
ÉTAT DE SANTÉ DES POUSSINS DE LA MÉSANGE MAGHRÉBINE Cyanistes teneriffae
15
Société d’Études Ornithologiques de France Muséum National d'Histoire Naturelle Volume 79 Numéro 1 Année 2011 Alauda SEOF Revue internationale d’Ornithologie www.mnhn.fr/assoc/seof/
-
Upload
univ-annaba -
Category
Documents
-
view
0 -
download
0
Transcript of ÉTAT DE SANTÉ DES POUSSINS DE LA MÉSANGE MAGHRÉBINE Cyanistes teneriffae

Société d’Études Ornithologiques de FranceMuséum National d'Histoire Naturelle
Volume 79 Numéro 1 Année 2011
AlaudaS E O FRevue internationale d’Ornithologie
www.mnhn.fr/assoc/seof/
3990. MONTADERT (M.) & LÉONARD (P.).– Biologie de la reproduction de la Gélinotte des bois Bonasabonasia dans les Alpes-de-Haute-Provence (France) (1ère partie) . . . . . . . . . . . . . . . . . . . . . . . . . . . .
3991. HANANE (S.), BERGIER (P.) & THÉVENOT(M.).– La reproduction de la Tourterelle mailléeStreptopelia senegalensis dans la plaine du Tadla (Maroc Central) : analyse comparée avec laTourterelle des bois Streptopelia turtur . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
3992. DENIS (P.).– Étude quantitative de l’avifaune nicheuse d’un grand massif forestier en plained’Alsace (Haguenau, France). . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
3993. STRENNA (L.) & BOILEAU (N.).– La capture du Faucon crécerelle Falco tinnunculus par la méthodedu bal-chatri en Bourgogne (1992-2005). . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
3994. PROVOST (P.), BARGAIN (B.) & CHEVEAU (P.).– Écologie du Phragmite aquatique Acrocephaluspaludicola sur deux sites de halte majeurs pendant le passage postnuptial dans l’Ouest de laFrance . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
3996 DJENIDI (R.), BOUSLAMA (Z.), HOUHAMDI (M.) & SOUALAH ALILA (H.).– État de santé des poussinsde la mésange maghrébine Cyanistes teneriffae . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
NOTES
3995 MATTHES (H.).– *Anomalie des rémiges d’un Martinet noir Apus apus. . . . . . . . . . . . . . . . . . . . . . . 3997 MOULAY-MELIANI (K.), MOALI (A.) & ISENMANN (P.).– *Première nidification de la Mouette rieuse
Chroicocephalus (Larus) ridibundus en Algérie . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3998 BREHME (S.), HERING (J.) & FUCHS (E.).– * Données sur la reproduction de la Tourterelle maillée
Streptopelia senegalensis dans le Fezzan et le Sud-Ouest de la Lybie . . . . . . . . . . . . . . . . . . . . . . . . 3999 BIBLIOGRAPHIE . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
CONTENTS3990. MONTADERT (M.) & LÉONARD (P.).– Breeding biology of Hazel Grouse Bonasa bonasia in the
Southeastern French Alps . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3991. HANANE (S.), BERGIER (P.) & THÉVENOT(M.).– Breeding biology of Laughing Dove Streptopelia
senegalensis in the Tadla lowland (Central Morocco): a comparative analysis with Turtle DoveStreptopelia turtur . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
3992. DENIS (P.).– Breeding avifauna census of a large lowland forest in Alsace (North-Eastern France) . . 3993. STRENNA (L.) & BOILEAU (N.).– The capture of the Common Kestrel Falco tinnunculus with bal-
chatri traps in Burgundy (1992-2005) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3994. PROVOST (P.), BARGAIN (B.) & CHEVEAU (P.).– Ecological observations on Aquatic Warbler
Acrocephalus paludicola at two major staging areas on postnuptial in western France . . . . . . . . . 3996 DJENIDI (R.), BOUSLAMA (Z.), HOUHAMDI (M.) & SOUALAH ALILA (H.).– Health status of nestlings
of the African Blue Tit Cyanistes teneriffae . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
NOTES
3995 MATTHES (H.).– *Abnormal wing primaries in a young Common Swift Apus apus. . . . . . . . . . . . . . 3997 MOULAY-MELIANI (K.), MOALI (A.) & ISENMANN (P.).– *First breeding of Black-headed Gull
Chroicocephalus (Larus) ridibundus in Algeria. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3998 BREHME (S.), HERING (J.) & FUCHS (E.).– *Palm Dove Streptopelia senegalensis breeding records
in Fezzan and Southwestern Libya . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3999 REVIEW . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
SOMMAIRE
1-16
17-28
29-42
43-52
53-63
65-78
64
79
80-8485-88
1-16
17-2829-42
43-52
53-63
65-78
64
79
80-8485-88
LXXIX. — 1. 2011
ALA
UD
A (
no
uv
ell
e s
érie
) LX
XIX
. – 1
. 2
01
1 Couv alauda (1) 2011 23/02/11 18:01 Page 1

Alauda 79 (1), 2011 : 65-783996
ÉTAT DE SANTÉ DES POUSSINS DE LA MÉSANGEMAGHRÉBINE Cyanistes teneriffae
Rédha DJENIDI(1), Zihad BOUSLAMA(2), Slim BENYACOUB(2), Moussa HOUHAMDI(2),Hana SOUALAH ALILA(2) et Adnène BELABED(2)
Health status of nestlings of the AfricanBlue Tit Cyanistes tenerif fae. Someaspects of the ecology of an African BlueTit population breeding in a lowlandCork Oak Quercus suber woodland innorth-eastern Algeria and the impact ofparasite load on the body conditions ofnestlings were studied during three years(1998 to 2000). The mean clutch sizewas 6.8 eggs and the mean number offledged young was 4.5 per nest, thusindicating a fairly good breeding successin that Mediterranean oak woodland.The inventory of ectoparasites in nestsand their impact on nestlings showedthat 80-100% of nests were infested bymites (Dermanyssus), t icks ( Ixodes),diptera (Protocal l iphora ) and f leas(Ceratophyllus). Moreover, this heavyparasite load seems to have not negativelyaffected the body parameters of the nestlings(tarsus length, mass at 15 days) and it had onlya little effect on mortality. These data show thatparents must have s trongly invested inparasitized broods to compensate the potentialcosts of parasite load by increasing feedingfrequency and care to nestlings (verified bysuccessful feeding frequency). Our resul ts
suggest that the host species shows behaviouraldefence strategies against pressures by parasite. Finally, Diptera larvae do not seem to haveharmed nestlings in the same way than it hasbeen shown for nestlings in Corsica. In thisAlgerian population the direct impact of parasitelarvae on nestlings could not be reported duringthe three study years.
Mots clés : Cyanistes teneriffae, Nord-Est algérien, Reproduction, Morphométrie, Ectoparasites.
Key words: Cyanistes teneriffae, North-eastern Algeria, Reproduction, Morphometry, Ectoparasites.
(1) Département de Biologie. Université d’Annaba. BP 12. El Hadjar. Algérie ([email protected]).(2) Laboratoire de recherche des Écosystèmes terrestres et aquatiques (EcoSTAq). Université d’Annaba. BP 12.El Hadjar. Algérie ([email protected]).
Dessin François Lovaty
ALAUDA(1)2011 14/04/11 14:11 Page 65

INTRODUCTION
Certaines études ont montré l’impact négatifdes ectoparasites sur les traits d’histoire de vie desoiseaux hôtes (MØLLER et al., 1990 ; LOYE & ZUK,1991 ; MØLLER, 1997). Notamment leur influenceest négative sur les conditions corporelles despoussins, en affectant la masse pondérale, la lon-gueur du tarse et le taux d’hématocrite ce quihypothèque la survie des jeunes parasités(RICHNER et al., 1993 ; MERINO & POTTI, 1995 ;MØLLER, 1997). Ces travaux ont concerné surtoutles ectoparasites du nid comme les Puces et lesMites, qui affectent souvent à la fois les poussinset leurs parents. C’est alors que les mauvaisesconditions corporelles des jeunes résultent nonseulement de l’impact direct des parasites maiségalement de l’investissement parental quidevient plus faible (MERINO & POTTI, 1995). Enabsorbant de l’énergie et d’autres ressources deleurs hôtes les parasites exercent leur impactnégatif (LOYE & ZUK, 1991 ; FORBES, 1993 ;CLAYTON & MOORE, 1997). Pourtant, des travauxanciens stipulaient que les parasites n’avaient pasun impact négatif important sur l’hôte en raisondes processus de co-évolution entre les deux(ALEXANDER, 1981). Cependant, des observationsrécentes suggèrent que la virulence du parasiteserait importante, notamment lorsqu’il s’agit d’ec-toparasites hématophages s’attaquant durant unecourte période aux oisillons nidicoles (LEHMANN,1993 ; MØLLER, 1997 ; MERINO & MØLLER, 1999)
Des travaux antérieurs ont aussi montré l’im-pact négatif des ectoparasites sur les traits d’his-toire de vie des oiseaux (MØLLER et al., 1990 ;RICHNER et al., 1993 ; MØLLER, 1997). Ces ecto-parasites des nids peuvent être des vecteurs demaladies (LOYE & ZUK, 1991 ; CLAYTON &MOORE, 1997) et influencer l’effort reproductif deleurs hôtes en entraînant un coût important dansleur reproduction (FORBES, 1993 ; ILMOMEN et al.,2000 ; RABERG et al., 2000). Une conséquencedirecte du parasitisme chez les oiseaux est l’aug-mentation des besoins alimentaires suite à unecarence due aux parasites. Les effets du parasi-tisme sont encore plus déterminants lorsque leurshôtes sont exposés à des conditions environne-mentales sévères telles qu’une disponibilité ali-
mentaire faible (DE LOPE et al., 1993 ; DUFVA &ALLANDER, 1996). Une nourriture correcte pour-rait apporter des éléments nécessaires à l’activa-tion du système immunitaire mais un déficit pour-rait réduire la résistance des organismes à l’actiondes parasites (LOCHMILLER et al., 1993 ; SAINO etal., 1997 ; BRINKHOF et al., 1999).
