
AChR/ϵ-AChR Switch at Agrin-Induced Postsynaptic-like Apparatus in Skeletal Muscle
10
g-AChR/e-AChR Switch at Agrin-Induced Postsynaptic-like Apparatus in Skeletal Muscle Mendell Rimer,* Iacob Mathiesen, ² Terje Lømo, ² and U. J. McMahan* *Department of Neurobiology, Stanford University School of Medicine, Stanford, California 94305; and ² Department of Neurophysiology, University of Oslo, Oslo, Norway We transfected the extrajunctional region of denervated soleus muscles in adult rats with neural agrin cDNA to induce myofibers to form postsynaptic-like apparatus containing acetylcholine receptor (AChR) aggregates. By 1 week D30% of the AChR aggregates contained a mixture of e-AChRs and g-AChRs while D70% had only g-AChRs. If the transfected muscles were reinnervated in the origi- nal junctional region, the postsynaptic-like apparatus, despite the absence of apposed axon terminals, gradually came to have only e-AChRs. We conclude that at the postsynaptic apparatus of ectopic neuromuscular junc- tions formed by a foreign nerve implanted into the extra- junctional region of denervated muscles, agrin secreted by the axon terminal plays a direct role in the g-AChR/e- AChR switch that occurs as the apparatus reaches matu- rity. Our findings, together with results from other studies, indicate further that agrin and acetylcholine are the only nerve-derived factors required for this switch. INTRODUCTION The subunit composition of acetylcholine receptors (AChRs) that first aggregate in the postsynaptic appara- tus at developing neuromuscular junctions in the mam- malian embryo is a2bgd. As the junctions mature, these AChRs are gradually replaced by receptors composed of a2bed which have different channel conductance prop- erties (Fischbach and Schuetze, 1980; Mishina et al., 1986). A similar g-AChR/e-AChR switch occurs at ectopic neuromuscular junctions that form when the central stump of a foreign nerve is implanted into the extrajunctional region of denervated muscles in adult rats (Brenner et al., 1987, 1994). In both cases the switch is induced by the axon terminal; electrical activity in the muscle fibers generated by the interaction of the neuro- transmitter acetylcholine with g-AChRs blocks the gene expression of the g-subunit while another axon released factor(s) upregulates the local expression of the e-sub- unit gene which is apparently unaffected by electrical activity (reviewed in Duclert and Changeux, 1995). Because of their accessibility for electrophysiological experiments, the ectopic neuromuscular junctions have been particularly useful for examining mechanisms involved in the formation of the AChR aggregates (e.g., Lømo and Slater, 1978, 1980a; Brenner et al., 1987, 1994; Lømo et al., 1988; Rotzler and Brenner, 1990). This study concerns the number and nature of axon- provided factors that in addition to acetylcholine induce the switch from g-AChRs to e-AChRs at ectopic neuro- muscular junctions formed in the denervated soleus muscles of the adult rat. As demonstrated in the accom- panying report (Cohen et al., 1997) we found that if cDNA encoding an agrin isoform specific to neurons, agrin-Y4Z8, was injected into the extrajunctional region of these muscles, it was taken up by some of the muscle fibers. The neural agrin was expressed at histochemi- cally detectable levels in the sarcoplasm of such fibers and secreted into the extracellular space where it acted on agrin receptors in the plasma membrane of these and nearby untransfected fibers, causing them to form on their surface postsynaptic-like apparatus. The appara- tus was apparent by 1 week after cDNA injection and was still evident, although at a lower frequency, after 6 weeks. As found in the postsynaptic apparatus of normal and ectopic neuromuscular junctions, the post- synaptic-like apparatus contained extracellular, plasma membrane, and cytoplasmic protein aggregates, plasma membrane infoldings, and accumulations of myonuclei. Such findings are consistent with the agrin hypothesis (Nitkin et al., 1987; McMahan, 1990; see also Reist et al., 1992; DeChaira et al., 1996; Gautam et al., 1996), which predicts that agrin released by the axon terminal in- duces the aggregation of postsynaptic proteins during MCN Molecular and Cellular Neuroscience 9, 254–263 (1997) Article No. CN970622 254 1044-7431/97 $25.00 Copyright r 1997 by Academic Press All rights of reproduction in any form reserved.
Transcript of AChR/ϵ-AChR Switch at Agrin-Induced Postsynaptic-like Apparatus in Skeletal Muscle

g-AChR/e-AChR Switch at Agrin-InducedPostsynaptic-like Apparatus in Skeletal Muscle
Mendell Rimer,* Iacob Mathiesen,† Terje Lømo,†
and U. J. McMahan**Department of Neurobiology, Stanford University School of Medicine, Stanford, California94305; and †Department of Neurophysiology, University of Oslo, Oslo, Norway
We transfected the extrajunctional region of denervatedsoleus muscles in adult rats with neural agrin cDNA toinduce myofibers to form postsynaptic-like apparatuscontaining acetylcholine receptor (AChR) aggregates. By1 week D30% of the AChR aggregates contained a mixtureof e-AChRs and g-AChRs while D70% had only g-AChRs.If the transfected muscles were reinnervated in the origi-nal junctional region, the postsynaptic-like apparatus,despite the absence of apposed axon terminals, graduallycame to have only e-AChRs. We conclude that at thepostsynaptic apparatus of ectopic neuromuscular junc-tions formed by a foreign nerve implanted into the extra-junctional region of denervated muscles, agrin secretedby the axon terminal plays a direct role in the g-AChR/e-AChR switch that occurs as the apparatus reaches matu-rity. Our findings, together with results from other studies,indicate further that agrin and acetylcholine are the onlynerve-derived factors required for this switch.
