
Formaldehyde Cycle and the Phases of Stress Syndrome
12
Formaldehyde Cycle and the Phases of Stress Syndrome ERN ˝ O TYIHÁK, a,b LAJOS TRÉZL, c AND BÉLA SZENDE d b Plant Protection Institute, Hungarian Academy of Sciences, P.O. Box 102, H-1525 Budapest, Hungary c Department of Organic Chemical Technology, Technical University of Budapest, P.O. Box 91, H-1521 Budapest, Hungary d I. Institute of Pathology and Experimental Cancer Research, Semmelweis University Medical School, Üll˝ oi út 26., H-1085 Budapest, Hungary INTRODUCTION A detectable amount of formaldehyde (HCHO) is found in biological systems that originate from both methylation 1 and demethylation. 2 Radiolabeled formaldemethone is formed from the S-CH 3 - 3 H of S-adenosyl-L-methionine (SAM) in the course of the enzymatic conversion of histamine to N τ -methylhista- mine in the presence of dimedone (added as a HCHO capture molecule). The for- mation of HCHO is probably linked to the enzymatic transmethylation of histamine. 1 The high level of HCHO and the accumulation of N-methylated sub- stances during the early stages of plant growth and in rapidly dividing cells are the result of intensive transmethylation reactions. 3 The amount of HCHO increased considerably in virus-infected tobacco leaves, and with L-methionine and SAM as substrates the demethylase activity was very high (biotic stress). 4 A strong correlation also exists between external temperature and the amount of measurable HCHO in pinto bean leaf tissues. 5 High levels were detected after heat-shock treatment of the leaves, which also led to the levels of three potential HCHO generators (trigonelline, choline, and N ε -trimethyl-L-lysine [TML]) being decreased moderately (abiotic stress). This paper summarizes recent results in connection with the relationship between formaldehyde cycle and the phases of stress syndrome. FORMALDEHYDE IN THE BIOSPHERE SAM serves as a methyl donor in virtually all of the vast number of enzymatic transmethylation reactions, including DNA. Earlier results 1,6 query the existence of a methyl radical or cation in the enzymatic transmethylation reaction, clearly detecting a formation of HCHO from the methyl moiety of SAM. 1 In the demethylation process of SAM as a sulfonium ion with an optically active center, the first biotransformation pathway for SAM containing the 259 a Additional correspondence information: Telephone: 36-1-1-558-722/315; Fax: 36-1-1-56- 3698; e-mail: [email protected]
-
Upload
independent -
Category
Documents
-
view
8 -
download
0
Transcript of Formaldehyde Cycle and the Phases of Stress Syndrome

Formaldehyde Cycle and the Phases ofStress Syndrome
ERNO TYIHÁK,a,b LAJOS TRÉZL,c AND BÉLA SZENDEd
bPlant Protection Institute, Hungarian Academy of Sciences,P.O. Box 102, H-1525 Budapest, HungarycDepartment of Organic Chemical Technology, Technical University ofBudapest, P.O. Box 91, H-1521 Budapest, HungarydI. Institute of Pathology and Experimental Cancer Research,Semmelweis University Medical School, Ülloi út 26., H-1085 Budapest, Hungary
INTRODUCTION
A detectable amount of formaldehyde (HCHO) is found in biological systems thatoriginate from both methylation1 and demethylation.2 Radiolabeledformaldemethone is formed from the S-CH3-
3H of S-adenosyl-L-methionine(SAM) in the course of the enzymatic conversion of histamine to Nτ-methylhista-mine in the presence of dimedone (added as a HCHO capture molecule). The for-mation of HCHO is probably linked to the enzymatic transmethylation ofhistamine.1 The high level of HCHO and the accumulation of N-methylated sub-stances during the early stages of plant growth and in rapidly dividing cells arethe result of intensive transmethylation reactions.3
The amount of HCHO increased considerably in virus-infected tobacco leaves,and with L-methionine and SAM as substrates the demethylase activity was veryhigh (biotic stress).4 A strong correlation also exists between external temperatureand the amount of measurable HCHO in pinto bean leaf tissues.5 High levels weredetected after heat-shock treatment of the leaves, which also led to the levels ofthree potential HCHO generators (trigonelline, choline, and Nε-trimethyl-L-lysine[TML]) being decreased moderately (abiotic stress).
