Expression and Developmental Regulation of the Hydra-RFamide and Hydra-LWamide Preprohormone Genes...
15
Expression and Developmental Regulation of the Hydra-RFamide and Hydra-LWamide Preprohormone Genes in Hydra: Evidence for Transient Phases of Head Formation Christian Mitgutsch, Frank Hauser, and Cornelis J. P. Grimmelikhuijzen 1 Department of Cell Biology, Zoological Institute, University of Copenhagen, Universitetsparken 15, DK-2100 Copenhagen Ø, Denmark Hydra magnipapillata has three distinct genes coding for preprohormones A, B, and C, each yielding a characteristic set of Hydra-RFamide (Arg-Phe-NH 2 ) neuropeptides, and a fourth gene coding for a preprohormone that yields various Hydra-LWamide (Leu-Trp-NH 2 ) neuropeptides. Using a whole-mount double-labeling in situ hybridization technique, we found that each of the four genes is specifically expressed in a different subset of neurons in the ectoderm of adult Hydra. The preprohormone A gene is expressed in neurons of the tentacles, hypostome (a region between tentacles and mouth opening), upper gastric region, and peduncle (an area just above the foot). The preprohormone B gene is exclusively expressed in neurons of the hypostome, whereas the preprohormone C gene is exclusively expressed in neurons of the tentacles. The Hydra-LWamide preprohormone gene is expressed in neurons located in all parts of Hydra with maxima in tentacles, hypostome, and basal disk (foot). Studies on animals regenerating a head showed that the prepro-Hydra-LWamide gene is expressed first, followed by the preprohormone A and subsequently the preprohormone C and the preprohormone B genes. This sequence of events could be explained by a model based on positional values in a morphogen gradient. Our head-regeneration experiments also give support for transient phases of head formation: first tentacle-specific preprohormone C neurons (frequently associated with a small tentacle bud) appear at the center of the regenerating tip, which they are then replaced by hypostome-specific preprohormone B neurons. Thus, the regenerating tip first attains a tentacle-like appearance and only later this tip develops into a hypostome. In a developing bud of Hydra, tentacle-specific preprohormone C neurons and hypostome-specific preprohormone B neurons appear about simultaneously in their correct positions, but during a later phase of head development, additional tentacle-specific preprohormone C neurons appear as a ring at the center of the hypostome and then disappear again. Nerve-free Hydra consisting of only epithelial cells do not express the preprohormone A, B, or C or the LWamide preprohormone genes. These animals, however, have a normal phenotype, showing that the preprohormone A, B, and C and the LWamide genes are not essential for the basic pattern formation of Hydra. © 1999 Academic Press Key Words: Hydra; development; pattern formation; neuropeptide gene expression. INTRODUCTION Cnidarians are primitive animals such as sea anemones, corals, and jellyfishes and are closely related to the first metazoans that evolved a nervous system. The primitive nervous systems of cnidarians produce large amounts of a variety of neuropeptides (for a review see Grimmelikhuij- zen et al., 1996). These neuropeptides may function as neurotransmitters or neuromodulators, but also as neuro- hormones, controlling developmental processes such as morphogenesis (Grimmelikhuijzen et al., 1992, 1996; Leitz et al., 1994; Takahashi et al., 1997). Of all cnidarians, Hydra is the most widely used model to study development. Recently, we have isolated four peptides from Hydra that all have the C-terminal sequence Arg-Phe-NH 2 (RFamide) in common (Moosler et al., 1996). Using immunocyto- chemistry and immunoelectron microscopy, we have shown that these RFamide peptides are produced by neu- rons in Hydra and that they are localized in neuronal 1 To whom correspondence should be addressed. Fax: 0045- 35321200. E-mail: [email protected]. Developmental Biology 207, 189 –203 (1999) Article ID dbio.1998.9150, available online at http://www.idealibrary.com on 0012-1606/99 $30.00 Copyright © 1999 by Academic Press All rights of reproduction in any form reserved. 189
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Expression and Developmental Regulation of the Hydra-RFamide and Hydra-LWamide Preprohormone Genes...

(
Developmental Biology 207, 189–203 (1999)Article ID dbio.1998.9150, available online at http://www.idealibrary.com on
Expression and Developmental Regulation of theHydra-RFamide and Hydra-LWamide PreprohormoneGenes in Hydra: Evidence for TransientPhases of Head Formation
Christian Mitgutsch, Frank Hauser, andCornelis J. P. Grimmelikhuijzen1
Department of Cell Biology, Zoological Institute, University of Copenhagen,Universitetsparken 15, DK-2100 Copenhagen Ø, Denmark
Hydra magnipapillata has three distinct genes coding for preprohormones A, B, and C, each yielding a characteristic set ofHydra-RFamide (Arg-Phe-NH2) neuropeptides, and a fourth gene coding for a preprohormone that yields various Hydra-LWamideLeu-Trp-NH2) neuropeptides. Using a whole-mount double-labeling in situ hybridization technique, we found that each of thefour genes is specifically expressed in a different subset of neurons in the ectoderm of adult Hydra. The preprohormone A geneis expressed in neurons of the tentacles, hypostome (a region between tentacles and mouth opening), upper gastric region, andpeduncle (an area just above the foot). The preprohormone B gene is exclusively expressed in neurons of the hypostome, whereasthe preprohormone C gene is exclusively expressed in neurons of the tentacles. The Hydra-LWamide preprohormone gene isexpressed in neurons located in all parts of Hydra with maxima in tentacles, hypostome, and basal disk (foot). Studies on animalsregenerating a head showed that the prepro-Hydra-LWamide gene is expressed first, followed by the preprohormone A andsubsequently the preprohormone C and the preprohormone B genes. This sequence of events could be explained by a model basedon positional values in a morphogen gradient. Our head-regeneration experiments also give support for transient phases of headformation: first tentacle-specific preprohormone C neurons (frequently associated with a small tentacle bud) appear at the centerof the regenerating tip, which they are then replaced by hypostome-specific preprohormone B neurons. Thus, the regenerating tipfirst attains a tentacle-like appearance and only later this tip develops into a hypostome. In a developing bud of Hydra,tentacle-specific preprohormone C neurons and hypostome-specific preprohormone B neurons appear about simultaneously intheir correct positions, but during a later phase of head development, additional tentacle-specific preprohormone C neuronsappear as a ring at the center of the hypostome and then disappear again. Nerve-free Hydra consisting of only epithelial cells donot express the preprohormone A, B, or C or the LWamide preprohormone genes. These animals, however, have a normalphenotype, showing that the preprohormone A, B, and C and the LWamide genes are not essential for the basic pattern formationof Hydra. © 1999 Academic Press
Key Words: Hydra; development; pattern formation; neuropeptide gene expression.
nhmeiRai
INTRODUCTION
Cnidarians are primitive animals such as sea anemones,corals, and jellyfishes and are closely related to the firstmetazoans that evolved a nervous system. The primitivenervous systems of cnidarians produce large amounts of avariety of neuropeptides (for a review see Grimmelikhuij-zen et al., 1996). These neuropeptides may function as
1 To whom correspondence should be addressed. Fax: 0045-
35321200. E-mail: [email protected].0012-1606/99 $30.00Copyright © 1999 by Academic PressAll rights of reproduction in any form reserved.
