
Excitatory amino acids and synaptic transmission in embryonic rat brainstem motoneurons in...
11
European Journal of Neuroscience, Vol. 11, pp. 1324–1334, 1999 © European Neuroscience Association Excitatory amino acids and synaptic transmission in embryonic rat brainstem motoneurons in organotypic culture Thomas Launey,* Anton Ivanov, Ga ` bor Kapus, 1,² Nadine Ferrand, Istvan Tarnawa 1,‡ and Jean-Patrick Gueritaud Unite ´ de Neurocyberne ´ tique Cellulaire, CNRS UPR 9041, 280, Bd. Sainte Marguerite, 13009, Marseille, France 1 Institute for Drug Research, Budapest, Hungary Keywords: AMPA receptor, EPSPs, excitatory amino acids, glutamate receptor, iontophoresis, kainate receptor, NMDA receptor Abstract We used brainstem motoneurons recorded in organotypic slice co-cultures maintained for more than 18 days in vitro, together with multibarrel ionophoretic applications of glutamate receptor agonists and bath applications of specific blocking agents, to study the responses of rat brainstem motoneurons to glutamate receptor activation, and the contribution of these receptors to synaptic transmission. Differentiated brainstem motoneurons in vitro are depolarized by glutamate, N-methyl-D-aspartate (NMDA) and DL-α-amino-3-hydroxy-5-methyl-4-isoxazole-propionic acid (AMPA) iontophoresis, and express NMDA, AMPA and also specific kainate receptors, as evidenced by (1/–)2-amino-5-phosphonovaleric acid (APV)- and (–)1-(4-aminophenyl)-3-methyl-carbamoyl- 4-methyl-7,8-methylenedioxy-3,4-dihydro-5H-2,3-benzo-diazepine [GYKI 53784 (LY303070)]-resistant depolarizations. Electrical stimulations applied to the dorsal part of the explant trigger excitatory synaptic potentials with latencies distributed in three regularly spaced groups. Excitatory postsynaptic potentials (EPSPs) in the earliest group have a similar latency and time course and correspond to monosynaptic activation. EPSPs in later groups have more scattered latencies and time courses and correspond to polysynaptic activation. Monosynaptic EPSPs are insensitive to the specific NMDA blocker APV, and are completely and reversibly suppressed by the non-competitive AMPA receptor antagonist GYKI 53784 (LY303070). Detailed analysis of the spontaneous excitatory synaptic activity shows that APV decreases the frequency of spontaneous EPSPs without modifying their shape or amplitude. We conclude that excitatory synapses on brainstem motoneurons in vitro are mainly activated through AMPA receptors (AMPA-Rs). NMDA receptors (NMDA-Rs) are present in the membrane, but are located either at extrasynaptic sites or silent synapses, and are not directly involved in synaptic transmission on motoneurons. On the contrary, NMDA receptors contribute to synaptic transmission within the premotor interneuronal network. Introduction A great deal of attention has been paid to the involvement of excitatory amino acids (EAA) in synaptic transmission on spinal motoneurons (Jahr & Yoshioka, 1986; Collingridge & Lester, 1989; Engberg et al., 1993; Walmsley & Bolton, 1994). DL-α-amino-3- hydroxy-5-methyl-4-isoxazole-propionic acid (AMPA) is generally considered to mediate fast excitatory transmission in mature moto- neurons (Jahr & Yoshioka, 1986; Engberg et al., 1993), whereas N-methyl-D-aspartate (NMDA) would be involved in slow synaptic events (Dale & Roberts, 1985), but the relative contribution of NMDA receptors subtypes (NMDA-Rs) and non-NMDA-Rs to synaptic inputs is still unclear, depending on the preparation and experimental paradigm (Nelson et al., 1986; O’Brien & Fischbach, 1986; Jiang Correspondence: J.-P. Gueritaud, as above. E-mail: [email protected] *Present address: Laboratory for Memory and Learning, Brain Science Institute, RIKEN, 2-1 Hirosawa, Wako-shi 351–0198, Saitama, Japan. ² Present address: Chinoin Pharmaceutical Work Co. Ltd., Discovery Research, P.O. Box 110, H-1325, Budapest, Hungary. ‡ Present address: Chemical Works of Gedeon Richter Ltd., Pharmacological Research Center, P.O. Box 27, H-1475, Budapest, Hungary. Received 10 June 1998, revised 13 October 1998, accepted 16 November 1998 et al., 1990; Ziskind-Conhaim, 1990; Trueblood et al., 1996; O’Brien et al., 1997). EAAs have also been shown to be involved in excitotoxic damage and degenerative disorders (Frandsen & Schousboe, 1993), as well as in many processes linked to motor system maturation. Recent studies have shown that low concentrations of glutamate agonist are required to support motoneuronal survival, and that blocking NMDA- Rs during the postnatal period during which mature properties are acquired may lead to dramatic reduction of motoneuronal survival and maturation, both in vitro (Brenneman et al., 1990) and in vivo (Kalb, 1994). As calcium-related events are thought to control phenomena, e.g. morphogenesis, synaptic stabilization or gene expres- sion, the calcium-permeable NMDA-Rs have been naturally suspected to provide the link with activity-dependent neuronal maturation (Brenneman et al., 1990). Recent studies have shown that the time course of expression of glutamate receptor subunits (GLU-Rs) is strictly regulated during maturation (Jakowec et al., 1995), and that NMDA-Rs show transient overexpression during the critical period of motoneuronal development in early postnatal life (Hori & Kanda, 1996). In a recent work on dissociated spinal neurons in culture, Robert et al. (1998) have shown evidence in favour of regulation of transmitter release at developing AMPA synapses by NMDA receptors activation.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Excitatory amino acids and synaptic transmission in embryonic rat brainstem motoneurons in...

European Journal of Neuroscience, Vol. 11, pp. 1324–1334, 1999 © European Neuroscience Association
Excitatory amino acids and synaptic transmission inembryonic rat brainstem motoneurons in organotypicculture
Thomas Launey,* Anton Ivanov, Gabor Kapus,1,† Nadine Ferrand, Istvan Tarnawa1,‡ and Jean-Patrick GueritaudUnite de Neurocybernetique Cellulaire, CNRS UPR 9041, 280, Bd. Sainte Marguerite, 13009, Marseille, France1Institute for Drug Research, Budapest, Hungary
Keywords: AMPA receptor, EPSPs, excitatory amino acids, glutamate receptor, iontophoresis, kainate receptor, NMDA receptor
Abstract
We used brainstem motoneurons recorded in organotypic slice co-cultures maintained for more than 18 days in vitro, together withmultibarrel ionophoretic applications of glutamate receptor agonists and bath applications of specific blocking agents, to study theresponses of rat brainstem motoneurons to glutamate receptor activation, and the contribution of these receptors to synaptictransmission. Differentiated brainstem motoneurons in vitro are depolarized by glutamate, N-methyl-D-aspartate (NMDA) andDL-α-amino-3-hydroxy-5-methyl-4-isoxazole-propionic acid (AMPA) iontophoresis, and express NMDA, AMPA and also specifickainate receptors, as evidenced by (1/–)2-amino-5-phosphonovaleric acid (APV)- and (–)1-(4-aminophenyl)-3-methyl-carbamoyl-4-methyl-7,8-methylenedioxy-3,4-dihydro-5H-2,3-benzo-diazepine [GYKI 53784 (LY303070)]-resistant depolarizations. Electricalstimulations applied to the dorsal part of the explant trigger excitatory synaptic potentials with latencies distributed in three regularlyspaced groups. Excitatory postsynaptic potentials (EPSPs) in the earliest group have a similar latency and time course andcorrespond to monosynaptic activation. EPSPs in later groups have more scattered latencies and time courses and correspond topolysynaptic activation. Monosynaptic EPSPs are insensitive to the specific NMDA blocker APV, and are completely and reversiblysuppressed by the non-competitive AMPA receptor antagonist GYKI 53784 (LY303070). Detailed analysis of the spontaneousexcitatory synaptic activity shows that APV decreases the frequency of spontaneous EPSPs without modifying their shape oramplitude. We conclude that excitatory synapses on brainstem motoneurons in vitro are mainly activated through AMPA receptors(AMPA-Rs). NMDA receptors (NMDA-Rs) are present in the membrane, but are located either at extrasynaptic sites or silentsynapses, and are not directly involved in synaptic transmission on motoneurons. On the contrary, NMDA receptors contribute tosynaptic transmission within the premotor interneuronal network.
