
Evaluation of a predictive in vitro Leydig cell assay for anti-androgenicity of phthalate esters in...
8
Evaluation of a predictive in vitro Leydig cell assay for anti-androgenicity of phthalate esters in the rat Pergentino Balbuena, Jerry Campbell Jr., Harvey J. Clewell III, Rebecca A. Clewell ⇑ The Hamner Institutes for Health Sciences, Research Triangle Park, NC 27709, United States article info Article history: Received 9 January 2013 Accepted 31 March 2013 Available online 8 April 2013 Keywords: Phthalates R2C MEHP Antiandrogenicity MEHP metabolites MBP abstract An in vitro assay using the rat Leydig cell line R2C was evaluated for its ability to quantitatively predict inhibition of testosterone synthesis. Results obtained for endocrine active phthalates (MEHP, MBP), and inactive phthalates (MMP and MEP) were highly consistent with in vivo results based on tissue and media concentrations. Statistically significant inhibition of testosterone synthesis (p < 0.05, 1-way ANOVA) was observed at 1 lM MBP and 3 lM MEHP, while MEP and MMP did not affect inhibition of testosterone syn- thesis until much higher concentrations (100 lM). Concentrations causing 50% inhibition of testoster- one synthesis for MBP and MEHP (3 and 6 lM respectively), were similar to in vivo values (3 and 7 lM). The R2C assay was used to determine the relative potency of 14 structurally diverse monoesters and oxi- dative metabolites of MEHP. Monoesters with alkyl chains 4–5 carbons in length had the highest potency for testosterone inhibition, while 0–2 carbon alkyl chains were least potent. Phase I metabolism did not completely inactivate MEHP, underscoring the need for metabolism data when interpreting in vitro phar- macodynamic data. This steroid inhibition assay provides a predictive in vitro alternative to expensive and timeconsuming developmental rat studies for phthalate-induced antiandrogenicity. Ó 2013 Elsevier Ltd. All rights reserved. 1. Introduction The phthalate diesters (phthalates) are a class of chemicals in which phthalic acid is esterified to two alkyl chains that may vary in carbon chain length and structure (Fig. 1). The phthalates are used to increase flexibility and longevity of plastics and are present in a variety of industrial and consumer products, such as industrial flooring, sealants, cosmetics, pharmaceuticals, children’s toys and food packaging (Hernandez-Diaz et al., 2009, Christen et al., 2012). Because they are not covalently bound to the plastics, the phthalates can leach into food and consumer products from plastic containers. Upon ingestion, the diester phthalates are quickly hydrolyzed into their monoester metabolites (Rowland et al., 1977; White et al., 1980), which are generally considered the tox- icologically active species (Clewell et al., 2010; Jones et al., 1993; Parks et al., 2000). High doses of some phthalates adversely affect male rat sexual development when administered during gestation. Perinatal administration of di-n-butyl phthalate (DBP: >100 mg/kg/day) and di-2,4-ethylhexyl phthalate (DEHP: >300 mg/kg/day) cause re- duced anogenital distance (AGD), increased nipple retention, hypo- spadias (malformed penis), cryptorchid (undescended) testes, and incomplete development or agenesis of some reproductive organs in male rats (Clewell et al., 2013a; Gray et al., 2000; Lehmann et al., 2004; Mylchreest et al., 1998, 1999). Many of these reproductive effects are thought to result from inhibition of testosterone synthe- sis in the fetal rat testes by the monoester phthalates (Akingbemi et al., 2004; Oishi and Hiraga, 1980; Parks et al., 2000). In vivo and in vitro studies confirm that the monoester metabolites of the endocrine active phthalates DBP (monobutyl phthalate; MBP) and DEHP (mono-2-ethylhexyl phthalate; MEHP) reduce testoster- one synthesis in vivo and in cultured Leydig cells (Clewell et al., 2010; Jones et al., 1993; Kwack et al., 2009; Lehmann et al., 2004; Liu et al., 2005). It is generally accepted that the downstream metabolites resulting from glucuronide conjugation and oxidative metabolism are inactive (Ema and Miyawaki, 2001; Silva et al., 2003). Recently, however, it has been suggested that one or more of the oxidative metabolites of MEHP may have anti-androgenic activity (Chauvigné et al., 2009; Koch et al., 2005). Although the endocrine active phthalates (DEHP, DBP, etc.) ap- pear to act by the same mechanism (Liu et al., 2005), not all phtha- lates are endocrine active (Gray et al., 2000). The ability of the phthalates to inhibit testosterone synthesis and disrupt male rat reproductive tract development is determined by the structure of alkyl side chains (Gray et al., 2000; Hannas et al., 2011; Liu et al., 2005). The more potent phthalates have 3–5 carbons in their alkyl chains. Dipentyl phthalate (DPP; C5) is the most potent of the 0887-2333/$ - see front matter Ó 2013 Elsevier Ltd. All rights reserved. http://dx.doi.org/10.1016/j.tiv.2013.03.015 ⇑ Corresponding author. Address: The Hamner Institutes for Health Sciences, 6 Davis Drive, Research Triangle Park, NC 27709, United States. Tel.: +1 919 558 1307; fax: +1 919 558 1200. E-mail address: [email protected] (R.A. Clewell). Toxicology in Vitro 27 (2013) 1711–1718 Contents lists available at SciVerse ScienceDirect Toxicology in Vitro journal homepage: www.elsevier.com/locate/toxinvit
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Evaluation of a predictive in vitro Leydig cell assay for anti-androgenicity of phthalate esters in...

Toxicology in Vitro 27 (2013) 1711–1718
Contents lists available at SciVerse ScienceDirect
Toxicology in Vitro
journal homepage: www.elsevier .com/locate / toxinvi t
Evaluation of a predictive in vitro Leydig cell assay for anti-androgenicityof phthalate esters in the rat
0887-2333/$ - see front matter � 2013 Elsevier Ltd. All rights reserved.http://dx.doi.org/10.1016/j.tiv.2013.03.015
⇑ Corresponding author. Address: The Hamner Institutes for Health Sciences,6 Davis Drive, Research Triangle Park, NC 27709, United States. Tel.: +1 919 5581307; fax: +1 919 558 1200.
E-mail address: [email protected] (R.A. Clewell).
Pergentino Balbuena, Jerry Campbell Jr., Harvey J. Clewell III, Rebecca A. Clewell ⇑The Hamner Institutes for Health Sciences, Research Triangle Park, NC 27709, United States
a r t i c l e i n f o
Article history:Received 9 January 2013Accepted 31 March 2013Available online 8 April 2013
Keywords:PhthalatesR2CMEHPAntiandrogenicityMEHP metabolitesMBP
a b s t r a c t
An in vitro assay using the rat Leydig cell line R2C was evaluated for its ability to quantitatively predictinhibition of testosterone synthesis. Results obtained for endocrine active phthalates (MEHP, MBP), andinactive phthalates (MMP and MEP) were highly consistent with in vivo results based on tissue and mediaconcentrations. Statistically significant inhibition of testosterone synthesis (p < 0.05, 1-way ANOVA) wasobserved at 1 lM MBP and 3 lM MEHP, while MEP and MMP did not affect inhibition of testosterone syn-thesis until much higher concentrations (�100 lM). Concentrations causing 50% inhibition of testoster-one synthesis for MBP and MEHP (3 and 6 lM respectively), were similar to in vivo values (3 and 7 lM).The R2C assay was used to determine the relative potency of 14 structurally diverse monoesters and oxi-dative metabolites of MEHP. Monoesters with alkyl chains 4–5 carbons in length had the highest potencyfor testosterone inhibition, while 0–2 carbon alkyl chains were least potent. Phase I metabolism did notcompletely inactivate MEHP, underscoring the need for metabolism data when interpreting in vitro phar-macodynamic data. This steroid inhibition assay provides a predictive in vitro alternative to expensiveand timeconsuming developmental rat studies for phthalate-induced antiandrogenicity.