INTRODUCTION
The subunit composition of acetylcholine receptors(AChRs) that first aggregate in the postsynaptic appara-tus at developing neuromuscular junctions in the mam-malian embryo is a2bgd. As the junctions mature, theseAChRs are gradually replaced by receptors composed ofa2bed which have different channel conductance prop-erties (Fischbach and Schuetze, 1980; Mishina et al.,1986). A similar g-AChR/e-AChR switch occurs atectopic neuromuscular junctions that form when thecentral stump of a foreign nerve is implanted into theextrajunctional region of denervated muscles in adultrats (Brenner et al., 1987, 1994). In both cases the switchis induced by the axon terminal; electrical activity in themuscle fibers generated by the interaction of the neuro-transmitter acetylcholine with g-AChRs blocks the geneexpression of the g-subunit while another axon released
factor(s) upregulates the local expression of the e-sub-unit gene which is apparently unaffected by electricalactivity (reviewed in Duclert and Changeux, 1995).Because of their accessibility for electrophysiologicalexperiments, the ectopic neuromuscular junctions havebeen particularly useful for examining mechanismsinvolved in the formation of the AChR aggregates (e.g.,Lømo and Slater, 1978, 1980a; Brenner et al., 1987, 1994;Lømo et al., 1988; Rotzler and Brenner, 1990).This study concerns the number and nature of axon-
provided factors that in addition to acetylcholine inducethe switch from g-AChRs to e-AChRs at ectopic neuro-muscular junctions formed in the denervated soleusmuscles of the adult rat. As demonstrated in the accom-panying report (Cohen et al., 1997) we found that ifcDNA encoding an agrin isoform specific to neurons,agrin-Y4Z8, was injected into the extrajunctional regionof these muscles, it was taken up by some of the musclefibers. The neural agrin was expressed at histochemi-cally detectable levels in the sarcoplasm of such fibersand secreted into the extracellular space where it actedon agrin receptors in the plasma membrane of these andnearby untransfected fibers, causing them to form ontheir surface postsynaptic-like apparatus. The appara-tus was apparent by 1 week after cDNA injection andwas still evident, although at a lower frequency, after 6weeks. As found in the postsynaptic apparatus ofnormal and ectopic neuromuscular junctions, the post-synaptic-like apparatus contained extracellular, plasmamembrane, and cytoplasmic protein aggregates, plasmamembrane infoldings, and accumulations of myonuclei.Such findings are consistent with the agrin hypothesis(Nitkin et al., 1987; McMahan, 1990; see also Reist et al.,1992; DeChaira et al., 1996; Gautam et al., 1996), whichpredicts that agrin released by the axon terminal in-duces the aggregation of postsynaptic proteins during
MCN Molecular and Cellular Neuroscience 9, 254–263 (1997)
Article No. CN970622
254
1044-7431/97 $25.00Copyright r 1997 by Academic Press
All rights of reproduction in any form reserved.

the formation of ectopic neuromuscular junctions in theadult as well as at developing junctions in the embryo,and they indicate further that neural agrin alone may besufficient to induce much if not all of the other structuralspecializations in the developing postsynaptic appara-tus. Among the proteins aggregated in the agrin-induced postsynaptic-like apparatus are AChRs. In thisstudy, we stained AChR aggregates in the postsynaptic-like apparatus in cross sections of denervated andreinnervated soleus muscles with both a-bungarotoxin,which marked all AChR aggregates, and antiserum tog-AChRs or e-AChRs to determine by fluorescencehistochemistry whether the aggregates exhibit ag-AChR/e-AChR switch. Our results indicate that indenervated muscles all such aggregates initially haveg-AChRs while some also have e-AChRs. If the musclesbecome reinnervated at the original synaptic sites,g-AChRs are eliminated from the agrin-induced AChRaggregates, as they are from the AChR aggregates atsynaptic sites so that by 4 weeks the aggregates in bothcases have only e-AChRs. A report that bears on thefindings presented here has recently appeared else-where (Jones et al., 1997); see Discussion.
RESULTS
All muscles used for this study were initially dener-vated. Three days after denervation they were injectedwith cDNA encoding rat neural agrin Y4Z8. Previousstudies on such muscles (Cohen et al., 1997) have shownthat by 7 days after injection (10 days after denervation),myofibers have expressed the neural agrin, and thenumber of agrin-induced AChR aggregates has reachedits maximum level. About 1% of the muscle fibersbecome transfected and they continue to express theneural agrin for more than 6 weeks after injection. Thereis a gradual attrition in the number of AChR aggregatesin denervated muscles so that by 6 weeks it is 25% ofthat at 1 week.The postsynaptic apparatus in denervated muscles
persists for more than a month in the absence of axonterminals (e.g., Fambrough, 1979). However within afew days after denervation, owing to the absence ofelectrical activity in the muscle fibers, g-AChRs areexpressed and accumulate in the postsynaptic appara-tus along with e-AChRs (Gu and Hall, 1988). When thenerve is crush-damaged, allowing the regenerating axonsto reestablish neuromuscular activity at the originalsynaptic sites (Tello, 1917; Letinsky et al., 1976), the
postsynaptic apparatus again comes to have onlye-AChRs (Witzemann et al., 1991). There is evidence(Cohen et al., 1997) that in transfected muscles like thosewe used for this study, some myofibers are focallydamaged during cDNA injection. Although repair isapparently rapid, we were uncertain what effect suchdamage and the overexpression of agrin might have onthe expression of g-AChRs and e-AChRs in transfectedmuscles following denervation and reinnervation. Fig-ure 1a–1l demonstrates that the AChR subunit composi-tion of postsynaptic apparatus in denervated and crush-denervated/reinnervated soleus muscles transfectedwith agrin cDNA was the same as that reported foruninjected muscles (Gu and Hall, 1988; Witzemann etal., 1991). Thus, neither injecting nor transfecting soleusmuscle had an obvious effect on the g-AChR/e-AChRswitches at the original postsynaptic apparatus.As shown in Figs. 1n, 1t, and 2, by 10 days after
denervating the muscles in a way that inhibited reinner-vation, all of the AChR aggregates induced by thesecreted neural agrin had g-AChRs. About one-third ofthese aggregates also had relatively low levels ofe-AChRs (Figs. 1m, 1s, and 2). Thus, neural agrin aloneis sufficient to induce a postsynaptic-like apparatushaving both g-AChRs and e-AChRs as found at thepostsynaptic apparatus in denervated adult muscles.Electrophysiological studies have revealed that when
the soleus nerve is crushed near its entry into themuscle, as we did for these studies, the regeneratingaxons begin to reestablish neuromuscular transmissionabout 10 days after the crush (Andreose et al., 1993). By17 days reinnervation is nearly complete. When wetreated cross sections of transfected muscles 17 daysafter crush-denervation with antibody for synapsin I, tomark axon terminals (Greengard et al., 1993), all of thepostsynaptic apparatus we observed by staining AChRaggregates with Texas red a-bungarotoxinwere coexten-sive with synapsin I staining (data not shown). Thus,neither injection nor transfection had an obvious effecton the timing of reinnervation of the soleus musclesafter nerve crush.Most of the neural agrin-induced AChR aggregates at
17–24 days after crush-denervation/reinnervation had amixture of g-AChRs and e-AChRs (Figs. 1o, 1p, 1u, 1v,and 2) as did most of theAChR aggregates at neuromus-cular junctions at this time (Figs. 1c, 1d, 1i, and 1j). At45–66 days, when AChR aggregates at the neuromuscu-lar junctions had only e-AChRs (Figs. 1e, 1f, 1k, and 1l)so did the agrin-induced AChR aggregates (Figs. 1q, 1r,1w, 1x, and 2).
