
Epithelial-to-mesenchymal transition confers resistance to apoptosis in three murine mammary...
11
ORIGINAL ARTICLE Ewan J. D. Robson . Walid T. Khaled . Kathrine Abell . Christine J. Watson Epithelial-to-mesenchymal transition confers resistance to apoptosis in three murine mammary epithelial cell lines Received December 6, 2005; accepted in revised form February 16, 2006 Abstract Epithelial-to-mesenchymal transition (EMT) is an essential embryogenic and developmental process, characterized by altered cellular morphology, loss of cell adhesion, and gain of migratory ability. Dysregu- lation of this process has been implicated in tumori- genesis, mediating the acquisition of migratory and invasive phenotypes by tumor cells. Mammary epithe- lial cells provide an excellent model in which to study the process, being derived from mammary gland tissue that utilizes EMT to facilitate branching morphogenesis through which the developing gland migrates into and invades the fat pad. Inappropriate EMT has been heav- ily implicated in the progression of ductal hyperplasia and mammary tumor metastasis. We examined the morphological and molecular changes of three murine mammary epithelial cell lines following EMT induction. EMT was induced in the EpH-4 and NMuMG cell lines by transforming growth factor (TGF)-b1 but not by ethanol, while the KIM-2 cell line was partially resistant to TGF-b1 but responded fully to ethanol. The response to EMT-inducing reagent was shown to be critically dependent on the time of treatment, with confluent cells failing to respond. Timelapse photography identified increased motility during wound healing in cells pre- treated with EMT-inducing reagent compared with un- treated controls. Furthermore, EMT conferred resist- ance to UV-induced apoptosis. Our data indicate that evaluation of characteristics other than loss and gain of phenotypic markers may be of benefit when assessing EMT, and contribute to the evidence suggesting that inappropriate EMT facilitates the acquisition of resist- ance to apoptosis, a key characteristic required for tumor survival. Key words epithelial to mesenchymal transition mammary epithelial cell apoptosis wound healing TGF-b1 Introduction Epithelial–mesenchymal transition (EMT) facilitates crucial functions during embryogenesis and subsequent development. As a result of EMT, polarized epithelial cells lose their polarity, undergo cytoskeletal re-organ- ization, and disruption of cell–cell/cell–substratum con- tacts, and become migratory. These phenotypic changes are associated with an altered profile of cell surface markers, for example increased expression of vimentin. During embryo development, transient acquisition of mesenchymal characteristics confers motility onto epi- thelial cells, enabling spatial reorganization (Hay, 1995). EMT activity is implicated in facilitating em- bryo implantation (Vicovac and Aplin, 1996), palatal fusion (Bloch-Zupan et al., 2001), formation of the heart valves (Wagner et al., 2001), formation of the pe- ripheral nervous system (PNS) (Selleck and Bronner- Fraser, 2000), and in branching morphogenesis of the mammary gland (Fata et al., 2004). Dysregulation of EMT has major consequences for cells and tissues. Failure of normal EMT and activation of inappropriate EMT are both implicated in the development of diverse pathologies including tumor formation and the gener- ation of fibroblasts during organ fibrosis (Guarino Authors’ contribution: E.J.D.R. carried out the majority of the work. W.T.K. contributed to the time-lapse video analysis, K.A. identified the use of ethanol as an inducer of E.M.T. and C.J.W. participated in the design of the study. Ewan J. D. Robson Walid T. Khaled Kathrine Abell Christine J. Watson ( . *) Mammary Apoptosis and Development Group Department of Pathology, University of Cambridge Tennis Court Road Cambridge CB2 1QP, UK E-mail: [email protected] U.S. Copyright Clearance Center Code Statement: 0301–4681/2006/7405–254 $ 15.00/0 Differentiation (2006) 74:254–264 DOI: 10.1111/j.1432-0436.2006.00075.x r 2006, Copyright the Authors Journal compilation r 2006, International Society of Differentiation
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Epithelial-to-mesenchymal transition confers resistance to apoptosis in three murine mammary...

ORIGINAL ARTICLE
Ewan J. D. Robson . Walid T. Khaled . Kathrine Abell .
Christine J. Watson
Epithelial-to-mesenchymal transition confers resistance to apoptosisin three murine mammary epithelial cell lines
Received December 6, 2005; accepted in revised form February 16, 2006
Abstract Epithelial-to-mesenchymal transition (EMT)is an essential embryogenic and developmental process,characterized by altered cellular morphology, loss ofcell adhesion, and gain of migratory ability. Dysregu-lation of this process has been implicated in tumori-genesis, mediating the acquisition of migratory andinvasive phenotypes by tumor cells. Mammary epithe-lial cells provide an excellent model in which to studythe process, being derived from mammary gland tissuethat utilizes EMT to facilitate branching morphogenesisthrough which the developing gland migrates into andinvades the fat pad. Inappropriate EMT has been heav-ily implicated in the progression of ductal hyperplasiaand mammary tumor metastasis. We examined themorphological and molecular changes of three murinemammary epithelial cell lines following EMT induction.EMT was induced in the EpH-4 and NMuMG cell linesby transforming growth factor (TGF)-b1 but not byethanol, while the KIM-2 cell line was partially resistantto TGF-b1 but responded fully to ethanol. The responseto EMT-inducing reagent was shown to be criticallydependent on the time of treatment, with confluent cellsfailing to respond. Timelapse photography identifiedincreased motility during wound healing in cells pre-treated with EMT-inducing reagent compared with un-treated controls. Furthermore, EMT conferred resist-
ance to UV-induced apoptosis. Our data indicate thatevaluation of characteristics other than loss and gain ofphenotypic markers may be of benefit when assessingEMT, and contribute to the evidence suggesting thatinappropriate EMT facilitates the acquisition of resist-ance to apoptosis, a key characteristic required fortumor survival.
Key words epithelial to mesenchymal transition �mammary epithelial cell � apoptosis � wound healing �TGF-b1
Introduction
Epithelial–mesenchymal transition (EMT) facilitatescrucial functions during embryogenesis and subsequentdevelopment. As a result of EMT, polarized epithelialcells lose their polarity, undergo cytoskeletal re-organ-ization, and disruption of cell–cell/cell–substratum con-tacts, and become migratory. These phenotypic changesare associated with an altered profile of cell surfacemarkers, for example increased expression of vimentin.
During embryo development, transient acquisition ofmesenchymal characteristics confers motility onto epi-thelial cells, enabling spatial reorganization (Hay,1995). EMT activity is implicated in facilitating em-bryo implantation (Vicovac and Aplin, 1996), palatalfusion (Bloch-Zupan et al., 2001), formation of theheart valves (Wagner et al., 2001), formation of the pe-ripheral nervous system (PNS) (Selleck and Bronner-Fraser, 2000), and in branching morphogenesis of themammary gland (Fata et al., 2004). Dysregulation ofEMT has major consequences for cells and tissues.Failure of normal EMT and activation of inappropriateEMT are both implicated in the development of diversepathologies including tumor formation and the gener-ation of fibroblasts during organ fibrosis (Guarino
Authors’ contribution: E.J.D.R. carried out the majority of thework. W.T.K. contributed to the time-lapse video analysis,K.A. identified the use of ethanol as an inducer of E.M.T. andC.J.W. participated in the design of the study.
Ewan J. D. Robson � Walid T. Khaled � Kathrine Abell �Christine J. Watson ( .*)Mammary Apoptosis and Development GroupDepartment of Pathology, University of CambridgeTennis Court RoadCambridge CB2 1QP, UKE-mail: [email protected]
U.S. Copyright Clearance Center Code Statement: 0301–4681/2006/7405–254 $ 15.00/0
Differentiation (2006) 74:254–264 DOI: 10.1111/j.1432-0436.2006.00075.xr 2006, Copyright the AuthorsJournal compilationr 2006, International Society of Differentiation

et al., 1999; Lee and Joo, 1999; Iwano et al., 2002;Kalluri and Neilson, 2003). Where inappropriate EMTarises, the mesenchymal cells produced are typicallyhighly motile and more predisposed to promote inva-sion and metastasis (Oft et al., 1996). This subornmentof the EMT process by tumor cells may be a necessarypre-requisite for invasion and metastasis.
