
Expression Analysis of the Prostaglandin E 2 Production Pathway in Human Pancreatic Cancers
7
Expression Analysis of the Prostaglandin E 2 Production Pathway in Human Pancreatic Cancers Sascha Hasan, MS,* Makoto Satake, MD, PhD,* David W. Dawson, MD, PhD,Þþ Hitoshi Funahashi, MD, PhD,* Eliane Angst, MD,*§ Vay Liang W. Go, MD,|| Howard A. Reber, MD,*þ Oscar J. Hines, MD,*þ and Guido Eibl, MD*þ Objectives: There is strong evidence for an important role of cyclooxygenase (COX) 2 and COX-2Ygenerated PGE 2 during pancreatic tumorigenesis. Cyclooxygenase 2 has therefore become a potential chemotherapeutic target for pancreatic cancer. However, recent studies raised concerns regarding the safety of selective COX-2 inhibitors. Although the benefits of COX-2 inhibition may eventually outweigh the associated cardiovascular risks, there are a number of alternative targets for inhibiting the formation of PGE 2 in human tumors that may prove less harmful to the patient. This study aimed at analyzing the expression of various proteins involved in the generation of PGE 2 in human pancreatic cancers. Methods and Results: Real-time polymerase chain reaction and Western blot analyses demonstrated overexpression of cytoplasmic phospholipase A2, COX-2, cytoplasmic prostaglandin E synthase, and microsomal prostaglandin E synthases 1 and 2 in most human pancreatic cancers when compared with matched normal pancreas. Immunohistochemistry revealed expression of these proteins predomi- nantly by pancreatic cancer cells. Variable expression of these proteins was also confirmed in several human pancreatic cancer cell lines. Conclusions: Our studies demonstrated for the first time that various proteins involved in the generation of PGE 2 are overexpressed in human pancreatic cancers. These proteins may represent potentially novel targets for the therapy of pancreatic cancers. Key Words: pancreatic cancer, prostaglandin E 2 , COX-2, PGE synthases, phospholipase A2 (Pancreas 2008;37:121Y127) P ancreatic ductal adenocarcinoma is an extremely lethal disease that is characterized by its propensity to infiltrate adjacent tissue and to metastasize even at early stages. 1 Absence of specific symptoms, lack of early detection markers, aggressive tumor growth, and resistance to conven- tional chemotherapy and radiotherapy conspire to culminate in a median overall survival rate of less than 9 months and annual mortality figures virtually equaling incidence numbers. 2 Surgery is the only potentially curative option, but only 10% to 15% of patients at the time of diagnosis are candidates for surgical resection. As of today, only very few chemothera- peutic treatment options exist, which disappointingly offer only a marginal survival benefit to the patients. In recent years, cyclooxygenase (COX) 2, the inducible isoform of COX enzymes, has gained strong interest as a potential molecular target in the therapy and prevention of human cancers. 3,4 The protumorigenic properties of COX-2 are generally thought to be mediated by COX-2Ygenerated prostanoids, of which prostaglandin E 2 (PGE 2 ) is the most abundant prostanoid species in many tumors. 4 Prostaglandin E 2 has been shown to promote tumor growth and spread by multiple mechanisms, including stimulation of cellular pro- liferation, invasion, and angiogenesis, inhibition of apoptotic cell death, and modulation of immune response. 4 In pancreatic cancers, COX-2 and COX-2Yderived PGE 2 have been found to stimulate growth, invasion, and angiogenesis. 5Y7 The generation of PGE 2 begins with the liberation of arachidonic acid (AA), an omega-6 polyunsaturated fatty acid, from the sn-2 position of major membrane phospholipids by phospholipase A 2 enzymes (PLA 2 ), in particular by the 85- to 110-kDa group IV cytosolic PLA 2 (cPLA 2 ). 8 Arachidonic acid is the preferred substrate for the 72-kDa COX enzymes, which catalyze the oxygenation and reduction of AA to the unstable hydroxy endoperoxide PGH 2 . Although the COX-1 isoform is normally expressed constitutively in most mamma- lian tissues, the COX-2 enzyme can be rapidly induced by various inflammatory and oncogenic stimuli. 8 Isomerization of PGH 2 to PGE 2 is catalyzed by specific synthases, namely microsomal PGE 2 synthase (mPGES) 1, mPGES-2, and cytoplasmic PGES (cPGES). 9 Microsomal PGES-1 is a 15- to 16-kDa membrane-associated protein, the expression of which is often concordantly induced together with COX-2 and that seems to preferentially couple with COX-2 to generate PGE 2 . 8 In contrast, the 26-kDa cytoplasmic PGES seems to couple preferentially to COX-1, and its expression generally is unaffected by inflammatory stimuli. 8 The membrane- associated 33-kDa mPGES-2 is synthesized as a Golgi membraneYassociated protein, and the proteolytic removal of ORIGINAL ARTICLE Pancreas & Volume 37, Number 2, August 2008 121 Received for publication August 8, 2007; accepted December 20, 2007. From the *Department of Surgery, Hirshberg Laboratories for Pancreatic Cancer Research; †Department of Pathology and Laboratory Medicine, and ‡Jonsson Comprehensive Cancer Center, David Geffen School of Medicine at UCLA, Los Angeles, CA; §Department of Visceral and Transplant Surgery, University Hospital, Berne, Switzerland; and ||Depart- ment of Medicine, David Geffen School of Medicine at UCLA, Los Angeles, CA. This work was supported by National Institutes of Health Grants R01 CA104027, R01 CA122042, and P01 AT003960 (to G.E. and V.L.W. Go), the Hirshberg Foundation for Pancreatic Cancer Research (H.A.R., D.W.D.), and a personal grant from the Swiss Scientific Foundation (PBZHB-117008; E.A.). The Tissue Procurement Core Laboratory services were partially funded by Grant CA16042 from the National Cancer Institute. Reprints: Guido Eibl, MD, Department of Surgery, David Geffen School of Medicine at UCLA, 675 Charles E. Young Drive South, MRL 2535, Los Angeles, CA 90095 (e-mail: [email protected]). Copyright * 2008 by Lippincott Williams & Wilkins Copyright @ 2008 Lippincott Williams & Wilkins. Unauthorized reproduction of this article is prohibited.
