
Arsenic stress activates MAP kinase in rice roots and leaves
10
Arsenic stress activates MAP kinase in rice roots and leaves Kudupudi Prabhakara Rao a,1 , Gubbala Vani a,b,1 , Kundan Kumar a,2 , Dhammaprakash Pandhari Wankhede a , Mohit Misra a , Meetu Gupta b,⇑ , Alok Krishna Sinha a,⇑ a National Institute of Plant Genome Research, Aruna Asaf Ali Marg, New Delhi 110067, India b Department of Biotechnology, Jamia Millia Islamia, Central University, New Delhi 110025, India article info Article history: Received 26 August 2010 and in revised form 1 November 2010 Available online 21 November 2010 Keywords: Arsenic Mitogen activate protein kinase (MAPK) Nitric oxide Reactive oxygen species Rice abstract The toxic metalloid arsenite has become a potential threat to rice growing regions leading to serious con- tamination in food chain. In the present study effect of different physiological concentration of arsenite that is toxic and triggers the molecular events were evaluated in rice seedlings. Along with severe effect on the growth of rice seedling, production of reactive oxygen species (ROS) and nitric oxide (NO) in arse- nite treated rice roots was also observed. Activation of a 42 kDa mitogen activated protein kinase (MAPK/ MPK) by arsenite was observed in rice leaves and 42 and 44 kDa in roots in dose dependent manner. The activated MAPK could be immunoprecipitated with anti-phospho-tyrosine antibody, 4G10. The kinetic of MAPK activation by arsenite was found to be dose dependent. Transcript analysis of MAPK family and immunokinase assay in arsenite treated rice seedling revealed significant level of induction in OsMPK3 transcripts in leaves and OsMPK3, OsMPK4 transcripts in roots. Among MAPK kinase (MKKs) gene family, OsMKK4 transcripts were found to be induced in arsenite treated rice leaves and roots. In-silico homology modeling and docking analysis supported OsMPK3–OsMKK4 interaction. The data indicates that arsenite stress is transduced through MAPK signaling cascade in rice. Ó 2010 Elsevier Inc. All rights reserved. Introduction Heavy metal toxicity is one of the major environmental health problems in modern society, with potentially dangerous bioaccu- mulation through the food chain. Rapid industrialization and urbanization have enhanced the levels of toxic heavy metals in the environment, posing a potential health hazard for all living organisms [1,2]. When accumulated in excess in plant tissues these metals can function as stressors causing physiological constraints and alterations in various vital growth processes [3]. The metalloid arsenic is a toxic pollutant ubiquitous in the environment. It is present both as arsenite (AsIII) and arsenate (AsV) in the environ- ment, with arsenate being more prevalent than arsenite in soils [4,5]. In response to toxic levels of heavy metals, plants synthesize Cys-rich, metal-binding peptides including phytochelatins and metallothioneins. Heavy metals are detoxified by chelation and sequestration by Cys rich metal-binding peptides in the vacuole [6,7] while, various membrane transport systems play an important role in metal ion homeostasis and tolerance [8]. Gene expression patterns change when plants encounter exces- sive amounts of heavy metals [9]. Heavy metals are also reported to activate MAPKs in higher plants [10,11]. Exposure of Medicago seedlings to excess copper or cadmium ions resulted in a complex activation pattern of four distinct MAPKs: SIMK, MMK2, MMK3 and SAMK [10] Agrawal et al. [12,13] reported that the amount of OsMSRMK2, OsMSRMK3 and OsWJUMK1 transcripts increased in re- sponse to Cu and Cd treatments in rice seedlings. It was reported that OsMAPK2 transcripts increased within 12 h upon Cu treatment in rice suspension cells [14]. Further, activation of MAPK 3 by differ- ent heavy metals like Cd and Cu [11,15] and Zn [16] mostly in the range of 40 and 42 kDa was reported from rice. Tsai and Huang [17] showed that iron at higher concentrations activated a 42 kDa MAPK. Using different inhibitors it was demonstrated that MAPK, reactive oxygen species, protein phosphatase, and H + -ATPase might function in the plant iron-triggered signaling pathway in rice roots. Studies from mammalian system reveals that non-cytotoxic concen- trations of As 3+ ,V 1+ and Zn 2+ induced a rapid phosphorylation of MAPK in BEAS cells [18]. They suggested that metal-induced activa- tion of MAPK was due to inhibition of phosphatase activity. Simi- larly, trivalent As 3+ reportedly activates JNK and P38 in HeLa cells. In case of plants, arsenic (As) toxicity and its biochemical effects have been mostly evaluated in ferns and a few higher plants. In a 0003-9861/$ - see front matter Ó 2010 Elsevier Inc. All rights reserved. doi:10.1016/j.abb.2010.11.006 ⇑ Corresponding authors. Fax: +91 1126741658 (A.K. Sinha). E-mail addresses: [email protected] (M. Gupta), [email protected] (A.K. Sinha). 1 These authors have contributed equally to this work. 2 Present address: Department of Plant, Soil, and Insect Sciences, University of Massachusetts, Amherst, MA 01002, USA. 3 Abbreviations used: MAPK/MPK, mitogen activated protein kinase; MAPKK/MKK, MAPK kinase; ROS, reactive oxygen species; NO, nitric oxide. Archives of Biochemistry and Biophysics 506 (2011) 73–82 Contents lists available at ScienceDirect Archives of Biochemistry and Biophysics journal homepage: www.elsevier.com/locate/yabbi
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Arsenic stress activates MAP kinase in rice roots and leaves

Archives of Biochemistry and Biophysics 506 (2011) 73–82
Contents lists available at ScienceDirect
Archives of Biochemistry and Biophysics
journal homepage: www.elsevier .com/ locate/yabbi
Arsenic stress activates MAP kinase in rice roots and leaves
Kudupudi Prabhakara Rao a,1, Gubbala Vani a,b,1, Kundan Kumar a,2, Dhammaprakash Pandhari Wankhede a,Mohit Misra a, Meetu Gupta b,⇑, Alok Krishna Sinha a,⇑a National Institute of Plant Genome Research, Aruna Asaf Ali Marg, New Delhi 110067, Indiab Department of Biotechnology, Jamia Millia Islamia, Central University, New Delhi 110025, India
a r t i c l e i n f o
Article history:Received 26 August 2010and in revised form 1 November 2010Available online 21 November 2010
Keywords:ArsenicMitogen activate protein kinase (MAPK)Nitric oxideReactive oxygen speciesRice
0003-9861/$ - see front matter � 2010 Elsevier Inc. Adoi:10.1016/j.abb.2010.11.006
⇑ Corresponding authors. Fax: +91 1126741658 (A.E-mail addresses: [email protected] (M. Gu
(A.K. Sinha).1 These authors have contributed equally to this wo2 Present address: Department of Plant, Soil, and
Massachusetts, Amherst, MA 01002, USA.
a b s t r a c t
The toxic metalloid arsenite has become a potential threat to rice growing regions leading to serious con-tamination in food chain. In the present study effect of different physiological concentration of arsenitethat is toxic and triggers the molecular events were evaluated in rice seedlings. Along with severe effecton the growth of rice seedling, production of reactive oxygen species (ROS) and nitric oxide (NO) in arse-nite treated rice roots was also observed. Activation of a 42 kDa mitogen activated protein kinase (MAPK/MPK) by arsenite was observed in rice leaves and 42 and 44 kDa in roots in dose dependent manner. Theactivated MAPK could be immunoprecipitated with anti-phospho-tyrosine antibody, 4G10. The kinetic ofMAPK activation by arsenite was found to be dose dependent. Transcript analysis of MAPK family andimmunokinase assay in arsenite treated rice seedling revealed significant level of induction in OsMPK3transcripts in leaves and OsMPK3, OsMPK4 transcripts in roots. Among MAPK kinase (MKKs) gene family,OsMKK4 transcripts were found to be induced in arsenite treated rice leaves and roots. In-silico homologymodeling and docking analysis supported OsMPK3–OsMKK4 interaction. The data indicates that arsenitestress is transduced through MAPK signaling cascade in rice.
