
Experience-Dependent Transfer of Otx2 Homeoprotein into the Visual Cortex Activates Postnatal...
13
Experience-Dependent Transfer of Otx2 Homeoprotein into the Visual Cortex Activates Postnatal Plasticity Sayaka Sugiyama, 1,6 Ariel A. Di Nardo, 2 Shinichi Aizawa, 3 Isao Matsuo, 4 Michel Volovitch, 2 Alain Prochiantz, 2,5, * and Takao K. Hensch 1,6,7, * 1 Lab for Neuronal Circuit Development, RIKEN Brain Science Institute, 2-1 Hirosawa, Wako-shi, Saitama 351-0198, Japan 2 CNRS UMR 8542, Ecole Normale Supe ´ rieure, 46 rue d’Ulm, 75230 Paris Cedex 05, France 3 RIKEN Center for Developmental Biology, 2-2-3 Minatojima-minamimachi, Chuo-ku, Kobe 650-0047, Japan 4 Department of Molecular Embryology, Osaka Medical Center and Research Institute for Maternal and Child Health, 840 Murodo-cho, Izumi, Osaka 594-1101, Japan 5 Colle ` ge de France, Paris, France 6 Department of Neurology, Children’s Hospital Boston, Harvard Medical School, 300 Longwood Avenue Boston, MA 02115, USA 7 Department of Molecular & Cellular Biology, Harvard University, 7 Divinity Avenue, Cambridge, MA 02138, USA *Correspondence: [email protected] (T.K.H.), [email protected] (A.P.) DOI 10.1016/j.cell.2008.05.054 SUMMARY Neural circuits are shaped by experience in early postnatal life. Distinct GABAergic connections within visual cortex determine the timing of the critical period for rewiring ocular dominance to establish visual acuity. We find that maturation of the parvalbu- min (PV)-cell network that controls plasticity onset is regulated by a selective re-expression of the embry- onic Otx2 homeoprotein. Visual experience pro- moted the accumulation of non-cell-autonomous Otx2 in PV-cells, and cortical infusion of exogenous Otx2 accelerated both PV-cell development and crit- ical period timing. Conversely, conditional removal of Otx2 from non-PV cells or from the visual pathway abolished plasticity. Thus, the experience-depen- dent transfer of a homeoprotein may establish the physiological milieu for postnatal plasticity of a neural circuit. INTRODUCTION Neural circuits are shaped by experience in postnatal life. For ex- ample, inputs from the two eyes first converge in the primary vi- sual cortex (Wiesel and Hubel, 1963), where competitive interac- tions determine which eye will eventually dominate both functionally and anatomically. From mouse to man, early monoc- ular occlusion permanently reduces visual acuity (amblyopia) through the deprived eye (Daw, 1995; Prusky and Douglas, 2003). Unbalanced sensory experience leads to a rapid shift of neuronal spiking response in favor of the open eye (ocular dom- inance) concomitant with a rapid pruning of dendritic spines and later rewiring of thalamo-cortical afferents (Wiesel and Hubel, 1963; Antonini et al., 1999; Hensch, 2005). Recently, the maturation of particular inhibitory connections within the neocortex has been shown to drive this sensitivity (Hensch, 2005). Mice lacking the synaptic isoform of GABA-syn- thetic enzyme, glutamic acid decarboxylase (GAD65), fail to shift visual responsiveness (Hensch et al., 1998) or to lose dendritic spines (Mataga et al., 2004) upon brief monocular deprivation (MD). Both can be rescued by concurrent benzodiazepine treat- ment (to enhance GABAergic transmission). Raising animals in complete darkness from birth (dark-rearing) naturally delays the maturation of intracortical inhibition and ocular dominance plas- ticity (Mower, 1991; Morales et al., 2002; Iwai et al., 2003; Katagiri et al., 2007). Conversely, benzodiazepines (BDZ) induce a prema- ture plasticity in young wild-type mice just after eye-opening (postnatal day, P13) (Fagiolini and Hensch, 2000; Fagiolini et al., 2004). Inhibition triggers plasticity through GABA A receptors con- taining the a1-subunit (Fagiolini et al., 2004; Katagiri et al., 2007). These are enriched at somatic synapses on pyramidal cells made by large-basket cells containing the calcium-bind- ing protein parvalbumin (PV) (Klausberger et al., 2002). Unlike other GABAergic interneurons, PV-cells emerge with a late postnatal timecourse in anticipation of critical period onset (del Rio et al., 1994). Overexpression of brain-derived neurotro- phic factor (BDNF) accelerates both PV-cell development and critical period timecourse (Huang et al., 1999; Hanover et al., 1999). Gradually with age, peri-neuronal nets (PNNs) of extracellular matrix molecules condense around PV-cells in parallel with the decline of critical period plasticity (Celio et al., 1998; Pizzorusso et al., 2002). Importantly, chondroitinase treatment to destroy these nets reactivates visual cortical plasticity in adult animals (Pizzorusso et al., 2002). Moreover, the wide basket-cell axon plexus stretches horizontally across ocular dominance columns in the cat and is ideally situated to shape their final spacing through competitive mechanisms in development (Buza ´ s et al., 2001; Hensch and Stryker, 2004). 508 Cell 134, 508–520, August 8, 2008 ª2008 Elsevier Inc.
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Experience-Dependent Transfer of Otx2 Homeoprotein into the Visual Cortex Activates Postnatal...

Experience-Dependent Transfer ofOtx2 Homeoprotein into the VisualCortex Activates Postnatal PlasticitySayaka Sugiyama,1,6 Ariel A. Di Nardo,2 Shinichi Aizawa,3 Isao Matsuo,4 Michel Volovitch,2 Alain Prochiantz,2,5,*and Takao K. Hensch1,6,7,*1Lab for Neuronal Circuit Development, RIKEN Brain Science Institute, 2-1 Hirosawa, Wako-shi, Saitama 351-0198, Japan2CNRS UMR 8542, Ecole Normale Superieure, 46 rue d’Ulm, 75230 Paris Cedex 05, France3RIKEN Center for Developmental Biology, 2-2-3 Minatojima-minamimachi, Chuo-ku, Kobe 650-0047, Japan4Department of Molecular Embryology, Osaka Medical Center and Research Institute for Maternal and Child Health, 840 Murodo-cho, Izumi,
Osaka 594-1101, Japan5College de France, Paris, France6Department of Neurology, Children’s Hospital Boston, Harvard Medical School, 300 Longwood Avenue Boston, MA 02115, USA7Department of Molecular & Cellular Biology, Harvard University, 7 Divinity Avenue, Cambridge, MA 02138, USA
*Correspondence: [email protected] (T.K.H.), [email protected] (A.P.)
DOI 10.1016/j.cell.2008.05.054
SUMMARY
Neural circuits are shaped by experience in earlypostnatal life. Distinct GABAergic connections withinvisual cortex determine the timing of the criticalperiod for rewiring ocular dominance to establishvisual acuity. We find that maturation of the parvalbu-min (PV)-cell network that controls plasticity onset isregulated by a selective re-expression of the embry-onic Otx2 homeoprotein. Visual experience pro-moted the accumulation of non-cell-autonomousOtx2 in PV-cells, and cortical infusion of exogenousOtx2 accelerated both PV-cell development and crit-ical period timing. Conversely, conditional removal ofOtx2 from non-PV cells or from the visual pathwayabolished plasticity. Thus, the experience-depen-dent transfer of a homeoprotein may establish thephysiological milieu for postnatal plasticity of a neuralcircuit.
INTRODUCTION
Neural circuits are shaped by experience in postnatal life. For ex-
ample, inputs from the two eyes first converge in the primary vi-
sual cortex (Wiesel and Hubel, 1963), where competitive interac-
tions determine which eye will eventually dominate both
functionally and anatomically. From mouse to man, early monoc-
ular occlusion permanently reduces visual acuity (amblyopia)
through the deprived eye (Daw, 1995; Prusky and Douglas,
2003). Unbalanced sensory experience leads to a rapid shift of
neuronal spiking response in favor of the open eye (ocular dom-
inance) concomitant with a rapid pruning of dendritic spines and
later rewiring of thalamo-cortical afferents (Wiesel and Hubel,
1963; Antonini et al., 1999; Hensch, 2005).
508 Cell 134, 508–520, August 8, 2008 ª2008 Elsevier Inc.
Recently, the maturation of particular inhibitory connections
within the neocortex has been shown to drive this sensitivity
(Hensch, 2005). Mice lacking the synaptic isoform of GABA-syn-
thetic enzyme, glutamic acid decarboxylase (GAD65), fail to shift
visual responsiveness (Hensch et al., 1998) or to lose dendritic
spines (Mataga et al., 2004) upon brief monocular deprivation
(MD). Both can be rescued by concurrent benzodiazepine treat-
ment (to enhance GABAergic transmission). Raising animals in
complete darkness from birth (dark-rearing) naturally delays the
maturation of intracortical inhibition and ocular dominance plas-
ticity (Mower, 1991; Morales et al., 2002; Iwai et al., 2003; Katagiri
et al., 2007). Conversely, benzodiazepines (BDZ) induce a prema-
ture plasticity in young wild-type mice just after eye-opening
(postnatal day, P13) (Fagiolini and Hensch, 2000; Fagiolini
et al., 2004).
Inhibition triggers plasticity through GABAA receptors con-
taining the a1-subunit (Fagiolini et al., 2004; Katagiri et al.,
2007). These are enriched at somatic synapses on pyramidal
cells made by large-basket cells containing the calcium-bind-
ing protein parvalbumin (PV) (Klausberger et al., 2002). Unlike
other GABAergic interneurons, PV-cells emerge with a late
postnatal timecourse in anticipation of critical period onset
(del Rio et al., 1994). Overexpression of brain-derived neurotro-
phic factor (BDNF) accelerates both PV-cell development and
critical period timecourse (Huang et al., 1999; Hanover et al.,
1999).
Gradually with age, peri-neuronal nets (PNNs) of extracellular
matrix molecules condense around PV-cells in parallel with the
decline of critical period plasticity (Celio et al., 1998; Pizzorusso
et al., 2002). Importantly, chondroitinase treatment to destroy
these nets reactivates visual cortical plasticity in adult animals
(Pizzorusso et al., 2002). Moreover, the wide basket-cell axon
plexus stretches horizontally across ocular dominance columns
in the cat and is ideally situated to shape their final spacing
through competitive mechanisms in development (Buzas et al.,
2001; Hensch and Stryker, 2004).