� 2013 Elsevier Ltd. All rights reserved.
1. Introduction
The phthalate diesters (phthalates) are a class of chemicals inwhich phthalic acid is esterified to two alkyl chains that may varyin carbon chain length and structure (Fig. 1). The phthalates areused to increase flexibility and longevity of plastics and are presentin a variety of industrial and consumer products, such as industrialflooring, sealants, cosmetics, pharmaceuticals, children’s toys andfood packaging (Hernandez-Diaz et al., 2009, Christen et al.,2012). Because they are not covalently bound to the plastics, thephthalates can leach into food and consumer products from plasticcontainers. Upon ingestion, the diester phthalates are quicklyhydrolyzed into their monoester metabolites (Rowland et al.,1977; White et al., 1980), which are generally considered the tox-icologically active species (Clewell et al., 2010; Jones et al., 1993;Parks et al., 2000).
High doses of some phthalates adversely affect male rat sexualdevelopment when administered during gestation. Perinataladministration of di-n-butyl phthalate (DBP: >100 mg/kg/day)and di-2,4-ethylhexyl phthalate (DEHP: >300 mg/kg/day) cause re-duced anogenital distance (AGD), increased nipple retention, hypo-
spadias (malformed penis), cryptorchid (undescended) testes, andincomplete development or agenesis of some reproductive organsin male rats (Clewell et al., 2013a; Gray et al., 2000; Lehmann et al.,2004; Mylchreest et al., 1998, 1999). Many of these reproductiveeffects are thought to result from inhibition of testosterone synthe-sis in the fetal rat testes by the monoester phthalates (Akingbemiet al., 2004; Oishi and Hiraga, 1980; Parks et al., 2000). In vivoand in vitro studies confirm that the monoester metabolites ofthe endocrine active phthalates DBP (monobutyl phthalate; MBP)and DEHP (mono-2-ethylhexyl phthalate; MEHP) reduce testoster-one synthesis in vivo and in cultured Leydig cells (Clewell et al.,2010; Jones et al., 1993; Kwack et al., 2009; Lehmann et al.,2004; Liu et al., 2005). It is generally accepted that the downstreammetabolites resulting from glucuronide conjugation and oxidativemetabolism are inactive (Ema and Miyawaki, 2001; Silva et al.,2003). Recently, however, it has been suggested that one or moreof the oxidative metabolites of MEHP may have anti-androgenicactivity (Chauvigné et al., 2009; Koch et al., 2005).
Although the endocrine active phthalates (DEHP, DBP, etc.) ap-pear to act by the same mechanism (Liu et al., 2005), not all phtha-lates are endocrine active (Gray et al., 2000). The ability of thephthalates to inhibit testosterone synthesis and disrupt male ratreproductive tract development is determined by the structure ofalkyl side chains (Gray et al., 2000; Hannas et al., 2011; Liu et al.,2005). The more potent phthalates have 3–5 carbons in their alkylchains. Dipentyl phthalate (DPP; C5) is the most potent of the

Fig. 1. Generic structure for the diester and monoester phthalates. Hydrolysis of thediester phthalates by intestinal lipases is rapid and nearly complete prior tosystemic distribution (White et al., Rowland et al.). R indicates alkyl chains P1Carbon. Alkyl side chains of the diesters are not necessarily identical depending onthe diester.
1712 P. Balbuena et al. / Toxicology in Vitro 27 (2013) 1711–1718
phthalates tested in vivo (Hannas et al., 2011; Howdeshell et al.,2008). However, since dose–response and pharmacokinetic dataare only available for a few phthalates in vivo (Clewell et al.,2013b; Howdeshell et al., 2008; Hannas et al., 2011; Lehmannet al., 2004; Mylchreest et al., 1999), it is difficult to predict the rel-ative potency of untested phthalates based on available animalstudies.
The extensive use of phthalates in consumer products makesthe study of the dose–response and the mechanism of action ofthe phthalates a priority. The current recommended method foridentifying endocrine active phthalates is a rodent developmentalstudy, wherein pregnant rats are administered diester phthalatesduring gestation and lactation and alterations in the reproductivetract of the post-pubertal male offspring are evaluated (Boberget al., 2011; Clewell et al., 2013b; Fisher et al., 2003; Foster et al.,2001; Gray et al., 2000; Gray et al., 2009; Plummer et al., 2007).Since rats produce multiple offspring per litter, the number of ani-mals used and the cost for these types of studies are prohibitive ofin-depth dose–response and mechanistic studies. Thus, the devel-opment of in vitro assays to replace in vivo rat studies of phthal-ate-mediated anti-androgenicity is an important step in reducingthe cost and use of animals for phthalate screening and mechanis-tic studies.
A mouse Leydig cell (MA-10) assay was previously shown to bea useful tool for testing anti-androgenic activity of the phthalateesters in vitro, showing a high degree of correlation with inhibitionof testosterone synthesis in the fetal rat (Clewell et al., 2010).Although this assay proved reliable, two important differences ex-ist between this cell line and the in vivo models for phthalate ef-fects. MA-10s are derived from adult mouse testes and requireluteinizing hormone (LH) receptor stimulation to induce testoster-one production. In contrast, in vivo anti-androgenic effects are ob-served in the male rat, not the mouse. Furthermore, LH does notregulate testosterone in the rat fetus during development, and istherefore not required for phthalate inhibition of testosterone inthe fetal testes. In this paper, we evaluated the use of a rat Leydigcell line (R2C) to predict inhibition of testosterone synthesis in fe-tal rats. The R2C cell line is derived from a Leydig cell tumor from a2-month-old male Wistar-Furth rat. Like the fetal rat Leydig cell,the R2Cs do not require either hormonal stimulation or cyclicadenosine monophosphate (cAMP) induction for testosteronesecretion (Freeman, 1987; Stocco and Chen, 1991), though this cellline was derived from an adult, not fetal, rat.
R2Cs have been used to assess the effects of various compoundson cell physiology and to some extent, the steroidogenic pathway(Freeman, 1987; Rao et al., 2003). Furthermore, some studies haveshown that phthalates can inhibit steroid hormone production in
the R2Cs, though these studies measured the testosterone precur-sor progesterone for a single phthalate (MEHP) and not testoster-one itself (Gazouli et al., 2002). To date, no effort has been madeto show whether the inhibition observed in the R2C cells is indic-ative of an in vivo response in rat.