Introduction
A great deal of attention has been paid to the involvement ofexcitatory amino acids (EAA) in synaptic transmission on spinalmotoneurons (Jahr & Yoshioka, 1986; Collingridge & Lester, 1989;Engberg et al., 1993; Walmsley & Bolton, 1994).DL-α-amino-3-hydroxy-5-methyl-4-isoxazole-propionic acid (AMPA) is generallyconsidered to mediate fast excitatory transmission in mature moto-neurons (Jahr & Yoshioka, 1986; Engberget al., 1993), whereasN-methyl-D-aspartate (NMDA) would be involved in slow synapticevents (Dale & Roberts, 1985), but the relative contribution of NMDAreceptors subtypes (NMDA-Rs) and non-NMDA-Rs to synaptic inputsis still unclear, depending on the preparation and experimentalparadigm (Nelsonet al., 1986; O’Brien & Fischbach, 1986; Jiang
Correspondence: J.-P. Gueritaud, as above.E-mail: [email protected]
*Present address: Laboratory for Memory and Learning, Brain ScienceInstitute, RIKEN, 2-1 Hirosawa, Wako-shi 351–0198, Saitama, Japan.†Present address: Chinoin Pharmaceutical Work Co. Ltd., Discovery Research,P.O. Box 110, H-1325, Budapest, Hungary.‡Present address: Chemical Works of Gedeon Richter Ltd., PharmacologicalResearch Center, P.O. Box 27, H-1475, Budapest, Hungary.
Received 10 June 1998, revised 13 October 1998, accepted 16 November 1998
et al., 1990; Ziskind-Conhaim, 1990; Truebloodet al., 1996; O’Brienet al., 1997).
EAAs have also been shown to be involved in excitotoxic damageand degenerative disorders (Frandsen & Schousboe, 1993), as wellas in many processes linked to motor system maturation. Recentstudies have shown that low concentrations of glutamate agonist arerequired to support motoneuronal survival, and that blocking NMDA-Rs during the postnatal period during which mature properties areacquired may lead to dramatic reduction of motoneuronal survivaland maturation, bothin vitro (Brennemanet al., 1990) andin vivo(Kalb, 1994). As calcium-related events are thought to controlphenomena, e.g. morphogenesis, synaptic stabilization or gene expres-sion, the calcium-permeable NMDA-Rs have been naturally suspectedto provide the link with activity-dependent neuronal maturation(Brennemanet al., 1990). Recent studies have shown that the timecourse of expression of glutamate receptor subunits (GLU-Rs) isstrictly regulated during maturation (Jakowecet al., 1995), and thatNMDA-Rs show transient overexpression during the critical periodof motoneuronal development in early postnatal life (Hori & Kanda,1996). In a recent work on dissociated spinal neurons in culture,Robertet al. (1998) have shown evidence in favour of regulation oftransmitter release at developing AMPA synapses by NMDA receptorsactivation.

Synaptic transmission in rat brainstem motoneurons 1325
Cranial motoneurons are functionally distinct from spinal moto-neurons. They maturate earlier and are involved in life-supportingactivities that must be functional at birth. Although all motoneurontypes are involved in some type of rhythmic activity, cranial moto-neurons are inserted in neuronal networks with a completely differentarchitecture from those at the spinal level, and little is known aboutthe neurotransmitters involved at their level.
Pharmacological studies have shown that cranial motoneuronsexpress NMDA-Rs (Durandet al., 1987; Chandler, 1989; McCrimmonet al., 1989; Widmeret al., 1992; Reckling, 1992). Although O’Brienet al. (1997) have described spontaneous miniature EPSCs sensitiveto APV, there is no clear evidence for the role of NMDA-Rs inafferent synaptic transmission. Ouardouz & Durand (1994) on ocularmotoneurons, and Funket al. (1997) on hypoglossal motoneurons,have shown fast excitatory synaptic transmission to be mediated byAMPA receptor subtypes (AMPA-Rs), and we have recently shownthat facial and hypoglossal motoneurons in organotypic culturesexpress Ca21-permeable AMPA-Rs (Launeyet al., 1997b).
In the present work, we used the same preparation to study theinvolvement of ionotropic GLU-Rs in excitatory transmission ondeveloping rat motoneurons in an organotypic slice co-culture ofbrainstem slice and muscle tissue. This preparation was described inprevious papers (Gueritaud & Seyfritz, 1992; Eustache & Gueritaud,1995; Launeyet al., 1997a), and showed that facial and hypoglossalmotoneurons can develop functional connections to newly formedstriated muscle fibres and receive monosynaptic excitatory synapticinputs from sensory structures located in the dorsal part of the explant.Here, we show that cranial motoneurons are depolarized by NMDAand AMPA iontophoresis, thus expressing functional NMDA andAMPA-Rs. We report for the first time that these motoneurons alsoexpress specific kainate receptor subtypes (KAI-Rs), and provideelectrophysiological evidence that synaptic transmission at themotoneuron level is dependent on AMPA-Rs. NMDA-Rs, althoughpresent on the motoneuron membrane, are not activated by afferentsynapses.
Part of this work has been published in abstract form (Launeyet al., 1996).
Materials and methods
Brainstem organotypic cultures
Cultures were prepared as described in detail in previous papers(Gueritaud & Seyfritz, 1992; Eustache & Gueritaud, 1995). Briefly,pregnant Wistar rats were purchased from Iffa Credo (France).Housing and experimental animal protocols strictly conformed to theguidelines of the French Agriculture and Forestry Ministry, and wereapproved by the Centre National de la Recherche Scientifique.Following 17–19 days of gestation, rats were killed by decapitation.The two uterine horns were aseptically removed and transferred to aPetri dish under a laminar flow hood for further processing. Completeembryos were extracted and kept intact in cool saline, to be processedone at a time. Brainstems were isolated and transverse slices, 275µmthick, were obtained using a McIlwain tissue chopper. Brainstemexplants containing the facial or hypoglossal motor nuclei wereobtained by trimming the corresponding slices from unnecessarytissue. Muscle explants were obtained from the tongues of the sameembryos. A brainstem explant and muscle explant were placed in adrop of reconstituted chicken plasma (Sigma-Aldrich, France) on aclean glass coverslip. The plasma was then coagulated with Thrombin(Sigma-Aldrich) and the explants quickly positioned during theprocess of coagulation. The muscle explant was placed 0.7 mm
© 1999 European Neuroscience Association,European Journal of Neuroscience, 11, 1324–1334
FIG. 1. Motoneuron identification. (A) Low-power dark-field photomicrographof a brainstem–muscle co-culture. BS, brainstem explant (transverse slice atthe level of the hypoglossal nucleus); M, tongue muscle explant. Noticethe monolayer outgrowth connecting the two explants. Carbocyanin DiIC18microcrystals deposited on muscle tissue are indicated by asterisks. The dottedline indicates the brainstem midline. Positions of stimulating electrode (1)and recording electrode (2) are overdrawn. (B) Phase contrast high-powerphotomicrograph of a motoneuron (arrows). Notice myotube crossing overthe primary dendrite. (C) Fluorescence photomicrograph of the same neuronretrogradely labelled with carbocyanin DiIC18 uptake from muscle explant.Calibration bars: 1 mm (A); 25µm (B and C).
ventrally to the brainstem. After coagulation, the coverslip wasinserted in a 15 mL conical polystyrene tube (Falcon, Becton Dickin-son, France) with 1.5 mL culture medium. Culture medium wascomposed of 49% Dulbecco’s Modified Eagle’s Medium withGlutamax-I, 24% Hanks Balanced Salt Solution, 8% foetal calf serum,2.5% glucose solution at 20%, all supplied by Gibco (Gibco BRL,MD, USA). The osmolarity was adjusted to 290–300 mOsm. Tubeswere then placed on a motor-driven roller drum, tilted 5 ° fromhorizontal and set to 2 rev/min in an incubator. Culture tubes werekept in a dry atmosphere at 36 °C, with a controlled level of carbon

1326 T. Launeyet al.
dioxide set to 5%. Culture medium was replaced twice a week. Itwas supplemented with 20 ng/mL nerve growth factor during the firstweek in vitro and 5 ng/mL for an additional week. No growth-promoting agents were added thereafter. On day 5, the medium wassupplemented for 24 h with antimitotics to prevent further growth ofnon-neuronal cells. Cultures were maintained for 2–8 weeksin vitro.
Intracellular recording technique
Cultures selected for recording displayed a muscle region with strongirregular contractions, a brainstem region with a clear organotypicstructure, and a flat monolayer between the brainstem motor regionand the newly formed muscle fibres (Fig. 1A). Although intracellularrecordings were performed on cultures between 13 and 60 daysin vitro (DIV), optimum cultures were in the range 21–35 DIV. Forrecordings, the coverslip was taken out of the culture tube, transferredto the recording chamber and superfused with Gey’s balanced saltsolution (GBSS) at a rate of 1 mL/min. Standard GBSS contains(in mM): NaCl, 137; KCl, 5; CaCl2, 1.5; MgCl2, 1; MgSO4, 0.3;KH2PO4, 0.2; NaH2PO4, 0.8; NaHCO3, 2.7; glucose, 5.5. When lowMg21 solutions were used, MgSO4 was omitted and MgCl2 wasreduced to 0.1 mM. EAA receptor antagonists were applied bysuperfusion using a perfusion line-switching device. EAAs wereapplied by micro-ionophoresis using a six-channel multibarrel elec-trode, positioned less than 20µm away from the soma of the recordedneuron and connected to an ionophoresis current pump (Bionic Inst.,France). Moving the electrode more than one cell diameter awayfrom the neuron usually resulted in the absence of a detectable effectof the applied substance. Barrels were filled, respectively, with NaCl(1 M), NMDA (150 mM), glutamate (1.5M), NaCl (1M), AMPA(20 mM) and kainate (20 mM) in that order, each compound beingdissolved in equimolar NaOH. EAA stimulations were obtained byapplying 5 or 10 s negative current pulses with current intensities upto 100 nA. By passing the same current intensities through the NaClbarrel, we verified the absence of a direct effect of the electricalstimulation. Glutamate and agonists were applied precisely at 1 minintervals in the following order: glutamate, NMDA, kainate, AMPA.We verified that this protocol did not induce any cross-desensitization.The lack of direct electrical stimulation was checked by applying thesame current through the NaCl barrel.