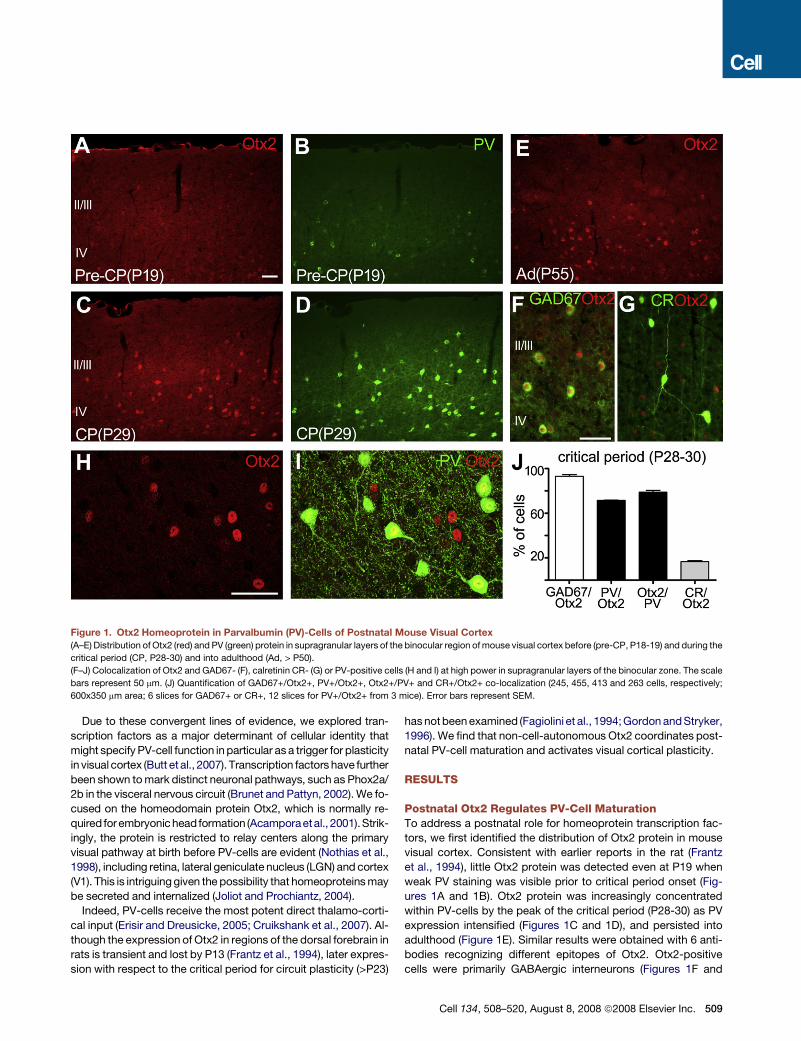
Figure 1. Otx2 Homeoprotein in Parvalbumin (PV)-Cells of Postnatal Mouse Visual Cortex
(A–E) Distribution of Otx2 (red) and PV (green) protein in supragranular layers of the binocular region of mouse visual cortex before (pre-CP, P18-19) and during the
critical period (CP, P28-30) and into adulthood (Ad, > P50).
(F–J) Colocalization of Otx2 and GAD67- (F), calretinin CR- (G) or PV-positive cells (H and I) at high power in supragranular layers of the binocular zone. The scale
bars represent 50 mm. (J) Quantification of GAD67+/Otx2+, PV+/Otx2+, Otx2+/PV+ and CR+/Otx2+ co-localization (245, 455, 413 and 263 cells, respectively;
600x350 mm area; 6 slices for GAD67+ or CR+, 12 slices for PV+/Otx2+ from 3 mice). Error bars represent SEM.
Due to these convergent lines of evidence, we explored tran-
scription factors as a major determinant of cellular identity that
might specify PV-cell function in particular as a trigger for plasticity
in visual cortex (Butt et al., 2007). Transcription factors have further
been shown to mark distinct neuronal pathways, such as Phox2a/
2b in the visceral nervous circuit (Brunet and Pattyn, 2002). We fo-
cused on the homeodomain protein Otx2, which is normally re-
quired forembryonic head formation (Acamporaetal., 2001).Strik-
ingly, the protein is restricted to relay centers along the primary
visual pathway at birth before PV-cells are evident (Nothias et al.,
1998), including retina, lateral geniculate nucleus (LGN) and cortex
(V1). This is intriguing given the possibility that homeoproteins may
be secreted and internalized (Joliot and Prochiantz, 2004).
Indeed, PV-cells receive the most potent direct thalamo-corti-
cal input (Erisir and Dreusicke, 2005; Cruikshank et al., 2007). Al-
though the expression of Otx2 in regions of the dorsal forebrain in
rats is transient and lost by P13 (Frantz et al., 1994), later expres-
sion with respect to the critical period for circuit plasticity (>P23)
has not been examined (Fagiolini et al., 1994; Gordon and Stryker,
1996). We find that non-cell-autonomous Otx2 coordinates post-
natal PV-cell maturation and activates visual cortical plasticity.
RESULTS
Postnatal Otx2 Regulates PV-Cell MaturationTo address a postnatal role for homeoprotein transcription fac-
tors, we first identified the distribution of Otx2 protein in mouse
visual cortex. Consistent with earlier reports in the rat (Frantz
et al., 1994), little Otx2 protein was detected even at P19 when
weak PV staining was visible prior to critical period onset (Fig-
ures 1A and 1B). Otx2 protein was increasingly concentrated
within PV-cells by the peak of the critical period (P28-30) as PV
expression intensified (Figures 1C and 1D), and persisted into
adulthood (Figure 1E). Similar results were obtained with 6 anti-
bodies recognizing different epitopes of Otx2. Otx2-positive
cells were primarily GABAergic interneurons (Figures 1F and
Cell 134, 508–520, August 8, 2008 ª2008 Elsevier Inc. 509
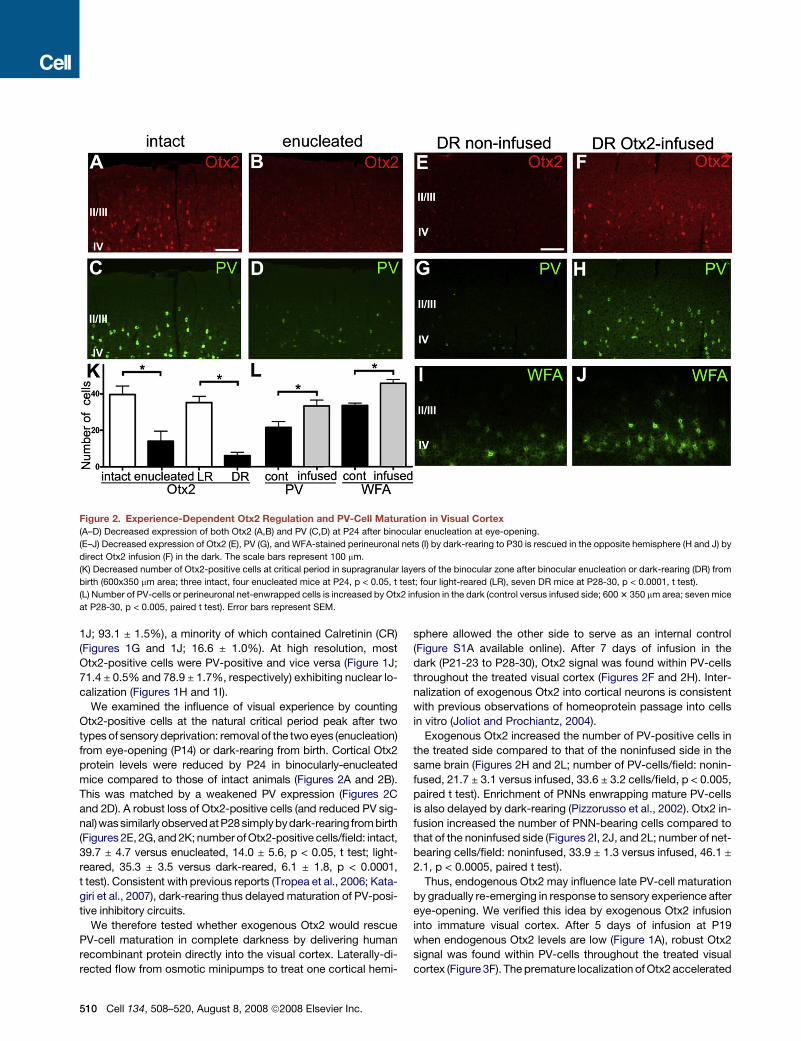
Figure 2. Experience-Dependent Otx2 Regulation and PV-Cell Maturation in Visual Cortex
(A–D) Decreased expression of both Otx2 (A,B) and PV (C,D) at P24 after binocular enucleation at eye-opening.
(E–J) Decreased expression of Otx2 (E), PV (G), and WFA-stained perineuronal nets (I) by dark-rearing to P30 is rescued in the opposite hemisphere (H and J) by
direct Otx2 infusion (F) in the dark. The scale bars represent 100 mm.
(K) Decreased number of Otx2-positive cells at critical period in supragranular layers of the binocular zone after binocular enucleation or dark-rearing (DR) from
birth (600x350 mm area; three intact, four enucleated mice at P24, p < 0.05, t test; four light-reared (LR), seven DR mice at P28-30, p < 0.0001, t test).
(L) Number of PV-cells or perineuronal net-enwrapped cells is increased by Otx2 infusion in the dark (control versus infused side; 600 3 350 mm area; seven mice
at P28-30, p < 0.005, paired t test). Error bars represent SEM.
1J; 93.1 ± 1.5%), a minority of which contained Calretinin (CR)
(Figures 1G and 1J; 16.6 ± 1.0%). At high resolution, most
Otx2-positive cells were PV-positive and vice versa (Figure 1J;
71.4 ± 0.5% and 78.9 ± 1.7%, respectively) exhibiting nuclear lo-
calization (Figures 1H and 1I).
We examined the influence of visual experience by counting
Otx2-positive cells at the natural critical period peak after two
types of sensory deprivation: removal of the two eyes (enucleation)
from eye-opening (P14) or dark-rearing from birth. Cortical Otx2
protein levels were reduced by P24 in binocularly-enucleated
mice compared to those of intact animals (Figures 2A and 2B).
This was matched by a weakened PV expression (Figures 2C
and 2D). A robust loss of Otx2-positive cells (and reduced PV sig-
nal)was similarly observedatP28 simplybydark-rearing from birth
(Figures 2E, 2G, and 2K; number of Otx2-positive cells/field: intact,
39.7 ± 4.7 versus enucleated, 14.0 ± 5.6, p < 0.05, t test; light-
reared, 35.3 ± 3.5 versus dark-reared, 6.1 ± 1.8, p < 0.0001,
t test). Consistent with previous reports (Tropea et al., 2006; Kata-
giri et al., 2007), dark-rearing thus delayed maturation of PV-posi-
tive inhibitory circuits.
We therefore tested whether exogenous Otx2 would rescue
PV-cell maturation in complete darkness by delivering human
recombinant protein directly into the visual cortex. Laterally-di-
rected flow from osmotic minipumps to treat one cortical hemi-
510 Cell 134, 508–520, August 8, 2008 ª2008 Elsevier Inc.
sphere allowed the other side to serve as an internal control
(Figure S1A available online). After 7 days of infusion in the
dark (P21-23 to P28-30), Otx2 signal was found within PV-cells
throughout the treated visual cortex (Figures 2F and 2H). Inter-
nalization of exogenous Otx2 into cortical neurons is consistent
with previous observations of homeoprotein passage into cells
in vitro (Joliot and Prochiantz, 2004).