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Expression Analysis of the Prostaglandin E 2 Production Pathway in Human Pancreatic Cancers

Expression Analysis of the Prostaglandin E2 ProductionPathway in Human Pancreatic Cancers
Sascha Hasan, MS,* Makoto Satake, MD, PhD,* David W. Dawson, MD, PhD,ÞþHitoshi Funahashi, MD, PhD,* Eliane Angst, MD,*§ Vay Liang W. Go, MD,|| Howard A. Reber, MD,*þ
Oscar J. Hines, MD,*þ and Guido Eibl, MD*þ
Objectives: There is strong evidence for an important role ofcyclooxygenase (COX) 2 and COX-2Ygenerated PGE2 duringpancreatic tumorigenesis. Cyclooxygenase 2 has therefore becomea potential chemotherapeutic target for pancreatic cancer. However,recent studies raised concerns regarding the safety of selective COX-2inhibitors. Although the benefits of COX-2 inhibition may eventuallyoutweigh the associated cardiovascular risks, there are a number ofalternative targets for inhibiting the formation of PGE2 in humantumors that may prove less harmful to the patient. This study aimed atanalyzing the expression of various proteins involved in thegeneration of PGE2 in human pancreatic cancers.
Methods and Results: Real-time polymerase chain reaction andWestern blot analyses demonstrated overexpression of cytoplasmicphospholipase A2, COX-2, cytoplasmic prostaglandin E synthase, andmicrosomal prostaglandin E synthases 1 and 2 in most humanpancreatic cancers when compared with matched normal pancreas.Immunohistochemistry revealed expression of these proteins predomi-nantly by pancreatic cancer cells. Variable expression of these proteinswas also confirmed in several human pancreatic cancer cell lines.
Conclusions: Our studies demonstrated for the first time that variousproteins involved in the generation of PGE2 are overexpressed inhuman pancreatic cancers. These proteins may represent potentiallynovel targets for the therapy of pancreatic cancers.
Key Words: pancreatic cancer, prostaglandin E2, COX-2,PGE synthases, phospholipase A2
(Pancreas 2008;37:121Y127)
Pancreatic ductal adenocarcinoma is an extremely lethaldisease that is characterized by its propensity to infiltrate
adjacent tissue and to metastasize even at early stages.1
Absence of specific symptoms, lack of early detectionmarkers, aggressive tumor growth, and resistance to conven-tional chemotherapy and radiotherapy conspire to culminate ina median overall survival rate of less than 9 months and annualmortality figures virtually equaling incidence numbers.2
Surgery is the only potentially curative option, but only 10%to 15% of patients at the time of diagnosis are candidates forsurgical resection. As of today, only very few chemothera-peutic treatment options exist, which disappointingly offeronly a marginal survival benefit to the patients.
In recent years, cyclooxygenase (COX) 2, the inducibleisoform of COX enzymes, has gained strong interest as apotential molecular target in the therapy and prevention ofhuman cancers.3,4 The protumorigenic properties of COX-2are generally thought to be mediated by COX-2Ygeneratedprostanoids, of which prostaglandin E2 (PGE2) is the mostabundant prostanoid species in many tumors.4 ProstaglandinE2 has been shown to promote tumor growth and spread bymultiple mechanisms, including stimulation of cellular pro-liferation, invasion, and angiogenesis, inhibition of apoptoticcell death, and modulation of immune response.4 In pancreaticcancers, COX-2 and COX-2Yderived PGE2 have been found tostimulate growth, invasion, and angiogenesis.5Y7
The generation of PGE2 begins with the liberation ofarachidonic acid (AA), an omega-6 polyunsaturated fatty acid,from the sn-2 position of major membrane phospholipids byphospholipase A2 enzymes (PLA2), in particular by the 85- to110-kDa group IV cytosolic PLA2 (cPLA2).
8 Arachidonicacid is the preferred substrate for the 72-kDa COX enzymes,which catalyze the oxygenation and reduction of AA to theunstable hydroxy endoperoxide PGH2. Although the COX-1isoform is normally expressed constitutively in most mamma-lian tissues, the COX-2 enzyme can be rapidly induced byvarious inflammatory and oncogenic stimuli.8 Isomerization ofPGH2 to PGE2 is catalyzed by specific synthases, namelymicrosomal PGE2 synthase (mPGES) 1, mPGES-2, andcytoplasmic PGES (cPGES).9 Microsomal PGES-1 is a 15-to 16-kDa membrane-associated protein, the expression ofwhich is often concordantly induced together with COX-2 andthat seems to preferentially couple with COX-2 to generatePGE2.
8 In contrast, the 26-kDa cytoplasmic PGES seemsto couple preferentially to COX-1, and its expression generallyis unaffected by inflammatory stimuli.8 The membrane-associated 33-kDa mPGES-2 is synthesized as a GolgimembraneYassociated protein, and the proteolytic removal of
ORIGINAL ARTICLE
Pancreas & Volume 37, Number 2, August 2008 121
Received for publication August 8, 2007; accepted December 20, 2007.From the *Department of Surgery, Hirshberg Laboratories for Pancreatic
Cancer Research; †Department of Pathology and Laboratory Medicine,and ‡Jonsson Comprehensive Cancer Center, David Geffen School ofMedicine at UCLA, Los Angeles, CA; §Department of Visceral andTransplant Surgery, University Hospital, Berne, Switzerland; and ||Depart-ment of Medicine, David Geffen School of Medicine at UCLA, LosAngeles, CA.
This work was supported by National Institutes of Health Grants R01CA104027, R01 CA122042, and P01 AT003960 (to G.E. and V.L.W. Go),the Hirshberg Foundation for Pancreatic Cancer Research (H.A.R.,D.W.D.), and a personal grant from the Swiss Scientific Foundation(PBZHB-117008; E.A.). The Tissue Procurement Core Laboratoryservices were partially funded by Grant CA16042 from the NationalCancer Institute.
Reprints: Guido Eibl, MD, Department of Surgery, David Geffen School ofMedicine at UCLA, 675 Charles E. Young Drive South, MRL 2535, LosAngeles, CA 90095 (e-mail: [email protected]).
Copyright * 2008 by Lippincott Williams & Wilkins
Copyright @ 2008 Lippincott Williams & Wilkins. Unauthorized reproduction of this article is prohibited.
the N-terminal hydrophobic domain leads to the formation ofa mature cytosolic enzyme.10 Microsomal PGE2 synthase 2 isconstitutively expressed in most tissues and has been shown tocouple with both COX-1 and COX-2, with a presumablymodest preference for COX-2.8 In contrast to the abundantliterature on the role of COX-2 in human cancer biology,reports describing the importance of cPLA2, mPGES-1,mPGES-2, and cPGES in human cancers are scarce. Elevatedlevels of cPLA2 and mPGES-1 are generally found to correlatewith COX-2 expression and PGE2 production in experimentalmodels and human cancers.11Y15 Although cPGES andmPGES-2 are generally thought of being expressed constitu-tively in most tissues, elevated levels of mPGES-2 have beendescribed in colorectal cancer specimens, possibly indicating afunctional role in cancer biology.16
To date, most studies have focused on inhibiting COX-2in an effort to reduce protumorigenic PGE2 levels. However,reasonable concerns about the cardiovascular safety of se-lective COX-2 inhibitors seem to warrant a broader analysis ofalternative proteins involved in the generation of PGE2.
