
Exposure to appetitive food stimuli markedly activates the human brain
8
Exposure to appetitive food stimuli markedly activates the human brain Gene-Jack Wang, a, * Nora D. Volkow, a,1 Frank Telang, a Millard Jayne, a Jim Ma, a Manlong Rao, b Wei Zhu, b Christopher T. Wong, a Naomi R. Pappas, a Allan Geliebter, c and Joanna S. Fowler d a Medical Department, Brookhaven National Laboratory, Upton, NY 11973, USA b Department of Applied Mathematics and Statistics, S.U.N.Y., Stony Brook, NY 11794, USA c St. Luke’s/Roosevelt Hospital, New York, NY 10025, USA d Chemistry Department, Brookhaven National Laboratory, Upton, NY 11973, USA Received 22 September 2003; revised 18 November 2003; accepted 20 November 2003 Objective: The increased incidence of obesity most likely reflects changes in the environment that had made food more available and palatable. Here we assess the response of the human brain to the presentation of appetitive food stimuli during food presentation using PET and FDG. Method: Metabolic changes in response to food presentation were done in 12 healthy normal body weight subjects who were food deprived before the study. Results: Food presentation significantly increased metabolism in the whole brain (24%, P < 0.01) and these changes were largest in superior temporal, anterior insula, and orbitofrontal cortices. The increases in the right orbitofrontal cortex were the ones that correlated significantly with the increases in self-reports of hunger and desire for food. Discussion: The marked increase in brain metabolism by the presentation of food provides evidence of the high sensitivity of the human brain to food stimuli. This high sensitivity coupled with the ubiquitousness of food stimuli in the environment is likely to contribute to the epidemic of obesity. In particular, the activation of the right orbitofrontal cortex, a brain region involved with drive, may underlie the motivation to procure food, which may be subjectively experienced as ‘‘desire for food’’ and ‘‘hunger’’ when exposed to food stimuli. D 2004 Elsevier Inc. All rights reserved. Keywords: PET; FDG; Food; Orbitofrontal cortex; Insula; Motivation Introduction The incidence of obesity in the United States and in the world has reached epidemic proportions and continues to rise (i.e., approximately 30% of adults in the United States are obese) (Flegal et al., 2002). The increased morbidity and mortality associated with obesity places a sense of urgency to understand the processes that have contributed to this epidemic. Of particular relevance is the environment, which has made food not only widely available but also increasingly more varied and palatable. Although many signals that regulate food intake originate from internal sources that control caloric and nutrient intake (i.e., leptin, insulin, ghrelin, PYY), variables other than caloric intake and satiety have profound effects on food intake. These include the pleasurable sensory responses from food (i.e., palatability), emo- tional variables (i.e., stress, depression), and environmental factors (i.e., food availability, food related cues, alternative reinforcers) (Patel and Schlundt, 2001). Disruption in the sensitivity of the brain to these other variables could result in excessive eating and obesity. A particularly relevant variable is the sensory appeal and the conditioned responses that the food conveys to the subject. In a prior study with PET and [ 11 C]raclopride, we showed that striatal dopamine release was associated with the desire for food during presentation of palatable food stimuli (Volkow et al., 2002a). This response was consistent with dopamine’s (DA) role in motivation for food (Martel and Fantino, 1996). However, DA’s role is to regulate the activity of brain regions, which in turn are the ones that are likely to modulate the motivation for food consumption (nucleus accumbens, dorsal striatum, orbitofrontal cortex, and insula) (Bassareo et al., 2002; Chikama et al., 1997). Here we assessed the brain circuits activated by the presentation of food stimuli using the experimental paradigm that we had used to show DA increases in striatum with exposure to palatable food stimuli (Volkow et al., 2002a). Methods Twelve healthy right-handed subjects (seven female and five male subjects, age 28 F 5 years old) were recruited for the study. Subjects with body mass index greater than 30 kg/m 2 , history of eating disorders, surgical/medical treatment for weight control, dependence on alcohol or other drugs of abuse (except for caffeine <5 cups/day or nicotine <1 pack/day), neurological or psychiatric disorder, use of prescription (nonpsychiatric) medication(s) that can affect brain function in the past 2 weeks, medical conditions 1053-8119/$ - see front matter D 2004 Elsevier Inc. All rights reserved. doi:10.1016/j.neuroimage.2003.11.026 * Corresponding author. Fax: +1-631-344-5260. E-mail address: [email protected] (G.-J. Wang). 1 N.D. Volkow’s current affiliations are NIDA and NIAA, 6001 Executive Dr., Bethesda MD 20892-9581. Available online on ScienceDirect (www.sciencedirect.com.) www.elsevier.com/locate/ynimg NeuroImage 21 (2004) 1790 – 1797
Transcript of Exposure to appetitive food stimuli markedly activates the human brain

www.elsevier.com/locate/ynimg
NeuroImage 21 (2004) 1790–1797
Exposure to appetitive food stimuli markedly activates
the human brain
Gene-Jack Wang,a,* Nora D. Volkow,a,1 Frank Telang,a Millard Jayne,a Jim Ma,a Manlong Rao,b