eurotransmitters or neuromodulators, but also as neuro-ormones, controlling developmental processes such asorphogenesis (Grimmelikhuijzen et al., 1992, 1996; Leitz
t al., 1994; Takahashi et al., 1997). Of all cnidarians, Hydras the most widely used model to study development.ecently, we have isolated four peptides from Hydra thatll have the C-terminal sequence Arg-Phe-NH2 (RFamide)n common (Moosler et al., 1996). Using immunocyto-
chemistry and immunoelectron microscopy, we haveshown that these RFamide peptides are produced by neu-
rons in Hydra and that they are localized in neuronal189

aHfHmnA
GeqmRsfon
csq
T
c
fcgo
tcqm
l((
(
190 Mitgutsch, Hauser, and Grimmelikhuijzen
dense-cored vesicles (Grimmelikhuijzen, 1985; Koizumi etl., 1989). We have also cloned the preprohormones for theydra-RFamides and found that three different genes code
or three different preprohormones, each containing severalydra-RFamide variants (Darmer et al., 1998). Preprohor-one A contains single unprocessed copies of the isolated
europeptides Hydra-RFamide I (,Glu-Trp-Leu-Gly-Gly-rg-Phe-NH2), Hydra-RFamide II (,Glu-Trp-Phe-Asn-Gly-
Arg-Phe-NH2), Hydra-RFamide III/IV (Lys-Pro-His-Leu-Arg-ly-Arg-Phe-NH2/His-Leu-Arg-Gly-Arg-Phe-NH2), and sev-
ral other, not yet identified, putative neuropeptide se-uences (Fig. 1A). Preprohormone B resembles preprohor-one A and contains single unprocessed copies of Hydra-Famide I and II and various other, putative neuropeptideequences (Fig. 1B). Preprohormone C is rather differentrom the other two preprohormones and contains one copyf unprocessed Hydra-RFamide I and seven other, putativeeuropeptide sequences (Fig. 1C).In addition to the RFamide preprohormones, we have also
loned a preprohormone from Hydra that gives rise to aeries of putative neuropeptides with the C-terminal se-uence Leu-Trp-NH2 (LWamide; Leviev et al., 1997; Fig.
1D). Independent of our own cloning work, five of theseputative neuropeptides have recently been isolated andsequenced from Hydra extracts (Takahashi et al., 1997).
FIG. 1. Schematic representation of four preprohormones in Hy-dra. (A) Preprohormone A, containing a signal sequence (black box),one unprocessed Hydra-RFamide I (hatched and marked by 1),Hydra-RFamide II (2), Hydra-RFamide III/IV (3, 4), and severalputative neuropeptide sequences (hatched, without numbers). (B)Preprohormone B, containing a signal sequence (black) and oneunprocessed sequence of Hydra-RFamide I (1), Hydra-RFamide II(2), and several putative neuropeptide sequences (hatched, withoutnumbers). (C) Preprohormone C, containing a signal sequence(black), one copy of unprocessed Hydra-RFamide I (1), and sevenputative neuropeptide sequences (hatched, without numbers). (D)The Hydra-LWamide precursor, containing a signal sequence(black), unprocessed sequences of the isolated peptides Hydra-LWamide I–V (hatched and marked by 5–9), and three putativeneuropeptide sequences (hatched, without numbers). The data ofA–C are from Darmer et al. (1998), those of D from Leviev et al.1997) and Takahashi et al. (1997).
he LWamide peptides from Hydra and other cnidarian h
Copyright © 1999 by Academic Press. All right
species induce metamorphosis in planula larvae from themarine hydroid Hydractinia echinata, but also stimulatemuscle contraction (Leitz et al., 1994; Takahashi et al.,1997).
It is currently not known whether the genes coding forthe Hydra-RFamide preprohormones A, B, and C and theHydra-LWamide preprohormone are expressed in the sameor in different sets of neurons and it is also unknown howthese genes are regulated during developmental processessuch as head regeneration and bud development. Further-more, it is not known to what extent these genes, via theirneuropeptide products, control basic pattern formation inHydra. All these questions are addressed in the presentpaper.
MATERIALS AND METHODS
Animals
Hydra magnipapillata strain 105 (isolated by Drs. T. Sugiyamaand T. Fujisawa, National Institute of Genetics, Mishima, Japan)was cultured as described earlier (Leviev et al., 1997). Nerve-free,epithelial H. magnipapillata were obtained in two ways. First, achimera (A10; a kind gift from Dr. T. Fujisawa) that consisted ofepithelial cells from H. magnipapillata strain 105 and I cells froma temperature-sensitive mutant (sf-1; Sugiyama and Fujisawa,1978) was used. At normal culture temperature (20°C), the chimerahas the cell composition of a normal Hydra, but at elevatedtemperatures, it loses its I cells and all their differentiation prod-ucts (such as nerve cells and nematocytes) and turns into anerve-free, epithelial Hydra consisting only of wild-type strain 105epithelial cells. This chimera was kept at 28°C for 10 days in Hydramedium (Leviev et al., 1997) supplemented with 50 mg/L rifampi-in and 50 mg/L kanamycin without feeding. The animals were
subsequently returned to 20°C and force-fed during 2 months withnauplii of Artemia salina (which had been killed and disrupted byreezing), using a glass capillary. Six hours after feeding, the gastricavities of the animals were rinsed using a fine water jet from alass capillary. In the course of these 2 months, the third generationf buds became fully nerve-free (as determined by in situ hybrid-
ization, see below).The second method of obtaining nerve-free Hydra consisted of a
reatment of wild-type H. magnipapillata with 0.4–0.8% colchi-ine (Sigma) for 7 h (Campbell, 1976). The animals were subse-uently washed and kept in Hydra medium supplemented with 50g/L rifampicin and 50 mg/L kanamycin for 10 days without
feeding, but with a regular change of culture medium. The animalswere then force-fed as described for the chimerae.
For regeneration experiments, we used Hydra without budsstarved for 1 day. They were transversely cut, either just below thetentacle zone or in the middle of the body column, using a finepiece of sharp razor blade. After cutting they were transferred tofresh Hydra medium.
In Situ Hybridization
Whole-mount in situ hybridizations using single, digoxigenin-abeled cRNA probes were carried out as described by Grens et al.1996) with the following slight modifications. pBluescript vectorsStratagene) containing cDNA insertions coding for either prepro-
ormone A (nucleotide positions 2–537 of Fig. 1 from Darmer et al.,s of reproduction in any form reserved.

L
ias
gbhMwtlceSfgsbcd
191Developmental Expression of Hydra Neuropeptide Genes
1998), preprohormone B (nucleotide positions 146–614 of Fig. 2from Darmer et al., 1998), preprohormone C (nucleotide positions1–1303 of Fig. 3 from Darmer et al., 1998), or prepro-Hydra-
Wamide (nucleotide positions 765–1377 from Leviev et al., 1997)were used. These vectors were linearized by cleavage at a suitableposition. Sense or antisense in vitro transcripts were obtained usingT3 or T7 RNA polymerase and the DIG RNA Labeling Kit(Boehringer Mannheim). Hybridization was performed for 3 days at55°C in 50% formamide, 53 SSC (13 SSC is 150 mM NaCl, 15 mMsodium citrate, pH 7.0), 0.1% Tween 20, 0.1% Chaps {3-[(3-cholamidopropyl)dimethylammonio]-1-propane sulfonate; Sigma},13 Denhardt’s solution (13 Denhardt’s solution is 0.02% bovineserum albumin, 0.02% polyvinylpyrrolidone, 0.02% Ficoll), 0.01%heparin, and 0.2 mg/ml labeled RNA probe. Staining was carried outn the dark using Fab fragments of a monoclonal digoxigeninntibody coupled to alkaline phosphatase and BM purple as aubstrate (Boehringer Mannheim).
