
Axon collaterals of cat medial rectus motoneurons
8
Brain Research, 174 (1979) 153-160 153 © Elsevier/North-Holland Biomedical Press Short Communications Axon collaterals of cat medial rectus motoneurons C. EVINGER, R. BAKER and R. A. McCREA Department of Physiology and Biophysics, New York University Medical Center, 550 First Avenue, New York, N.Y. 10016 (U.S.A.) (Accepted May 31st, 1979) Unlike studies in the mammalian spinal cord, there is neither anatomical nor physiological evidence for the presence of axon collaterals from cranial motoneurons. Anatomical studies have shown extensive collateralization of spinal a-motoneu- rons 7-1°As,1a,23,24 and physiological evidence has demonstrated recurrent collateral activation of small neurons (Renshaw cells; lg,14) that in turn produce inhibition of a- motoneurons and spinal interneurons20,21,2L Comparable recurrent synaptic effects have not been observed in cranial motoneurons. In particular, extensive electro- physiological investigations of extraocular motoneurons have failed to detect recur- rent synaptic effectsz-6. In support of the latter observations, intracellular horse- radish peroxidase (HRP) studies of abducens motoneurons did not reveal axon collaterals 4 despite the technique's ability to demonstrate their presence in the spinal cord 7-10. Somewhat surprisingly, axon collaterals were observed following HRP injection into cat medial rectus motoneurons. Although these collaterals were recurrent to the oculomotor complex, they did not produce recurrent synaptic effects in medial rectus or other extraocular motoneurons. The experiments were performed on 5 cats anesthetized with sodium pento- barbital (35 mg/kg~. Additional anesthetic and fluids were administered throughout the experiment to maintain stable physiological conditions. Following surgery, all animals were immobilized with Flaxedil and artificially respired. Electrodes were implanted to permit bilateral electrical stimulation of the VIIIth nerve. The nerve to the medial rectus muscle was dissected free within the orbit and placed on small silver hook electrodes and two silver ball electrodes were placed in the posterior part of the orbit in order to activate the entire oculomotor nerve. The oculomotor nucleus was approached through the superior colliculus following removal of the posterior cerebral cortex and hippocampus (details in ref. 3). Glass microelectrodes were filled with a 7 HRP (Sigma VI) solution (in equal parts of 1.0 M KC1 and 0.1 M Tris buffer) with pH adjusted to 7.4. After physiologically identifying medial rectus motoneurons, HRP was injected with steps of positive current (10-30 nA) applied through the micro-
-
Upload
independent -
Category
Documents
-
view
6 -
download
0
Transcript of Axon collaterals of cat medial rectus motoneurons

Brain Research, 174 (1979) 153-160 153 © Elsevier/North-Holland Biomedical Press
Short Communica t ions
Axon collaterals of cat medial rectus motoneurons
C. EVINGER, R. BAKER and R. A. McCREA
Department of Physiology and Biophysics, New York University Medical Center, 550 First Avenue, New York, N.Y. 10016 (U.S.A.)
(Accepted May 31st, 1979)
Unlike studies in the mammalian spinal cord, there is neither anatomical nor physiological evidence for the presence of axon collaterals from cranial motoneurons. Anatomical studies have shown extensive collateralization of spinal a-motoneu- rons 7-1°As,1a,23,24 and physiological evidence has demonstrated recurrent collateral activation of small neurons (Renshaw cells; lg,14) that in turn produce inhibition of a- motoneurons and spinal interneurons20,21,2L Comparable recurrent synaptic effects have not been observed in cranial motoneurons. In particular, extensive electro- physiological investigations of extraocular motoneurons have failed to detect recur- rent synaptic effects z-6. In support of the latter observations, intracellular horse- radish peroxidase (HRP) studies of abducens motoneurons did not reveal axon collaterals 4 despite the technique's ability to demonstrate their presence in the spinal cord 7-10. Somewhat surprisingly, axon collaterals were observed following HRP injection into cat medial rectus motoneurons. Although these collaterals were recurrent to the oculomotor complex, they did not produce recurrent synaptic effects in medial rectus or other extraocular motoneurons.