Wei Zhu,b Christopher T. Wong,a Naomi R. Pappas,a Allan Geliebter,c and Joanna S. Fowlerd
aMedical Department, Brookhaven National Laboratory, Upton, NY 11973, USAbDepartment of Applied Mathematics and Statistics, S.U.N.Y., Stony Brook, NY 11794, USAcSt. Luke’s/Roosevelt Hospital, New York, NY 10025, USAdChemistry Department, Brookhaven National Laboratory, Upton, NY 11973, USA
Received 22 September 2003; revised 18 November 2003; accepted 20 November 2003
Objective: The increased incidence of obesity most likely reflects
changes in the environment that had made food more available and
palatable. Here we assess the response of the human brain to the
presentation of appetitive food stimuli during food presentation using
PET and FDG. Method: Metabolic changes in response to food
presentation were done in 12 healthy normal body weight subjects
who were food deprived before the study. Results: Food presentation
significantly increased metabolism in the whole brain (24%, P < 0.01)
and these changes were largest in superior temporal, anterior insula,
and orbitofrontal cortices. The increases in the right orbitofrontal
cortex were the ones that correlated significantly with the increases in
self-reports of hunger and desire for food. Discussion: The marked
increase in brain metabolism by the presentation of food provides
evidence of the high sensitivity of the human brain to food stimuli. This
high sensitivity coupled with the ubiquitousness of food stimuli in the
environment is likely to contribute to the epidemic of obesity. In
particular, the activation of the right orbitofrontal cortex, a brain
region involved with drive, may underlie the motivation to procure
food, which may be subjectively experienced as ‘‘desire for food’’ and
‘‘hunger’’ when exposed to food stimuli.
D 2004 Elsevier Inc. All rights reserved.
Keywords: PET; FDG; Food; Orbitofrontal cortex; Insula; Motivation
Introduction
The incidence of obesity in the United States and in the world
has reached epidemic proportions and continues to rise (i.e.,
approximately 30% of adults in the United States are obese)
(Flegal et al., 2002). The increased morbidity and mortality
associated with obesity places a sense of urgency to understand
the processes that have contributed to this epidemic. Of particular
1053-8119/$ - see front matter D 2004 Elsevier Inc. All rights reserved.
doi:10.1016/j.neuroimage.2003.11.026
* Corresponding author. Fax: +1-631-344-5260.
E-mail address: [email protected] (G.-J. Wang).1 N.D. Volkow’s current affiliations are NIDA and NIAA, 6001
Executive Dr., Bethesda MD 20892-9581.
Available online on ScienceDirect (www.sciencedirect.com.)
relevance is the environment, which has made food not only
widely available but also increasingly more varied and palatable.
Although many signals that regulate food intake originate from
internal sources that control caloric and nutrient intake (i.e., leptin,
insulin, ghrelin, PYY), variables other than caloric intake and
satiety have profound effects on food intake. These include the
pleasurable sensory responses from food (i.e., palatability), emo-
tional variables (i.e., stress, depression), and environmental factors
(i.e., food availability, food related cues, alternative reinforcers)
(Patel and Schlundt, 2001). Disruption in the sensitivity of the
brain to these other variables could result in excessive eating and
obesity. A particularly relevant variable is the sensory appeal and
the conditioned responses that the food conveys to the subject. In a
prior study with PET and [11C]raclopride, we showed that striatal
dopamine release was associated with the desire for food during
presentation of palatable food stimuli (Volkow et al., 2002a). This
response was consistent with dopamine’s (DA) role in motivation
for food (Martel and Fantino, 1996). However, DA’s role is to
regulate the activity of brain regions, which in turn are the ones that
are likely to modulate the motivation for food consumption
(nucleus accumbens, dorsal striatum, orbitofrontal cortex, and
insula) (Bassareo et al., 2002; Chikama et al., 1997). Here we
assessed the brain circuits activated by the presentation of food
stimuli using the experimental paradigm that we had used to show
DA increases in striatum with exposure to palatable food stimuli
(Volkow et al., 2002a).
Methods
Twelve healthy right-handed subjects (seven female and five
male subjects, age 28 F 5 years old) were recruited for the study.
Subjects with body mass index greater than 30 kg/m2, history of
eating disorders, surgical/medical treatment for weight control,
dependence on alcohol or other drugs of abuse (except for caffeine
<5 cups/day or nicotine <1 pack/day), neurological or psychiatric
disorder, use of prescription (nonpsychiatric) medication(s) that
can affect brain function in the past 2 weeks, medical conditions

G.-J. Wang et al. / NeuroImage 21 (2004) 1790–1797 1791
that may alter cerebral function, cardiovascular disease and
diabetes, and head trauma with loss of consciousness of more
than 30 min were excluded from the study. Urine screening tests
for psychoactive drugs (including PCP, cocaine, amphetamine,
opiates, barbiturates, benzodiazepine, and THC) were performed
to corroborate lack of drug use. Written informed consents were
obtained after the experimental procedure was explained and after
the subjects had read the consent form. The protocol was
approved by the Institutional Review Board at Brookhaven
National Laboratory.