17Y19
Although the biological significance of COX-2 and COX-2Ygenerated PGE2 in pancreatic cancers is widely recognized,a comprehensive analysis of enzymes involved in PGE2
production other than COX-2 in human pancreatic cancers hasnot been reported.
MATERIALS AND METHODS
ReagentsThe following antibodies were used for immunoblotting
and immunohistochemistry: rabbit polyclonal cPLA2 (no. 2832)from Cell Signaling Technology, Inc. (Danvers, Mass); rabbitmonoclonal COX-2 (clone SP21) from Lab Vision (Fremont,Calif ); rabbit polyclonal mPGES-1 (no. 160140), rabbit poly-clonal mPGES-2 (no. 160145), and rabbit polyclonal cPGES (no.160150) from Cayman Chemical (Ann Arbor, Mich).
Cell CultureThe human pancreatic cancer cell lines AsPC-1 (well to
poorly differentiated), BxPC-3 (well to poorly differentiated),Capan-2 (well differentiated), HPAF-II (moderately differen-tiated), MIA PaCa-2 (undifferentiated), and PANC-1 (poorlydifferentiated) were obtained from the American Type CultureCollection (Rockville, Md) and cultured as describedpreviously.6
Tissue SamplesAll studies were approved by the institutional review
board. Seven fresh-frozen tissue samples of resected humanductal adenocarcinomas and matched, histologically tumor-free resection margins were obtained from the PancreaticTumor Tissue Bank at University of California at Los Angeles.The histopathology of all specimens was confirmed by agastrointestinal pathologist (D.W.D.). Frozen tissues werepulverized in liquid nitrogen with a mortar and pestle andimmediately processed for protein and RNA extraction.
RNA IsolationTotal RNA was extracted using the Aurum total RNA
mini kit (Bio-Rad Laboratories, Hercules, Calif ). RNA qualityand quantity were measured using the Experion automatedelectrophoresis system (Bio-Rad). RNA concentrations wereadditionally confirmed spectrophotometrically.
Real-Time Polymerase Chain ReactionRNA (0.5 Kg) was reverse transcribed using the iScript
cDNA synthesis kit (Bio-Rad). Real-time polymerase chainreaction (PCR) was performed on an iQ5 real-time PCRdetection system (Bio-Rad) using SYBR Green chemistry andspecific intron-spanning primers designed by Beacon Designer6.0 (PREMIER Biosoft International, Palo Alto, Calif ).Optimal primer concentrations and cycling conditions weredetermined for each primer set (Table 1). Polymerase chainreaction efficiencies for each primer pair were calculated froma standard dilution series. Amplification of unspecific productswas excluded by melt curve analysis and agarose electrophor-esis of PCR products. Gene expression was normalized toglyceraldehyde-3-phosphate dehydrogenase as a housekeep-ing gene and calculated using the $$CT method.
Protein ExtractionPulverized frozen tissues were homogenized (Tissuemi-
ser; Fisher Scientific, Chino, Calif ) and sonicated (Vibra-Cell;Sonics & Materials, Inc., Newtown, Conn) in phosphate-buffered saline containing the Complete protease inhibitorcocktail (Roche Applied Science, Indianapolis, Ind). Thehomogenate was centrifuged, and the protein concentration inthe supernatant was determined by the BCA protein assay kit(Pierce, Rockford, Ill).
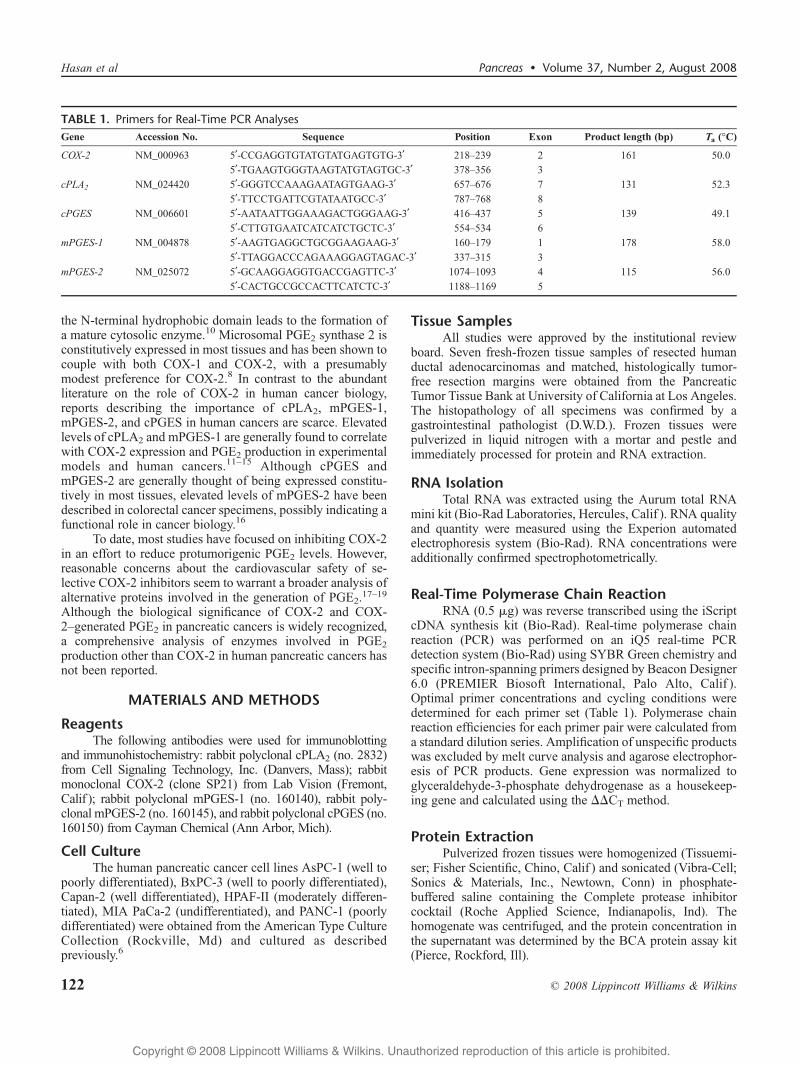
TABLE 1. Primers for Real-Time PCR AnalysesGene Accession No. Sequence Position Exon Product length (bp) Ta (-C)
COX-2 NM_000963 5¶-CCGAGGTGTATGTATGAGTGTG-3¶ 218Y239 2 161 50.0
5¶-TGAAGTGGGTAAGTATGTAGTGC-3¶ 378Y356 3
cPLA2 NM_024420 5¶-GGGTCCAAAGAATAGTGAAG-3¶ 657Y676 7 131 52.3
5¶-TTCCTGATTCGTATAATGCC-3¶ 787Y768 8
cPGES NM_006601 5¶-AATAATTGGAAAGACTGGGAAG-3¶ 416Y437 5 139 49.1
5¶-CTTGTGAATCATCATCTGCTC-3¶ 554Y534 6
mPGES-1 NM_004878 5¶-AAGTGAGGCTGCGGAAGAAG-3¶ 160Y179 1 178 58.0
5¶-TTAGGACCCAGAAAGGAGTAGAC-3¶ 337Y315 3
mPGES-2 NM_025072 5¶-GCAAGGAGGTGACCGAGTTC-3¶ 1074Y1093 4 115 56.0
5¶-CACTGCCGCCACTTCATCTC-3¶ 1188Y1169 5
Hasan et al Pancreas & Volume 37, Number 2, August 2008
122 * 2008 Lippincott Williams & Wilkins
Copyright @ 2008 Lippincott Williams & Wilkins. Unauthorized reproduction of this article is prohibited.