For the simultaneous visualization of the expression of twoenes, we also synthesized RNA probes that were labeled withiotin, using the Biotin RNA Labeling Mix (Boehringer Mann-eim), or with fluorescein, using the Fluorescein RNA Labelingix (Boehringer Mannheim). The hybridization was carried outith a 1:1 mixture of two differently labeled probes, corresponding
o two different preprohormone genes. Staining of the digoxigenin-abeled probe was carried out using the digoxigenin antibody-oupled alkaline phosphatase described above, using as substrateither BM purple (Boehringer Mannheim) or Fast Red (Sigma).taining of the fluorescein-labeled probe was performed using Fabragments of a monoclonal fluorescein antibody that were conju-ated to alkaline phosphatase (Boehringer Mannheim). As a sub-trate, either BM purple or Fast Red was used. Staining of theiotin-labeled probe was performed using Fab fragments of a mono-lonal biotin antibody conjugated to peroxidase andiaminobenzidine/H2O2 as substrate (Boehringer Mannheim). For
staining of samples that were double-hybridized with biotin anddigoxigenin probes, we added a mixture of the peroxidase-coupledbiotin antibody and phosphatase-coupled digoxigenin antibody,incubated and washed the sample as described by Grens et al.(1996), and then stained it with one of the phosphatase substrates(BM purple or Fast Red) as mentioned above. The phosphatasereaction was stopped by washing away the substrate, after whichthe buffer was changed for that necessary for the peroxidasereaction (DAB substrate; Boehringer Mannheim). For staining ofsamples that were double-hybridized with digoxigenin and fluores-cein probes, we first incubated them with a phosphatase-coupleddigoxigenin antibody and carried out one phosphatase reactionusing one of the phosphatase substrates (BM purple or Fast Red).We stopped this reaction with 100 mM glycine–HCl, pH 2.2/0.1%Tween 20 (Hauptmann and Gerster, 1994), washed the sample, andthen incubated it with the phosphatase-coupled fluorescein anti-body followed by the second phosphatase reaction using the othersubstrate (or vice versa).
Because the probe for preprohormone A mRNA slightly crossre-acts with preprohormone B mRNA, and the preprohormone B probewith preprohormone A mRNA (Darmer et al., 1998), we added thetwo differentially labeled probes together in one hybridizationreaction, but only stained for one of them. In this way, we alwaysobtained a specific preprohormone B staining (because the prepro-hormone A mRNA was competed away by the unstained prepro-hormone A mRNA probe) or, vice versa, a specific preprohormoneA staining (because the preprohormone B mRNA was competedaway by the unstained preprohormone B mRNA probe).
Stained animals were mounted either in Euparal or in buffered
Copyright © 1999 by Academic Press. All right
glycerol (after Fast Red staining) and examined and photographedusing a Leica DMRB microscope.
RESULTS
Expression of the Four Preprohormone Genes inAdult Hydra
Using digoxigenin-labeled antisense RNA probes corre-sponding to the whole cDNA coding for the Hydra-RFamidepreprohormone A, we found hybridization in neurons of theectoderm of the peduncle (a region just above the foot),upper gastric region (just below the tentacles), hypostome (aregion between tentacles and mouth opening), and tentacles(Fig. 2A). Hybridization was clearly in neurons, but it couldnot always be excluded that other cell types, for exampleepithelial cells, were also stained. By changing the labelingor staining methods, however, it became clear that thehybridization was exclusively in neurons (Figs. 3E and 4A).
Using digoxigenin-labeled antisense RNA probes corre-sponding to preprohormone B cDNA, we found exclusivestaining in the ectoderm of the hypostome (Fig. 2B). Thisstaining was caused by neurons that were mostly elongatedin shape and projected toward the outer surface of theectoderm and, thus, clearly were sensory neurons. Bychanging the labeling or staining methods, it became clearthat the hybridization was confined to neurons and that noother cell types were stained (Fig. 4B).
With antisense probes corresponding to preprohormone CcDNA, we found exclusive staining of neurons in thetentacles (Fig. 2C) with a sharp border of preprohormone Cexpression between tentacle bases and hypostome.
When we used an antisense probe corresponding to theHydra-LWamide precursor cDNA, we obtained strong hy-bridization in neurons of the tentacles, hypostome, gastricregion, and pedal disk (foot) (Fig. 2D) with clear maxima ofneuronal cell densities in head (tentacles and hypostome)and pedal disk. However, in contrast to the other threepreprohormone genes, we found that Hydra-LWamide geneexpression in the midgastric area and in very early budsstrongly varied. In some batches of animals, we foundstrong staining in these regions, whereas in other batchesthese regions were virtually devoid of hybridizing neurons.The reason for these differences is unclear. For head regen-eration and other developmental experiments (see below),we always analyzed batches of animals that initially werefree of hybridizing neurons in the midgastric region.
When we used sense instead of antisense probes in our insitu hybridizations, or when we used no probes at all, nostaining was observed.
Different Sets of Neurons Express the FourPreprohormone Genes
To find out whether the same or different neurons ex-press the four preprohormone genes, we carried out double-labeling experiments, using digoxigenin-labeled RNA
probes corresponding to one preprohormone cDNA ands of reproduction in any form reserved.

192

tobwcan(dddo
Fssnnciroshabaccpm
trrTft
y
193Developmental Expression of Hydra Neuropeptide Genes
either biotin-labeled or fluorescein-labeled RNA probescorresponding to another preprohormone cDNA. In thisway we found that the four preprohormone genes wereexpressed in four different sets of neurons. Examples of thisdouble-labeling in the tentacles are given in Figs. 3A–3D.Figure 4 gives examples of whole-mount double-labelingexperiments in other regions of Hydra.
The cDNA probes coding for the preprohormones A and Bslightly crossreact (Darmer et al., 1998), giving a potentialproblem with the identification of neurons expressing thepreprohormone A or B gene. For in situ hybridizations usingsingle staining, however, we could circumvent this problemby adding the two differentially labeled probes together, butstaining only one of them. In this way, we always obtaineda specific staining for preprohormone B mRNA, because thepreprohormone A mRNA was competed away by the un-stained preprohormone A mRNA probe or vice versa (seeMaterials and Methods). Using this technique, we foundthat the preprohormone B gene was exclusively expressedin a specific set of neurons in the hypostome, whereas thepreprohormone A gene was expressed in a specific set ofneurons in the peduncle, upper gastric region, tentacles,and, again, hypostome (see above). Therefore, the onlyneurons in which the preprohormone A and B genes couldbe coexpressed would be neurons of the hypostome. Here,however, we could not decide, using our double-labelingtechnique with both cRNA probes visualized, whether thepreprohormone A and B genes were expressed in the sameor in different sets of neurons.
Expression of the Four Preprohormone Genes inDeveloping Buds
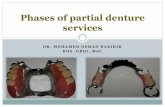
In developing buds, the four preprohormone genes areexpressed in a fixed, temporal pattern. First, neurons con-taining mRNAs for the Hydra-LWamide preprohormone(“LWamide neurons”) emerge at the tip of a very young bud,followed by neurons expressing the preprohormone A gene(“A neurons”; Fig. 5A). Then, at about the same time,neurons expressing the preprohormone B gene (“B neu-rons”) and those expressing the preprohormone C gene (“Cneurons”) appear. The B neurons originate in a disk-likearea located around the center of the tip of the developingbud (Figs. 5D and 7E), whereas the C neurons appear at theperiphery of the bud tip, at positions at which later the
FIG. 2. Whole-mount in situ hybridization of Hydra with cRHydra-LWamide preprohormone. (A) Hybridization with the preprhypostome, tentacles, upper gastric region, and peduncle. The animnot yet expressed the preprohormone A gene (365). (B) Hybridizaneurons in the ectoderm of the hypostome (370). (C) Staining withthe ectoderm of the tentacles. Note staining in the tentacles of twoStaining is seen in ectodermal neurons occurring all over the Hydroung developing bud at the lower right side that is heavily staine
Copyright © 1999 by Academic Press. All right
entacles emerge (Figs. 6A and 7A). At a later stage, clustersf C neurons form, each associated with a small tentacleud located at the margin of the Hydra bud tip (Fig. 7B),hile the B neurons increase in number and occupy the
enter of the tip (Fig. 7F). Subsequently, when the tentaclesnd associated C neurons have become more prominent, aew ring of C neurons originates in the center of the bud tip
Figs. 3F, 6B, and 7C). This central ring of C neuronsisappears again when the bud grows older (Fig. 7D). Lessramatic changes occur with the B neurons during final budevelopment (Figs. 5E, 7G, and 7H), since they graduallyccupy the positions that we know from adult animals.