g-AChR/e-AChR Switch 255
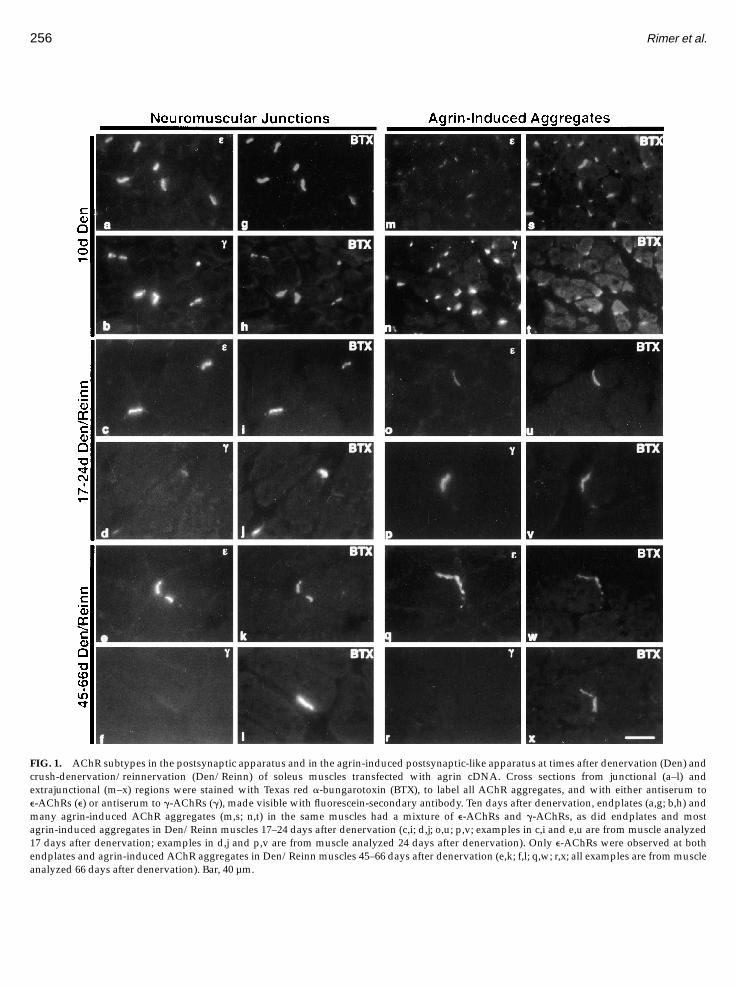
FIG. 1. AChR subtypes in the postsynaptic apparatus and in the agrin-induced postsynaptic-like apparatus at times after denervation (Den) andcrush-denervation/reinnervation (Den/Reinn) of soleus muscles transfected with agrin cDNA. Cross sections from junctional (a–l) andextrajunctional (m–x) regions were stained with Texas red a-bungarotoxin (BTX), to label all AChR aggregates, and with either antiserum toe-AChRs (e) or antiserum to g-AChRs (g), made visible with fluorescein-secondary antibody. Ten days after denervation, endplates (a,g; b,h) andmany agrin-induced AChR aggregates (m,s; n,t) in the same muscles had a mixture of e-AChRs and g-AChRs, as did endplates and mostagrin-induced aggregates in Den/Reinn muscles 17–24 days after denervation (c,i; d,j; o,u; p,v; examples in c,i and e,u are from muscle analyzed17 days after denervation; examples in d,j and p,v are from muscle analyzed 24 days after denervation). Only e-AChRs were observed at bothendplates and agrin-inducedAChR aggregates in Den/Reinn muscles 45–66 days after denervation (e,k; f,l; q,w; r,x; all examples are frommuscleanalyzed 66 days after denervation). Bar, 40 µm.
256 Rimer et al.

In one set of experiments we took eight or more serialsections from the transfected region of crush-dener-vated/reinnervated muscles 17–24 days after nervedamage and from chronically denervated muscles 10and 17 days after nerve damage. Each was stained withTexas red a-bungarotoxin to label all AChR aggregatesand with antibody Agrin-86 to label the myofibersexpressing neural agrin (Cohen et al., 1997). We foundthat the number of neural agrin transfected fibers percross section for crush-denervated/reinnervated andchronically denervated muscles varied within only an,2-fold range (19.1–8.5; 12.1 6 2.5, mean 6 SEM,musclen 5 6). However, the number of AChR aggregates percross section in the crush-denervated/reinnervatedmuscles (2.4 6 0.18, mean 6 SEM, muscle n 5 3) was,50-fold lower than in 10-day denervated muscles (102,muscle n 5 2; 119.1, 84.5) and ,20-fold lower than in17-day denervated muscles (50 6 13, mean 6 SEM,muscle n 5 3). Since functional reinnervation of thecrush-denervated muscles began ,10 days after nervedamage, these results indicate that reinnervation bringsabout a marked reduction in the number of agrin-induced AChR aggregates. This reduction accounts forthe lower number of total agrin-induced AChR aggre-gates counted for reinnervated muscles in Fig. 2.
We checked muscles 45–66 days after crush-denerva-tion/reinnervation to see whether or not axons hadsprouted from the reinnervated original synaptic sites toinnervate the agrin-induced postsynaptic-like appara-tus by staining for the axon terminals’ synaptic vesicleprotein SV2 (Buckley and Kelly, 1985) and e-AChRs.While SV2 staining was coextensive with AChR aggre-gates in the postsynaptic apparatus at the several tens oforiginal synaptic sites examined, it was not observed atany of the AChR aggregates of the agrin-induced post-synaptic-like apparatus (Fig. 3). Identical results wereobtained when serial cross sections were stained forsynapsin I and a-bungarotoxin (not shown). Thus, theg-AChR/e-AChR switch in the postsynaptic-like appa-ratus in reinnervated muscles occurred in the absence ofa directly apposed axon terminal.We also checked muscles chronically denervated for
at least 4 weeks to determine the nature of the AChRs inagrin-induced aggregates in long-term inactive muscles.Interestingly,,82% of Texas red a-bungarotoxin stainedaggregates at these times also stained for e-AChR whileonly 10% also stained for g-AChR (Fig. 4). In each case,at least 200 aggregates in two different muscles werescored. All of the AChR aggregates at the sites of the
FIG. 2. Quantitative presentation of data from AChR aggregates in postsynaptic-like apparatus summarized in Fig. 1. The percentage of allAChR aggregates (labeled with Texas red a-bungarotoxin) that stained for either g-AChRs or e-AChRs was determined by counting the number ofagrin-induced aggregates under Texas red (all AChRs) or fluorescein (g-AChRs or e-AChRs) optics. Values presented as mean percentageco-localization 6 range (highest–lowest value). Underlined number above bars represents the total number of AChR aggregates counted for eachcase. Muscle n 5 2, except for those in which e-AChRs were counted 17–24 days after denervation, muscle n 5 3.
g-AChR/e-AChR Switch 257

former neuromuscular junctions in the same musclestained for both e-AChR and g-AChR (data not shown).