The recording chamber was fixed on a vibration-free stand in thefocal plane of an inverted microscope (Nikon, Japan) which couldbe moved with a micropositioning platform independently from therecording chamber and electrodes. The culture was observed withphase contrast or Hoffman modulation contrast optics.
Recordings were performed using fine tip microelectrodes. Pipetteswere pulled from standard 1.2 mm o.d. filamented glass capillaries(Clark, Reading, UK) on a programmable micropipette puller (modelP-87, Sutter Instr., USA) and filled with 1M K-acetate. Theseelectrodes had a tip resistance of 50–90 MΩ. Membrane potentialswere recorded using a conventional bridge amplifier (Axoclamp-2A,Axon Instruments, CA, USA) allowing the simultaneous injection ofconstant currents through the electrode. Recordings were digitized at44 kHz and stored with Pulse Code Modulation on video tapes(VR100-A, Instrutech Corp., NY, USA) for later analysis, or digitizedat 10–20 kHz and displayed on-line and stored on an IBM compatiblemicrocomputer using a 1401-plus digital interface and dedicatedsoftware (CED, Cambridge, UK).
Extracellular microstimulation was performed using glass pipetteswith fire-polished tips 20–50µm in diameter, filled with GBSS. Short(0.1–0.5 ms) voltage pulses of varying amplitude (0.1–10 V) weredelivered by a conventional stimulator.
© 1999 European Neuroscience Association,European Journal of Neuroscience, 11, 1324–1334
Analysis of EPSPs
Surface area
Ten consecutive digitized monosynaptic excitatory postsynaptic poten-tials (EPSPs) without contaminating overlying spontaneous or polys-ynaptic activity were selected and averaged in each case. The surfacearea was measured by automatic integration of the trace between twocursors. The initial cursor was positioned on the last digitized pointbefore the start of the potential. The final cursor was positioned100 ms later. We verified that this position was always clearlyposterior to the return of the potential to base line level.
Extraction of individual spontaneous EPSPs
A semi-manual fitting procedure was used to extract spontaneousEPSPs from digitized continuous recordings. A complete descriptionand discussion of this procedure is given elsewhere (Larkumet al.,1998). Briefly, selected EPSPs were fitted, using a computer-assistedmethod, with a generalizedα-function (a–b function) of the form
f(t) 5 ctae–bt
which was shown to describe the shape of EPSPs accurately. Charac-teristic parameters of fitted EPSPs, e.g. time-to-peak, half-width, risetime and amplitude, could be computed from the fitted function andsaved for further analysis. The quality of the fit was checked bysubtraction of the summed fits from the original data. The fit wasaccepted when this procedure produced a straight line (Fig. 8B). Toavoid spontaneous variations of the EPSP frequency, a sample of 600EPSPs was obtained in each case by fitting all the EPSPs fromsuccessive 400 ms periods of digitized tracings taken at 3 s intervals.
Motoneuron identification
In previous studies (Gueritaud & Seyfritz, 1992; Eustache &Gueritaud, 1995; Launeyet al., 1997a), we demonstrated the existenceof functional motoneurons in our cultures and described the maincriteria that can be used to identify them. We showed that, under ourculture conditions, motoneurons derived from facial or hypoglossalstructures displayed identical electrophysiological properties(Eustache & Gueritaud, 1995). Motoneurons are identified as largecells with an eccentric nucleus and a single dark nucleolus (Fig. 1B),always located in the motor region or at the ventral periphery of thebrainstem explant, in the monolayer region between the brainstemand the muscle tissue (Fig. 1A). Such morphological and topographicalidentification was further confirmed in this work using retrogradefluorescent labelling (Fig. 1A,C) with the carbocyanin dye DiIc18(1,19-dioctadecyl-3,3,39,39-tetramethyl-indocarbocyanine perchlorate,Molecular Probes Europe, Leiden, The Netherlands), applied onmuscle tissue (Fig. 1A) 24 h before the recording session to demon-strate that the axon leaves the brainstem explant and reaches themuscle tissue. This procedure consistently resulted in 20–60 labelledmotoneurons per culture. Unambiguous functional identification wasobtained through antidromic activation from muscle tissue stimulation(Launeyet al., 1997a), but this was not possible to perform for eachrecorded motoneuron.
Results
The database for the present report stems from 102 stable intracellularrecordings obtained from identified motoneurons in organotypiccultures maintained from 12 to 60 DIV. The majority (n 5 62)were recorded between 20 and 37 DIV. Accessing motoneurons for
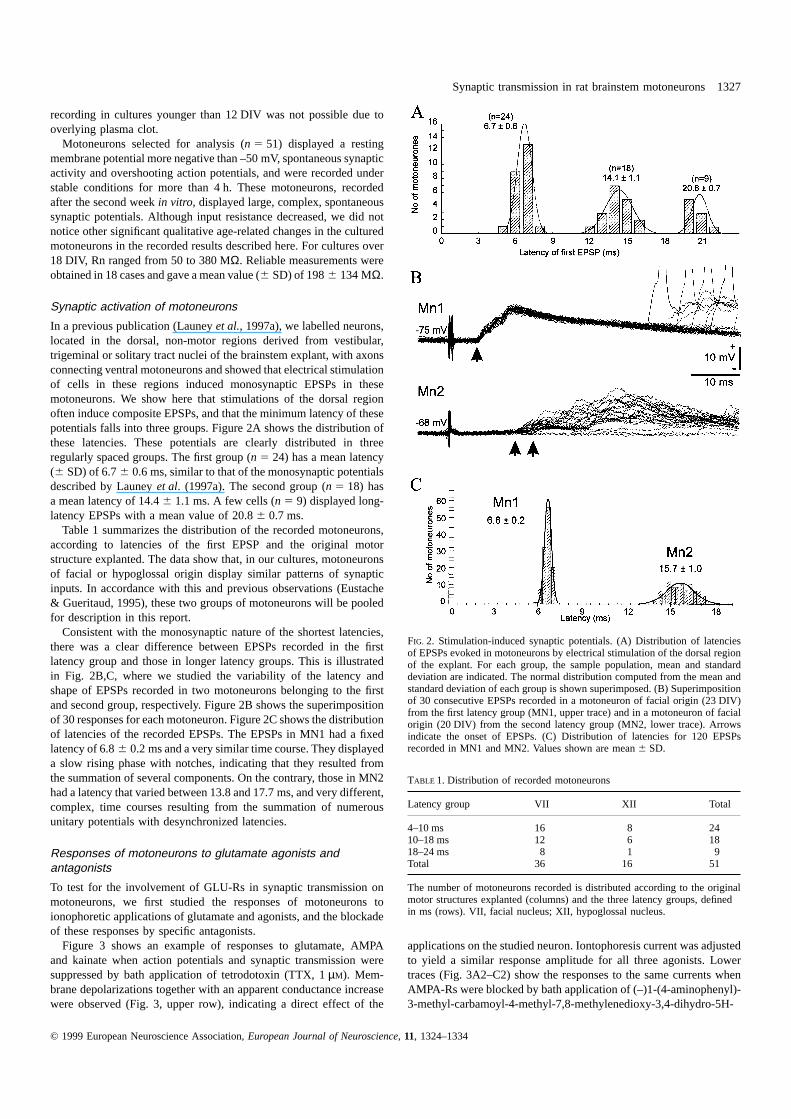
Synaptic transmission in rat brainstem motoneurons 1327
recording in cultures younger than 12 DIV was not possible due tooverlying plasma clot.
Motoneurons selected for analysis (n 5 51) displayed a restingmembrane potential more negative than –50 mV, spontaneous synapticactivity and overshooting action potentials, and were recorded understable conditions for more than 4 h. These motoneurons, recordedafter the second weekin vitro, displayed large, complex, spontaneoussynaptic potentials. Although input resistance decreased, we did notnotice other significant qualitative age-related changes in the culturedmotoneurons in the recorded results described here. For cultures over18 DIV, Rn ranged from 50 to 380 MΩ. Reliable measurements wereobtained in 18 cases and gave a mean value (6 SD) of 1986 134 MΩ.