Expression of the Four Preprohormone Genes inHydra Regenerating a Head
To see whether the same temporal pattern of neuropep-tide gene expression and tentacle development also can beobserved in other developmental processes involving headformation, we have investigated Hydra regenerating a head.or this purpose, Hydra heads were cut off by a transverseection just below the tentacle zone. By this, all tentacle-pecific C neurons were removed and the reoccurrence of Ceurons and tentacle buds was subsequently monitored. Ceurons appear at the head-regenerating tip about 20 h afterutting (Fig. 8A). They then increase in number and spreadn a rather irregular fashion over the apical surface of theegenerating head (Figs. 8B and 8C). Subsequently, clustersf C neurons form: one central cluster at the tip and five orix peripheral clusters at the basis of the area regenerating aead (Figs. 8D and 8E). At this stage, all C neuron clustersre very regularly spaced, having about the same distanceetween each other. Furthermore, small tentacle buds haveppeared both in the peripheral and in the central C neuronlusters (Fig. 8D). Subsequently, the central C neuronluster and associated tentacle bud disappear, while theeripheral C neuron clusters and tentacle buds becomeore prominent and develop into tentacles (Fig. 8E and 8F).Because the regenerates described above still contained
he “old” A and LWamide neurons of their upper gastricegion, no conclusion could be drawn regarding the tempo-al appearance of these neurons in the regenerating heads.o obtain a more complete picture of the reappearance of all
our neuronal cell types, we have cut Hydra in the middle ofheir body columns and followed the regeneration of heads
probes coding for the preprohormones A, B, and C and themone A probe. Staining occurs in neurons in the ectoderm of thedeveloping two buds at the lower right and left sides, which havewith the preprohormone B probe. Staining occurs exclusively inpreprohormone C probe. Staining occurs exclusively in neurons inr buds (340). (D) Staining with the prepro-Hydra-LWamide probe.y with maxima in tentacles, hypostome, and pedal disk. Note theits tip (365).
NAohoral is
tiontheolde
a bodd at
s of reproduction in any form reserved.

194 Mitgutsch, Hauser, and Grimmelikhuijzen
Copyright © 1999 by Academic Press. All rights of reproduction in any form reserved.

ati
apTp( 314
pLdt(s
195Developmental Expression of Hydra Neuropeptide Genes
in the lower body column parts. We found that first theLWamide neurons emerge at the tip of the regeneratinghead, followed by A, C, and then B neurons. We also found
FIG. 4. Simultaneous visualization of the expression of two preprohdult Hydra and a connecting young bud. The preprohormone A geeduncle of the adult Hydra and in ectodermal neurons of the hypostohe preprohormone C gene (blue) is exclusively expressed in the tentreprohormone B gene (brown) is exclusively expressed in neurons loca
red) is expressed in neurons of the tentacles (somewhat out of focus;
FIG. 3. Simultaneous visualization of the expression of two prereprohormone A (red) and C (blue) genes in different neurons of tWamide (brown) gene in different neurons of the tentacles (3350ifferent neurons of the tentacles (3350). (D) Expression of the prephe tentacles (3350). (E) Staining of the preprohormone A gene expnote the stained processes). The staining was carried out by alkalinequential use of the two substrates gave both a high resolution an
of a developing bud, showing expression of the preprohormone Cneurons at the tip of the bud (380).
Copyright © 1999 by Academic Press. All right
pattern in the appearance of C neurons similar to that inhe above-mentioned regenerates. First, the C neuronsncrease in number and spread over a larger surface of the
ne genes in Hydra whole mounts. (A) Peduncle and basal disk of aned) is expressed in an ectodermal dense network of neurons in thetentacles, subhypostomal region, and developing peduncle of the bud.ips of the bud (380). (B) Hypostome of an adult Hydra. Note that then the ectoderm of the hypostome, whereas the preprohormone C gene0).
ormone genes in whole mounts of Hydra. (A) Expression of thentacles (3350). (B) Expression of the preprohormone A (blue) andExpression of the preprohormone A (brown) and C (blue) gene inrmone C (blue) and LWamide (brown) gene in different neurons ofn in cells of the tentacle base to show that these cells are neurons
osphatase and Fast Red, followed by BM purple as a substrate. Thisintensive staining of the cells (3350). (F) Lower-power micrograph) and A (red) gene. Note the occurrence of a central cluster of C
ormone (rme,
acle tted i
prohhe te). (C)roho
ressioe phd an(blue
s of reproduction in any form reserved.

196 Mitgutsch, Hauser, and Grimmelikhuijzen
FIG. 5. Whole-mount hybridizations of developing buds of Hydra. (A) A very young bud hybridized with a probe coding for preprohormoneA. Note hybridizing cells clustered around the tip of the bud (3180). (B) A young bud just starting to develop tentacles hybridized with aprobe coding for preprohormone A. Strong hybridization occurs all over the tip of the bud (3145). (C) A late bud with several tentacles,hybridized with a probe coding for preprohormone A. Hybridizing neurons occur in the hypostome, in the tentacles, and in a regioncorresponding to the peduncle (3180). (D) A young bud hybridized with a probe coding for preprohormone B. Hybridizing neurons occurexclusively at the tip (3200). (E) A late bud with several tentacles hybridized with a probe coding for preprohormone B. Stained neurons
exclusively occur in the hypostome (3175).Copyright © 1999 by Academic Press. All rights of reproduction in any form reserved.
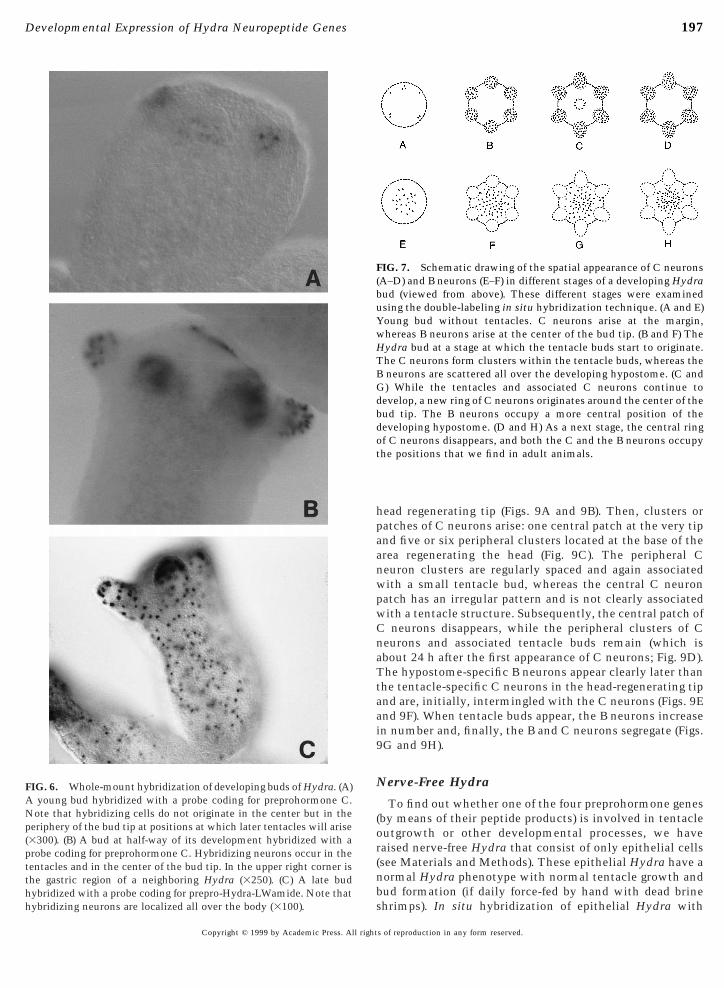
hybridizing neurons are localized all over the body (3100).