This paper summarizes recent results in connection with the relationshipbetween formaldehyde cycle and the phases of stress syndrome.
FORMALDEHYDE IN THE BIOSPHERE
SAM serves as a methyl donor in virtually all of the vast number of enzymatictransmethylation reactions, including DNA. Earlier results1,6 query the existenceof a methyl radical or cation in the enzymatic transmethylation reaction, clearlydetecting a formation of HCHO from the methyl moiety of SAM.1
In the demethylation process of SAM as a sulfonium ion with an opticallyactive center, the first biotransformation pathway for SAM containing the
259
aAdditional correspondence information: Telephone: 36-1-1-558-722/315; Fax: 36-1-1-56-3698; e-mail: [email protected]

S-methyl moiety is the hydroxylation of the carbon atom of the chiral methylgroup with subsequent cleavage of the chiral methyl group in the form ofmethanol, which is further metabolized to HCHO.6 HCHO reacts with the aminogroup of an amino acid (for example, arginine) at the active site of transmethylase,translating the hydroxymethyl (-CH2OH) group in reduction condition to theacceptor molecule.
There is a HCHO cycle in biological systems6,7 in which formation of the methylgroup of L-methionine takes place through HCHO and the formation of HCHOfrom SAM is linked to different enzymic transmethylation reactions1,7 (FIG. 1). Anumber of rapid formaldehyde pathways in different tissues exists throughhydroxymethyl groups linked to various acceptor molecules.8
FIGURE 2 illustrates an example for the measurement of HCHO in its dimedoneadduct form and some fully N-methylated substances (betaines) in the freshleaves of a tree species.
FIGURE 3 summarizes schematically the quantitative ratios of HCHO and itspotential generators during seasonal changes studying 31 different tree species.The level of HCHO was very high in spring and autumn leaves. The amount ofbetaines as marker molecules also was very high in spring. In this case the HCHOis originated from methylation processes, whereas in autumn the large amount ofHCHO can be formed in demethylation processes.9
Recently, HCHO, as its dimedone adduct (formaldemethone), has been iso-lated from and identified in, for example, human urine,10 as well as Ascophyllumnodosum (alga)11 and callus tissue culture of Datura innoxia.12
260 ANNALS NEW YORK ACADEMY OF SCIENCES
FIGURE 1. Biotransformation steps of HCHO cycle. Examples of transmethylase acceptors:nucleic acids, proteins (Lys, Arg), ethanolamine, norepinephrine, normorphine, and hista-mine. Examples of methylation products: Nε-methylated lysines, phosphatidylcholine,morphine, Nτ-Me-histamine. Examples of natural CH2O generators: N5-CH3THF andbetaine.

TYIHÁK et al.: FORMALDEHYDE CYCLE 261
FIGURE 2. Change of HCHO and its potential generators in Tilia platyphyllos depending onthe season.8,9 Experimental conditions: plant parts were frozen with liquid nitrogen and thefrozen material (250 mg) was mixed with 0.7 ml 0.2% methanolic dimedone solution. Theclear supernatant was used for OPLC consecutive separation8,9: TLC silica gel F254 withimpregnated edges, chloroform–methylene chloride (35 : 65, vol/vol) for formaldemethoneand then i-propanol–methanol–0.1M Na-acetate (20 : 3 : 30, vol/vol) for betaines.Densitometric evaluation was performed at 260 nm for formaldemethone and at 525 nm forbetaines after Dragendorff reaction. May, July, , October.
FIGURE 3. Change of HCHO and its potential generators in trees during the vegetation.8,9
Experimental conditions as in FIGURE 2.