DISCUSSION
Our previous studies on the postsynaptic-like appara-tus in denervated adult soleus muscles transfected withneural agrin cDNA indicate that agrin, secreted by theaxon terminals, plays a crucial role in the formation ofthe postsynaptic apparatus at ectopic neuromuscularjunctions by directing the assembly of most, if not all,postsynaptic structures (Cohen et al., 1997). In theexperiments described here, by 10 days after denervat-ing soleus muscles and 7 days after injecting them withneural agrin cDNA, all of the AChR aggregates in theagrin-induced postsynaptic-like apparatus had g-AChRswhile ,30% also had e-AChRs. If the muscles werereinnervated at the original synaptic sites, 1–2 weekslater there was an ,50-fold reduction in the number ofagrin-induced AChR aggregates, at least ,20-fold ofwhich was due to reinnervation rather than spontane-ous attrition, but of those that persisted ,70% had amixture of g-AChRs and e-AChRs while the remainderhad only g-AChRs. By 4 weeks after reinnvervation allof the AChR aggregates in postsynaptic-like apparatus,despite the absence of a directly apposed axon terminal,
had only e-AChRs. Altogether, these results indicatethat at the postsynaptic apparatus of ectopic neuromus-cular junctions, agrin secreted by the axon terminal alsoplays a direct role in the g-AChR/e-AChR switch as theapparatus matures.As we were preparing this paper for publication,
Jones et al. (1997) reported that expression of chickneural agrin in the extrajunctional region of soleusmuscles of rat and mouse was accompanied by theexpression of the e-AChR subunit gene and the forma-tion of e-AChR aggregates on the surface of musclefibers. However, it is not clear from such experimentswhether the response was due to chick agrin actingdirectly or alone on myofibers. As we point out else-where (Cohen et al., 1997), the expression of chick agrinin rat muscle can generate a substantial inflammatoryresponse in the region of transfection. Although theiranimals were fed an immunosuppressant, there wasneither acknowledgment of the problem nor assessmentof whether the immunosuppressant had any effect on
FIG. 3. Regenerated axons are confined to the junctional region.Cross sections from junctional (a,b) and extrajunctional region (c,d) ofa crush-denervated/reinnervated transfected muscle 66 days afterdenervation. Each was stained with a mouse monoclonal antibody toSV2, made visible with a Texas-red anti-mouse secondary antibodyand antiserum to e-AChRs visualized with a fluorescein anti-rabbitsecondary antibody. SV2 staining is co-localized with e-AChR stainingat the reinnervated neuromuscular junctions (a,b) while there is noSV2 staining at the agrin-induced e-AChR aggregate (c,d). Bar, 40 µm.
FIG. 4. AChR subtypes in agrin-induced postsynaptic-like appara-tus after a long period of chronic denervation. Cross sections from thetransfected region of 31-day denervated soleus muscles were stainedfor either e-AChRs (a) or g-AChRs (b) and with Texas red-a-bungarotoxin (c,d) as described in the legend to Fig. 1. All AChRaggregates in these examples stained strongly for e-AChRs andweakly or not at all for g-AChRs. Bar, 40 µm.
258 Rimer et al.

the inflammation. Muscle inflammation, alone, can in-crease the expression of AChRs (reviewed in Gordon etal., 1976). Such inflammation might explain, for ex-ample, their paradoxical but undiscussed observationthat innervated rat muscles expressing chick agrin showthe sameAChR response as denervated rat muscles. Theextrajunctional region of normal muscles do not havedetectable levels of MuSK, a tyrosine kinase whichmediates agrin’s AChR aggregating activity, and theydo not show any AChR response to agrin provided bythe axons of an implanted foreign nerve (Valenzuela etal., 1995; DeChiara et al., 1996; Cohen et al., 1997). Analternative explanation may be that the innervatedmuscles were damaged during penetration by theircDNA pipet; muscle fiber damage by itself can increaseAChR expression (Lømo and Westgaard, 1975; Gold-man et al., 1991; Brenner et al., 1992). Our experiments,which relied on the expression of noninflammatory ratneural agrin as well as a variety of other controls, leavelittle doubt that neural agrin acted directly and alone onmuscle fibers to induce the extrajunctional aggregationofAChRs in this study (Cohen et al., 1997). Furthermore,we sought to determine whether there is a g-AChR/e-AChR switch in agrin-induced postsynaptic-like appara-tus as in the postsynaptic apparatus at neuromuscularjunctions by examining both denervated and reinner-vated muscles for prolonged periods after transfectionand by using probes specific for g-AChR as well ase-AChR. This enabled us to avoid any transient effects ofdamage caused by penetration of the muscle with thecDNA-containing pipet, and it made it possible toexamine the time course and stability of the switch inthe presence and absence of muscle activity.mRNAs for the g- and e-AChR subunits in extrajunc-
tional regions of normal soleus muscles, as in othermuscles, are undetectable by in situ hybrization (Witze-mann et al., 1991; Kues et al., 1995). Upon denervation ofsoleus muscles, the extrajunctional region uniformlyexhibits readily detectable levels of g-subunit mRNAand marginally detectable levels of e-subunit mRNA(Witzemann et al., 1991), although for the extrajunc-tional region of other denervated muscles higher levelsof e-subunit mRNA have been observed (Witzemann etal., 1987). Accordingly, the g-AChRs and e-AChRs thatwe found in the neural agrin-induced postsynaptic-likeapparatus in denervated myofibers and in denervated/reinnervatedmyofibers shortly after reinnervation couldhave been derived, at least in part, from the uniformexpression and the lateral migration of theAChRs in theplasma membrane. Such lateral migration is a knownmechanism of agrin-inducedAChR aggregation inmyo-tube cultures (Godfrey et al., 1984). However, several
lines of evidence indicate that after electrical activity indenervated muscles is restored by reinnervation, theextrajunctional expression of AChRs returns to normallevels (Lømo andWestgaard, 1975; Goldman et al., 1988;Witzemann et al., 1991; Brenner et al., 1992). Our findingthat AChR aggregates at both postsynaptic apparatusand agrin-induced postsynaptic-like apparatus in rein-nervatedmuscles contained g-AChRs aswell as e-AChRsuntil at least 24 days after the nerve was crushedindicates that normal levels of neural inhibition of theuniform expression of g-subunit and extrajunctionale-subunit were not reestablished until after that time.The absence of g-AChRs in aggregates at 45 daysindicates further that such expression had ceased bythen as did any lateral migration of either g- or e-AChRsfrom beyond the immediate vicinity of the postsynaptic-like apparatus. Accepting a half-life of 10 days fore-AChRs in the postsynaptic-like apparatus, as found atthe postsynaptic apparatus in normal, denervated/reinnervated or denervated/electrically stimulatedmuscles (Andreose et al., 1993), a single exponentialequation predicts that at least 88% of the e-AChRs weobserved at the agrin-induced postsynaptic-like appara-tus in reinnervated muscles 66 days after the nervecrushwere derived from the local expression of e-AChRsthat had begun at 45 days. That a local transcriptionalmechanism is likely to mediate this induction is sug-gested by the finding that there is a 3.5-fold increase inAChR e-subunit transcripts after injecting muscle celllines permanently transfected with chick agrin Y4Z19into denervated mouse soleus muscles (Jones et al.,1997).The general appearance of muscle fibers in the dener-