Les parents développent une série de réponsescomportementales pour réduire l’impact de lacharge parasitaire (LOYE & ZUK, 1991; CLAYTON &MOORE, 1997) et augmenter le fitness de leurs des-cendants (CLUTTON-BROCK, 1991 ; CLUTTON-BROCK & GODFRAY, 1991). Leur investissement setraduit par un apport accru de nourriture aux jeunesparasités. C’est ainsi que les parents consacrent unegrande partie de leur temps et de leur énergie à cetteactivité (DRENT & DAAN, 1980; BRYANT & TATNER,1988) pour compenser les effets dépresseurs duparasitisme et assurer la survie de leur descen-dance. La condition des poussins à l’envol dépen-drait donc de l’interaction entre l’investissementparental et la contrainte parasitaire. En plus dunourrissage, les parents peuvent avoir recours àd’autres moyens pour défendre leur nichée contreles ectoparasites, tels que l’incorporation dans leurnid de végétaux pouvant avoir soit une actioninsecticide, soit une action répulsive pouvant limi-ter l’abondance des ectoparasites (WIMBERGER,1984; CLARK & MASON, 1985 et 1988; COWIE &HINSLEY, 1988 ; CLARK, 1991 ; BANBURA &ZIELINSKI, 1995; LAMBRECHTS & DOS SANTOS,2000). Ils peuvent aussi augmenter les conditionssanitaires (déparasiter le nid et les poussins enconsommant les parasites ou en les expulsant).
Dans ce travail nous nous proposons d’étudierles conditions morphologiques et physiologiquesdes poussins de la Mésange maghrébine pour desnichées témoins et des nichées déparasitées.
ZONE D’ÉTUDE
L’étude a été réalisée au niveau du ParcNational d’El-Kala (Algérie). Cette région (FIG. 1)a fait l’objet de nombreux travaux, principalementceux de DE BELAIR (1990), BENYACOUB (1993),CHABI (1998) et BENYACOUB & CHABI (2000). Larégion est localisée à l’extrême Nord-Est algérien
Alauda 79 (1), 201166
ALAUDA(1)2011 14/04/11 14:11 Page 66

et a été érigée en 1991 par l’UNESCO en zoneprotégée. Elle est limitée par la mer Méditerranéeau Nord, les monts de Medjerda au Sud, la fron-tière algéro-tunisienne à l’Est et les plainesd’Annaba à l’Ouest. Les nichoirs ont été installésdans la réserve de Brabtia (30 m d’altitude), loca-lisée au Sud du parc, dans une subéraie avec sous-bois de plusieurs centaines d’hectares à 8 km àl’Ouest de la ville d’El-Kala (36°53N-8°30E).
MATÉRIEL ET MÉTHODES
Le travail a été réalisé durant trois annéessuccessives (1998 à 2000). Nous avons installéchaque année 132 à 154 nichoirs. Ces dernierssont des boîtes en bois de 2 cm d’épaisseur per-mettant un bon isolement thermique similaire àcelui des cavités naturelles, avec un volume inter-ne suffisant pour la construction du nid. Le dia-mètre du trou d’envol est égal à 26 mm, et permetl’accès uniquement à la Mésange maghrébine.Les nichoirs ont été installés avant le début de lasaison de reproduction (fin février) sur des arbresayant le même âge à environ 2 mètres du sol etdistants de 35 à 40 m. Les nichoirs ont été visitésune fois par semaine. Après l’éclosion, lesnichoirs ont été visités tous les deux jours et lesnichées ont été divisées en deux lots : un lottémoin et un lot traité.
Dans le lot traité, le nid a été remplacé tousles deux jours depuis l’éclosion et jusqu’à l’âge de15 jours par un autre nid stérile. La stérilisation
consiste à passer le nid dans un four à micro-ondesà 830 W pendant 3 minutes (HURTREZ-BOUSSÈS etal. 1997a, b). Les poussins des deux lots sontpesés tous les deux jours jusqu’à l’âge de 15 jours.Lorsque les poussins sont âgés de 15 jours, nousavons mesuré le tarse et prélevé 20 µl de sang parpoussin dans un tube capillaire hépariné qui estimmédiatement conservé dans une glacière. Lesang a été centrifugé à 12000 tours/min et nousavons estimé le taux d’hématocrite. Parallèlement,nous avons réalisé un frottis sanguin pour estimerles taux de globules rouges et blancs. Les lamesont subi une double coloration qui consiste, dansun premier temps, à un traitement à base de “May-Grunwald” qui permet de mettre en évidence lesérythrocytes et “Giemsa” qui permet la colorationdes noyaux des globules blancs.
La lame a été balayée dans un premier tempssous microscope photonique à faible grossis-sement (objectif x 10) pour juger de la qualité decette dernière. Par la suite le comptage des cel-lules sanguines a été effectué à l’objectif (x 100).Sur 100 cellules observées, nous avons calculé lepourcentage de globules rouges et celui des glo-bules blancs. Nous avons également calculé letaux de croissance des poussins depuis l’âge de 2 jours à 15 jours en mesurant le gain de massecorporelle par jour et par poussin.
La moyenne et l’écart-type pour chaque para-mètre ont été calculés et les résultats sont traitéspar des tests paramétriques. Les abondances desectoparasites ont été transformées en Log10 afind’atténuer les variations de l’écart-type.
État de santé des poussins de la Mésange maghrébine 67
FIG. 1.– Carte de situation du Parc National d’El Kala (BENYACOUB et al., 1998).Location of the Parc National d’El Kala in Algeria.
Cap de Garde
M E R M É D I T E R R A N É E
A L G É R I E
T U N I S I E
Annaba
0 20 km
El-KalaCap Rosa Cap Segleb
Cap de Fer �
ALAUDA(1)2011 14/04/11 14:11 Page 67
Nous avons également utilisé l’analyse encomposantes principales (ACP) pour traiter lestableaux de contingence (observation : nichoir -charge parasitaire et observation : nichoir - va-riable). Cette analyse pouvant se faire sur diffé-rents types de données, permet d’étudier la dépen-dance entre les paramètres morpho-physiolo-giques des poussins et la charge parasitaire.
RÉSULTATS
Traitement des nidsL’analyse statistique montre que la stérilisa-
tion des nids a un effet hautement significatif surl’intensité des différents groupes de parasites. Elleest significativement plus faible dans les nids trai-tés (TAB. I).
Paramètres morphologiquesLa masse des poussins à l’âge de 15 jours
(TAB. II) ne présente pas de différence significativeentre les deux lots (F1,51 = 3,39, p > 0,05). En revan-che, la longueur du tarse est significativement plusélevée dans le lot témoin (F 1,49 = 35,08, p < 0,01).