197Developmental Expression of Hydra Neuropeptide Genes
Copyright © 1999 by Academic Press. All right
head regenerating tip (Figs. 9A and 9B). Then, clusters orpatches of C neurons arise: one central patch at the very tipand five or six peripheral clusters located at the base of thearea regenerating the head (Fig. 9C). The peripheral Cneuron clusters are regularly spaced and again associatedwith a small tentacle bud, whereas the central C neuronpatch has an irregular pattern and is not clearly associatedwith a tentacle structure. Subsequently, the central patch ofC neurons disappears, while the peripheral clusters of Cneurons and associated tentacle buds remain (which isabout 24 h after the first appearance of C neurons; Fig. 9D).The hypostome-specific B neurons appear clearly later thanthe tentacle-specific C neurons in the head-regenerating tipand are, initially, intermingled with the C neurons (Figs. 9Eand 9F). When tentacle buds appear, the B neurons increasein number and, finally, the B and C neurons segregate (Figs.9G and 9H).
Nerve-Free Hydra
To find out whether one of the four preprohormone genes(by means of their peptide products) is involved in tentacleoutgrowth or other developmental processes, we haveraised nerve-free Hydra that consist of only epithelial cells(see Materials and Methods). These epithelial Hydra have anormal Hydra phenotype with normal tentacle growth andbud formation (if daily force-fed by hand with dead brine
FIG. 7. Schematic drawing of the spatial appearance of C neurons(A–D) and B neurons (E–F) in different stages of a developing Hydrabud (viewed from above). These different stages were examinedusing the double-labeling in situ hybridization technique. (A and E)Young bud without tentacles. C neurons arise at the margin,whereas B neurons arise at the center of the bud tip. (B and F) TheHydra bud at a stage at which the tentacle buds start to originate.The C neurons form clusters within the tentacle buds, whereas theB neurons are scattered all over the developing hypostome. (C andG) While the tentacles and associated C neurons continue todevelop, a new ring of C neurons originates around the center of thebud tip. The B neurons occupy a more central position of thedeveloping hypostome. (D and H) As a next stage, the central ringof C neurons disappears, and both the C and the B neurons occupythe positions that we find in adult animals.
FIG. 6. Whole-mount hybridization of developing buds of Hydra. (A)A young bud hybridized with a probe coding for preprohormone C.Note that hybridizing cells do not originate in the center but in theperiphery of the bud tip at positions at which later tentacles will arise(3300). (B) A bud at half-way of its development hybridized with aprobe coding for preprohormone C. Hybridizing neurons occur in thetentacles and in the center of the bud tip. In the upper right corner isthe gastric region of a neighboring Hydra (3250). (C) A late budhybridized with a probe coding for prepro-Hydra-LWamide. Note that
shrimps). In situ hybridization of epithelial Hydra with
s of reproduction in any form reserved.

198 Mitgutsch, Hauser, and Grimmelikhuijzen
FIG. 8. Whole-mount hybridization of Hydra regenerating a head, using a cRNA probe coding for preprohormone C. Hydra heads were cut offby a transverse section just below the tentacle zone and the regeneration of heads in the remaining body columns was monitored. (A) Side viewof a regenerating tip 24 h after removal of the head. Scattered C neurons have appeared at the tip (3250). (B) A regenerating tip 24 h after removalof the head. After the hybridization procedure was finished, this tip was cut off and mounted in such a way that it could be viewed from the top.Note the irregular distribution of C neurons (3250). (C) A regenerating tip 32 h after removal of the head. The view is from the top. Note the Cneuron clusters that, at three or four peripheral positions, are already part of small tentacle buds (3250). (D) A regenerating tip 40 h after removalof the head and viewed from the top. Note the one central and six peripheral tentacle buds, each containing C neurons. The central tentacle budhas the same size as the peripheral ones (3200). (E) Side view of a regenerating tip 44 h after removal of the head. The peripheral tentacles arenow well developed, while a central patch of C neurons is still present (3150). (F) Side view of a regenerating tip 48 h after removal of the head.
This animal has well-developed peripheral tentacles, while the central patch of C neurons has now disappeared (3200).Copyright © 1999 by Academic Press. All rights of reproduction in any form reserved.

nRbLpirprilt
rHBtihdetiaaanncbet
tdtntarafr(ffiwrteprs1ma
199Developmental Expression of Hydra Neuropeptide Genes
probes corresponding to each of the four preprohormonemRNAs showed that this type of Hydra did not express anyof the preprohormone genes. This clearly shows that thefour preprohormone genes are not essential for the basicpattern formation of Hydra.
DISCUSSION
The distributions of the A, B, and C neurons, when added
FIG. 9. Schematic drawing of the temporal and spatial appearanceof C neurons (A–D) and B neurons (E–H) during head regenerationof the lower half of a Hydra, of which the upper half, includinghead, has been removed (viewed from above). The time scale givesa rough indication of the time points during which the differentdevelopmental stages appear. All stages were examined using thedouble-labeling in situ hybridization technique. (A and E) First Cneurons appear at the tip of the regenerating head, while B neuronsare still absent. (B and F) The C neurons increase in number and arespread in an irregular fashion over the surface of the regeneratingtip. The first B neurons appear, intermingled with the C neurons.(C and G) An irregular patch of C neurons has appeared in thecenter of the regenerating tip, together with five or six small,regularly spaced tentacle buds in the periphery, each associatedwith a cluster of C neurons. The B neurons have increased innumber and are localized all over the center of the regenerating tip.(D and H) The peripheral tentacle buds and associated C neuronshave become more prominent, while the central C neuron patchhas disappeared. The B neurons occupy a region lying apical of thetentacle basis. The distribution of C and B neurons now resemblesthe pattern found in adult Hydra. In stage (A 1 E), the center of theregenerating tip has pure tentacle-like properties; in stage (B 1 F)and (C 1 G), there is a mixture of tentacle- and hypostome-likecharacteristics; and in stage (D 1 H), the center of the regeneratingtip resembles a normal hypostome.
together, correspond well with the distributions of those e
Copyright © 1999 by Academic Press. All right
eurons in Hydra that we visualized previously usingFamide antibodies (Grimmelikhuijzen, 1985). The distri-ution of the neurons expressing the prepro-Hydra-Wamide gene, however, is somewhat different from thatublished by Leitz and Lay (1995), using LWamide antibod-es. Whereas Leitz and Lay found LWamide-positive neu-ons only in the tentacles and basal disk, we find strongrepro-Hydra-LWamide gene expression in numerous neu-ons distributed all over the Hydra body (Fig. 2D). However,n some batches of Hydra, we find that the gastric regionsack prepro-Hydra-LWamide expression. The reasons forhese variations are unknown.