The goal of the current study was to determine whether the R2Ccell line, which is a more appropriate in vitro model for the rat,would provide an in vitro assay for phthalate inhibition that is asrobust as the MA-10 cell assay, and more importantly, that willcorrelate with in vivo effects observed in the fetal rat. The R2C tes-tosterone synthesis assay was tested with several phthalates,including known ‘‘endocrine active’’ phthalates (MEHP, MBP, andmono-n-octyl phthalate; MnOP), as well as known ‘‘inactive’’ com-pounds (monoethyl phthalate; MEP and monomethyl phthalate;MMP). Inhibition of testosterone synthesis in the R2C cell linewas then compared to data from MA-10 cells and fetal ratsin vivo. The R2C assay was then used for two predictive assess-ments. First, the potency of a battery of phthalates with differentalkyl chain structures was evaluated for inhibition of testosteronesynthesis, including 14 commercially available monoester phtha-lates. Second, the role of Phase I (P450) metabolism in detoxifica-tion of the phthalate monoesters was evaluated by testing theability of MEHP and its oxidative metabolites to inhibit testoster-one synthesis.
2. Methods
2.1. Chemicals
Mono-n-octyl phthalate (MnOP), monobenzyl phthalate(MBeP), mono-2-ethylhexyl phthalate (MEHP), monobutyl phthal-ate (MBP), monoethyl phthalate (MEP), and monomethyl phthalate(MMP) were purchased from AccuStandard, Inc. (New Haven, CT).DL-mono-1-methylhexyl phthalate (MMHP), mono(1,2-dimethyl-propyl) phthalate (MdMPP), (+)-mono(1,2,2-trimethylpropyl)phthalate (MtMPP), mono-n-hexyl phthalate (MnHP), (+)-mono-1-tert-butyl-3-methylbutyl phthalate (MtBMBP), DL-mono-1-tert-butylpentyl phthalate (MtBPP), and (�)mono-2,2-dimethyl-1-iso-propylpropyl phthalate (MdMIPP) were purchased from the rarechemical library at Sigma Aldrich Co., LLC (St. Louis, MO). Mono-(2-ethyl-5-carboxy-pentyl) phthalate (5-Cx-MEPP), mono-(2-ethyl-5-hydroxyhexyl) phthalate (5-Hx-MEHP), mono-[(2-car-boxymethyl) hexyl] phthalate (2-Cx-MMHP), mono-(2-ethyl-5-ox-ohexyl) phthalate (5-Oxo-MEHP), monoisodecyl phthalate (MiDP),and monopentyl phthalate (MPP) were purchased from CambridgeIsotopes, Inc. (Andover, MA). Phthalic acid (PA) was purchasedfrom Sigma–Aldrich Co., LLC (St Louis, MO). Human luteinizinghormone (whole molecule, >95% purity) was purchased fromScripps Laboratories (San Diego, CA).
2.2. Cell culture
The mouse Leydig tumor cell line (MA-10) was generously pro-vided by Dr. Mario Ascoli (Department of Pharmacology, Universityof Iowa, College of Medicine, Iowa City). MA-10 cell culture wasperformed as previously described (Ascoli, 1981; Clewell et al.,2010). Briefly, 150 mm � 20 mm Polystyrene tissue-culture dishes(Sigma–Aldrich) were coated with 0.1% gelatin. Cells were culturedin these dishes at a concentration of 1.5 � 106 cells per culture dishuntil they reached 80% confluence. Cells were grown in Way-mouth’s MB 752/1 cell media (Invitrogen) containing 0.002 MHEPES (Invitrogen), 50 lg/ml Gentamicin (Invitrogen), 15% horseserum (Atlanta Biologicals), and 0.4% Fungizone (Invitrogen) andkept at 37 �C with a mixture of air and 5% CO2. When cells reached80% confluence, they were trypsinized and reseeded in gelatin-

P. Balbuena et al. / Toxicology in Vitro 27 (2013) 1711–1718 1713
coated 24-well plates at a concentration of 1.0 � 105 cells per wellfor an additional 24 h. After 24 h, the media was removed and cellswere treated with different concentrations of the phthalate mono-esters in serum free media. DMSO was used as the vehicle control.The concentration of DMSO was 0.1% v/v in all samples. After 24 h,the media was replaced with serum free media containing DMSOor the appropriate phthalate plus 100 ng/mL LH. The cells wereincubated in LH for 6 h and the media was removed and assayedfor testosterone.
The rat Leydig tumor cell line (R2C) was obtained from theAmerican Type Culture Collection (ATCC). Cells were cultured in150 mm � 20 mm polystyrene tissue-culture treated dishes coatedwith 5 lg/cm2 collagen I and maintained with Ham’s F10 (+L-glu-tamine, HEPES) containing 2.5% fetal calf serum, 15% horse serum,0.4% fungizone and 1% penicillin/streptomycin, (10,000 U/ml). Cellswere allow to reach 80% confluence, and then were trypsinized andreseeded in 24-well plates coated with 5 lg/cm2 collagen I at aconcentration of 1.3 � 105 cells per well for an additional 24 h.After 24 h in the 24-well plates, the media was removed and theR2C cells were treated with different concentrations of the phthal-ate esters in serum free media. DMSO was used as the vehicle con-trol with concentrations of 0.1% v/v in all control samples. Samplesfrom these media were removed after 24 h and assayed fortestosterone.
2.3. Testosterone analysis
Testosterone in the medium was measured using an ELISA assay(Neogen Corporation, Lexington, KY) according to the manufac-turer’s directions. Medium was diluted in assay buffer providedin the kit prior to analysis.
2.4. Protein content and ATP viability analysis
After samples for testosterone were collected, cells from eachwell were treated with 200 lL of lysis buffer (0.1 M Tris–HCl(pH = 8.0), 0.05 M EDTA, 0.1 M NaCl, 1% sodium dodecyl sulfate,1% sarcosyl) supplemented with HALT Protease Inhibitor Cocktail(10 lL/1 mL buffer) (Thermo Fisher Scientific Inc., Rockford, IL). Ly-sis buffer components were purchased from Sigma–Aldrich. Theresulting lysate was homogenized 5 times with a 21-gauge needle(Fisher Scientific, Waltham MA), and the total protein content wasmeasured in an spectrophotometer utilizing the bicinchoninic acid(BCA) assay (Sigma–Aldrich) according to the manufacturer’sdirections. Cell viability was assessed using total protein contentand intracellular ATP content (ATPLite, PerkinElmer Life Sciences).None of the tested compounds showed >10% loss of total protein atconcentrations up to 300 lM (see Supplementary Data, Table S1).Most samples showed less than 10% reduction in ATP (see Supple-mentary Data, Table S2).
2.5. Relative potency calculations
Media concentrations associated with 50% inhibition of testos-terone (IC50) were calculated using Prism 6.0 (GraphPad Software,Inc., LaJolla, CA) using a one-site inhibition curve. IC50 values forpublished in vivo studies were calculated as follows. The adminis-tered (external) dose required to cause 50% reduction in testoster-one (ED50) was determined based on published dose response datafor DBP (Clewell et al., 2008; Lehmann et al., 2004) and DEHP(Borch et al., 2004; Parks et al., 2000) using the one-site inhibitioncurve in Prism. Fetal testes monoester concentrations associatedwith the ED50s, (IC50s for in vivo data) were then calculated usingpublished physiologically based pharmacokinetic (PBPK) modelsas described previously (Clewell et al., 2010, 2012). Developmentand validation of the PBPK models are described elsewhere
(Clewell et al., 2008; Gentry et al., 2011). In vivo tissue metabolitedata was available at similar doses to the ED50s for DBP and DEHP,which supported the accuracy of calculated fetal metabolite con-centrations from the PBPK models (Clewell et al., 2009; Kurataet al., 2012).