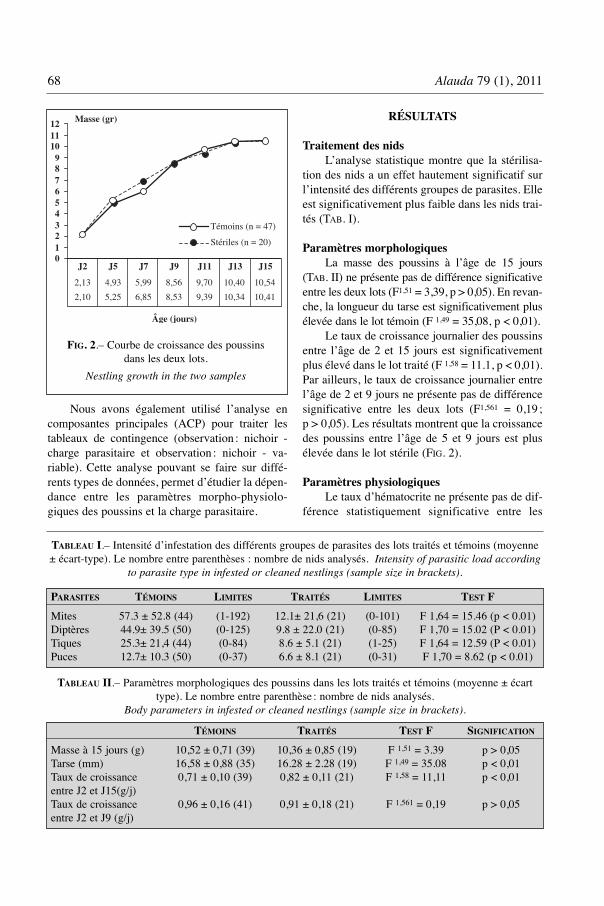
Le taux de croissance journalier des poussinsentre l’âge de 2 et 15 jours est significativementplus élevé dans le lot traité (F 1,58 = 11.1, p < 0,01).Par ailleurs, le taux de croissance journalier entrel’âge de 2 et 9 jours ne présente pas de différencesignificative entre les deux lots (F1,561 = 0,19 ;p > 0,05). Les résultats montrent que la croissancedes poussins entre l’âge de 5 et 9 jours est plusélevée dans le lot stérile (FIG. 2).
Paramètres physiologiquesLe taux d’hématocrite ne présente pas de dif-
férence statistiquement significative entre les
Alauda 79 (1), 201168
Âge (jours)
Masse (gr)
Témoins (n = 47)
2,13 4,93 5,99 8,56 9,70 10,40 10,54
Stériles (n = 20)
2,10 5,25 6,85 8,53 9,39 10,34 10,41
J2 J5 J7 J9 J11 J13 J150123456789
121110
FIG. 2.– Courbe de croissance des poussinsdans les deux lots.
Nestling growth in the two samples
PARASITES TÉMOINS LIMITES TRAITÉS LIMITES TEST F
Mites 57.3 ± 52.8 (44) (1-192) 12.1± 21,6 (21) (0-101) F 1,64 = 15.46 (p < 0.01)Diptères 44.9± 39.5 (50) (0-125) 9.8 ± 22.0 (21) (0-85) F 1,70 = 15.02 (P < 0.01)Tiques 25.3± 21,4 (44) (0-84) 8.6 ± 5.1 (21) (1-25) F 1,64 = 12.59 (P < 0.01)Puces 12.7± 10.3 (50) (0-37) 6.6 ± 8.1 (21) (0-31) F 1,70 = 8.62 (p < 0.01)
TABLEAU I.– Intensité d’infestation des différents groupes de parasites des lots traités et témoins (moyenne± écart-type). Le nombre entre parenthèses : nombre de nids analysés. Intensity of parasitic load according
to parasite type in infested or cleaned nestlings (sample size in brackets).
TÉMOINS TRAITÉS TEST F SIGNIFICATION
Masse à 15 jours (g) 10,52 ± 0,71 (39) 10,36 ± 0,85 (19) F 1,51 = 3.39 p > 0,05Tarse (mm) 16,58 ± 0,88 (35) 16.28 ± 2.28 (19) F 1,49 = 35.08 p < 0,01Taux de croissance 0,71 ± 0,10 (39) 0,82 ± 0,11 (21) F 1,58 = 11,11 p < 0,01entre J2 et J15(g/j)Taux de croissance 0,96 ± 0,16 (41) 0,91 ± 0,18 (21) F 1,561 = 0,19 p > 0,05entre J2 et J9 (g/j)
TABLEAU II.– Paramètres morphologiques des poussins dans les lots traités et témoins (moyenne ± écarttype). Le nombre entre parenthèse : nombre de nids analysés.
Body parameters in infested or cleaned nestlings (sample size in brackets).
ALAUDA(1)2011 14/04/11 14:11 Page 68
deux lots (F1,26 = 24,6 ; p > 0,001). En revanche, lepourcentage des cellules sanguines est significati-vement différent entre les deux (TAB. III). Lepourcentage de globules blancs est significati-vement plus élevé dans le lot témoin (F1,62
= 185,06 ; p < 0,001). Le taux de globules rougesest significativement plus élevé dans le lot traité(F1.62 = 163,37, p < 0,001).
Analyse en composantes principales (ACP)Nous avons soumis la matrice nichoirs-varia-
bles morpho-physiologiques et de reproduction àune ACP réduite, à laquelle nous avons couplé lamatrice nichoirs-parasites comme variables expli-catives. L’analyse a été effectuée sous le logicielPC-Ord “ V 4.10” (Mc CUNE & MEFFORD, 1999).
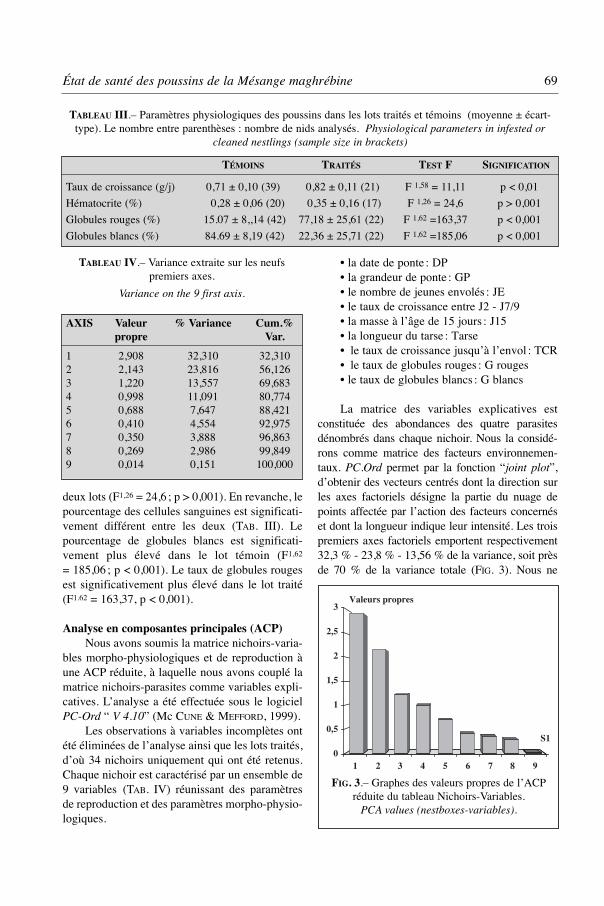
Les observations à variables incomplètes ontété éliminées de l’analyse ainsi que les lots traités,d’où 34 nichoirs uniquement qui ont été retenus.Chaque nichoir est caractérisé par un ensemble de9 variables (TAB. IV) réunissant des paramètresde reproduction et des paramètres morpho-physio-logiques.
• la date de ponte : DP• la grandeur de ponte : GP• le nombre de jeunes envolés : JE• le taux de croissance entre J2 - J7/9• la masse à l’âge de 15 jours : J15• la longueur du tarse : Tarse• le taux de croissance jusqu’à l’envol : TCR• le taux de globules rouges : G rouges• le taux de globules blancs : G blancs
La matrice des variables explicatives estconstituée des abondances des quatre parasitesdénombrés dans chaque nichoir. Nous la considé-rons comme matrice des facteurs environnemen-taux. PC.Ord permet par la fonction “joint plot”,d’obtenir des vecteurs centrés dont la direction surles axes factoriels désigne la partie du nuage depoints affectée par l’action des facteurs concernéset dont la longueur indique leur intensité. Les troispremiers axes factoriels emportent respectivement32,3 % - 23,8 % - 13,56 % de la variance, soit prèsde 70 % de la variance totale (FIG. 3). Nous ne
État de santé des poussins de la Mésange maghrébine 69
1 2 3 4 5 6 7 8 9
S1
Valeurs propres
0
0,5
1
1,5
2
2,5
3
FIG. 3.– Graphes des valeurs propres de l’ACPréduite du tableau Nichoirs-Variables.
PCA values (nestboxes-variables).
TÉMOINS TRAITÉS TEST F SIGNIFICATION
Taux de croissance (g/j) 0,71 ± 0,10 (39) 0,82 ± 0,11 (21) F 1,58 = 11,11 p < 0,01
Hématocrite (%) 0,28 ± 0,06 (20) 0,35 ± 0,16 (17) F 1,26 = 24,6 p > 0,001
Globules rouges (%) 15.07 ± 8,,14 (42) 77,18 ± 25,61 (22) F 1,62 =163,37 p < 0,001
Globules blancs (%) 84.69 ± 8,19 (42) 22,36 ± 25,71 (22) F 1,62 =185,06 p < 0,001
TABLEAU III.– Paramètres physiologiques des poussins dans les lots traités et témoins (moyenne ± écart-type). Le nombre entre parenthèses : nombre de nids analysés. Physiological parameters in infested or
cleaned nestlings (sample size in brackets)
AXIS Valeur % Variance Cum.% propre Var.
