
Renal Intercalated Cells Sense and Mediate Inflammation via the P2Y14 Receptor
Upload
independentCategory
view
1download
0
Bone 48 (2011) 533ndash542
Contents lists available at ScienceDirect
Bone
j ourna l homepage wwwe lsev ie rcom locate bone
EphBephrin-B interactions mediate human MSC attachment migration andosteochondral differentiation
Agnieszka Arthur a1 Andrew Zannettino b2 Romana Panagopoulos a Simon A Koblar c2 Natalie A Sims d3Con Stylianou e4 Koichi Matsuo f4 Stan Gronthos a5
a Mesenchymal Stem Cell Group Department of Haematology Institute of Medical and Veterinary ScienceHanson Institute and Centre for Stem Cell ResearchRobinson InstituteUniversity of Adelaide Adelaide SA Australiab Myeloma Research Program Department of Haematology Institute of Medical and Veterinary Science Centre for Cancer BiologySA Pathology and Centre for Stem Cell ResearchRobinson Institute University of Adelaide Adelaide SA Australiac School of Molecular and Biomedical Science School of Medicine Centre for Stem Cell ResearchRobinson Institute University of Adelaide Adelaide SA Australiad Bone Cell Biology and Disease Unit St Vincents Institute of Medical Research and The Department of Medicine at St Vincents Hospital The University of Melbourne Australiae Stem Cells and Tissue Repair (VNSC) Institute of Medical Biology Immunos Biomedical Grove Singaporef Center for Integrated Medical Research Graduate School of Medicine Keio University Shinjuku-ku Tokyo Japan
Funding source This work is supported by a granMedical Research Council Conflict of interest There was no conflict of inter
Corresponding author Mesenchymal Stem Cell Grogy Institute of Medical and Veterinary ScienceHanson5000 SA Australia Fax +61 8 8222 3139
E-mail addresses agnesarthurhealthsagovau (andrewzannettinoimvssagovau (A Zannettino) sim(SA Koblar) nsimssvieduau (NA Sims) ConStylian(C Stylianou) matsuoscitckeioacjp (K Matsuo) sta(S Gronthos)
1 Conception and design collection assemblyinterpretation manuscript writing
2 Conception and design data analysis and interpretprovision of study material
3 Manuscript writing provision of study mateinterpretation
4 Manuscript writing and provision of study materi5 Conception and design assembly of data data
manuscript writing provision of study material finalfinancial support
8756-3282$ ndash see front matter copy 2010 Elsevier Inc Aldoi101016jbone201010180
a b s t r a c t
a r t i c l e i n f o
Article historyReceived 5 May 2010Revised 29 October 2010Accepted 29 October 2010Available online 5 November 2010
Edited by J Aubin
KeywordsStem cellBoneCartilageEphEphrinMesenchymal stem cell
Bone marrow derived mesenchymal stemstromal cells (MSC) contribute to skeletal tissue formation and theregulation of haematopoiesis The Ephephrin family of receptor tyrosine kinases is potentially important inthe maintenance of the stem cell niche within neural intestinal and dental tissues and has recently beenshown to play a role in regulating bone homeostasis However the contribution of EphBephrin-B moleculesin human MSC function remains to be determined In the present study EphB and ephrin-B molecules wereexpressed by ex vivo expanded human MSC populations and within human bone marrow trephine samplesTo elucidate the contribution of EphBephrin-B molecules in MSC recruitment we performed functionalspreading and migration assays and showed that reverse ephrin-B signalling inhibited MSC attachment andspreading by activating Src- PI3Kinase- and JNK-dependent signalling pathways In contrast forward EphB2signalling promoted MSC migration by activating the Src kinase- and Abl-dependent signalling pathwaysFurthermore activation of ephrin-B1 andor ephrin-B2 molecules expressed by MSC was found to increaseosteogenic differentiation while ephrin-B1 activation promoted chondrogenic differentiation Theseobservations suggest that EphBephrin-B interactions may mediate the recruitment migration anddifferentiation of MSC during bone repair
t from the National Health and
est in this studyoup Department of Haematol-Institute Frome Road Adelaide
A Arthur)onkoblaradelaideeduauouimba-staredusgngronthoshealthsagovau
of data data analysis and
ation manuscript writing and