The experiments were performed on 5 cats anesthetized with sodium pento- barbital (35 mg/kg~. Additional anesthetic and fluids were administered throughout the experiment to maintain stable physiological conditions. Following surgery, all animals were immobilized with Flaxedil and artificially respired. Electrodes were implanted to permit bilateral electrical stimulation of the VIIIth nerve. The nerve to the medial rectus muscle was dissected free within the orbit and placed on small silver hook electrodes and two silver ball electrodes were placed in the posterior part of the orbit in order to activate the entire oculomotor nerve. The oculomotor nucleus was approached through the superior colliculus following removal of the posterior cerebral cortex and hippocampus (details in ref. 3). Glass microelectrodes were filled with a 7 HRP (Sigma VI) solution (in equal parts of 1.0 M KC1 and 0.1 M Tris buffer) with pH adjusted to 7.4. After physiologically identifying medial rectus motoneurons, HRP was injected with steps of positive current (10-30 nA) applied through the micro-

154
'\.?
Fig. 1. Morphophysiological profile of a medial rectus motoneuron. The reconstructed somadendritic tree and axon with one collateral are shown in the center of the figure. In the upper right inset the axo~ collateral is shown with only the soma and proximal dendrites at the same scale as in the main drawing. The upper left inset shows the location of the motoneuron in the oculomotor nucleus. Electrophysio- logical records from this motoneuron are presented in the lower right inset. A: response to ipsilateral (Vi) and contralateral (Vc) vestibular nerve stimulation followed by medial rectus (MR) stimulation at axon straddling intensity. B: antidromic spike after 3 min of HRP injection. C: membrane hyper- polarization (10 nA) blocked the IS-SD invasion revealing an M-spike. Arrows indicate the absence of recurrent synaptic effects. Upper and lower traces are high (AC) and low (DC) gain, respectively. a, axon; AQ, aqueduct, c, axon collateral; d, dendrite; MLF, medial longitudinal fasciculus; MR, medial rectus nerve stimulation; PAG, periaqueductal gray; t, collateral terminals; Vc, contralateral vestibular nerve stimulation; Vi, ipsilateral vestibular nerve stimulation; 3N, oculomotor nucleus; 3n, oculomotor nerve.

155
1111111
.2mm
i 2mV ! | 20 mV J
"r'-"----F'--"-'--
Fig. 2. Morphophysiological profile of a medial rectus motoneuron. The reconstructed somadendritic tree and axon with two collaterals are shown in the center of the figure. Location of the motoneuron in the oculomotor nucleus is shown on the upper left. The lower left inset shows electrophysiological records obtained after 3 min of HRP injection. The two upper traces are the high gain (AC) records, the next two are low gain (DC) records and the lowest trace is the extracellular field potentials. MR, Vi and Vc responses are similar to that described in Fig. 1.
156

157
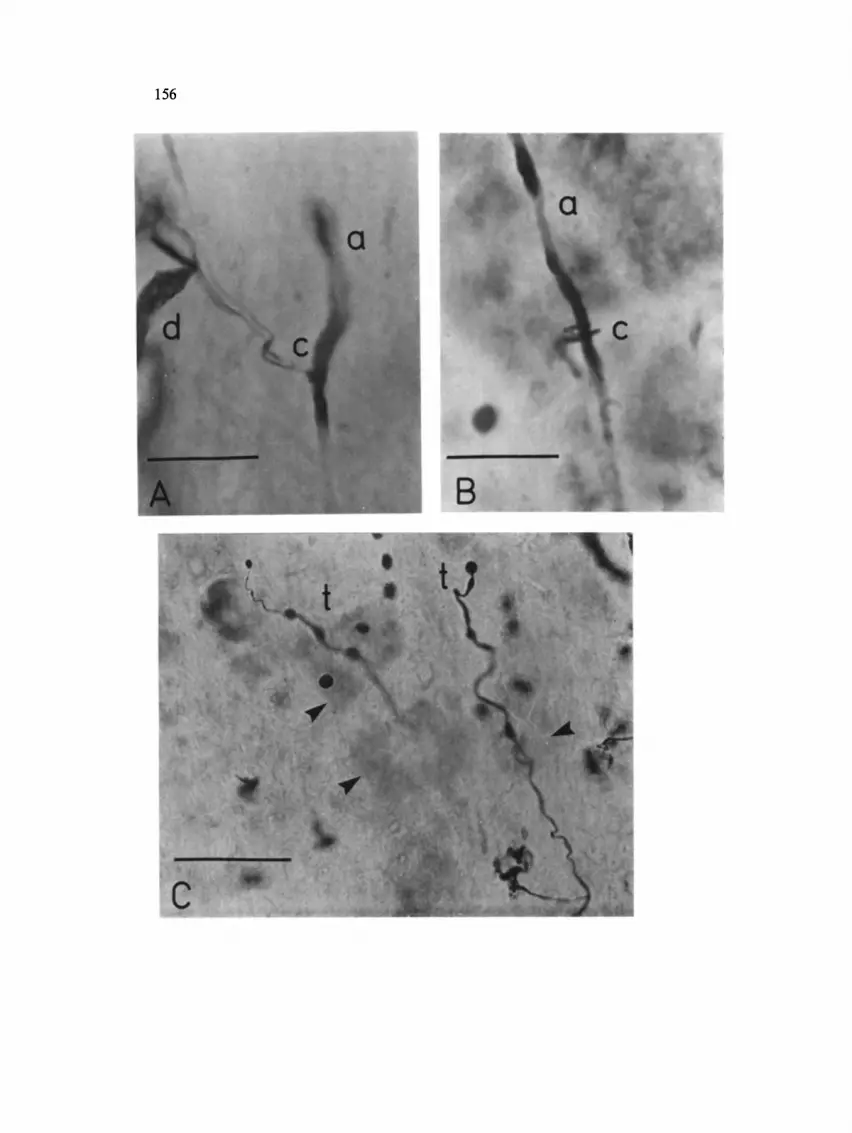