Epithelial, nerve-free Hydra have a phenotype thatoughly resembles that of wild-type Hydra. In epithelialydra, we did not find expression of the preprohormone A,, or C or the LWamide preprohormone genes, which meanshat the four genes and their neuropeptide products are notnvolved in basic pattern formation of Hydra. It could be,owever, that the four neuropeptide genes control theifferentiation of nonepithelial cells in wild-type Hydra,.g., the differentiation of stem cells (I cells) into the variousypes of nerve cells. In this context, it was interesting tonvestigate the order in which the four types of neuronsppeared during head formation. In both head-regeneratingnimals and developing buds, this order was LWamide, A,nd then C and B neurons. This already means that theeuropeptide products of the Hydra-RFamide (A, B, C)eurons cannot be responsible for the differentiation of Iells into LWamide neurons, but it leaves open the possi-ility that the LWamide neurons are responsible for themergence of the A, B, or C neurons (or other combina-ions).
In a previous study, Bode and co-workers investigated theemporal pattern of early tentacle and hypostome formationuring head regeneration in Hydra (Bode et al., 1988). As aentacle marker, they used a monoclonal antibody recog-izing the ectodermal surface of epithelial cells from theentacles (and, to a lesser extent, also of cells from the foot),nd as a hypostome marker, they used an RFamide antibodyecognizing ectodermal hypostomal sensory cells. This RF-mide antibody, however, also recognized ganglion cellsrom the lower hypostomal region, tentacles, upper gastricegion, and foot and was, therefore, not hypostome-specificGrimmelikhuijzen, 1985). Using the two markers, theyound that, during head regeneration, the tentacle antigenrst appeared at the tip of the regenerating head togetherith RFamide-positive ganglion cells, which then spread
adially over a larger surface. Once the tentacle buds startedo originate and form a ring around the basis of the regen-rating head, both the tentacle antigen and the RFamide-ositive ganglion cells disappeared from the center of theegenerating tip and were replaced by RFamide-positiveensory cells characteristic for the hypostome (Bode et al.,988). From these data, Bode and co-workers proposed aodel in which the tip of the regenerating head first gainscertain positional value (or positional information based,
.g., on a morphogen gradient; see Wolpert, 1971, 1989;
s of reproduction in any form reserved.

(h
200 Mitgutsch, Hauser, and Grimmelikhuijzen
Wolpert et al., 1974) that corresponds to that of a tentacle.Later, the tip attains a higher positional value correspond-ing to that of a hypostome, thereby causing the prematuretentacle cells to disappear from the hypostome, but toremain further away from the tip at a position at which thepositional value is lower and corresponds to that of atentacle.
A second model for pattern formation exists that is alsobased on a morphogen (or “source”) gradient. Here, everystructure (e.g., Hydra hypostome or tentacles) has its ownshort-range autocatalytic activator and long-range rapidlydiffusible inhibitor (Gierer and Meinhardt, 1972; Mein-hardt, 1993, 1996). Also, this model can explain the findingsof Bode and co-workers (Meinhardt, 1993, 1996).
More recently, two other groups have confirmed theobservations of Bode and co-workers: Weinzinger et al.(1994) used as a tentacle marker an RNA probe correspond-ing to a newly cloned gene, ks1, coding for a tentacleepithelial cell-specific protein, of which the function isunknown. They found that 2 days after removal of the head(by a transverse section just below the tentacle ring), ks1expression appeared as a patch at the center of the head-regenerating tip, at a time point at which the basal tentacleswere already beginning to evaginate. Subsequently, thiscentral ks1 expression disappeared, whereas it remained inthe tentacles. Technau and Holstein (1995) applied thesame tentacle-specific monoclonal antibody as Bode andco-workers (1988), but used as a hypostomal marker amonoclonal antibody directed against epithelial cells fromthe hypostomal endoderm. In accordance with Bode et al.1988), these authors observed that, after removal of theead from Hydra (by a transverse cut just below the
tentacle zone), first a patch of tentacle-specific antigenappears at the center of the regenerating tip, followed by theappearance of the basal ring of tentacles and the disappear-ance of the central patch of tentacle antigen. The endoder-mal hypostome marker appears considerably later than thetentacle marker, namely more than 12 h after the appear-ance of the basal tentacle ring. The combined use of anendodermal hypostome marker and an ectodermal tentaclemarker might be dangerous, because it could theoreticallybe possible that the hypostomal endoderm follows differentregeneration kinetics than the hypostomal ectoderm. Themost elegant way to compare hypostome and tentacledevelopment, therefore, is to use markers for the sametissue layer.
Recently two other hypostome markers, hyp-1 and bud-head, that appear as early as 8 h after removal of the headhave been published (Hermans-Borgmeyer et al., 1996;Martinez et al., 1997). Both markers, however, are endoder-mal. Furthermore, budhead is not fully hypostome-specific(Martinez et al., 1997).
Our present experimental data support a model thatresembles the original two-part pattern model for headregeneration, as proposed by Bode et al. (1988). The markersthat we used, however, were different from and superior to
the ones used by Bode et al. (1988), Weinzinger et al. (1994),Copyright © 1999 by Academic Press. All right
and Technau and Holstein (1995) because: (i) They weremore specific; both preprohormone B and C probes bind toa mRNA species coding for a known protein with a knownfunction. Furthermore, the preprohormone B probe stainsexclusively neurons of the ectoderm of the hypostome (Figs.2B, 4B, 5D,and 5E), while the preprohormone C probe stainsexclusively neurons of the ectoderm of the tentacles (Fig.2C). (ii) Our tentacle-specific marker is an early marker fortentacle ectoderm. This marker could already recognizeneurons in an apical head-regenerating tip 20 h after headremoval and 12 h before the first peripheral tentacle struc-tures became visible. It is, therefore, 12–28 h earlier thanthe tentacle marker used by Weinzinger et al. (1994).However, our tentacle-specific marker might not be signifi-cantly earlier than the one used by Bode and co-workers(1988) and Technau and Holstein (1995). (iii) Ourhypostome-specific marker is an early marker for hypos-tome ectoderm. This marker could already recognize neu-rons in the tip of a head-regenerating lower half of Hydra8 h before the first tentacle buds became apparent (Fig. 9F).The marker, therefore, is at least 16 h earlier than thehypostome-specific marker used by Technau and Holstein(1995). Furthermore, because our hypostome-specificmarker recognizes mRNA coding for RFamide peptidesproduced by the hypostomal sensory neurons, it must alsobe significantly earlier than the RFamide antibody markerused by Bode and co-workers (1988). (iv) Our two mRNAmarkers enabled us to develop a double-labeling technique.Double-labeling techniques were not used by the threeother research groups in their regeneration experiments.
Using our two early and specific markers, we found that,during head regeneration of the lower half of Hydra, firstthe tentacle-specific C neurons appear at the center of theregenerating tip (Fig. 9A). These neurons subsequentlyincrease in number and spread radially, thereby occupying alarger surface (Fig. 9B). At this point, the first B neuronsarise, intermingled with the C neurons (Fig. 9F). Then theperipheral C neurons become organized in five or sixregularly spaced clusters, each associated with a smalltentacle bud, while the central C neurons form an irregularpatch around the center of the regenerating tip (Fig. 9C).Subsequently, the central patch of C neurons disappearsand is replaced by hypostome-specific B neurons (Figs. 9Dand 9H). At this final stage, the whole center of theregenerating tip is occupied by hypostome-specific B neu-rons (Fig. 9H), where about 1 day earlier this region onlycontained tentacle-specific C neurons (Fig. 9A). Thus, alsowith our own tentacle and hypostome markers, we find thathead formation in the head-regenerating tip of Hydra passesthrough at least two phases, a tentacle- and a hypostome-specific phase.