Western BlottingTotal cell lysates were fractionated on sodium dodecyl
sulfateYpolyacrylamide gel electrophoresis and transferred tonitrocellulose membranes. Cyclooxygenase 2, cPLA2,mPGES-1, mPGES-2, and cPGES were detected usingspecific antibodies, and horseradish peroxidaseYconjugatedantimouse or antirabbit immunoglobulins as secondaryantibodies (Pierce). Protein-antibody complexes were visua-lized with the SuperSignal West Pico ChemiluminescentSubstrate (Pierce).
ImmunohistochemistryParaffin-embedded tissue samples were sectioned,
deparaffinized in xylene, and rehydrated in a graded ethanolseries. Sections were subjected to heat-induced epitoperetrieval using boiling citrate buffer (pH 6.0). Endogenousperoxidase activity was quenched by 0.3% H2O2, andunspecific binding sites were blocked with 5% bovine serumalbumin. Immunostainings were performed with primaryantibodies against COX-2 (1:100), cPGES (1 Kg/mL),mPGES-1 (2 Kg/mL), and mPGES-2 (2 Kg/mL) for 1 hourat room temperature and cPLA2 (2 Kg/mL) overnight at 4-C.
Immunocomplexes were visualized by corresponding horse-radish peroxidaseYconjugated secondary antibodies and 3-amino-9-ethylcarbazole as a chromogen using the UltraVisiondetection system (Lab Vision). Sections were counterstainedwith Hematoxyllin QS (Vector Laboratories, Burlingame,Calif ). As a control, nonspecific antibodies at a concentrationcorresponding to that of the primary antibodies were used tostain all sections using the identical protocol.
Statistical AnalysisData are presented as mean T SD. Differences in the
mean of 2 samples were analyzed by an unpaired t test. An >
value of 0.05 was used to determine significant differences.All statistics were done in SigmaStat 3.1 (Systat Software,Inc., Chicago, Ill).
RESULTSTo determine the expression pattern of several enzymes
involved in PGE2 biosynthesis in human pancreatic cancers,7 pancreatic ductal adenocarcinoma tissue samples fromsurgically resected tumor specimens were obtained from theUniversity of California at Los Angeles Pancreatic Cancer
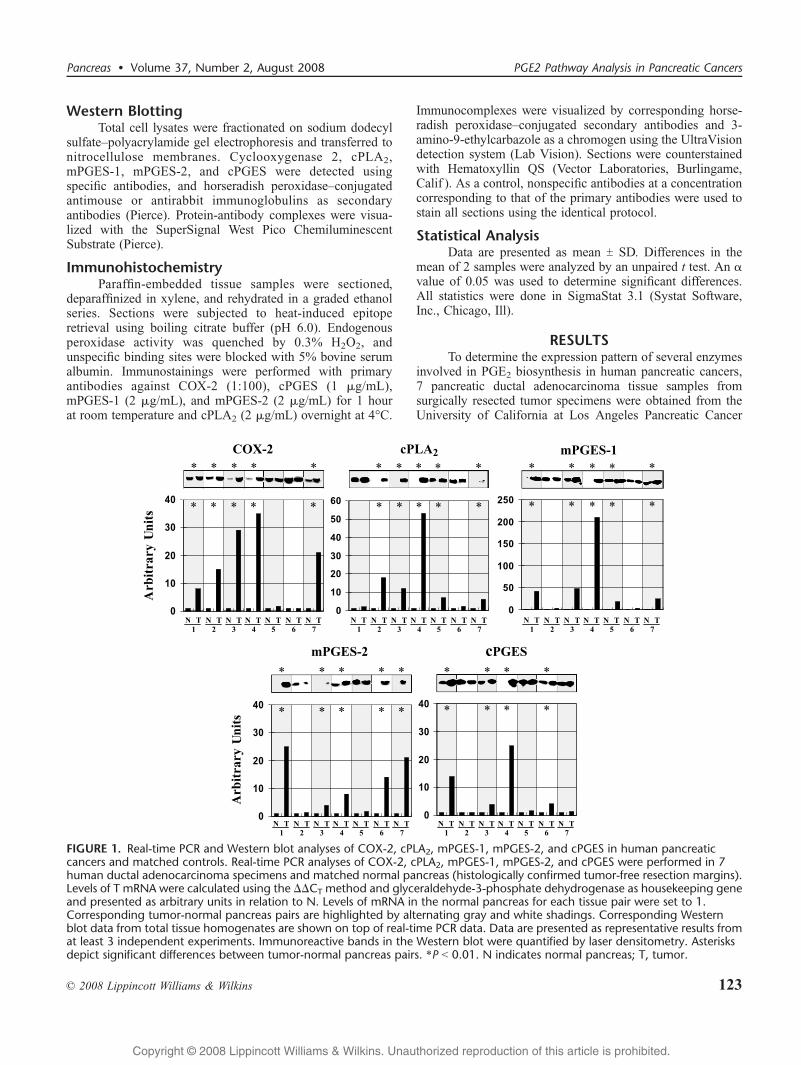
FIGURE 1. Real-time PCR and Western blot analyses of COX-2, cPLA2, mPGES-1, mPGES-2, and cPGES in human pancreaticcancers and matched controls. Real-time PCR analyses of COX-2, cPLA2, mPGES-1, mPGES-2, and cPGES were performed in 7human ductal adenocarcinoma specimens and matched normal pancreas (histologically confirmed tumor-free resection margins).Levels of T mRNA were calculated using the $$CT method and glyceraldehyde-3-phosphate dehydrogenase as housekeeping geneand presented as arbitrary units in relation to N. Levels of mRNA in the normal pancreas for each tissue pair were set to 1.Corresponding tumor-normal pancreas pairs are highlighted by alternating gray and white shadings. Corresponding Westernblot data from total tissue homogenates are shown on top of real-time PCR data. Data are presented as representative results fromat least 3 independent experiments. Immunoreactive bands in the Western blot were quantified by laser densitometry. Asterisksdepict significant differences between tumor-normal pancreas pairs. *P G 0.01. N indicates normal pancreas; T, tumor.
