
Some applications of excited-state-excited-state transition densities

www.elsevier.com/locate/brainres
Brain Research 1065
Research Report
Exercise affects glutamate receptors in postsynaptic densities
from cortical mice brain
Marcelo O. Dietrich a, Carlos E. Mantese a, Lisiane O. Porciuncula a, Gabriele Ghisleni a,
Lucia Vinade a,b, Diogo O. Souza a, Luis V. Portela a,*
aDepartamento de Bioquımica, Instituto de Ciencias Basicas da Saude, Universidade Federal do Rio Grande do Sul,
Rua Ramiro Barcelos, 2600-Anexo 90035-003, Porto Alegre/RS, BrazilbLaboratory of Neurobiology, National Institute of Neurological Disorders and Stroke, National Institutes of Health, Bethesda, Maryland, USA
Accepted 27 September 2005
Available online 18 November 2005
Abstract
Physical activity has been proposed as a behavior intervention that promotes mental health and some of the benefits induced by exercise
have been related to the glutamatergic system. Indeed, glutamate is the most abundant excitatory neurotransmitter in brain. Thus, we
evaluated if voluntary exercise in mice could modulate glutamatergic synapses at level of postsynaptic density (PSD). Through Western blot,
we found that exercise during 1 month increased glutamatergic-related protein content in PSD from cortex of mice. Exercise increased the
immunocontent of GluR1 (129%), SAP-97 (179%), GRIP-1 (129%), and in less extent, GluR2/3 (118%) and PSD-95 (112%) proteins. The
overall content of NMDA subunits R1, R2A and R2B were not altered in mice that had exercised, however, the phosphorylated NMDA
subunits, phospho-NMDAR1 (150%), and phospho-NMDAR2B (183%) showed a strong increase. Because exercise increased the content of
phosphorylated forms of NMDA receptors, we evaluated the binding of MK-801, a specific ligand that binds to open NMDA channel.
Exercise increased the binding of MK-801 in cortical cellular membranes in 51%. Altogether, our results point to a modulation of
glutamatergic synapses by exercise with likely implications in the exercise-induced mental health.
D 2005 Elsevier B.V. All rights reserved.
Theme: Excitable membranes and synaptic transmission
Topic: Postsynaptic mechanisms
Keywords: Voluntary exercise; Brain; Glutamate receptor; NMDA; Phosphorylation
1. Introduction
Glutamate is the most abundant neurotransmitter in the
mammalian central nervous system (CNS), acting on iono-
0006-8993/$ - see front matter D 2005 Elsevier B.V. All rights reserved.
doi:10.1016/j.brainres.2005.09.038
Abbreviations: AMPA, a-amino-3-hydroxi-5-methyl-4-isoxazol pro-
pionic acid; GluR1, GluR2/3, GluR6/7, glutamate receptor subunit type
1, 2/3 and 6/7, respectively; GRIP-1, glutamate receptor interacting protein
1; MAGUK, membrane-associated guanylyl kinase; NMDA, N-methyl-d-
aspartate; p-NR1 and p-NR2B, phosphorylated NMDA receptor subunit
type 1 and type 2B, respectively; PSD, postsynaptic density; PSD-95,
postsynaptic density protein of 95 kDa; SAP-97, synapse associated protein
97
* Corresponding author. Fax: +55 51 3316 5540.
E-mail address: [email protected] (L.V. Portela).
tropic N-methyl-d-aspartate (NMDA), a-amino-3-hydroxi-
5-methyl-4-isoxazol propionic acid (AMPA) and kainate
(KA) receptors, and on metabotropic G protein-coupled
receptors, all of which are essential receptors for normal brain
functions [35]. However, as overstimulation of glutamatergic
receptors triggers neuronal damage involved in acute and
chronic brain diseases [28,31,35,46], a fine modulation of
glutamatergic synapses activity is pivotal for CNS function.