1 2,908 32,310 32,310 2 2,143 23,816 56,126 3 1,220 13,557 69,683 4 0,998 11,091 80,774 5 0,688 7,647 88,421 6 0,410 4,554 92,975 7 0,350 3,888 96,863 8 0,269 2,986 99,849 9 0,014 0,151 100,000
TABLEAU IV.– Variance extraite sur les neufspremiers axes.
Variance on the 9 first axis.
ALAUDA(1)2011 14/04/11 14:11 Page 69
retiendrons donc que les trois premiers axes pourl’interprétation de l’analyse. Nous avons choisi deprésenter pour chaque variable les plans factorielsles plus significatifs afin d’alléger le texte. Lasuperficie des symboles-points est proportionnelleaux valeurs prises par les variables.
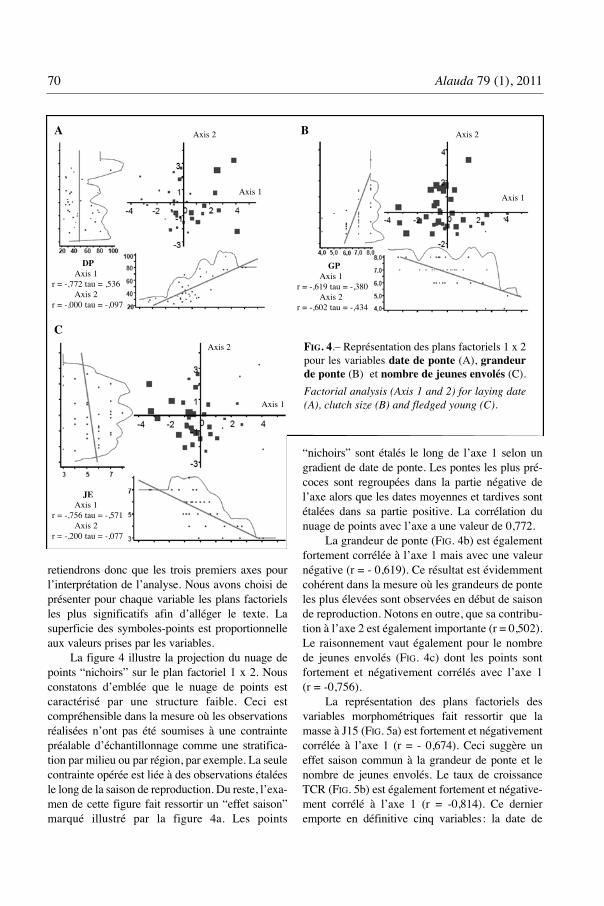
La figure 4 illustre la projection du nuage depoints “nichoirs” sur le plan factoriel 1 x 2. Nousconstatons d’emblée que le nuage de points estcaractérisé par une structure faible. Ceci estcompréhensible dans la mesure où les observationsréalisées n’ont pas été soumises à une contraintepréalable d’échantillonnage comme une stratifica-tion par milieu ou par région, par exemple. La seulecontrainte opérée est liée à des observations étaléesle long de la saison de reproduction. Du reste, l’exa-men de cette figure fait ressortir un “effet saison”marqué illustré par la figure 4a. Les points
“nichoirs” sont étalés le long de l’axe 1 selon ungradient de date de ponte. Les pontes les plus pré-coces sont regroupées dans la partie négative del’axe alors que les dates moyennes et tardives sontétalées dans sa partie positive. La corrélation dunuage de points avec l’axe a une valeur de 0,772.
La grandeur de ponte (FIG. 4b) est égalementfortement corrélée à l’axe 1 mais avec une valeurnégative (r = - 0,619). Ce résultat est évidemmentcohérent dans la mesure où les grandeurs de ponteles plus élevées sont observées en début de saisonde reproduction. Notons en outre, que sa contribu-tion à l’axe 2 est également importante (r = 0,502).Le raisonnement vaut également pour le nombrede jeunes envolés (FIG. 4c) dont les points sontfortement et négativement corrélés avec l’axe 1(r = -0,756).
La représentation des plans factoriels desvariables morphométriques fait ressortir que lamasse à J15 (FIG. 5a) est fortement et négativementcorrélée à l’axe 1 (r = - 0,674). Ceci suggère uneffet saison commun à la grandeur de ponte et lenombre de jeunes envolés. Le taux de croissanceTCR (FIG. 5b) est également fortement et négative-ment corrélé à l’axe 1 (r = -0,814). Ce dernieremporte en définitive cinq variables : la date de
Alauda 79 (1), 201170
A
C
B
DPAxis 1
r = -,772 tau = ,536Axis 2
r = -,000 tau = -,097
JEAxis 1
r = -,756 tau = -,571Axis 2
r = -,200 tau = -,077
GPAxis 1
r = -,619 tau = -,380Axis 2
r = -,602 tau = -,434
Axis 2
Axis 1
Axis 2
Axis 1
Axis 2
Axis 1
FIG. 4.– Représentation des plans factoriels 1 x 2pour les variables date de ponte (A), grandeurde ponte (B) et nombre de jeunes envolés (C).
Factorial analysis (Axis 1 and 2) for laying date(A), clutch size (B) and fledged young (C).
ALAUDA(1)2011 14/04/11 14:11 Page 70
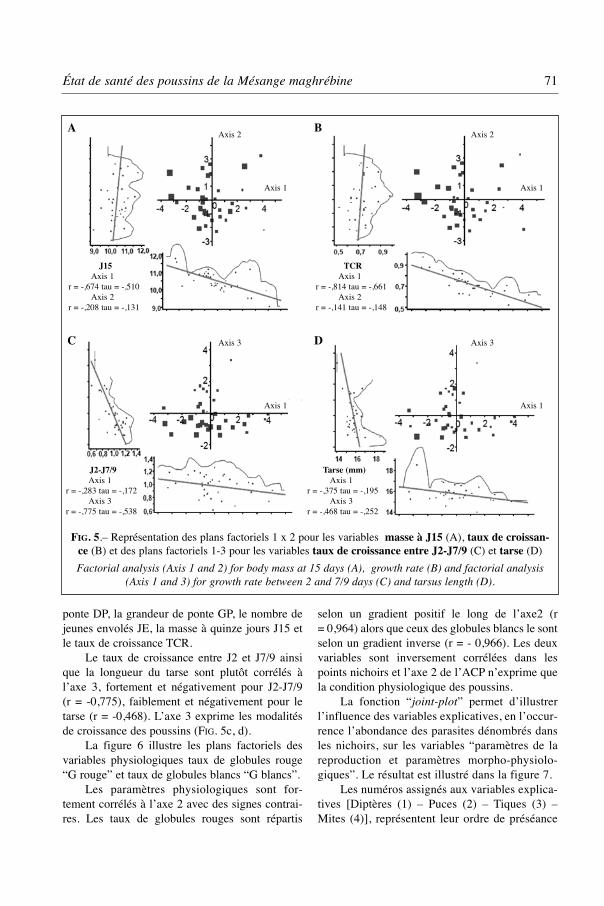
ponte DP, la grandeur de ponte GP, le nombre dejeunes envolés JE, la masse à quinze jours J15 etle taux de croissance TCR.
Le taux de croissance entre J2 et J7/9 ainsique la longueur du tarse sont plutôt corrélés àl’axe 3, fortement et négativement pour J2-J7/9(r = -0,775), faiblement et négativement pour letarse (r = -0,468). L’axe 3 exprime les modalitésde croissance des poussins (FIG. 5c, d).
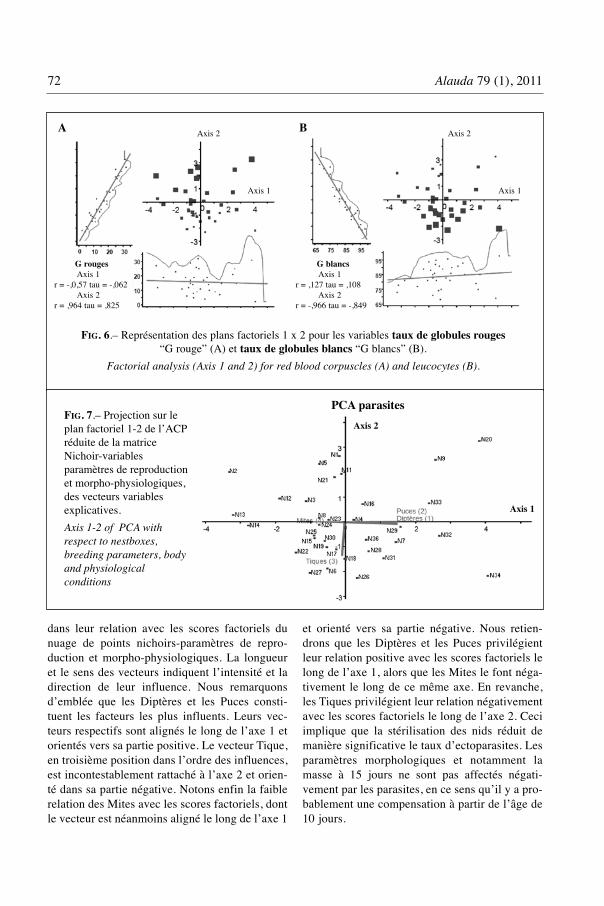
La figure 6 illustre les plans factoriels desvariables physiologiques taux de globules rouge“G rouge” et taux de globules blancs “G blancs”.