Subjects were scanned with 2-deoxy-2[18F]fluoro-D-glucose
(FDG) using a Siemens HR + positron emission tomography
(PET) scanner. Details on procedures for positioning of the sub-
jects, arterialized venous and venous catheterization, quantification
of radiotracer, and transmission and emission scans have been
published (Wang et al., 2003). Briefly, one emission scan (20 min)
was taken 35 min after an intravenous injection of 4–6 mCi of
FDG. During the study, subjects were positioned supine in the PET
camera with their eyes open; the room was dimly lit and noise was
kept to a minimum. Subjects were scanned twice with FDG in two
different days under the following conditions. (1) Day A: Food
presentation started 15 min before FDG injection and continued for
a total of 45 min. (2) Day B: Neutral intervention started 15 min
before FDG injection and continued for a total of 45 min. The
sequence was randomized so that for half of the subject the first
day was day A, while for the other half it was day B.
The subjects were asked to fill out a questionnaire, which
contained the following information on the day of screening: a
rating of the subject’s overall interest in food; what the subject’s
favorite foods were; what food smells stimulated the subject’s
appetite; what food smells diminished the subject’s appetite; and to
rate a list of foods for their preferences on a scale from 1 to 10, 10
being the highest. The food items with the highest ratings were
Fig. 1. Transaxial FDG-PET brain images of a subject during food presentation and
cortex, insula, and orbitofrontal cortex. The metabolic images for both scans are sc
neutral intervention and presented using the rainbow scale, where red represents
then selected to be presented to the subject during the food
stimulation condition.
For the food stimulation condition, the subjects were asked to
describe their favorite foods and how they like to eat them while
they were presented with foods that they had reported as among
their favorite ones. The food was warmed to enhance the smell and
the subjects were presented with it so that they could view it and
smell it. A cotton swab impregnated with the food was placed in
their tongues so they could taste it. A given food item was
presented for 5 min and then it was exchanged for a new one.
For the neutral intervention, subjects were asked to describe in as
much detail as possible their family genealogy while they were
presented with nonfood related items (paper photographs of nature
scenes, of humans, and of animals), which they were allowed to
smell, and a cotton swab impregnated with water was placed in
their tongues and lips as was done for the food condition. The food
and the neutral interventions were started 15 min before radiotracer
injection and were continued for a total of 45 min. Subjects were
asked to have their last meal at 7 PM the evening before the day of
the study and were studied between 17 and 19 h after the last meal.
During the PET studies, participants were instructed to orally
respond to each descriptor using a whole number between 1 and 10
for the self-report of ‘‘hunger’’ and ‘‘desire of food’’, which were
obtained before the food presentation and then at 5-min intervals
for a total of 45 min.
Regions of interest (ROI) in orbitofrontal cortex, anterior
insula, parietal cortex, temporal cortex, caudate, putamen, and
thalamus were obtained using a template, which we had previously
published (Wang et al., 2003). We also computed a value for global
metabolism by averaging the activity in the 63 planes scanned. To
minimize the effects of overall changes in brain metabolism on the
regional measures, we normalized them using the ratio of the
region to the global metabolic measures (‘‘relative’’ measures).
during neutral intervention at levels of postcentral gyrus, superior temporal
aled with respect to the maximum absolute metabolic value obtained on the
the highest value and dark violet represents the lowest value.

G.-J. Wang et al. / NeuroImage 21 (2004) 1790–17971792
Differences in metabolism between the measures obtained in
the food presentation and neutral intervention conditions were
tested with paired samples t tests. Pearson product moment
correlations were used to assess the relationship between the
changes in metabolism and the change in the behavioral measures
between the food and neutral interventions.
Differences in metabolism between the two conditions were
also tested on the voxel level using the software package for
Statistical Parametric Mapping (SPM99 Software, 1999). Before
the analysis, each subject’s PET image was mapped onto the
Montreal Neurological Institute template, which closely resembles
the Talairach brain and smoothed via a Gaussian kernel with full
width half maximum (FWHM) at 16 mm. The relative (normal-
ized) metabolic image was obtained by dividing the signal level of
each voxel with the global mean, which is the average signal level
of all voxels in the PET image. The analysis, which was
essentially a paired samples t test performed on each voxel, was
Fig. 2. Absolute brain metabolic changes between food presentation and neutral i
right somatosensory cortex; LIN: left insula; RIN: right insula; LST: left superior
cortex; ROF: right orbitofrontal cortex; LOC: left occipital cortex; ROC: right oc
thalamus; RTH: right thalamus. A, P < 0.005; B, P < 0.01; C, P < 0.05.
performed using both the absolute and the relative metabolic
images. Multiple-test correction was performed via the random
field theory less conservative than the traditional approach such as
the Bonferroni method (Worsley et al, 1996). Pixels that were
significantly different from those in the neutral presentation (P <
0.05) were identified with respect to the Talairach and Tournoux
stereotactic coordinates (Talairach and Tournoux, 1988) and dis-
played on the axial MR images. The threshold for the cluster size
was set at 100 voxels. Only regions with corrected P values <
0.05 at the cluster level were considered significantly activated or
deactivated.