Regular physical activity, an essential component of a
healthy lifestyle, recently has been shown to mediate central
nervous system (CNS) adaptations, up to now more
extensively reported in hippocampus [1,6]. Animal studies
have demonstrated that exercise protects neurons from
various brain insults [4,18,47,54], activates neuronal cells
(2005) 20 – 25
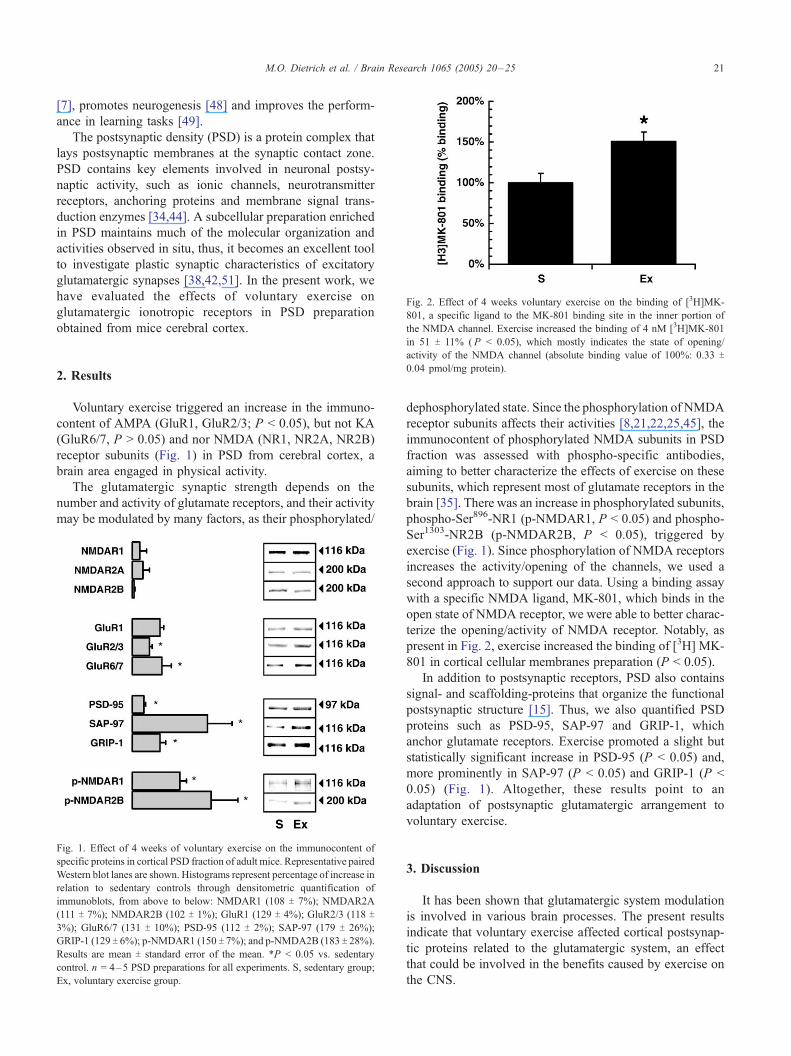
Fig. 2. Effect of 4 weeks voluntary exercise on the binding of [3H]MK-
801, a specific ligand to the MK-801 binding site in the inner portion of
the NMDA channel. Exercise increased the binding of 4 nM [3H]MK-801
M.O. Dietrich et al. / Brain Research 1065 (2005) 20–25 21
[7], promotes neurogenesis [48] and improves the perform-
ance in learning tasks [49].
The postsynaptic density (PSD) is a protein complex that
lays postsynaptic membranes at the synaptic contact zone.
PSD contains key elements involved in neuronal postsy-
naptic activity, such as ionic channels, neurotransmitter
receptors, anchoring proteins and membrane signal trans-
duction enzymes [34,44]. A subcellular preparation enriched
in PSD maintains much of the molecular organization and
activities observed in situ, thus, it becomes an excellent tool
to investigate plastic synaptic characteristics of excitatory
glutamatergic synapses [38,42,51]. In the present work, we
have evaluated the effects of voluntary exercise on
glutamatergic ionotropic receptors in PSD preparation
obtained from mice cerebral cortex.
in 51 T 11% ( P < 0.05), which mostly indicates the state of opening/activity of the NMDA channel (absolute binding value of 100%: 0.33 T
0.04 pmol/mg protein).
2. ResultsVoluntary exercise triggered an increase in the immuno-
content of AMPA (GluR1, GluR2/3; P < 0.05), but not KA
(GluR6/7, P > 0.05) and nor NMDA (NR1, NR2A, NR2B)
receptor subunits (Fig. 1) in PSD from cerebral cortex, a
brain area engaged in physical activity.
The glutamatergic synaptic strength depends on the
number and activity of glutamate receptors, and their activity
may be modulated by many factors, as their phosphorylated/
Fig. 1. Effect of 4 weeks of voluntary exercise on the immunocontent of
specific proteins in cortical PSD fraction of adult mice. Representative paired
Western blot lanes are shown. Histograms represent percentage of increase in
relation to sedentary controls through densitometric quantification of
immunoblots, from above to below: NMDAR1 (108 T 7%); NMDAR2A
(111 T 7%); NMDAR2B (102 T 1%); GluR1 (129 T 4%); GluR2/3 (118 T
3%); GluR6/7 (131 T 10%); PSD-95 (112 T 2%); SAP-97 (179 T 26%);
GRIP-1 (129 T 6%); p-NMDAR1 (150 T 7%); and p-NMDA2B (183 T 28%).