Exogenous Otx2 increased the number of PV-positive cells in
the treated side compared to that of the noninfused side in the
same brain (Figures 2H and 2L; number of PV-cells/field: nonin-
fused, 21.7 ± 3.1 versus infused, 33.6 ± 3.2 cells/field, p < 0.005,
paired t test). Enrichment of PNNs enwrapping mature PV-cells
is also delayed by dark-rearing (Pizzorusso et al., 2002). Otx2 in-
fusion increased the number of PNN-bearing cells compared to
that of the noninfused side (Figures 2I, 2J, and 2L; number of net-
bearing cells/field: noninfused, 33.9 ± 1.3 versus infused, 46.1 ±
2.1, p < 0.0005, paired t test).
Thus, endogenous Otx2 may influence late PV-cell maturation
by gradually re-emerging in response to sensory experience after
eye-opening. We verified this idea by exogenous Otx2 infusion
into immature visual cortex. After 5 days of infusion at P19
when endogenous Otx2 levels are low (Figure 1A), robust Otx2
signal was found within PV-cells throughout the treated visual
cortex (Figure 3F). The premature localization of Otx2 accelerated
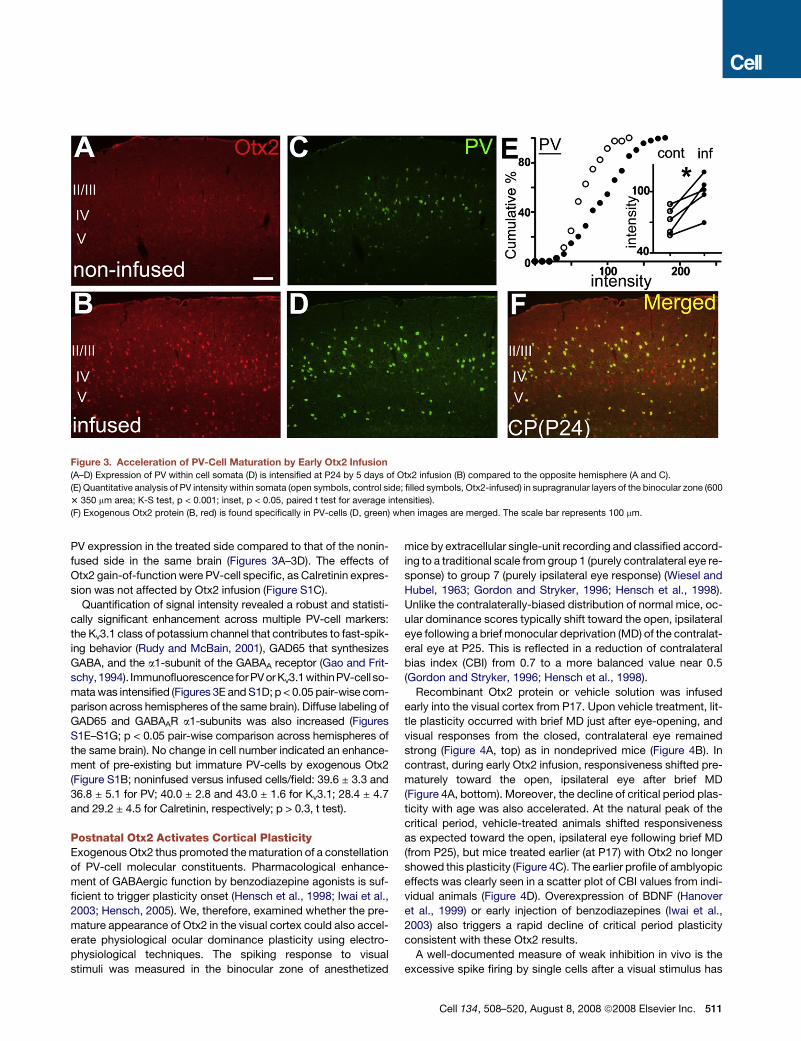
Figure 3. Acceleration of PV-Cell Maturation by Early Otx2 Infusion
(A–D) Expression of PV within cell somata (D) is intensified at P24 by 5 days of Otx2 infusion (B) compared to the opposite hemisphere (A and C).
(E) Quantitative analysis of PV intensity within somata (open symbols, control side; filled symbols, Otx2-infused) in supragranular layers of the binocular zone (600
3 350 mm area; K-S test, p < 0.001; inset, p < 0.05, paired t test for average intensities).
(F) Exogenous Otx2 protein (B, red) is found specifically in PV-cells (D, green) when images are merged. The scale bar represents 100 mm.
PV expression in the treated side compared to that of the nonin-
fused side in the same brain (Figures 3A–3D). The effects of
Otx2 gain-of-function were PV-cell specific, as Calretinin expres-
sion was not affected by Otx2 infusion (Figure S1C).
Quantification of signal intensity revealed a robust and statisti-
cally significant enhancement across multiple PV-cell markers:
the Kv3.1 class of potassium channel that contributes to fast-spik-
ing behavior (Rudy and McBain, 2001), GAD65 that synthesizes
GABA, and the a1-subunit of the GABAA receptor (Gao and Frit-
schy,1994). Immunofluorescence forPV or Kv3.1withinPV-cell so-
mata was intensified (Figures 3E and S1D; p < 0.05 pair-wise com-
parison across hemispheres of the same brain). Diffuse labeling of
GAD65 and GABAAR a1-subunits was also increased (Figures
S1E–S1G; p < 0.05 pair-wise comparison across hemispheres of
the same brain). No change in cell number indicated an enhance-
ment of pre-existing but immature PV-cells by exogenous Otx2
(Figure S1B; noninfused versus infused cells/field: 39.6 ± 3.3 and
36.8 ± 5.1 for PV; 40.0 ± 2.8 and 43.0 ± 1.6 for Kv3.1; 28.4 ± 4.7
and 29.2 ± 4.5 for Calretinin, respectively; p > 0.3, t test).
Postnatal Otx2 Activates Cortical PlasticityExogenous Otx2 thus promoted the maturation of a constellation
of PV-cell molecular constituents. Pharmacological enhance-
ment of GABAergic function by benzodiazepine agonists is suf-
ficient to trigger plasticity onset (Hensch et al., 1998; Iwai et al.,
2003; Hensch, 2005). We, therefore, examined whether the pre-
mature appearance of Otx2 in the visual cortex could also accel-
erate physiological ocular dominance plasticity using electro-
physiological techniques. The spiking response to visual
stimuli was measured in the binocular zone of anesthetized
mice by extracellular single-unit recording and classified accord-
ing to a traditional scale from group 1 (purely contralateral eye re-
sponse) to group 7 (purely ipsilateral eye response) (Wiesel and
Hubel, 1963; Gordon and Stryker, 1996; Hensch et al., 1998).
Unlike the contralaterally-biased distribution of normal mice, oc-
ular dominance scores typically shift toward the open, ipsilateral
eye following a brief monocular deprivation (MD) of the contralat-
eral eye at P25. This is reflected in a reduction of contralateral
bias index (CBI) from 0.7 to a more balanced value near 0.5
(Gordon and Stryker, 1996; Hensch et al., 1998).
Recombinant Otx2 protein or vehicle solution was infused
early into the visual cortex from P17. Upon vehicle treatment, lit-
tle plasticity occurred with brief MD just after eye-opening, and
visual responses from the closed, contralateral eye remained
strong (Figure 4A, top) as in nondeprived mice (Figure 4B). In
contrast, during early Otx2 infusion, responsiveness shifted pre-
maturely toward the open, ipsilateral eye after brief MD
(Figure 4A, bottom). Moreover, the decline of critical period plas-
ticity with age was also accelerated. At the natural peak of the
critical period, vehicle-treated animals shifted responsiveness
as expected toward the open, ipsilateral eye following brief MD
(from P25), but mice treated earlier (at P17) with Otx2 no longer
showed this plasticity (Figure 4C). The earlier profile of amblyopic
effects was clearly seen in a scatter plot of CBI values from indi-
vidual animals (Figure 4D). Overexpression of BDNF (Hanover
et al., 1999) or early injection of benzodiazepines (Iwai et al.,
2003) also triggers a rapid decline of critical period plasticity
consistent with these Otx2 results.
A well-documented measure of weak inhibition in vivo is the
excessive spike firing by single cells after a visual stimulus has
Cell 134, 508–520, August 8, 2008 ª2008 Elsevier Inc. 511
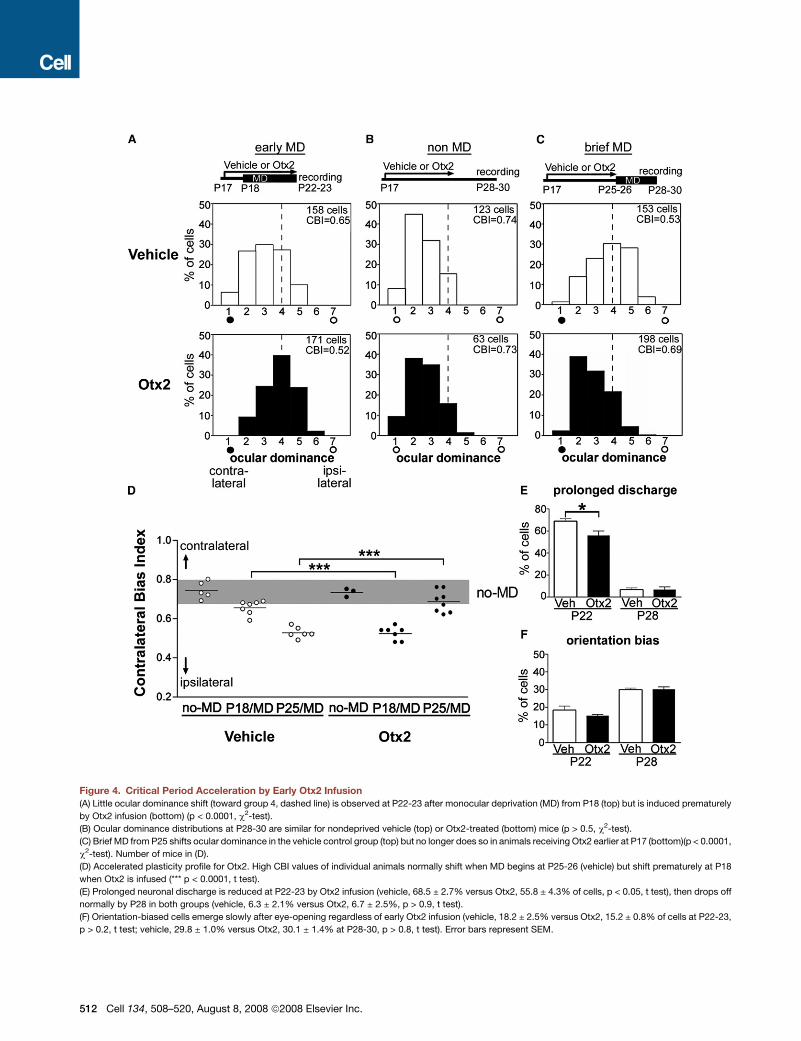
Figure 4. Critical Period Acceleration by Early Otx2 Infusion
(A) Little ocular dominance shift (toward group 4, dashed line) is observed at P22-23 after monocular deprivation (MD) from P18 (top) but is induced prematurely
by Otx2 infusion (bottom) (p < 0.0001, c2-test).