� 2010 Elsevier Inc. All rights reserved.
Introduction
Heavy metal toxicity is one of the major environmental healthproblems in modern society, with potentially dangerous bioaccu-mulation through the food chain. Rapid industrialization andurbanization have enhanced the levels of toxic heavy metals inthe environment, posing a potential health hazard for all livingorganisms [1,2]. When accumulated in excess in plant tissues thesemetals can function as stressors causing physiological constraintsand alterations in various vital growth processes [3]. The metalloidarsenic is a toxic pollutant ubiquitous in the environment. It ispresent both as arsenite (AsIII) and arsenate (AsV) in the environ-ment, with arsenate being more prevalent than arsenite in soils[4,5]. In response to toxic levels of heavy metals, plants synthesizeCys-rich, metal-binding peptides including phytochelatins andmetallothioneins. Heavy metals are detoxified by chelation andsequestration by Cys rich metal-binding peptides in the vacuole[6,7] while, various membrane transport systems play animportant role in metal ion homeostasis and tolerance [8].
ll rights reserved.
K. Sinha).pta), [email protected]
rk.Insect Sciences, University of
Gene expression patterns change when plants encounter exces-sive amounts of heavy metals [9]. Heavy metals are also reportedto activate MAPKs in higher plants [10,11]. Exposure of Medicagoseedlings to excess copper or cadmium ions resulted in a complexactivation pattern of four distinct MAPKs: SIMK, MMK2, MMK3 andSAMK [10] Agrawal et al. [12,13] reported that the amount ofOsMSRMK2, OsMSRMK3 and OsWJUMK1 transcripts increased in re-sponse to Cu and Cd treatments in rice seedlings. It was reportedthat OsMAPK2 transcripts increased within 12 h upon Cu treatmentin rice suspension cells [14]. Further, activation of MAPK3 by differ-ent heavy metals like Cd and Cu [11,15] and Zn [16] mostly in therange of 40 and 42 kDa was reported from rice. Tsai and Huang[17] showed that iron at higher concentrations activated a 42 kDaMAPK. Using different inhibitors it was demonstrated that MAPK,reactive oxygen species, protein phosphatase, and H+-ATPase mightfunction in the plant iron-triggered signaling pathway in rice roots.Studies from mammalian system reveals that non-cytotoxic concen-trations of As3+, V1+ and Zn2+ induced a rapid phosphorylation ofMAPK in BEAS cells [18]. They suggested that metal-induced activa-tion of MAPK was due to inhibition of phosphatase activity. Simi-larly, trivalent As3+ reportedly activates JNK and P38 in HeLa cells.In case of plants, arsenic (As) toxicity and its biochemical effectshave been mostly evaluated in ferns and a few higher plants. In a
3 Abbreviations used: MAPK/MPK, mitogen activated protein kinase; MAPKK/MKK,MAPK kinase; ROS, reactive oxygen species; NO, nitric oxide.

74 K.P. Rao et al. / Archives of Biochemistry and Biophysics 506 (2011) 73–82
genome wise comparative study, Chakrabarty et al. [19] has shownthat the different sets of genes are activated by arsenate an arsenitestress in rice. Arsenite has also shown to activate MAPK activity inBrassica juncea [20]. However a complete analysis of MAPK genefamily involved in arsenic stress signal transduction has not been re-ported so far.
In the present investigation we have studied the effect of arse-nite on the growth of rice seedling and its photosynthetic capabil-ity. To gain further insight into early events of arsenic stresssignaling, involvement of ROS, NO, activation of MAPK and its cor-relation with increasing arsenite concentration in leaf and rootswere also analyzed. Transcript analysis of all the members of MAPKand MKK gene family in rice under arsenic stress revealed thegenes involved in this heavy metal signaling.
Materials and methods
Plant growth and treatments
Rice (Oryza sativa L. indica cultivar group var Pusa Basmati-1)was grown in growth chamber (SCILAB instrument, Taiwan) hydro-ponically at 28 �C with 16 h light and 8 h dark period. For Arsenicstress, different concentrations of sodium meta-arsenite were pre-pared by dissolving in water and added to the 1/10th of the Hoa-gland solution in which plants were growing. The plant sampleswere harvested at different time points mentioned in the legendof respective figures by snap freezing in liquid N2 and stored in�80 �C for further analysis.
Measurement of chlorophyll fluorescence
Chlorophyll fluorescence was measured by using imaging PAMchlorophyll flurometer (Walz, Effeltrich, Germany). PAM stands forthe pulse amplitude modulation in which fluorescence is excitedby very brief, but strong light pulses from light emitting diodes.The leaf of the plant for which measurement has to be made waskept on the stage of instrument and initiated by default parameterof the instrument. Effective quantum yield of PSII (YII) was calcu-lated as: YII = DF/Fm0 = (Fm0–Ft)/Fm0, where Ft was the fluores-cence yield at any time (steady state) and Fm0 was the maximumyield reached in a pulse of saturating light during illumination.Fv/Fm ratio is calculated as Fv/Fm = (Fm–Fo)/Fm where ‘Fo’ and‘Fm’ are the minimal and maximal fluorescence yields of a darkadapted sample. All the measurements were done in a closedchamber under controlled growth conditions. At least four leaveswere selected from each plants and within each leaf three differentpositions were selected for data recording.
Evaluation of cell death using Evans blue assay
Two weeks old rice seedlings were treated with 50 lM sodiummeta-arsenite for 15 min. The roots were stained in 0.25% solutionof Evans blue for 15 min at room temperature, and then washedtwice for 15 min with distilled water to remove the excess stainand observed under microscope (Leica CM 1510S, Leica Microsys-tems GmBH).
Detection of reactive oxygen species (ROS) and nitric oxide (NO) in riceroots
Two weeks old rice seedlings were used to localize the genera-tion of ROS and NO in rice roots. For ROS determination segmentsof rice roots of approximately 20 mm2 from the apex were incu-bated for 1 h at 25 �C in dark with 25 lM 20,70-dichloro dihydrofluorescein diacetate, acetyl ester (CM-H2DCFDA; Sigma). While
for NO determination the segment of rice roots were incubatedwith 10 lM 4,5-diaminofluorescin diacetate (DAF-2 DA, Sigma) un-der similar condition as above. The rice root segments after incuba-tion were treated with 50 lM sodium meta-arsenite for 15 minand observed under fluorescence microscope. A Leica MPS60 fluo-rescent microscope equipped with a green fluorescent protein fil-ter (excitation 450–490 nm, emission 500–530 nm) was used forfluorescence images. Exposure times were equal for all samples.Autofluorescence was not observed in unstained controls.