rial and data analysis and
alanalysis and interpretationapproval of manuscript and
l rights reserved
copy 2010 Elsevier Inc All rights reserved
Introduction
The bone marrow harbours a non-hematopoietic population ofstromal progenitor cells referred to as mesenchymal stemstromalcells (MSC) that reside within a perivascular niche [12] MSC arecentral to skeletal tissue homeostasis [34] and serve as a primarysource of osteogenic progenitor cells to the bone forming osteo-blasts [5] Various factors including parathyroid hormone humangrowth hormone 125-dihydroxyVitaminD3 prostaglandin E2transforming growth factor-β and bone morphogenetic proteinfamily members regulate osteoblast differentiation [6ndash8] Cellularsignalling cascades activated by these osteogenic factors promoteosteoblast function through the production of mature osteoblasticproteins such as bone sialoprotein osteopontin osteocalcinalkaline phosphatase and type I collagen [6ndash8] To date little isknown about the molecular signals that maintain multi-potentialMSC populations within the stem cell niche and what mechanismsdrive the mobilization of MSC towards the bone surfaces tocontribute to bone formation
534 A Arthur et al Bone 48 (2011) 533ndash542
Recent studies suggest that Ephephrin molecules of bothsubclasses regulate bone homeostasis [9ndash11] These membranebound receptor tyrosine kinases predominantly function by inhibitoryor repulsive cellular responses via cellndashcell contact where theresponses can be mediated through the receptor (forward signalling)andor the ligand (reverse signalling) [1213] The involvement of theEph family in cell motility and cell fate decisions is well defined duringaxon guidance [1415] neural progenitor migration [16] and neuralcrest boundary formation [1718] EphBephrin-B signalling alsomodulates hematopoietic cell migration where EphB4-expressinghematopoietic cell migration is repressed by ephrin-B2 expressingstromal cells [19]
We have previously demonstrated that the bi-directional interac-tion between the receptor and ligand restricts human dental pulpstem cells (DPSC) to their perivascular niche under normal physio-logical conditions wheremobilisation of DPSC occurs following injurythrough the down regulation of ephrin-B1 [2021] The Eph family ofreceptor tyrosine kinases have also been implicated in cell fatedecisions [22] of neural crest cells [23] neuronal precursors [24]intestinal epithelial cells [25] hematopoietic lineages [26] skeletaldevelopment [27ndash30] and now in bone remodelling [9ndash1131]
The importance of Ephephrin molecules during bone formation[3233] and bone homeostasis [9] has recently been highlighted by thestudies showing that the osteogenic potential of osteoblasts wasenhanced by forward signalling via EphB4 while the differentiationand function of osteoclasts was inhibited by reverse signalling throughephrin-B2 [9] EphB2 and ephrin-B1were also implicated in this processhowever to a lesser extent Moreover Allan et al [10] have reported anadditional layer of complexity in this process whereby osteoblastexpressionof ephrin-B2was regulatedbyparathyroidhormone receptor1 in a dose dependent manner which in turn may regulate boneformationor osteogenesis in aparacrineor autocrinemanner [10]Whilestudies examining the role of Ephephrinmolecules in bone homeostasishave focused on EphB4 and ephrin-B2 ephrin-B1 expression within thetumour tissue and the surrounding blood vessels has been correlated topoor clinical prognosis for osteosarcoma [31] Moreover completeablation of ephrin-B1 results in perinatal lethality associated with arange of phenotypes including defects in neural crest cell-derivedtissues incomplete body wall closure and abnormal skeletal patterning[28] More specifically ephrin-B1 is important for the correct formationof cranial sutures digit and rib formation and sternebrae fusion [2734]Twigg et al [34] have also demonstrated that unrelated females withcraniofrontonasal syndrome (CFNS) displayed heterozygous ephrin-B1loss of function which was correlated to disrupted boundary formationin developing coronal sutures [34] Furthermore a recent report by Xinget al [33] suggests that ephrin-B1 reverse signalling is important forbone formation [33] with in vivo and in vitro experiments showing thatephrin-B1 osteogenesis is mediated by the activation of the ephrin-B1PDZ domain Dephosphorylation of PDZ bindingmotif TAZ results in itsnuclear translocation and subsequent activation of Osterix an osteo-blast-specific transcription factor [33]