CHANGE OF HCHO CYCLE IN STRESS SITUATIONS
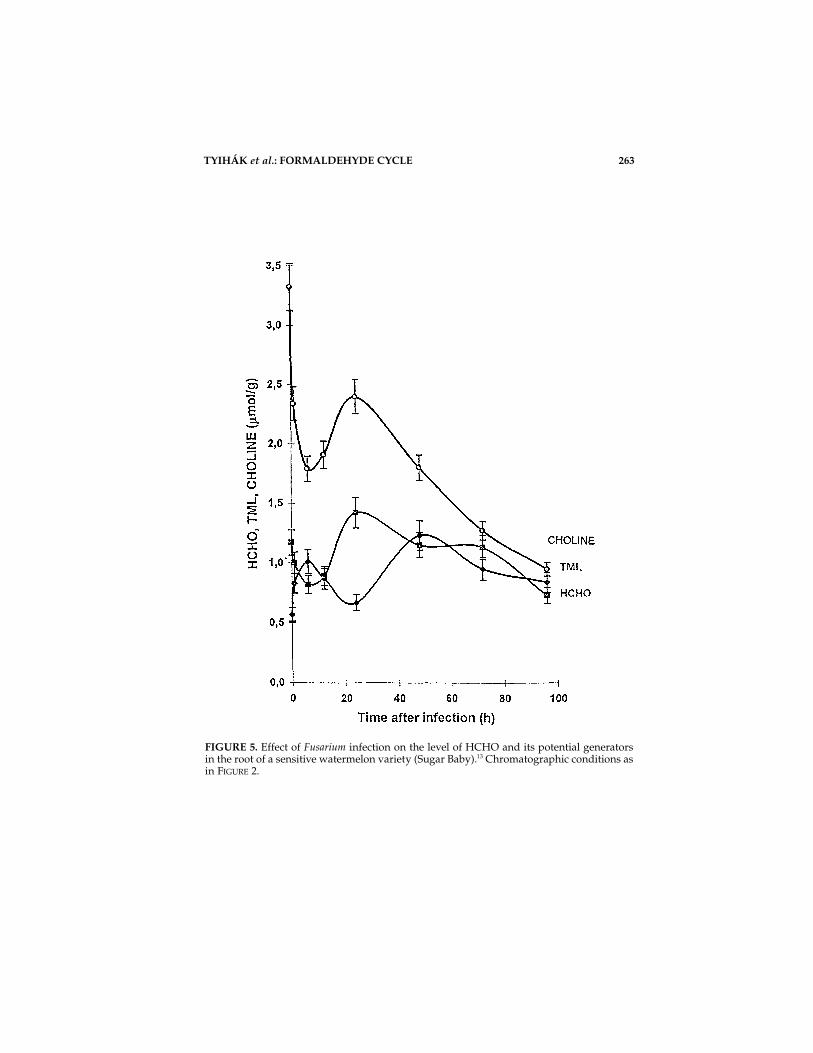
FIGURE 4 shows the phases of the stress syndrome according to Selye and oth-ers. HCHO (mainly in -CH2OH groups) and its potential generators (N-, S-, O-methylated small and large molecules) constitute a part of the resistance potentialThe measurable HCHO level is considerably elevated in the parts of watermelon(Citrullus vulgaris L.) immediately after a nonlethal infection with Fusarium oxys-porum f. sp. niveum. At the same time the level of some betaines (TML, choline) aspotential HCHO generators (gene products) is decreased (biotic stress).13
However, the biochemical “response reactions” of the two watermelon varieties ofdifferent disease sensitivity during the stress syndrome were characteristic (FIG. 5and FIG. 6).