Protein extraction and MAPK activity assay
Rice leaf or root tissue were ground in liquid nitrogen andhomogenized in extraction buffer containing 50 mM HEPES, pH7.5, 5 mM EDTA, 5 mM EGTA, 50 mM b-glycerol phosphate, 1 mMphenylmethylsulfonyl fluoride, 10 mM DTT, 10 mM Na3VO4,10 mM NaF, 10% glycerol and 0.74 g ml�1 Polyvinyl polypyrroli-done (PVPP). After centrifugation at 16,000g, aliquots of superna-tant were used either for in-gel kinase or in-solution assay orimmunokinase assay for MAPK. The protein concentration wasdetermined by Bradford method using BSA as standard. In-gel ki-nase assay was performed as described previously [21]. Briefly,50 lg of total protein was fractionated on a 12% polyacrylamidegel containing 0.1% SDS and 0.5 mg ml�1 bovine brain myelin basicprotein (MBP) (Sigma Aldrich). After electrophoresis, the SDS wasremoved by washing the gel three times (30 min each) at roomtemperature with buffer containing 25 mM Tris, pH 7.5, 0.5 mMDTT, 0.1 mM Na3VO4 and 5 mM NaF, 0.5 mg ml�1 BSA and 0.1% Tri-ton X-100. The kinases were allowed to renature overnight at 4 �Cwith three changes of renaturing buffer (25 mM Tris, pH 7.5,0.5 mM DTT, 5 mM NaF and 0.1 mM Na3VO4) 1.5 h, overnight, 2 hinterval. The phosphorylation of MBP was performed in 20 mL ofreaction buffer (25 mM Tris, pH 7.5, 2 mM EGTA, 12 mM MgCl2,1 mM DTT, 0.1 mM Na3VO4, 1 lM ATP and 50 lCi of c32P-ATP(3000 Ci mmol�1) at room temperature for 60 min. The gel wastransferred into washing buffer (5% trichloroacetic acid and 1% so-dium pyrophosphate) at room temperature for 15 min with 3–4changes of the washing buffer. Finally the gel was dried on filterpaper and autoradiographed in phosphorimager (Typhoon, GEhealth care).
For in-solution kinase determination, 5 lL of sample was mixedwith reaction buffer to give a final volume of 15 lL containing25 mm Tris/HCl, pH 7.5, 5 mm MgCl2, 1 mm EGTA, 1 mm DTT,0.5 mg mL�1 MBP, 25 lm ATP, and 1 lCi c32P-ATP. Incubation atroom temperature was stopped after 20 min by addition of 10 lLof 5X SDS sample buffer. The unincorporated c32P-ATP was sepa-rated from the MBP by SDS-gel electrophoresis. Activities werevisualized and quantified using a phosphor imager (Typhoon, GEhealth care).
Immunokinase assay was performed by immunoprecipitationof the activated MAPK followed by an in-gel kinase assay. For theimmunoprecipitation 100 lg of total protein in a crude extractwas brought to 150 mM NaCl and 0.1% (v/v) Nonidet P40. Afteraddition of 1 lg of the phospho-Tyr specific monoclonal antibody4G10 (Upstate biotechnology) or AtMPK3 or AtMPK4 antibodies(Sigma chemical, USA) the assay was shaken at 4 �C for 2 h andafter the subsequent addition of 15 lL of protein A-Sepharose(GE healthcare) incubated at 4 �C with shaking for another 4 h.The protein A-Sepharose with the bound antigen was pelleted at20,000g for 2 min and washed with 500 lL of 100 mM HEPES/KOH, pH 7.5, 5 mM EDTA, 5 mM EGTA, 10 mM DTT, 10 mMNa3VO4, 10 mM NaF, 50 mM b-glycerol phosphate, 10% (w/v) glyc-erol, 1 lg mL�l antipain, 0.1 mM phenyl methyl sulfonyl fluoride,1 mm benzamidine, 150 mm NaCl, and 1% (v/v) Nonidet P40. Thepellet was resuspended in 40 lL of SDS-loading buffer and incu-bated at 37 �C for 15 min and at 65 �C for another 15 min. After

K.P. Rao et al. / Archives of Biochemistry and Biophysics 506 (2011) 73–82 75
centrifugation, the supernatant was analyzed by an in-gel kinase orin-solution assay with MBP as substrate.
Quantitative real time PCR (qRT-PCR) analysis
Specific primer pairs were designed for all the members ofMAPK and MAPKK gene family (Supplementary Table 1) for qRT-PCR analysis. The specificities of the primers were confirmed byrunning the RT-PCR product in agarose gel, before they were usedfor qRT-PCR analysis. The amplicon product of each of them wasaround 200 bp. Each PCR reaction consisted of 10 ll of SYBR GreenPCR master mix (SYBR Green mix, Applied Biosystems), 0.9 lL ofeach primer (20 mM), 6.2 lL of water and 2 lL of each reversetranscribed cDNA product. Quantitative real time PCR was carriedout using step one real time PCR systems (Applied Biosystems) asmentioned earlier in Kumar et al. [22]. The thermocycler programhad an initial 95 �C for 10 min followed by 40 cycles consisting of15 s denaturation at 95 �C, 60 s annealing and extension at 60 �C.At the end of each reaction, a melting curve was created using asingle cycle consisting of 15 s denaturation at 95 �C and a 60 sannealing at 60 �C. This was followed by a slow temperature in-crease to 95 �C at the rate of 0.3 �C/s. The melting curve was usedto detect the presence of primer dimer or other unwanted ampli-fied products that would produce a detectable CT value and nega-tively skew the results.
In-silico homology modeling and protein–protein docking
For selection of templates for homology modeling of OsMKK4–OsMPK3 and OsMPK4, PSI BLAST was performed against the PDBdatabase (http://www.rcsb.org/pdb/welcome.do) [23]. The signifi-cant hits with >30% sequence identity were selected. Proteins usedas templates for homology modeling, along with PDB IDs and theiridentity with the target proteins are shown in Supplementary Ta-ble 2. We have used the Discovery studio 2.5.5.9350 (http://www.accelrys.com/dstdio) suit with align 123, which is a ClustalWhybrid for sequence alignment and modeler 9v8 [24] for thehomology model building. 3-D Models were refined with the helpof loop refinement (MODELER and looper algorithm based) andside chain refinement protocols. Evaluation of 3D models was doneby drawing Ramachandran plot and running verify protein (Profiles3D) protocol. Prepare protein protocol was finally run on proteinmodels. The protocol executes the following steps, some of whichcan be controlled by parameters: (i) clean the protein, (ii) optimizeside-chain conformation for residues with inserted atoms, (iii) re-move water molecules (optional), (iv) break bonds between metaland protein atoms (optional), (v) model missing loop regions basedon SEQRES information or by user-definition (optional).
For protein–protein docking, the Z DOCK and R DOCK programswere used. Z DOCK is rigid body protein docking algorithm thatexplicitly searches rotational space and uses a Fast Fourier Trans-formation (FFT) algorithm to significantly speed up searching intranslational space [25]. R DOCK program is an energy minimiza-tion algorithm [26] designed as refinement re-ranking tool for ZDOCK’s top predictions. In initial stage the protein receptor(OsMPK3, OsMPK4) and protein ligand (OsMKK4) were treated asrigid bodies and all rotational and translational degrees of freedomwere fully explored with scoring function that is tolerant to confor-mational changes. An angular step of 15� was used which results in2600 poses. In the refinement stage, R DOCK top poses of near na-tive structure obtained in the initial stage were refined and re-ranked. R DOCK minimization of the complexes generated by ZDOCK comprised small clashes removal to allow small conforma-tional changes, optimization of polar interactions and optimizationof charged interactions.
Results
Arsenite severely effect the growth of rice seedlings
In order to evaluate the toxicity of arsenite on the growth of riceseedlings, 3 day old rice seedlings were treated with 0.01, 0.05, 0.1,1, 10, 50 and 100 lM sodium meta-arsenite, respectively. Thelength of the shoot and roots were measured after 14 days of treat-ment. As shown in Fig. 1A, arsenite inhibited the growth of riceseedlings including shoot and root in a dose dependent manner.The growth of rice seedling was inhibited under 10 lM arsenitetreatment, and its effect was further increased on the length ofshoot and root when treated with 50 and 100 lM arsenite(Fig. 1B). Further, viability of the rice root cells was examined usingEvans blue staining assay. One-week-old rice seedlings were trea-ted with 50 lM arsenite for 1, 3, 9 and 24 h before staining withEvans blue dye. The cells with damaged membranes only, incorpo-rates the Evans blue dye. The results showed that the cell death in-creases with the duration of arsenite incubation (Fig. 1C),indicating that arsenite at higher concentration are toxic to cells.