Although our observations confirm the conclusions ofBode and co-workers (1988), they are much more detailed,partly due to the specificity and early appearance of ourtentacle and hypostome markers. First, we found that thereis a broader overlap between the tentacle- and hypostome-
specific phases in the head-regenerating tip: the hypostome-s of reproduction in any form reserved.

ahtzttdp
Cvtrihntid9ptepfa
201Developmental Expression of Hydra Neuropeptide Genes
specific B neurons already coexist with the tentacle-specificC neurons at a very early stage of head regeneration, whententacle buds have not yet appeared (Figs. 9B and 9F). Thisis 8–16 h earlier than described by Bode et al. (1988). Theoverlap persists until the central patch of C neurons disap-pears from the hypostome (Figs. 9D and 9H). Therefore,there is not a clear-cut two-part pattern of head regenera-tion such as Bode et al. (1988) propose, but it would be moreppropriate to speak of several transient phases leading toead formation. Second, in animals in which we removedhe head by a transverse section just below the tentacleone, we observed that during head regeneration a realentacle structure appeared in the center of the regeneratingip (Fig. 8D), which during the progress of head formationisappeared again. This phenomenon has never been re-orted before.It is interesting that the peripheral and central clusters ofneurons, arising during head regeneration, are spaced in a
ery regular way (Fig. 8D). This spacing might be caused byhe “tentacle inhibitor,” which is a hypothetical, short-anged morphogen released by a developing tentacle andnhibiting the formation of other, nearby tentacles (Mein-ardt, 1993, 1996). The appearance of regularly spaced Ceuron clusters, therefore, might be caused by the inhibi-ion of preprohormone C gene expression in neurons lyingn between the tentacle “anlagen.” It might also be that theisappearance of the central C neuron cluster (Figs. 8D andC) is caused by inhibition coming from the other five or sixeripheral tentacle anlagen, because the perceived inhibi-ion in the central anlage might be expected to be consid-rably higher (two to three times) than that in one of theeripheral anlagen. Thus, two explanations might be givenor the localization of C neurons and associated tentacles indult Hydra. One is based on the above-mentioned assump-
tion that C neuron and tentacle formation occurs betweentwo thresholds (between high and very high positionalvalues). In the other explanation, there is only one lowerthreshold (high positional value), and C neurons and asso-ciated tentacles are not expressed at very high positionalvalues (e.g., in the hypostome), because during head devel-opment, the peripheral tentacle anlagen inhibit the forma-tion or maintenance of other tentacles in that region.
Of course, there are other possible explanations for thepositions of the tentacles and associated C neurons. One ofthem has been worked out in a computer model by Mein-hardt (1993, 1996). Here, the tentacles have, again, only onelower threshold (high positional value or, in the terminol-ogy of Meinhardt, high “source density”) and the expressionof tentacles at very high positional values is inhibited bythe developing hypostome itself (via its hypostome “acti-vator”). For our head-regeneration experiments, this wouldmean that there is a competition between the tentacle-specific cells (among them C neurons) and hypostome-specific cells (among them B neurons) in the early phases ofa regenerating head (Figs. 9B and 9F). As the number ofhypostome-specific cells increases in the center of the
head-regenerating tip (Figs. 9F–9H), due to the increase inCopyright © 1999 by Academic Press. All right
positional value, the number of tentacle-specific cellswould decrease in this area (Figs. 9C and 9D), thus leadingto a sorting out of tentacle- and hypostome-specific cells. Inagreement with our experimental data, Meinhardt’s com-puter simulations predict the emergence of transienttentacle-specific structures in the center of the regeneratingtip during head regeneration (Figs. 8D and 9C; Meinhardt,1993).
It is assumed that the above-mentioned positional valuefor head formation in adult Hydra is highest at the hypos-tomal apex, followed by the hypostomal base, tentacles,upper gastric region, lower gastric region, and foot (Wolpert,1971, 1989; Gierer and Meinhardt, 1972; Wolpert et al.,1974; Meinhardt, 1993, 1996). Thus, as discussed above,preprohormone B expression would reflect the highest po-sitional value, since it is specific for the hypostome,whereas preprohormone C expression would represent asomewhat lower value, since it is specific for the tentacles.It would be interesting, then, to understand how the expres-sion of the preprohormone A gene is regulated, since thisgene is expressed in the hypostomal apex, hypostomal base,tentacles, and upper gastric region (Fig.2). Expression in thehypostomal apex points to a very high positional value,whereas expression in the upper gastric region points to amedial value. The answer, therefore, could be that thepreprohormone A gene is activated by a wide range ofpositional values, from medial to very high. A similarexplanation could be given for the expression of the prepro-Hydra-LWamide gene, which is found all over the Hydrabody. This gene would be activated by an even wider rangeof positional values, from low to very high.
These interpretations are supported by our head-regeneration experiments. One can assume that after theremoval of the head and upper gastric region, the posi-tional value in the tip of the remaining half of the Hydrabody column is low and that this value gradually in-creases again during head regeneration (Wolpert, 1971,1989; Gierer and Meinhardt, 1972; Wolpert et al., 1974;Meinhardt, 1993, 1996). In agreement with this, LW-amide neurons originate early (at a low positional value)during head regeneration and continue to be present untilthe regeneration is complete (also in the newly regener-ated hypostome, where there is a very high positionalvalue). In our head-regenerating animals, the A neuronsappear shortly after the emergence of LWamide neurons(at medial positional value) and they continue to bepresent until the regeneration is complete (also in thehypostome, where there is a very high positional value).As mentioned earlier, the C neurons follow the A neu-rons, and the B neurons follow the C neurons, in head-regenerating Hydra. Thus, both in intact and in head-regenerating animals, the expression of the four differentpreprohormone genes can be explained by a specific po-sitional value (high or low; with a broad or a narrowrange) required to activate them. This does not exclude,however, that certain “fine tunings” exist, e.g., that one
of the neuropeptide gene products could influence thes of reproduction in any form reserved.

rttt
pha
taHtc
C
D
202 Mitgutsch, Hauser, and Grimmelikhuijzen
expression of its own or other neuropeptide genes. Such afine tuning could occur at a certain positional value andcould create a second pattern in a second and third di-mension (see, for example, the regularly-spaced distribu-tions of A, C, and LWamide neurons at a certain posi-tional value, that of the tentacles; Fig. 3).
Our explanation, namely that the expression of the fourdifferent preprohormone genes is simply based on posi-tional values, appears to be valid only for the head andgastric regions. The situation in the foot might be morecomplicated, since other morphogen gradients are likely toplay a role in this area (Meinhardt, 1993).
The development of a Hydra bud might also representa special situation. The initial positional value of a Hydrabud is low, because it starts to grow at a position locatedat the lower one-fourth to one-third of the Hydra bodycolumn. It is assumed that the positional value of theapex of the developing bud subsequently increases duringbud growth (Gierer and Meinhardt, 1972; Meinhardt,1993, 1996). Therefore, one would expect that the fourtypes of neurons would appear in the same temporalorder during bud outgrowth as during head regeneration.In accordance with this is the emergence of first theLWamide and then the A neurons. However, in contrastto head regenerates, the C and B neurons appear at aboutthe same time (Figs. 7A and 7E). Also the C and B neuronsare not intermingled as we found in head regenerates(Figs. 9B and 9F), but they have already been sorted outwhen they appear: the C neurons originate at the periph-ery of the bud tip (Figs. 6A and 7A), whereas the Bneurons emerge at the center (Fig. 7E). Both observationsare not conflicting with our earlier conclusions fromintact animals and head regenerates, namely that the Cneurons appear at high and the B neurons at very highpositional values. However, in contrast to head regener-ates, it seems that the center of the tip of the Hydra budemains only for a short period of time at a high posi-ional value and quickly passes to a very high value,hereby not allowing its neurons enough time to expressheir preprohormone C gene.