(B) Ocular dominance distributions at P28-30 are similar for nondeprived vehicle (top) or Otx2-treated (bottom) mice (p > 0.5, c2-test).
(C) Brief MD from P25 shifts ocular dominance in the vehicle control group (top) but no longer does so in animals receiving Otx2 earlier at P17 (bottom)(p < 0.0001,
c2-test). Number of mice in (D).
(D) Accelerated plasticity profile for Otx2. High CBI values of individual animals normally shift when MD begins at P25-26 (vehicle) but shift prematurely at P18
when Otx2 is infused (*** p < 0.0001, t test).
(E) Prolonged neuronal discharge is reduced at P22-23 by Otx2 infusion (vehicle, 68.5 ± 2.7% versus Otx2, 55.8 ± 4.3% of cells, p < 0.05, t test), then drops off
normally by P28 in both groups (vehicle, 6.3 ± 2.1% versus Otx2, 6.7 ± 2.5%, p > 0.9, t test).
(F) Orientation-biased cells emerge slowly after eye-opening regardless of early Otx2 infusion (vehicle, 18.2 ± 2.5% versus Otx2, 15.2 ± 0.8% of cells at P22-23,
p > 0.2, t test; vehicle, 29.8 ± 1.0% versus Otx2, 30.1 ± 1.4% at P28-30, p > 0.8, t test). Error bars represent SEM.
512 Cell 134, 508–520, August 8, 2008 ª2008 Elsevier Inc.

left the receptive field (prolonged discharge). This is typically
prominent in GAD65 knockout (Hensch et al., 1998; Iwai et al.,
2003) or young wild-type mice before the critical period
(Iwai et al., 2003; Fagiolini et al., 2004). Like benzodiazepine
treatment (Hensch et al., 1998), early Otx2 infusion significantly
reduced this immature property in vivo, indicating a rapid func-
tional maturation of inhibitory circuits by this homeoprotein
(Figure 4E). Other receptive field properties, such as low orienta-
tion bias at young ages, were not affected by early Otx2 infusion
(Figure 4F and Table S1), consistent with a molecular dissocia-
tion of ocular dominance and orientation development as re-
ported previously (Hensch et al., 1998; Fagiolini et al., 2003).
Taken together, Otx2 gain-of-function promoted critical period
onset in conjunction with PV-cell maturation.
To confirm whether Otx2 is required for PV-cell maturation and
critical period plasticity, we next carried out Otx2 loss-of-func-
tion experiments. Due to the embryonic lethality of constitutive
Otx2 deletion (Acampora et al., 2001), we turned to a conditional
approach. Otx2 protein was detected within PV-cells at P28 in
Otx2 flox/flox mice in which the Otx2 gene was flanked by LoxP
sites (Tian et al., 2002) (Figure 5A). In contrast, when one Otx2
allele was excised by Cre recombinase under the control of
a postnatal a-calcium/calmodulin-dependent protein kinase II
(aCaMKII) promoter, cortical Otx2 signal was strongly reduced
(Figure 5B). Strikingly, PV expression was also reduced when en-
dogenous Otx2 was disrupted (Figures 5C and 5D), retaining an
immature appearance (see Figure 1B). Immunofluorescence
intensity within PV-cell somata was significantly decreased in
the cumulative distribution or comparisons of average intensity
for individual mice (Figure 5G).
Loss of Otx2 also delayed PNN formation and reduced the
number of cells labeled by WFA (Figures 5F and 5H; control,
43.9 ± 1.8 versus hetKO, 32.0 ± 1.8 cells/field at P28-30; control,
59.9 ± 1.7 versus hetKO, 44.6 ± 1.7 for adults, > P60, respec-
tively; p < 0.0005, t test) or Cat-301-antigen (Figures S2F and
S2G; control, 30.1 ± 1.9 versus hetKO, 15.6 ± 1.8 cells/field at
P28-30; control, 56.8 ± 3.4 versus hetKO, 37.2 ± 2.7 for adults, >
P60, respectively; p < 0.0005, t test). Moreover, PV-expressing
cell number was also reduced by the loss of endogenous Otx2
(Figure S2D; control, 38.0 ± 4.4 versus hetKO, 28.6 ± 1.0 cells/
field, p < 0.05, t test). These effects were specific to PV-cells
since Calretinin-cells were not affected in expression level (Fig-
ures S2A–S2C) or number (Figure S2D; control, 18.5 ± 1.3 versus
hetKO, 20.3 ± 2.3 cells/field, p > 0.5, t test).
Postnatal Otx2 loss-of-function would then be expected to
prevent plasticity if somatic inhibition is compromised (Katagiri
et al., 2007), as for global GAD65 disruption (Hensch et al.,
1998; Hensch, 2005). The frequency of miniature IPSCs, arising
primarily from peri-somatic inputs (Soltesz et al., 1995), was sig-
nificantly reduced in slices from Otx2 KO mice (A. Scheuber and
T.K.H., unpublished data). Ocular dominance plasticity failed to
occur concomitant with immature PV expression (Figure 5D) in
mice lacking even one Otx2 allele (Figures 5I, 5J, and 5L), which
had no noticeable effect on other receptive field properties (re-
sponsiveness, orientation bias, habituation; Table S1). We then
attempted to rescue plasticity in vivo using benzodiazepine ag-
onists to enhance GABAA receptor currents. Ocular dominance
shifts were restored by diazepam treatment during brief MD (Fig-
ures 5K and 5L) as in GAD65 KO mice (Hensch et al., 1998). En-
dogenous Otx2 may thus activate critical period plasticity
through the functional maturation of inhibitory circuits.
Non-Cell-Autonomous Otx2 in PV-CellsRemarkably, inhibitory interneurons in the cortex do not express
aCaMKII (Liu and Jones, 1996; Sik et al., 1998). Crossing
Otx2flox/flox mice with those expressing a Cre-recombinase
driven by the aCaMKII promoter above (Minichiello et al., 1999)
thus results in a conditional disruption of Otx2 synthesis outside
cortical GABA cells. The impairment of PV-cell maturation and
critical period plasticity (Figure 5) must therefore reflect a non-
cell-autonomous action of Otx2.
Consistent with this idea, Otx2 transcripts were not detected in
the visual cortex by in situ hybridization analyses (Figure 6A) or
RT-PCR (Figure S3A) during the critical period. Green fluores-
cent protein (GFP) transcribed from the Otx2 locus in Otx2-
GFP knock-in mice (c/o Drs. D. Acampora and A. Simeone)
also failed to show any cortical signal (Figure 6B). Instead,
both Otx2 mRNA and Otx2-GFP expression were observed as
expected at sub-cortical stages of the visual pathway in the lat-
eral geniculate nucleus (LGN, Figures 6D and 6E), superior colli-
culus (Figures S3A and S3B) and retina (Figures 6G, 6H, S4A,
and S4C) (Bovolenta et al., 1997; Baas et al., 2000; Frantz
et al., 1994; Nothias et al., 1998). These observations confirm
that Otx2 is not synthesized in cortical PV-cells themselves.
Homeoprotein family members have been shown to contain
secretion and internalization signals in the homeodomain (Joliot
and Prochiantz, 2004) and are found in the secretory compart-
ment in vivo (Joliot et al., 1997). This would allow potential trans-
fer from cell to cell as observed in vitro (Joliot and Prochiantz,
2004), raising the possibility that Otx2 homeoprotein may be
propagated into the visual cortex along the visual pathway
(Figure 6C). Even within the retina Otx2 protein was found in gan-
glion cells (Figure 6I), downstream of the bipolar cells containing
its mRNA in the inner nuclear layer (Figure 6G). In the LGN, Otx2
protein was diffusely spread over multiple cells (Figure 6F) unlike
Otx2 transcripts found within discrete GABAergic interneurons
(Figure 6D; GABA+/Otx2+, 93.7 ± 1.4%, 283 cells; Otx2+/
GABA+, 92.0 ± 2.4%, 287 cells; 4 mice; Sugino et al., 2006).
Otx2 along the Visual PathwayTo demonstrate a potential route of transport for Otx2 protein
in vivo along the visual pathway (Figure 6C), we introduced
a tagged protein into the eyes. After an intraocular injection of
biotinylated Otx2, labeled protein was detected in the visual
cortex, where the biotin tag was internalized into cortical cells
(Figure 6J). Notably, the proportion of cells that were both
Otx2-labeled and PV-positive (Figures 6K and 6L; 77.7 ± 3.6%)
was identical to the endogenous ratio (Figure 1J). Consistent
with its non-cell-autonomous distribution in the visual cortex,
neither uninjected nor BSA-biotin injected animals exhibited
such a cortical label (Figures 6M–6O).
Otx2 expression at sub-cortical stages persists from early
postnatal days before eye-opening (Frantz et al., 1994; Baas
et al., 2000). In our conditional deletion mice, Otx2 transcripts
were reduced in the inner nuclear layer of the retina by aCaMKII
promoter-mediated Cre expression (Figures S4A and S4B; Liu
Cell 134, 508–520, August 8, 2008 ª2008 Elsevier Inc. 513
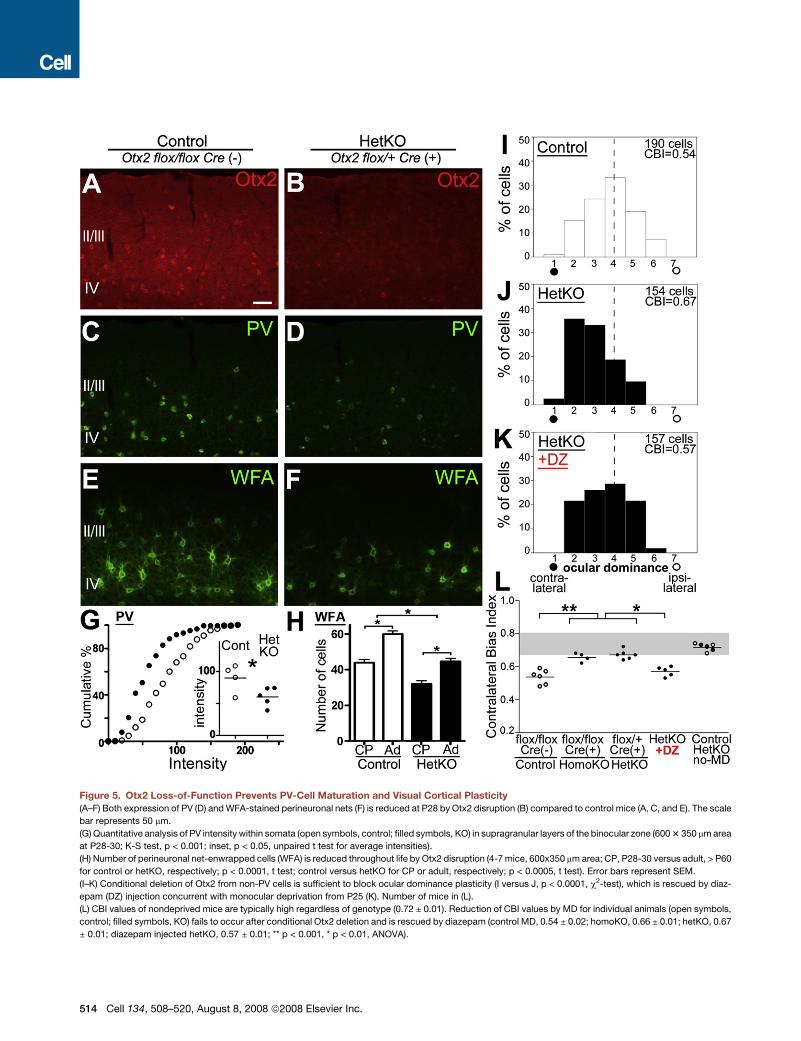
Figure 5. Otx2 Loss-of-Function Prevents PV-Cell Maturation and Visual Cortical Plasticity
(A–F) Both expression of PV (D) and WFA-stained perineuronal nets (F) is reduced at P28 by Otx2 disruption (B) compared to control mice (A, C, and E). The scale
bar represents 50 mm.