Arsenite adversely effect photosynthetic efficiency in rice seedlings
By measuring the yield of chlorophyll fluorescence, informationabout changes in the efficiency of photochemistry and heat dissipa-tion can be gained [27]. To ascertain the effect of arsenite on the phys-iology of the rice seedlings, chlorophyll fluorescence was measured.The Fv/Fm value, which reflects the maximum quantum efficiencyof PSII was found to be around 0.74–0.77 in treated and controlsamples, respectively (Fig. 2). Overall photosynthetic performanceof plant was measured by measuring effective quantum yield (yieldII) of photochemical energy conversion in light adapted state. Theeffective yield of photosynthesis was reduced with increase of arse-nite concentration in rice seedlings (Fig. 2). The data indicated thatthough photochemistry of the plants remains unaffected by arsenitetreatment, the effective photosynthesis gets severely inhibited.
ROS and NO are generated in arsenite treated rice seedlings
Reactive oxygen species (ROS) are thought to display pleiotropiceffect in plant response. It has been realized that ROS act as defensemolecule against pathogen infection and act as intermediate sig-naling molecules in many abiotic stresses to regulate the expres-sion of genes. To determine whether arsenite treatment inducesROS production, a ROS-sensitive dye, 5-(and-6)-chloromethyl-20,70-dichloro dihydro fluorescein diacetate acetyl ester (CM-H2DCFDA), was used. This compound is non-fluorescent but is rap-idly oxidized to the highly fluorescent DCF by intracellular ROS.Rice roots were treated with 50 lM arsenite for 15 min and ob-served for DCF fluorescence. As shown in Fig. 3a–d the levels ofDCF fluorescence increased significantly after arsenite treatmentsin root tip regions. By contrast, relatively lower levels of DCF fluo-rescence were produced in the control roots. These results suggestthat arsenite induce ROS production in rice root tips.
Nitric oxide (NO) as a radical participates in plant innate immu-nity. The NO burst was detected by 4,5-diaminofluorescein diace-tate (DAF-2DA) mediated fluorescence. Production of nitric oxideconverts the non-fluorescent dye, DAF-2, to its fluorescent triolederivative, DAF-2T. In order to investigate the occurrence of NOburst with arsenite, rice roots were treated with 50 lM arsenitefor 15 min. As shown in Fig. 3e–h the levels of DAF-2T fluorescenceincreased significantly after arsenite treatments in root tip regions.By contrast, relatively lower levels of DAF-2T fluorescence wereproduced in the control roots which indicated the generation ofNO with arsenite treatment.

Fig. 1. Effect of arsenite on the growth of rice seedlings. (A) Three day old seedlings were treated with different concentrations of arsenite and allowed to grow for 14 days. (B)Shoot and root lengths of rice seedlings grown in different concentrations of arsenite for 14 days were measured and depicted as bar graph for comparison. Values arerepresented as mean and SD of three independent samples. (C) Cell death assay of arsenite treated rice roots by Evans staining. Two weeks old rice seedlings were treatedwith 50 lM arsenite for respective time points and the roots were stained in 0.25% solution of Evans blue for 15 min at room temperature. Excess stain was removed bywashing with distilled water and observed under microscope. All the experiments were repeated at least three times independently.
Fig. 2. Chlorophyll fluorescence of arsenite treated rice seedlings. Three days old riceseedlings were treated with different concentrations of arsenite for 7 days followedby measuring the maximum quantum yield (Fv/Fm) and effective quantum yield.Values are represented as mean and SD of three independent samples.
76 K.P. Rao et al. / Archives of Biochemistry and Biophysics 506 (2011) 73–82
Arsenite activates MAP kinase in rice roots and leaves
To determine whether arsenic treatment activates MAPK, riceseedlings were treated with 50 lM sodium meta-arsenite in twoseparate experiments. Samples were harvested at different timepoints from leaves and roots independently. The crude extracts
from arsenite treated leaf and root samples were separated on a10% SDS–PAGE embedded with myelin basic protein (MBP) as sub-strate and in-gel kinase assay was carried out. The results indicatedactivation of 44 kDa MBP phosphorylating kinase in leaf samples3 h after arsenite treatment and remained high up to 9 h(Fig. 4A). In case of roots activation of two MBP phosphorylating ki-nases with an apparent molecular weight of 42 and 44 kDa wereobserved with in 5 min of arsenite treatment, which remained highup to 1 h thereby exhibiting a continuous decline in the activity(Fig. 4A). A known feature of MAPKs is their activation upon phos-phorylation on the conserved sequence motif TXY, both at Tyr andThr residues. In order to confirm the activity of MBP phosphorylat-ing kinase is due to a MAP kinase, an immunokinase assay was per-formed. For the purpose, immuno precipitation with anti-phospho-tyrosine antibody (4G10) was carried out in a few representativetime points from arsenite treated leaves and roots. As shown inFig. 4B, immunoprecipitation of leaf samples treated with arsenitefor 30 min, 1 and 3 h and root with 5, 30 and 60 min followed byin-gel kinase assay resulted in precipitating the increasing MBP ki-nase activity. The results obtained confirmed that the activation ofMBP phosphorylating kinase by arsenite treatment is indeed amember of MAP kinase.
Kinetics of MAPK activation by arsenite is dose dependent
Rice seedlings treated with 50 lM arsenite resulted in the acti-vation of MAP kinase in leaves and roots. To investigate the doseresponse of arsenite on the activation of MAPK, 2 weeks old riceseedlings were treated with 0.1, 10, 50, 100, 150 and 300 lM of

Fig. 3. Generation of ROS and NO in arsenite treated rice roots. (a–d) Generation of ROS (a) and (c) are the bright field, DCF (20 ,70-dichlorofluorescin) fluorescence image ofcontrol rice roots treated with ROS-sensitive dye, respectively. (b) and (d) are the bright field, DCF fluorescence image of rice roots treated with 50 lM arsenite followed byROS-sensitive dye, respectively. (e–h) Generation of nitric oxide (NO) in arsenite treated rice roots. (e) and (g) are the bright field, DAF-2T (diamino fluorescein) fluorescenceimage of control rice roots treated with NO sensitive dye, respectively. (g) and (h) are the bright field, DAF-2T fluorescence image of rice roots treated with 50 lM arsenitefollowed by ROS-sensitive dye, respectively.
Fig. 4. Activation of MAPK by arsenite in rice seedlings. (A) Two week old riceseedlings were treated with 50 lM arsenite samples, harvested at respective timepoints and in-gel kinase assay was performed using MBP as substrate. (B) Crudeprotein extracts of rice leaf and root samples were immuno precipitated with 4G10antibody and analyzed by in-gel kinase assay. Myelin basic protein was used assubstrate in the assay. The experiments were repeated at least three times withsimilar results.