Results are mean T standard error of the mean. *P < 0.05 vs. sedentary
control. n = 4–5 PSD preparations for all experiments. S, sedentary group;
Ex, voluntary exercise group.
dephosphorylated state. Since the phosphorylation of NMDA
receptor subunits affects their activities [8,21,22,25,45], the
immunocontent of phosphorylated NMDA subunits in PSD
fraction was assessed with phospho-specific antibodies,
aiming to better characterize the effects of exercise on these
subunits, which represent most of glutamate receptors in the
brain [35]. There was an increase in phosphorylated subunits,
phospho-Ser896-NR1 (p-NMDAR1, P < 0.05) and phospho-
Ser1303-NR2B (p-NMDAR2B, P < 0.05), triggered by
exercise (Fig. 1). Since phosphorylation of NMDA receptors
increases the activity/opening of the channels, we used a
second approach to support our data. Using a binding assay
with a specific NMDA ligand, MK-801, which binds in the
open state of NMDA receptor, we were able to better charac-
terize the opening/activity of NMDA receptor. Notably, as
present in Fig. 2, exercise increased the binding of [3H] MK-
801 in cortical cellular membranes preparation (P < 0.05).
In addition to postsynaptic receptors, PSD also contains
signal- and scaffolding-proteins that organize the functional
postsynaptic structure [15]. Thus, we also quantified PSD
proteins such as PSD-95, SAP-97 and GRIP-1, which
anchor glutamate receptors. Exercise promoted a slight but
statistically significant increase in PSD-95 (P < 0.05) and,
more prominently in SAP-97 (P < 0.05) and GRIP-1 (P <
0.05) (Fig. 1). Altogether, these results point to an
adaptation of postsynaptic glutamatergic arrangement to
voluntary exercise.
3. Discussion
It has been shown that glutamatergic system modulation
is involved in various brain processes. The present results
indicate that voluntary exercise affected cortical postsynap-
tic proteins related to the glutamatergic system, an effect
that could be involved in the benefits caused by exercise on
the CNS.

M.O. Dietrich et al. / Brain Research 1065 (2005) 20–2522
AMPA receptors are involved in fast synaptic trans-
mission at glutamatergic synapses with consequent depola-
rization of the cellular membrane, promoting the opening of
NMDA receptors, a mechanism that has been related to
long-term potentiation (LTP) [17], neuroprotection [14], and
neurotrophic processes [24]. Additionally, AMPA receptors
have a particular and complex system of trafficking,
involved in the synaptic plasticity [2,27,36,43]. Although
KA and AMPA receptors have been difficult to be differ-
entiated, it has been shown that at postsynaptic membranes
KA receptors modulate the glutamatergic neurotransmission
[20]. Recently, it has been evidenced that specific membrane
proteins are able to connect KA to AMPA receptors,
mediating synaptic transmission in some neuronal networks
[5]. Our results suggest that regular physical exercise
promotes synapse plasticity by increasing the content of
AMPA in the PSD (Fig. 1). However, the importance of
increased AMPA receptors content in postsynaptic sites
needs to be further evaluated to access their function under
physiological adaptations.
Some benefits induced by exercise seem to involve
NMDA receptors, as the increase in neurogenesis [16] and
improvement in long-term potentiation [49]. In addition,
molecular approaches demonstrated that induction by
exercise of striatal Fos expression is dependent of NMDA
receptor activity [26]. Recently, it has been suggested that
NMDA receptors activity is an essential step to exercise-
induced synaptic plasticity in hippocampus [50]. NMDA
receptors, mostly studied due to their wide-ranging involve-
ment in many brain functions, have been associated with
many physiological and pathological brain conditions.
NMDA receptors exert a key role on neuronal excitability
and synaptic signaling through a fine control of its channel
activity. Accordingly, phosphorylation of NMDA receptors
increases the ion current through the receptor, whereas their
activity decreases by action of protein phosphatases
[23,41,53]. Moreover, phosphorylation of NMDA receptors
modulates their trafficking [39]. The present study gives
evidences of NMDA receptor modulation by exercise
through an increase in the phosphorylation of the subunits
NR1 and NR2B, and an increase in the binding of a channel
ligand (MK-801), which binds to the open NMDA channel,
with likely implications in brain physiology. Both data
together should indicate that our protocol of exercise
induced an opening of NMDA receptors and consequently
should increase their activities.