Table 1RT-PCR primers generated for human specific sequences
Gene Accession Forwar
β-actin NM_001101 GATCAEphB1 AF037331 GTGGCEphB2 AF025304 ATGAAEphB3 NM_004443 CACTGEphB4 NM_004444 GCCGCEphB6 NM_004445 TGGGAephrin-B1 NM_004429 AGCTCephrin-B2 NM_004093 CCTCTCephrin-B3 NM_001406 AGGTG
Collectively these studies implicate a role of Ephephrinmoleculesin postnatal haematopoietic and skeletal tissue homeostasis Howev-er the role of Ephephrin family members has not been addressed inhuman MSC based systems of migration and osteogenic differentia-tion In this study the expression of EphBephrin-B molecules wasexamined in cultured human MSC Functional studies were alsoperformed to determine the potential role of EphBephrin-B interac-tions in human MSC attachment migration and osteochondraldifferentiation
Materials and methods
Cell culture
STRO-1 immunoselected MSC [35] were prepared from bonemarrow mononuclear cells isolated from normal human BM aspiratesas previously described [35ndash37] Primary cultures were established byculturing 5times104cm2 STROminus1+MNC in alpha Modification of EaglesMedium(αMEM) supplementedwith 20 (vv) fetal calf serum 2 mML-glutamine 1 mM sodium pyruvate 100 μM L-ascorbate-2-phosphateand penicillin (50 iumL)streptomycin sulphate (50 μgmL) at 37 degChumidified atmosphere in the presence of 5 CO2 Upon achieving 90confluence adherent cells were liberated by trypsinEDTA treatmentcounted and then re-plated at 8times103cm2 in αMEM growth mediumsupplemented with 10 FCS
Real-time polymerase chain reaction (RT-PCR)
Total RNA was extracted from human culture expanded MSC togenerate cDNA as previously described [20] Two micrograms of RNAwas reverse transcribed with superscript III (Invitrogen MulgraveVictoria Australia) Real-time PCR reactions were performed usingSyberGreen (SABiosciences Fredrick MD USA) and Rotorgene 3000series (Corbett Life Sciences Sydney NSW Australia) Primer sets(Table 1) were used at a final concentration of 100 ngμL and reactionsfor each sample and were performed in triplicate
Immunocytochemistry
Human bone marrow trephine samples were fixed in 10 bufferedformalin for 48 h at room temperature then decalcified in 10 EDTApH 80 and 005 PFA at 4 degC prior to paraffin embeddingDeparaffinised 5 μm sections were blocked for 30 min at roomtemperature with 3 normal horse serum and either 3 normalrabbit or goat serum depending on the secondary antibody Sectionswere stained with either 1ndash2 μgL goat IgG1 anti-EphB1 EphB2ephrin-B3 or rabbit IgG anti-EphB4 ephrin-B1 ephrin-B2 and thecorresponding isotype negative controls rabbit IgG or Goat IgG incorresponding blocking buffer overnight at 4 degC Samples werewashed and incubated with either rabbit anti-goat biotin (VectorLaboratory) or goat anti-rabbit biotin (Vector Laboratory) in
d 5prime-3prime Reverse 5prime-3prime
TTGCTCCTCCTGAGC GTCATAGTCCGCCTAGAAGCTACGATGAAAACCTGAAC CTGGTTGGGCTCGAAGACATCACGATCCGCACGTA TTGGTCCGTAGCCAGTTGTTCTAGTTCATGGAAAACTGC GTAGGATCGGAGGGGTCATCAGCTTTGGAAGAG CATCCAGGCCGCTCAGTTCGAGGTGAGTGTTCTG CCACATGACATGCCTCAAAGGCCTCAACCCCAAGTT GGCAGATGATGTCCAGCTTGTCTCAACTGTGCCAAA CCCAGAGGTTAGGGCTGAATTCTTCTCCGAGTGG GGAGGTTGCATTGCTGGT
535A Arthur et al Bone 48 (2011) 533ndash542
corresponding blocking buffer for two hours at room temperatureSections were washed with PBS and incubated with Streptavidin-HRP(Pierce Thermo Scientific) for one hour at room temperature in thecorresponding blocking buffer Samples were washed again andvisualised with DAB substrate (DAKO Glostrup Denmark) Sectionswere counterstained with haematoxylin and mounted with DePix(BDH MERCK Whitehouse Station NJ USA)
Western blot