K.P. Rao et al. / Archives of Biochemistry and Biophysics 506 (2011) 73–82 77
sodium meta-arsenite and water as mock treatment as control inseparate set of experiments. Leaf and root samples were harvestedat different time points up to 9 h and analyzed by in-gel kinase as-say. In case of leaf, activation of MAPK was observed from 3 h with0.1, 10 and 50 lM arsenite treatment, whereas early activation ofMAPK with in 30 min to 1 h was observed with higher concentra-tions of 100, 150 and 300 lM arsenite treatment (Fig. 5). In case ofroots the response was very quick and activation of MAPK was ob-served with in 5 min of adding arsenite at 50 and 100 lM concen-trations. The activation continued up to 3 h with peak of activity at1 h. When rice plants were treated with 150 and 300 lM arsenitethe activity of MAPK increased in 5 min with maximum activityshifted to 30 min to 1 h and continued up to 9 h. Very less or faintactivity of MAP kinase was observed with 0.1 and 10 lM of arse-nite treatment in 30 min to 1 h. In most of the treatments the acti-vated activity of MAP kinase was maintained up to 9 h. No activityof MAP kinase was observed in rice seedlings mock treated withwater. Overall this result indicates that kinetics of MAPK activitywas strongly correlated with the increasing concentration of arse-nite treatment in leaves and roots.
OsMPK3 is transcriptionally upregulated both in leaves and roots byarsenite stress
To study the transcript regulation of MAPKs in arsenite stress,2 weeks old rice plants were treated with 50 lM arsenite and sam-ples were collected at 30 min, 3 and 9 h. Transcript analysis of theMAPKs in arsenite treated leaves by quantitative real time PCR re-vealed that among the five members present in group A, B and Conly OsMPK3 showed increase in transcript level from 30 min to9 h. Almost 7 folds increase in transcript level of OsMPK3 was ob-served compared to control after 9 h of arsenite treatment(Fig. 6A). From group D, which comprises of 11 membersOsMPK17-1 showed increase in transcript level at 3 h followed bya decrease at 9 h. While OsMPK21-1 and OsMPK21-2 transcript lev-els remain unchanged up to 3 h and showed increase at 9 h.
In case of roots, two MAPKs, OsMPK3 and OsMPK4 belonging togroup A and B, respectively, responded at transcript level to arse-nite treatment (Fig. 6B). There was a continuous increase in thetranscript level of OsMPK3 up to 3 h, thereafter a slight decreasewas observed at 9 h. In case of OsMPK4 there was 13 folds increasein transcript level at 3 h followed by decrease at 9 h, which was 8fold higher than the control. Among the group D, OsMPK17-2 andOsMPK20-1 showed 20 fold increase at 3 h followed by 44 folds in-crease at 9 h. An up-regulation in the transcript levels of OsMPK16-2, OsMPK17-1, OsMPK21-1 and OsMPK21-2 were also observed. Itwas clear from these results that OsMPK3 a member of group AMAPKs has shown correlated transcript regulation with arsenitetreatment in both leaves and roots.
To validate further the results obtained by qRT-PCR, the pro-teins from leaf tissues at 180 min and root tissues at 60 min treatedwith arsenite were immunoprecipitated with Arabidopsis AtMPK3and AtMPK4 antibodies. These antibodies showed cross-reactionwith OsMPK3 and OsMPK4 proteins expressed in bacteria (datanot shown). The immunoprecipitated proteins were subjected toin-solution kinase assay. Interestingly, it was observed that inleaves OsMPK3 gets activated and not OsMPK4 (Fig 6C) while inroots both OsMPK3 and OsMPK4 showed activity (Fig. 6D).
Transcripts of OsMKK gene family respond differentially in arsenitetreated rice leaves and roots
MAP Kinase Kinase (MKK) is the up stream component of theMAP kinase in MAPK signaling cascade. There are eight membersof MKK in rice, grouped into four groups A, B, C and D. Out of eightOsMKKs, seven were analyzed for the transcript regulation under

Fig. 5. Dose response study of activation of MAPK by arsenite in rice leaves and roots. Two weeks old rice seedlings were treated with 0.1, 1, 10, 50, 100 and 300 lM of sodiummeta-arsenite, leaves and roots were sampled at respective time points in individual experiments. Crude protein extracts were analyzed by in-gel kinase assay for MAP kinaseactivity using MBP as substrate. The experiments were repeated at least three times with similar results.
Fig. 6. Relative gene expression of MAPKs in rice leaves (A) and roots (B) treated with arsenite. Rice seedlings were treated with 50 lM arsenite. Leaves and roots weresampled at 30 min, 3 and 9 h in independent experiments. Transcript regulations were analyzed by quantitative real time PCR. The data represented as an average of threeindependent experiments. In-solution assay after immunoprecipitation with orthologs antibody from Arabidopsis, AtMPK3 and AtMPK4 from rice leaf (C) and root (D)samples. MBP was used as substrate for in-solution kinase assay. The lower panel shows Coomassie Brilliant Blue (CBB) staining for equal loading of the proteins. Theexperiment was repeated twice with similar result.
78 K.P. Rao et al. / Archives of Biochemistry and Biophysics 506 (2011) 73–82
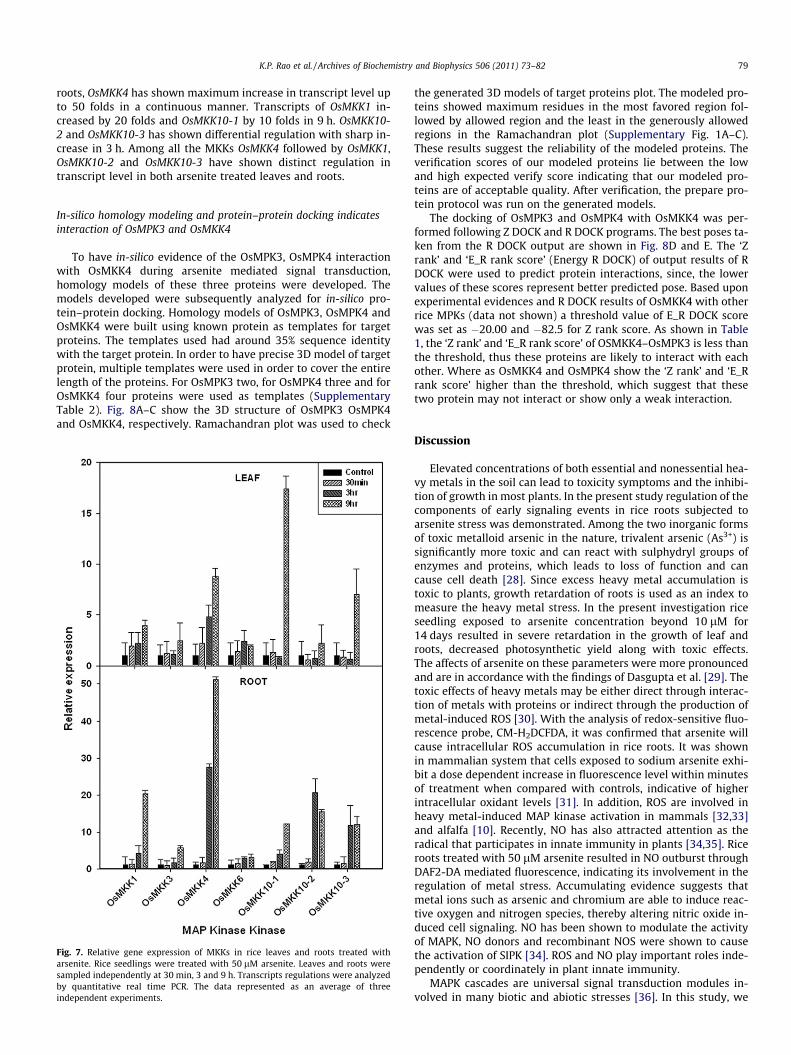
arsenite stress. OsMKK4 and OsMKK5 members of group B MKKsshared 95% sequence homology and hence they were consideredone and named as OsMKK4/5 for the present analysis. Transcriptanalysis of all the MKKs were carried out in the leaves and rootsof 50 lM arsenite treated rice plants by quantitative real time
PCR. OsMKK4 showed 10 fold increase in transcript level up to9 h in leaves, whereas OsMKK10-1 and OsMKK10-3 showed sharpincrease in the transcript level at 9 h (Fig. 7). Another member ofMKK family, OsMKK1 has also shown continuous increase in tran-script level by four folds in 9 h compare to the control. In case of
K.P. Rao et al. / Archives of Biochemistry and Biophysics 506 (2011) 73–82 79
roots, OsMKK4 has shown maximum increase in transcript level upto 50 folds in a continuous manner. Transcripts of OsMKK1 in-creased by 20 folds and OsMKK10-1 by 10 folds in 9 h. OsMKK10-2 and OsMKK10-3 has shown differential regulation with sharp in-crease in 3 h. Among all the MKKs OsMKK4 followed by OsMKK1,OsMKK10-2 and OsMKK10-3 have shown distinct regulation intranscript level in both arsenite treated leaves and roots.