In addition to the evaluation of glutamate receptors, we
also studied the content of glutamate-related proteins, which
anchor receptors, maintaining the architecture and function-
ality of postsynaptic densities. Accordingly, the content of
anchoring proteins (PSD-95, SAP-97 and GRIP-1) accom-
panied the increase of the glutamate receptors subunits.
PSD-95 and SAP-97 are members of the MAGUK
(membrane-associated guanylyl kinase) proteins. PSD-95
has been described to anchor NMDA receptors (through
NR2 subunits) [42] and kainate receptors (through GluR6
subunits) [10]. SAP-97 anchors both AMPA (through
GluR1 subunit) [19] and kainate receptors (GluR6 subunit)
[30]. Similarly, GRIP-1 anchors the GluR2/3 subunits of
AMPA receptors [9] and the GluR5 subunit of kainate
receptors, as well [12]. These complex interactions among
anchoring proteins and glutamatergic receptors are involved
in the synaptic plasticity. Therefore, our results suggest that
voluntary exercise modulate these interactions.
Even though the majority of molecular and neuro-
chemical alterations were reported in the hippocampus, we
cannot exclude the importance of the cortical region in the
exercise-mediated effects. In fact, monitoring of local
cerebral glucose utilization showed that running wheel
exercise increase this index in some cortical regions at the
same magnitude as in the hippocampus [52]. Moreover,
exercise promotes increase in the cortical mRNA BDNF, the
molecule proposed as responsible for the exercise benefits
[32]. Recently, the levels of proteins involving in the
intracellular signaling including the presynaptic protein
synapsin I were found to be increase in cortical regions
from exercised animals [11]. Despite these observations,
studies exploring the effects of physical activity in other
brain regions and in different synaptic compartments are
very scarce. Thus, our study highlights some alterations in
synaptic proteins in a very specialized region of the synapse,
which could reflect on the brain function.
Summarizing, our results demonstrate that voluntary
physical exercise affected the immunocontent of glutamate
receptors type AMPA and related-proteins, and the opening/
activity of NMDA (through the modulation of the phos-
phorylation/dephosphorylation state), suggesting that the
glutamatergic system may be a putative target for modu-
lation by regular exercise activity.
It has been shown that the activation of the glutamatergic
neurotransmission may be beneficial or detrimental, depend-
ing among other factors on the degree of activation. Slight
activation may beneficial, as observed in memory studies
(see [13]), while over stimulation may be detrimental, as
observed in various acute and chronic brain injuries (see
[28,29]). Concerning the involvement of the glutamatergic
neurotransmission on brain effects of exercise, this duality is
also present, the exercise potentially being beneficial [1] or
detrimental [40].
Thus, the functional relevance of the effects of exercise
on the glutamatergic receptors presented in this study is
opened and under investigation by our group.
4. Experimental procedures
4.1. Animals
Male albino CF1 mice, 2 months old (from Fundacao
Estadual de Producao e Pesquisa do Rio Grande do Sul,
Porto Alegre, Brazil), were housed in standard cages (48 �26 cm), 3 to 4 animals per cage. Animals were allocated in a

M.O. Dietrich et al. / Brain Research 1065 (2005) 20–25 23
room with controlled temperature (22 T 1 -C), under a 12-hlight/12-h dark cycle. Mice were divided in two groups:
sedentary (S) and voluntarily physical active (exercise, Ex)
groups. Animals from Ex group had free access, for 1 month,
to a running wheel connected to an electronic counter, which
was located within the living cage; S group stayed for the
same time in similar conditions without running wheel. The
animals were controlled for a distance. After 4 weeks of
running wheel, the average distance was 3500 m. All
experiments were in agreement with Committee on Care
and Use of Experimental Animal Resources, UFRGS, Brazil.
4.2. Materials
Antibodies against GRIP-1 (glutamate receptor interact-
ing protein 1), GluR1, GluR2/3, GluR6/7 (glutamate
receptor subunit type 1, 2/3 and 6/7, respectively),
phospho-NR1ser896 and phospho-NR2Bser1303 (phosphory-
lated subunits of NMDA receptor type 1 and type 2B,
respectively) were obtained from Upstate (Lake Placid, NY,
USA). Antibodies against PSD-95 (postsynaptic density
protein of 95 kDa) and SAP-97 (synapse associated protein
97) were obtained from ABR (Golden, CO, USA). Anti-
bodies against NR1, NR2A and NR2B (NMDA receptor
subunits type 1, 2A, and 2B respectively) were purchased
from Chemicon Int. (Temecula, CA, USA). [3H] MK-801
(24.2Ci/mmol) was purchased from Perkin-Elmer Life
Sciences, Ltd., USA.