Synaptic activation of motoneurons
In a previous publication (Launeyet al., 1997a), we labelled neurons,located in the dorsal, non-motor regions derived from vestibular,trigeminal or solitary tract nuclei of the brainstem explant, with axonsconnecting ventral motoneurons and showed that electrical stimulationof cells in these regions induced monosynaptic EPSPs in thesemotoneurons. We show here that stimulations of the dorsal regionoften induce composite EPSPs, and that the minimum latency of thesepotentials falls into three groups. Figure 2A shows the distribution ofthese latencies. These potentials are clearly distributed in threeregularly spaced groups. The first group (n 5 24) has a mean latency(6 SD) of 6.76 0.6 ms, similar to that of the monosynaptic potentialsdescribed by Launeyet al. (1997a). The second group (n 5 18) hasa mean latency of 14.46 1.1 ms. A few cells (n 5 9) displayed long-latency EPSPs with a mean value of 20.86 0.7 ms.
Table 1 summarizes the distribution of the recorded motoneurons,according to latencies of the first EPSP and the original motorstructure explanted. The data show that, in our cultures, motoneuronsof facial or hypoglossal origin display similar patterns of synapticinputs. In accordance with this and previous observations (Eustache& Gueritaud, 1995), these two groups of motoneurons will be pooledfor description in this report.
Consistent with the monosynaptic nature of the shortest latencies,there was a clear difference between EPSPs recorded in the firstlatency group and those in longer latency groups. This is illustratedin Fig. 2B,C, where we studied the variability of the latency andshape of EPSPs recorded in two motoneurons belonging to the firstand second group, respectively. Figure 2B shows the superimpositionof 30 responses for each motoneuron. Figure 2C shows the distributionof latencies of the recorded EPSPs. The EPSPs in MN1 had a fixedlatency of 6.86 0.2 ms and a very similar time course. They displayeda slow rising phase with notches, indicating that they resulted fromthe summation of several components. On the contrary, those in MN2had a latency that varied between 13.8 and 17.7 ms, and very different,complex, time courses resulting from the summation of numerousunitary potentials with desynchronized latencies.
Responses of motoneurons to glutamate agonists andantagonists
To test for the involvement of GLU-Rs in synaptic transmission onmotoneurons, we first studied the responses of motoneurons toionophoretic applications of glutamate and agonists, and the blockadeof these responses by specific antagonists.
Figure 3 shows an example of responses to glutamate, AMPAand kainate when action potentials and synaptic transmission weresuppressed by bath application of tetrodotoxin (TTX, 1µM). Mem-brane depolarizations together with an apparent conductance increasewere observed (Fig. 3, upper row), indicating a direct effect of the
© 1999 European Neuroscience Association,European Journal of Neuroscience, 11, 1324–1334
FIG. 2. Stimulation-induced synaptic potentials. (A) Distribution of latenciesof EPSPs evoked in motoneurons by electrical stimulation of the dorsal regionof the explant. For each group, the sample population, mean and standarddeviation are indicated. The normal distribution computed from the mean andstandard deviation of each group is shown superimposed. (B) Superimpositionof 30 consecutive EPSPs recorded in a motoneuron of facial origin (23 DIV)from the first latency group (MN1, upper trace) and in a motoneuron of facialorigin (20 DIV) from the second latency group (MN2, lower trace). Arrowsindicate the onset of EPSPs. (C) Distribution of latencies for 120 EPSPsrecorded in MN1 and MN2. Values shown are mean6 SD.
TABLE 1. Distribution of recorded motoneurons
Latency group VII XII Total
4–10 ms 16 8 2410–18 ms 12 6 1818–24 ms 8 1 9Total 36 16 51
The number of motoneurons recorded is distributed according to the originalmotor structures explanted (columns) and the three latency groups, definedin ms (rows). VII, facial nucleus; XII, hypoglossal nucleus.
applications on the studied neuron. Iontophoresis current was adjustedto yield a similar response amplitude for all three agonists. Lowertraces (Fig. 3A2–C2) show the responses to the same currents whenAMPA-Rs were blocked by bath application of (–)1-(4-aminophenyl)-3-methyl-carbamoyl-4-methyl-7,8-methylenedioxy-3,4-dihydro-5H-

1328 T. Launeyet al.
FIG. 3. Depolarizing responses to ionophoreticapplications of glutamate and agonists on amotoneuron during bath application of TTX(1 µM). Recording from a motoneuron of facialorigin after full blockade of action potentialsby bath application of TTX (1µM). (A1, B1and C1) Control responses to glutamate,AMPA and kainate, respectively. (A2, B2 andC2) The same responses after bath applicationof GYKI 53784 (10µM).
2,3-benzo-diazepine (GYKI 53784 or LY303070, 10µM). GYKI53764 is a powerful specific non-competitive antagonist of AMPA-Rswhich was preferred to 6-cyano-7-nitroquinoxaline-2,3-dione (CNQX)because of its selectivity for AMPA-Rs and its non-competitive action(Bleakmanet al., 1996; Lodgeet al., 1996; Vizi et al., 1996). Theglutamate response (A2) was reduced by 81%, whereas the AMPA(B2) and kainate (C2) responses were reduced by 57% and 56%,respectively, compared to the control.
As we were mostly concerned with the expression of GLU receptorsand their implication in synaptic transmission, TTX was not used inmost experiments, although this may impair the precise quantificationof the depolarizing effect of the drugs.
The responses induced by ionophoretic application of glutamateand agonists on a motoneuron at resting potential are further detailedin Fig. 4. A 50 nA current pulse through the glutamate barrel of theelectrode induced a membrane depolarization with superimposedspiking and an apparent input resistance decrease (Fig. 4A1). Uponcessation of glutamate ejection, spiking stopped and membranepotential returned to base line. The amplitude of the membranedepolarization, the apparent decrease of input resistance and thefrequency of the spike burst were dose dependent.
These glutamate responses were modified by bath applicationsof the NMDA-Rs antagonist (1/–)2-amino-5-phosphonovaleric acid(APV) or the AMPA-Rs antagonist GYKI 53784.
Figure 4A2 shows that when NMDA-Rs were blocked by bathapplication of APV (50µM), the glutamate response to the samestimulus was slightly modified. The apparent input resistance wasfurther reduced and the firing frequency decreased by approximately20%. When AMPA-Rs were blocked by addition of GYKI 53784(10 µM) to the APV-containing bath (Fig. 4A3), this response wasfurther reduced, but not completely suppressed.
Ionophoretic application of NMDA (50 nA) from the same multi-barrel electrode in a standard Mg21 GBSS (Fig. 4B1) induced anincrease in the firing frequency together with a slight decreasein input resistance, with no clear change in membrane potential.Preliminary bath application of APV (50µM) (Fig. 4B2) stronglyreduced this response. A slight membrane depolarization remainedtogether with anodal break rebound spikes at the brake of theconductance pulses. The specificity of NMDA-Rs activation wasdemonstrated by the fact that addition of GYKI 53784 (10µM) to
© 1999 European Neuroscience Association,European Journal of Neuroscience, 11, 1324–1334
the bath (Fig. 4B3) did not modify the response. After washout ofthe drugs, the full NMDA response was recovered (Fig. 4B4).
In the same cell, kainate (50 nA) applied from the same multibarrelelectrode (Fig. 4C1) induced a response very similar to the glutamateresponse by producing a strong increase in firing frequency, togetherwith membrane depolarization, sharp apparent input resistance decreaseand spike inactivation. Bath application of APV (50µM) (Fig. 4C2)suppressed spontaneous firing but did not modify the pattern of responseto kainate. Addition of GYKI 53784 (10µM) (Fig. 4C3) to the bathstrongly inhibited this response. In this condition, kainate induced aslight (5 mV) depolarization with moderate spiking and no changein input resistance. With larger doses (20µM) of GYKI 53784 (notillustrated), the kainate response was not completely suppressed. Wash-out of the drugs restored the control response (Fig. 4C4).
The specific response of motoneurons to AMPA application isillustrated with another cell. Figure 5A1–A4 shows the responses toprogressively increased AMPA ejection currents on a motoneuron atresting potential (–57 mV). At low doses, AMPA induced a slowdepolarization. Increasing the current intensity increased the amplitudeand rising speed of the depolarization that eventually triggered aburst of action potentials. The frequency of action potentials increasedin a dose-dependent way during AMPA ejection and decreased uponcurrent cessation. Bath application of GYKI 53784 (5µM) partiallyinhibited the response (Fig. 5B1–B4). With an ejection currentintensity of 100 nA, spiking was suppressed, but a sub-thresholddepolarization remained. Increasing AMPA ejection current intensityincreased this depolarization and restored spiking at a low frequency.