In-silico homology modeling and protein–protein docking indicatesinteraction of OsMPK3 and OsMKK4
To have in-silico evidence of the OsMPK3, OsMPK4 interactionwith OsMKK4 during arsenite mediated signal transduction,homology models of these three proteins were developed. Themodels developed were subsequently analyzed for in-silico pro-tein–protein docking. Homology models of OsMPK3, OsMPK4 andOsMKK4 were built using known protein as templates for targetproteins. The templates used had around 35% sequence identitywith the target protein. In order to have precise 3D model of targetprotein, multiple templates were used in order to cover the entirelength of the proteins. For OsMPK3 two, for OsMPK4 three and forOsMKK4 four proteins were used as templates (SupplementaryTable 2). Fig. 8A–C show the 3D structure of OsMPK3 OsMPK4and OsMKK4, respectively. Ramachandran plot was used to check
Fig. 7. Relative gene expression of MKKs in rice leaves and roots treated witharsenite. Rice seedlings were treated with 50 lM arsenite. Leaves and roots weresampled independently at 30 min, 3 and 9 h. Transcripts regulations were analyzedby quantitative real time PCR. The data represented as an average of threeindependent experiments.
the generated 3D models of target proteins plot. The modeled pro-teins showed maximum residues in the most favored region fol-lowed by allowed region and the least in the generously allowedregions in the Ramachandran plot (Supplementary Fig. 1A–C).These results suggest the reliability of the modeled proteins. Theverification scores of our modeled proteins lie between the lowand high expected verify score indicating that our modeled pro-teins are of acceptable quality. After verification, the prepare pro-tein protocol was run on the generated models.
The docking of OsMPK3 and OsMPK4 with OsMKK4 was per-formed following Z DOCK and R DOCK programs. The best poses ta-ken from the R DOCK output are shown in Fig. 8D and E. The ‘Zrank’ and ‘E_R rank score’ (Energy R DOCK) of output results of RDOCK were used to predict protein interactions, since, the lowervalues of these scores represent better predicted pose. Based uponexperimental evidences and R DOCK results of OsMKK4 with otherrice MPKs (data not shown) a threshold value of E_R DOCK scorewas set as �20.00 and �82.5 for Z rank score. As shown in Table1, the ‘Z rank’ and ‘E_R rank score’ of OSMKK4–OsMPK3 is less thanthe threshold, thus these proteins are likely to interact with eachother. Where as OsMKK4 and OsMPK4 show the ‘Z rank’ and ‘E_Rrank score’ higher than the threshold, which suggest that thesetwo protein may not interact or show only a weak interaction.
Discussion
Elevated concentrations of both essential and nonessential hea-vy metals in the soil can lead to toxicity symptoms and the inhibi-tion of growth in most plants. In the present study regulation of thecomponents of early signaling events in rice roots subjected toarsenite stress was demonstrated. Among the two inorganic formsof toxic metalloid arsenic in the nature, trivalent arsenic (As3+) issignificantly more toxic and can react with sulphydryl groups ofenzymes and proteins, which leads to loss of function and cancause cell death [28]. Since excess heavy metal accumulation istoxic to plants, growth retardation of roots is used as an index tomeasure the heavy metal stress. In the present investigation riceseedling exposed to arsenite concentration beyond 10 lM for14 days resulted in severe retardation in the growth of leaf androots, decreased photosynthetic yield along with toxic effects.The affects of arsenite on these parameters were more pronouncedand are in accordance with the findings of Dasgupta et al. [29]. Thetoxic effects of heavy metals may be either direct through interac-tion of metals with proteins or indirect through the production ofmetal-induced ROS [30]. With the analysis of redox-sensitive fluo-rescence probe, CM-H2DCFDA, it was confirmed that arsenite willcause intracellular ROS accumulation in rice roots. It was shownin mammalian system that cells exposed to sodium arsenite exhi-bit a dose dependent increase in fluorescence level within minutesof treatment when compared with controls, indicative of higherintracellular oxidant levels [31]. In addition, ROS are involved inheavy metal-induced MAP kinase activation in mammals [32,33]and alfalfa [10]. Recently, NO has also attracted attention as theradical that participates in innate immunity in plants [34,35]. Riceroots treated with 50 lM arsenite resulted in NO outburst throughDAF2-DA mediated fluorescence, indicating its involvement in theregulation of metal stress. Accumulating evidence suggests thatmetal ions such as arsenic and chromium are able to induce reac-tive oxygen and nitrogen species, thereby altering nitric oxide in-duced cell signaling. NO has been shown to modulate the activityof MAPK, NO donors and recombinant NOS were shown to causethe activation of SIPK [34]. ROS and NO play important roles inde-pendently or coordinately in plant innate immunity.
MAPK cascades are universal signal transduction modules in-volved in many biotic and abiotic stresses [36]. In this study, we

Fig. 8. Theoretical 3D models of OsMPK3 (A), OsMPK4 (B) and OsMKK4 (C) build by homology modeling and used for protein–protein docking. The red region refers to thealpha helices, sky blue region are the beta sheets, green color depicts the turn whereas the grey color represents the loops. Top docking poses of OsMKK4–OsMPK3 (D) andOsMKK4–OsMPK4 (E). OsMKK4 is represented as molecular pattern where as OsMPK3 and OsMPK4 as ribbon.
Table 1OsMKK4–OsMPK3 and OsMPK4 interaction interphase scores. DOPE – Discrete Optimized Protein Energy, PDF – Probability Density Function, E_R – Energy R DOCK score. Lowervalues of DOPE score, PDF total energy, Z rank score and E_R DOCK refer to the top pose of the complex while higher Z DOCK score refer the same. 0 Clash means there is no stearicclash between the proteins after refinement by R DOCK protocol.