After a pattern had been established with five or sixeripheral tentacles containing C neurons (Fig. 7B), and aypostome containing B neurons (Fig. 7F), we observednother interesting phenomenon in Hydra buds, namely
the transient appearance of a secondary ring of C neuronsat the center of its hypostome (Figs. 3F, 6B, and 7C). Thisring of C neurons clearly indicates the presence oftentacle cells and suggests that pattern formation has notyet been completed. We have no good explanations forthis transient appearance of the C neuron ring, and also,a computer simulation of head formation in developingHydra buds does not predict the emergence of tentacle-like structures at the center of its developing hypostome(Meinhardt, 1993, 1996). Thus, although theoreticallyunexplained, we can clearly see that the tip of a Hydrabud also passes through several phases of development, of
which some have mixed tentacle and hypostome fea-Copyright © 1999 by Academic Press. All right
ures. Therefore, our findings in both head regeneratesnd developing buds clearly show that head formation inydra involves several phases of development, in which
entacle- and hypostome-specific gene expressions arelosely coexisting.
ACKNOWLEDGMENTS
We thank Dr. T. Fujisawa for the kind gift of Hydra strain A10,Dr. H. Meinhardt (Max Planck Institute for Developmental Biol-ogy, Tubingen, Germany) for his comments on the manuscript,Astrid Juel Jensen and Winnie Larsen for typing the manuscript,Beth Beyerholm and Michael Williamson for drawing Figs. 1, 7, and9, and the Danish Natural Science Research Council for financialsupport. Christian Mitgutsch was an Erasmus Student from theUniversity of Jena, Germany.
REFERENCES
Bode, P. M., Awad, T. A., Koizumi, O., Nakashima, Y., Grimmelik-huijzen, C. J. P., and Bode, H. R. (1988). Development of thetwo-part pattern during regeneration of the head in Hydra.Development 102, 223–235.ampbell, R. D. (1976). Elimination of Hydra interstitial and nervecells by means of colchicine. J. Cell Sci. 21, 1–13.armer, D., Hauser, F., Nothacker, H.-P., Bosch, T. C. G., Williamson,M., and Grimmelikhuijzen, C. J. P. (1998). Three different prohor-mones yield a variety of Hydra-RFamide (Arg-Phe-NH2) neuropep-tides in Hydra magnipapillata. Biochem. J. 332, 403–412.
Gierer, A., and Meinhardt, H. (1972). A theory of biological patternformation. Kybernetik 12,30–39.
Grens, A., Gee, L., Fisher, D. A., and Bode, H. R. (1996). CnNK-2, anNK-2 homeobox gene, has a role in patterning the basal end ofthe axis in Hydra. Dev. Biol. 180, 473–488.
Grimmelikhuijzen, C. J. P. (1985). Antisera to the sequence Arg-Phe-amide visualize neuronal centralization in hydroid polyps.Cell Tissue Res. 241, 171–182.
Grimmelikhuijzen, C. J. P., Carstensen, K., Darmer, D., Moosler, A.,Nothacker, H.-P., Reinscheid, R. K., Schmutzler, C., Vollert, H.,McFarlane, I. D., and Rinehart, K. L. (1992). Coelenterate neuropep-tides: Structure, action and biosynthesis. Am. Zool. 32, 1–12.
Grimmelikhuijzen, C. J. P., Leviev, I., and Carstensen, K. (1996).Peptides in the nervous systems of cnidarians: Structure, func-tion and biosynthesis. Int. Rev. Cytol. 167, 37–89.
Hauptmann, G., and Gerster, T. (1994). Two-color whole-mount insitu hybridization to vertebrate and Drosophila embryos. TrendsGenet. 10, 266.
Hermans-Borgmeyer, I., Schinke, B., Schaller, H. C., andHoffmeister-Ullerich, S. A. H. (1996). Isolation of a marker forhead-specific cell differentiation in Hydra. Differentiation 61,95–101.
Koizumi, O., Wilson, J. D., Grimmelikhuijzen, C. J. P., and West-fall, J. A. (1989). Ultrastructural localization of RFamide-likepeptides in neuronal dense-cored vesicles in the peduncle ofHydra. J. Exp. Zool. 249, 17–22.
Leitz, T., Morand, K., and Mann, M. (1994). Metamorphosin A: Anovel peptide controlling development of the lower metazoanHydractinia echinata (Coelenterata, Hydrozoa). Dev. Biol. 163,440–446.
Leitz, T., and Lay, M. (1995). Metamorphosin A is a neuropeptide.
Roux’s Arch. Dev. Biol. 204, 276–279.s of reproduction in any form reserved.

W
W
203Developmental Expression of Hydra Neuropeptide Genes
Leviev, I., Williamson, M., and Grimmelikhuijzen, C. J. P. (1997).Molecular cloning of a preprohormone from Hydra magnipapil-lata containing multiple copies of Hydra-LWamide (Leu-Trp-NH2) neuropeptides: Evidence for processing at Ser and Asnresidues. J. Neurochem. 68, 1319–1325.
Martinez, D. E., Dirksen, M. L., Bode, P. M., Jamrich, M., Steele,R. E., and Bode, H. R. (1997). Budhead, a fork head/HNF-3homologue, is expressed during axis formation and head specifi-cation in Hydra. Dev. Biol. 192, 523–536.
Meinhardt, H. (1993). A model for pattern formation of hypostome,tentacles, and foot in Hydra: How to form structures close toeach other, how to form them at a distance. Dev. Biol. 157,321–333.
Meinhardt, H. (1996). Models of biological pattern formation:Common mechanism in plant and animal development. Int. J.Dev. Biol. 40, 123–134.
Moosler, A., Rinehart, K. L., and Grimmelikhuijzen, C. J. P. (1996).Isolation of four novel neuropeptides, the Hydra-RFamides I–IV,from Hydra magnipapillata. Biochem. Biophys. Res. Commun.229, 596–602.
Sugiyama, T., and Fujisawa, T. (1978). Genetic analysis of develop-mental mechanisms in Hydra. V. Cell lineage and development
of chimera Hydra. J. Cell Sci. 32, 215–232.Copyright © 1999 by Academic Press. All right
Takahashi, T., Muneoka, Y., Lohmann, J., Lopez de Haro, M. S.,Solleder, G., Bosch, T. C. G., David, C. N., Bode, H. R., Koizumi,O., Shimizu, H., Hatta, M., Fujisawa, T., and Sugiyama, T. (1997).Systematic isolation of peptide signal molecules regulating de-velopment in Hydra: LWamide and PW families. Proc. Natl.Acad. Sci. USA 94, 1241–1246.
Technau, U., and Holstein, T. W. (1995). Head formation in Hydrais different at apical and basal levels. Development 121, 1273–1282.
Weinzinger, R., Salgado, L. M., David, C. N., and Bosch, T. C. G.(1994). Ks1, an epithelial cell-specific gene, responds to early signalsof head formation in Hydra. Development 120, 2511–2517.
Wolpert, L. (1971). Positional information and pattern formation.Curr. Top. Dev. Biol. 6, 183–224.olpert, L. (1989). Positional information revisited. Development107 (Suppl.), 3–12.olpert, L., Hornbruch, A., and Clarke, M. R. B. (1974). Positionalinformation and positional signalling in Hydra. Am. Zool. 14,647–663.
Received for publication July 24, 1998Revised November 12, 1998
Accepted November 12, 1998
s of reproduction in any form reserved.




