In the adult, EMT has a role in wound healing(Iwano et al., 2002). The wound healing mechanism in-volves an inflammatory response and activation ofEMT, leading to a reduction in cell adhesion and pro-motion of cellular motility allowing formation of a cel-lular sheet across the injured tissue (Bissell andRadisky, 2001). In normal circumstances, these chang-es are temporary, reverting after healing. However, sus-tained inflammation can lead to an imbalancedmicroenvironment, with constitutive activation of cyto-kine signalling systems potentially inducing aberrantEMT (Gotzmann et al., 2004). Macrophages releasedduring the wound healing process have the potential toaid tumor progression through secretion of growth andchemotactic factors (Lin and Pollard, 2004), as well asgiving rise to mutagenic oxygen and nitrogen radicalsthat have carcinogenic properties (Pollard, 2004).
The physical processes through which EMT is effect-ed are loss of cell-to-cell adhesion (notably disruption ofthe cadherin–catenin complex; Kim et al., 2002; Egeret al., 2004), disruption of cell-to-substratum adhesion(through matrix metalloproteinase (MMP) activity;Yang and Liu, 2001; Strutz et al., 2002; Zeisberget al., 2002), and cytoskeletal reorganization. EMT ischaracterized by a change in gross morphology, fromthe typical cobblestone-like appearance of the epithelialmonolayer to a spindly, elongated, mesenchymal cellpopulation, and by the loss and gain of various cyto-skeletal proteins. No single set of markers is adoptedthroughout the literature, but commonly cited epithelialmarkers include cytokeratins 8 and 18 and membranelocalized E-cadherin, while vimentin expression, mem-brane localized N-cadherin, and cytoplasmic relocationof E-cadherin are indicative of the mesenchymal phe-notype (O 0Hare et al., 1991; Miettinen et al., 1994;Somasiri et al., 2000; Bhowmick et al., 2001a, 2001b;Dandachi et al., 2001; Zavadil et al., 2001). However,studies vary widely in their stringency of application ofthe various criteria for identification of EMT (Vincent-Salomon and Thiery, 2003). Our investigation aimed toestablish a means of assessing EMT based upon char-acteristics acquired by cell populations following in-duction of phenotypic transition, with an emphasis onfeatures contributing to tumor survival.
Substantial in vitro work has focussed on EMT in-duced in mammary cell lines, particularly in response totransforming growth factor (TGF)-b1, a cytokine in-volved in maintaining homeostasis in a variety of cellsby regulating cell growth and apoptosis (Siegel andMassague, 2003). Interestingly, TGF-b1 has been
shown to induce EMT in cancer cells, to promote an-giogenesis and immunosuppression, and to mediatemetastasis (Li et al., 2001; McEarchern et al., 2001). Weused TGF-b1 as one EMT inducing reagent, assessingits effect in three different mammary epithelial celllines—KIM-2, NmuMG, and EpH-4. A second reagent,ethanol, was identified serendipitously to induce EMTin KIM-2 cells.
The induction and maintenance of EMT was initiallyassessed on the basis of gross morphological changes andgain of vimentin expression in the mesenchymal popula-tion. In order to investigate the additional characteristicsconferred by EMT in addition to the loss/gain ofphenotypic markers, we analysed the effect of EMT onwound healing, using timelapse video microscopy.Wound healing assays have been performed previouslywith EpH-4 cells (Wechselberger et al., 2001), using pho-tographic stills taken several hours apart. The advantageof the timelapse approach, used in recent studies of hu-man corneal cells (Hardarson et al., 2004), was to allowcontinuous observation of the healing process, as well asproviding data on the rate of wound repopulation. Wethen determined the effect of EMT on the apoptotic re-sponse to UV exposure, using immunofluorescence anal-ysis of cleaved caspase 3. These assays indicated thatEMT induction conferred both increased migratory abil-ity and resistance to apoptosis on all three cell lines, twodictators of malignant growth (Hanahan and Weinberg,2000). Thus, these data indicate that evaluation of char-acteristics other than loss and gain of epithelial/me-senchymal markers may be of benefit when assessingEMT, and add to the evidence highlighting the potentialfor dysregulated EMT to promote tumor survival.
Materials and methods
Unless stated otherwise, all reagents were obtained from Sigma(Poole, U.K.).
Cell culture
Three established mouse mammary epithelial cell lines were usedduring the course of this study, KIM-2 (Gordon et al., 2000),NMuMG (Owens, 1974), and EpH-4 (Reichmann et al., 1989).KIM-2 cells are a normal mouse mammary epithelial cell line de-rived by this laboratory from mid-pregnant mice and conditionallyimmortalized using the Simian virus (SV)-40 large T-antigen (Gor-don et al., 2000). Confluent populations of KIM-2 cells were pass-aged as described previously (Gordon et al., 2000) every 2–4 dayswith cells grown on type I collagen every second passage. NMuMGcells are a normal mammary epithelial cell line derived from themammary gland tissue of an adult female NAMRU mouse (Owens,1974), and spontaneously immortalized. NMuMG cells were pass-aged using a dilution ratio of 1:10 every 2–3 days and grown solelyon plastic. EpH-4 cells are a normal mammary epithelial cell linederived from the mammary gland of a mid-pregnant BALB/cmouse and spontaneously immortalized (Reichmann et al., 1989).Confluent populations of EpH-4 cells were passaged as described
255

(Reichmann et al., 1989). EpH-4 cells were passaged using a dilu-tion ratio of 1:10, every 2–4 days and grown solely on plastic.
Induction of EMT
Cells were seeded in six-well plates (Falcon, Franklin Lakes, NJ)and treated either with 2–20 ng/ml TGF-b1 (R&D Systems) or0.1–5.0% ethanol (Fisher Scientific, Loughborough, UK), concur-rent with seeding or 24 hr post-seeding.
Microscopy
All microscopic work was carried out using a Zeiss AxiovertS100TV microscope (Zeiss, Gottingen, Germany) equipped with aHamamatsu C4742-95 ORCA1 digital camera (Hamamatsu Photo-tonics Ltd., Welwyn Garden City, UK). Images were visualized andmanipulated using AQM 2000 Kinetic Acquisition Manager soft-ware (Kinetic Imaging Ltd., UK). Cells were generally fixed inmethanol/acetone before microscopic study, although some exper-iments required images of live cells. In these cases, cells were trans-ferred from the incubator to the microscope and the time spentoutside the incubator kept to a maximum 30min.
Immunofluorescence analysis
Cells for immunofluorescence analysis were fixed in methanol/ac-etone (1:1, v/v), exposed to a blocking solution (phosphate-bufferedsaline (PBS) containing either bovine serum albumin (BSA) or goatserum) for 1–2 hr at room temperature to prevent nonspecific bind-ing, incubated overnight with primary antibody at 41C, washed,incubated with secondary antibody (at RT) and washed again. Cellswere stored in sterile 1� PBS containing gentamycin (1:1,000, LifeTechnologies, Paisley, UK), and viewed within 3 days of immuno-staining.
Vimentin immunofluorescence
Cells were blocked in PBS with 5% BSA (w/v). The primary an-tibody used was a mouse monoclonal anti-vimentin clone,VIM13.2, made up in PBS–BSA at a dilution of 1:200. The sec-ondary antibody used was the goat anti-mouse Ig(Ig)G Alexa Fluor488 (Molecular Probes, Invitrogen Ltd., Paisley, UK). Betweensecondary and primary antibody incubation, cells were washed3� 10min in PBS–BSA. Following secondary antibody incubation,cells were washed for 10min in PBS, followed by 10min incubationin Hoescht dye 33,342 made up in PBS (2 mg/ml), and a final 10minwash in PBS.