Relative potency factors (RPF) were calculated using MBP as thereference compound. The large amount of historical data for MBPin our laboratory for both cell lines, and in the fetal rat, supportsthe accuracy of the calculated IC50s for MBP (Clewell et al., 2009,2010; unpublished data). The reproducibility and variability inthe calculation of the IC50s were tested by calculating the IC50sfor 3 independent experiments with MBP. Each assay includedthree biological (independent) assays run on different weeks. TheIC50s for the individual runs were 2.98, 2.55 and 3.03 lM. The aver-age and SEM of the 3 runs were 2.85 ± 0.15, indicating that the as-say is consistent over time. RPFs were calculated by dividing theIC50 for MBP by the IC50 for the test chemical. Thus, compoundswith RPFs less than 1.0 would be less potent than MBP, and thosewith RPFs greater than 1.0 would be more potent than MBP.
2.6. Statistical analysis
All results are expressed as means ± SEM of three or more bio-logical (independent) replicates that were performed as separateexperiments. Each biological replicate also consisted of three tech-nical (in plate) replicates. Statistical analysis for dose–responsedata was performed using one-way analysis of variance (ANOVA),followed by Dunnett’s post-test to compare chemical treatmentsto the DMSO controls. ‘‘p’’ values less than 0.05 were consideredstatistically significant. For direct comparisons between two sam-ples (i.e., inhibition in MA-10 vs. R2C cell at a specific dose), a stu-dent’s t-test was used with a p-value cut-off of 0.05. All statisticalanalyses were performed using GraphPad Prism version 6.0(GraphPad software, San Diego, CA).
3. Results
3.1. Comparison of phthalate-induced inhibition of testosteronesynthesis in R2C and MA-10 cell lines
The positive controls (MEHP and MBP) showed a similar ability toinhibit testosterone in both the MA-10 and the R2C Leydig cell lines(Fig. 2; Table 1), with IC50s of 3 and 6 lM. The negative controls, MEPand MMP, showed very little response, with only a slight reduction intestosterone production at P100 lM. Dose–response data for thefollowing phthalates have previously been collected in the MA-10mouse Leydig cell line: MnOP, MEHP, MBeP, MBP, MEP, and MMP.The inhibition curves were highly consistent between the MA-10and R2C cell lines (Fig. 2). No statistical significance was observed be-tween the two cell lines at identical phthalate doses (p > 0.05, stu-dent’s t-test). This was true for all of the tested compounds.
Comparison of the relative potencies of the compounds in R2Ccells was also consistent with previous results in MA-10 Leydigcells (Table 1). The IC50 values for both cell lines show a strong cor-relation (R2 = 0.9915) (Fig. 3). The relative potency factors (RPF) forthe two cell lines are shown in Table 1. MEHP was similar in po-tency to MBP, while MnOP clearly showed a reduced ability to in-hibit testosterone, with RPFs of approximately 0.1 in both cell lines.MEP and MMP, which are expected to be inactive, showed very lit-tle response with RPFs less than 0.03 and 0.004, respectively.
3.2. Comparison of phthalate-induced inhibition of testosteronesynthesis in R2C cells to in vivo data
The IC50 values of the well-studied active phthalates (MBP,MEHP) and two phthalates considered to be inactive (MEP, MMP)
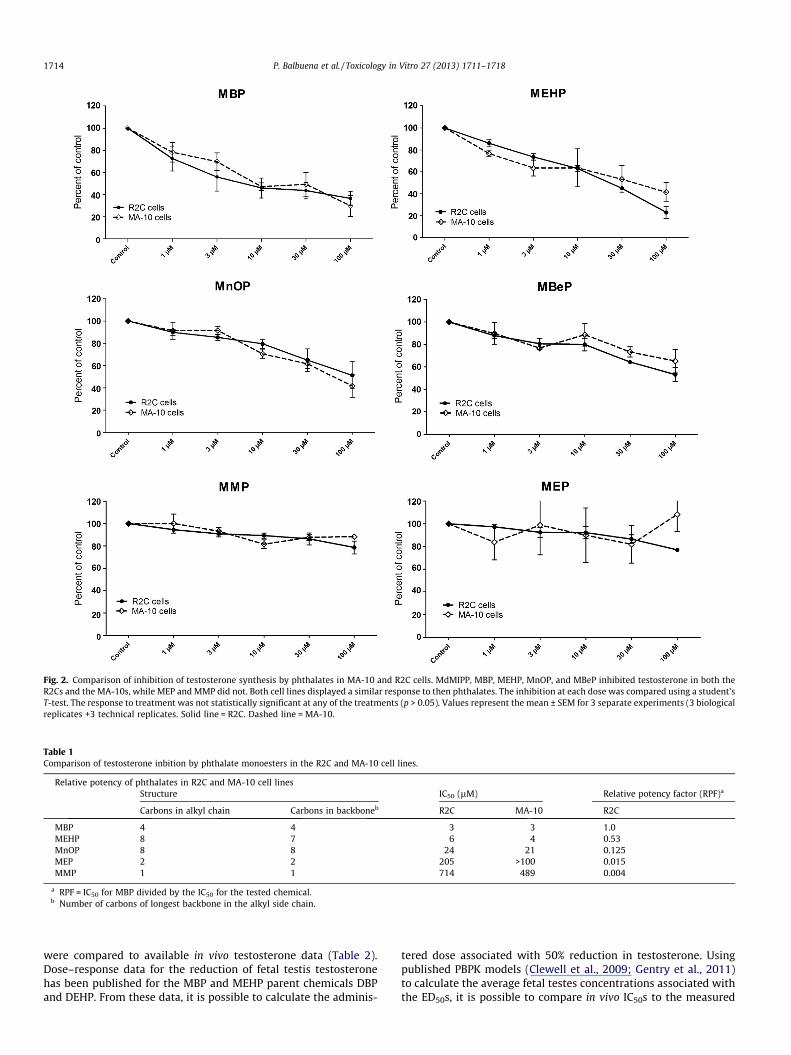
Fig. 2. Comparison of inhibition of testosterone synthesis by phthalates in MA-10 and R2C cells. MdMIPP, MBP, MEHP, MnOP, and MBeP inhibited testosterone in both theR2Cs and the MA-10s, while MEP and MMP did not. Both cell lines displayed a similar response to then phthalates. The inhibition at each dose was compared using a student’sT-test. The response to treatment was not statistically significant at any of the treatments (p > 0.05). Values represent the mean ± SEM for 3 separate experiments (3 biologicalreplicates +3 technical replicates. Solid line = R2C. Dashed line = MA-10.
Table 1Comparison of testosterone inbition by phthalate monoesters in the R2C and MA-10 cell lines.
Relative potency of phthalates in R2C and MA-10 cell linesStructure IC50 (lM) Relative potency factor (RPF)a
Carbons in alkyl chain Carbons in backboneb R2C MA-10 R2C
MBP 4 4 3 3 1.0MEHP 8 7 6 4 0.53MnOP 8 8 24 21 0.125MEP 2 2 205 >100 0.015MMP 1 1 714 489 0.004
a RPF = IC50 for MBP divided by the IC50 for the tested chemical.b Number of carbons of longest backbone in the alkyl side chain.