Fig. 3. Photomicrographs of axon collaterals and collateral terminals. A: axon collateral of the cell in Fig. 1. B: the dorsal axon collateral of the cell in Fig. 2. C: the axon collateral end branches and terminals labelled t in Fig. 2. The terminals are the irregularly shaped enlargements appearing through- out the collateral. D: the ventral axon collateral of the cell in Fig. 2. E: an enlargement of one of the terminals in C. F: an enlargement of 3 of the terminals shown in C. The calibrations are 10/~m for D-F, 20 pm for A-B and 50 pm for C. The arrows indicate somas shown by the Nissl counterstaining.

158
electrode at a 75 ~:o duty cycle for 2-3 min. Generally, 50 nA/min was sufficient to completely fill motoneurons. One to 3 h after staining, the animal was perfused with 1 ~ paraformaldehyde and 1.25 ~ glutaraldehyde. The brain was removed, blocked and placed in 30 ~ sucrose solution. After sinking, the midbrain was frozen and cut in 100 /~m coronal sections before reaction with the DAB method including cobalt intensificationlL Cells were reconstructed with the aid of a Zeiss microscope equipped with a drawing tube.
In this study, 6 medial rectus motoneurons were identified by employing the electrophysiological criteria described by Baker and Highstein 3 (Figs. 1A and 2). The motoneurons produced antidromic action potentials in response to medial rectus stim- ulation. In addition, they exhibited disynaptic EPSPs in response to ipsilateral vesti- bular nerve stimulation. Contralateral vestibular stimulation usually failed to produce synaptic responses. Both antidromic and orthodromic activation of medial rectus motoneurons still occurred after 50 nA min of intracellular HRP injection (Figs. 1B, 1C and 2).
Two representative medial rectus motoneurons of the 6 stained and recon- structed cells are shown in Figs. 1 and 2. Consistent with earlier anatomical findings 1,13, both cells were located rostrally in the dorsal subdivision of the oculo- motor nucleus. Their dendritic trees always extended out of the oculomotor complex. Some branches extended dorsally into the periaqueductal gray and some laterally to- wards, and in one case (Fig. 1), into the MLF. The main axon originated from the cell body in all neurons and descended to exit the nucleus via the ipsilateral IIIrd nerve (Figs. l and 2). Axon collaterals (Figs. 1 and 2c) arose from the main axon during its course through the oculomotor complex.
Photomicrographs (Fig. 3) show the recurrent axon collaterals drawn in Fig. 1 and 2. The collateral of Fig. 1 branches dorsolaterally from the parent axon and immediately bifurcates into two branches (Fig. 3A). The motoneuron in Fig. 2 had two collaterals. The dorsal collateral wrapped itself around the parent axon and followed a ventral trajectory before turning medially (Fig. 3B). The second collateral branched dorsolaterally from the parent axon, but did not bifurcate near the parent axon (Fig. 3D). The axon collaterals exhibited extensive branching. For instance, there were 13 end branches of the axon collateral in Fig. I. For the 6 medial rectus cells, the number of end branches per collateral ranged from 2 to 15. Despite such extensive branching, axon collaterals did not appear to leave the ipsilateral oculomotor nucleus. In fact, the majority of collaterals appeared to terminate within a few hundred #m of the injected motoneuron.