Cleaved caspase 3 immunofluorescence
Cells were blocked in PBS with 10% (v/v) goat serum (DakoCyto-mation, Dako N. America Inc., CA). The primary antibody usedwas a mouse monoclonal anti-cleaved caspase 3 antibody (CellSignalling, Danvers, MA), made up in PBS with 5% (v/v) goatserum at a dilution of 1:100. The secondary antibody used was thegoat anti-mouse IgG Alexa Fluor 488 (Molecular Probes). Betweensecondary and primary antibody incubation, cells were washed3� 5min in PBS. Following secondary antibody incubation, cellswere washed for 5min in PBS, followed by 5min incubation inHoescht 33,342 made up in PBS (2mg/ml), and a final 5min washin PBS.
Wound healing
Cells were seeded in six-well plates (Falcon) and grown to conflu-ency (day 3 for all control populations, days 6–9 for ethanol-treatedKIM-2 cells, days 4–6 for TGF-b1-treated NMuMG and EpH-4cells). Medium was aspirated and a gel filter tip drawn carefullythrough the monolayer to create a wound between cells. Cells werethen washed with sterile 1�PBS to remove any loose debris, andfresh medium applied (plus or minus ethanol or TGF-b1 accord-ingly). Plates were then transferred to a heated stage attached to theZeiss microscope, and conditions maintained as in the incubator;371C, 5% CO2, (using TempControl 37-2 digital [Zeiss], and CTI-Controller 3700 digital [Zeiss], respectively) and with a water res-ervoir to maintain humidity. The field of view was centred over asection of the wound, and the timelapse program initiated. Phaseimages were recorded every 8min for up to 48hr. Images wereprocessed to make a film using the Amira 3.0 software package(Template Graphics Software, San Diego, CA). To analyze thewound healing process, a snapshot was taken of the first frame(time5 0) and the distance between the edges of the wound meas-ured using Jasc Paint Shop Pro 7 (Corel, Maidenhead, UK). Thefilm was then advanced frame by frame until the two edges of thewound met. The frame number was recorded and the time for thewound to close established (time5 frame number � 8min). Divid-ing distance by time then gave a rate for the wound healing process.
Resistance to UV-induced apoptosis assay
Cells were seeded in six-well plates and grown to confluency (day 3for all control populations, days 6–9 for ethanol-treated KIM-2 cells,days 4–6 for TGF-b1-treated NMuMG and EpH-4 cells). Mediumwas then aspirated from control and experimental cells alike, whichwere then washed in sterile 1� PBS. UV treatment (UVP CL-100Ultraviolet Crosslinker, 600–6,000mJ/cm2, UVP Ltd., Cambridge,UK) was applied to cells in the absence of liquid to allow full ex-posure (media removed for less than 60 sec in total). Fresh media( � treatment) was added to cells, which were returned to the in-cubator. 6 hr following UV treatment, cells were fixed and subjectedto cleaved caspase 3 immunofluorescence. Five random fields of viewper population were analysed. Merged images were created fromblue (Hoescht 33342-stained nuclei) and green (Alexa Fluor-taggedcleaved caspase 3-positive cells) images. The total number of cellswas established by counting nuclei, and the number of apoptotic cellsdetermined by counting cleaved caspase 3-positive cells (Jasc PaintShop Pro 7). The percentage of cells undergoing apoptosis wascalculated.
Western blotting
Untreated and ethanol/TGF-b1-treated cells were grown to conflu-ency as above. Whole cell extracts were prepared using RIPA buffer(150mMNaCl, 1% NP40, 0.5% sodium deoxycholate, 0.1% sodiumdodecyl sulfate (SDS)) containing the protease inhibitors leupeptin(10mg/ml), pepstatin (1mM), aprotinin (4mg/ml), phenyl-methylsulfonyl fluoride (50mg/ml), and sodium vanadate (100mM).Protein concentrations were measured (Pierce), and 10mg protein foreach sample applied to 12% SDS-polyacrylamide gels. Followingprotein transfer, polyvinylidine difluoride membranes (Millipore,Billerica, MA) were blocked in TBS (5% milk, 0.1% Tween-20) for60min at RT, before overnight incubation at 41C with primary anti-phospho-Stat5 antibody (Cell Signalling, diluted according to man-ufacturer’s instructions). Specifically bound antibody was detectedwith peroxidase conjugated secondary antibody (Dakocytomation),using an enhanced chemiluminescence (ECL) kit (Amersham), andrecorded by exposure to ECL Hyperfilm (Amersham, Little Chalfont,UK). Membranes were stripped and reprobed with Stat5 antibody(Transduction Laboratories) to generate total Stat5 profiles for use asloading controls.
256

Results
Induction of EMT
Our first aim was to establish a robust system for in-vestigating EMT in mammary epithelial cells. In orderto eliminate cell line specific effects, we used three nor-mal immortalized murine mammary cell lines.
KIM-2 cells had been observed to spontaneously giverise to both myoepithelial and luminal cell types (Gor-don et al., 2000). Initial experiments demonstrated theKIM-2 cell line to be only partially responsive to TGF-b1 treatment, failing to generate a homogenous popu-lation of phenotypically switched cells (data notshown). A homogenous population of phenotypicallyswitched cells was generated following treatment with1% ethanol (Fig. 1A), while a 5% treatment inducedtotal cell death. Ethanol treatment induced a morpho-logical change from a classical epithelial cobblestone-like monolayer to a collection of elongated, spindlycells. Double-colour immunofluorescence analysis indi-cated strongly increased vimentin expression in treatedcells by comparison with controls (Fig. 1A). Treatmentof both NMuMG and EpH-4 cells with up to 2% eth-anol failed to induce EMT (data not shown), while 5%ethanol induced cell death.
The response of NMuMG cells to TGF-b1 has beencharacterized previously (Miettinen et al., 1994; Bhow-mick et al., 2001a, 2001b; Zavadil et al., 2001). We ob-served a similar response, with TGF-b1 treatmentresulting in the appearance of elongated cells and in-creased vimentin staining (Fig. 1B). TGF-b1-inducedphenotypic switching of Ha-Ras transformed EpH-4cells has been described (Oft et al., 1998), with a similar
response observed in EpH-4 cells under the control ofan inducible c-Fos oestrogen receptor oncoprotein(Eger et al., 2004). However, it was reported that non-transformed EpH-4 cells fail to undergo EMT inresponse to TGF-b1 (Brown et al., 2004). Surprisingly,we observed a marked response to TGF-b1 treatment(2–20ng/ml) in non-transformed EpH-4 cells, with cellspresenting a spindly, disorganized morphology andstrong vimentin positivity by comparison with untreat-ed cells (Fig. 1C).
The rate at which cell confluency was achieved by allthree cell lines was markedly retarded during inductionof EMT. Ethanol-treated KIM-2 cells became confluentby day 8–9, while untreated cells were confluent by day 3(data not shown). TGF-b1-treated EpH-4 and NMuMGcells reached confluency on day 6 as opposed to day 3 incontrol populations (data not shown). Interestingly,while KIM-2 and NMuMG cells appeared to retaincontact inhibition following EMT induction (as evi-denced by their continued confinement to a monolayer),EpH-4 cells lost such inhibition and layers of prolifer-ating cells piled up on top of each other (Fig. 2).
Response to EMT induction depends on the time oftreatment
In establishing our biological system, we observed anotable feature of EMT induction. With particular re-gard to EpH-4 cells, but also KIM-2 cells, cell popu-lations treated concurrently with seeding presented adifferent appearance to those treated 24 hr post-seeding.In cells treated post-seeding, small islands of cobble-stone-like cells were present throughout the population,
Control EtOH
KIM- 2
Control TGFββ1
EpH-4
Control TGFβ1
NMuMG
A B C
Fig. 1 Epithelial-to-mesenchymal transition (EMT) in three murinemammary epithelial cell lines. (A) KIM-2 cells were cultured ineither maintenance media (MM) or MM supplemented with 1%ethanol (v/v, start of treatment concurrent with seeding) until con-fluent (untreated, 3 days; ethanol-treated, 9 days). (B) NMuMGcells were cultured in either maintenance media (MM) or MMsupplemented with 2 ng/ml transforming growth factor-b1 (TGF)-b1 (start of treatment concurrent with seeding) until confluent (un-treated, 3 days; TGF-b1 treated, 6 days). (C) EpH-4 cells werecultured in either Dulbecco’s modified Eagle’s medium (DMEM) orDMEM supplemented with 2 ng/ml TGF-b1 (start of treatment
concurrent with seeding) until confluent (untreated, 3 days, TGF-b1-treated, 6 days). The upper panels show phase contrast imagesof untreated and treated cells. Comparison of the gross morphologyof these cells shows the change from a cobblestone-like monolayerto a collection of elongated, spindly cells. The lower panels showdouble-color immunofluorescence analysis of vimentin (visualizedwith an anti-vimentin antibody and anti-mouse Alexa fluor 488,green) and nuclei (Hoescht, blue). Low levels of vimentin were ob-served in untreated cells. By contrast, ethanol/TGF-b1-treated cellsare strongly positive for vimentin. Scale bar5 75 nm.