Results
The body weight of the subjects was between 110 and 210 lb
(average: 147.5 F 32.4 lb). The averaged body mass index of the
ntervention conditions. GLO: global; LSS: left somatosensory cortex; RSS:
temporal cortex; RST: left superior temporal cortex; LOF: left orbitofrontal
cipital cortex; LBG: left basal ganglia; RBG: right basal ganglia; LTH: left

G.-J. Wang et al. / NeuroImage 21 (2004) 1790–1797 1793
subjects was 24 F 2.6 (range: 20–29). The favorite food items
most frequently selected by the subjects were bacon–egg–cheese
sandwich, cinnamon bun, pizza, hamburger with cheese, fried
chicken, lasagna, Bar-Be-Que rib, ice cream, brownie, and choc-
olate cake.
Compared to the neutral intervention condition (36 F 2 Amol/
100g/min), the food presentation (45F 9 Amol 100 g�1 min�1, P <
0.01) significantly increased whole brain metabolism (+24F 21%).
The absolute regional metabolic measures were also significantly
higher for all brain regions (except in the occipital cortex) for the
food than the neutral condition (Figs. 1 and 2). Normalization by
whole brain metabolism (relative metabolic measures) revealed that
(Fig. 3) the largest increases were in the left and right postcentral
gyrus (left: +14.9 F 11.6%, P < 0.0001; right: +8.5 F 7.9%, P <
0.012), left superior temporal (+11.2 F 9.2%, P < 0.002), left
anterior insula (+9.6 F 14.6%, P < 0.03), and left orbitofrontal
cortex (+8.9 F 12.3%, P < 0.015).
The SPM analyses of the absolute metabolic values at a
significance level of P < 0.0001 (voxel level) showed a large
Fig. 3. Relative brain metabolic changes between food presentation a
contiguous cluster (72,865 voxels) in bilateral cortical (inferior
frontal, parietal, temporal, occipital, and cerebellar regions) and
subcortical structures with food presentation. The results from
SPM with the relative metabolic measures (Fig. 4) corroborated
the findings obtained using the ROI method. The SPM analyses
yielded a large contiguous cluster (8339 voxels) in the left
parietotemporal cortices and two smaller clusters: one in the right
parietotemporal cortex (1679 voxels) and one in the left orbito-
frontal cortex (499 voxels) with food presentation. The large
cluster in the left parietotemporal cortices encompassed three
subclusters that included the posterior central gyrus (Brodmann’s
areas 1): (�42,�26, 30); the left insula: (�44,�6, 2); and the left
superior temporal gyrus: (�60,�30,�2). The significance of this
cluster was P < 0.004 (corrected at the cluster level). The
significance for the cluster in the right parietotemporal cortex
(62, �16, 20) was P < 0.05 (uncorrected at the cluster level).
The cluster in the left orbitofrontal cortex was not significant at the
cluster level; however, there were two voxel-level significant
regions (�38, 44, 4, P < 0.01; �38, 56, 2, P < 0.02).
nd neutral intervention conditions. A, P < 0.005; B, P < 0.05.
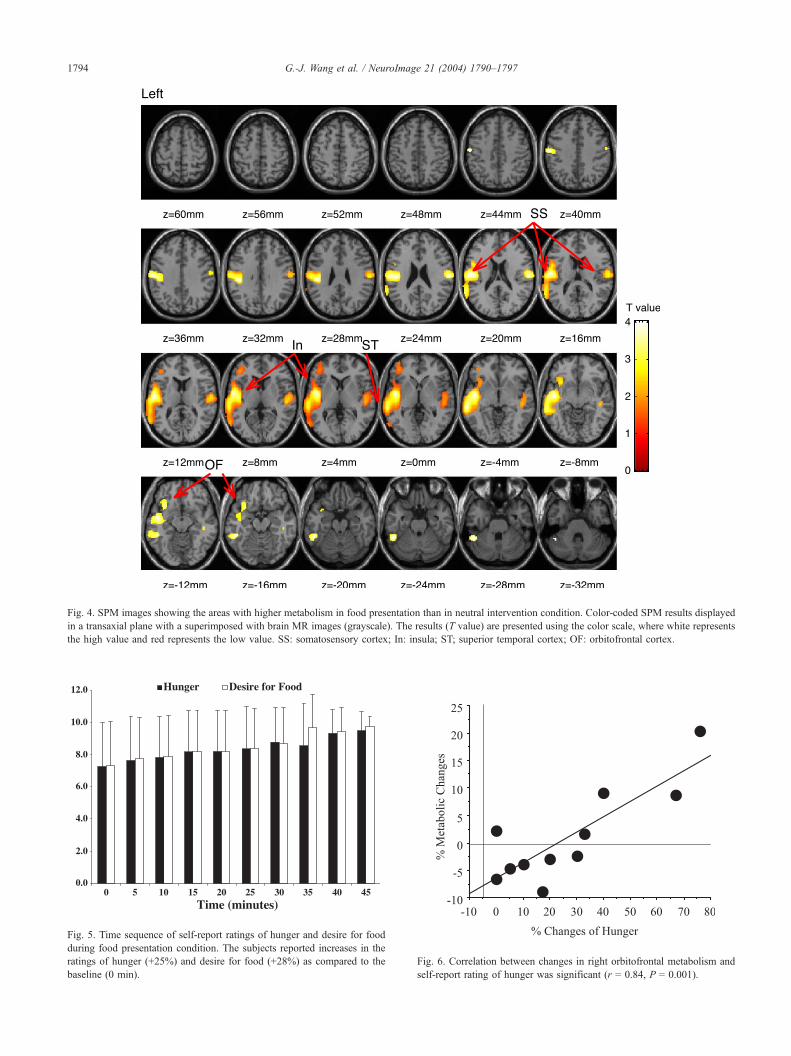
Fig. 4. SPM images showing the areas with higher metabolism in food presentation than in neutral intervention condition. Color-coded SPM results displayed
in a transaxial plane with a superimposed with brain MR images (grayscale). The results (T value) are presented using the color scale, where white represents
the high value and red represents the low value. SS: somatosensory cortex; In: insula; ST; superior temporal cortex; OF: orbitofrontal cortex.