Contribution of glutamate receptors to monosynaptic EPSPs
Figure 6A shows the superimposition of four monosynaptic responsesrecorded in a hypoglossal motoneuron and averaged over five sweepseach. When APV (20µM) was added to the bath, the monosynapticEPSP was unchanged, but it was completely blocked by GYKI 52466(60 µM). Washing out partly recovered the response. This resultremained unchanged when recordings were performed in a low-magnesium GBSS, with 10 mM glycine and 10 mM bicuculline added(not illustrated). Because magnesium could not be completely removedfrom the recording medium, NMDA-Rs might still be blocked by theremaining 0.1 mM. To uncover any late NMDA component in therecorded EPSPs, we measured the surface area of the monosynaptic

Synaptic transmission in rat brainstem motoneurons 1329
FIG. 4. Effects of glutamate agonists and antagonists. Responses of amotoneuron of facial origin (32 DIV) with a resting membrane potential of–75 mV to ionophoretic applications of glutamate 50 nA (left column), NMDA50 nA (middle column), and kainate 50 nA (right column). Drug applicationis indicated by the black bar below each trace. (A1–C1) Control responses.Notice the sharp apparent input resistance decrease for glutamate and kainateresponses. (A2–C2) Responses obtained during bath application of APV(50 µM in normal Mg21 solution). The spiking frequency in A2 is slightly(20%) reduced and the apparent input resistance squeezed. The kainateresponse is unaffected. (A3–C3) Responses obtained during bath applicationof APV (50 µM in normal Mg21 solution) and GYKI 53784 (10µM). Theglutamate and kainate responses are strongly reduced, but not completelysuppressed. Notice that the apparent input resistance is unchanged. TheNMDA response is identical to the APV response in (B2). (A4–C4) Returnto control conditions after washout of the drugs.
response at different membrane potentials (Fig. 6B). The experimentaldata points could be fitted with a single regression line, and noevidence was observed of the uncovering of a late component atdepolarized levels.
Figure 6C–E illustrates the concomitant blockade of the monosynap-tic potentials (Fig. 6C1–E1) and the AMPA response (Fig. 6C2–E2)by bath application of GYKI 53784 (5µM). In order to enhance EPSPcomponents and to block spike generation, EPSPs were triggered athyperpolarized levels during each conductance pulse. Ten EPSPswere averaged before ionophoretic ejection of AMPA. GYKI 53784reversibly suppressed both the EPSP and the AMPA response withoutchanging the input resistance of the cell. As already shown in Figs3 and 5, the AMPA response was not completely suppressed. Blockadeof the EPSP at hyperpolarized levels was also incomplete and a slightdepolarization remained.
Contribution of glutamate receptors to polysynaptic EPSPs
Figure 7 shows the effect of APV on polysynaptic potentials. InFig. 7A, 50 control responses are superimposed to show the variability
© 1999 European Neuroscience Association,European Journal of Neuroscience, 11, 1324–1334
FIG. 5. Dose-dependent responses to ionophoretic applications of AMPA nearthe soma of a motoneuron. Drug application is indicated by the black bar beloweach trace, together with the corresponding current intensity. Motoneuron offacial origin (23 DIV). (A1–A4) Responses to increasing doses of AMPA incontrol saline. (B1–B4) Responses to increasing doses of AMPA during bathapplication of GYKI 53784 (5µM).
of the latency (20.26 2.4 ms) and the composite shape of theseEPSPs. In Fig. 7B, the same situation is shown after bath applicationof 50 µM of APV. Notice the large increase in response failures andthe important variability of latency and shape of the recordedpotentials. The histogram in Fig. 7C shows the distribution ofamplitudes of the first component, measured at the dashed line inFig. 7A,B (32 ms), for 150 successive stimulations, before and afterAPV application. APV induced a significant change in distribution(P , 0.001, Mann–Whitney U-test) characterized by a sharp decreasein the mean amplitude of the potentials.
These results are further supported by a detailed analysis of the effectsof APV and GYKI on spontaneous EPSPs recorded in motoneurons.Brainstem motoneurons in organotypic cultures display regular epochsof intense synaptic activity lasting a few minutes, interspaced withquiet periods. Figure 8A shows that APV (50µM) reduced but did notsuppress spontaneous synaptic activity recorded during active periods.The spontaneous EPSP’s frequency was sharply reduced, but largepotentials were not suppressed. Further addition of GYKI 53784(20 µM) in the perfusion completely and reversibly blocked these poten-tials. These two patterns of activity were clearly different from theepisodic firing of a single isolated EPSP observed during the quietperiods.
Using a dedicated computer program, individual spontaneousEPSPs were extracted by successive peeling, using an a–b functionfitting procedure. The procedure is illustrated in Fig. 8B. The uppertrace shows the original recording. The middle trace shows the sumof a–b fits on this recording. The lower trace shows the result of thesubtraction of the sum of a–b fits from the original data, demonstratingthe validity of the procedure to describe the shape of the recordedspontaneous EPSPs. A sample of 600 individual EPSPs (see Materialsand methods) was collected during periods of spontaneous activitybefore and after APV application, and the interevent delay, amplitudeand half-width distributions of each population were compared(Fig. 8C–E). The delay between single EPSPs is increased (from

1330 T. Launeyet al.
FIG. 6. Effects of EAA antagonists onmonosynaptic EPSPs. (A) Compositemonosynaptic EPSPs recorded in a motoneuronof hypoglossal origin. Each trace is theaverage of 10 responses. Averaged responsesin different conditions are superimposed forcomparison. Control, control response instandard GBSS; APV, response during bathapplication of APV (20µM), the response isidentical to the control; GYKI, response duringbath application of APV (20µM) and GYKI52466 (60µM), the response is suppressed;Wash, response after washout of the drugs.(B) Plot of the surface of the monosynapticEPSP recorded at different membranepotentials. Surface measurements are indicatedas a percentage of the maximum value. Theblack dots correspond to recordings in controlconditions and empty squares to recordingsduring bath application of GYKI 53784(5 µM). (C–E) Concomitant blockade of theEPSP and AMPA response by GYKI 53784.EPSPs are triggered during a hyperpolarizingconductance pulse to suppress spiking andenhance amplitude. Upper traces are averagesof 10 responses taken immediately before theAMPA application shown at a slower sweepspeed on lower traces. The black bar below thelower traces indicates ionophoretic currentapplication. (C1 and C2) Control responses.(D1 and D2) Responses after bath applicationof GYKI 53784 (5µM). Notice that the inputresistance is unchanged. (E1 and E2) Recoveryof responses after washout of the drug.
49.8 ms to 131.3 ms in the illustrated example in Fig. 8C) and thetwo distributions are significantly different (P , 0.001,Mann–Whitney test). The shape and amplitude of single events arenot modified, as illustrated in Fig. 8D,E. The distributions are notsignificantly different (P . 0.2 andP . 0.05, respectively, Mann–Whitney test). This indicates that APV acted at a site remote fromthe motoneuron somatodendritic membrane by reducing the frequencyof synaptic potentials reaching the motoneuron. Thus, contrary to theresults on motoneurons membrane, NMDA-Rs would contribute tosynaptic activity within the premotor network.
Discussion
Our results show that identified brainstem motoneurons differentiatedin organotypic culturesin vitro express the main types of ionotropicGLU-Rs, i.e. NMDA, AMPA and also specific kainate types, andthat NMDA-Rs seem to be involved in synaptic transmission at thepremotor level but not on the motoneurons membrane, where synaptictransmission depends mainly on AMPA-Rs.
Before discussing our results in more detail, we should considersome particularities of our preparation, i.e. the fact that motoneuronsderived from facial or hypoglossal nuclei express similar electro-
© 1999 European Neuroscience Association,European Journal of Neuroscience, 11, 1324–1334
physiological properties (Eustache & Gueritaud, 1995) and synapticorganization, and that inhibitory postsynaptic potentials (IPSPs) areonly exceptionally observed. It is well known that a variable degreeof synaptic reorganization takes place in all organotypic preparations,and that the degree of tissue organization in the final culture isdependent to a large extent on the maturity of the tissue at the timeof explantation (Ga¨hwiler et al., 1991). In our case, the brainstemtissue at E18–E19 is still largely immature. Neurons have terminatedtheir migrations and nuclear structures can be identified (Paxinoset al., 1991), but synaptic connections are not established and mostneurons are still devoid of dendritic arbor. Furthermore, motoneuronsare dependent on trophic support from muscle tissue for survival andproper differentiation (Nurcombe & Bennett, 1982; Openheim, 1989;Eustacheet al., 1994). In such conditions, it is most likely thatmotoneurons will develop similar properties, dependent on the premo-tor structures available in the slice and the type of muscle tissueco-explanted. The cultures reach an optimal condition for electro-physiological studies between 3 and 6 weeksin vitro (Gahwiler et al.,1991; Spengeret al., 1991; Gueritaud & Seyfritz, 1992), after whichthey start to degenerate. The results described in this paper wereobtained during this optimal period. IPSPs have been observed in asimilar organotypic preparation obtained from spinal cord slices

Synaptic transmission in rat brainstem motoneurons 1331
FIG. 7. Influence of APV on polysynaptic EPSPs. (A) Superimposition of 50synaptic responses with a minimum latency of 206 3 ms. (B) The samesituation during bath application of APV (50µM). Notice the large increasein stimulation failures as well as the great variability of shapes and latenciesof EPSPs. (C) Distribution of the amplitude of responses measured at 32 ms(dashed line in A and B) before and during APV application.