257
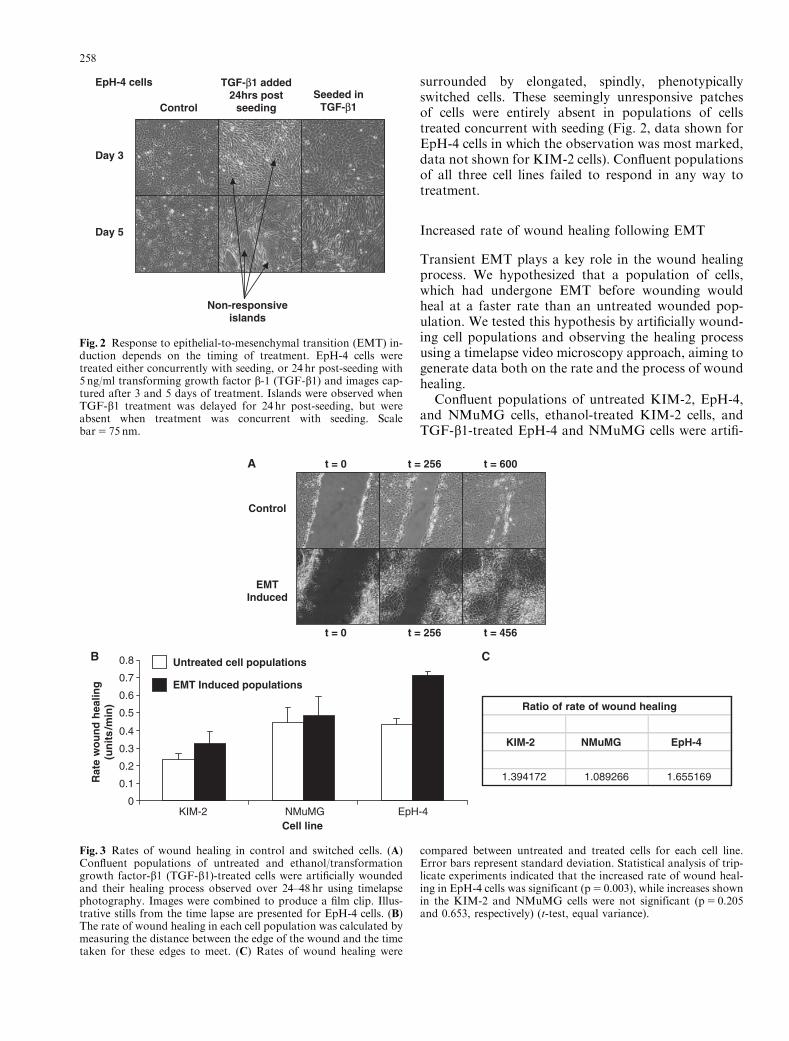
surrounded by elongated, spindly, phenotypicallyswitched cells. These seemingly unresponsive patchesof cells were entirely absent in populations of cellstreated concurrent with seeding (Fig. 2, data shown forEpH-4 cells in which the observation was most marked,data not shown for KIM-2 cells). Confluent populationsof all three cell lines failed to respond in any way totreatment.
Increased rate of wound healing following EMT
Transient EMT plays a key role in the wound healingprocess. We hypothesized that a population of cells,which had undergone EMT before wounding wouldheal at a faster rate than an untreated wounded pop-ulation. We tested this hypothesis by artificially wound-ing cell populations and observing the healing processusing a timelapse video microscopy approach, aiming togenerate data both on the rate and the process of woundhealing.
Confluent populations of untreated KIM-2, EpH-4,and NMuMG cells, ethanol-treated KIM-2 cells, andTGF-b1-treated EpH-4 and NMuMG cells were artifi-
Day 3
Day 5
ControlSeeded in
TGF-β1
TGF-β1 added 24hrs post
seeding
EpH-4 cells
Non-responsive islands
Fig. 2 Response to epithelial-to-mesenchymal transition (EMT) in-duction depends on the timing of treatment. EpH-4 cells weretreated either concurrently with seeding, or 24 hr post-seeding with5 ng/ml transforming growth factor b-1 (TGF-b1) and images cap-tured after 3 and 5 days of treatment. Islands were observed whenTGF-b1 treatment was delayed for 24 hr post-seeding, but wereabsent when treatment was concurrent with seeding. Scalebar5 75 nm.
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
KIM-2 NMuMG EpH-4Cell line
Rat
e w
ou
nd
hea
ling
(un
its
/min
)
t = 0 t = 256 t = 600
t = 0 t = 256 t = 456
Control
EMT Induced
Untreated cell populations
EMT Induced populations
KIM-2 NMuMG EpH-4
1.394172 1.089266 1.655169
Ratio of rate of wound healing
A
B C
Fig. 3 Rates of wound healing in control and switched cells. (A)Confluent populations of untreated and ethanol/transformationgrowth factor-b1 (TGF-b1)-treated cells were artificially woundedand their healing process observed over 24–48 hr using timelapsephotography. Images were combined to produce a film clip. Illus-trative stills from the time lapse are presented for EpH-4 cells. (B)The rate of wound healing in each cell population was calculated bymeasuring the distance between the edge of the wound and the timetaken for these edges to meet. (C) Rates of wound healing were
compared between untreated and treated cells for each cell line.Error bars represent standard deviation. Statistical analysis of trip-licate experiments indicated that the increased rate of wound heal-ing in EpH-4 cells was significant (p5 0.003), while increases shownin the KIM-2 and NMuMG cells were not significant (p5 0.205and 0.653, respectively) (t-test, equal variance).
258

cially wounded by scratching a gel loading tip across themonolayer, and their response recorded over 24–48 hrusing timelapse photography, with images combined toproduce a film clip. The rate of wound healing was cal-culated for each population, and the nature of theprogress of the healing process assessed. Illustrativestills from the time lapse are presented in Fig. 3A. Theseexperiments indicated that wound healing occurred at asignificantly faster rate in EpH-4 cells (p5 0.002) in-duced to undergo EMT by comparison with untreatedcell populations (Figs. 3B and 3C). Critically, while theincreased rate of wound healing in KIM-2 andNMuMG cells was not statistically significant, thesecell lines demonstrated marked differences in the natureof the wound healing process when treated with eth-anol/TGF-b1. Repeated observations of timelapse foot-age indicated that the movement displayed by untreatedKIM-2 and NMuMG cells resembled the epithelial cellsheet migration model described for MDCK cells(Matsubayashi et al., 2004; Farooqui and Fenteany,2005), with cells at each side of the wound moving as acohesive unit. This organized migration was lost in cellsthat had undergone EMT, with cells appearing to moveindependently of their neighbors. Videos of the woundhealing timelapse experiments on which these conclu-sions are based accompany this manuscript (Supple-mentary videos). Descriptions illustrating these videos,and analysis of the timelapse data, are presented by cellline below.
KIM-2 cells
Control: Cells on either edge of the wound movedslowly together, with each side appearing to act as acohesive unit, giving rise to an organised wound healingprocess.
Ethanol-treated: Ethanol-treated KIM-2 cells dem-onstrated markedly different behavior to untreated cellsduring wound healing. Cells moved more quickly, ex-tending pseudopodia to a greater length to bridge thewound with a network of interconnecting spindly cells,yielding a mesh-like monolayer. Cells appeared to moveindependently, and not as part of an organized cell sheetmigration. The remaining gaps in the monolayer werefilled either through continued cell migration, or as aresult of cell division.