Pancreas & Volume 37, Number 2, August 2008 PGE2 Pathway Analysis in Pancreatic Cancers
* 2008 Lippincott Williams & Wilkins 123
Copyright @ 2008 Lippincott Williams & Wilkins. Unauthorized reproduction of this article is prohibited.
Tissue Bank. Each tumor sample was matched with thehistologically tumor-free pancreatic resection margin of thesame patient. These resection margin samples were consideredBnormal[ and used to compare expression patterns betweentumor and normal pancreas. The tissue specimens were againevaluated by a gastrointestinal pathologist to confirm histo-pathology. All tumor samples comprised 20% to 40%malignant cells, whereas some normal resection margintissues contained variably low-grade pancreatic intraepithelialneoplasias (PanINs) besides predominantly normal pancreaticparenchyma. Real-time PCR analysis of total RNA extractedfrom the whole tissue samples demonstrated a statisticallysignificant overexpression of COX-2 in 5 of 7 tissue pairs(5/7), cPLA2 (5/7), mPGES-1 (5/7), mPGES-2 (5/7), andcPGES (4/7) in tumor tissues compared with matched normalpancreas (Fig. 1). A greater than 10-fold increase in COX-2,cPLA2, mPGES-1, mPGES-2, and cPGES mRNA levels wasdetected in 4, 3, 5, 3, and 2 tumor samples, respectively.Interestingly, 2 tumor samples (3 and 4) had a significantmRNA overexpression of all 5 enzymes tested, whereas tumorsamples 2, 5, and 6 had an increase in mRNA of only 2enzymes (Fig. 1). Real-time PCR data were validated byWestern blot analyses of total tissue homogenates. Again,compared with matched normal pancreas, overexpression ofCOX-2, cPLA2, mPGES-1, mPGES-2, and cPGES protein was
seen in 5, 5, 5, 5, and 4 tumors, respectively (Fig. 1). Tumors 3and 4 displayed significant overexpression of all 5 proteins.Western blot analyses of A-actin demonstrated equal proteinloading (not shown).
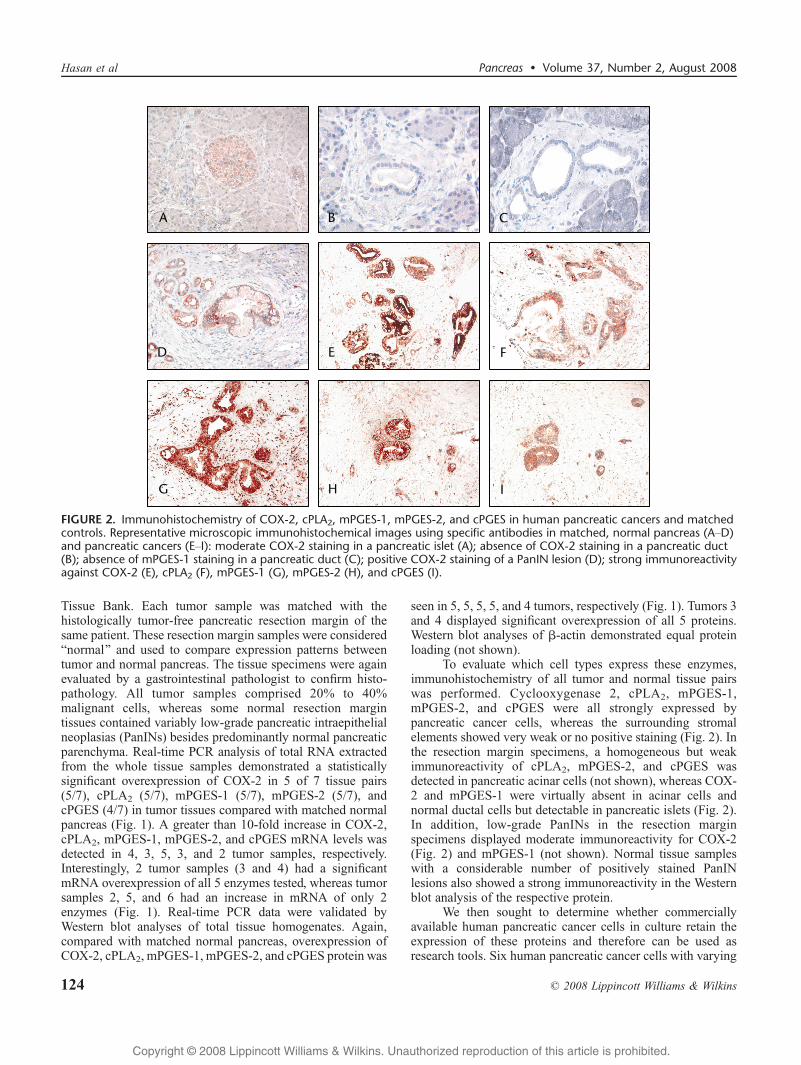
To evaluate which cell types express these enzymes,immunohistochemistry of all tumor and normal tissue pairswas performed. Cyclooxygenase 2, cPLA2, mPGES-1,mPGES-2, and cPGES were all strongly expressed bypancreatic cancer cells, whereas the surrounding stromalelements showed very weak or no positive staining (Fig. 2). Inthe resection margin specimens, a homogeneous but weakimmunoreactivity of cPLA2, mPGES-2, and cPGES wasdetected in pancreatic acinar cells (not shown), whereas COX-2 and mPGES-1 were virtually absent in acinar cells andnormal ductal cells but detectable in pancreatic islets (Fig. 2).In addition, low-grade PanINs in the resection marginspecimens displayed moderate immunoreactivity for COX-2(Fig. 2) and mPGES-1 (not shown). Normal tissue sampleswith a considerable number of positively stained PanINlesions also showed a strong immunoreactivity in the Westernblot analysis of the respective protein.