(G) Quantitative analysis of PV intensity within somata (open symbols, control; filled symbols, KO) in supragranular layers of the binocular zone (600 3 350 mm area
at P28-30; K-S test, p < 0.001; inset, p < 0.05, unpaired t test for average intensities).
(H) Number of perineuronal net-enwrapped cells (WFA) is reduced throughout life by Otx2 disruption (4-7 mice, 600x350 mm area; CP, P28-30 versus adult, > P60
for control or hetKO, respectively; p < 0.0001, t test; control versus hetKO for CP or adult, respectively; p < 0.0005, t test). Error bars represent SEM.
(I–K) Conditional deletion of Otx2 from non-PV cells is sufficient to block ocular dominance plasticity (I versus J, p < 0.0001, c2-test), which is rescued by diaz-
epam (DZ) injection concurrent with monocular deprivation from P25 (K). Number of mice in (L).
(L) CBI values of nondeprived mice are typically high regardless of genotype (0.72 ± 0.01). Reduction of CBI values by MD for individual animals (open symbols,
control; filled symbols, KO) fails to occur after conditional Otx2 deletion and is rescued by diazepam (control MD, 0.54 ± 0.02; homoKO, 0.66 ± 0.01; hetKO, 0.67
± 0.01; diazepam injected hetKO, 0.57 ± 0.01; ** p < 0.001, * p < 0.01, ANOVA).
514 Cell 134, 508–520, August 8, 2008 ª2008 Elsevier Inc.
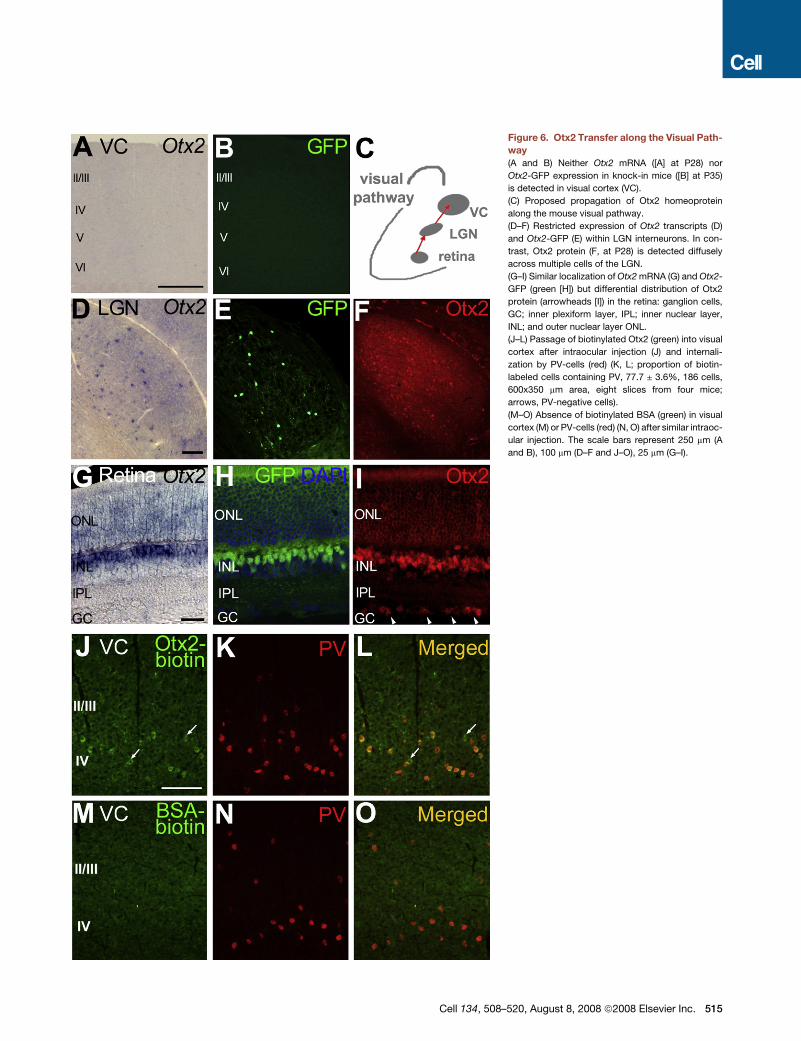
Figure 6. Otx2 Transfer along the Visual Path-
way
(A and B) Neither Otx2 mRNA ([A] at P28) nor
Otx2-GFP expression in knock-in mice ([B] at P35)
is detected in visual cortex (VC).
(C) Proposed propagation of Otx2 homeoprotein
along the mouse visual pathway.
(D–F) Restricted expression of Otx2 transcripts (D)
and Otx2-GFP (E) within LGN interneurons. In con-
trast, Otx2 protein (F, at P28) is detected diffusely
across multiple cells of the LGN.
(G–I) Similar localization of Otx2 mRNA (G) and Otx2-
GFP (green [H]) but differential distribution of Otx2
protein (arrowheads [I]) in the retina: ganglion cells,
GC; inner plexiform layer, IPL; inner nuclear layer,
INL; and outer nuclear layer ONL.
(J–L) Passage of biotinylated Otx2 (green) into visual
cortex after intraocular injection (J) and internali-
zation by PV-cells (red) (K, L; proportion of biotin-
labeled cells containing PV, 77.7 ± 3.6%, 186 cells,
600x350 mm area, eight slices from four mice;
arrows, PV-negative cells).
(M–O) Absence of biotinylated BSA (green) in visual
cortex (M) or PV-cells (red) (N, O) after similar intraoc-
ular injection. The scale bars represent 250 mm (A
and B), 100 mm (D–F and J–O), 25 mm (G–I).
Cell 134, 508–520, August 8, 2008 ª2008 Elsevier Inc. 515

et al., 2000). Although visual experience through the eyes was re-
quired for cortical Otx2 appearance, retinal Otx2 synthesis was
unaffected by dark-rearing (Figures S3C and S3D). The emer-
gence of endogenous Otx2 within visual cortex may then include
an experience-dependent transport or transfer, which might
explain homeoprotein accumulation within cortical PV-cells
only after eye-opening and the onset of vision.
We directly addressed the putative passage of Otx2 by local
infusion of antagonists either into the eye or cortex (Figure 7A).
To spatio-temporally intercept the extracellular transfer of Otx2
we used a specific antibody. Seven-day infusion of antibody
into visual cortex (P21-23 to P28-30) was sufficient to capture
and clear Otx2 in the extracellular space (Figures S4E and
S4F). To knock-down the synthesis of homeoprotein we gener-
ated short-interfering RNA (siRNA) constructs coupled to pene-
tratin (Joliot and Prochiantz, 2004). siRNA efficacy was verified in
HeLa cells expressing Otx2 in vitro and in the inner nuclear layer
of the mouse retina in vivo (Figures 7E, S4C, and S4D). These
treatments showed no general effect upon receptive field prop-
erties in visual cortex (Table S1).
Instead, both early intraocular injection and intracortical infu-
sion of anti-Otx2 antibody successfully prevented plasticity (Fig-
ures 7B and 7C). Ocular dominance remained skewed in favor of
the contralateral eye (left of group 4) even after brief MD at P25
(Figure 7F ‘‘a-Otx2’’). In contrast, intracortical siRNA injection
did not block critical period plasticity (Figure 7D), consistent
with the absence of Otx2 mRNA in the cortex (Figure 6A). To pre-
vent plasticity, siRNA was effective only in the eye (Figure 7F
‘‘siRNA’’). Taken together, these results support the idea that
Otx2 is synthesized and transferred from outside the visual
cortex to enable visual cortical plasticity.
DISCUSSION
Our results demonstrate a novel postnatal role for an embryonic
homeoprotein, Otx2, in critical period brain development. Its
mechanism of action is entirely consistent with recent under-
standing of plasticity in the visual cortex based upon the activa-
tion of particular GABAergic connections (Hensch, 2005). Endur-
ing behavioral amblyopia (loss of visual acuity after MD) reflects
a shift in spiking output from V1 during a restricted critical period
(Gordon & Stryker, 1996; Prusky and Douglas, 2003; Tohmi et al.,
2006; Morishita and Hensch, 2008). Spike timing is dictated by
the maturation of GABAergic transmission at somatic synapses
containing the GABAA a1- receptor subunit (Fagiolini et al., 2004;
Katagiri et al., 2007), typically innervated by large-basket PV-
cells which mature postnatally in anticipation of the critical pe-
riod (del Rio et al., 1994; Huang et al., 1999; Di Cristo et al., 2007).
Direct Otx2 gain- or loss-of-function, respectively, promoted
or hindered both the expression of PV-cell molecular constitu-
ents and ocular dominance plasticity. Surprisingly, these bidirec-
tional actions were non-cell-autonomous. Selective deletion of
the Otx2 gene outside cortical interneurons successfully re-
moved Otx2 protein from PV-cells. Moreover, intercepting
Otx2 by infusion of antibodies or blocking its synthesis by siRNA
injection into the eye prevented critical period plasticity. It is in-
triguing that passage of extra-cortical factors may establish
the milieu for cortical plasticity in vivo. Robust Otx2 mRNA ex-
516 Cell 134, 508–520, August 8, 2008 ª2008 Elsevier Inc.
pression is observed in the retina, LGN and superior colliculus,
demarcating a possible conduit for propagation along the visual
pathway (Bovolenta et al., 1997; Baas et al., 2000; Frantz et al.,
1994; Nothias et al., 1998; Sugino et al., 2006; Rath et al.,
2007). The mechanisms of transport and transfer in vivo will be
an important line of investigation.