Similar effects can be seen in abiotic stress (e.g. cold shock) situations as well.14
FORMATION OF EXCITED FORMALDEHYDE AND SINGLET OXYGEN INA L-LYSINE-MEDIATED TERNARY SYSTEM
Under physiological conditions HCHO can be activated by H2O2 in the pres-ence of the ε-amino group of L-lysine with simultaneous formation of 1O2, andchemiluminescence (CL). The measured CL spectrum can be seen in FIGURE 7.However, the light emission analysis at different wavelengths shows a red lightemission (530, 633, 705 nm) for 1O2 and at the same time a blue light emission (430nm) for excited group in HCHO ( > *C = O ).15
The chemiluminescence emission from the reaction between HCHO and H2O2
is low (TABLE 1). However, under the same conditions the presence of L-lysine infree or bound form considerably increases CL emission. Additionally, the forma-tion of CL emission in this L-lysine-mediated ternary system can be influenced bydifferent inhibitors and accelerators alike (TABLE 2).7,8,15,16
262 ANNALS NEW YORK ACADEMY OF SCIENCES
FIGURE 4. Phases of stress syndrome according to Selye and others.
TYIHÁK et al.: FORMALDEHYDE CYCLE 263
FIGURE 5. Effect of Fusarium infection on the level of HCHO and its potential generatorsin the root of a sensitive watermelon variety (Sugar Baby).13 Chromatographic conditions asin FIGURE 2.

264 ANNALS NEW YORK ACADEMY OF SCIENCES
FIGURE 6. Effect of Fusarium infection on the level of HCHO and its potential generatorsin the root of a resistant watermelon variety (Charleston).13 Chromatographic conditions asin FIGURE 2.

TYIHÁK et al.: FORMALDEHYDE CYCLE 265
FIGURE 7. Light emission analysis at different wavelengths in a mixture of L-lysine :HCHO : H2O2 (1 mM : 1 mM : 1 mM) at T = 298°K, pH = 7.4 in 0.1 M Sörensen buffer.8,15,16
TABLE 1. Chemiluminescense Emission Values of Different Systemsa
Reaction Mixture Count/min(cpm)
1 mM L-lysine + 1 mM HCHO 5.00 · 102
1 mM L-lysine + 1 mM H2O2 5.10 · 102
1 mM HCHO + 1 mM H2O2 2.50 · 103
1 mM L-lysine + 1 mM HCHO + 1 mM H2O2 1.05 · 105
1 mM N-acetyl-L-lysine-methylamide + 1 mM HCHO + 1 mM H2O2 1.36 · 105
1 mM Lys-Lys-Arg + 1 mM HCHO + H2O2 8.00 · 105
10 mg cytochrome C + 1 mM HCHO + 1 mM H2O2 5.50 · 105
aReaction conditions: 0.1 M Sörensen buffer (pH = 7.4), 25°C. Reaction time: 1 min.

The possible reaction steps in L-lysine, HCHO, and H2O2 systems with the new-type generation of 1O2 are as follows:
LYSINE - εNH2 + H2O2 → LYSINE - εN*H + H2O + *OHLYSINE - εN*H + HCHO → LYSINE - εNH2 + *CHO*CHO + O2 → *O-O-CHO
peroxi-formyl radicalO H O
// //LYSINE - εN*H + *O-O-C → 1O2 + LYSINE-εN - C
\ \H H
Nε-formyl-L-lysine*OH + *CHO → HCOOH
We have proved in model reactions that excited HCHO (H2C*O) can also be
formed in the presence of L-lysine and H2O2.8,15,16
HCHOLYSINE -εNH2 + HCHO → lysine - εNH - CH2OH →LYSINE - εNH - CH3 + HCOOHLYSINE - εNH - CH3 + H2O2 → lysine - εNH2 + CH3OOH
Decomposition methyl peroxideCH3OOH → CH3O
* + *OHMethoxy HydroxyRadical RadicalH2O + CH3OH
CH3O* + *OH
H*CHO + H2Oexcited formaldehyde
To this ternary system forming H*CHO and 1O2, the free or bound L-lysine arepresent as common components in practically all biological systems, while HCHOforms with the dissociation of hydroxymethyl groups from methylation anddemethylation reactions,1,2 and H2O2 also is a normal metabolite.17
The biotic and abiotic stress conditions provide favorable opportunities for theinvasive formation of these excited molecules (H*CHO and 1O2) and the existence
266 ANNALS NEW YORK ACADEMY OF SCIENCES
TABLE 2. Effect of Inhibitors and Accelerators on the Chemiluminescence Emission in L-Lysine-HCHO-H2O2 Reaction Mixture under Physiological Conditionsa
Reaction Mixture Accelerator/Inhibitor Count/min(cpm)
1 mM L-lysine + 1 mM HCHO 0 1.05 · 105
+ 1 mM H2O2 (RM)RM 5 mg horseradish peroxidase 1.25 · 105
RM 1 mM L-tryosine 2.00 · 105
RM 1 mM Na-azid (NaN3) 3.10 · 104
RM 1 mM L-ascorbic acid + 6.00 · 103
1 mM glutathione
aReaction conditions: 0.1 M Sörensen buffer (pH = 7.4), 25°C.