(Spengeret al. 1991; Streitet al., 1991). These authors have shownthat an organotypic structure develops, with the morphological andfunctional correlates of the spinal reflex arc, including IPSPs. It ismost likely that the absence of IPSPs in our preparation can beexplained by the fact that inhibitory premotor structures, located morerostrally, are excluded from the original brainstem slice.
Glutamate receptors activation
Ionophoretic application of glutamate in the immediate proximity ofthe soma of brainstem motoneurons in culture induces a dose-dependentconductance increase, together with a membrane depolarization, andburst of action potentials that can be precisely compared with responsesrecorded on brainstem or spinal motoneuronsin vivo (Durandet al.,1987; Engberget al., 1993; Ouardouz & Durand, 1994). The fact thatthis response was only observed when the multibarrel electrode waslocated less than 20µm away from the soma, and the fact that blockadeof spike generation and synaptic transmission by TTX did not suppress
© 1999 European Neuroscience Association,European Journal of Neuroscience, 11, 1324–1334
these responses, are strong argumentsagainst an effect of the applicationon nearby neurons.
Glutamate is known to activate a variety of ionotropic membranereceptor subtypes named according to their specific agonists NMDA,kainate or AMPA (Watkins & Evans, 1981). Our results show thationophoretic application of NMDA in the vicinity of the soma has adepolarizing effect. It should be noticed that these NMDA responsescould be recorded despite the fact that we used a standard (1.3 mM)Mg21 solution. Receptors of the NMDA type are selectively antagon-ized by APV (Davieset al., 1981). Because APV has a minor inhibitoryeffect on the glutamate-induced response of motoneurons in culture, itmay be assumed that functional NMDA-Rs have a low membranedensity.
Our results show a major contribution of AMPA-Rs to the glutamateresponse. This is in accordance with the results of ionophoretic studieson brainstem motoneuronsin vivo (Durandet al., 1987; Ouardouz &Durand, 1994). This correlates well with our previous study (Launeyet al., 1997b), which shows, using EAA-induced cobalt uptake, thatbrainstem motoneurons in organotypic culture express calcium-per-meable AMPA-Rs on their primary dendrites.
Non-NMDA-Rs have been separated in KAI-Rs and AMPA-Rs.Kainate activates both KAI-Rs and AMPA-Rs, but these two familiesof receptors can be distinguished on the basis of their molecular subunitsconstitution (Hollmann & Heinemann, 1994), as well as their desensit-ization properties (Zorumskiet al., 1989) and sensitivity to 2,3-benzo-diazepine blockers (Donevan & Rogawski, 1993; Viziet al., 1996;Lodgeet al., 1996).
GYKI 53784 (LY303070), a compound belonging to the 2,3-benzo-diazepine family, is the active enantiomer of the racemic GYKI 53655,which was shown to be a potent non-competitive antagonist of AMPA-mediated receptor activation (Viziet al., 1996) and to have a clearselectivity for AMPA-Rs over KAI-Rs in cortical and DRG cells (Wild-ing & Huettner, 1995) or hippocampal neurons (Lermaet al., 1997), asdoes LY303070 (GYKI 53784) on cerebellar cells (Bleakmanet al.,1996).
The existence of specific functional receptors of the kainate typeexpressed by native brain cells has long been controversial (Lermaet al., 1997), but such receptors have recently been shown to mediatesynaptic activation in CA3 hippocampal neurons (Vignes &Collingridge, 1997; Castilloet al., 1997). Kainate is a strong agonist ofthe GLU-Rs on our brainstem motoneurons, and the kainate responseis not completely suppressed by high doses (up to 10 mM) of GYKI53784 (LY303070). Donevan & Rogawski (1993) and Zorumskiet al.(1993) have shown that less than 50 mM of the less potent GYKI 52466blocks more than 90% of kainate or AMPA responses on AMPA-Rs incultured hippocampal neurons, and Wilding & Huettner (1995) considerthat 10 mM GYKI 53655, which they consider about 20-fold morepotent than GYKI 52466, should block more than 90% of AMPA-preferring receptors. Thus, the remaining response observed with highdoses (10 or 20 mM) of the most potent GYKI 53784 could correspondto the activation of kainate-preferring GYKI 53784-resistant receptors(Lermaet al., 1997), although the similarity of the responses to kainateand AMPA recorded under TTX and GYKI (Fig. 3B2,C2) would indi-cate a preferential effect of kainate on AMPA-Rs.
Synaptic activation of motoneurons
Stimulations of the explant at sites in the dorsal part of the explantlocated well outside of the motor region induced EPSPs in identifiedmotoneurons with latencies distributed in three regularly spaced groups.The first group had a narrow distribution, and for each recorded motone-uron within this group the shape and latency of the EPSP were constant.These EPSPs correspond to the monosynaptic potentials described

1332 T. Launeyet al.
FIG. 8. Analysis of spontaneous synapticactivity. (A) Modifications of synaptic noise.Control, synaptic noise in standard GBSSsolution; APV, effect of bath application of50 µM APV; GYKI, suppression of synapticnoise by bath application of APV (50µM) andGYKI 53784 (20µM); Wash, recovery afterwashout of the drugs. (B) ‘a–b’ fitting (seetext) to extract individual EPSPs. Upper trace:original recording. Middle trace: sum of thesuccessive a–b functions. Lower trace: resultof the subtraction of the sum of a–b functionsfrom the original recording. (C) Distribution ofintervals between successive EPSPs before andduring APV application. The distributionmeans (vertical dotted lines) shift from 49.8 to131.3 ms. (D and E) Distribution of EPSPshalf-width and amplitude, respectively, beforeand during APV application.
previously (Launeyet al., 1997a). The time course of monosynapticpotentials sometimes displayed a composite nature resulting from thesummation of a few unitary EPSPs. The other two groups correspondto polysynaptic EPSPs with more scattered latencies and variable timecourses. Mean latency values were roughly multiples of the monosynap-tic one. Observed latencies imply rather slow conduction velocities andsynaptic delays, but these are in agreement with conduction velocitiesand synaptic delays determined in other organotypic preparations (Streitet al., 1991).
Contribution of glutamate receptors to synaptic activation
Our results show that GLU-Rs are involved in synaptic transmissionon motoneurons in culture, as AMPA-R antagonists suppress synapticpotentials. This was expected because glutamate is a major neuro-transmitter in the motor systemin vivo (Collingridge & Lester, 1989).The lack of effect of APV on monosynaptic EPSPs and their completeblockade by AMPA-R antagonists tends to demonstrate that NMDA-Rs are not involved in synaptic transmission at the motoneuron level andthat synaptic transmission on brainstem motoneurons in organotypicculture is mostly dependent on AMPA-Rs.
© 1999 European Neuroscience Association,European Journal of Neuroscience, 11, 1324–1334
The fact that responses to ionophoretic applications of NMDA wererecorded at normal Mg21 concentrations shows that these receptors arenot completely blocked by Mg21 at resting potential. Yet, it may beargued that a small contribution of NMDA-Rs to the EPSPs might stillbe masked by partial Mg21 blockade, voltage dependency or superim-position of an inhibitory component. This hypothesis is not supportedby our experimental data. EPSPs recorded in a low Mg21 solutionsupplemented with glycine and bicuculline did not display any modi-fications of shape. As we could not completely suppress magnesiumfrom the recorded solution, we tried to detect small voltage-dependentchanges of the EPSPs shapes using surface measurements. This verysensitive test showed a clear linear variation of the EPSPs with mem-brane depolarization and a complete blockade of the response by GYKIcompounds at all potentials. The unmasking of a voltage-dependentNMDA component would have induced a change in slope. Thus, itseems most likely that NMDA-Rs are not involved in these responses.
Although APV does not modify the monosynaptic EPSP, it doeschange the shape of polysynaptic potentials and the pattern of synapticnoise. Using a fitting procedure to extract the shape of individualspontaneous synaptic potentials, we described a change in the delay

Synaptic transmission in rat brainstem motoneurons 1333
between successive EPSPs, corresponding to a lowering of frequencyfor the EPSPs. As no modification in half-width or amplitude of thepotentials was seen, this clearly indicates that these modifications occurat the premotor level. It may be argued that the fitting procedure maynot be able to extract all the EPSPs occurring with high frequency(Larkumet al., 1998). This procedure may in fact lead to an underevalu-ation of the frequency of EPSPs, if this frequency is high, as in controlconditions. Because the EPSP frequency under APV is lower, its meas-urement can be expected to be more accurate. This would mean that thedecrease in EPSP frequency reported here is an underestimate of theactual one.