Fig. 5. Time sequence of self-report ratings of hunger and desire for food
during food presentation condition. The subjects reported increases in the
ratings of hunger (+25%) and desire for food (+28%) as compared to the
baseline (0 min).
Fig. 6. Correlation between changes in right orbitofrontal metabolism and
self-report rating of hunger was significant (r = 0.84, P = 0.001).
G.-J. Wang et al. / NeuroImage 21 (2004) 1790–17971794

G.-J. Wang et al. / NeuroImage 21 (2004) 1790–1797 1795
The food presentation condition resulted in significant increases
in the ratings of hunger (25 F 26%, P = 0.006) and desire for food
(28 F 27%, P = 0.007, Fig. 5). The correlation analyses showed a
significant association between relative metabolic changes in the
right orbitofrontal cortex and the self-reports of hunger (r = 0.84,
P = 0.001, Fig. 6) and desire for food (r = 0.77, P = 0.006). No
significant correlations were found between the behavioral meas-
ures and the metabolic changes in the insula, postcentral gyrus, and
superior temporal regions.
Discussion
This study shows that presentation of food stimuli to fasting
subjects resulted in marked increases in whole brain metabolism
(24%). The increases in metabolism were largest in left cortical
regions (postcentral gyrus, superior temporal cortex, insula, and
orbitofrontal cortex). There is no conclusive evidence regarding a
lateralized representation for processing of chemosensory stimuli
in the brain. Indeed, some studies have reported right hemisphere
dominance (Small et al., 1999), while others reported bilateral
(Faurion et al., 1999) or left hemispheric (Del Parigi et al., 2002)
activation in response to chemosensory stimulation. In our study,
the predominant activation in the left hemisphere could reflect the
experimental conditions used for the food and the neutral
stimulations.
Food stimulation resulted in activation of the postcentral region
on the lateral surface of the parietal cortex, which is where the
somatosensory map of the tongue is located and is an area involved
with taste perception (Heimer, 1995). Thus, its activation is consis-
tent with its known role in the somatosensory perception of food.
The reward associated with the taste of food (influenced by palat-
ability) is distinctively different from that associated with food
ingestion (influenced by hunger/satiety). Palatability increases food
intake through a positive-feedback reward mechanism that involves
opioid and GABA/benzodiazepine systems (Cooper, 1989; Yeo-
mans and Gray, 2002). We had previously observed during non-
stimulation conditions (resting state) enhanced brain activity in this
somatosensory region in morbidly obese subjects when compared to
normal-weight subjects (Wang et al., 2002). This led us to postulate
that enhanced activity in this region could make an individual more
sensitive to the rewarding properties of food related to palatability
and could be one of the variables contributing to excess food
consumption. Similarly, in obese women, cerebral blood flow in
parietal cortex was found to be significantly higher than in normal-
weight subjects (Karhunen et al., 1997).
The anterior insula showed activation during the food presen-
tation, which is consistent with its role as a primary gustatory
cortex (O’Doherty et al., 2001; Yaxley et al., 1990; Zald et al.,
1998). The activation of the insula has been observed with fasting
(Morris and Dolan, 2001; Tataranni et al., 1999) and during
activation with unpleasant and aversive food-eating conditions
(O’Doherty et al., 2001; Zald et al., 1998). The anterior insular
region has also been implicated in the processing of visceral
sensations (Allen et al., 1991; Small et al., 1999). Indeed, insular
activation has been reported during temperature and pain percep-
tion (Casey et al., 1996).
The OFC has been identified as a secondary gustatory cortex
(Rolls et al., 1990). It is believed that the OFC is involved with the
rapid learning of visual, olfactory, and taste associations (Rolls,
1997). The OFC receives projections from the insula, striatum, and
amygdala. These connections place the OFC in a position to
process information about the motivational value of gustatory
stimuli. Neurons in the OFC are modulated by the motivational
state of the animal, which respond to the sight or taste of food
when the animal is hungry but do not respond when the animal is
satiated (Rolls, 1989). The neurons in the OFC respond selectively
to rewards or aversive stimuli and process the relative preference
for food rewards (Tremblay and Schultz, 1999). The OFC is also
involved in processing tastes that have both positive and negative
affective valence (O’Doherty et al., 2001). Primate studies have
shown not only taste but also smell-responsive cells throughout the
posterior OFC (Rolls et al., 1999). The present study showed
increased metabolic activity in the left posterior OFC in response to
the food presentation, which included tasting and smelling of food.