Les paramètres physiologiques sont for-tement corrélés à l’axe 2 avec des signes contrai-res. Les taux de globules rouges sont répartis
selon un gradient positif le long de l’axe2 (r= 0,964) alors que ceux des globules blancs le sontselon un gradient inverse (r = - 0,966). Les deuxvariables sont inversement corrélées dans lespoints nichoirs et l’axe 2 de l’ACP n’exprime quela condition physiologique des poussins.
La fonction “joint-plot” permet d’illustrerl’influence des variables explicatives, en l’occur-rence l’abondance des parasites dénombrés dansles nichoirs, sur les variables “paramètres de lareproduction et paramètres morpho-physiolo-giques”. Le résultat est illustré dans la figure 7.
Les numéros assignés aux variables explica-tives [Diptères (1) – Puces (2) – Tiques (3) –Mites (4)], représentent leur ordre de préséance
État de santé des poussins de la Mésange maghrébine 71
FIG. 5.– Représentation des plans factoriels 1 x 2 pour les variables masse à J15 (A), taux de croissan-ce (B) et des plans factoriels 1-3 pour les variables taux de croissance entre J2-J7/9 (C) et tarse (D)
Factorial analysis (Axis 1 and 2) for body mass at 15 days (A), growth rate (B) and factorial analysis(Axis 1 and 3) for growth rate between 2 and 7/9 days (C) and tarsus length (D).
A B
C D
J15Axis 1
r = -,674 tau = -,510Axis 2
r = -,208 tau = -,131
TCRAxis 1
r = -,814 tau = -,661Axis 2
r = -,141 tau = -,148
J2-J7/9Axis 1
r = -,283 tau = -,172Axis 3
r = -,775 tau = -,538
Tarse (mm)Axis 1
r = -,375 tau = -,195Axis 3
r = -,468 tau = -,252
Axis 2Axis 2
Axis 3Axis 3
Axis 1 Axis 1
Axis 1 Axis 1
ALAUDA(1)2011 14/04/11 14:11 Page 71
dans leur relation avec les scores factoriels dunuage de points nichoirs-paramètres de repro-duction et morpho-physiologiques. La longueuret le sens des vecteurs indiquent l’intensité et ladirection de leur influence. Nous remarquonsd’emblée que les Diptères et les Puces consti-tuent les facteurs les plus influents. Leurs vec-teurs respectifs sont alignés le long de l’axe 1 etorientés vers sa partie positive. Le vecteur Tique,en troisième position dans l’ordre des influences,est incontestablement rattaché à l’axe 2 et orien-té dans sa partie négative. Notons enfin la faiblerelation des Mites avec les scores factoriels, dontle vecteur est néanmoins aligné le long de l’axe 1
et orienté vers sa partie négative. Nous retien-drons que les Diptères et les Puces privilégientleur relation positive avec les scores factoriels lelong de l’axe 1, alors que les Mites le font néga-tivement le long de ce même axe. En revanche,les Tiques privilégient leur relation négativementavec les scores factoriels le long de l’axe 2. Ceciimplique que la stérilisation des nids réduit demanière significative le taux d’ectoparasites. Lesparamètres morphologiques et notamment lamasse à 15 jours ne sont pas affectés négati-vement par les parasites, en ce sens qu’il y a pro-bablement une compensation à partir de l’âge de10 jours.
Alauda 79 (1), 201172
FIG. 6.– Représentation des plans factoriels 1 x 2 pour les variables taux de globules rouges“G rouge” (A) et taux de globules blancs “G blancs” (B).
Factorial analysis (Axis 1 and 2) for red blood corpuscles (A) and leucocytes (B).
Axis 2
Axis 1
Axis 2
Axis 1
Axis 2
Axis 1
G rougesAxis 1
r = -,0,57 tau = -,062Axis 2
r = ,964 tau = ,825
G blancsAxis 1
r = ,127 tau = ,108Axis 2
r = -,966 tau = -,849
A B
PCA parasitesFIG. 7.– Projection sur leplan factoriel 1-2 de l’ACPréduite de la matriceNichoir-variablesparamètres de reproductionet morpho-physiologiques,des vecteurs variablesexplicatives.
Axis 1-2 of PCA withrespect to nestboxes,breeding parameters, bodyand physiologicalconditions
ALAUDA(1)2011 14/04/11 14:11 Page 72

Les paramètres physiologiques sont, enrevanche, affectés. Le taux de croissance journa-lier ainsi que celui des globules rouges est plusélevé chez les poussins des nichées stérilisées, cequi montre que ceux-ci sont en meilleures condi-tions que ceux des nichées-témoins. Paral-lèlement, le taux de globules blancs est plus élevéchez les poussins des nichées-témoins, ce quitémoigne d’une réaction immunitaire importantechez ces derniers ou alors d’une infection. Cesrésultats montrent que l’effet des ectoparasitessemble influencer les conditions des poussins enparticulier entre l’âge de 5 et 10 jours.
DISCUSSION ET CONCLUSION
Les résultats ne permettent pas de conclure àun impact de la charge parasitaire sur les conditionscorporelles des poussins durant la période de nour-rissage. Nous n’observons pas à ce titre de diffé-rence significative entre les lots témoins et traités.La charge parasitaire, bien que plus élevée que celleobservée en Corse, n’a donc pas affecté la massedes oisillons âgés de 15 jours. Cette absence d’effetdes ectoparasites peut être attribuée à une compen-sation des pertes énergétiques et ce, par l’augmen-tation de l’investissement parental.
En effet, il est bien admis que les ectopara-sites et les micro-organismes du nid jouent unrôle-clé dans l’évolution des soins parentaux desoiseaux nidicoles (HART, 1997). Ces soins semanifestent par des comportements de protectiondes poussins vis-à-vis des ectoparasites ou par unapport de plantes aromatiques qui ont un effetrépulsif et / ou pesticide (CLARK & MASON,1995 ; LAMBRECHTS & DOS SANTOS, 2000 ;LAFUMA et al., 2001). La défense peut aussi pren-dre la forme d’une stimulation du système immu-nitaire des poussins (GWINNER et al., 2000), del’abstention à nicher dans des nichoirs infestés(BROWN & BROWN, 1986 ; MERINO & POTTI,1995), du choix d’un partenaire non parasité(MØLLER et al., 1994), d’une autovaccination despoussins (HEEB et al., 1998), de la “sanitation”des nids (HURTREZ –BOUSSÈS et al., 2000), et/oude l’augmentation des fréquences de nourrissage.
Peu d’études expérimentales concernantl’impact de la charge parasitaire sur l’investis-
sement des parents dans le nourrissage des jeunesont été réalisées (CHRISTE et al., 1996). Certainsauteurs ont observé une augmentation de la fré-quence des apports alimentaires (RICHNER et al.,1995 ; CHRISTE et al., 1996 ; HURTREZ-BOUSSÈS etal., 1998) alors que d’autres ont noté une diminu-tion (MØLLER et al., 1994). Enfin, ROGERS et al.(1991) et SAINO et al. (1998) n’ont observé aucunchangement en réponse à une infestation de para-sites (TRIPET et al., 2002). Cette différence decomportement des parents est attribuée à uncompromis entre la reproduction en cours et lareproduction future (MØLLER, 1997 ; CHRISTE etal., 1996 ; SZÈP & MØLLER, 1999).
Nos résultats sont en accord avec les travauxde RICHNER et al. (1995), CHRISTE et al. (1996) etHURTREZ-BOUSSÈS et al. (1998). En effet, nouspouvons émettre l’hypothèse que les parents s’in-vestissent de manière importante pour l’entretienet l’état sanitaire des nichées, afin de compenserles effets délétères des ectoparasites. Cet investis-sement aura permis ici d’éviter aux poussins undéficit de croissance.
En revanche, l’analyse des résultats sanguinsdémontre de manière incontestable un impactphysiologique sévère. En effet, les différenceshautement significatives entre le taux de globulesrouges, de globules blancs et de l’hématocrite desdeux lots, militent fortement d’une part, en faveurd’une anémie importante des poussins avant l’en-vol et d’autre part d’une réponse non moinsimportante à une infection de ces mêmes indivi-dus. Du reste, la différence significative entre lesnichées parasitées et traitées pour l’hématocriteconfirme ce constat.