Homeoprotein family members contain secretion and internal-
ization sequences in the homeodomain, allowing potential trans-
fer from cell to cell as observed in vitro (Joliot and Prochiantz,
2004; Brunet et al., 2007). Recent studies have demonstrated
that embryonic morphogens like WNTs, FGFs and TGFb can
be re-used as anterograde or retrograde factors like neurotro-
phins for neuronal outgrowth and juxtaposition of synaptic com-
ponents in the formation of mature synaptic connections
(Marshak et al., 2007; Mandolesi et al., 2005; Packard et al.,
2003; Salinas, 2005). Thus, the conventional strategy to search
for ‘plasticity genes’ from cortical cDNA libraries (Majdan and
Shatz, 2006; Tropea et al., 2006) may need to be expanded to
include early embryonic factors that originate sub-cortically.
It is generally thought that the number of interneurons and their
diversification has increased with evolution to regulate cortical
function. Cortical interneurons originate within the subcortical
telencephalon (for PV-cells the medial ganglionic eminence)
and then migrate tangentially into the overlying cortex but never
express Otx2 (Butt et al., 2007; Wonders and Anderson, 2006).
Otx2 signals anterior ‘‘head’’ position in all metazoans, starting
with the hydra. Like many homeoproteins (Brunet et al., 2007),
its role has preceded that of classical signaling molecules and
mediators of Otx2 expression in the cortex (e.g., activity–depen-
dent BDNF is absent even in arthropods).
Homeoproteins may act both as transcription factors and reg-
ulators of local protein synthesis (Dubnau and Struhl, 1996;
Nedelec et al., 2004; Brunet et al., 2005), perhaps through direct
interaction with eukaryotic translation initiation factor 4E (eIF4E)
(Nedelec et al., 2004; Topisirovic and Borden, 2005). Once inter-
nalized by PV cells, Otx2 has direct access to the cytoplasm,
where it can engage the protein translation apparatus as shown
for Engrailed in migrating growth cones (Brunet et al., 2005).
A dual action at either the transcriptional or translational level
could accelerate PV-cell maturation and critical period onset
simply by enhancing their known sensitivity to ambient neurotro-
phins such as BDNF (e.g., by upregulation of TrkB receptor)
(Huang et al., 1999; Marty et al., 1997).
Interestingly, PNN structures consisting of cell-adhesion mol-
ecules, hyaluronic acids and chondroitin sulfate proteoglycans
(CSPGs) tightly enwrap mature PV-cells specifically (Celio
et al., 1998). Our gain-of-function and loss-of-function results
show that Otx2 accumulation can induce the maturation of
PNNs. Several putative Otx2 binding sites are found in the
promoter region for Tenascin (Briata et al., 1999), a major extra-
cellular matrix component which may directly regulate PNN
development (Nguyen Ba-Charvet et al., 1998). Indeed, Tenas-
cin-R KO mice exhibit severe defects in PNN formation on
PV-cells as well as weakened GABA function (Bruckner et al.,
2000; Nikonenko et al., 2003; Saghatelyan et al., 2000).
The emergence of Otx2 within PV-cells may further serve as
a positive feedback loop: promoting PNN maturation that conse-
quently promotes Otx2 uptake and the strengthening of somatic
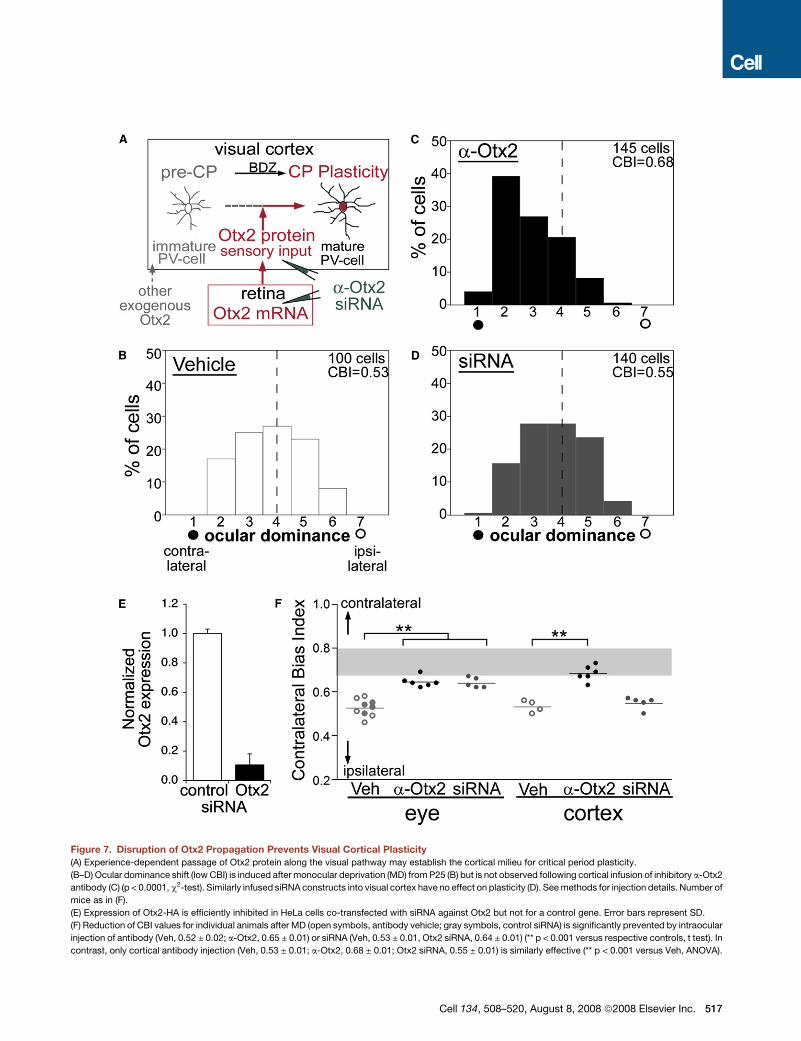
Figure 7. Disruption of Otx2 Propagation Prevents Visual Cortical Plasticity
(A) Experience-dependent passage of Otx2 protein along the visual pathway may establish the cortical milieu for critical period plasticity.
(B–D) Ocular dominance shift (low CBI) is induced after monocular deprivation (MD) from P25 (B) but is not observed following cortical infusion of inhibitory a-Otx2
antibody (C) (p < 0.0001, c2-test). Similarly infused siRNA constructs into visual cortex have no effect on plasticity (D). See methods for injection details. Number of
mice as in (F).
(E) Expression of Otx2-HA is efficiently inhibited in HeLa cells co-transfected with siRNA against Otx2 but not for a control gene. Error bars represent SD.
(F) Reduction of CBI values for individual animals after MD (open symbols, antibody vehicle; gray symbols, control siRNA) is significantly prevented by intraocular
injection of antibody (Veh, 0.52 ± 0.02; a-Otx2, 0.65 ± 0.01) or siRNA (Veh, 0.53 ± 0.01, Otx2 siRNA, 0.64 ± 0.01) (** p < 0.001 versus respective controls, t test). In
contrast, only cortical antibody injection (Veh, 0.53 ± 0.01; a-Otx2, 0.68 ± 0.01; Otx2 siRNA, 0.55 ± 0.01) is similarly effective (** p < 0.001 versus Veh, ANOVA).
Cell 134, 508–520, August 8, 2008 ª2008 Elsevier Inc. 517

inhibition at critical period onset. Negatively charged CSPG
sugars in the PNN bind plant lectins such as WFA with high
affinity and may do the same with positively-charged, homeodo-
main-containing proteins to facilitate internalization. The endog-
enous preference of Otx2 for PV-cells was unexpected from
in vitro studies (Joliot and Prochiantz, 2004). We found that other
infused homeoproteins (Engrailed-2) fail to accumulate in corti-
cal PV-cells (data not shown). Specific sugar epitopes may be
recognized by distinct homeoproteins according to a putative
‘sugar code’ for future study (Holt and Dickson, 2005).
Importantly, the removal of PNNs with chondroitinase injection
reactivates critical period plasticity in adulthood (Pizzorusso
et al., 2002). We found that similar treatment depleted adult
PV-cells of their Otx2 content, resetting them to an immature
state (M. Beurdeley and S.S., data not shown; Saghatelyan
et al., 2000). Instead, high polysialic acid expression during the
pre-critical period would nonspecifically trap Otx2 (Joliot et al.,
1991), leading to a reduction of homeoprotein internalization by
PV-cells, delaying their maturation and visual cortical plasticity
(Di Cristo et al., 2007). Plasticity onset may therefore reflect the
emergence of PV-cell function from a weakened state rather
than as a consequence of PNN removal per se. A dynamic, bidi-
rectional plasticity of afferent input onto PV-cells appears to
underlie critical period plasticity then becomes fixed with age
(Y. Yazaki-Sugiyama and T.K.H., unpublished data). It will be in-
teresting to map the detailed time-course of Otx2 accumulation
and critical period reactivation specifically in PV-cells when
PNNs are dissolved by chondroitinase treatment in the adult.
Persistent passage of Otx2 may further explain two visual phe-
nomena across the life spectrum. Dark-rearing from birth classi-
cally delays critical period onset (Cynader and Mitchell, 1980;
Fagiolini et al., 1994, 2003) by a specific reduction of somatic in-
hibition (Iwai et al., 2003; Katagiri et al., 2007). Here, we find that
Otx2 transport or accumulation into PV-cells is also experience-
dependent and appears only gradually in visual cortex after eye
opening with the onset of natural vision. Critical period time-
course may thus shift with impoverished or enriched environ-
ments (Cancedda et al., 2004) perhaps by regulating delivery
of morphogenic factors from the periphery.
Conversely, intracortical inhibition deteriorates upon aging,
degrading vision but paradoxically improving motion detection
(Leventhal et al., 2003; Betts et al., 2005). Failing eyesight with
senescence could reduce the visually-driven, sustained passage
of Otx2 into cortical interneurons leading to these effects.
Indeed, the dosage effect of Otx2 (one allele is not enough to
activate plasticity) confirms that variations in activity, paralleled
by variation in Otx2 transport, may have graded effects on the re-
finement of cortical architecture. The distribution of factors could
be variable among individual animals exposed to distinct sen-
sory experiences.
Modality-specific transport of a molecular messenger by ac-
tivity from the periphery into the respective cortical area may
be a generally safe way to signal when the sensory epithelium
is fully functional (e.g., whisking before eye opening) prior to en-
gaging local cortical plastic machinery. This would give rise to
differential critical period timecourse across brain regions and
may contribute to functional decline with aging when the sensory
periphery begins to deteriorate. Access to the cortical milieu for
518 Cell 134, 508–520, August 8, 2008 ª2008 Elsevier Inc.
plasticity from the periphery may in turn offer a powerful thera-
peutic tool for recovery of function. It will be of great interest to
confirm whether postnatal plasticity in each sensory modality re-
lies on a unique homeoprotein.