NMuMG cells
Control: Cells at either edge of the wound elongatedvery slightly, into the gap. Wound closure was achievedlargely through a slow flattening of cells, evidenced byincreasing distances between cell nuclei, starting at theedge of the wound and moving into the body of theunwounded monolayer. The two edges of the woundwere thus pushed toward each other as organized units.
Cells remained attached throughout, with none detach-ing to migrate independently. Following the re-estab-lishment of cell-to-cell contact, the distance betweennuclei decreased as the cells regained their originalshape.
TGF-b1-treated: NMuMG cells treated with TGF-b1 quickly moved into the wound, accompanied by veryrapid and extreme elongation of pseudopodia. Muchquicker re-establishment of cell-to-cell contact was ob-served than in the untreated population, with cellsclearly migrating independently rather than as a cellsheet. This yielded a mesh of elongated cells, with sub-sequent gap filling achieved through continued migra-tion and/or cell division. The cell flattening feature ofthe control population was not observed in the treatedpopulation.
EpH-4 cells
Control: The wound healing process in untreatedEpH-4 cells occurred in a broadly similar fashion tothat observed in untreated NMuMG cells.
TGF-b1-treated: Treated EpH-4 cells healed in es-sentially the same pattern as that described for untreat-ed EpH-4 cells, with no marked elongation of the cellsinvolved in closing the gap. Cells simply moved at amarkedly faster rate. An interesting feature of thesecells was the impact of the multilayered structure ofEpH-4 populations post EMT. Cells in the bottom layerflattened and moved into the gap, while the upper layersremained static; as the wound closed, the upper layer ofcells began to move over the top of the bottom layer,which in turn was covered by the next layer.
Resistance to UV-induced apoptosis following EMT
Resistance to apoptosis is a common feature in the de-velopment of human cancers, and in tumors that do notrespond to radiation treatment or cytotoxic drugs thatrely on inducing apoptosis (Osford et al., 2004; Vareset al., 2004). We investigated the effect of EMT on theapoptotic response of these cell lines to UV to deter-mine the effect of EMT on resistance to apoptosis.
Confluent populations of untreated KIM-2,NMuMG, and EpH-4 cells, ethanol-treated KIM-2cells (day 9 post-treatment), TGF-b1-treated NMuMGand EpH-4 cells (day 6 post-treatment), and sub-con-fluent treated KIM-2 and NMuMG cells (day 6 and 4post-treatment respectively) were exposed to 1,500 mJ/cm2 UV. This level of UV exposure induced a moderateapoptotic response in untreated populations, anticipat-ed to be sufficiently low to avoid masking any resistanceresulting from EMT in treated populations. Separateplates of cells derived at the same time were not exposedto UV and used as controls. Immunofluorescence anal-
259
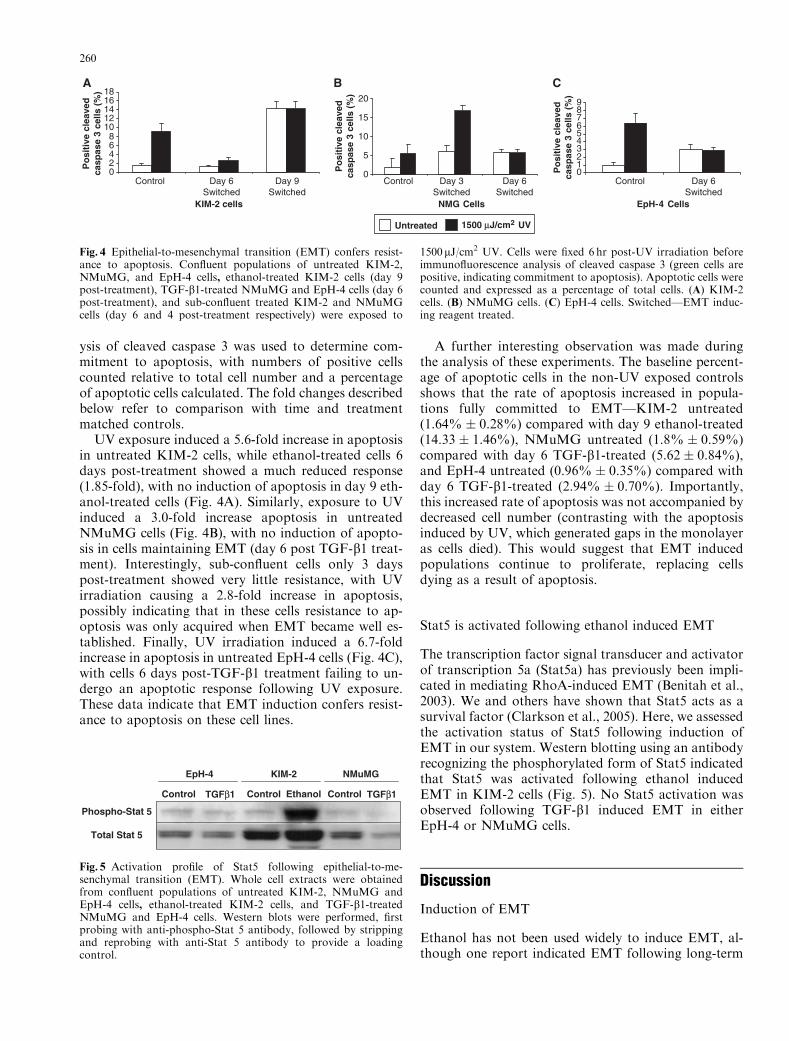
ysis of cleaved caspase 3 was used to determine com-mitment to apoptosis, with numbers of positive cellscounted relative to total cell number and a percentageof apoptotic cells calculated. The fold changes describedbelow refer to comparison with time and treatmentmatched controls.
UV exposure induced a 5.6-fold increase in apoptosisin untreated KIM-2 cells, while ethanol-treated cells 6days post-treatment showed a much reduced response(1.85-fold), with no induction of apoptosis in day 9 eth-anol-treated cells (Fig. 4A). Similarly, exposure to UVinduced a 3.0-fold increase apoptosis in untreatedNMuMG cells (Fig. 4B), with no induction of apopto-sis in cells maintaining EMT (day 6 post TGF-b1 treat-ment). Interestingly, sub-confluent cells only 3 dayspost-treatment showed very little resistance, with UVirradiation causing a 2.8-fold increase in apoptosis,possibly indicating that in these cells resistance to ap-optosis was only acquired when EMT became well es-tablished. Finally, UV irradiation induced a 6.7-foldincrease in apoptosis in untreated EpH-4 cells (Fig. 4C),with cells 6 days post-TGF-b1 treatment failing to un-dergo an apoptotic response following UV exposure.These data indicate that EMT induction confers resist-ance to apoptosis on these cell lines.
A further interesting observation was made duringthe analysis of these experiments. The baseline percent-age of apoptotic cells in the non-UV exposed controlsshows that the rate of apoptosis increased in popula-tions fully committed to EMT—KIM-2 untreated(1.64% � 0.28%) compared with day 9 ethanol-treated(14.33 � 1.46%), NMuMG untreated (1.8% � 0.59%)compared with day 6 TGF-b1-treated (5.62 � 0.84%),and EpH-4 untreated (0.96% � 0.35%) compared withday 6 TGF-b1-treated (2.94% � 0.70%). Importantly,this increased rate of apoptosis was not accompanied bydecreased cell number (contrasting with the apoptosisinduced by UV, which generated gaps in the monolayeras cells died). This would suggest that EMT inducedpopulations continue to proliferate, replacing cellsdying as a result of apoptosis.
Stat5 is activated following ethanol induced EMT
The transcription factor signal transducer and activatorof transcription 5a (Stat5a) has previously been impli-cated in mediating RhoA-induced EMT (Benitah et al.,2003). We and others have shown that Stat5 acts as asurvival factor (Clarkson et al., 2005). Here, we assessedthe activation status of Stat5 following induction ofEMT in our system. Western blotting using an antibodyrecognizing the phosphorylated form of Stat5 indicatedthat Stat5 was activated following ethanol inducedEMT in KIM-2 cells (Fig. 5). No Stat5 activation wasobserved following TGF-b1 induced EMT in eitherEpH-4 or NMuMG cells.