vated/reinnervated muscles injected with agrin cDNAwas not obviously different from that of muscle fibers inuninjected denervated/reinnervated muscles or in nor-mal muscles. However, in transfected muscles inspected31 days or later after chronic denervation and transfec-tion, the muscle fibers were, as expected (e.g., Sola andMartin, 1953), severely atrophied from the lack ofelectrical activity. We noted that while there was amixture of g-AChRs and e-AChRs at the original postsyn-aptic apparatus of such muscles, there were onlye-AChRs at most of the agrin-induced postsynaptic-likeapparatus. Studies on g-subunit mRNA expression inthe extrajunctional region of atrophic soleus muscleshave demonstrated that it becomes much reduced be-low that in the junctional region (Kues et al., 1995),which probably accounts for the absence of g-AChRs inthe postsynaptic-like apparatus of these muscles. Thelong-term effect of atrophy on the extrajunctional expres-sion of the e-subunit in the soleus muscle is not known.
g-AChR/e-AChR Switch 259

However, Adams et al. (1995) have reported that whileexpression of g-subunit RNA is progressively reducedin tibialis anterior muscles chronically denervated forseveral months, expression of the e-subunit RNA re-mains elevated. Thus, in muscle fibers of severelyatrophied muscles, unlike those in normal and reinner-vated muscles, g-subunit expression is inhibited byactivity-independentmechanisms.Moreover, the persis-tence of e-AChRs in the postsynaptic-like apparatusunder atrophic conditions could be due to an agrin-induced lateral migration of e-AChRs uniformly ex-pressed in the extrajunctional region, as discussed above,as well as an agrin-induced increase in local expressionof the e-subunit.One mechanism by which agrin could bring about
local expression of e-AChRs at the agrin-induced post-synaptic-like apparatus is by inducing a mechanismthat enhances gene expression in local nuclei. Severallines of evidence suggest that, at developing neuromus-cular junctions, neuregulin in the synaptic cleft acts byway of erbB receptors in the postsynaptic apparatus toincrease the local gene expression of certain proteins(Martinou et al., 1991; Corfas and Fischbach, 1993; Fallset al., 1993; Jo et al., 1995; Chu et al., 1995; Altiok et al.,1995; Zhu et al., 1995; Missias et al., 1996). One suchfinding is that the application of neuregulin tomyotubesin culture results in a 10-fold increase in e-AChR subunitexpression (Martinou et al., 1991; Chu et al., 1995).Moreover, motor neurons and muscle fibers synthesizeand secrete neuregulin into the synaptic cleft at theneuromuscular junction (Falls et al., 1993; Goodearl et al.,1995; Moscoso et al., 1995; M. Rimer, I. Cohen, T. Lømo,S. J. Burden, and U. J. McMahan, in preparation). As wewill show elsewhere (M. Rimer, I. Cohen, T. Lømo, S. J.Burden, and U. J. McMahan, in preparation), muscleneuregulin and erbB2 and erbB3 receptors aggregate atthe agrin-induced postsynaptic-like apparatus in dener-vated transfected soleus muscles, suggesting that neuralagrin by itself induces the assembly of a neuregulin-based autocrine pathway for enhancing the gene expres-sion of e-AChRs.An additional mechanism by which agrin might
increase the local expression of e-AChRs is by inducingthe accumulation of protein-making machinery. At nor-mal mammalian neuromuscular junctions, in general,and ectopic neuromuscular junctions in the rat’s soleusmuscles, in particular, the postsynaptic apparatus in-cludes an accumulation of myonuclei and other organ-elles involved in the expression of AChRs (Korneliussenand Sommerschild, 1976; Brenner et al., 1994). Indeed,we and others have demonstrated elsewhere (Cohen etal., 1997; Jones et al., 1997) that at the postsynaptic-like
apparatus in denervated and innervated soleus musclesinjectedwith agrin cDNAthere are aggregates ofmyonu-clei. Because nuclei are always accompanied by otherorganelles, this finding indicates that agrin may inducethe accumulation of all postsynaptic organelles requiredfor a local increase in expression of proteins.The persistence of e-AChR aggregates in the postsyn-
aptic-like apparatus in reinnervated muscles after theuniform gene expression of the g- and e-AChR subunitsreturned to undetectable levels (Lømo and Westgaard,1975; Goldman et al., 1988; Witzemann et al., 1991;Brenner et al., 1992) could have resulted from agrininducing the local expression of the e-subunit gene in away that was independent of electrical activity. Alterna-tively, the axon terminals at the reinnervated originalpostsynaptic apparatus could have secreted unknownsubstances that resulted in the local expression byjunctional nuclei of diffusible factors required for mak-ing the e-AChR expression at the agrin-induced postsyn-aptic-like apparatus resistant to activity. In studies to bereported elsewhere (I. Mathiesen, M. Rimer, I. Cohen,U. J. McMahan, and T. Lømo, in preparation), we foundthat agrin-induced AChR aggregates in denervatedsoleus muscles made active by direct electrical stimula-tion can persist for more than 10 weeks after agrincDNA injection. This indicates that the axon terminal atectopic neuromuscular junctions that form in dener-vated adult muscles needs to provide only acetylcho-line, which suppresses g-subunit expression by generat-ing electrical activity in myofibers, and agrin, whichgenerates the local expression of the e-subunit that isresistant to electrical activity, to bring about the g-AChR/e-AChR switch in the postsynaptic apparatus.Axons of a foreign nerve implanted into the extrajunc-
tional region of denervated soleus muscles initially formnascent neuromuscular junctions at several sites on amuscle fiber. Over the next few weeks the number ismarkedly reduced so that only one or a few junctionsper myofiber persist and reach maturity. Muscle activityis likely to play an important role in this synapseelimination. For example, if the foreign axon terminalsare caused to degenerate prior to the elimination ofjunctions, direct electrical stimulation of the musclesresults in the elimination of former synaptic sites untilonly one or a few per fiber remain. Our finding that thenumber of agrin-induced aggregates in the extrajunc-tional regions of 17-day crush-denervated/reinnervatedsoleus muscles was ,20-fold less than in 17-day dener-vated muscles and ,50-fold less than in 10-day dener-vated muscles, together with the finding that someaggregates persist indefinitely, provides further evi-dence that agrin-induced AChR aggregates are similar