4.3. Postsynaptic density (PSD) preparation
The PSD fraction was prepared as previously described
[3] with some modifications. Cerebral cortices from Wistar
rats were dissected at 0–4 -C, manually homogenized in
0.32 M sucrose, 1 mM MgCl2 and 1 Ag/ml leupeptin in 1
mM of HEPES (pH 7.4) (1 g of tissue/4 ml) and
synaptosomes were obtained by differential and density-
gradient centrifugations as follows: sucrose gradients
containing 8 ml of the resuspended brain preparation and
gradients containing 10 ml each of 1.25 M, 1.0 M and
0.85 M sucrose solutions were centrifuged for 2 h at
80,000 � g using SW 28 rotor (Beckman Instruments,
Spinco Division, Palo Alto, CA, USA). The synaptosomal
fraction was collected from the interface of 1.25 M and 1.0
M sucrose and diluted with 1 mM of HEPES, pH 7.4, to
11 ml/g of original tissue. An equal volume of 1% Triton
X-100 in 0.32 M sucrose was added and stirred at 4 -C for
15 min. This suspension was centrifuged at 36,000 � g for
30 min and the pellets were resuspended into 0.32 M sucrose
(0.4 ml/g of original tissue). Two milliliters of this material
layered onto gradients in polyallomer tubes containing 4 ml
of 2.1 M sucrose and 3 ml each of 1.5 M and 1.0 M sucrose.
The gradients were centrifuged for 2 h at 200,000 � g in a
SW 40 rotor (Beckman Instruments). The postsynaptic
fraction was collected from the 2.1–1.5 M interface with a
siliconized plastic pipette and diluted with 20 mM of HEPES
and an equal volume of 1% Triton X-100 in 150 mM KCl to
obtain 0.5 ml/g of original tissue. The PSD fraction was
collected on the 2.1 M sucrose cushion by centrifugation at
20,0000 � g for 40 min. This interface was then
resuspended in 20 mM HEPES, pH 7.4, and again collected
on the 2.1 M sucrose by centrifugation at 200,000 � g for 30
min. The PSD fraction from the interface was resuspended
by stirring in 40% glycerol and stored at �20 -C.
4.4. Synaptic membrane preparation
Synaptic membrane preparation was obtained from the
cerebral cortices, as described by Jones and Matus [15], and
stored frozen at �70 -C for up to 2 months. On the day of
assay, membranes were thawed at 37 -C, for 30 min, washed
3 times with 5 mM Tris–HCl, pH 7.4 at 27,000 � g for 15
min. The final pellet was resuspended in the same buffer to
yield a protein concentration of 0.5–2 mg/ml and used for the
[3H]MK-801 binding assay.
4.5. Protein measurement
The protein content of PSD fraction and of synaptic
membrane preparation was estimated by the method of
Peterson [37].
4.6. [3H] MK-801 binding assay
Incubations were started by adding 50 Ag of membrane
protein to a medium containing 5 mM Tris–HCl, pH 7.4
and 4 nM [3H] MK-801. Following 60 min of incubation at
37 -C, bound and free [3H] MK-801 were separated by
filtration through GF/B Whatman filters. Radioactivity was
determined using a Wallac liquid scintillation counter.
Specific binding was calculated as the difference between
total binding and non-specific binding, which was measured
in the presence of 4AM MK-801. Non-specific binding
typically amounted to 20–30% of the total binding [33].
4.7. Electrophoresis and Western blot analysis
Proteins from PSD preparations were separated by 7.5%
SDS-PAGE mini-gels and transferred to nitrocellulose
membrane using a Trans-Blot system (Bio-Rad, Hercules
CA). Membranes were processed as follow: (1) blocking
with 2% bovine serum albumin for 2 h; (2) incubation with
primary antibody for 1 h: 1:5000 anti-PSD-95; 1:1000, anti-
SAP-97; 1:1000 anti-GRIP-1; 1:1000 anti-GluR1; 1:1000
anti-GluR2/3; 1:1000 anti-GluR 6/7; 1:5000 anti-NR1;
1:1000 anti-p-NR1; 1:100 anti-NR2B; 1:1000 anti-p-
NR2B; 1:1000 anti-NR2A; (3) incubation with alkaline
phosphatase-conjugated secondary antibody (1:5000 anti-
mouse or 1:10,000 anti-rabbit) for 1 h; (4) developing with
alkaline phosphatase substrate. Membranes were then dried
and scanned. Measurement of bands intensity through total
area of the peaks was performed using the public domain

M.O. Dietrich et al. / Brain Research 1065 (2005) 20–2524
NIH Image program (Developed at the U.S. National
Institutes of Health and available on the Internet at http://
rsb.info.nih.gov/nih-image/). An internal control in PSD is
unknown, i.e., a protein which content does not vary with
exercise; thus it was performed additional examination of
the total amount of protein loaded on the gel. We used
Coomassie blue stain in gels, analyzed by NIH Image, and
Ponceau S red stain in membranes to be sure the same
quantity of protein was loaded in each lane (5 Ag/lane).When protein stained among lanes differed more than 5%
we did not develop the membrane.