1714 P. Balbuena et al. / Toxicology in Vitro 27 (2013) 1711–1718
were compared to available in vivo testosterone data (Table 2).Dose–response data for the reduction of fetal testis testosteronehas been published for the MBP and MEHP parent chemicals DBPand DEHP. From these data, it is possible to calculate the adminis-
tered dose associated with 50% reduction in testosterone. Usingpublished PBPK models (Clewell et al., 2009; Gentry et al., 2011)to calculate the average fetal testes concentrations associated withthe ED50s, it is possible to compare in vivo IC50s to the measured

Fig. 3. Correlation between IC50 values determined for phthalate monoesters in theR2C and MA-10 cell lines. Correlation analysis between the cell lines showed an R2
of 0.9915.
Table 2Comparison of in vitro and in vivo IC50 values.
Testosterone inhibition in vitro vs. in vivoIC50 (lM)
In vivo (fetal rat testes)a In vitro (R2Cs)
MBP 3 3MEHP 7 6MEP >306b 205MMP �409b 713.9
a Calculated from PBPK models (Clewell et al., 2008; Gentry et al., 2011) and datain rat fetal testes (Clewell et al., 2009; Kurata et al., 2012), unless otherwisespecified.
b No testosterone reduction measured at highest tested doses (750 mg/kg/day)in vivo. Fetal testes monoester concentrations at 750 mg/kg/day DEP and DMPmaternal dose (Clewell et al., 2010).
P. Balbuena et al. / Toxicology in Vitro 27 (2013) 1711–1718 1715
IC50s in R2C cells. The in vitro IC50 values were remarkably similarto the in vivo IC50s for the two active phthalates. For MEHP, the cal-culated in vitro IC50 was 6 lM compared to 7 lM in vivo. In the caseof MBP, the calculated IC50s were identical (3 lM) in the R2C cellsand the fetal testes. The two inactive phthalates were also com-pared to in vivo data to the extent possible. The in vivo data formetabolite distribution and dose–response for fetal effects of DEPand DMP are relatively sparse, due to their lack of effect on fetaltestosterone. The best achievable estimates of fetal testes concen-trations are based on previous studies by Clewell et al., 2010 andLiu et al., 2005. In these studies, concentrations of MEP and MMPin fetal testes were measured at a single time point after maternaladministration of 500 mg/kg/day DEP and DMP. These doses werenot associated with altered steroidogenesis. Peak testes concentra-tions were 396 and 409 lM for MEP and MMP, respectively. Fromthis data, it can be inferred that the average concentrations wouldhave to be very high (>>>10 lM) in the fetal testes if either of thesephthalates were to inhibit testosterone at doses >500 mg/kg/day.The in vitro values measured in the current study are consistentwith this observation; IC50 values were well over 100 lM for bothMEP and MMP (205 and 714 lM respectively).
3.3. Predictive use of the R2C inhibition of testosterone synthesis assay
Qualitative structure activity analysis: The R2C assay was used totest a battery of phthalates with alky chain lengths ranging from 1to 10 carbons (C1 – C10) and various degrees of branching (Table 3).Dose response curves are provided in the Supplementary Data(Figs. S1A and S1B). While all of the compounds showed somedose-dependent decreases in testosterone at the tested concentra-tions (6300 lM), both chain length and branching had a clear ef-fect on phthalate potency. MBP, MPP and MdMIPP inducedstatistically significant inhibition starting at 1 lM. MEHP, MnHP,
MtBPP, MiDP and MdMPP significantly (p < 0.05) inhibited testos-terone beginning at 3 lM. For MtBMBP, and MMHP significantinhibition of testosterone started at 10 lM. For MBeP and MnOPstatistically significant inhibition started at 30 lM. MEP, MtMPP,and MMP inhibition was significant only at the highest concentra-tions (100 lM, 300 lM and 10 lM, respectively) (SupplementaryData, Figs. S1A and S1B). When the RPFs were compared, all ofthe phthalates showed reduced potency compared to MBP. As ex-pected, MEP and MMP were the least potent phthalates (Table 3).
In order to visualize the relationship between potency andeither (1) the total number of carbons in the alkyl chain or (2)the number of carbons in the alkyl chain backbones, both parame-ters were compared against the measured RPFs for all phthalates(see Supplementary Data, Fig. S2). There was no clear relationshipbetween the total number of carbons in the alkyl chain or the num-ber of carbons in the longest backbone of the alkyl chain and theability of the phthalate to inhibit testosterone. It is apparent, how-ever, that both the alkyl chain length and the degree of branchingare important factors in phthalate potency. When branched iso-mers were excluded from the analysis, a pattern was much moreapparent, with the C4 and C5 phthalates being most potent and asteep decline in potency when the alkyl chains had less than 4,or more than 5, carbons (Fig. 4).
Analysis of anti-androgenic activity of MEHP oxidative metabo-lites: Dose–response data are provided in the Supplementary Data(Fig. S3) and summarized in Table 4. 5-Hx-MEHP was the most po-tent metabolite of MEHP, inhibiting testosterone synthesis at rela-tively low doses (9 lM vs. 6 lM for MEHP). Phthalic acid was theleast potent metabolite, with significant inhibition only at thehighest concentrations (Table 4).
4. Discussion
Because of their common use in many different branches ofindustry, human exposure to phthalates is widespread. This, to-gether with rodent studies showing effects of some phthalates ontestosterone-dependent fetal development, underscores the needfor a better understanding of the dose–response for effects of dif-ferent phthalates on testosterone synthesis. Within the contextof the reduction of animal use in research, the identification ofnew assay systems and models for the assessment of phthalate ef-fects in vitro is an important task.
This paper demonstrates that the R2C cell line is a viable modelto assess phthalate effects on testosterone synthesis in the rat Ley-dig cell. Comparison of the R2Cs with the MA-10s (a cell line thatcan be induced to produce testosterone by LH) did not show anystatistically significant differences in their responses to phthalatesexposure, indicating that the in vitro testosterone synthesis modelsare robust. More importantly, the dose–response obtained in vitrofor known endocrine active phthalates (MEHP, MBP) and inactivephthalates (MMP, MEP) were remarkably similar to in vivo resultswhen compared based on tissue/media monoester concentrations.Monoester concentrations causing 50% inhibition of testosteroneproduction in the R2C cells were 3 and 6 lM for MBP and MEHPvs. 3 and 7 lM in vivo. Likewise, the inactive phthalates did not af-fect testosterone synthesis in vitro until monoester doses weremuch higher (>> 100 lM MMP or MEP). These results indicate thatthe R2C in vitro model is a good predictive assay for phthalate med-iated inhibition of testosterone synthesis in the fetal rat testes.
The assessment of a battery of different phthalates confirmedthat the R2C assay is a sensitive test for differences in anti-andro-genic potency among the phthalate family. Based on preliminaryanalyses, a simple relationship between the alkyl chain lengthand potency across all phthalate monoesters was not determineddue to the confounding effects of branching. A more sophisticated

Table 3Comparison of potency for testosterone inhibition across phthalates with varying alkyl chain structures. RPF = Relative potency factor in relation to MBP potency.