All collateral end branches gave rise to several irregularly shaped enlargements or swellings (Figs. 1-3). Cullheim and Kellerth 7 have demonstrated that these terminal swellings are for the most part synaptic boutons. Fig. 3C shows two axon collateral end branches with terminal swellings from the motoneuron in Fig. 2. The presence of Nissl counterstain demonstrates that the terminal swellings occur near somas and proximal dendrites (Fig. 3C). The higher magnification photomicrographs show the large size of these terminals (Fig. 3E, F). The terminal of Fig. 3E was 5 #m in diameter. Mean terminal diameter (major and minor diameter/2) ranged from 0.8 to 8

159
/zm and averaged 3.49 ± 1.46; n = 114 terminals. Cullheim and Kellerth reported the same range for axon collaterals of a-motoneurons contributing to the sciatic nerve.
Despite the fact that every medial rectus motoneuron exhibited axon collaterals with extensive terminals, within and near, the medial rectus subdivision, no evidence was found for recurrent synaptic activation of medial rectus motoneurons. Recurrent collaterals were never observed to appose the somadendritic trees of stained extra- ocular motoneurons. In addition, the electrophysiological data conclusively demonstr- ated the absence of any recurrent synaptic activation. The typical 30-40 msec after- hyperpolarization following antidromic activation at straddling intensity was uninter- rupted by synaptic potentials (Fig. 1 B; arrows). Membrane hyperpolarization (10 nA) of the medial rectus motoneuron blocked the IS-SD invasion so that maximal nerve stimulation only elicited an M-spike. Nevertheless, there were no recurrent synaptic effects superimposed on the M-spike (Fig. 1C; arrows). Previous electrophysiological studies of medial rectus motoneurons also failed to find evidence for recurrent activ- ation 3.
These results demonstrate the existence of one or more collaterals emanating f rom axons of medial rectus motoneurons. These collaterals terminate in the oculomotor nucleus, but unlike the axon collaterals of spinal motoneurons, they do not appear to activate neurons recurrent to extraocular motoneurons. I t is likely that the collaterals terminate on the oculomotor internuclear interneurons described by Maciewicz et a1.15, 16. In support of this idea, the first electrophysiological study of the oculomotor nu- cleus 2~ and our recent work (unpublished observations) have found recurrent synaptic activation of interneurons within the oculomotor nucleus following I I I rd nerve stim- ulation. I f medial rectus axon collaterals synapse on these cells, then the signal sent to other brain stem nuclei could be, in part, similar to that arriving at the extraocular muscles. This type of organization contrasts sharply with that of the abducens nucleus. The latter contains a population of motoneurons and internuclear neurons that exhibit qualitatively identical discharge patterns n , but which do not have recurrent axon collaterals 4. The contrasting organization between the internuclear neurons in the abducens and oculomotor nuclei could reflect a difference in function for the two classes of neurons. This latter suggestion is likely because vertical motoneurons in the oculomotor nucleus also have axon collaterals. This result may indicate a role for internuclear neurons in both horizontal and vertical eye movement.
Research supported by U.S.P.H.S. Grants EY-02007, NS-13742 and Fellow- ships NS-05857, NS-06163.
1 Akagi, Y., The localization of the motor neurons innervating the extraocular muscles in the oculo- motor nuclei of the cat and rabbit, using horseradish peroxidase, J. comp. Neurol., 181 (1978) 745-762.
2 Baker, R. and Highstein, S. M., Physiological identification of interneurons and motoneurons in the abducens nucleus, Brain Research, 83 (1975) 292-298.
3 Baker, R. and Highstein, S. M., The vestibular projections to medial rectus subdivision of the oculomotor complex, J. Neurophysiol., 41 (1978) 1629-1646.

160
4 Baker, R. and McCrea, R., The parabducens nucleus. In V. Wilson and H. Asanuma (Eds.), Integration in the Nervous System, Igakaku Shoin, Tokyo, 1979, pp. 97-123.
5 Baker, R. and Precht, W., Electrophysiological properties of trochlear motoneurons as revealed by lVth nerve stimulation, Exp. Brain Res., 14 (1972) 127-157.
