator in the brain and endocrine system, Annu. Rev.
Ž .Med. 47 1996 219–227.w x16 J.A. del Rio, B. Heimrich, V. Borrell, E. Forster, A. Drakew, S.¨
Alcantara, K. Nakajima, T. Miyata, M. Ogawa, K. Mikoshiba, P.Derer, M. Frotscher, E. Soriano, A role for Cajal–Retzius cells andreelin in the development of hippocampal connections, Nature 385Ž .1997 70–74.
w x17 C.R. Dermon, A. Stamataki, Laminar pattern of NADPH-diaphoraseŽ .activity in the developing avian cerebellum, Neuroreport 5 1994
1941–1945.w x18 L. Frazier-Cierpial, P.C. Brunjes, Early postnatal cellular prolifera-
tion and survival in the olfactory bulb and rostral migratory streamof normal and unilaterally odor deprived rats, J. Comp. Neurol. 289Ž .1989 481–492.
w x19 J.A. Gally, P.R. Montague, G.N. Reeke, G.M. Edelman, The NOhypothesis: possible effects of a short-lived, rapidly diffusible signalin the development and function of the nervous system, Proc. Natl.
Ž .Acad. Sci. 87 1990 3547–3551.w x20 M.A. Gates, L.B. Thomas, E.M. Howard, E.D. Laywell, B. Sajin, A.
Faissner, B. Gotz, J. Silver, D.A. Steindle, Cell and molecularanalysis of the developing and adult mouse subventricular zone of
Ž .the cerebral hemispheres, J. Comp. Neurol. 361 1995 249–266.w x21 G. Giuili, A. Luzi, M. Poyard, G. Guellaen, Expression of mouse
brain soluble guanylyl cyclase and NO synthase during ontogeny,Ž .Dev. Brain Res. 81 1994 269–283.
w x22 J.E. Haley, G.L. Wilcox, P.F. Chapman, The role of nitric oxide inŽ .hippocampal long-term potentiation, Neuron 8 1992 211–216.
w x23 R.D. Hawkins, M. Zhuo, O. Arancio, Nitric oxide and carbon
monoxide in hippocampal long-term potentiation, J. Neurobiol. 25Ž .1994 652–665.
w x24 D.T. Hess, S.I. Patterson, J.H.P. Skene, Neuronal growth conecollapse and inhibition of protein fatty acylation by nitric oxide,
Ž .Nature 366 1993 562–565.w x25 Y. Higuchi, H. Hattori, R. Hattori, K. Furusho, Increased neurons
containing neuronal nitric oxide synthase in the brain of a hypoxic–Ž .ischemic neonatal rat model, Brain Dev. 18 1996 369–375.
w x26 C. Iadecola, Bright and dark sides of nitric oxide in ischemic brainŽ .damage, Trends Neurosci. 20 1997 132–139.
w x27 L.J. Ignarro, G.M. Buga, K.S. Wood, R.E. Byrns, G. Chaudhuri,Endothelium-derived relaxing factor produced and released from
Ž .artery and vein is nitric oxide, Proc. Natl. Acad. Sci. USA 84 19879265–9269.
w x28 G.O. Ivy, H.P. Killakey, The ontogeny of the distribution of callosalprojection neurons in the rat parietal cortex, J. Comp. Neurol. 195Ž .1981 367–389.
w x29 Y. Izumi, D.B. Clifford, C.F. Zorumski, Inhibition of long-termpotentiation by NMDA-mediated nitric oxide release, Science 257Ž .1992 1273–1276.
w x30 A. Kalsbeek, P. Voorn, R.M. Buijs, C.W. Pool, H.B.M. Uylings,Development of monoaminergic innervation in the prefrontal cortex
Ž .of the rat, J. Comp. Neurol. 269 1988 58–72.w x31 G. Keilhoff, B. Seidel, H. Noack, W. Tischmeyer, D. Stanek, G.
Wolf, Patterns of nitric oxide synthase at the messenger RNA andprotein levels during early rat brain development, Neuroscience 75Ž .1996 1193–1201.
w x32 K. Kishi, J.Y. Peng, S. Kakuta, K. Murakami, M. Kuroda, S.Yokota, S. Hayakawa, T. Kuge, T. Asamaya, Migration of bipolarsubependymal cells, precursors of the granule cells of the rat olfac-tory bulb, with reference to the arrangement of the radial glial fibers,
Ž .Arch. Histol. Cytol. 53 1990 219–226.w x33 R.G. Knowles, S. Moncada, Nitric oxide synthase in mammals,
Ž .Biochem. J. 298 1994 249–258.w x34 S. Koh, R. Loy, Localization and development of nerve growth
factor-sensitive rat basal forebrain neurons and their afferent projec-Ž .tions to hippocampus and neocortex, J. Neurosci. 9 1989 2999–
3018.w x35 P. Kugler, D. Drenckhahn, Astrocytes and Bergman glia as an
Ž .important site of nitric oxide synthase I, Glia 16 1996 165–173.w x36 D.J. Linden, Long-term synaptic depression in the mammalian brain,
Ž .Neuron 12 1994 457–472.w x37 R.D. Lund, M.J. Mustari, Development of the geniculocortical path-
Ž .way in rats, J. Comp. Neurol. 173 1977 289–305.w x38 M.B. Luskin, Restricted proliferation and migration of postnatally
generated neurons derived from the forebrain subventricular zone,Ž .Neuron 11 1993 173–189.