EXPERIMENTAL PROCEDURES
Animals
Conventionally raised C57Bl/6 mice (12:12 hours light:dark cycle) were pur-
chased from Japan SLC. Otx2flox/flox mice (Tian et al., 2002) were crossed
with aCaMKII-Cre transgenic mice (Minichiello et al., 1999) to generate an
Otx2 conditional knockout mouse line. GFP knock-in mice were kindly pro-
vided by Drs. D. Acampora and A. Simeone (MRC, King’s College London,
UK).
cDNA Amplification
Mouse Otx2 and GABAA receptor a2 subunit cDNA was amplified by one-step
RT-PCR (Invitrogen) with degenerate oligonucleotides (see Supplemental
Data). Each cDNA was inserted into pBluescript to confirm the sequence
and to generate the RNA probe.
In Situ Hybridization and Immunohistochemistry
In situ hybridization on floating sections was performed according to methods
described previously (Sugiyama and Nakamura, 2003). Mice were perfused
and post-fixed with 4% paraformaldehyde (PFA). Coronal sections (20 mm)
were hybridized at 65�C with a Digoxigenin (DIG)-labeled RNA probe for
Otx2, then detected by alkaline phosphate (AP)-conjugated anti-DIG antibody.
Primary antibodies (see Supplemental Data) or Biotin-labeled lectin from
Wisteria Floribunda agglutinin (WFA, Sigma) were incubated with floating cor-
onal sections (20 mm) at 4�C over night. Specificity of anti-Otx2 antibodies was
tested by preincubation with human Otx2 protein (2 mg/ml, produced in bacu-
lovirus) for 30 min before immunoreaction with tissues.
Quantification
For quantitative analysis of fluorescence intensity, immunostaining was per-
formed concurrently with the same solutions, and images were photographed
in one sitting with the same gain and exposure time.
The number of cells or intensity of immunofluorescence was measured in
a 600 3 350 mm area covering the supragranular layers of the binocular
zone. The intensity per cell was plotted on a cumulative histogram for compar-
ison across groups (Kolmogorov-Smirnov test). Averaged intensity per hemi-
sphere or for each mouse was compared pair-wise across the same brain or
unpaired between two groups of animals, respectively (t test).
The intensity of Otx2 immunofluorescence in retina was measured per pixel
across a 200 3 20 mm area, covering the inner nuclear layer. Averaged intensity
per mouse was compared between the two groups (t test).
Drug Infusion
Otx2 protein, inhibitory antibody against Otx2 (a-Otx2), vehicle solution or pen-
etratin-coupled siRNA against Otx2 or control (Muratovska and Eccles, 2004)
was infused into the right visual cortex. Cortical infusion was performed by os-
motic minipumps (Alzet1007D, Alza) connected to cannulae stereotaxically
implanted in mice at ages indicated in the text (Hensch et al., 1998). Diazepam
(DZ) or vehicle solution was injected daily into both lateral ventricles, starting
one day before monocular deprivation until the day before recording (Hensch
et al., 1998).
Biotinylated human Otx2 protein or bovine serum albumin (BSA), a-Otx2, ve-
hicle solution or penetratin-coupled siRNA against Otx2 or control was
injected into both eyes by micropipette (at P21 and P23 for proteins; at P16,
P19 and P22 for a-Otx2; at P18, P19, P21, and P22 for siRNA).
Single-Unit Electrophysiology In Vivo
Electrophysiological recordings were performed using standard techniques
(Hensch et al., 1998). Five to seven single units were recorded for each pene-
tration (>3 spaced at 200 mm intervals) across the medio-lateral axis of the bin-
ocular zone to avoid sampling bias. Multiple receptive field properties

(prolonged discharge, orientation bias, response strength, habituation and ret-
inotopy) were evaluated for each mouse (see Supplemental Data; Gordon and
Stryker, 1996; Hensch et al., 1998; Taha et al., 2002). Responses from single
cells were assigned ocular dominance scores using a seven-point classifica-
tion scheme (Wiesel and Hubel, 1963; Hensch et al., 1998). Ocular dominance
in the binocular zone from each mouse was calculated as a contralateral bias
index (CBI, see Supplemental Data). Visual responses after brief (3–5 days)
monocular deprivation (MD) were recorded from the hemisphere contralateral
to the deprived eye.
SUPPLEMENTAL DATA
Supplemental Data include Supplemental Experimental Procedures, Sup-
plemental References, four figures, and one table and can be found with this
article online at http://www.cell.com/cgi/content/full/134/3/508/DC1/.
ACKNOWLEDGMENTS
We thank D. Acampora and A. Simeone for graciously providing the Otx2-GFP
mouse line and the original Otx2 plasmids; A. Ruiz i Altaba and M. Wassef for
Otx2 antibody; S. Dupas for recombinant Otx2 protein; G. Corte for recombi-
nant baculovirus; K. Moya, M. Beurdeley, and A. Scheuber for help in some ex-
periments and discussion. Funded by the Human Frontiers Science Program
(HFSP, RGP0026/2002), Fondation pour La Recherche Medicale (Longevite
Cognitive et Sensorielle), and in part by the Special Postdoctoral Research
Program (RIKEN) and Japanese Ministry of Science, Education and Technol-
ogy (MEXT, IBR 17021047). S.S. is a Research Fellow of the Japan Society
for the Promotion of Science (JSPS). A.A.D. is a fellow of the Canadian Insti-
tutes of Health Research (CIHR).
Received: June 22, 2007
Revised: April 22, 2008
Accepted: May 5, 2008
Published: August 7, 2008
REFERENCES
Acampora, D., Gulisano, M., Broccoli, V., and Simeone, A. (2001). Otx genes in
brain morphogenesis. Prog. Neurobiol. 64, 69–95.
Antonini, A., Fagiolini, M., and Stryker, M.P. (1999). Anatomical correlates of
functional plasticity in mouse visual cortex. J. Neurosci. 19, 4388–4406.
Baas, D., Bumstead, K.M., Martinez, J.A., Vaccarino, F.M., Wikler, K.C., and
Barnstable, C.J. (2000). The subcellular localization of Otx2 is cell-type specific
and developmentally regulated in the mouse retina. Brain Res. Mol. Brain Res.
78, 26–37.
Betts, L.R., Taylor, C.P., Sekuler, A.B., and Bennett, P.J. (2005). Aging reduces
center-surround antagonism in visual motion processing. Neuron 45, 361–366.
Bovolenta, P., Mallamaci, A., Briata, P., Corte, G., and Boncinelli, E. (1997). Im-
plication of OTX2 in pigment epithelium determination and neural retina differ-
entiation. J. Neurosci. 17, 4243–4252.
Briata, P., Ilengo, C., Bobola, N., and Corte, G. (1999). Binding properties of the
human homeodomain protein OTX2 to a DNA target sequence. FEBS Lett.
445, 160–164.
Bruckner, G., Grosche, J., Schmidt, S., Hartig, W., Margolis, R.U., Delpech, B.,
Seidenbecher, C.I., Czaniera, R., and Schachner, M. (2000). Postnatal devel-
opment of perineuronal nets in wild-type mice and in a mutant deficient in te-
nascin-R. J. Comp. Neurol. 428, 616–629.
Brunet, I., Weinl, C., Piper, M., Trembleau, A., Volovitch, M., Harris, W., Pro-
chiantz, A., and Holt, C. (2005). The transcription factor Engrailed-2 guides ret-
inal axons. Nature 438, 94–98.
Brunet, I., Di Nardo, A., Sonnier, L., Beurdeley, M., and Prochiantz, A. (2007).
Shaping neural pathways with messenger homeoproteins. Trends Neurosci.
30, 260–267.
Brunet, J.F., and Pattyn, A. (2002). Phox2 genes - from patterning to connec-
tivity. Curr. Opin. Genet. Dev. 12, 435–440.
Butt, S.J., Cobos, I., Golden, J., Kessaris, N., Pachnis, V., and Anderson, S.
(2007). Transcriptional regulation of cortical interneuron development. J. Neu-
rosci. 27, 11847–11850.
Buzas, P., Eysel, U.T., Adorjan, P., and Kisvarday, Z.F. (2001). Axonal topog-
raphy of cortical basket cells in relation to orientation, direction, and ocular
dominance maps. J. Comp. Neurol. 437, 259–285.
Cancedda, L., Putignano, E., Sale, A., Viegi, A., Berardi, N., and Maffei, L.
(2004). Acceleration of visual system development by environmental enrich-
ment. J. Neurosci. 24, 4840–4848.
Celio, M.R., Spreafico, R., De Biasi, S., and Vitellaro-Zuccarello, L. (1998).
Perineuronal nets: past and present. Trends Neurosci. 21, 510–515.
Cruikshank, S.J., Lewis, T.J., and Connors, B.W. (2007). Synaptic basis for in-
tense thalamocortical activation of feedforward inhibitory cells in neocortex.
Nat. Neurosci. 10, 462–468.
Cynader, M., and Mitchell, D.E. (1980). Prolonged sensitivity to monocular
deprivation in dark-reared cats. J. Neurophysiol. 43, 1026–1040.
Daw, N. (1995). Visual Development (New York: Plenum).
del Rio, J.A., de Lecea, L., Ferrer, I., and Soriano, E. (1994). The development
of parvalbumin-immunoreactivity in the neocortex of the mouse. Brain Res.
Dev. Brain Res. 81, 247–259.
Di Cristo, G., Chattopadhyaya, B., Kuhlman, S.J., Fu, Y., Belanger, M.C., Wu,
C.Z., Rutishauser, U., Maffei, L., and Huang, Z.J. (2007). Activity-dependent
PSA expression regulates inhibitory maturation and onset of critical period
plasticity. Nat. Neurosci. 10, 1569–1577.
Dubnau, J., and Struhl, G. (1996). RNA recognition and translational regulation
by a homeo-domain protein. Nature 379, 694–699.
Erisir, A., and Dreusicke, M. (2005). Quantitative morphology and postsynaptic
targets of thalamocortical axons in critical period and adult ferret visual cortex.
J. Comp. Neurol. 485, 11–31.
Fagiolini, M., Pizzorusso, T., Berardi, N., Domenici, L., and Maffei, L. (1994).
Functional postnatal development of the rat primary visual cortex and the
role of visual experience: dark rearing and monocular deprivation. Vision
Res. 34, 709–720.
Fagiolini, M., and Hensch, T.K. (2000). Inhibitory threshold for critical-period
activation in primary visual cortex. Nature 404, 183–186.
Fagiolini, M., Katagiri, H., Miyamoto, H., Mori, H., Grant, S.G., Mishina, M., and
Hensch, T.K. (2003). Separable features of visual cortical plasticity revealed by
N-methyl-D-aspartate receptor 2A signaling. Proc. Natl. Acad. Sci. USA 100,
2854–2859.
Fagiolini, M., Fritschy, J.M., Low, K., Mohler, H., Rudolph, U., and Hensch, T.K.
(2004). Specific GABAA circuits for visual cortical plasticity. Science 303,
1681–1683.
Frantz, G.D., Weimann, J.M., Levin, M.E., and McConnell, S.K. (1994). Otx1
and Otx2 define layers and regions in developing cerebral cortex and cerebel-
lum. J. Neurosci. 14, 5725–5740.