Protein pair DOPE score PDF total energy Z DOCK score Z rank score E_R DOCK Clash
OsMKK4–OsMPK3 �39379.9 13306.10 15.26 �99.423 �25.1994 0OSMKK4–OSMPK4 �42867.3 �11614.00 14.68 �82.402 �18.5373 0
80 K.P. Rao et al. / Archives of Biochemistry and Biophysics 506 (2011) 73–82
have demonstrated that arsenite stimulate a rapid activation oftwo MBP phosphorylating kinases with approximate molecularweight of 42 and 44 kDa in rice roots and 42 kDa in leaves. Furtherthe MBP kinases were confirmed as MAP kinases by their recogni-tion with anti-phospho-tyrosine antibody in an immuno kinase as-say. Immunoprecipitation with 4G10 antibody followed by in-gelkinase assay reveals 42 kDa kinase more strongly then 44 kDa ki-nase in roots. This may be because of the competition for phosphotyrosine residue in 4G10 antibody by the two activated kinase.However, both activities were observed in roots when it waschecked independently with antibodies AtMPK3 and AtMPK4 (Fig6D). MAPK gets activated with in 5 min, as a quick response toarsenite treatment and remains high up to 1–3 h in roots. Whereasin leaves the activation was observed only after 3 h of arsenitetreatment. It may be possible that since roots are in immediatecontact with the arsenite, fast and transient response of MAPKwas induced as a part of early signaling events and subsequentlystimulated delayed response in leaves. Recently, Gupta et al. [20]has shown activation of MAPK by arsenite in B. juncea. In thekinetic analysis with increasing concentration of arsenite revealedthat MAPK activity also progressively increased in both roots andleaves, correlating the activity of MAPK with the concentration ofarsenite. This further strengthens the involvement of MAPKcascade in arsenite stress signaling. Previously it was reported thatexposure of alfalfa (Medicago sativa) seedlings to excess Cd2+ andCu2+ activated four distinct MAP kinases: SAMK, SIMK, MMK2,and MMK3 [10]. SAMK and SIMK are the orthologs of rice OsMPK3and OsMPK6, respectively. Moreover heavy metal-induced MAP ki-nase gene expression [11,12,14,37,38] and MAP kinase activation[14–17] have been reported in rice.
Along with the activation of MAPK in response to stress, it wasevident from the previous reports that they do regulate at tran-script level [39].Transcript analysis of MAPK and MKK gene familywas analyzed in response to arsenite stress by quantitative realtime PCR. There are 15 members of MAPK being reported in rice[40,41], while our analysis in indica cultivar group cv PusaBasmati-1 revealed sixteen members (unpublished data). TheseMAPKs are being classified into four groups, A–D based on their se-quence similarity. In Oryza OsMPK3 and OsMPK6 belongs to groupA, OsMPK4 to group B, OsMPK7 and OsMPK14 to group C whileOsMPKs 16-1, 16-2, 17-1, 17-2, 20-1, 20-2, 20-3, 20-4, 21-1 and21-2 belonged to group D [36,37]. Whereas eight members ofMKKs are being reported from rice which are again classified intofour groups, A–D [22,40]. Out of these eight MKKs, OsMKK1 andOsMKK6 belongs to group A, OsMKK3 to group B, OsMKK4 andOsMKK5 to group C while OsMKK10-1, OsMKK10-2 andOsMKK10-3 belongs to group D. Our quantitative real time PCRanalysis during arsenite stress revealed that OsMPK3, a memberof group A MAPKs, has shown progressive accumulation both inroots and leaves. In roots OsMPK4 transcripts, a member of groupB also showed continuous accumulation in response to arsenitestress. Interestingly we observed two bands in in-gel kinase assayfrom root samples while only one band in leaf samples. Since thepredicted molecular weight of OsMPK3 and OsMPK4 are in therange of 42 and 44 kDa, respectively, it can be speculated thatOsMPK3 gets activated in leaf while both OsMPK3 and OsMPK4gets activated in root samples. To confirm the speculation, we per-formed an immunoprecipitation of arsenic activated MAP kinasesin rice using Arabidopsis AtMPK3 and AtMPK4. Interestingly, inrice leaf immunoprecipitation followed by in-solution kinase assay

K.P. Rao et al. / Archives of Biochemistry and Biophysics 506 (2011) 73–82 81
revealed the activity of only OsMPK3 and not OsMPK4 while inroots activity of both OsMPK3 and OsMPK4 could be observed.
Apart from OsMPK3 and OsMPK4, there are a few MAPKs fromgroup D that showed prominent increase in transcript levels uponarsenite treatment. The predicted molecular weight of group DMAPKs are more than 50 kDa and the activity observed in the pres-ent study were 42 and 44 kDa upon arsenite treatments in in-gelkinase assay. Therefore it can be interpreted that the activity ofgroup D MAPKs was not observed in in-gel kinase assays may bedue to the regulation restricted at transcript level only.
Previous studies have shown that the expression of a rice MAPkinase gene, OsMPK3 is inducible by cold, salinity, Cu2+, and Cd2+
[11,14,15,42]. The deduced amino acid sequence of OsMPK3 com-prises 369 amino acid residues. Its amino acid sequence is identicalto that of OsMSRMK2 [12], OsMAP1 [43], OsBIMK1 [44], and Os-MAPK5 [45]. In these studies, the OsMPK3 gene was also inducedby various biotic and abiotic stresses and was suggested to play arole in stress signal transduction.
In the MAPK signaling cascade MAP Kinase Kinase (MKK) actsupstream to the MAP kinases. Transcript analysis of MKK genefamily revealed the regulatory pattern of MKKs in arsenite trea-ted rice plants. Among the MKK gene family, OsMKK4 has shownprogressive increase in transcript level followed by OsMKK1 inleaves and roots. In a separate yeast two-hybrid screen, Dinget al. [46] reported that OsMKK4 (Os02g54600) was linked toOsMPK3 (Os03g17700), which has been implicated in stress tol-erance and host resistance [45]. Thus, OsMKK4 might functionas an upstream regulator for OsMPK3. Moreover OsMKK4 isorthologs to the Arabidopsis AtMKK4, which has been identifiedas the upstream kinases of AtMPK3 and AtMPK6 in the flagel-lin-triggered pathway [47]. Given that OsMPK3 is orthologs toAtMPK3 [40], the proposed OsMKK4–OsMPK3 cascade in arseniteinduced signal in rice appears to be parallel to the AtMKK4–AtMPK3/AtMPK6 pathway reported in Arabidopsis innate immu-nity [47]. Previously Jonak et al. [10] reported that SIMKK specif-ically mediates the activation of SIMK and SAMK in response tocopper stress in alfa alfa cells. OsMKK4 is an ortholog of SIMKKin rice. Since MAPK cascades are highly conserved across theplants it can be speculated that OsMKK4 may act upstream tothe OsMPK3 in heavy metal stress signaling in particular and ar-senic stress signaling in rice. Regulation of OsMKK1, OsMKK10-2and OsMKK10-3 was also observed in rice roots and leaves.
To have further insight into OsMPK3 and OsMPK4 activation isdependent on activation of upstream OsMKK4, in-silico homologymodel of these three proteins were generated and analyzed fordocking study. If the binding interface of the modeled protein isin its near native conformation then the protein–protein interac-tions can be predicted more precisely. So it is not necessary thatwe find the whole length of protein in its native conformationsto proceed with protein–protein docking. That is why homologymodels can be used for the interaction predictions. The dockingstudy revealed good interaction between OsMPK3 and OsMKK4.However the interaction of OsMPK4 with OsMKK4 was predictedto be weak. These predictions are in agreement with the recentstudy [48] where OsMKK4 is shown to activate OsMPK3 andOsMK6 in vivo. However, in vitro OsMKK4 shows phosphorylationof OsMPK3, OsMPK6 and also OsMPK4. It is plausible that, OsMKK4differentially phosphorylate these groups A and B OsMPKs in re-sponse to different stimuli.
The results presented showed arsenite has detrimental effect onthe photosynthetic efficiency and growth of the rice seedlings. Fur-ther, it revealed the involvement of MAP kinase cascade along withROS and NO production in arsenite mediated stress signal trans-duction in plants. The results also indicated that possibly OsMPK3and OsMPK4 are the MAPKs responsible for this signal transduc-tion along with OsMKK4 as an upstream kinase of the cascade.
Though a genetic analysis needs to be carried out to confirm theobservation.
Acknowledgments
K.P.R., K.K. and D.P.W. acknowledge the fellowship provided byUniversity Grant Commission, India. The work is financially sup-ported by a grant from Department of Biotechnology, Govt. ofIndia.