Cell lysates were prepared as previously described [38] andequivalent amounts of protein (40 μg) separated on 8 or 10 SDSPAGE gels and transferred to PVDF membranes Immunoblotting wasperformed as previously described [38] with the following antibodiesgoat anti-EphB1 (Santa Cruz) EphB2 EphB3 EphB4 EphB6 ephrin-B1ephrin-B2 and ephrin-B3 (RampD Systems Minneapolis MN USA)Following incubationwith either anti-goat Ig (Millipore) or anti-mouseIgG (Chemicon Melbourne Victoria Australia) alkaline phosphataseconjugated secondary antibodies (Millipore Billerica USA) mem-branes were developed with enhanced chemifluorescence substrate(GE Healthcare Little Chalfont USA) Membranes were washed andthen incubated in stripping buffer (Western Blot Recycling Kit AlphaDiagnostic Intl Inc) for 15 min atRT beforebeingused to re-probewithmouse anti-human β-actin (Sigma) Quantitative analysis was per-formed using ImageQuant software (GE Healthcare)
Spreading assay
The spreading assay was conducted as previously described [20]Briefly Fc-fusion proteins (Human-IgG-Fc 1 mgmL EphB-Fc andephrin-B-Fc 200 μgmL RampD Systems) were pre-clustered with a 10-fold concentration of Human anti-Goat IgG (24 mgmL) in PBS for 1 hat room temperature Ninety-six well flat-bottom plates were coatedwith Poly-L-lysine (001 solution Sigma) for 5 min at roomtemperature Following the pre-clustering of the Fc the sampleswere incubated in coated wells for 2 h at 37 degC in a 5 CO2 incubatorthen washed with PBS Cells were liberated using PUCKS-EDTA (5 mMKCl 130 mMNaCl 3 mMNaHCO3 5 mM D-glucose 10 mMHEPES (pH73) and 1 mMEDTA) and resuspended in growthmedia 8times103cellswell was incubated for 3 h at 37 degC in 5 CO2 For the inhibitor assaycells were incubated for 30 min at room temperature with signallinginhibitor PP2 (CalBiochem San Diago CA USA) LY294002 (Sigma)SP600125 (Sigma) or 01 DMSO prior to being added to the well Thecultures were then fixed and stained with phalloidin-TRITC Forconsistency images of the cultures were taken in the centre of eachwell
Migration assay
The migration assay was conducted as previously described [20]Briefly pre-clustered Fc-fusion proteins (5 μgmL) were added to thebottom chambers of transwells Cultured MSC were liberated usingPUCKS-EDTA washed and resuspended in growthmedia then seededat 3times104cellswell into the top chamber of each transwell (CorningCorning NY USA) for 24 h at 37 degC in 5 CO2 For the inhibitor assaycells were incubated for 30 min at room temperature with signallinginhibitor U0126 (Cell Signalling Technology Danvers MA USA) PP2SB203580 (CalBiochem) LY Imatinib (Kindly provided by NovartisBasel Switzerland) 01 DMSO or pre-incubated with 100 μMblocking peptides specific for EphB1 (EWLSPNLAPSVR) EphB2(SNEWILPRLPQH) EphB4 (TNYLFSPNGPIARAW) or control peptide(RTVAHHGGLYHTNAEVK) (Mimotopes Clayton Victoria Australia)for one hour on ice prior to being added to the transwell The cultureswere then fixed and the inside of the transwell was cleared beforestaining with DAPI The assay was repeated in duplicate wells andimages were taken at 3 random positions within the well for each
well The number of nuclei were counted in each image andrepresented as the average cell countfield of view
Mineralisation assay
Single cell suspensions were seeded at 8000 cellscm2 in osteo-genic-induction media [37] in the presence of either pre-clusteredEphB2-Fc EphB4-Fc human-Fc (5 μgmL) or EphB blocking peptides(50 μM) After 4 weeks the mineralisedmatrix was dissolved in 06 MHCl and calcium levels quantitated by the Cresolphthalein Complex-one assay (Thermo Electron Corporation Melbourne Victoria USA)Calcium levels were normalised to cell number by quantitation of DNAcontent per well using Hoescht as described previously [39] Inaddition mineralised matrix formation was visualised by Alizarin RedStaining as previously described [39]
Glycoasminoglycan synthesis assay
Cultured MSC were liberated using PUCKS-EDTA and seeded as asingle cell suspension in duplicate 96-well plate at 5times104cellswell inchondrogenic media [40] containing pre-clustered humanEphBephrin-B-Fc (5 μgmL) or EphB blocking peptides (50 μM) and10 ngmL TGF-β1 at 37 degC 5 CO2 After 48 h the media was replacedwith a final concentration of 1 μCi100 μLwell Sulfur-35 radionuclide(35SO4