Discussion
Induction of EMT
Ethanol has not been used widely to induce EMT, al-though one report indicated EMT following long-term
Control TGFβ1
EpH-4
Control TGFβ1
NMuMG
Control Ethanol
KIM-2
Phospho-Stat 5
Total Stat 5
Fig. 5 Activation profile of Stat5 following epithelial-to-me-senchymal transition (EMT). Whole cell extracts were obtainedfrom confluent populations of untreated KIM-2, NMuMG andEpH-4 cells, ethanol-treated KIM-2 cells, and TGF-b1-treatedNMuMG and EpH-4 cells. Western blots were performed, firstprobing with anti-phospho-Stat 5 antibody, followed by strippingand reprobing with anti-Stat 5 antibody to provide a loadingcontrol.
02468
1012141618
Control Day 6Switched
Day 9Switched
Control Day 3 Switched
Day 6Switched
KIM-2 cells
Po
siti
ve c
leav
edca
spas
e 3
cells
(%
)
Po
siti
ve c
leav
edca
spas
e 3
cells
(%
)
Po
siti
ve c
leav
edca
spas
e 3
cells
(%
)
0
5
10
15
20
NMG Cells
0123456789
Control Day 6Switched
EpH-4 Cells
Untreated 1500 µJ/cm UV
A B C
Fig. 4 Epithelial-to-mesenchymal transition (EMT) confers resist-ance to apoptosis. Confluent populations of untreated KIM-2,NMuMG, and EpH-4 cells, ethanol-treated KIM-2 cells (day 9post-treatment), TGF-b1-treated NMuMG and EpH-4 cells (day 6post-treatment), and sub-confluent treated KIM-2 and NMuMGcells (day 6 and 4 post-treatment respectively) were exposed to
1500mJ/cm2 UV. Cells were fixed 6 hr post-UV irradiation beforeimmunofluorescence analysis of cleaved caspase 3 (green cells arepositive, indicating commitment to apoptosis). Apoptotic cells werecounted and expressed as a percentage of total cells. (A) KIM-2cells. (B) NMuMG cells. (C) EpH-4 cells. Switched—EMT induc-ing reagent treated.
260

treatment of immortalized human gingival keratin-ocytes with ethanol (Chamulitrat et al., 2003). Here,we demonstrate induction of EMT in one mammaryepithelial cell line (KIM-2) in response to ethanol, withconcurrent activation of Stat5, a transcription factorpreviously identified as a mediator of RhoA-inducedEMT (Benitah et al., 2003). Stat5 is a known survivalfactor, and an important future goal will be to ascertainits contribution to the apoptotic resistance gained fol-lowing ethanol-induced EMT. Interestingly, Stat5 wasnot activated in response to TGF-b1 in NMuMG orEpH-4 cells, an observation which suggests potentialmechanistic differences between ethanol and TGF-b1-induced EMT.
The 1% ethanol treatment we used is equivalent to aconcentration of 794mg/dl. In blood, more than500mg/dl alcohol is potentially fatal (Birnbaumer andBessen, 1992). Interestingly, we observed around 50%EMT using 0.2% ethanol (data not shown), a concen-tration of 159mg/dl. Blood alcohol of 150mg/dl resultsin difficulties with walking and balance (Birnbaumerand Bessen, 1992). Chronic ethanol abuse in humanpopulations is associated with various pathologies, in-cluding increased risk of cancer (Aroor and Shukla,2004). The role of ethanol or its metabolites in mam-mary neoplasm has not yet been characterized, despiteepidemiological evidence of a link between ethanol in-take increased risk of breast cancer (Fan et al., 2000;Izevbigie et al., 2002; Aroor and Shukla, 2004) and thesuggestion that combining oestrogen replacement ther-apy and alcohol intake synergistically enhances therisk of breast cancer. We suggest that investigatingethanol-induced EMT may yield useful information
regarding the role of alcohol in increasing the risk ofbreast cancer.
The response of NMuMG cells to TGF-b1 has beenwell documented (Miettinen et al., 1994; Piek et al.,1999; Bhowmick et al., 2001a, 2001b). Our demonstra-tion of TGF-b1-induced EMT in EpH-4 cells is novel,contrasting with the results of Brown et al. (2004). Thereason for this discrepancy is not clear, but our dataindicate that confluent cells fail to undergo EMT inresponse to TGF-b1, suggesting that induction of EMTis critically dependent on either cell cycle status or theextent of cell-to-cell contact. Interestingly, in both theKIM-2 and EpH-4 cell lines, patches of non-responsivecells occur in populations where ethanol/TGF-b1treatment was delayed until 24 hr post-seeding, whichdid not arise in populations treated concurrent withseeding.
One explanation for this observation might be thatthe islands arise from a more strongly epithelial sub-population, growth of which is inhibited owing to thepresence of ethanol or TGF-b1, and that the pheno-typically switched cell populations arise from a sub-population already of a more mesenchymal character(both cell lines are heterogeneously populated). How-ever, this appears unlikely, given the very small fibro-blast sub-populations present in either cell line. Whileproliferation is retarded in treated cells compared withuntreated cells, confluency is still achieved at a ratesuggesting involvement of the bulk of the seeded cells,i.e. the epithelial population.
An alternative model is presented schematically (Fig.6). KIM-2 and EpH-4 cells are seeded in clusters ofseveral cells with partial cell-to-cell contact, and treat-
Seeded directly with EMTinduction treatment
Cells seeded as a
clusterProliferation innormal media Addition EMT
induction treatment
1
2
34
Fig. 6 Model of resistance to epithelial-to-mesenchymal transition(EMT) induction in cell clusters. 1, Epithelial cells are seeded inclusters to maintain some degree of cell-to-cell contact. 2, Cellsseeded in clusters are not cell-to-cell contacted around their entireperiphery. Addition of EMT inducting reagent causes these cells toundergo EMT, giving rise to a homogeneous monolayer of
switched cells. 3, Cells seeded in normal growth media proliferate;the center of these clusters becomes extensively cell-to-cell contact-ed. 4, Cells in extensive cell-to-cell contact do not respond to EMTinduction reagent, while surrounding cells with less cell-to-cell con-tact undergo EMT, forming islands of non-responsive cells withinswathes of phenotypically switched cells.
261

ment with ethanol or TGF-b1 at this time yields con-fluent populations demonstrating complete EMT. Incontrast, after 24 hr of proliferation, cells at the centerof the clusters have established extensive cell-to-cellcontact, a state reported to inhibit EMT (Jenndahlet al., 2005). This could explain the appearance of non-responsive islands of cells. Further support for thismodel is provided by the absence of a response to EMTinducing reagents shown by confluent, fully cell-to-cellcontacted populations.
Characterization of EMT
Basic morphological and cell marker characterizationwas carried out to confirm data on EMT induction re-ported by previous groups using the NMuMG cell linewas reproducible in our hands, thus providing a refer-ence point. We then completed the picture by generat-ing novel data for the KIM-2 and EpH-4 cell lines.However, we also generated new data with respect tothe NMuMG cell line. Miettinen et al. (1994) observedthe response of NMuMG cells treated with TGF-b1 for24–48 hr. Piek et al. (1999) treated cells for 36 hr, andXie et al. (2003) carried out microarray experiments onTGF-b1 responses at 1, 6, and 24 hr post-treatment. Wereport observations arising from cells treated for up to 6(NMuMG and EpH-4) or 9 days (KIM-2). It is clearfrom the results of the apoptotic resistance assays pre-sented here that resistance developed in the period be-tween 3 and 6 days post-treatment in NMuMG cells.While previous studies have focused on early responsedata, we have demonstrated that later responses are alsoimportant.
EMT contribution to tumorigenesis
Hanahan and Weinberg (2000) described six character-istics as key to dictating malignant growth. We ob-served up to three of these that were affected as a resultof EMT induction in all three cell lines.