Gao, B., and Fritschy, J.M. (1994). Selective allocation of GABAA receptors
containing the alpha 1 subunit to neurochemically distinct subpopulations of
rat hippocampal interneurons. Eur. J. Neurosci. 6, 837–853.
Gordon, J.A., and Stryker, M.P. (1996). Experience-dependent plasticity of
binocular responses in the primary visual cortex of the mouse. J. Neurosci.
16, 3274–3286.
Hanover, J.L., Huang, Z.J., Tonegawa, S., and Stryker, M.P. (1999). Brain-
derived neurotrophic factor overexpression induces precocious critical period
in mouse visual cortex. J. Neurosci. 19, RC40.
Hensch, T.K., Fagiolini, M., Mataga, N., Stryker, M.P., Baekkeskov, S., and
Kash, S.F. (1998). Local GABA circuit control of experience-dependent plas-
ticity in developing visual cortex. Science 282, 1504–1508.
Hensch, T.K., and Stryker, M.P. (2004). Columnar architecture sculpted by
GABA circuits in developing cat visual cortex. Science 303, 1678–1681.
Hensch, T.K. (2005). Critical period plasticity in local cortical circuits. Nat. Rev.
Neurosci. 6, 877–888.
Cell 134, 508–520, August 8, 2008 ª2008 Elsevier Inc. 519

Holt, C.E., and Dickson, B.J. (2005). Sugar codes for axons? Neuron 46, 169–
172.
Huang, Z.J., Kirkwood, A., Pizzorusso, T., Porciatti, V., Morales, B., Bear, M.F.,
Maffei, L., and Tonegawa, S. (1999). BDNF regulates the maturation of inhibi-
tion and the critical period of plasticity in mouse visual cortex. Cell 98, 739–
755.
Iwai, Y., Fagiolini, M., Obata, K., and Hensch, T.K. (2003). Rapid critical period
induction by tonic inhibition in mouse visual cortex. J. Neurosci. 23, 6695–
6702.
Joliot, A.H., Triller, A., Volovitch, M., Pernelle, C., and Prochiantz, A. (1991).
alpha- 2,8-polysialic acid is the neuronal surface receptor of antennapedia
homeobox peptide. New Biol. 3, 1121–1134.
Joliot, A., Trembleau, A., Raposo, G., Calvet, S., Volovitch, M., and Prochiantz,
A. (1997). Association of Engrailed homeoproteins with vesicles presenting
caveolae-like properties. Development 124, 1865–1875.
Joliot, A., and Prochiantz, A. (2004). Transduction peptides: from technology
to physiology. Nat. Cell Biol. 6, 189–196.
Katagiri, H., Fagiolini, M., and Hensch, T.K. (2007). Optimization of somatic in-
hibition at critical period onset in mouse visual cortex. Neuron 53, 805–812.
Klausberger, T., Roberts, J.D., and Somogyi, P. (2002). Cell type- and input-
specific differences in the number and subtypes of synaptic GABA(A) recep-
tors in the hippocampus. J. Neurosci. 22, 2513–2521.
Leventhal, A.G., Wang, Y., Pu, M., Zhou, Y., and Ma, Y. (2003). GABA and its
agonists improved visual cortical function in senescent monkeys. Science 300,
812–815.
Liu, L.O., Li, G., McCall, M.A., and Cooper, N.G.F. (2000). Photoreceptor reg-
ulated expression of Ca2+/Calmodulin-dependent protein kinase II in the
mouse retina. Brain Res. Mol. Brain Res. 82, 150–166.
Liu, X.B., and Jones, E.G. (1996). Localization of alpha type II calcium calmod-
ulin-dependent protein kinase at glutamatergic but not gamma-aminobutyric
acid (GABAergic) synapses in thalamus and cerebral cortex. Proc. Natl.
Acad. Sci. USA 93, 7332–7336.
Majdan, M., and Shatz, C.J. (2006). Effects of visual experience on activity-de-
pendent gene regulation in cortex. Nat. Neurosci. 9, 650–659.
Mandolesi, G., Menna, E., Harauzov, A., von Bartheld, C.S., Caleo, M., and
Maffei, L. (2005). A role for retinal brain-derived neurotrophic factor in ocular
dominance plasticity. Curr. Biol. 15, 2119–2124.
Marshak, S., Nikolakopoulou, A.M., Dirks, R., Martens, G.J., and Cohen-Cory,
S. (2007). Cell-autonomous TrkB signaling in presynaptic retinal ganglion cells
mediates axon arbor growth and synapse maturation during the establishment
of retinotectal synaptic connectivity. J. Neurosci. 27, 2444–2456.
Marty, S., da Penha Berzaghi, M., and Berninger, B. (1997). Neurotrophins and
activity-dependent plasticity of cortical interneurons. Trends Neurosci. 20,
198–202.
Mataga, N., Mizuguchi, Y., and Hensch, T.K. (2004). Experience-dependent
pruning of dendritic spines in visual cortex by tissue plasminogen activator.
Neuron 44, 1031–1041.
Minichiello, L., Korte, M., Wolfer, D., Kuhn, R., Unsicker, K., Cestari, V., Rossi-
Arnaud, C., Lipp, H.P., Bonhoeffer, T., and Klein, R. (1999). Essential role for
TrkB receptors in hippocampus-mediated learning. Neuron 24, 401–414.
Morales, B., Choi, S.Y., and Kirkwood, A. (2002). Dark rearing alters the devel-
opment of GABAergic transmission in visual cortex. J. Neurosci. 22, 8084–
8090.
Morishita, H., and Hensch, T.K. (2008). Critical period revisited: impact on vi-
sion. Curr. Opin. Neurobiol. 18, 101–107.
Mower, G.D. (1991). The effect of dark rearing on the time course of the critical
period in cat visual cortex. Dev Brain Res. 58, 151–158.
Muratovska, A., and Eccles, M.R. (2004). Conjugate for efficient delivery of
short interfering RNA (siRNA) into mammalian cells. FEBS Lett. 558, 63–68.
Nedelec, S., Foucher, I., Brunet, I., Bouillot, C., Prochiantz, A., and Trembleau,
A. (2004). Emx2 homeodomain transcription factor interacts with eukaryotic
520 Cell 134, 508–520, August 8, 2008 ª2008 Elsevier Inc.
translation initiation factor 4E (eIF4E) in the axons of olfactory sensory neurons.
Proc. Natl. Acad. Sci. USA 101, 10815–10820.
Nguyen Ba-Charvet, K.T., von Boxberg, Y., Guazzi, S., Boncinelli, E., and God-
ement, P. (1998). A potential role for the OTX2 homeoprotein in creating early
‘highways’ for axon extension in the rostral brain. Development 125,
4273–4282.
Nikonenko, A., Schmidt, S., Skibo, G., Bruckner, G., and Schachner, M. (2003).
Tenascin-R-deficient mice show structural alterations of symmetric periso-
matic synapses in the CA1 region of the hippocampus. J. Comp. Neurol.
456, 338–349.
Nothias, F., Fishell, G., and Ruiz i Altaba, A. (1998). Cooperation of intrinsic and
extrinsic signals in the elaboration of regional identity in the posterior cerebral
cortex. Curr. Biol. 8, 459–462.
Packard, M., Mathew, D., and Budnik, V. (2003). Wnts and TGF beta in synap-
togenesis: old friends signalling at new places. Nat. Rev. Neurosci. 4, 113–120.
Pizzorusso, T., Medini, P., Berardi, N., Chierzi, S., Fawcett, J.W., and Maffei, L.
(2002). Reactivation of ocular dominance plasticity in the adult visual cortex.
Science 298, 1248–1251.
Prusky, G.T., and Douglas, R.M. (2003). Developmental plasticity of mouse vi-
sual acuity. Eur. J. Neurosci. 17, 167–173.
Rath, M.F., Morin, F., Shi, Q., Klein, D.C., and Møller, M. (2007). Ontogenetic
expression of the Otx2 and Crx homeobox genes in the retina of the rat.
Exp. Eye Res. 85, 65–73.
Rudy, B., and McBain, C.J. (2001). Kv3 channels: voltage-gated K+ channels
designed for high-frequency repetitive firing. Trends Neurosci. 24, 517–526.
Saghatelyan, A.K., Gorissen, S., Albert, M., Hertlein, B., Schachner, M., and
Dityatev, A. (2000). The extracellular matrix molecule tenascin-R and its
HNK-1 carbohydrate modulate perisomatic inhibition and long-term potentia-
tion in the CA1 region of the hippocampus. Eur. J. Neurosci. 12, 3331–3342.
Salinas, P.C. (2005). Signaling at the vertebrate synapse: new roles for embry-
onic morphogens? J. Neurobiol. 64, 435–445.
Sik, A., Hajos, N., Gulacsi, A., Mody, I., and Freund, T.F. (1998). The absence of
a major Ca2+ signaling pathway in GABAergic neurons of the hippocampus.
Proc. Natl. Acad. Sci. USA 95, 3245–3250.
Soltesz, I., Smetters, D.K., and Mody, I. (1995). Tonic inhibition originates from
synapses close to the soma. Neuron 14, 1273–1283.
Sugino, K., Hempel, C.M., Miller, M.N., Hattox, A.M., Shapino, P., Wu, C.,
Huang, Z.J., and Nelson, S.B. (2006). Molecular taxonomy of major neuronal
classes in the adult mouse forebrain. Nat. Neurosci. 9, 99–107.
Sugiyama, S., and Nakamura, H. (2003). The role of Grg4 in tectal laminar for-
mation. Development 130, 451–462.
Taha, S., Hanover, J.L., Silva, A.J., and Stryker, M.P. (2002). Autophosphory-
lation of alphaCaMKII is required for ocular dominance plasticity. Neuron 36,
483–491.
Tian, E., Kimura, C., Takeda, N., Aizawa, S., and Matsuo, I. (2002). Otx2 is
required to respond to signals from anterior neural ridge for forebrain specifi-
cation. Dev. Biol. 242, 204–223.
Topisirovic, I., and Borden, K.L. (2005). Homeodomain proteins and eukaryotic
translation initiation factor 4E (eIF4E): an unexpected relationship. Histol.
Histopathol. 20, 1275–1284.
Tohmi, M., Kitaura, H., Komagata, S., Kudoh, M., and Shibuki, K. (2006).
Enduring critical period plasticity visualized by transcranial flavoprotein
imaging in mouse primary visual cortex. J. Neurosci. 26, 11775–11785.
Tropea, D., Kreiman, G., Lyckman, A., Mukherjee, S., Yu, H., Horng, S., and
Sur, M. (2006). Gene expression changes and molecular pathways mediating
activity-dependent plasticity in visual cortex. Nat. Neurosci. 9, 660–668.
Wiesel, T.N., and Hubel, D.H. (1963). Single-cell responses in striate cortex of
kittens deprived of vision in one eye. J. Neurophysiol. 26, 1003–1017.
Wonders, C.P., and Anderson, S.A. (2006). The origin and specification of
cortical interneurons. Nat. Rev. Neurosci. 7, 687–696.




