D’après CLARK (1991) l’anémie consécutiveà des exsanguinations par des ectoparasites héma-tophages réduit les possibilités d’apport d’oxy-gène aux cellules et limite des efforts intenses. Ilest clair dans ce cas que les oisillons anémiésseront très handicapés pour des activités derecherche de la nourriture et auront une faibleaptitude à échapper à l’attaque des prédateurs.
L’analyse en composantes principales de lamatrice (nichoir- variables morpho-physiolo-giques et paramètres de la reproduction) couplée àla matrice (nichoirs-parasites) comme variablesexplicatives, a révélé un important effet de saison-nalité sur le premier axe factoriel. Cet axe
État de santé des poussins de la Mésange maghrébine 73
ALAUDA(1)2011 14/04/11 14:11 Page 73

regroupe les différents paramètres de reproductionainsi que ceux qui représentent les conditionsmorphologiques. Il semble ici que nous soyonsconfrontés à un regroupement factoriel des para-mètres sans signification particulière autre queleur auto-corrélation d’une part et leur corrélationà la saisonnalité d’autre part.
Le couplage à la matrice des variables expli-catives (nichoirs - parasites) permet d’identifierpar ordre de préséance les Diptères et les Puces etdans une moindre mesure les Mites comme fac-teurs influençant les paramètres de reproduction etmorphométriques. Nous pensons que ce résultatne constitue qu’ “un bruit de fond” général, for-tement lié à la saisonnalité des phénomènesincluant la phénologie d’apparition des Diptères,Puces et Mites.
L’axe 2 du plan factoriel de l’ACP isole etrévèle, en revanche, deux gradients de signesopposés : l’un concernant les érythrocytes etl’autre les leucocytes. Cet axe concernerait doncles conditions physiologiques des poussins avantl’envol. Le couplage de la matrice des variablesexplicatives révèle que ce sont uniquement lestaux d’infestation par les Tiques qui expliquentces gradients de conditions physiologiques. Cesrésultats ne confortent pas ceux obtenus dans lespopulations européennes et corses chez qui la pré-sence de ces ectoparasites est anecdotique et leurimpact sur les paramètres morpho-physiologiquesest donc négligeable. Nous serions donc face àune expérience franchement différente de l’expé-rience européenne dans la mesure où l’impactparasitaire est la conséquence de la présence deTiques. Rappelons à ce titre que 10 années d’ob-servation n’ont jamais révélé une action de parasi-tisme de la part des Diptères sous la forme de lar-ves accrochées à la peau des oisillons, alors quecelle des Tiques est plus évidente.
L’augmentation de la température au cours dela saison favorise l’abondance des ectoparasitesreprésentés par les Arthropodes. Ces derniers syn-chronisent leur cycle de vie en fonction de celui deleurs hôtes. Les résultats obtenus montrent que lesnids de la Mésange maghrébine sont beaucoupplus infestés que ceux des autres populations quinichent en Europe continentale et méditerra-néenne (HARPER et al., 1992 ; RICHNER & TRIPET,1999 ; EAVA et al., 1994 ; MERINO & POTTI, 1995,
HURTREZ-BOUSSES et al., 1999 ; ALLANDER, 1998 ;BAUCHAU, 1998 ; HEEB et al., 2000). Ceci montreque chaque région présente une diversité en ecto-parasites qui lui est spécifique. En effet, si enCorse, les Diptères sont dominants (HURTREZ-BOUSSES et al., 1999), en Algérie c’est un “cock-tail” d’ectoparasites qui infeste les nids de laMésange maghrébine. Leur taux d’infestation estinfluencé par les paramètres comme la date deponte et la hauteur du nid pour les Mites et lesDiptères, la grandeur de ponte pour les différentsparasites. À leur tour, ces parasites ont un impactsur les conditions physiologiques des poussins,puisque le taux de globules rouges est plus élevéchez les poussins des nichées déparasitées, alorsque celui des globules blancs est plus élevé dansles nichées parasitées (BOUSLAMA et al., 2001 et2002). Il est montré que les poussins issus desnichées parasitées présentent une anémie relati-vement importante et peuvent donc, d’une part,être fortement pénalisés après l’envol (périodependant laquelle leurs besoins énergétiques aug-mentent en raison de la mue), tandis que la dispo-nibilité alimentaire décroît et d’autre part, êtreincapables de se disperser.
En revanche, à l’âge de 15 jours, la massecorporelle des poussins et le taux de croissance nesont en général pas significativement influencéspar les ectoparasites, ce qui suggère fortement queles parents des nichées parasitées se sont investispour compenser les effets dépresseurs des ectopa-rasites sur la croissance de leurs jeunes.
En effet, certains travaux ont révélé que lesectoparasites des nids ainsi que d’autres micro-organismes pourraient jouer un rôle dans l’évolu-tion de l’investissement parental (HART, 1997).Les adultes développent une série de stratégiescomportementales pour protéger leur descendancecontre les parasites afin de leur assurer une survieultérieure (HART, 1992).
La condition des poussins à l’envol dépendde l’interaction entre l’investissement parental etla contrainte parasitaire. Selon la qualité des soinsparentaux, les effets du parasitisme peuvent êtreaggravés ou atténués. En effet, comme l’activitéde nourrissage est conditionnée par la demandedes poussins (BENGTSSON & RYDEN, 1983 ;GODFRAY, 1991) il faut s’attendre ainsi à deuxtypes de réponses (CHRISTE et al., 1996) :
Alauda 79 (1), 201174
ALAUDA(1)2011 14/04/11 14:11 Page 74

- un nourrissage important des jeunes afin decompenser les méfaits du parasitisme quelle quesoit la qualité des nichées ;
- le refus de s’investir dans l’élevage desnichées parasitées qui seraient de mauvaise quali-té, ce qui impliquerait une amplification de l’im-pact des parasites.
Par ailleurs, la charge parasitaire demanderaitun investissement dans la défense immunitaire(RABERG et al., 2000). Plusieurs études ont eneffet montré une relation positive entre l’effortreproducteur et la susceptibilité aux endoparasitestels que Plasmodium qui affaiblissent par exemplela Mésange charbonnière et le Lézard vivipareLacerta vivipara (SORCI et al., 1996). En plus del’endoparasitisme, les adultes subissent égalementles attaques des ectoparasites lors de leurs visitesau nid et notamment lorsque la femelle abrite sespoussins. RICHNER & HEEB (1995) ont proposé unmodèle dans lequel l’évolution de la grandeur deponte est en relation avec la capacité qu’a ou nonle parasite à ajuster sa démographie à celle de sonhôte. Ils proposent deux types de parasites :
- ceux dont le nombre est indépendant de lagrandeur de la nichée (le cas des parasites à cyclelong, comme les Puces), chez qui on observe unediminution de la charge parasitaire avec une aug-mentation du nombre de poussins. L’évolutiondevrait conduire à une augmentation de la gran-deur de ponte ;
- ceux dont la démographie s’ajuste aunombre de poussins (parasites à cycle court,comme les Acariens).
Les résultats obtenus par HURTREZ-BOUSSÈS
(1996) ont vérifié ce modèle et comme RICHNER &HEEB (1995) la démographie du parasite est ajus-tée à la grandeur de la nichée de l’hôte. Toutefoisnos observations révèlent que nous sommes enprésence d’un véritable cocktail de parasites(cycle court et cycle long), des études portantessentiellement sur les contraintes parasitairesimposées aux adultes (en prenant en considérationleur sexe) et notamment sur les paramètres démo-graphiques tels que la date de ponte, la grandeurde ponte et leurs évolutions dans le temps s’avére-raient nécessaires.
En Corse, les larves de Diptères sont abon-dantes et présentent un impact direct sur les pous-sins. En réponse, les parents utilisent une stratégie
de défense comportementale avec apport deplantes aromatiques, “pot-pourri”, à effet pesticidedurant la période de présence des poussins dans lenid, pour diminuer leur impact (LAMBRECHTS &DOS SANTOS, 2000). Pour les populations algé-riennes, nous n’avons noté ni la présence de cesplantes aromatiques dans les nids, ni aucun effetapparent de ces larves durant ces trois années d’é-tude, ou durant les dix dernières années de re-cherche en biologie des populations de laMésange maghrébine. Ceci laisse supposer que lesDiptères qui seraient hématophages verraient leurvirulence diminuer en raison de la présence simul-tanée d’autres ectoparasites ou alors seraient, eux-mêmes, parasités. La première hypothèse peut êtrevérifiée expérimentalement en élevant, par exem-ple, ces ectoparasites en captivité et en les intro-duisant par la suite dans des nichées complète-ment stérilisées pour vérifier leurs effets sur lespoussins. En revanche, nos résultats suggèrent for-tement que les tiques auraient un effet direct surl’état général des poussins notamment en affectantleur qualité physiologique, composante essentiellede leur fitness, car elle affecte leur survie aprèsleur envol. Les oisillons s’envolent considérable-ment anémiés et affectés d’une hyperleucocytose.L’anémie est expliquée par le fait que l’espèceresponsable du genre Ixodes est strictement héma-tophage à tous ses stades de développement etprovoquerait, vu son poids et son éthologie, uneexsanguination importante. Celle-ci entraîneraitdonc une diminution du nombre de globules rou-ges et de l’hématocrite. Quant à l’augmentation dutaux de leucocytes, deux hypothèses peuvent êtreémises :
- développement d’une activation accrue deleur système immunitaire ;
- infection des oisillons car les tiques sontconnues pour être responsables de plusieursmaladies bactériennes et parasitaires.