The first of these characteristics was cell migration.Wound healing assays demonstrated a significantly in-creased rate of healing in EpH-4 cells treated to undergoEMT. While EMT induction did not significantly in-crease the rate of wound healing in KIM-2 andNMuMG cells, a distinct change in the healing proc-ess was observed. The organized cell sheet migrationobserved in untreated cells following wounding waslost, and ethanol/TGF-b1-treated cells appeared to gainthe ability to move independently of the cell sheet. Therecapitulation of any such dysregulated cell migrationcould clearly contribute to increased invasive or meta-static potential in vivo.
A second characteristic was gain of resistance to ap-optosis. In all three cell lines, resistance to UV-inducedapoptosis was conferred following induction of EMT.
Gain of resistance to apoptosis promotes a greatly in-creased risk of tumorigenic initiation events leading tothe establishment of a viable tumor. Previous reportsindicate that TGF-b treatment of rat fetal hepatocytesinduced EMT in a sub-population which subsequentlydemonstrated resistance to TGF-b-mediated apoptosis(Valdes et al., 2002), while Snail, a transcription factorheavily implicated in the promotion of EMT duringdevelopment, conferred resistance to apoptosis inducedby TNF-a and serum deprivation in MDCK cells (Vegaet al., 2004). In our preliminary studies, KIM-2 cells,which differentiate in response to lactogenic hormonetreatment and are normally subject to extensive apop-tosis following hormone withdrawal (Gordon et al.,2000), became resistant following partial EMT inducedby TGF-b1 (data not shown). Our data provide a usefulcontribution to the evidence indicating a role for EMTin the mediation of anti-apoptotic mechanism(s).
Our apoptosis resistance assay indicated that thebaseline rate of apoptosis increased in EMT inducedpopulations of all three cell lines by comparison withuntreated cells, but this feature was not accompanied byreduced cell number (apoptosis assay raw data, notshown). This suggests that while there was an increaseddeath signal in these populations, any cells that diedwere replaced. Continued investigation is required toestablish precisely how cells are replaced in this situa-tion, but it is interesting to speculate that a third char-acteristic of malignant growth, disrupted regulation ofproliferation, may be a contributory factor.
Conclusion
All three cell lines used in this study demonstrated in-creased tumorigenic characteristics in response to theinduction of EMT. Given the associated biological im-portance of the two different EMT inducing reagentsused, further investigation of the signaling pathwaysand downstream genetic targets involved is warranted.This study highlights an alternative approach to char-acterising EMT, suggesting that cell behavior, such asacquisition of increased motility and resistance to ap-optosis, is useful additional information for assessingphenotype in addition to surface marker profile.
Acknowledgments Competing interests: none declared.
Supplementary material
The following supplementary material is available forthis article online:
Video Clip S1. Wound healing in normal EpH4 cells.Video Clip S2.Wound healing in switched EpH4 cells.
262

Video Clip S3. Wound healing in normal KIM-2cells.
Video Clip S4. Wound healing in switched KIM-2cells.
Video Clip S5. Wound healing in normal NMuMGcells.
Video Clip S6. Wound healing in switched NMuMGcells.
This material is available as part of the online articlefrom http://www.blackwell-synergy.com
References
Aroor, A.R. and Shukla, S.D. (2004) MAP kinase signaling in di-verse effects of ethanol. Life Sci 74:2339–2364.
Benitah, S.A., Valeron, P.F., Rui, H. and Lacal, J.C. (2003)STAT5a activation mediates the epithelial to mesenchymal tran-sition induced by oncogenic RhoA. Mol Biol Cell 14:40–53.
Bhowmick, N.A., Ghiassi, M., Bakin, A., Aakre, M., Lundquist,C.A., Engel, M.E., Arteaga, C.L. and Moses, H.L. (2001a)Transforming growth factor-beta1 mediates epithelial to me-senchymal transdifferentiation through a RhoA-dependentmechanism. Mol Biol Cell 12:27–36.
Bhowmick, N.A., Zent, R., Ghiassi, M., McDonnell, M. and Mo-ses, H.L. (2001b) Integrin beta 1 signaling is necessary for trans-forming growth factor-beta activation of p38MAPK andepithelial plasticity. J Biol Chem 276:46707–46713.
Birnbaumer and Bessen (1992). Alcohols and glycols. In: Emer-gency medicine—concepts and clinical practice, 3rd edition.p. 2520.
Bissell, M.J. and Radisky, D. (2001) Putting tumors in context. NatRev Cancer 1:46–54.
Bloch-Zupan, A., Hunter, N., Manthey, A. and Gibbins, J. (2001)R-twist gene expression during rat palatogenesis. Int J Dev Biol45:397–404.
Brown, K.A., Aakre, M.E., Gorska, A.E., Price, J.O., Eltom, S.E.,Pietenpol, J.A. and Moses, H.L. (2004) Induction by transform-ing growth factor-beta1 of epithelial to mesenchymal transition isa rare event in vitro. Breast Cancer Res 6:R215–R231.
Chamulitrat, W., Schmidt, R., Chunglok, W., Kohl, A. and Toma-kidi, P. (2003) Epithelium and fibroblast-like phenotypes derivedfrom HPV16 E6/E7-immortalized human gingival keratino-cytes following chronic ethanol treatment. Eur J Cell Biol 82:313–322.
Clarkson, R.W., Boland, M.P., Kritikou, E.A., Lee, J.M., Free-man, T.C., Tiffen, P.G. and Watson, C.J. (2005) The genes in-duced by Stat3 and Stat5 in mammary epithelial cells definethe roles of these stats in mammary development. MolEndocrinol.
Dandachi, N., Hauser-Kronberger, C., More, E., Wiesener, B.,Hacker, G.W., Dietze, O. and Wirl, G. (2001) Co-expression oftenascin-C and vimentin in human breast cancer cells indicatesphenotypic transdifferentiation during tumor progression: cor-relation with histopathological parameters, hormone receptors,and oncoproteins. J Pathol 193:181–189.
Eger, A., Stockinger, A., Park, J., Langkopf, E., Mikula, M.,Gotzmann, J., Mikulits, W., Beug, H. and Foisner, R. (2004)beta-Catenin and TGFbeta signalling cooperate to maintain amesenchymal phenotype after FosER-induced epithelial to me-senchymal transition. Oncogene 23:2672–2680.
Fan, S., Meng, Q., Gao, B., Grossman, J., Yadegari, M., Goldberg,I.D. and Rosen, E.M. (2000) Alcohol stimulates estrogen recep-tor signaling in human breast cancer cell lines. Cancer Res60:5635–5639.
Farooqui, R. and Fenteany, G. (2005) Multiple rows of cells behindan epithelial wound edge extend cryptic lamellipodia to collec-tively drive cell-sheet movement. J Cell Sci 118:51–63.
Fata, J.E., Werb, Z. and Bissell, M.J. (2004) Regulation of mam-mary gland branching morphogenesis by the extracellular matrixand its remodeling enzymes. Breast Cancer Res 6:1–11.
Gordon, K.E., Binas, B., Chapman, R.S., Kurian, K.M., Clarkson,R.W., Clark, A.J., Lane, E.B. and Watson, C.J. (2000) A novelcell culture model for studying differentiation and apoptosis inthe mouse mammary gland. Breast Cancer Res 2:222–235.
Gotzmann, J., Mikula, M., Eger, A., Schulte-Hermann, R.,Foisner, R., Beug, H. and Mikulits, W. (2004) Molecular as-pects of epithelial cell plasticity: implications for local tumor in-vasion and metastasis. Mutat Res 566:9–20.
Guarino, M., Micheli, P., Pallotti, F. and Giordano, F. (1999)Pathological relevance of epithelial and mesenchymal phenotypeplasticity. Pathol Res Pract 195:379–389.
Hanahan, D. and Weinberg, R.A. (2000) The hallmarks of cancer.Cell 100:57–70.
Hardarson, T., Hanson, C., Claesson, M. and Stenevi, U. (2004)Time-lapse recordings of human corneal epithelial healing. ActaOphthalmol Scand 82:184–188.
Hay, E.D. (1995) An overview of epithelio–mesenchymal transfor-mation. Acta Anat (Basel) 154:8–20.