w x39 L. Ma, Y. Ishisaki, I. Morita, S. Murota, Presence of nitric oxidesynthase activity in the neurons of the rat embryonal cerebrum,
Ž .Neurosci. Lett. 132 1991 23–25.w x40 M. Marin-Padilla, T.M. Marin-Padilla, Origin, prenatal development
Ž .and structural organization of layer I of the human cerebral motorŽ .cortex. A Golgi study, Anat. Embryol. 164 1982 161–206.
w x41 M. Marin-Padilla, Ontogenesis of the pyramidal cell of the mam-malian neocortex and developmental cytoarchitectonics: a unifying
Ž .theory, J. Comp. Neurol. 321 1992 223–240.w x42 E. Meisami, S. Hamedi, Relative contribution of brain and periph-
eral connections to postnatal growth and cell accretion in the ratŽ .olfactory bulb, Dev. Brain Res. 29 1986 282–286.
w x43 G. Meyer, J.M. Soria, J.R. Martinez-Galan, B. Martin-Clemente, A.Fairen, Different origins and developmental histories of transientneurons in the marginal zone of the fetal and neonatal cortex of the
Ž .rat, J. Comp. Neurol. 397 1998 493–518.w x44 G. Meyer, T. Gonzalez-Hernandez, Developmental changes in layer
I of the human neocortex during prenatal life: a DiI-tracing andAChE and NADPH-d histochemistry study, J. Comp. Neurol. 338Ž .1993 317–336.

( )M. Santacana et al.rDeÕelopmental Brain Research 111 1998 205–222222
w x45 R.R. Mize, F.T. Banfro, C.A. Scheine, Pre- and postnatal expressionof amino acid neurotransmitters, calcium binding proteins, and nitricoxide synthase in the developing superior colliculus, Prog. Brain
Ž .Res. 108 1996 313–332.w x46 S. Moncada, R.M. Palmer, E.A. Higgs, Nitric oxide: physiology,
Ž .pathophysiology and pharmacology, Pharmacol. Rev. 43 1991109–142.
w x47 M.A. Moro, J. de Alba, J.C. Leza, L. Bosca, A.P. Fernandez, M.L.Bentura, J. Rodrigo, I. Lizasoain, Neuronal expression of type IInitric oxide synthase in an ex vivo model of brain ischemia, Eur. J.
Ž .Neurosci. 10 1998 445–456.w x48 F.J. Northington, R.C. Koehler, R.J. Traystman, L.J. Martin, Nitric
oxide synthase 1 and nitric oxide synthase 3 protein expression isregionally and temporally regulated in fetal brain, Dev. Brain Res.
Ž .95 1996 1–14.w x49 M. Ogawa, T. Miyata, K. Nakajima, K. Yagyu, M. Seike, K.
Ikenaka, H. Yamamoto, K. Mikoshiba, The reeler gene-associatedantigen on Cajal–Retzius neurons is a crucial molecule for laminar
Ž .organization of cortical neurons, Neuron 14 1995 899–912.w x50 O. Palluy, M. Rigaud, Nitric oxide induces cultured cortical neuron
Ž .apoptosis, Neurosci. Lett. 208 1996 1–4.w x51 R.M. Palmer, A.G. Ferrige, S. Moncada, Nitric oxide release ac-
counts for the biological activity of endothelium-derived relaxingŽ .factor, Nature 327 1987 524–526.
w x52 J.G. Parnavelas, G.C. Papadopoulos, M.E. Cavanagh, Changes inneurotransmitters during development, in: A. Peters, E.G. JonesŽ .Eds. , Cerebral Cortex, Vol. 7, Development and Maturation ofCerebral Cortex, Plenum, New York, 1988, pp. 177–209.
w x53 J. Rodrigo, D.R. Springall, O. Uttenthal, M.L. Bentura, F. Abadia-Molina, V. Riveros-Moreno, R. Martinez-Murillo, J. Polak, S. Mon-cada, Localization of nitric oxide synthase in the adult rat brain,
Ž .Philos. Trans. R. Soc. Lond. B 345 1994 175–221.w x54 A.J. Roskams, D.S. Bredt, T.M. Dawson, G.V. Ronnett, Nitric oxide
mediates the formation of synaptic connections in developing andŽ .regenerating olfactory receptor neurons, Neuron 13 1994 289–299.
w x55 A.F. Samdani, T.M. Dawson, V.L. Dawson, Nitric oxide in modelsŽ .of focal ischemia, Stroke 28 1997 1283–1288.
w x56 M. Santacana, A.G. de la Vega, M. Heredia, Peripherin fibers in themain olfactory bulb are different from olfactory fibers and fromLHRH fibers: an immunocytochemical and DiI study, Brain Res.