4.8. Statistical analysis
Optical densities (OD) and binding values were com-
pared through paired t test. It was necessary 8 animals for
each PSD preparation. We performed 4 PSD preparations in
each experiment (two exercise and two sedentary). Thus, we
used unpaired t test (two-tailed) comparing samples
prepared in the same day to avoid bias due to PSD
preparation. Additionally, these pairs of samples were run
in the same gel in paired lanes. The results are presented as
percent average T standard error of the mean (SEM),
considering the S group as 100%. P < 0.05 was considered
statistically significant.
Acknowledgments
This work was support by Brazilian agencies, CNPq,
CAPES and FAPERGS (BPV 04/60002.2 for LV) and USA/
NIH intramural program.
References
[1] N.C. Berchtold, G. Chinn, M. Chou, J.P. Kesslak, C.W. Cotman,
Exercise primes a molecular memory for brain-derived neurotrophic
factor protein induction in the rat hippocampus, Neuroscience 133
(2005) 853–861.
[2] D.S. Bredt, R.A. Nicoll, AMPA receptor trafficking at excitatory
synapses, Neuron 40 (2003) 361–379.
[3] R.K. Carlin, D.J. Grab, R.S. Cohen, P. Siekevitz, Isolation and
characterization of postsynaptic densities from various brain-
regions—Enrichment of different types of postsynaptic densities,
J. Cell Biol. 86 (1980) 831–843.
[4] E. Carro, J.L. Trejo, S. Busiguina, I. Torres-Aleman, Circulating
insulin-like growth factor I mediates the protective effects of
physical exercise against brain insults of different etiology and
anatomy, J. Neurosci. 21 (2001) 5678–5684.
[5] G.L. Collingridge, J.T. Isaac, Functional roles of protein interactions
with AMPA and kainate receptors, Neurosci. Res. 47 (2003) 3–15.
[6] C.W. Cotman, N.C. Berchtold, Exercise: a behavioral intervention to
enhance brain health and plasticity, Trends Neurosci. 25 (2002)
295–301.
[7] A. Czurko, H. Hirase, J. Csicsvari, G. Buzsaki, Sustained activation of
hippocampal pyramidal cells by Fspace clamping_ in a running wheel,
Eur. J. Neurosci. 11 (1999) 344–352.
[8] M. Di Luca, F. Gardoni, A. Finardi, S. Pagliardini, F. Cattabeni, G.
Battaglia, C. Missale, NMDA receptor subunits are phosphorylated by
activation of neurotrophin receptors in PSD of rat spinal cord,
NeuroReport 12 (2001) 1301–1305.
[9] H. Dong, R.J. O’Brien, E.T. Fung, A.A. Lanahan, P.F. Worley, R.L.
Huganir, GRIP: a synaptic PDZ domain-containing protein that
interacts with AMPA receptors, Nature 386 (1997) 279–284.
[10] E.P. Garcia, S. Mehta, L.A. Blair, D.G. Wells, J. Shang, T. Fukushima,
J.R. Fallon, C.C. Garner, J. Marshall, SAP90 binds and clusters
kainate receptors causing incomplete desensitization, Neuron 21
(1998) 727–739.
[11] G.S. Griesbach, F. Gomez-Pinilla, D.A. Hovda, The upregulation of
plasticity-related proteins following TBI is disrupted with acute
voluntary exercise, Brain Res. 1016 (2004) 154–162.
[12] H. Hirbec, J.C. Francis, S.E. Lauri, S.P. Braithwaite, F. Coussen, C.
Mulle, K.K. Dev, V. Coutinho, G. Meyer, J.T. Isaac, G.L. Colling-
ridge, J.M. Henley, V. Couthino, Rapid and differential regulation of
AMPA and kainate receptors at hippocampal mossy fibre synapses by
PICK1 and GRIP, Neuron 37 (2003) 625–638.
[13] I. Izquierdo, J.H. Medina, Memory formation: the sequence of
biochemical events in the hippocampus and its connection to activity
in other brain structures, Neurobiol. Learn Mem. 68 (1997) 285–316.