We then sought to determine whether commerciallyavailable human pancreatic cancer cells in culture retain theexpression of these proteins and therefore can be used asresearch tools. Six human pancreatic cancer cells with varying
FIGURE 2. Immunohistochemistry of COX-2, cPLA2, mPGES-1, mPGES-2, and cPGES in human pancreatic cancers and matchedcontrols. Representative microscopic immunohistochemical images using specific antibodies in matched, normal pancreas (AYD)and pancreatic cancers (EYI): moderate COX-2 staining in a pancreatic islet (A); absence of COX-2 staining in a pancreatic duct(B); absence of mPGES-1 staining in a pancreatic duct (C); positive COX-2 staining of a PanIN lesion (D); strong immunoreactivityagainst COX-2 (E), cPLA2 (F), mPGES-1 (G), mPGES-2 (H), and cPGES (I).
Hasan et al Pancreas & Volume 37, Number 2, August 2008
124 * 2008 Lippincott Williams & Wilkins
Copyright @ 2008 Lippincott Williams & Wilkins. Unauthorized reproduction of this article is prohibited.
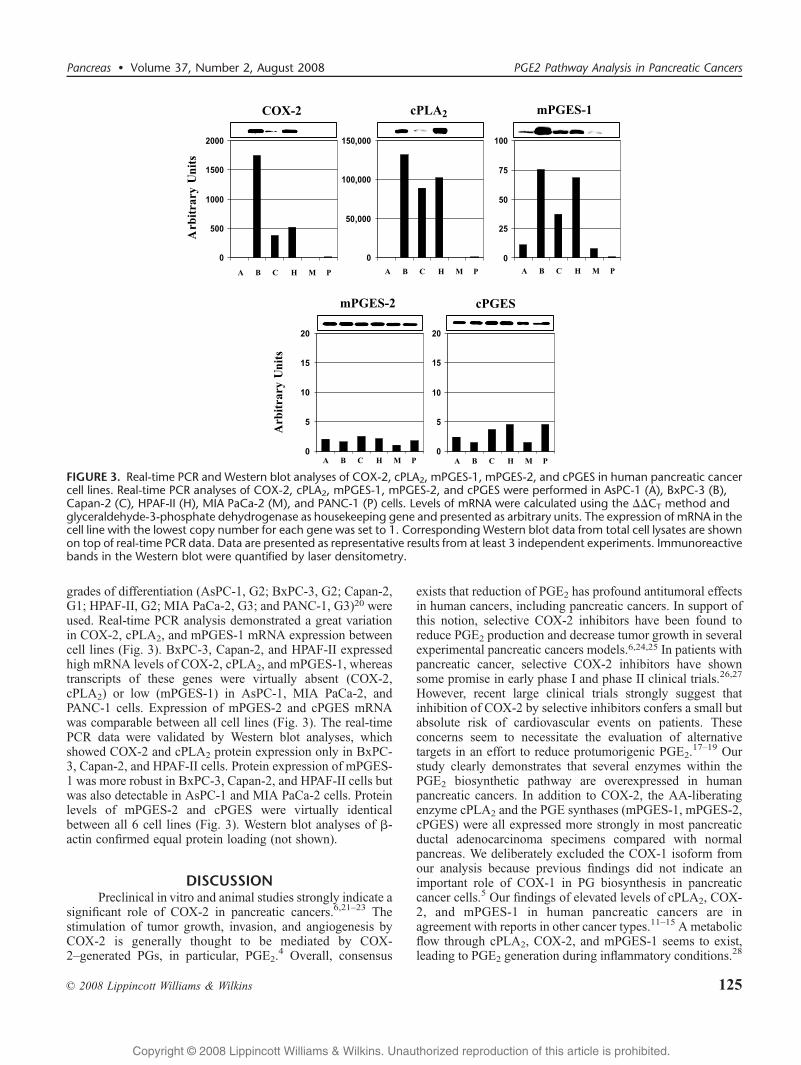
grades of differentiation (AsPC-1, G2; BxPC-3, G2; Capan-2,G1; HPAF-II, G2; MIA PaCa-2, G3; and PANC-1, G3)20 wereused. Real-time PCR analysis demonstrated a great variationin COX-2, cPLA2, and mPGES-1 mRNA expression betweencell lines (Fig. 3). BxPC-3, Capan-2, and HPAF-II expressedhigh mRNA levels of COX-2, cPLA2, and mPGES-1, whereastranscripts of these genes were virtually absent (COX-2,cPLA2) or low (mPGES-1) in AsPC-1, MIA PaCa-2, andPANC-1 cells. Expression of mPGES-2 and cPGES mRNAwas comparable between all cell lines (Fig. 3). The real-timePCR data were validated by Western blot analyses, whichshowed COX-2 and cPLA2 protein expression only in BxPC-3, Capan-2, and HPAF-II cells. Protein expression of mPGES-1 was more robust in BxPC-3, Capan-2, and HPAF-II cells butwas also detectable in AsPC-1 and MIA PaCa-2 cells. Proteinlevels of mPGES-2 and cPGES were virtually identicalbetween all 6 cell lines (Fig. 3). Western blot analyses of A-actin confirmed equal protein loading (not shown).
DISCUSSIONPreclinical in vitro and animal studies strongly indicate a
significant role of COX-2 in pancreatic cancers.6,21Y23 Thestimulation of tumor growth, invasion, and angiogenesis byCOX-2 is generally thought to be mediated by COX-2Ygenerated PGs, in particular, PGE2.
4 Overall, consensus
exists that reduction of PGE2 has profound antitumoral effectsin human cancers, including pancreatic cancers. In support ofthis notion, selective COX-2 inhibitors have been found toreduce PGE2 production and decrease tumor growth in severalexperimental pancreatic cancers models.6,24,25 In patients withpancreatic cancer, selective COX-2 inhibitors have shownsome promise in early phase I and phase II clinical trials.26,27
However, recent large clinical trials strongly suggest thatinhibition of COX-2 by selective inhibitors confers a small butabsolute risk of cardiovascular events on patients. Theseconcerns seem to necessitate the evaluation of alternativetargets in an effort to reduce protumorigenic PGE2.