Ž .734 1996 337–340.w x57 K. Schilling, H.H. Schmidt, S.L. Baader, Nitric oxide synthase
expression reveals compartments of cerebellar granule cells andsuggests a role for mossy fibers in their development, Neuroscience
Ž .59 1994 893–903.w x58 S.H. Snyder, Nitric oxide, first in a new class of neurotransmitters,
Ž .Science 257 1992 494–496.w x59 D.R. Springall, V. Riveros-Moreno, L. Buttery, A. Suburo, A.E.
Bishop, M. Merret, S. Moncada, J.M. Polak, Immunological detec-Ž .tion of nitric oxide synthase s in human tissues using heterologous
Ž .antibodies suggesting different isoforms, Histochemistry 98 1992259–266.
w x60 M. Tanaka, S. Yoshida, M. Yano, F. Hanaoka, Roles of endogenousnitric oxide in cerebellar cortical development in slice cultures,
Ž .Neuroreport 5 1994 2049–2052.w x61 H. Togashi, M. Sasaki, E. Frohman, E. Taira, R.R. Ratan, T.M.
Ž .Dawson, V.L. Dawson, Neural type I nitric oxide synthase regu-Ž .lates nuclear factor kB activity and immunologic type II nitric
Ž .oxide synthase expression, Proc. Natl. Acad. Sci. USA 94 19972676–2680.
w x62 L.O. Uttenthal, D. Alonso, A.P. Fernandez, R.O. Campbell, M.A.Moro, J.C. Leza, I. Lizasoain, F.J. Esteban, J.B. Barroso, R. Valder-rama, J.A. Pedrosa, M.A. Peinado, J. Serrano, A. Richart, M.L.Bentura, M. Santacana, R. Martinez-Murillo, J. Rodrigo, Neuronaland inducible nitric oxide synthases and nitrotyrosine immunoreac-
Žtivities in the cerebral cortex of aging rats, Microsc. Res. Tech. in.press .
w x63 K.L. Valentino, E.G. Jones, The early formation of the corpuscallosum: a light and electron microscope study in foetal and
Ž .neonatal rats, J. Neurocytol. 11 1982 583–609.w x64 F. Valverde, M.V. Facal-Valverde, M. Santacana, H. Heredia, De-
velopment and differentiation of early generated cells of sublayerVIb in the somatosensory cortex of the rat: a correlated Golgi and
Ž .autoradiographic study, J. Comp. Neurol. 290 1989 118–140.w x65 C.V. Williams, D. Nordquist, S.C. McLoon, Correlation of nitric
oxide synthase expression with changing patterns of axonal projec-Ž .tions in the developing visual system, J. Neurosci. 14 1994 1746–
1755.w x66 M.A. Wilson, S. Tonegawa, Synaptic plasticity, place cells and
spatial memory: study with second generation knockouts, TrendsŽ .Neurosci. 20 1997 102–106.
w x67 S.P. Wise, E.G. Jones, Developmental studies of thalamocortical andcommissural connections in the rat somatic sensory cortex, J. Comp.
Ž .Neurol. 178 1978 187–208.w x68 W. Wu, F.J. Liuzzi, F.P. Schinco, A.S. Depto, Y. Li, J.A. Mong,
T.M. Dawson, S.H. Snyder, Neuronal nitric oxide synthase is in-Ž .duced in spinal neurons by traumatic injury, Neuroscience 61 1994
719–726.w x69 X.X. Yan, L.J. Garey, L.S. Jen, Prenatal development of NADPH-
diaphorase-reactive neurons in human frontal cortex, Cereb. CortexŽ .6 1996 737–745.
w x70 G.M. Young, S.W. Levison, Persistence of multipotential progeni-Ž .tors in the juvenile rat subventricular zone, Dev. Neurosci. 18 1996
255–265.w x71 H.Y. Yun, V.L. Dawson, T.M. Dawson, Nitric oxide in health and
Ž .disease of the nervous system, Mol. Psychiatr. 2 1997 300–310.w x72 Z.G. Zhang, M. Chopp, S. Gautman, C. Zaloga, R.L. Zhang,
H.H.H.W. Schmidt, J.S. Pollock, U. Forstermann, Upregulation ofneuronal nitric oxide synthase and mRNA, and selective sparing ofnitric synthase-containing neurons after focal cerebral ischemia in
Ž .rat, Brain Res. 654 1994 85–95.




