Iwano, M., Plieth, D., Danoff, T.M., Xue, C., Okada, H. and Ne-ilson, E.G. (2002) Evidence that fibroblasts derive from epithe-lium during tissue fibrosis. J Clin Invest 110:341–350.
Izevbigie, E.B., Ekunwe, S.I., Jordan, J. and Howard, C.B. (2002)Ethanol modulates the growth of human breast cancer cells invitro. Exp Biol Med (Maywood) 227:260–265.
Jenndahl, L.E., Isakson, P. and Baeckstrom, D. (2005) c-erbB2-induced epithelial–mesenchymal transition in mammary epithe-lial cells is suppressed by cell–cell contact and initiated prior to E-cadherin downregulation. Int J Oncol 27:439–448.
Kalluri, R. and Neilson, E.G. (2003) Epithelial–mesenchymal tran-sition and its implications for fibrosis. J Clin Invest 112:1776–1784.
Kim, K., Lu, Z. and Hay, E.D. (2002) Direct evidence for a role ofbeta-catenin/LEF-1 signaling pathway in induction of EMT. CellBiol Int 26:463–476.
Lee, E.H. and Joo, C.K. (1999) Role of transforming growth fac-tor-beta in transdifferentiation and fibrosis of lens epithelial cells.Invest Ophthalmol Vis Sci 40:2025–2032.
Li, C., Guo, B., Bernabeu, C. and Kumar, S. (2001) Angiogenesis inbreast cancer: the role of transforming growth factor beta andCD105. Microsc Res Tech 52:437–449.
Lin, E.Y. and Pollard, J.W. (2004) Macrophages: modulators ofbreast cancer progression. Novartis Found Symp 256:158–168;discussion 168–172, 259–269.
Matsubayashi, Y., Ebisuya, M., Honjoh, S. and Nishida, E. (2004)ERK activation propagates in epithelial cell sheets and regulatestheir migration during wound healing. Curr Biol 14:731–735.
McEarchern, J.A., Kobie, J.J., Mack, V., Wu, R.S., Meade-Tollin,L., Arteaga, C.L., Dumont, N., Besselsen, D., Seftor, E., Hen-drix, M.J., Katsanis, E. and Akporiaye, E.T. (2001) Invasion andmetastasis of a mammary tumor involves TGF-beta signaling.Int J Cancer 91:76–82.
Miettinen, P.J., Ebner, R., Lopez, A.R. and Derynck, R. (1994)TGF-beta induced transdifferentiation of mammary epithelialcells to mesenchymal cells: involvement of type I receptors. J CellBiol 127:2021–2036.
O0Hare, M.J., Ormerod, M.G., Monaghan, P., Lane, E.B. andGusterson, B.A. (1991) Characterization in vitro of luminal andmyoepithelial cells isolated from the human mammary gland bycell sorting. Differentiation 46:209–221.
Oft, M., Heider, K.H. and Beug, H. (1998) TGFbeta signaling isnecessary for carcinoma cell invasiveness and metastasis. CurrBiol 8:1243–1252.
Oft, M., Peli, J., Rudaz, C., Schwarz, H., Beug, H. and Reichmann,E. (1996) TGF-beta1 and Ha-Ras collaborate in modulating the
263

phenotypic plasticity and invasiveness of epithelial tumor cells.Genes Dev 10:2462–2477.
Osford, S.M., Dallman, C.L., Johnson, P.W., Ganesan, A. andPackham, G. (2004) Current strategies to target the anti-apoptoticBcl-2 protein in cancer cells. Curr Med Chem 11:1031–1039.
Owens, R.B. (1974) Glandular epithelial cells from mice: amethod for selective cultivation. J Natl Cancer Inst 52:1375–1378.
Piek, E., Moustakas, A., Kurisaki, A., Heldin, C.H. and ten Dijke,P. (1999) TGF-(beta) type I receptor/ALK-5 and Smad proteinsmediate epithelial to mesenchymal transdifferentiation inNMuMG breast epithelial cells. J Cell Sci 112(Part 24): 4557–4568.
Pollard, J.W. (2004) Tumor-educated macrophages promote tumorprogression and metastasis. Nat Rev Cancer 4:71–78.
Reichmann, E., Ball, R., Groner, B. and Friis, R.R. (1989) Newmammary epithelial and fibroblastic cell clones in coculture formstructures competent to differentiate functionally. J Cell Biol108:1127–1138.
Selleck, M.A. and Bronner-Fraser, M. (2000) Avian neural crestcell fate decisions: a diffusible signal mediates induction of neuralcrest by the ectoderm. Int J Dev Neurosci 18:621–627.
Siegel, P.M. and Massague, J. (2003) Cytostatic and apoptotic ac-tions of TGF-beta in homeostasis and cancer. Nat Rev Cancer3:807–821.
Somasiri, A., Wu, C., Ellchuk, T., Turley, S. and Roskelley, C.D.(2000) Phosphatidylinositol 3-kinase is required for adherensjunction-dependent mammary epithelial cell spheroid formation.Differentiation 66:116–125.
Strutz, F., Zeisberg, M., Ziyadeh, F.N., Yang, C.Q., Kalluri, R.,Muller, G.A. and Neilson, E.G. (2002) Role of basic fibroblastgrowth factor-2 in epithelial–mesenchymal transformation. Kid-ney Int 61:1714–1728.
Valdes, F., Alvarez, A.M., Locascio, A., Vega, S., Herrera, B.,Fernandez, M., Benito, M., Nieto, M.A. and Fabregat, I. (2002)The epithelial mesenchymal transition confers resistance to theapoptotic effects of transforming growth factor Beta in fetal rathepatocytes. Mol Cancer Res 1:68–78.
Vares, G., Ory, K., Lectard, B., Levalois, C., Altmeyer-Morel, S.,Chevillard, S. and Lebeau, J. (2004) Progesterone prevents ra-diation-induced apoptosis in breast cancer cells. Oncogene23:4603–4613.
Vega, S., Morales, A.V., Ocana, O.H., Valdes, F., Fabregat, I. andNieto, M.A. (2004) Snail blocks the cell cycle and confers resist-ance to cell death. Genes Dev 18:1131–1143.
Vicovac, L. and Aplin, J.D. (1996) Epithelial–mesenchymal tran-sition during trophoblast differentiation. Acta Anat (Basel)156:202–216.
Vincent-Salomon, A. and Thiery, J.P. (2003) Host microenvironmentin breast cancer development: epithelial–mesenchymal transi-tion in breast cancer development. Breast Cancer Res 5:101–106.
Wagner, M., Miles, K. and Siddiqui, M.A. (2001) Early deve-lopmental expression pattern of retinoblastoma tumor suppres-sor mRNA indicates a role in the epithelial-to-mesenchymetransformation of endocardial cushion cells. Dev Dyn 220:198–211.
Wechselberger, C., Ebert, A.D., Bianco, C., Khan, N.I., Sun, Y.,Wallace-Jones, B., Montesano, R. and Salomon, D.S. (2001)Cripto-1 enhances migration and branching morphogenesis ofmouse mammary epithelial cells. Exp Cell Res 266:95–105.
Xie, L., Law, B.K., Aakre, M.E., Edgerton, M., Shyr, Y.,Bhowmick, N.A. and Moses, H.L. (2003) Transforming growthfactor beta-regulated gene expression in a mouse mammarygland epithelial cell line. Breast Cancer Res 5:R187–R198.
Yang, J. and Liu, Y. (2001) Dissection of key events in tubularepithelial to myofibroblast transition and its implications in renalinterstitial fibrosis. Am J Pathol 159:1465–1475.
Zavadil, J., Bitzer, M., Liang, D., Yang, Y.C., Massimi, A., Kneitz,S., Piek, E. and Bottinger, E.P. (2001) Genetic programs of ep-ithelial cell plasticity directed by transforming growth factor-be-ta. Proc Natl Acad Sci USA 98:6686–6691.
Zeisberg, M., Maeshima, Y., Mosterman, B. and Kalluri, R. (2002)Renal fibrosis. Extracellular matrix microenvironment regulatesmigratory behavior of activated tubular epithelial cells. Am JPathol 160:2001–2008.
264




