260 Rimer et al.

to those induced by axon terminals of a foreign nerve(Cohen et al., 1997). Indeed direct stimulation of dener-vated soleus muscles brings about the elimination ofagrin-induced AChR aggregates until only one or a fewper muscle fiber persist (I. Mathiesen, M. Rimer, I.Cohen, U. J. McMahan, and T. Lømo, in preparation).Thus, it may well be that acetylcholine and agrin are theonly signals required from the axon terminals in theestablishment of the ultimate number of neuromuscularjunctions per myofiber in ectopically innervated soleusmuscles.Myotubes raised in culture express higher levels of
e-AChR mRNA when they are grown on substratecontaining agrin than they do when grown on substratelacking it (Jones et al., 1996). The region of agrin thatinduces such expression in culture is apparently differ-ent from the region that induces aggregation of AChRsand many other proteins. For example, an agrin isoformlacking AChR-aggregating activity is as effective ininducing the increased expression of e-AChRs as anisoform which is known to have at least 104-fold greaterAChR-aggregating activity based on dose–responsemea-surements on soluble agrin inmyotube cultures (Jones etal., 1996). Myotube factors regulating the expression ofe-AChR mRNA levels may be different in vitro fromthose in vivo. Nevertheless, such findings raise forconsideration the possibility that in the postsynaptic-like apparatus induced to form in soleus muscles trans-fected with neural agrin cDNA, some region in theneural agrin other than that which triggers AChRaggregation and the assembly of other postsynapticcomponents, infoldings in the plasma membrane, andthe accumulation of organelles acts to induce the localexpression of e-AChRs.The lack of accessibility of embryonic muscles for in
vivo experiments has made it difficult to assess theextent of agrin’s role in the formation of the postsynap-tic apparatus in developing muscles beyond its beingessential for protein aggregation (DeChiara et al., 1996;Gautam et al., 1996). There is no reason to rule out thepossibility that as the postsynaptic apparatus matures indeveloping muscles neural agrin plays a role in theinduction of later events as we propose it does at ectopicneuromuscular junctions in adult muscles. However,because of different levels of protein expression betweendeveloping and adult muscle, neural agrin and acetyl-choline may not be sufficient to induce the g-AChR/e-AChR switch in embryos as they are in adults. Forexample, adult denervated muscles express low levelsof e-subunit mRNA while myotubes in developingmuscles do not express detectable levels of e-subunitmRNA until they become innervated (Brenner et al.,
1994). Thus, the axon terminal at developing neuromus-cular junctions may trigger e-subunit expression as wellas enhancing it and making it resistant to electricalactivity. It remains to be determined whether agrin orsome other neuron-derived factor is the trigger ofe-subunit expression. It may be possible to discernbetween these possibilities by producing transgenicmice that express neural agrin under the control of askeletal muscle-specific constitutive promoter. If neuralagrin triggers e-subunit expression, e-mRNA might bedetected in developing myotubes of such transgenicanimals before nerve–muscle contact is established.
EXPERIMENTAL METHODS
Animals, Denervation, and cDNA Injection
All experiments were done on adult male Wistar rats(,180 g). For surgery they were anesthesized withketamine (100–200 mg/kg ip) 1 chloral hydrate (300–600 mg/kg ip). Soleus muscles were denervated 3 daysbefore cDNA injection either by cutting out a 3- to 5-mmpiece of the sciatic nerve several centimeters away fromthe muscle, which prevented reinnervation, or by crush-ing with fine forceps the soleus nerve near the point ofits entry into the muscle, which allowed reinnervation.cDNA encoding rat agrin isoform Y4Z8 was injectedwith a fine glass capillary pipet inserted into the extra-junctional region about halfway between the neuromus-cular junctions and the proximal myotendon junctionsas described elsewhere (Cohen et al., 1997).