17Y19 Ourstudy clearly demonstrates that several enzymes within thePGE2 biosynthetic pathway are overexpressed in humanpancreatic cancers. In addition to COX-2, the AA-liberatingenzyme cPLA2 and the PGE synthases (mPGES-1, mPGES-2,cPGES) were all expressed more strongly in most pancreaticductal adenocarcinoma specimens compared with normalpancreas. We deliberately excluded the COX-1 isoform fromour analysis because previous findings did not indicate animportant role of COX-1 in PG biosynthesis in pancreaticcancer cells.5 Our findings of elevated levels of cPLA2, COX-2, and mPGES-1 in human pancreatic cancers are inagreement with reports in other cancer types.11Y15 A metabolicflow through cPLA2, COX-2, and mPGES-1 seems to exist,leading to PGE2 generation during inflammatory conditions.28
FIGURE 3. Real-time PCR and Western blot analyses of COX-2, cPLA2, mPGES-1, mPGES-2, and cPGES in human pancreatic cancercell lines. Real-time PCR analyses of COX-2, cPLA2, mPGES-1, mPGES-2, and cPGES were performed in AsPC-1 (A), BxPC-3 (B),Capan-2 (C), HPAF-II (H), MIA PaCa-2 (M), and PANC-1 (P) cells. Levels of mRNA were calculated using the $$CT method andglyceraldehyde-3-phosphate dehydrogenase as housekeeping gene and presented as arbitrary units. The expression of mRNA in thecell line with the lowest copy number for each gene was set to 1. Corresponding Western blot data from total cell lysates are shownon top of real-time PCR data. Data are presented as representative results from at least 3 independent experiments. Immunoreactivebands in the Western blot were quantified by laser densitometry.
Pancreas & Volume 37, Number 2, August 2008 PGE2 Pathway Analysis in Pancreatic Cancers
* 2008 Lippincott Williams & Wilkins 125
Copyright @ 2008 Lippincott Williams & Wilkins. Unauthorized reproduction of this article is prohibited.

There is now strong evidence that COX-2 and mPGES-1 arefunctionally coupled in the cell.28,29 Colocalization of COX-2and mPGES-1 in the perinuclear membrane may allow theirefficient functional coupling. Similar to COX-2, mPGES-1can be rapidly induced by inflammatory stimuli and has beenfound to be the main PGE synthase in inflammatorydiseases.28 The importance of mPGES-1 in tumorigenesishas been highlighted by studies using COX-2/mPGES-1transgenic mice, which developed gastric tumors.30 Theseobservations led recently to a growing interest in mPGES-1 asa potential molecular target in cancer therapy and preven-tion.31 Interestingly, we also found a more robust expression ofmPGES-2 and cPGES in pancreatic cancers compared withnormal pancreas. Both enzymes are generally thought to beconstitutively expressed in most tissues and not to beappreciably elevated during inflammation or tissue damage.28
However, elevated levels of mPGES-2 have recently beenfound in colorectal cancers, suggesting a potential role in thiscancer type.16 The biological significance of the observedoverexpression of mPGES-2 and cPGES in human pancreaticcancers is unclear at the moment and needs to be validated inlarger series and experimental models. Immunohistochemistryrevealed that all enzymes are predominantly expressed by thecancer cells and not by the stromal elements, includinginflammatory cells and fibroblasts. Interestingly, the histolo-gically confirmed tumor-free resection margins, which wereused as the matched normal pancreas, variably contained low-grade PanIN-1 and PanIN-2 lesions. These PanINs werestronger positive for COX-2 and mPGES-1 compared with thesurrounding pancreatic parenchyma, which may explain inpart the strong immunoreactivity of the normal pancreassample in Western blot analyses. In fact, we think that theconsiderable differences in protein expression between thenormal tissue samples stem from the variable amount ofpancreatic acini, islets, normal ductal cells, and PanIN lesionsin these tissue samples.
We also investigated the expression of COX-2, cPLA2,mPGES-1, mPGES-2, and cPGES in 6 human pancreaticcancer cell lines. Similar to COX-2, cPLA2 and mPGES-1were strongly expressed only in BxPC-3, Capan-2, and HPAF-II cells, suggesting the importance of these enzymes as acommon metabolic pathway for PGE2 production in pancrea-tic cancer cells. In contrast to BxPC-3, Capan-2, and HPAF-II,PGE2 production in AsPC-1, MIA PaCa-2, and PANC-1 cellsis virtually undetectable.5 Accordingly, cPLA2, COX-2, andmPGES-1 are expressed only at very low levels or areundetectable in AsPC-1, MIA PaCa-2, and PANC-1 cells. Incontrast, mPGES-2 and cPGES levels are virtually identicalin all 6 cell lines. Together with our previous results, these invitro studies suggest that pancreatic cancer cells generatePGE2 through a cPLA2, COX-2, and mPGES-1 pathway.Interestingly, PGE2 production and the expression of cPLA2,COX-2, and mPGES-1 thereby seem to correlate with thedegree of cell differentiation because AsPC-1, MIA PaCa-2,and PANC-1 are generally considered to be less differentiatedthan BxPC-3, Capan-2, and HPAF-II. This is in agreementwith a previous study that described a correlation of COX-2expression and grade of differentiation in pancreatic cancercell lines.32
The limited number of analyzed samples did not allowfor meaningful correlations between the expression status ofthe examined proteins and clinicopathological parameters.However, that was not the purpose of this study, which wasessentially to determine for the first time whether enzymesinvolved in PGE2 biosynthesis in pancreatic cancers areoverexpressed; these results would then serve as a basis forfuture larger series. In summary, we described for the first timethat several enzymes involved in PGE2 biosynthesis areoverexpressed in most human pancreatic cancers and mayrepresent intriguing alternative molecular targets to reduceprotumorigenic PGE2 levels in pancreatic cancer.
ACKNOWLEDGMENTSThe authors thank the Department of Pathology’s Tissue
Procurement Core Laboratory (TPCL) at UCLA for providingthe tissue samples.
REFERENCES1. Bardeesy N, DePinho RA. Pancreatic cancer biology and genetics.
Nat Rev Cancer. 2002;2:897Y909.2. Jemal A, Siegel R, Ward E, et al. Cancer statistics, 2007. CA Cancer
J Clin. 2007;57:43Y66.3. Liao Z, Mason KA, Milas L. Cyclo-oxygenase-2 and its inhibition in
cancer: is there a role? Drugs. 2007;67:821Y845.4. Wang D, Dubois RN. Prostaglandins and cancer. Gut. 2006;55:
115Y122.5. Eibl G, Bruemmer D, Okada Y, et al. PGE(2) is generated by specific
COX-2 activity and increases VEGF production in COX-2Yexpressinghuman pancreatic cancer cells. Biochem Biophys Res Commun. 2003;306:887Y897.
6. Eibl G, Takata Y, Boros LG, et al. Growth stimulation of COX-2Ynegativepancreatic cancer by a selective COX-2 inhibitor. Cancer Res. 2005;65:982Y990.