→
→

of these opportunities explains the intensive burst under extreme environmentalcircumstances. Under stress-free conditions a noninvasive, continuous formationof H*CHO and 1O2 is probable.
FORMALDEHYDE AND APOPTOSIS
Interestingly, the term apoptosis, or programmed cell death (pcd), was derivedfrom the Greek by Kerr in 1972 to reflect the process by which leaves fall fromtrees to emphasize the natural process of a programmed form of cell death he wasstudying in liver tissue.18 In the actual apoptosis (the fall of autumn leaves fromtrees) the level of HCHO increases dramatically before leaf fall (e.g., compared tothe summer HCHO level, autumn HCHO levels are 5–10 thousand percent higher,and at the same time the amount of choline, trigonelline, and so forth decreases incorrelation with the HCHO formation (see FIG. 3).9
Using 1′-methyl-ascorbigen (Me-Asc), an N-methylindole-containing deriva-tive of L-ascorbic-acid for the treatment of PC-3 human prostate carcinoma cells, itresulted in a significant decrease in tumor cell number in parallel with an increasein apoptotic cells (TABLE 3).19
The administration of the unmethylated ascorbigen didn’t result in a similareffect. Accordingly, the N-methyl group was hypothesized to play an essential rolein biological activities of Me-Asc. Dimedone, forming formaldemethone with
TYIHÁK et al.: FORMALDEHYDE CYCLE 267
TABLE 3. Level of HCHO in the Culture Medium of PC-3 Prostate Cancer Cells 48 Hoursafter Treatment with 100 µg/ml of Me-Asc
Treatment HCHO (µg/ml) Percent of ControlControl 2.2 ± 0.1 100Me-Asc 3.6 ± 0.3 164
TABLE 4. Effect of Methyl-Ascorbigen (Me-Asc), Dimedone (Di) and Me-Asc+Di on theProliferation of PC-3 Human Prostate Cancera
Treatment 24 Hours 48 Hours 72 HoursControl 9.2 ± 0.1 13.6 ± 0.4 15.3 ± 0.9Me-Asc 5.8 ± 0.1b 4.7 ± 1.5c 7.5 ± 1.2d
Di 9.4 ± 1.2 14.2 ± 2.2 17.8 ± 9.3Me-Asc+Di 7.1 ± 1.7 9.9 ± 2.5e 9.0 ± 1.2ViabilityControl 91 ± 5 99 ± 2 94 ± 3Me-Asc 88 ± 4 71 ± 6 65 ± 2Di 94 ± 6 98 ± 1 96 ± 6Me-Asc+Di 84 ± 2 88 ± 0 88 ± 2
aDose: Me-Asc: 100 µg/ml; Di: 10 µg/ml. Hours after treatment: cell number and viabil-ity (V).
bp < 0.005 vs. control.cp < 0.0025 vs. control.dp < 0.01 vs. control.ep < 0.01 vs. Me-Asc.