De ce point de vue, il reste à mesurer, chez lespopulations locales de la mésange, l’impact desendoparasites. En effet, le site d’étude est localiséau sein d’un complexe humide fréquenté par denombreux vecteurs de ces derniers, notamment lesDiptères du genre Aedes, Culex ou Anopheles, lar-gement répandus dans la région (HASSAINE, 2002).Les conditions environnementales ne doivent pasêtre sans influence sur l’état sanitaire général des
État de santé des poussins de la Mésange maghrébine 75
ALAUDA(1)2011 14/04/11 14:11 Page 75

organismes et plus particulièrement des Vertébréshoméothermes et de leur aptitude à survivre. Danscette perspective, il nous semble que l’inventairedes endoparasites et de leurs vecteurs ainsi quel’étude des facteurs qui gouvernent leur actionsoient particulièrement appropriés.
BIBLIOGRAPHIE
• ALEXANDER (M.) 1981.– Why microbial predatorsand parasites do not eliminate their prey and host.Ann. Rev. Microbiol., 35: 113-133.
• ALLANDER (K.) 1998.– The effects of an ectoparasi-te on reproductive success in the Great Tit : a 3-yearexperimental study. Can. J. Zool., 76: 19-25.
• BANBURA (J.) & ZIELINSKI (P.) 1995.– The influenceof laying sequence and ambient temperature on eggsize variation in the Swallow Hirundo rustica. J.Ornithol., 136: 453-460.
• BAUCHAU (V.) 1998.– Comparison of parasitismlevel in two sympatric passerines : the PiedFlycatcher and the Great Tit. Ecoscience, 5 : 164-171.
• BENGTSSON (H.) & RYDEN (O.) 1983.– Parental fee-ding rate in relation to begging behaviour in asyn-chronously hatched broods of Great Tit (Parusmajor). Behav. Ecol. Sociobiol., 12: 243-251.
• BENYACOUB (S.) 1993.– Ecologie de l’avifauneforestière nicheuse de la région d’El Kala (Nord-Est algérien). Thèse Doct. Univ. Bourgogne. Dijon.
• BENYACOUB (S.), LOUANCHI (M.), BABA AHMED (R.),BENHOUHOU (S.), BOULAHBEL (R.), CHALABI (B.),HAOU (F.), ROUAG (R.) & ZIANE (N.) 1998.– Plandirecteur de gestion du parc national d’El-Kala etdu complexe de zones humides (wilaya d’El Tarf).
• BENYACOUB (S.) & CHABI (Y.) 2000.– Diagnose éco-logique de l’avifaune du Parc National d’El-Kala.Numéro spécial juin. Publication Univ. Annaba.
• BOUSLAMA (Z.), LAMBRECHTS (M.M.) & CHABI (Y.)2001.– Nest ectoparasites and consequences fornestling in algerian Blue Tits : impacts of ticks,blowflies and fleas. Ecoscience, 8 (3) 320-324.
• BOUSLAMA (Z.), LAMBRECHTS (M.M.), ZIANE (N.),DJENIDI (R.) & CHABI (Y). 2002.– Parental foodprovisioning of chicks and nest ectoparasites in anorth-african population of Blue Tits. Ibis, 144 (online), E73-E78.
• BRINKHOF (M.W.G.), HEEB (P.), KÖLLIKER (M.) &RICHNER (H.) 1999.– Immunocompetence of nest-ling Great Tits in relation to rearing environmentand parentage. Proc. Roy. Soc. Lond. B., 266:2315-2322
• BROWN (C.R.) & BROWN (M.B.) 1986.– Ecto-parasitism as a cost of coloniality in Cliff Swallows(Hirundo pyrrhonota). Ecology, 67: 1206-1218.
• BRYANT (D.M.) & TATNER (P.) 1988.– Energetics ofthe annual cycle of Dippers Cinclus cinclus. Ibis,130 : 17-38.
• CHABI (Y.) 1998.– Étude de l’écologie de la repro-duction des populations de mésanges (Parus) dansles chênaies du Nord-Est de l’Algérie. Thèse Doct.Univ. Annaba.
• CHRISTE (P.), RICHNER (H.) & OPPLIGER (H.) 1996.–Begging, food provisioning and nestling competi-tion in Great Tit broods infested with parasites.Behav. Ecol., 7 : 127-131.
• CLARK (L.) 1991.– The nest protection hypothesis :the adaptive use of plant secondary compounds byEuropean Starlings. In LOYE (J.E.) & ZUK (M.)Bird-parasite interactions: ecology, evolution andbehaviour, Oxford Univ. Press.
• CLARK (L.) & MASON (J.R.) 1985.– Use of nestmaterial as insecticidal and antipathogenic agentsby European Starling. Oecologia, 67: 169-170.
• CLAYTON (D.H.) & MOORE (J.) 1997.– Host-parasiteEvolution. Oxford University Press.
• COWIE (R.J.) & HINSLEY (S.A.) 1988.– Feeding eco-logy of Great Tits (Parus major) and Blue Tits(Parus caeruleus) breeding in suburban gardens. J.Anim. Ecol., 57: 611-626.
• DE BELAIR (G.) 1990.– Structure, fonctionnement etperspective de gestion de 4 éco-complexes lacustreset marécageux (El-Kala, Est algérien). Thèse Doct.Univ. Montpellier II.
• DE LOPE (F.), GONZALES (G.), PEREZ (J.J.) & MØLLER
(A.P.) 1993.– Increased detrimental effects of ecto-parasites on their bird hosts during adverse envi-ronmental conditions. Oecologia, 95: 234-240.
• DRENT (R.H.) & DAAN (S.) 1980.– The prudentparent : energetic adjustments in avian breeding.Ardea, 68: 225-252.
• DUFVA (R.) & ALLANDER (K.) 1996.– Variableeffects of the Hen Flea Ceratophyllus gallinae onthe breeding success of the Great Tit Parus majorin relation to weather conditions. Ibis, 138 (4) : 772-777.
• EEVA (T.), LEHIKOINEN (E.) & NURMI (J.) 1994.–Effects of ectoparasites on breeding success ofGreat Tits (Parus major) and Pied Flycatchers(Ficedula hypoleuca) in an air pollution gradient.Can. J. Zool., 72: 624-635.
• FORBES (M.R.L.) 1993.– Parasitism and host repro-ductive effort. Oikos, 67: 444-450.
• GODFRAY (H.C.J.) 1991.– Signaling of need by
Alauda 79 (1), 201176
ALAUDA(1)2011 14/04/11 14:11 Page 76

offspring to their parents. Nature, 352 (6333): 328-330.
• GWINNER (H.), OLTROGGE (M.), TROST (L.) &NIENABER (U.) 2000.– Green plants in Starlingnests : effects on nestlings. Anim. Behav., 59: 301-309.
• HARPER (G.H.), MARCHANT (A.) & BODDINGTON
(D.G.) 1992.– The ecology of the Hen FleaCerotophyllus gallinae and the Moorhen FleaDasypsyllus gallinulae in nestboxes. J. Anim. Ecol.,61: 317-327
• HART (B.) 1992.– Behavioural adaptations to parasi-tes : an ethological approach. J. Parasitol., 78: 256-265.
• HART (B.) 1997.– Behavioural defence. In CLAYTON
(D.H.) & MOORE (J.) Host – parasite evolution.General principles and avian models, OxfordUniversity Press.
• HASSAINE (K.) 2002.– Biogéographie et biotypolo-gie des Culicidés (Diptera-Neurotocera) del’Afrique méditerranéenne. Bioécologie des espè-ces les plus vulnérantes de la région occidentalealgérienne. Thèse Doct. Univ. Tlemcen.
• HEEB (P.), WERNER (I.) & KOLLIKER (M.) 1998.–Benefits of induced host responses against an ecto-parasite. P. Roy. Soc. Lond. Bio., 265 (1390): 51-56.
• HEEB (P.), KÖLLIGER (M.) & RICHNER (H.) 2000.–Bird-ectoparasite interactions, nest humidity, andectoparasite community structure. Ecology, 81:958-968.
• HURTREZ-BOUSSÈS (S.) 1996.– Interactions hôteparasite : le système Mésange bleue - Protocal-liphora en région méditerranéenne. Thèse Doct.Univ. Montpellier II.