The analysis of the shape of high-frequency EPSPs may also beimpaired by non-linear summation of synaptic events (Rall, 1964). Thiswould mainly affect the fast (AMPA) events and less prominently theslow (NMDA) events, and application of APV, blocking NMDA events,would induce a change in the distributions of half-width, affected by apresumed NMDA component, or amplitudes (mainly AMPA compon-ents). This is not the case. It may thus be concluded that APV-inducedchanges do not take place on the motoneuron membrane. Thus, NMDA-dependent synaptic transmission would take place on neurons involvedin the premotor network.
This demonstrates that the lack of NMDA components in mono-synaptic EPSPs on motoneurons is not due to a culture artefact inducingthe non-expression or downregulation of NMDA-Rs at the synapticlevel (Giffardet al., 1990). As ionophoretic applications of NMDA onmotoneurons demonstrate the existence of NMDA-Rs in the motoneu-ron membrane, these receptors might be located either at extrasynapticsites or silent synapses. Such synapses were described bothin vivo(Hennemanet al., 1984) andin vitro (Punet al., 1986), where they maybe involved in a trophic process (Kalb, 1994, Durandet al., 1996).
NMDA-Rs are known to mediate slow synaptic responses at manysynapses in the brain (Collingridge & Lester, 1989). The balancebetween NMDA and AMPA-Rs was shown to change from one synapseto thenext (Bekkers&Stevens,1989).ThecontributionofNMDA-Rs tosynaptic transmission inmaturespinalmotoneurons isstill controversial(Jahr & Yoshioka, 1986; Walmsley & Bolton, 1994). NMDA-Rs havebeen shown to contribute to synaptic responses at the spinal level incats (Engberget al., 1993), chick embryonic (O’Brien & Fischbach,1986), or rat embryonic (Ziskind-Conhaim, 1990) or neonatal (Pinco& Lev-Tov, 1994) motoneurons. Yet, their implication in synaptictransmission seems to be age dependent. Ziskind-Conhaim (1990) andHori & Kanda (1996) show that slow NMDA-dependent EPSPs appearearly and tend to diminish during the two first postnatal weeks to bereplaced by fast AMPA-dependent EPSPs (Nelsonet al., 1986; Robertset al., 1998). A similar pattern seems to apply to brainstem motoneu-rons. In neonatal rat brainstem slices, trigeminal and hypoglossalmotoneurons apparently express both NMDA and non-NMDA-Rs atsynaptic sites (Truebloodet al., 1996; O’Brienet al., 1997), but record-ings obtainedin vivoon adult rat abducens motoneurons (Ouardouz &Durand, 1994) suggest that NMDA-Rs are not involved in synaptictransmission. The results reported here on developing brainstemmotoneuronsin vitro correlate well with this observation, but as wecould not record from motoneurons during the first 12 days in culture,it remains to be determined by other means whether NMDA-Rs werepresent at synaptic sites during the early phase of cultures. ExtrasynapticNMDA-Rs may play important roles, early as factors of motoneuronsurvival (Brennemanet al., 1990) and morphogenesis (Kalb, 1994),and also later in signal processing by the adult dendritic membrane,possibly by amplifying individual, AMPA-mediated EPSPs (Sahet al.,1989). On dissociated spinal neurons recorded between 4 and 8 DIV,Robertet al. (1998) have shown evidence of control of AMPA receptor-dependent synaptic transmission by NMDA receptors, and suggest a
© 1999 European Neuroscience Association,European Journal of Neuroscience, 11, 1324–1334
possible feedback mechanism. In this work, we confirm the existenceof NMDA receptors on the motoneuron membrane, probably at extra-synaptic sites, and we also observed a modification of spontaneoussynaptic noise by APV, but our data on evoked EPSPs tend to favour arole for presynaptic NMDA receptors.
Acknowledgements
We wish to thank Prof. H.-R. Lu¨scher and Dr M. E. Larkum for allowing usto use their fitting program to analyse spontaneous EPSPs on our recordings.We are indebted to Dr S. Tyc-Dumont and Prof. H.-R. Lu¨scher for valuablecomments on the manuscript. This work was supported by doctoral grantsfrom Ministere de la Recherche et de la Technologie to T.L. and Ministe`redes Affaires E´ trangeres and French Embassay in Kiev to A.I., and by a grantfrom the Franco-Hungarian Scientific Cooperation Program BALATON 95,project No. 100.
Abbreviations
AMPA, DL-α-amino-3-hydroxy-5-methyl-4-isoxazole-propionic acid; AMPA-Rs, AMPA receptor subtypes; APV, (1/–)2-amino-5-phosphonovaleric acid;CNQX, 6-cyano-7-nitroquinoxaline-2,3-dione; DIV, daysin vitro; EAA, excit-atory amino acids; EPSP, excitatory postsynaptic potential; GBSS, Gey’sbalanced salt solution; GLU-Rs, glutamate receptor subtypes; GYKI 53784(or LY303070), (–)1-(4-aminophenyl)-3-methyl-carbamoyl-4-methyl-7,8-methylenedioxy-3,4-dihydro-5H-2,3-benzo-diazepine; IPSPs, inhibitory post-synaptic potentials; KAI-Rs, kainate receptor subtypes; NMDA,N-methyl-D-aspartate; NMDA-Rs, NMDA receptor subtypes; TTX, tetrodotoxin.
References
Bekkers, J.M. & Stevens, C.F. (1989) NMDA and non-NMDA receptors areco-localized at individual excitatory synapses in cultured rat hippocampus.Nature, 341, 230–233.
Bleakman, D., Ballyk, B.A., Schoepp, D.D., Palmer, A.J., Bath, C.P., Sharpe,E.F., Wooley, M.L., Bufton, H.R., Kamboj, R.K., Tarnawa, I. & Lodge, D.(1996) Activity of 2,3-benzodiazepines at native rat and recombinanthuman glutamate receptorsin vitro: stereospecificity and selectivity profiles.Neuropharmacology,35, 1689–1702.
Brenneman, D.E., Forsythe, I.D., Nicol, T. & Nelson, P.G. (1990)N-methyl-D-aspartate receptors influence neuronal survival in developing spinal cordcultures.Brain Res. Dev., 51, 63–68.
Castillo, P.E., Malenka, R.C. & Nicoll, R.A. (1997) Kainate receptors mediatea slow postsynaptic current in hippocampal CA3 neurones.Nature, 388,182–186.
Chandler, S.H. (1989) Evidence for excitatory amino acid transmissionbetween mesencephalic nucleus of V afferents and jaw-closer motoneuronsin the guinea pig.Brain Res., 477, 252–264.
Collingridge, G.L. & Lester, R.A. (1989) Excitatory amino acid receptors inthe vertebrate central nervous system.Pharmacol. Rev., 41, 143–210.
Dale, N. & Roberts, A. (1985) Dual-component amino-acid-mediated synapticpotentials: excitatory drive for swimming in Xenopus embryos.J. Physiol.(Lond.), 363, 35–59.
Davies, J., Francis, A.A., Jones, A.W. & Watkins, J.C. (1981) 2-Amino-5-phosphonovalerate (2APV), a potent and selective antagonist of amino acid-induced and synaptic excitation.Neurosci. Lett., 21, 77–81.
Donevan, S.D. & Rogawski, M.A. (1993) GYKI 52466, a 2,3-benzodiazepine,is a highly selective, noncompetitive antagonist of AMPA/kainate receptorresponses.Neurone, 10, 51–59.
Durand, G.M., Kovalchuk, Y. & Konnerth, A. (1996) Long-term potentiationand functional synapse induction in developing hippocampus.Nature, 381,71–75.
Durand, J., Engberg, I. & Tyc-Dumont, S. (1987)L-Glutamate andN-methyl-D-aspartate actions on membrane potential and conductance of cat abducensmotoneurones.Neurosci. Lett., 79, 295–300.
Engberg, I., Tarnawa, I., Durand, J. & Ouardouz, M. (1993) An analysis ofsynaptic transmission to motoneurones in the cat spinal cord using a newselective receptor blocker.Acta Physiol. Scand., 148, 97–100.
Eustache, I. & Gueritaud, J.P. (1995) Electrical properties of embryonic ratbrainstem motoneurones in organotypic slice culture.Brain Res. Dev., 86,187–202.

1334 T. Launeyet al.
Eustache, I., Seyfritz, N. & Gueritaud, J.P. (1994) Effects of insulin-likegrowth factors on organotypic cultures of embryonic rat brainstem slicesand skeletal muscle fibres.Dev. Brain Res., 81, 284–292.
Frandsen, A. & Schousboe, A. (1993) Excitatory amino acid-mediatedcytotoxicity and calcium homeostasis in cultured neurones.J. Neurochem.,60, 1202–1211.
Funk, G.D., Johnson, S.M., Smith, J.C., Dong, X.W., Lai, J. & Feldman, J.L.(1997) Functional respiratory rhythm generating networks in neonatal micelacking NMDAR1 gene. J. Neurophysiol., 78, 1414–1420.
Gahwiler, B.H., Thompson, S.M., Audinat, E. & Robertson, R.T. (1991)Organotypic slice culture of neural tissue. In Banker, G. & Goslin, K. (eds),Culturing Nerve Cells. MIT Press, Cambridge, MA, USA, pp. 379–411.