Antibodies and Immunohistochemistry
For examination of AChR aggregates by fluorescenceimmunohistochemistry muscles were pinned out ondental wax and frozen in isopentane cooled in liquid N2,and 10-µm serial cross sections were made through thetransfected region on a cryostat. Neural agrin wasstained with monoclonal antibody Agrin-86 (StressGen,Victoria, BC, Canada) as described in Cohen et al. (1997).g-AChR and e-AChR aggregates were stained withrabbit antisera raised against unique peptide sequencesof themouseAChR g and e-subunits, purified by affinitychromatographywith peptides that included part (anti-g)or the whole (anti-e) of the immunizing peptides. Theywere a kind gift of Dr. Josh Sanes and have beenpreviously described (Missias et al., 1996). The anti-gserum was used at 1/100 and the anti-e at 1/300. SV2was stained with hybridoma supernatant, used at 1/10,from cells producing monoclonal antibody (Buckley
g-AChR/e-AChR Switch 261

and Kelly, 1985). It was a kind gift of Dr. Kathy Buckley.Anti-synapsin I antiserum (574777), used at 1/1000, waspurchased from Calbiochem–Novabiochem Corp. (LaJolla, CA). For staining with all antibodies, except theanti-g-AChR, sections were fixed in 1% paraformalde-hyde for 5 min at room temperature, washed with PBScontaining 0.1% Triton X-100 (PBS-T), blocked for ,10min in PBS-T 10% normal goat serum (NGS), andincubated overnight at 4°C with primary antibodydiluted in PBS-T 10% NGS. The sections were washedwith PBS-T and incubated with fluorescein- or Texasred-conjugated secondary antibodies (Cappel OrganonTeknika Corp., Durham, NC) at 1/200, and Texas reda-bungarotoxin (Molecular Probes, Eugene, OR) at 0.5µg/ml, in PBS-T 10% NGS for 1 h at room temperature.After washing with PBS-T, sections were mounted withVectashield (Vector Laboratories, Burlingame, CA). Forstaining with anti-g-AChR sections were fixed for 10min with methanol at 220°C and treated with 50 mMethylamine–HCl (pH 11) at room temperature as indi-cated by Missias et al. (1996) before incubation withprimary antibody. Blocking and dilution of both theanti-g-AChR and the secondary antibodies were per-formed with PBS containing 10% NGS and 1% bovineserum albumin. Washes were done with PBS.
ACKNOWLEDGMENTS
Robert M. Marshall provided excellent technical support through-out the study. This work was supported by a generous gift from theM.A.Antelman Foundation and by NIHGrant NS14506 (U.J.M.), NIHTraining Grant NS07158 (M.R.), and a grant from the NorwegianResearch Council (T.L., I.M.).
REFERENCES
Adams, L., Carlson, B. M., Henderson, L., and Goldman, D. (1995).Adaptation of nicotinic acetylcholine receptor, myogenin, and mrf4gene expression to long-term muscle denervation. J. Cell Biol. 131:1341–1349.
Altiok, N., Bessereau, J.-L., and Changeux, J.-P. (1995). ErbB3 anderbB2/neu mediate the effect of heregulin on acetylcholine receptorgene expression in muscle: Differential expression at the endplate.EMBO J. 14: 4258–4266.
Andreose, J. S., Xu, R., Lømo, T., Salpeter, M. M., and Fumagalli, G.(1993). Degradation of twoAChR populations at rat neuromuscularjunctions: Regulation in vivo by electrical stimulation. J. Neurosci.13: 3433–3438.
Brenner, H. R., Lømo, T., and Williamson, R. (1987). Control ofend-plate channel properties by neurotrophic effects and by muscleactivity in rats. J. Physiol. 388: 367–381.
Brenner, H. R., Herczeg, A., and Slater, C. R. (1992). Synapse-specificexpression of acetylcholine receptor genes and their products atoriginal synaptic sites in rat soleus muscle fibers regenerating in theabsence of innervation.Development 116: 41–53.
Brenner, H. R., Rotzler, S., Kues, W. A., Witzemann, V., and Sakmann,B. (1994). Nerve-dependent induction of AChR e-subunit geneexpression in muscle is independent of the state of differentiation.Dev. Biol. 165: 527–536.
Buckley, K., and Kelly, R. B. (1985). Identification of a transmembraneglycoprotein specific for secretory vesicles of neural and endocrineorigin. J. Cell Biol. 100: 1284–1294.
Chu, G. C., Moscoso, L. M., Sliwkowski, M. X., and Merlie, J. P. (1995).Regulation of the acetylcholine receptor e-subunit gene by recombi-nant ARIA: An in vitro model for transynaptic gene regulation.Neuron 14: 329–339.
Cohen, I., Rimer, M., Lømo, T., and McMahan, U. J. (1997). Agrin-induced postsynaptic-like apparatus in skeletal muscle fibers invivo.Mol. Cell. Neurosci. 9: 327–253.
Corfas, G., and Fischbach, G. D. (1993). The number of Na1 channelsin cultured chick muscle is increased by ARIA, an acetylcholinereceptor-inducing activity. J. Neurosci. 13: 2118–2125.
DeChiara, T. M., Bowen, D. C., Valenzuela, D. M., Simmons, M. V.,Poueymirou, W. T., Thomas, S., Kinetz, E., Compton, D. L., Rojas, E.,Park, J. S., Smith, C., DiStefano, P. S., Glass, D. J., Burden, S. J., andYancopoulos, G. D. (1996). The receptor tyrosine kinase MuSK isrequired for neuromuscular junction formation in vivo. Cell 85:501–512.
Duclert, A., and Changeux, J. P. (1995). Acetylcholine receptor geneexpression at the developing neuromuscular junction. Physiol. Rev.75: 339–368.
Falls, D. L., Rosen, K. M., Corfas, G., Lane, W. S., and Fischbach, G. D.(1993). ARIA, a protein that stimulates acetylcholine receptorsynthesis, is a member of the neu ligand family. Cell 72: 801–815.
Fambrough, D. M. (1979). Control of acetylcholine receptors in skeletalmuscle. Physiol. Rev. 59: 165–227.
Fischbach, G. D., and Schuetze, S. M. (1980). A post-natal decrease inacetylcholine receptor open time in rat end-plates. J. Physiol. 303:125–137.
Gautam, M., Noakes, P. G., Moscoso, L., Rupp, F., Scheller, R. H.,Merlie, J. P., and Sanes, J. R. (1996). Defective neuromuscularsynaptogenesis in agrin-deficient mutant mice. Cell 85: 525–535.
Godfrey, E. W., Nitkin, R. M., Wallace, B. G., Rubin, L. L., andMcMahan, U. J. (1984). Components of the Torpedo electric organand muscle that cause aggregation of acetylcholine receptors oncultured muscle cells. J. Cell Biol. 99: 615–627.
Goldman, D., Brenner, H. R., and Heinemann, S. (1988). Acetylcholinereceptor a-, b-, g-, and d-subunit mRNA levels are regulated bymuscle activity.Neuron 1: 329–333.
Goldman, D., Carlson, B. M., and Staple, J. (1991). Induction ofadult-type acetylcholine receptor gene expression in non-inner-vated regenerating muscle.Neuron 7: 649–658.
Goodearl, A. D., Yee, A. G., Sandrock, A. A., Jr., Corfas, G., andFischbach, G. D. (1995). ARIA is concentrated in the synaptic basallamina of the developing chick neuromuscular junction. J. Cell Biol.130: 1423–1424.
Gordon, T., Jones, R., and Vrbova, G. (1976). Changes in chemosensitiv-ity of skeletal muscles as related to endplate formation. Prog.Neurobiol. 6: 103–136.