HCHO, applied simultaneously with Me-Asc diminished the apoptosis-inducingeffect of Me-Asc (TABLE 4).19
These results indicate that the HCHO generated in situ from the N-methyl groupof Me-Asc is linked to the induction of pcd and the retardation of cell proliferation.It is supposed that in autumn leaves the accumulated HCHO is excited.8,16
BIOCHEMICAL IMMUNIZATION OF PLANTS
The HCHO cycle as a fundamentally new finding can be used for the inductionof disease resistance in plants against biotrophic fungi using endogenous N-methylated substances as HCHO-donating inducers.20
FIGURE 8 illustrates the effect of D,L-carnitine on infection frequency in beanplants to Uromyces phaseoli. It can be seen that a relatively long induction time (6days) results in two dose-dependent, induced-resistance ranges (10–3–10–5 mol/land 10–9–10–10 mol/l of D,L-carnitine) to biotrophic fungi.
Using other N-methylated substances and other host–parasite relationshipsthis concentration dependent, aspecific “double immune response” could also
268 ANNALS NEW YORK ACADEMY OF SCIENCES
FIGURE 8. Effect of D,L-carnitine on the infection frequency of Uromyces phaseoli at beanplants. Bean variety: Saxa. Time interval between pretreatment and infection: 6 days. Fifth-degree polynomial regression analysis.

be demonstrated in every case (e.g., 1′-methyl-ascorbigen, wheat–Pucciniarelationship21).
There is a time- and dose-dependent induced resistance using TML or anotherendogenous N-methylated substance as a natural inducer.22 It seems that there isa “pharmacological” dose-dependent induced resistance (high concentration) anda “physiological” dose-dependent induced resistance (very low concentration23;that is, the resistance phase of the stress syndrome is divisible for two ranges.
FORMALDEHYDE AS A REGULATOR OF OSCILLATIONS IN OXIGEN-AND PHOTO-RELATED PROCESS OF DEMETHYLATION
The oscillation changes in accumulation of oxygen by cells and in concentrationof reaction products (e.g., vanillate, formaldehyde) as well as in the level of effec-tors (e.g., H2O2 and free radicals) were observed during a 24-hour period of incu-bation of Pseudococcus erythropolis cells with K-veratrate. The close relationshipbetween the demethylation process and the rate of aeration had been observed.24
REFERENCES
1. HUSZTI, Z. & E. TYIHÁK. 1986. Formation of formaldehyde from S-adenosyl-L-(methyl-3H)methionine during enzymatic transmethylation of histamine. FEBS Lett. 209: 362–366.
2. CHELVARAJAN, R. L., et al. 1993. Study of nicotine demethylation in Nicotiana otophora.J. Agric. Food Chem. 41: 858–862.
3. TYIHÁK, E., et al. 1992. Formaldehyde cycle and the cell proliferation: Plant tissue as amodel. In Proceedings of the Third International Conference on the Role ofFormaldehyde in Biological Systems. E. Tyihák, Ed.: 139–144. Hungarian BiochemicalSociety. Budapest, Hungary.
4. BURGYÁN, J., et al. 1982. Increased formaldehyde production from L-methionine (S-14CH3) by crude enzyme of TMV infected tobacco leaves. Acta Phytopathol. Acad. Sci.Hung. 17: 11–15.
5. TYIHÁK, E., et al. 1989. Temperature-dependent formaldehyde metabolism in beanplants. The heat shock response. Plant Sci. 59: 133–139.
6. TYIHÁK, E. 1987. Is there a formaldehyde cycle in biological systems? In Proceedings ofthe Second International Conference on the Role of Formaldehyde in BiologicalSystems. Keszthely, E. Tyihák and G. Gullner, Eds.: 137–144. SOTE Press. Hungary.
7. TYIHÁK , E., et al. 1993. Formaldehyde cycle and possibility of formation of singlet oxi-gen in plant tissue. In Proceedings of the International Symposium on Oxygen FreeRadicals and Scavangers in the Natural Sciences. Gy Mózsik, et al., Eds.: 21–28.Akadémiai Kiadó. Budapest, Hungary.