• HURTREZ-BOUSSÈS (S.), BLONDEL (J.), PERRET (P.) &RENAUD (F.) 1997a.– Relationship between inten-sity of blowfly infestation and reproductive successin a Corsican population of Blue Tits. J. Avian Biol.,28: 267-270.
• HURTREZ- BOUSSÈS (S.), PERRET (P.), RENAUD (F.) &BLONDEL (J.) 1997 b.– High blowfly parasiticloads affect breeding success in a Mediterraneanpopulation of Blue Tits. Oecologia, 112: 514-517.
• HURTREZ-BOUSSÈS (S.), BLONDEL (J.), PERRET (P.),FABREGUETTES (J.) & RENAUD (F.) 1998.– Chickparasitism by blowflies affects feeding rates in aMediterranean population of Blue Tits. Ecol. Lett.,1 : 17-20.
• HURTREZ-BOUSSES (S.), RENAUD (F.), BLONDEL (J.),PERRET (P.) & GALAN (M.J.) 2000.– Effects ofectoparasites of young on parents behaviour in aMediterranean population of Blue Tits. J. Avian
Biol., 31: 266-269.• HURTREZ-BOUSSÈS (S.), DE GARINE-WICHATITSKY
(M.), PERRET (P.), BLONDEL (J.) & RENAUD (F.)1999.– Variations in prevalence and intensity ofblow fly infestations in an insular Mediterraneanpopulation of Blue Tits. Can. J. Zool., 77: 337-341.
• ILMONEN (P.), TAARNA (T.) & HASSELQUIST (D.)2000.– Experimentally activated immune defencein female Pied Flycatchers results in reduced bree-ding success. Proc. Roy. Soc. Lond., B. 31: 266-269.
• LAFUMA (L.), LAMBRECHTS (M.M.) & RAYMOND (M.)2001.– Aromatic plants in bird nests as a protec-tion against blood-sucking flying insects. Behav.Proc., 56: 113-121.
• LAMBRECHTS (M.M.) & DOS SANTOS (A.) 2000.–Aromatic herbs in corsican Blue Tit nests : the‘Potpourri’ hypothesis. Acta Oecol., 21: 175-178.
• LEHMANN (T.) 1993.– Ectoparasites : direct impacton host fitness. Parasitol. Today, 9 : 8-13.
• LOCHMILLER (R.L.), VESTREY (M.R.) & BOREN (J.C.)1993.– Relationship between protein nutritionalstatus and immunocompetence in Northern Bob-white chicks. Auk, 110: 503-510.
• LOYE (J.E.) & ZUK (M.) 1991.– Bird-parasite inter-action. Ecology, evolution and behaviour. OxfordUniversity Press.
• MARRE (A.) 1987.– Etude géomorphologique du Telloriental algérien de Collo à la frontière tunisienne.Univ. Aix -Marseille II. U.E.R de Géographie.
• MERINO (S.) & POTTI (J.) 1995.– Mites and blowfliesdecrease growth and survival in nestling PiedFlycatchers. Oikos, 73: 95-103.
• MERINO (S.) & MOLLER (A.P.) 1999.– The coevolu-tion of virulence and immune defence in birds.Pages 2158-2162. In ADAMS (N.J.) & SLOTOW
(R.H.) BirdLife, Proc. 22 Int. Ornithol. Congr.Durban. Johannesburg: BirdLife South Africa.
• MC CUNE (B.) & MEFFORD (M.J.) 1999.–Multivariate analysis of ecological data. MgMsoftware, Gleneden Beach. Oregon. USA.
• MØLLER (A.P.), DE LOPE (F.), MORENO (J.), GONZALES
(G.) & PÉREZ (J.J.) 1994.– Ectoparasites and hostenergetics : House Martin bugs and House Martinnestlings. Oecologia, 98: 263-268.
• MØLLER (A.P.) 1997.– Parasitism and the evolutionof host life-history. In CLAYTON (D.H.) & MOORE
(J.) Host-parasite evolution: general principles andavian models, Oxford University Press.
• RABERG (L.), NILSSON (J.A.), ILMONEN (P.),STJERNMAN (M.) & HASSELQUIST (D.) 2000.– Thecost of an immune response: vaccination reducesparental effort. Ecol. Lett., 3 : 382-386.
État de santé des poussins de la Mésange maghrébine 77
ALAUDA(1)2011 14/04/11 14:11 Page 77

ICONOGRAPHIE DES OISEAUX DE FRANCE52 gravures originales de Paul BARRUEL en couleurs, sous jaquette, format 17,5 x 21,5 cm
Texte de P. BARRUEL, J. DORST, P. ENGELBACH, R. D. ETCHÉCOPAR, F. HUE,C. JOUANIN, G. OLIVIER, J. RAPINE sous la direction de J. BERLIOZ
Mémoires de la Société Ornithologique de France et de l’Union Française (1955)
100,00 € + 8 € de portÀ commander à SEOF, case postale 51, Muséum National d’Histoire Naturelle,
55 rue Buffon, F-75231 Paris cedex 05
• RICHNER (H.), OPPLINGER (A.) & CHRISTE (P.) 1993.–Effect of an ectoparasite on reproduction in GreatTits. J. Anim. Ecol., 62: 703-710.
• RICHNER (H.) & HEEB (P.) 1995.– Are clutch andbrood size patterns in birds shaped by ectoparasites.Oikos, 73 (3) : 435-441
• RICHNER (H.), OPPLINGER (A.) & CHRISTE (P.) 1995.–Parental investment affects prevalence of malaria.Proc. Natl. Acad. Sci. USA, 92: 1192-1194.
• RICHNER (H.) & TRIPET (F.) 1999.– Ectoparasitismand the trade-off between current and future repro-duction. Oikos, 86 (3) : 535-538.
• ROGERS (C.A.), ROBERTSON (R.J.) & STUTCHBURY
(B.J.) 1991.– Patterns and effects of parasitismby Protocalliphora sialia on Tree Swallow nest-lings. In LOYE (J.E.) & ZUK (M.) Bird-parasiteinteractions : ecology, evolution and behaviour,Oxford Ornithology Series.
• SAINO (N.), CALZA (S.) & MOLLER (A.P.) 1998.–
Effects of a dipteran ectoparasite on immuneresponse and growth trade-offs in Barn Swallow,Hirundo rustica, nestlings. Oikos, 81 : 217-228.
• SAINO (N.), CALZA (S.) & MOLLER (A.P.) 1997.–Immunocompetence of nestling Barn Swallow inrelation to brood size and parental effort. J. Anim.Ecol., 66: 825-836.
• SORCI (G.), CLOBERT (J.) & MICHALAKIS (Y.) 1996.–Cost of reproduction and cost of parasitism in theCommon Lizard, Lacerta vivipara. Oikos, 76 (1) :121-130
• SZÉP (T.) & MOLLER (A.P.) 1999.– Costs of parasi-tism and host immune defence in the Sand MartinRiparia riparia : a role for parent-offspringconflict ? Oecologia, 119 : 9-15.
• TRIPET (F.), GLASER (M.) & RICHNER (H.) 2002.–Behavioural responses to ectoparasites : time-bud-get adjustment and what matters to Blue Tits Paruscaeruleus infested by fleas. Ibis, 144: 135-143.
Alauda 79 (1), 201178
GÉRARD J. MOREL (1926 - 2011).– Pionnier dans l’étude de l’écologie des oiseaux sédentaires (afro-tropicaux) et visiteurs (paléarctiques) et des conditions de leur cohabitation dans les savanessahéliennes de l’Afrique de l’Ouest, Gérard J. MOREL a travaillé d’abord au sein de la Missiond’aménagement du Sénégal puis de l’Office de Recherche Scientifique et Technique d’Outre-Mer encollaboration avec sa femme Marie-Yvonne du Centre National de la Recherche Scientifique.Ensemble, ils ont animé pendant de très longues années la Station de recherche de Richard-Toll sur larive gauche du fleuve Sénégal. Leurs nombreuses publications ont porté sur l’écologie du Travailleur àbec rouge et du Moineau doré (deux fléaux des cultures en Afrique de l’Ouest), l’inventaire del’avifaune et les périodes de reproduction dans le Sahel sénégalais, le statut et la phénologie desmigrateurs et des hivernants paléarctiques, l’écologie hivernale de la Tourterelle des bois, les relationsentre l’Amarante du Sénégal et son parasite le Combassou du Sénégal… Ils ont aussi cosigné un trèsimportant ouvrage sur les oiseaux du Sénégal en 1990. Investi dans la Société d’Ornithologie de l’OuestAfricain qu’il a présidé et sa revue “Malimbus”, G. J. MOREL avait pris sa retraite en Normandie où ilavait apporté au Groupe Ornithologique Normand sa compétence et son soutien. Un grand monsieurinternationalement reconnu de l’ornithologie africaine s’en est allé…
Paul ISENMANN & Pierre NICOLAU-GUILLAUMET.
ALAUDA(1)2011 14/04/11 14:11 Page 78




