Greengard, P., Valtorta, F., Czernik, A. J., and Benfenati, F. (1993).Synaptic vesicle phosphoproteins and regulation of synaptic func-tion. Science 259: 780–785.
Gu, Y., and Hall, Z. W. (1988). Immunological evidence for a change insubunits of the acetylcholine receptor in developing and denervatedrat muscle.Neuron 1: 117–125.
Jo, S. A., Zhu, X., Marchionni, M. A., and Burden, S. J. (1995).
262 Rimer et al.

Neuregulins are concentrated at nerve–muscle synapses and acti-vate ACh-receptor gene expression.Nature 373: 158–161.
Jones, G., Herczeg, A., Ruegg, M. A., Lichtsteiner, M., Kroger, S., andBrenner, H. R. (1996). Substrate-bound agrin induces expression ofacetylcholine receptor e-subunit gene in culturedmammalianmusclecells. Proc. Natl. Acad. Sci. USA 93: 5985–5990.
Jones, G., Meier, T., Lichtsteiner, M., Witzemann, V., Sakmann, B., andBrenner, H. R. (1997). Induction by agrin of ectopic and functionalpostsynaptic-like membrane in innervated muscle. Proc. Natl. Acad.Sci. USA 94: 2654–2659.
Korneliussen, H., and Sommerschild, H. (1976). Ultrastructure of thenew neuromuscular junctions formed during reinnervation of ratsoleus muscle by a ‘‘foreign’’ nerve. Cell Tissue Res. 167: 439–452.
Kues, W. A., Brenner, H. R., Sakmann, B., and Witzemann, V. (1995).Local neurotrophic repression of gene transcripts encoding fetalAChRs at rat neuromuscular synapses. J. Cell Biol. 130: 949–957.
Letinsky, M. S., Fischbeck, K. H., and McMahan, U. J. (1976). Precisionof reinnervation of original postsynaptic sites in frog muscle after anerve crush. J. Neurocytol. 5: 691–718.
Lømo, T., and Westgaard, R. H. (1975). Further studies on the controlof ACh sensitivity by muscle activity in the rat. J. Physiol. 252:603–626.
Lømo, T., and Slater, C. R. (1978). Control of acetylcholine sensitivityand synapse formation by muscle activity. J. Physiol. 275: 391–402.
Lømo, T., and Slater, C. R. (1980). Acetylcholine sensitivity of develop-ing ectopic nerve–muscle junctions in adult rat soleus muscle. J.Physiol. 303: 73–189.
Lømo, T., Pockett, S., and Sommerschild, H. (1988). Control of numberand distribution of synapses during ectopic synapse formation inadult rat soleus muscles.Neuroscience 24: 673–686.
Martinou, J.-C., Falls, D. L., Fischbach, G. D., and Merlie, J. P. (1991).Acetylcholine receptor-inducing activity stimulates expression ofe-subunit gene of the muscle acetylcholine receptor. Proc. Natl. Acad.Sci. USA 88: 7669–7673.
McMahan, U. J. (1990). The agrin hypothesis. Cold Spring Harbor Symp.Quant. Biol. 50: 407–418.
Mishina, M., Takai, T., Imoto, K., Noda, M., Takahashi, T., Numa, S.,Methfessel, C., and Sakmann, B. (1986). Molecular distinctionbetween fetal and adult forms of muscle acetylcholine receptor.Nature 321: 406–411.
Missias, A. C., Chu, G. C., Klocke, B. J., Sanes, J. R., and Merlie, J. P.(1996). Maturation of the acetylcholine receptor in skeletal muscle:Regulation of the AChR g-to-e switch.Dev. Biol. 179: 223–238.
Moscoso, L. M., Chu, G. C., Gautam, M., Noakes, P. G., Merlie, J. P.,and Sanes, J. R. (1995). Synapse-associated expression of an acetyl-choline receptor-inducing protein, ARIA/heregulin, and its puta-tive receptors, erbB2 and erbB3, in developing mammalian muscle.Dev. Biol. 172: 158–169.
Nitkin, R. M., Smith, M. A., Magill, C., Fallon, J. R., Yao, Y.-M. M.,Wallace, B. G., and McMahan, U. J. (1987). Identification of agrin, asynaptic organizing protein from Torpedo electric organ. J. Cell Biol.105: 2471–2478.
Reist, N. E., Werle, M. J., and McMahan, U. J. (1992). Agrin released bymotor neurons induces the aggregation of AChRs at neuromuscularjunctions.Neuron 8: 865–868.
Rotzler, S., and Brenner, H. R. (1990). Metabolic stabilization ofacetylcholine receptors in vertebrate neuromuscular junction bymuscle activity. J. Cell Biol. 111: 655–661.
Sola, L. M., and Martin, A. W. (1953). Denervation hypertrophy andatrophy of the hemidiaphragm of the rat. Am. J. Physiol. 172:324–332.
Tello, J. F. (1917). Genesis de las terminaciones nerviosas motrices ysensitivas. I. En el sistema locomotor de los vertrebrados superiores.Histogenesis muscular. Trab. Inst. Cajal Invest. Biol. 15: 101–199.
Valenzuela, D. M., Stitt, T. N., DiStefano, P. S., Rojas, E., Mattson, K.,Compton, D. L., Nunez, L., Thomas, S., Le Beau, M. M., Fernald,A. A., Copeland, N. G., Jenkins, N. A., Burden, S. J., Glass, D. J.,and Yancopoulos, G. D. (1995). Receptor tyrosine kinase specific forthe skeletal muscle lineage: Expression in embryonic muscle, at theneuromuscular junction and after injury.Neuron 15: 573–584.
Witzemann, V., Barg, B., Nishikawa, Y., Sakmann, B., and Numa, S.(1987). Differential regulation of muscle acetylcholine receptorg- and e-subunit mRNAs. FEBS Lett. 223: 104–112.
Witzemann, V., Brenner, H. R., and Sakmann, B. (1991). Neural factorsregulate AChR subunit mRNAs at rat neuromuscular synapses. J.Cell Biol. 114: 125–141.
Zhu, X., Lai, C., Thomas, S., and Burden, S. J. (1995). Neuregulinreceptors, erbB3 and erbB4, are localized at neuromuscular syn-apses. EMBO J. 14: 5842–5848.
ReceivedApril 14, 1997Revised May 8, 1997Accepted May 8, 1997
g-AChR/e-AChR Switch 263




