8. TYIHÁK, E., et al. 1994. Possibility of formation of excited formaldehyde and singlet oxy-gen in biotic and abiotic stress situations. Acta Biol. Hung. 45: 3–10.
9. TYIHÁK, E. 1997. Study of formaldehyde cycle in budding and apoptosis of trees by per-sonal OPLC. In Chromatography. O. Kaiser et al., Eds.: 333–342. InCom Sonderband.Düsseldorf, Germany.
10. SÁRDI, É. & E. TYIHÁK. 1994. Simple determination of formaldehyde in dimedone adductform in biological samples by high performance liquid chromatography. Biomed.Chromatogr. 8: 313–314.
11. TYIHÁK, E., et al. 1996. Formaldehyde, as its dimedone adduct, from Ascophyllumnodosum. J. Appl. Phycol. 8: 211–215.
TYIHÁK et al.: FORMALDEHYDE CYCLE 269

12. LÁSZLÓ, I. & É. SZOKE. 1997. Collected abstracts of Stress of Life: Stress and Adaptationfrom Molecules to Man. International Congress (Budapest): #186. July 1–5, 1997.Budapest, Hungary.
13. SÁRDI, É. & J. BALLA. 1997. Collected abstracts of Stress of Life: Stress and Adaptationfrom Molecules to Man. International Congress (Budapest): #180. July 1–5, 1997.Budapest, Hungary.
14. ALBERT, L., et al. 1997. Collected abstracts of Stress of Life: Stress and Adaptation fromMolecules to Man. International Congress (Budapest): #180. July 1–5, 1997. Budapest,Hungary.
15. TRÉZL, L. & J. PIPEK. 1988. Formation of excited formaldehyde in model reactions simu-lating real biological systems. J. Mol. Struct. (Theochem.) 170: 213–223.
16 TRÉZL, L., et al. 1989. Enforcing effect of basic terminal group of L-lysine in the reactionsbetween formaldehyde and hydrogen peroxide. Magy. Kém. Folyóirat (HungarianChem. J.) 95: 131–139.
17. SPASETTO, P., et al. 1995. Human eosinophil peroxidase enhances tumor necrosis factorand hydrogen peroxide release by human monocyte-derived macrophages. Eur. J.Immunol. 25: 1366–1374.
18. KERR, J. F., et al. 1972. Apoptosis: A basic biological phenomenon with wide-rangingimplications in tissue kinetics. Br. J. Cancer 26: 239–257.
19. SZENDE, B., et al. 1995. Possible role of formaldehyde in the apoptotic and mitotic effectof 1-methyl-ascorbigen. Pathol. Oncol. Res. 1: 38–42.
20. TYIHÁK, E., et al. 1989. Induction of disease resistance by Nε-trimethyl-L-lysine in beanplants against Uromyces phaseoli. J. Phytopathol. 126: 253–256.
21. KÁTAY, GY., et al. 1997. Collected abstracts of Stress of Life: Stress and Adaptation fromMolecules to Man. International Congress (Budapest): #187. July 1–5, 1997. Budapest,Hungary.
22. TYIHÁK, E., et al. 1995. Formaldehyde cycle, “double immune response” and biochemi-cal immunization of plants. In Proceedings of the Third Conference of the EuropeanFoundation for Plant Pathology (EFPP). The Polish Phytopathology Society, Poznan.M. Manka, Ed.: 571–574.
23. TYIHÁK, E., et al. 1990. In Protein Methylation. W. K. Paik & S. Kim, Eds.: 364–388. CRCPress. Boca Raton, FL.
24. PAZDZIOCH, M., et al. 1997. Collected abstracts of Stress of Life: Stress and Adaptationfrom Molecules to Man. International Congress (Budapest): #187. July 1–5, 1997.Budapest, Hungary.
270 ANNALS NEW YORK ACADEMY OF SCIENCES




