Alkyl Side Chain Structure Number of Carbons in Alkyl Chain
Number of Carbons in
Longest Backbone
RPF
MiDP 10 8 0.28
MtBPP 9 5 0.18
MtBMBP 9 4 0.13
MnOP 8 8 0.17
MEHP 8 6 0.53
MdMiPP 8 3 0.39
MMHP 7 6 0.05
MnHP 6 6 0.13
MtMPP 6 3 0.05
MPP 5 5 0.61
MdMPP 5 3 0.03
MBP 4 4 1.00
MEP 2 2 0.01
MMP 1 1 0.00
CH2CH3
CH3 CH3
CH3
CH3CH3
CH3C
CH3
CH3CH3
CH3 CH3C
CH2CH3
CH2CH3
CH3
CH3 CH3
CH3CH3
CH3
C
CH2CH3
CH3
CH2CH3
CH3
CH3CH3
CH3
C
CH2 CH3
CH3
CH3
CH3
C
CH2CH3
CH2 CH3
CH3
Fig. 4. Phthalate potency for inhibition of testosterone synthesis vs. total number ofcarbons in straight alkyl chains.
1716 P. Balbuena et al. / Toxicology in Vitro 27 (2013) 1711–1718
analysis of molecular descriptors is clearly needed to predict po-tency based on chemical structure, particularly for phthalates withhighly branched alkyl chains. Nonetheless, a comparison of thesein vitro results with published in vivo data, demonstrates a similar
trend in potency. For phthalates with straight and minimallybranched alky chains, the phthalates with 4–5 carbons have thehighest potency for inhibition of testosterone synthesis, and thephthalates with 0–2 carbon alkyl chains are the least potent(essentially inactive) (Gray et al., 2000; Hannas et al., 2011; How-deshell et al., 2008). It should be noted, however, that MPP showedsomewhat reduced potency in vitro than has been suggested basedon in vivo studies. Hannas et al. (2012) found that DPP was approx-imately 3-fold more potent than DBP, while the current study pre-dicted a slightly reduced potency of MPP compared to MBP. Theobvious difference in these two studies is that the in vivo studycompared the inhibition of testosterone synthesis based on exter-nal dose of the diester, while this study tested the monoesters ex-pected after hydrolysis. It is possible that the differences in thetwo studies would be resolved by accounting for in vivo metabo-lism and comparing relative potency by target tissue, as is thecase with MEHP (Table 1). Unfortunately, there is no availabledata for DPP disposition in the fetal rat. This highlights the needfor both potency data and metabolism data when trying to com-pare the potency of different chemicals regardless of whether the

Table 4Testosterone inhibition by oxidative metabolites of MEHP in R2C cells.
Metabolite IC50 RPF (Compared to MBP)
MEHP mono-2-ethylhexyl phthalate 6 0.535-Hx-MEHP mono-(2-ethyl-5-hydroxyhexyl) phthalate 9 0.325-Oxo-MEHP mono-(2-ethyl-5-oxohexyl) phthalate 37 0.0805-Cx-MEPP mono-(2-ethyl-5-carboxy-pentyl) phthalate 122 0.0242-Cx-MMHP mono-[(2-carboxymethyl)hexyl] phthalate 174 0.017PA phthalic acid 325 0.009
P. Balbuena et al. / Toxicology in Vitro 27 (2013) 1711–1718 1717
assays are performed in vitro or in vivo. Nonetheless, the currentassay provides a relatively fast and inexpensive platform fortesting specific metabolites and predicting relative potencyin vitro that, together with in vitro or in vivo metabolism data,could provide a much more efficient way of screening newcompounds.
Analysis of the oxidative metabolites of MEHP indicates thatPhase I metabolism does not completely inactivate the phthalatemonoester. In fact, all four of the tested metabolites showed someability to inhibit testosterone in the R2C cells, though three of themetabolites were clearly less potent than the parent monoester.Interestingly, the 5-hydroxy-2-ethylhexyl phthalate metabolite,showed a similar potency compared to its parent monoester MEHP(9 vs. 6 lM, respectively). The R2C assay provides a convenientplatform for testing these metabolites as the dose can be easilycontrolled. In contrast, in vivo tissue dose is highly affected bymaternal and fetal kinetic processes including competition formetabolizing enzymes, chemical specific maternal-fetal transportand preferential clearance of water-soluble compounds. Thus, thein vitro assay allows the ‘‘true’’ relative potency of the metabolitesto be determined for the pharmacodynamic endpoint of interest.However, in order to evaluate the importance of these findingsfor risk assessment efforts, the in vitro results must be consideredin light of the in vivo pharmacokinetics. For example, despite thefact that 5-Hx-MEHP has a similar potency to MEHP, the tissueconcentrations of MEHP in rats are expected to be 4-fold higherthan 5-Hx-MEHP (Kurata et al., 2012). Thus, 5-Hx-MEHP wouldhave negligible effect on overall antiandrogen activity in anin vivo animal study.
In addition to the ability to screen phthalates for anti-androge-nicity, this in vitro assay also provides a means of studying themechanism of action for the phthalates. To date, the moleculartarget for the phthalates in the Leydig cell has not been identified,though studies have suggested a role for several key transcrip-tions factors (e.g., SREBP, PPARa and COUP-TF II) (Corton andLapinskas, 2005; Johnson et al., 2011; Lehmann et al., 2004;Plummer et al., 2007; van den Driesche et al., 2012). Identifyingthe mechanism of action for phthalate-induced inhibition of tes-tosterone synthesis is key to understanding species differencesin susceptibility to the phthalates. Recent studies have shown thatdespite similar fetal exposures to MBP, mice do not experienceinhibition of fetal testosterone production or the long-term repro-ductive malformations observed in rats (Gaido et al., 2007; Grayet al., 1982). Furthermore, studies using human fetal testes xeno-grafts in immunocompromised mice indicate that the response inhumans may be more similar to that of mice than rats (Hegeret al., 2012). In order to confirm the correct model for predictinghuman effects, the mechanism of inhibition of testosterone mustfirst be identified in the rat, and then it must be determined ifthe human is likely to be affected in the same way. Humanin vitro models, such as the human H295 adrenal cell that was val-idated by the OECD as screening assay for inhibition of steroidsynthesis (Rijk et al., 2012), may also be useful for comparingthe rat and human steroid synthesis pathways and their suscepti-bility to phthalate anti-androgenicity.
5. Conclusions
The R2C assay provides a means of screening new phthalates forpotential endocrine activity and studying the mechanism of actionfor phthalate induced inhibition of testosterone synthesis whilereducing the use of animals and the cost of experimentation. Whenin vitro data were compared to data obtained from the fetal rat tes-tes in vivo, results were highly consistent for both endocrine active(MEHP, MBP) and inactive phthalates (MMP and MEP) based oneffective concentrations in media and tissue. Testing with 14 struc-turally diverse monoesters showed that monoesters with alkylchains consisting of 4–5 carbons had the highest potency for tes-tosterone inhibition, while 0–2 carbon alkyl chains were least po-tent, which is consistent with predictions from in vivo studies ofphthalates with straight or minimally branched alkyl chains. How-ever, the current study also showed that potency is modified bybranching of the alkyl chains, making a more generalized structureactivity analysis problematic and underscoring the need for a high-throughput in vitro option for screening of new phthalates andphthalate alternatives. Application of the assay to known oxidativemetabolites of MEHP further demonstrated that phase I metabo-lism does not completely inactivate MEHP, emphasizing the needfor metabolism data when interpreting in vitro pharmacodynamicdata. Taken together, this data indicates that the R2C steroid inhi-bition assay provides a predictive in vitro alternative to rat studiesfor phthalate-induced antiandrogenicity. Development of relativepotency factors (RPFs) for phthalates from the R2C assay, in con-junction with human biomonitoring data, could provide an ap-proach for estimating cumulative risks from environmentalphthalate exposures without requiring additional animal studies.