Thus, its activation, which was not associated with the subjective
perception of hunger or desire of food, is likely to reflect its
involvement in processing the valence of the flavors and of the
smells from the stimuli. In contrast, the right OFC was significantly
associated with the perception of hunger and desire for food. A
similar correlation had previously been reported by a PET study
that showed a positive correlation between increases in cerebral
blood flow in right posterior OFC and hunger ratings during
presentation of food items (Morris and Dolan, 2001). This associ-
ation is consistent with the known role of the OFC in processing
expectation of food reward (Watababe, 1996). Wanting food (or
appetite) is modulated by DA pathways in part by modulating
activity in the OFC (Berridge, 1996; Spanagel and Weiss, 1999).
Indeed, imaging studies have shown that metabolic activity in the
OFC is in part regulated by DA activity (Volkow et al., 1993,
2002b).
Using the same food-stimulation paradigm, we previously
showed DA increases in the striatum during food stimulation that
were also associated with the ‘‘desire for food’’ and with the
perception of ‘‘hunger’’ (Volkow et al., 2002b). Thus, the current
studies suggest that the enhanced OFC activation by the food
stimulation may reflect downstream effects from DA stimulation
and that DA’s involvement in the drive for food consumption in
human subjects is in part mediated by its effects in the OFC. The
results could explain the deleterious effects of constant exposure to
food stimuli (e.g., advertisements, candy machines, food channels,
stores) in overeating (Coon and Tucker, 2002). It also raises the
possibility that being a food management professional (e.g., chef)
may pose a risk for overeating particularly in those at higher
genetic risk for obesity similar to the higher risk for drug abuse and
addiction in anesthesiologists who as part of their work are
constantly exposed to psychoactive drugs with abuse potential
(Tirrell, 1994). In nonhuman primates, lesions of the OFC produce
abnormal eating behavior (Baylis and Gaffan, 1991). The OFC is a
brain region that has been implicated in the compulsive behaviors
characteristic of drug addictive states (Volkow and Fowler 2000).
Indeed, activation of the right OFC cortex was shown to be
associated with cocaine craving induced by stimulant administra-
tion in cocaine-addicted subjects (Volkow et al. 1999). Markedly
increased metabolic activity in the OFC was also reported during
cue-induced cocaine craving in drug-addicted subjects (Bonson et
al., 2002; Grant et al., 1996; Wang et al., 1999). This suggests that
the same brain region, which is linked with the desire for natural
stimuli (food), is also linked with drug craving in drug-addicted
subjects. Thus, disrupted OFC activity could also possibly underlie
the compulsiveness and lack of control in the eating behaviors of
morbidly obese subjects.

G.-J. Wang et al. / NeuroImage 21 (2004) 1790–17971796
Although no studies have demonstrated abnormalities in the
OFC in obese subjects, we have reported decreases in DA D2
receptors in obese individuals (Wang et al., 2001). Since our
previous studies in drug-addicted subjects showed an association
between decreases in DA D2 receptors and metabolism in OFC
(Volkow et al., 1993, 2001), further studies are warranted to
assess if indeed there are abnormalities in the OFC in obese
subjects.
Conclusion
The marked activation of brain metabolism by the presentation
of food provides evidence of the high sensitivity of the human
brain to the presence of food stimuli. This high sensitivity coupled
with the ubiquitousness of food stimuli in the environment is likely
to contribute to the epidemic of obesity. In particular, the activation
in the right orbitofrontal cortex, a brain region involved with drive,
may underlie the motivation to procure food, which may be
subjectively experienced as ‘‘desire for food’’ and ‘‘hunger’’.
Acknowledgments
This research was carried out at Brookhaven National
Laboratory (BNL) under support by the U.S. Department of
Energy OBER (DE-ACO2-76CH00016) and by the National
Institute on Drug Abuse (DA 7092-01 and DA00280). We thank
David Schlyer and Michael Schueller for Cyclotron operations;
Donald Warner and David Alexoff for PET operations; Richard
Ferrieri, Colleen Shea, Youwen Xu, Victor Garza, and Payton King
for radiotracer preparation and analysis; Karen Apelskog for study
protocol preparation; and Noelwah Netusil, Pauline Carter, and
Naomi Pappas for patient care and recruitment.
References
Allen, G.V., Saper, C.B., Hurley, K.M., Cechetto, D.F., 1991. Organiza-
tion of visceral and limbic connections in the insular cortex of the rat.
J. Comp. Neurol. 311, 1–16.
Bassareo, V., De Luca, M.A., Di Chiara, G., 2002. Differential expression
of motivational stimulus properties by dopamine in nucleus accumbens
shell versus core and prefrontal cortex. J. Neurosci. 22, 4709–4719.