[14] S.S. Jayakar, M. Dikshit, AMPA receptor regulation mechanisms:
future target for safer neuroprotective drugs, Int. J. Neurosci. 114
(2004) 695–734.
[15] D.H. Jones, A.I. Matus, Isolation of synaptic plasma membrane from
brain by combined flotation–sedimentation density gradient centrifu-
gation, Biochim. Biophys. Acta 356 (1974) 276–287.
[16] T. Kitamura, M. Mishina, H. Sugiyama, Enhancement of neurogenesis
by running wheel exercises is suppressed in mice lacking NMDA
receptor epsilon 1 subunit, Neurosci. Res. 47 (2003) 55–63.
[17] R. Lamprecht, J. LeDoux, Structural plasticity and memory, Nat. Rev.,
Neurosci. 5 (2004) 45–54.
[18] J.O. Larsen, M. Skalicky, A. Viidik, Does long-term physical exercise
counteract age-related Purkinje cell loss? A stereological study of rat
cerebellum, J. Comp. Neurol. 428 (2000) 213–222.
[19] A.S. Leonard, M.A. Davare, M.C. Horne, C.C. Garner, J.W. Hell,
SAP97 is associated with the alpha-amino-3-hydroxy-5-methylisox-
azole-4-propionic acid receptor GluR1 subunit, J. Biol. Chem. 273
(1998) 19518–19524.
[20] J. Lerma, Roles and rules of kainate receptors in synaptic transmission,
Nat. Rev., Neurosci. 4 (2003) 481–495.
[21] E.S. Levine, J.E. Kolb, Brain-derived neurotrophic factor increases
activity of NR2B-containing N-methyl-d-aspartate receptors in
excised patches from hippocampal neurons, J. Neurosci. Res. 62
(2000) 357–362.
[22] E.S. Levine, R.A. Crozier, I.B. Black, M.R. Plummer, Brain-derived
neurotrophic factor modulates hippocampal synaptic transmission by
increasing N-methyl-d-aspartic acid receptor activity, Proc. Natl.
Acad. Sci. U. S. A. 95 (1998) 10235–10239.
[23] D.N. Lieberman, I. Mody, Regulation of NMDA channel function
by endogenous Ca(2+)-dependent phosphatase, Nature 369 (1994)
235–239.
[24] C. Limatola, Neurotrophic effects of AMPA, Cerebellum 3 (2004)
2–10.
[25] S.Y. Lin, K. Wu, E.S. Levine, H.T.J. Mount, P.C. Suen, I.B. Black,
BDNF acutely increases tyrosine phosphorylation of the NMDA
receptor subunit 2B in cortical and hippocampal postsynaptic
densities, Mol. Brain Res. 55 (1998) 20–27.
[26] I. Liste, M.J. Guerra, H.J. Caruncho, J.L. Labandeira-Garcia, Tread-
mill running induces striatal Fos expression via NMDA glutamate and
dopamine receptors, Exp. Brain Res. 115 (1997) 458–468.
[27] R.C. Malenka, Synaptic plasticity and AMPA receptor trafficking,
Ann. N. Y. Acad. Sci. 1003 (2003) 1–11.
[28] N.J. Maragakis, J.D. Rothstein, Glutamate transporters: animal models
to neurologic disease, Neurobiol. Dis. 15 (2004) 461–473.
[29] N.J. Maragakis, J.D. Rothstein, Glutamate transporters: animal models
to neurologic disease, Neurobiol. Dis. 15 (2004) 461–473.
[30] S. Mehta, H. Wu, C.C. Garner, J. Marshall, Molecular mechanisms

M.O. Dietrich et al. / Brain Research 1065 (2005) 20–25 25
regulating the differential association of kainate receptor subunits with
SAP90/PSD-95 and SAP97, J. Biol. Chem. 276 (2001) 16092–16099.
[31] B.S. Meldrum, Glutamate as a neurotransmitter in the brain: review of
physiology and pathology, J. Nutr. 130 (2000) 1007S–1015S.
[32] S.A. Neeper, J. Goauctemez-Pinilla, J. Choi, C. Cotman, Exercise and
brain neurotrophins, Nature 12 (6510) (1995) 109, (373).
[33] C.W. Nogueira, J.B. Rocha, D.O. Souza, Effect of dithiol chelating
agents on[3H]MK-801 and[3H]glutamate binding to synaptic plasma
membranes, Neurochem. Res. 26 (2001) 1305–1310.
[34] R.J. O’Brien, L.F. Lau, R.L. Huganir, Molecular mechanisms of
glutamate receptor clustering at excitatory synapses, Curr. Opin.
Neurobiol. 8 (1998) 364–369.