Acknowledgments
This work was supported by the American Chemistry CouncilLong Range Initiative (ACC–LRI) and the Environmental ProtectionAgency (EPA STAR grant 83386701–0).
Appendix A. Supplementary material
Supplementary data associated with this article can be found, inthe online version, at http://dx.doi.org/10.1016/j.tiv.2013.03.015.
References
Akingbemi, B.T., Ge, R., Klinefelter, G.R., Zirkin, B.R., Hardy, M.P., 2004. Phthalate-induced Leydig cell hyperplasia is associated with multiple endocrinedisturbances. Proc. Natl. Acad. Sci. U S A. 101 (3), 775–780.
Ascoli, M., 1981. Characterization of several clonal lines of cultured Leydig tumorcells: gonadotropin receptors and steroidogenic responses. Endocrinology 108(1), 88–95.
Boberg, J., Christiansen, S., Axelstad, M., Kledal, T.S., Vinggaard, A.M., Dalgaard, M.,Nellemann, C., Hass, U., 2011. Reproductive and behavioral effects of diisononylphthalate (DINP) in perinatally exposed rats. Reprod. Toxicol. 31 (2), 200–209.
Borch, J., Ladefoged, O., Hass, U., Vinggaard, A.M., 2004. Steroidogenesis in fetal malerats is reduced by DEHP and DINP, but endocrine effects of DEHP are notmodulated by DEHA in fetal, prepubertal and adult male rats. Reprod. Toxicol.18 (1), 53–61.

1718 P. Balbuena et al. / Toxicology in Vitro 27 (2013) 1711–1718
Chauvigné, F., Menuet, A., Lesné, L., Chagnon, M.C., Chevrier, C., Regnier, J.F.,Angerer, J., Jégou, B., 2009. Time- and dose-related effects of di-(2-ethylhexyl)phthalate and its main metabolites on the function of the rat fetal testis in vitro.Environ. Health Perspect. 117 (4), 515–521.
Christen, V., Crettaz, P., Oberli-Schrämmli, A., Fent, K., 2012. Antiandrogenic activityof phthalate mixtures: validity of concentration addition. Toxicol. Appl.Pharmacol. 259 (2), 169–176.
Clewell, R.A., Campbell, J.L., Ross, S.M., Gaido, K.W., Clewell 3rd, H.J., Andersen, M.E.,2010. Assessing the relevance of in vitro measures of phthalate inhibition ofsteroidogenesis for in vivo response. Toxicol. In vitro 24 (1), 327–334.
Clewell, R.A., Creasy, D.M., Willson, G., Edwards, K.E., Sochaski, M., Andersen, M.E.,2013a. Disposition of diiosononyl phthalate and its effects on sexualdevelopment of the male fetus following repeated dosing in pregnant rats.Reprod. Toxicol. 35, 56–69.
Clewell, R.A., Thomas, A., Willson, G., Creasy, D.M., Andersen, M.E., 2013b. A doseresponse study to assess effects after dietary administration of diisononylphthalate (DINP) in gestation and lactation on male rat sexual development.Reprod. Toxicol. 35, 70–80.
Clewell, R.A., Kremer, J.J., Williams, C.C., Campbell Jr., J.L., Andersen, M.E., Borghoff,S.J., 2008. Tissue exposures to free and glucuronidated monobutylyphthalate inthe pregnant and fetal rat following exposure to di-n-butylphthalate:Evaluation with a PBPK model. Toxicol. Sci. 103 (2), 241–259.
Clewell, R.A., Kremer, J.J., Williams, C.C., Campbell, J.L., Sochaski, M.A., Andersen,M.E., Borghoff, S.J., 2009. Kinetics of selected di-n-butyl phthalate metabolitesand fetal testosterone following repeated and single administration in pregnantrats. Toxicology 255 (1–2), 80–90.
Corton, J.C., Lapinskas, P.J., 2005. Peroxisome proliferator-activated receptors:mediators of phthalate ester-induced effects in the male reproductive tract?Toxicol. Sci. 83, 4–17.
Ema, M., Miyawaki, E., 2001. Adverse effects on development of the reproductivesystem in male offspring of rats given monobutyl phthalate, a metabolite ofdibutyl phthalate, during late pregnancy. Reprod Toxicol. 15 (2), 189–194.
Fisher, J.S., Macpherson, S., Marchetti, N., Sharpe, R.M., 2003. Human ’testiculardysgenesis syndrome’: a possible model using in-utero exposure of the rat todibutyl phthalate. Hum. Reprod. 18 (7), 1383–1394.
Foster, P.M., Mylchreest, E., Gaido, K.W., Sar, M., 2001. Effects of phthalate esters onthe developing reproductive tract of male rats. Hum. Reprod. Update. 7 (3),231–235.
Freeman, D.A., 1987. Constitutive steroidogenesis in the R2C Leydig tumor cell lineis maintained by the adenosine 39,59-cyclic monophosphateindependentproduction of a cycloheximide-sensitive factor that enhances mitochondrialpregnenolone biosynthesis. Endocrinology 120, 124–132.
Gaido, K.W., Hensley, J.B., Liu, D., Wallace, D.G., Borghoff, S., Johnson, K.J., Hall, S.J.,Boekelheide, K., 2007. Fetal mouse phthalate exposure shows that gonocytemultinucleation is not associated with decreased testicular testosterone.Toxicol. Sci. 97, 491–503.
Gazouli, M., Yao, Z.X., Boujrad, N., Corton, J.C., Culty, M., Papadopoulos, V., 2002.Effect of peroxisome proliferators on Leydig cell peripheral-typebenzodiazepine receptor gene expression, hormone-stimulated cholesteroltransport, and steroidogenesis: role of the peroxisome proliferator-activatorreceptor alpha. Endocrinology 143 (7), 2571–2583.
Gentry, P.R., Clewell 3rd, H.J., Clewell, R., Campbell, J., Van Landingham, C., Shipp,A.M., 2011. Challenges in the application of quantitative approaches in riskassessment: a case study with di-(2-ethylhexyl) phthalate. Crit. Rev. Toxicol. 41(2), 1–72.
Gray Jr., L.E., Ostby, J., Furr, J., Price, M., Veeramachaneni, D.N., Parks, L., 2000.Perinatal exposure to the phthalates DEHP, BBP, and DINP, but not DEP, DMP, orDOTP, alters sexual differentiation of the male rat. Toxicol. Sci. 58 (2), 350–365.
Gray Jr., L.E., Barlow, N.J., Howdeshell, K.L., Ostby, J.S., Furr, J.R., Gray, C.L., 2009.Transgenerational effects of Di (2-ethylhexyl) phthalate in the male CRL:CD(SD)rat: added value of assessing multiple offspring per litter. Toxicol. Sci. 110 (2),411–425.