7. Ito H, Duxbury M, Benoit E, et al. Prostaglandin E2 enhances pancreaticcancer invasiveness through an Ets-1Ydependent induction of matrixmetalloproteinase-2. Cancer Res. 2004;64:7439Y7446.
8. Park JY, Pillinger MH, Abramson SB. Prostaglandin E2 synthesisand secretion: the role of PGE2 synthases. Clin Immunol. 2006;119:229Y240.
9. Kudo I, Murakami M. Prostaglandin e synthase, a terminal enzyme forprostaglandin E2 biosynthesis. J Biochem Mol Biol. 2005;38:633Y638.
10. Tanikawa N, Ohmiya Y, Ohkubo H, et al. Identification andcharacterization of a novel type of membrane-associated prostaglandin Esynthase. Biochem Biophys Res Commun. 2002;291:884Y889.
11. Cohen EG, Almahmeed T, Du B, et al. Microsomal prostaglandin Esynthase-1 is overexpressed in head and neck squamous cell carcinoma.Clin Cancer Res. 2003;9:3425Y3430.
12. Panel V, Boelle PY, yala-Sanmartin J, et al. Cytoplasmic phospholipaseA2 expression in human colon adenocarcinoma is correlated withcyclooxygenase-2 expression and contributes to prostaglandin E2production. Cancer Lett. 2006;243:255Y263.
13. Shi Y, Cui L, Dai G, et al. Elevated prostaglandin E2 level viacPLA2YCOX-2YmPGES-1 pathway involved in bladder carcinogenesisinduced by terephthalic acid-calculi in Wistar rats. Prostaglandins LeukotEssent Fatty Acids. 2006;74:309Y315.
14. Yoshimatsu K, Altorki NK, Golijanin D, et al. Inducible prostaglandinE synthase is overexpressed in non-small cell lung cancer. Clin CancerRes. 2001;7:2669Y2674.
15. Yoshimatsu K, Golijanin D, Paty PB, et al. Inducible microsomalprostaglandin E synthase is overexpressed in colorectal adenomas andcancer. Clin Cancer Res. 2001;7:3971Y3976.
16. Murakami M, Nakashima K, Kamei D, et al. Cellular prostaglandin E2production by membrane-bound prostaglandin E synthaseY2 via bothcyclooxygenases-1 and -2. J Biol Chem. 2003;278:37937Y37947.
17. Chell S, Kadi A, Williams AC, et al. Mediators of PGE2 synthesis andsignalling downstream of COX-2 represent potential targets for the
Hasan et al Pancreas & Volume 37, Number 2, August 2008
126 * 2008 Lippincott Williams & Wilkins
Copyright @ 2008 Lippincott Williams & Wilkins. Unauthorized reproduction of this article is prohibited.

prevention/treatment of colorectal cancer. Biochim Biophys Acta. 2006;1766:104Y119.
18. Grosser T, Fries S, FitzGerald GA. Biological basis for the cardiovascularconsequences of COX-2 inhibition: therapeutic challenges andopportunities. J Clin Invest. 2006;116:4Y15.
19. Hermann M, Ruschitzka F. Cardiovascular risk of cyclooxygenase-2inhibitors and traditional non-steroidal anti-inflammatory drugs. AnnMed. 2007;39:18Y27.
20. Sipos B, Moser S, Kalthoff H, et al. A comprehensive characterizationof pancreatic ductal carcinoma cell lines: towards the establishmentof an in vitro research platform. Virchows Arch. 2003;442:444Y452.
21. Muller-Decker K, Furstenberger G, Annan N, et al. Preinvasiveduct-derived neoplasms in pancreas of keratin 5-promotercyclooxygenase-2 transgenic mice. Gastroenterology. 2006;130:2165Y2178.
22. Eibl G, Reber HA, Wente MN, et al. The selective cyclooxygenase-2inhibitor nimesulide induces apoptosis in pancreatic cancer cellsindependent of COX-2. Pancreas. 2003;26:33Y41.
23. Eibl G, Reber HA, Hines OJ, et al. COX and PPAR: possible interactionsin pancreatic cancer. Pancreas. 2004;29:247Y253.
24. Raut CP, Nawrocki S, Lashinger LM, et al. Celecoxib inhibitsangiogenesis by inducing endothelial cell apoptosis in human pancreatictumor xenografts. Cancer Biol Ther. 2004;3:1217Y1224.
25. Tseng WW, Deganutti A, Chen MN, et al. Selective cyclooxygenase-2inhibitor rofecoxib (Vioxx) induces expression of cell cycle arrest genes
and slows tumor growth in human pancreatic cancer. J Gastrointest Surg.2002;6:838Y843.
26. Ferrari V, Valcamonico F, Amoroso V, et al. Gemcitabine plus celecoxib(GECO) in advanced pancreatic cancer: a phase II trial. CancerChemother Pharmacol. 2006;57:185Y190.
27. Milella M, Gelibter A, Di CS, et al. Pilot study of celecoxib and infusional5-fluorouracil as second-line treatment for advanced pancreaticcarcinoma. Cancer. 2004;101:133Y138.
28. Ueno N, Takegoshi Y, Kamei D, et al. Coupling between cyclooxygenasesand terminal prostanoid synthases. Biochem Biophys Res Commun.2005;338:70Y76.
29. Murakami M, Naraba H, Tanioka T, et al. Regulation of prostaglandinE2 biosynthesis by inducible membrane-associated prostaglandin E2synthase that acts in concert with cyclooxygenase-2. J Biol Chem.2000;275:32783Y32792.
30. Oshima H, Oshima M, Inaba K, et al. Hyperplastic gastric tumorsinduced by activated macrophages in COX-2/mPGES-1 transgenic mice.EMBO J. 2004;23:1669Y1678.
31. Murakami M, Kudo I. Prostaglandin E synthase: a novel drugtarget for inflammation and cancer. Curr Pharm Des. 2006;12:943Y954.
32. Molina MA, Sitja-Arnau M, Lemoine MG, et al. Increasedcyclooxygenase-2 expression in human pancreatic carcinomas and celllines: growth inhibition by nonsteroidal anti-inflammatory drugs. CancerRes. 1999;59:4356Y4362.
Pancreas & Volume 37, Number 2, August 2008 PGE2 Pathway Analysis in Pancreatic Cancers
* 2008 Lippincott Williams & Wilkins 127
Copyright @ 2008 Lippincott Williams & Wilkins. Unauthorized reproduction of this article is prohibited.




