[35] S. Ozawa, H. Kamiya, K. Tsuzuki, Glutamate receptors in the ma-
mmalian central nervous system, Prog. Neurobiol. 54 (1998) 581–618.
[36] P. Perestenko, M.C. Ashby, J.M. Henley, Real-time imaging of alpha-
amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptor
(AMPA receptor) movements in neurons, Biochem. Soc. Trans. 31
(2003) 880–884.
[37] G.L. Peterson, A simplification of the protein assay method of Lowry
et al., which is more generally applicable, Anal. Biochem. 83 (1977)
346–356.
[38] L.O. Porciuncula, L. Vinade, S. Wofchuk, D.O. Souza, Guanine
based purines inhibit[3H]glutamate and[3H]AMPA binding at
postsynaptic densities from cerebral cortex of rats, Brain Res.
928 (2002) 106–112.
[39] K. Prybylowski, R.J. Wenthold, N-methyl-d-aspartate receptors:
subunit assembly and trafficking to the synapse, J. Biol. Chem. 279
(2004) 9673–9676.
[40] M. Ramsden, N.C. Berchtold, J. Patrick Kesslak, C.W. Cotman, C.J.
Pike, Exercise increases the vulnerability of rat hippocampal neurons
to kainate lesion, Brain Res. 971 (2003) 239–244.
[41] M.W. Salter, L.V. Kalia, Src kinases: a hub for NMDA receptor
regulation, Nat. Rev., Neurosci. 5 (2004) 317–328.
[42] R.H. Scannevin, R.L. Huganir, Postsynaptic organization and regu-
lation of excitatory synapses, Nat. Rev., Neurosci. 1 (2000) 133–141.
[43] M. Sheng, S.H. Lee, AMPA receptor trafficking and synaptic
plasticity: major unanswered questions, Neurosci. Res. 46 (2003)
127–134.
[44] M. Sheng, D.T. Pak, Glutamate receptor anchoring proteins and the
molecular organization of excitatory synapses, Ann. N. Y. Acad. Sci.
868 (1999) 483–493.
[45] P.C. Suen, K. Wu, E.S. Levine, H.T.J. Mount, J.L. Xu, S.Y. Lin, I.B.
Black, Brain-derived neurotrophic factor rapidly enhances phosphor-
ylation of the postsynaptic N-methyl-d-aspartate receptor subunit 1,
Proc. Natl. Acad. Sci. U. S. A. 94 (1997) 8191–8195.
[46] R.A. Swanson, W. Ying, T.M. Kauppinen, Astrocyte influences on
ischemic neuronal death, Curr. Mol. Med. 4 (2004) 193–205.
[47] J.L. Tillerson, W.M. Caudle, M.E. Reveron, G.W. Miller, Exercise
induces behavioral recovery and attenuates neurochemical deficits in
rodent models of Parkinson’s disease, Neuroscience 119 (2003)
899–911.
[48] H. van Praag, G. Kempermann, F.H. Gage, Running increases cell
proliferation and neurogenesis in the adult mouse dentate gyrus, Nat.
Neurosci. 2 (1999) 266–270.
[49] H. van Praag, B.R. Christie, T.J. Sejnowski, F.H. Gage, Running
enhances neurogenesis, learning, and long-term potentiation in mice,
Proc. Natl. Acad. Sci. U. S. A. 96 (1999) 13427–13431.
[50] S. Vaynman, Z. Ying, F. Gomez-Pinilla, Interplay between brain-
derived neurotrophic factor and signal transduction modulators in the
regulation of the effects of exercise on synaptic-plasticity, Neuro-
science 122 (2003) 647–657.
[51] L. Vinade, M. Chang, M.L. Schlief, J.D. Petersen, T.S. Reese, J.H.
Tao-Cheng, A. Dosemeci, Affinity purification of PSD-95-containing
postsynaptic complexes, J. Neurochem. 87 (2003) 1255–1261.
[52] J. Vissing, M. Andersen, N.H. Diemer, Exercise-induced changes in
local cerebral glucose utilization in the rat, J. Cereb. Blood Flow
Metab. 16 (1996) 729–736.
[53] Y.T. Wang, X.M. Yu, M.W. Salter, Ca(2+)-independent reduction of
N-methyl-d-aspartate channel activity by protein tyrosine phospha-
tase, Proc. Natl. Acad. Sci. U. S. A. 93 (1996) 1721–1725.
[54] R.Y. Wang, Y.R. Yang, S.M. Yu, Protective effects of treadmill
training on infarction in rats, Brain Res. 922 (2001) 140–143.
Copyright © 2022 FDOKUMEN




