Gray, T.J., Rowland, I.R., Foster, P.M., Gangolli, S.D., 1982. Species differences in thetesticular toxicity of phthalate esters. Toxicol. Lett. 11 (1–2), 141–147.
Hannas, B.R., Lambright, C.S., Furr, J., Howdeshell, K.L., Wilson, V.S., Gray Jr., L.E.,2011. Dose-response assessment of fetal testosterone production and geneexpression levels in rat testes following in utero exposure to diethylhexylphthalate, diisobutyl phthalate, diisoheptyl phthalate and diisononyl phthalate.Toxicol. Sci. 123 (1), 206–216.
Heger, N.E., Hall, S.J., Sandrof, M.A., McDonnell, E.V., Hensley, J.B., McDowell, E.N.,Martin, K.A., Gaido, K.W., Johnson, K.J., Boekelheide, K., 2012. Human fetal testisxenografts are resistant to phthalate-induced endocrine disruption. Environ.Health Perspect. 120 (8), 1137–1143.
Hernandez-Diaz, S., Mitchell, A.A., Kelley, K.E., Calafat, A.M., Hauser, R., 2009.Medications as a potential source of exposure to phthalates in the U.S.population. Environ. Health Perspect. 117, 185–189.
Howdeshell, K.L., Wilson, V.S., Furr, J., Lambright, C.R., Rider, C.V., Blystone, C.R.,Hotchkiss, A.K., Gray Jr., L.E., 2008. A mixture of five phthalate esters inhibitsfetal testicular testosterone production in the sprague-dawley rat in acumulative, dose-additive manner. Toxicol. Sci. 105 (1), 153–165.
Johnson, K.J., McDowell, E.N., Viereck, M.P., Xia, J.Q., 2011. Species-specific dibutylphthalate fetal testis endocrine disruption correlates with inhibition of SREBP2-dependent gene expression pathways. Toxicol. Sci. 120, 460–474.
Jones, H.B., Garside, D.A., Liu, R., Roberts, J.C., 1993. The influence of phthalate esterson Leydig cell structure and function in vitro and in vivo. Exp. Mol. Path. 58 (3),179–193.
Koch, H.M., Bolt, H.M., Preuss, R., Angerer, J., 2005. New metabolites of di(2-ethylhexyl)phthalate (DEHP) in human urine and serum after single oral dosesof deuterium-labelled DEHP. Arch. Toxicol. 79 (7), 367–376.
Kurata, Y., Makinodan, F., Shimamura, N., Katoh, M., 2012. Metabolism of di (2-ethylhexyl) phthalate (DEHP): comparative study in juvenile and fetalmarmosets and rats. J. Toxicol. Sci. 37 (1), 33–49.
Kwack, S.J., Kim, K.B., Kim, H.S., Lee, B.M., 2009. Comparative toxicologicalevaluation of phthalate diesters and metabolites in Sprague-Dawley male ratsfor risk assessment. J. Toxicol. Environ. Health A 72 (21–22), 1446–1454.
Lehmann, K.P., Phillips, S., Sar, M., Foster, P.M., Gaido, K.W., 2004. Dose-dependentalterations in gene expression and testosterone synthesis in the fetal testes ofmale rats exposed to di (n-butyl) phthalate. Toxicol. Sci. 81 (1), 60–68.
Liu, K., Lehmann, K.P., Sar, M., Young, S.S., Gaido, K.W., 2005. Gene expressionprofiling following in utero exposure to phthalate esters reveals new genetargets in the etiology of testicular dysgenesis. Biol. Reprod. 73 (1), 180–192.
Mylchreest, E., Cattley, R.C., Foster, P.M.D., 1998. Male reproductive tractmalformations in rats following gestational and lactational exposure to di(n-butyl) phthalate: an anti-androgenic mechanism? Toxicol. Sci. 43, 47–60.
Mylchreest, E., Sar, M., Cattley, R.C., Foster, P.M.D., 1999. Disruption of androgen-regulated male reproductive development by di(n-butyl) phthalate during lategestation in rats is different from flutamide. Toxicol. Appl. Pharmacol. 156, 81–95.
Oishi, S., Hiraga, K., 1980. Effects of phthalic acid monoesters on mouse testes.Toxicol. Lett. 6 (4–5), 239–242.
Parks, L.G., Ostby, J.S., Lambribght, C.R., Abbott, B.D., Klinefelter, G.R., Barlow, N.J.,Gray Jr., L.E., 2000. The plasticizer diethylhexyl phthalate inducesmalformations by decreasing fetal testosterone synthesis during sexualdifferentiation in the male rat. Toxicol. Sci. 58 (2), 339–349.
Plummer, S., Sharpe, R.M., Hallmark, N., Mahood, I.K., Elcombe, C., 2007. Time-dependent and compartment-specific effects of in utero exposure to di(n-butyl)phthalate on gene/protein expression in the fetal rat testis as revealed bytranscription profiling and laser capture microdissection. Toxicol. Sci. 97, 520–532.
Rao, R.M., Jo, Y., Leers-Sucheta, S., Bose, H.S., Miller, W.L., Azhar, S., Stocco, D.M.,2003. Differential regulation of steroid hormone biosynthesis in R2C and MA-10Leydig tumor cells: role of SR-B1-mediated selective cholesteryl ester transport.Biol. Reprod. 68 (1), 114–121.
Rijk, J.C., Peijnenburg, A.A., Blokland, M.H., Lommen, A., Hoogenboom, R.L., Bovee,T.F., 2012. Screening for modulatory effects on steroidogenesis using the humanH295R adrenocortical cell line: a metabolomics approach. Chem. Res. Toxicol.25 (8), 1720–1731.
Rowland, I.R., Cottrell, R.C., Phillips, J.C., 1977. Hydrolysis of phthalate esters bythe gastro-intestinal contents of the rat. Food Cosmet. Toxicol. 15 (1),17–21.
Silva, M.J., Barr, D.B., Reidy, J.A., Kato, K., Malek, N.A., Hodge, C.C., Hurtz 3rd, D.,Calafat, A.M., Needham, L.L., Brock, J.W., 2003. Glucuronidation patterns ofcommon urinary and serum monoester phthalate metabolites. Arch. Toxicol. 77(10), 561–567.
Stocco, D.M., Chen, W., 1991. Presence of identical mitochondrial proteins inunstimulated constitutive steroid-producing R2C rat Leydig tumor andstimulated nonconstitutive steroid producing MA-10 mouse Leydig tumorcells. Endocrinology 128, 1918–1926.
van den Driesche, S., Walker, M., McKinnell, C., Scott, H.M., Eddie, S.L., Mitchell, R.T.,Seckl, J.R., Drake, A.J., Smith, L.B., Anderson, R.A., Sharpe, R.M., 2012. Proposedrole for COUP-TFII in regulating fetal Leydig cell steroidogenesis, perturbation ofwhich leads to masculinization disorders in rodents. PLoS One 7 (5), e37064.
White, R.D., Carter, D.E., Earnest, D., Mueller, J., 1980. Absorption and metabolism ofthree phthalate diesters by rat small intestine. Food Cosmet. Toxicol. 18 (4),383–386.




