Baylis, L.L., Gaffan, D., 1991. Amygdalectomy and ventromedial prefrontal
ablation produce similar deficits in food choice and in simple object
discrimination learning for an unseen reward. Exp. Brain Res. 86,
617–622.
Berridge, K.C., 1996. Food reward: brain substrates of wanting and liking.
Neurosci. Biobehav. Rev. 20, 1–25.
Bonson, K.R., Grant, S.J., Contoreggi, C.S., Links, J.M., Metcalfe, J.,
Weyl, H.L., Kurian, V., Ernst, M., London, E.D., 2002. Neural systems
and cue-induced cocaine craving. Neuropsychopharmacology 26,
376–386.
Casey, K.L., Minoshima, S., Morrow, T.J., Koeppe, R.A., 1996. Compar-
ison of human cerebral activation pattern during cutaneous warmth, heat
pain, and deep cold pain. J. Neurophysiol. 76, 571–581.
Chikama, M., McFarland, N.R., Amaral, D.G., Haber, S.N., 1997. Insular
cortical projections to functional regions of the striatum correlate with
cortical cytoarchitectonic organization in the primate. J. Neurosci. 17,
9686–9705.
Coon, K.A., Tucker, K.L., 2002. Television and children’s consumption
patterns. A review of the literature. Minerva Pediatr. 54, 423–436.
Cooper, S.J., 1989. Benzodiazepine receptor-mediated enhancement and
inhibition of taste reactivity, food choice, and intake. Ann. N. Y. Acad.
Sci. 575, 321–336.
Del Parigi, A., Chen, K., Salbe, A.D., Gautier, J.F., Ravussin, E., Reiman,
E.M., Tataranni, P.A., 2002. Tasting a liquid meal after a prolonged fast
is associated with preferential activation of the left hemisphere. Neuro-
Report 13, 1141–1145.
Faurion, A., Cerf, B., Van De Moortele, P.F., Lobel, E., Mac Leod, P., Le
Bihan, D., 1999. Human taste cortical areas studied with functional
magnetic resonance imaging: evidence of functional lateralization relat-
ed to handedness. Neurosci. Lett. 277, 189–192.
Flegal, K.M., Carroll, M.D., Ogden, C.L., Johnson, C.L., 2002. Prevalence
and trends in obesity among US adults, 1999–2000. JAMA 288,
1723–1727.
Grant, S., London, E.D., Newlin, D.B., Villemagne, V.L., Liu, X., Conto-
reggi, C., Phillips, R.L., Kimes, A.S., Margolin, A., 1996. Activation of
memory circuits during cue-elicited cocaine craving. Proc. Natl. Acad.
Sci. U. S. A. 93, 12040–12045.
Heimer, L., 1995. The Human Brain and Spinal Cord. Second ed. Springer-
Verlag, New York, pp. 433–454.
Karhunen, L.J., Lappalainen, R.I., Vanninen, E.J., Kuikka, J.T., Uusitupa,
M.I., 1997. Regional cerebral blood flow during food exposure in obese
and normal-weight women. Brain 120 (Pt 9), 1675–1684.
Martel, P., Fantino, M., 1996. Mesolimbic dopaminergic system activity as
a function of food reward: a microdialysis study. Pharmacol., Biochem.
Behav. 53, 221–226.
Morris, J.S., Dolan, R.J., 2001. Involvement of human amygdala and orbi-
tofrontal cortex in hunger-enhanced memory for food stimuli. J. Neuro-
sci. 21, 5304–5310.
O’Doherty, J., Rolls, E.T., Francis, S., Bowtell, R., McGlone, F., 2001.
Representation of pleasant and aversive taste in the human brain.
J. Neurophysiol. 85, 1315–1321.
Patel, K.A., Schlundt, D.G., 2001. Impact of moods and social context on
eating behavior. Appetite 36 (2), 111–118.
Rolls, E.T., 1989. Information processing in the taste system of primates.
J. Exp. Biol. 146, 141–164.
Rolls, E.T., 1997. Taste and olfactory processing in the brain and its rela-
tion to the control of eating. Crit. Rev. Neurobiol. 11, 263–287.
Rolls, E.T., Yaxley, S., Sienkiewicz, Z.J., 1990. Gustatory responses of
single neurons in the caudolateral orbitofrontal cortex of the macaque
monkey. J. Neurophysiol. 64, 1055–1066.
Rolls, E.T., Critchley, H.D., Browning, A.S., Hernadi, I., Lenard, L., 1999.
Responses to the sensory properties of fat of neurons in the primate
orbitofrontal cortex. J. Neurosci. 19, 1532–1540.
Small, D.M., Zald, D.H., Jones-Gotman, M., Zatorre, R.J., Pardo, J.V.,
Frey, S., Petrides, M., 1999. Human cortical gustatory areas: a review
of functional neuroimaging data. NeuroReport 10, 7–14.
Spanagel, R., Weiss, F., 1999. The dopamine hypothesis of reward: past
and current status. Trends Neurosci. 22, 521–527.
SPM99 Software, 1999. MRC Cyclotron Unit, Hammersmith Hospital,
London, UK.