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.abb.2010.11.006.
References
[1] A. Di-Domenico, C.L. Rocca, C. Lintas, L.T. Baldassar, Bull. Environ. Contam.Toxicol. 43 (1998) 556–563.
[2] F.J. Zhao, J.F. Ma, A.A. Mehrag, S.P. Mcgrath, New Phytol. 181 (2009) 777–794.[3] N. Verbruggen, C. Hermans, H. Schat, Curr. Opin. Plant Biol. 12 (2009) 364–372.[4] R.S. Oremland, J.F. Stolz, Science 300 (2003) 939–944.[5] R.D. Tripathi, S. Srivastava, S. Mishra, N. Singh, R. Tuli, D.K. Gupta, F.J. Maathuis,
Trends Biotechnol. 25 (2007) 158–165.[6] S. Clemens, Planta 212 (2001) 475–486.[7] C. Cobbett, P. Goldsbrough, Annu. Rev. Plant Biol. 53 (2002) 159–182.[8] J.L. Hall, L.E. Williams, J. Exp. Bot. 54 (2003) 2601–2613.[9] M. Gupta, N.B. Sarin, J. Environ. Sci. 21 (2009) 686–690.
[10] C. Jonak, H. Nakagami, H. Hirt, Plant Physiol. 136 (2004) 3276–3283.[11] C.M. Yeh, L.J. Hsiao, H.J. Huang, Plant Cell Physiol. 45 (2004) 1306–1312.[12] G.K. Agrawal, R. Rakwal, H. Iwahashi, Biochem. Biophys. Res. Commun. 294
(2002) 1009–1016.[13] G.K. Agrawal, S.K. Agrawal, J. Shibato, H. Iwahashi, R. Rakwal, Biochem.
Biophys. Res. Commun. 300 (2003) 775–783.[14] C.M. Yeh, W.C. Hung, H.J. Huang, Physiol. Plant 119 (2003) 392–399.[15] C.M. Yeh, P.S. Chien, H.J. Huang, J. Exp. Bot. 58 (2007) 659–671.[16] C.W. Lin, H.B. Chang, H.J. Huang, Plant Physiol. Biochem. 43 (2005) 963–968.[17] T.M. Tsai, H.J. Huang, Physiol. Plant 127 (2005) 583–592.[18] J.M. Samet, L.M. Graves, J. Quay, L.A. Dailey, R.B. Devlin, A.J. Ghio, W. Wu, P.A.
Bromberg, W. Reed, Am. J. Physiol. Lung Cell Mol. Physiol. 275 (1998) L551–L558.
[19] D. Chakrabarty, P.K. Trivedi, P. Misra, M. Tiwari, M. Shri, D. Shukla, S. Kumar, A.Rai, A. Pandey, D. Nigam, R.D. Tripathi, R. Tuli, Chemosphere 74 (2009) 688–702.
[20] M. Gupta, P. Sharma, N.B. Sarin, A.K. Sinha, Chemosphere 74 (2009) 1201–1208.
[21] K.P. Rao, G. Vani, K. Kumar, A.K. Sinha, Mol. Cells 28 (2009) 417–422.[22] K. Kumar, K.P. Rao, P. Sharma, A.K. Sinha, Plant Physiol. Biochem. 46 (2008)
891–897.[23] H.M. Berman, J. Westbrook, Z. Feng, G. Gilliland, T.N. Bhat, H. Weissig, I.N.
Shindyalov, P.E. Bourne, Nucleic Acids Res. 28 (2000) 235–242.[24] A. Sali, L. Potterton, F. Yuan, H. van Vlijmen, M. Karplus, Proteins 23 (3) (1995)
318–326.[25] Rong Chen, Julian Mintseris, Joël Janin, Zhiping Weng, Proteins: Struct. Funct.
Bioinf. 52 (1) (2003) 88–91.[26] Li Li, Rong Chen, Zhiping Weng, Proteins: Struct. Funct. Bioinf. 53 (3) (2003)
693–707.[27] K. Maxwell, G.N. Johnson, J. Exp. Bot. 51 (2000) 659–668.[28] R. Requejo, M. Tena, Phytochemistry 66 (2005) 1519–1528.[29] T. Dasgupta, S.A. Hossain, A.A. Meharg, A.H. Price, New Phytol. 163 (2004) 45–
49.[30] S.S. Leonard, G.K. Harris, X.L. Shi, Free Radic. Biol. Med. 37 (2004) 1921–1942.[31] S.X. Liu, M. Athar, I. Lippai, C.A. Waldren, T.K. Hei, Proc. Natl. Acad. Sci. USA 98
(2001) 1643–1648.[32] S.R. Seo, S.A. Chong, S.I. Lee, J.Y. Sung, Y.S. Ahn, K.C. Chung, J.T. Seo, J.
Neurochem. 78 (2001) 600–610.[33] P. Rockwell, J. Martinez, L. Papa, E. Gomes, Cell. Signal. 16 (2004) 343–353.[34] D. Kumar, D.F. Klessig, Mol. Plant Microbe Interact. 13 (2000) 347–351.[35] S. Asai, H. Yoshioka, Plant Signal Behav. 3 (2008) 920–922.[36] M.C.S. Rodriguez, M. Petersen, J. Mundy, Annu. Rev. Plant Biol. 61 (2010) 621–649.[37] G.K. Agrawal, H. Iwahashi, R. Rakwal, Biochem. Biophys. Res. Commun. 302
(2003) 171–180.[38] W.C. Hung, D.D. Huang, C.M. Yeh, H.J. Huang, Plant Growth Regul. 45 (2005)
233–241.[39] L. Bogre, W. Ligterink, I. Meskiene, P.J. Barker, E. Heberle-Bors, N.S. Huskisson,
H. Hirt, Plant Cell 9 (1997) 75–83.[40] L.P. Hamel, M.C. Nicole, S. Sritubtim, M.J. Morency, M. Ellis, J. Ehlting, N.
Beaudoin, B. Barbazuk, D. Klessig, J. Lee, G. Martin, J. Mundy, Y. Ohashi, D.Scheel, J. Sheen, T. Xing, S. Zhang, A. Seguin, B.E. Ellis, Trends Plant Sci. 11(2006) 192–199.
[41] M.A.P.K. Group, Trends Plant Sci. 7 (2002) 301–308.

82 K.P. Rao et al. / Archives of Biochemistry and Biophysics 506 (2011) 73–82
[42] H.J. Huang, S.F. Fu, Y.H. Tai, W.C. Chou, D.D. Huang, Physiol. Plant 114 (2002)572–580.
[43] J.Q. Wen, K. Oono, R. Imai, Plant Physiol. 129 (2002) 1880–1891.[44] F. Song, R.M. Goodman, Planta 215 (2002) 997–1005.[45] L. Xiong, Y. Yang, Plant Cell 15 (2003) 745–759.[46] X. Ding, T. Richter, M. Chen, H. Fuji, Y.S. Seo, M. Xie, X. Zheng, S. Kenrar, et al.,
Plant Physiol. 149 (2009) 1478–1492.
[47] T. Asai, G. Tena, J. Plotnikova, M.R. Willmann, W.L. Chiu, L. Gomez-Gomez, T.Boller, F.M. Ausubel, J. Sheen, Nature 415 (2002) 977–983.
[48] M.K. Kaboshi, K. Okada, L. Kurimoto, S. Murakami, T. Umezawa, N. Shibuya, H.Yamane, A. Miyao, H. Takatsuji, A. Takahashi, H. Hirochika, Plant J. 63 (2010)599–612.




