6 Baker, R. G., Mano, N. and Shimazu, H., Intracellular recording of antidromic responses frcm abducens motoneurons in the cat, Brain Research, 15 (1969) 573-576.
7 Cullheim, S. and Kellerth, J.-O., Combined light and electron microscopic tracing of neurons, including axons and synaptic terminals, after intracellular injection of horseradish lceroxidase, Neurosci. Lett., 2 (1976) 307-313.
8 Cullheim, S. and Kellerth, J.-O., A morphological study of the axons and recurrent axon collaterals of cat sciatic a-motoneurons after intracellular staining with horseradish reroxidase, J. comp. Neurol., 178 (1978) 537-558.
9 Cullheim, S. and Kellerth, J.-O., A morphological study of the axons and recurrent axon collaterals of cat a-motoneurons supplying different hindlimb muscles, J. PhysioL (Lond.), 281 (1978) 285- 299.
10 Cullheim, S. and Kellerth, J.-O., A morphological study of the axons and recurrent axon collaterals of cat ct-motoneurons supplying different functional types of muscle unit, J. Physiol. (Lond.), 281 (1978) 301-313.
11 Delgado-Garcia, J., Baker, R. and Highstein, S. M., The activity of internuclear neurons identified within the abducens nucleus of the alert cat. In R. Baker and A. Berthoz (Eds.), Control of Gaze by Brain Stem Neurons, Elsevier, New York, 1977, 291-300.
12 Eccles, J. C., Fatt, P. and Koketsu, K., Cholinergic and inhibitory synapses in a pathway from motor-axon collaterals to motoneurons, J. PhysioL (Lond.), 126 (1954) 524-562.
13 Gacek, R. R., Localization of neurons supplying the extraocular muscles in the kitten using horseradish peroxidase, Exp. NeuroL, 44 (1974) 381-403.
14 Hultborn, H., Jankowska, E. and Lindstr6m, S., Recurrent inhibition from motor axon collaterals of transmission in the Ia inhibitory pathway to motoneurones, J. PhysioL (Lond.), 215 (1971) 391-412.
15 Maciewicz, R. J. and Spencer, R. F., Oculomotor and abducens internuclear pathway in the cat. In R. Baker and A. Berthoz (Eds.), Control of Gaze by Brain Stem Neurons, Elsevier, New York, 1977, pp. 99-108.
16 Maciewicz, R. J., Kaneko, C. R. S., Highstein, S. M. and Baker, R., Morphological identification of interneurons in the oculomotor nucleus that project to the abdncens nucleus in the cat, Brain Research, 96 (1975) 60-65.
17 McCrea, R. A., Bishop, G. A. and Kitai, S. T., Morphological and electrophysiological character- istics of projection neurons in the nucleus interpositus of the cat cerebellum, J. comp. NeuroL, 181 (1978) 397-420.
18 Prestige, M. C., lnitial collaterals ofmotor axons within the spinal cord of the cat, J. comp. Neurol., 126 (1966) 123-136.
19 Ram6n y Cajal, S., Histologie due Systdme Nerveux de l'Homme et des Vert~brEs, (reprinted 1972) Tome 1, C.S.I.C., Madrid, 1909, pp. 854-858.
20 Renshaw, B., Influence of discharge of motoneurons upon excitation of neighboring motoneurons, J. NeurophysioL, 4 (1941) 167-183.
21 Renshaw, B.,Centraleffectsofcentripetalimpulsesinaxonofspinalventralroots, J. NeurophysioL, 9 (1946) 191-204.
22 Sasaki, K., Electrophysiological studies on oculomotor neurons of the cat, Jap. J. PhysioL, 13 (1963) 287-302.
23 Scheibel, M. E. and Scheibel, A. B., Spinal motoneurons, interneurons and Renshaw cells. A Golgi study, Arch. ital. BioL, 104 (1966) 328-353.
24 Szentagothai, J., Synaptic architecture of the spinal motoneuron pool. In L. Widen (Ed.), Recent Advances in Clinical Neurophysiology, Elsevier, Amsterdam, 1967, pp. 4-19.
25 Wilson, V. J., Regulation and function of Renshaw cell discharge. In I. R. Granit (Ed.), Muscular Afferents and Motor Control Nobel Symposium, Almquist and Wiskell, Stockholm, 1966, pp. 317-329.




