Giffard, R.G., Monyer, H., Christine, C.W. & Choi, D.W. (1990) Acidosisreduces NMDA receptor activation, glutamate neurotoxicity, and oxygen-glucose deprivation neuronal injury in cortical cultures.Brain Res., 506,339–342.
Gueritaud, J.P. & Seyfritz, N. (1992) An organotypic co-culture of embryonicrat brainstem and tongue or skeletal muscle.Eur. J. Neurosci., 4,616–627.
Henneman, E., Lu¨scher, H.R. & Mathis, J. (1984) Simultaneously active andinactive synapses of single Ia fibres on cat spinal motoneurones.J. Physiol.(Lond.), 352, 147–161.
Hollmann, M. & Heinemann, S. (1994) Cloned glutamate receptors.Annu.Rev. Neurosci., 17, 31–108.
Hori, Y. & Kanda, K. (1996) Developmental alterations in NMDA receptor-mediated currents in neonatal rat spinal motoneurones.Neurosci. Lett., 205,99–102.
Jahr, C.E. & Yoshioka, K. (1986) Ia afferent excitation of motoneurones inthe in vitro new-born rat spinal cord is selectively antagonized by kynurenate.J. Physiol.(Lond.), 370, 515–530.
Jakowec, M.W., Fox, A.J., Martin, L.J. & Kalb, R.G. (1995) Quantitative andqualitative changes in AMPA receptor expression during spinal corddevelopment.Neuroscience, 67, 893–907.
Jiang, Z.G., Shen, E. & Dun, N.J. (1990) Excitatory and inhibitory transmissionfrom dorsal root afferents to neonate rat motoneurones in vitro.Brain Res.,535, 110–118.
Kalb, R.G. (1994) Regulation of motor neurone dendrite growth by NMDAreceptor activation.Development, 120, 3063–3071.
Larkum, M.E., Launey, T., Dityatev, A. & Lu¨scher, H.-R. (1998) Integrationof postsynaptic potentials in dendrites of motoneurons of rat spinal cordslice cultures.J. Neurophysiol., 80, 924–935.
Launey, T., Eustache, I., Ferrand, N. & Gueritaud, J.P. (1997a) Synaptic inputson rat brainstem motoneurones in organotypic slice culture.Neuroreport,8, 3287–3291.
Launey, T., Ivanov, A., Ferrand, N. & Gueritaud, J.P. (1997b) Developing ratbrainstem motoneurones in organotypic cultures express calcium permeableAMPA gated receptors.Brain Res. Dev., 781, 148–158.
Launey, T., Kapus, G., Seyfritz, N. & Gueritaud, J.P. (1996) Excitatory aminoacids responses of embryonic rat brainstem motoneurones in slice culture.Eur. J. Neurosci., 9 (Suppl.), 75.
Lerma, J., Morales, M., Vicente, M.A. & Herreras, O. (1997) Glutamatereceptors of the kainate type and synaptic transmission.TINS, 20, 9–12.
Lodge, D., Bond, A., O’Neill, M.J., Hicks, C.A. & Jones, M.G. (1996)Stereoselective effects of 2,3-benzodiazepines in vivo: electrophysiologyand neuroprotection studies.Neuropharmacology, 35, 1681–1688.
McCrimmon, D.R., Smith, J.C. & Feldman, J.L. (1989) Involvement ofexcitatory amino acids in neurotransmission of inspiratory drive to spinalrespiratory motoneurons.J. Neurosci., 9, 1910–1921.
Nelson, P.G., Pun, R.Y. & Westbrook, G.L. (1986) Synaptic excitation incultures of mouse spinal cord neurones: receptor pharmacology andbehaviour of synaptic currents.J. Physiol.(Lond.), 372, 169–190.
Nurcombe, V. & Bennett, M.R. (1982) Evidence for neuron-survival andneurite-promoting factors from skeletal muscle: their effects on embryonicspinal cord.Neurosci. Lett., 34, 89–93.
© 1999 European Neuroscience Association,European Journal of Neuroscience, 11, 1324–1334
O’Brien, J.A., Isaacson, J.S. & Berger, A.J. (1997) NMDA and non-NMDAreceptors are co-localized at excitatory synapses of rat hypoglossalmotoneurones.Neurosci. Lett., 227, 5–8.
O’Brien, R.J. & Fischbach, G.D. (1986) Modulation of embryonic chickmotoneurone glutamate sensitivity by interneurones and agonists.J. Neurosci., 6, 3290–3296.
Openheim, R.W. (1989) The neurotrophic theory and naturally occuringmotoneuron death.Trends Neurosci., 12, 252–255.
Ouardouz, M. & Durand, J. (1994) Involvement of AMPA receptors intrigeminal post-synaptic potentials recorded in rat abducens motoneuronesin vivo. Eur. J. Neurosci., 6, 1662–1668.
Paxinos, G., To¨rk, I., Tecott, L.H. & Valentino, A.L.R. (1991)Atlas of theDeveloping Rat Brain. Academic Press, San Diego, CA, USA.
Pinco, M. & Lev-Tov, A. (1994) Synaptic transmission between ventrolateralfuniculus axons and lumbar motoneurones in the isolated spinal cord of theneonatal rat.J. Neurophysiol., 72, 2406–2419.
Pun, R.Y., Neale, E.A., Guthrie, P.B. & Nelson, P.G. (1986) Active andinactive central synapses in cell culture.J. Neurophysiol., 56, 1242–1256.
Rall, W. (1964) Theoretical significance of dendrites for neuronal input–outputrelations. In Reiss, R. (ed.),Neuronal Theory and Modeling. StanfordUniversity Press, Stanford, USA, pp. 73–97.
Reckling, J.C. (1992) Interaction between thyrotropin-releasing hormone(TRH) and NMDA-receptor mediated responses in hypoglossalmotoneurones.Brain Res., 578, 289–296.
Robert, A., Black, J.A. & Waxman, S.G. (1998) Endogenous NMDA-receptor activation regulates glutamate release in cultures spinal neurons.J. Neurophysiol., 80, 196–208.
Sah, P., Hestrin, S. & Nicoll, R.A. (1989) Tonic activation of NMDA receptorsby ambient glutamate enhances excitability of neurones.Science, 246,815–818.
Spenger, C., Braschler, U.F., Streit, J. & Lu¨scher, H.-R. (1991) An organotypicspinal cord-dorsal root ganglion-skeletal muscle co-culture of embryonicrat. I. The morphological correlates of the synaptic reflex arc.Eur. J.Neurosci., 3, 1037–1053.
Streit, J., Spenger, C. & Lu¨scher, H.R. (1991) An organotypic spinal cord-dorsal root ganglion-skeletal muscle coculture of embryonic rat. II.Functional evidence for the formation of spinal reflex arcsin vitro. Eur. J.Neurosci., 3, 1054–1068.
Trueblood, P.R., Levine, M.S. & Chandler, S.H. (1996) Dual-componentexcitatory amino acid-mediated responses in trigeminal motoneurones andtheir modulation by serotonin in vitro.J. Neurophysiol., 76, 2461–2473.
Vignes, M. & Collingridge, G.L. (1997) The synaptic activation of kainatereceptors.Nature, 388, 179–182.
Vizi, E.S., Mike, A. & Tarnawa, I. (1996) 2,3-Benzodiazepines (GYKI 52466and analogs): negative allosteric modulators of AMPA receptors.CNS DrugRev., 2, 91–126.
Walmsley, B. & Bolton, P.S. (1994) Anin vivo pharmacological study ofsingle group Ia fiber contacts with motoneurones in the cat spinal cord.J .Physiol.(Lond.), 481, 731–741.
Watkins, J.C. & Evans, R.H. (1981) Excitatory amino acid transmitters.Annu.Rev. Pharmacol. Toxicol., 21, 165–204.
Widmer, H., Dreifuss, J.J. & Raggenbass, M. (1992)N-methyl-D-aspartateand vasopressin activate distinct voltage-dependent inward currents in facialmotoneurones.Brain Res., 593, 215–220.
Wilding, T.J. & Huettner, J.E. (1995) Differential antagonism of alpha-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid-preferring and kainate-preferring receptors by 2,3-benzodiazepines.Mol. Pharmacol., 47, 582–587.
Ziskind-Conhaim, L. (1990) NMDA receptors mediate poly- and monosynapticpotentials in motoneurones of rat embryos.J. Neurosci., 10, 125–135.
Zorumski, C.F., Yamada, K.A., Price, M.T. & Olney, J.W. (1993) Abenzodiazepine recognition site associated with the non-NMDA glutamatereceptor.Neurone, 10, 61–67.
Zorumski, C.F., Yang, J. & Fischbach, G.D. (1989) Calcium-dependent, slowdesensitization distinguishes different types of glutamate receptors.CellMol. Neurobiol., 9, 95–104.




