Talairach, J., Tournoux, P.A., 1988. Co-planar Stereotaxic Atlas of a Hu-
man Brain. Thieme, New York.
Tataranni, P.A., Gautier, J.F., Chen, K., Uecker, A., Bandy, D., Salbe, A.D.,
Pratley, R.E., Lawson, M., Reiman, E.M., Ravussin, E., 1999. Neuro-
anatomical correlates of hunger and satiation in humans using positron
emission tomography. Proc. Natl. Acad. Sci. U. S. A. 96, 4569–4574.
Tirrell, C.D., 1994. Psychoactive substance disorders among health care
professionals. Plast. Surg. Nurs. 14, 169–172.
Tremblay, L., Schultz, W., 1999. Relative reward preference in primate
orbitofrontal cortex. Nature 398, 704–708.
Volkow, N.D., Fowler, J.S., 2000. Addiction, a disease of compulsion
and drive: involvement of the orbitofrontal cortex. Cereb. Cortex 10,
318–325.
Volkow, N.D., Fowler, J.S., Wang, G.-J., Hitzemann, R.J., Logan, J.,
Schlyer, D.J., Dewey, S.L., Wolf, A.P., 1993. Decrease dopamine D2
receptor availability is associated with reduced frontal metabolism in
cocaine abuser. Synapse 14, 169–177.

G.-J. Wang et al. / NeuroImage 21 (2004) 1790–1797 1797
Volkow, N.D., Wang, G.-J., Fowler, J.S., Hitzemann, R.J., Angrist, B.,
Gatley, S.J., Logan, J., Ding, Y.-S., Pappas, N., 1999. Methylphenidate
induced craving in cocaine abusers is associated with changes in right
striato-orbitofrontal metabolism: implications in addiction. Am. J. Psy-
chiatry 156, 19–26.
Volkow, N.D., Chang, L., Wang, G.-J., Fowler, J.S., Ding, Y.-S., Sedler,
M., Logan, J., Franceschi, D., Gatley, J., Hitzemann, R., Gifford, A.,
Wong, C., Pappas, N., 2001. Low level of brain dopamine D2 receptors
in methamphetamine abusers: association with metabolism in the orbi-
tofrontal cortex. Am. J. Psychiatry 158, 2015–2021.
Volkow, N.D., Wang, G.-J., Fowler, J.S., Logan, J., Jayne, M., Franceschi,
D., Wong, C., Gatley, S.J., Gifford, A.N., Ding, Y.-S., Pappas, N.,
2002a. ‘‘Nonhedonic’’ food motivation in humans involves dopamine
in the dorsal striatum and methylphenidate amplifies this effect. Syn-
apse 44, 175–180.
Volkow, N.D., Fowler, J.S., Wang, G.-J., 2002b. Role of dopamine in drug
reinforcement and addiction in humans: results from imaging studies.
Behav. Pharmacol. 13, 355–366.
Wang, G.-J., Volkow, N.D., Fowler, J.S., Cervany, P., Hitzemann, R.J.,
Pappas, N.R., Wong, C.T., Felder, C., 1999. Regional brain metabolic
activation during craving elicited by recall of previous drug experien-
ces. Life Sci. 64, 775–784.
Wang, G.-J., Volkow, N.D., Logan, J., Pappas, N.R., Wong, C.T., Zhu, W.,
Netusil, N., Fowler, J.S., 2001. Brain dopamine and obesity. Lancet
357, 354–357.
Wang, G.-J., Volkow, N.D., Fowler, J.S., Felder, C., Levy, A.V., Pappas,
N.R., Wong, C.T., Zhu, W., Netusil, N., 2002. Enhanced metabolism in
oral regions of somatosensory cortex in obese individuals. NeuroReport
13, 1151–1155.
Wang, G.-J., Volkow, N.D., Fowler, J.S., Franceschi, D., Wong, C.T., Pap-
pas, N.R., Netusil, N., Zhu, W., Felder, C., Ma, Y., 2003. Alcohol
intoxication induces greater reductions in brain metabolism in male than
in female subjects. Alcohol, Clin. Exp. Res. 27, 909–917.
Watababe, M., 1996. Reward expectancy in primate prefrontal neurons.
Nature 382, 629–632.
Worsley, K.J., Marrett, S., Neelin, P., Vandal, A.C., Friston, K.J., Evans,
A.C., 1996. A unified statistical approach for determining significant
signals in images of cerebral activation. Hum. Brain Mapp. 4, 58–73.
Yaxley, S., Rolls, E.T., Sienkiewicz, Z.J., 1990. Gustatory responses of
single neurons in the insula of the macaque monkey. J. Neurophysiol.
63, 689–700.
Yeomans, M.R., Gray, R.W., 2002. Opioid peptides and the control of
human ingestive behaviour. Neurosci. Biobehav. Rev. 26, 713–728.
Zald, D.H., Lee, J.T., Fluegel, K.W., Pardo, J.V., 1998. Aversive gustatory
stimulation activates limbic circuits in humans. Brain 121 (Pt 6),
1143–1154.




