2minus Perkin Elmer Boston MA USA) The cells were thenincubated 37 degC 5 CO2 for a further 16 h Plates were treated withPapain digestion solution (Papain (20 UmL) NaAc (01 M) EDTA(24 mM) and N-Acetyl-L-Cysteine (20 mM) in water) at 65 degC for 3 hfollowed by Cetyl pyridinyl chloride (CPC) solution (25 CPC 40Ethanol in water) and Chondroitin sulphate A (CSA) solution (CSASigma 1 mgmL final) for a further 5 min Digested samples wereharvested onto glass filter membranes (Packard CT USA) using a cellharvester (Filtermate Packard) and then analysed using thescintillation counter (Top Count NXT Perkin Elmer Boston MAUSA) Top Count NTX program (v 253) GAG synthesis was quantifiedrelative to DNA concentration using replicate plates
shRNA knockdown
RNA duplexes targeting human EphB2 and ephrin-B2 were clonedinto the pFIV-H1-copGFP lentiviral vector as per manufacturersinstructions (System Biosciences Karrinyup WA Australia) TheshRNA oligonucleotide sequences used in this study were EphB2shRNA_1 (gatccggtgtgcaacgtgtttgagttcaagagactcaaacacgttgca-caccttttttgctagcg) EphB2 shRNA_2 (gatccgctgtgatttccagtgtcaatttcaa-gagaattgacactggaaatcacagcttttttgctagcg) ephrin-B2 shRNA_1 and(gatccccaggaataaagatccaacaattcaagagattgttggatctttattcctggttttttgc-tagcg) shRNA scramble nonsilencing control (gatccagttaatatcaa-gacgctgttcaagagacagcgtcttgatattaactttttttgctagcg) Viral supernatantcollected from transfected HEK-293T cells was used to infect primaryMSC from the top 10 of GFP-expressing cells using FACS aspreviously described [41] Cell lines with greater than 60 reductionin EphB2 and ephrin-B2 gene expressions compared to correspondingshRNA scramble MSC lines were used for the functional studies
Statistics
Treated groups were compared with their corresponding controlusing the paired Students t-tests Differences were considered to bestatistically significant when the P value was less than 005 Multipletreated groups were compared with the control group using one-wayANOVA where significance (P valueb005) was determined post hocusing the Dunnett test
536 A Arthur et al Bone 48 (2011) 533ndash542
Results
Characterisation of EphBephrin-B molecules expressed by human MSC
Gene expression analyses using real time PCR (Fig 1A) confirmedthat the B-subclass Ephephrin molecules were expressed by cultureexpanded human MSC At the protein level human MSC demonstrat-ed similar levels of ephrin-B1 (003plusmn0001 SD n=2 MSC donors)ephrin-B2 (003plusmn0002 SD n=2 MSC donors) and ephrin-B3(003plusmn0004 SD n=2 MSC donors) when compared to thecorresponding β-actin controls using ImageQuant software HigherEphB4 protein expression (393plusmn0075 SD n=2 MSC donors)correlated to high gene expression in contrast to EphB1 (048plusmn0216 SD n=2 MSC donors) EphB2 (012plusmn0055 SD n=2 MSCdonors) EphB3 (026plusmn0141 SD n=2 MSC donors) and EphB6(010plusmn0063 SD n=2 MSC donors) when compared to thecorresponding β-actin controls Immunohistochemical staining ofnormal human trephine biopsies obtained from MGUS patients (nopresence of lytic disease and less than 10 plasma cells in the bonemarrow) confirmed the expression of EphB and ephrin-B (Fig 2)molecules in vivo Consistent with the in vitro characterization EphB2EphB4 and ephrin-B1 expressions appeared most pronounced withinthe human bone localising to bone lining cells osteoblasts and cellswithin the marrow where ephrin-B1 was also found to be expressedby osteocytes The protein expression of ephrin-B2 was less intenselocalising to bone lining cells and osteoclasts (Fig 2) These findings
Fig 1 Expression of B-subclass Eph receptors and ephrin ligands by MSC (A) Real-time PCEphB4 and ephrin-B2 are most highly expressed by cultured MSC The histograms represenMSC-3 (B) The data depict representative Western blot analyses of different EphB and efollowing stripping and re-probing of each membrane
were inconsistent with the in vitro observations where ephrin-B3 wasexpressed by a large number of bone marrow cells (Fig 2)Furthermore these experiments demonstrated that EphB1 B2 B4and ephrin-B1 -B2 and -B3 molecules were expressed at varyingintensities around blood vessels similar to that observed for theperivascular and MSC associated marker CD146 (Fig 2) [43742]
EphBephrin-B interaction inhibit spreading via reverse signalling whilepromoting migration by forward signalling
Studies were conducted to determine the effect of forward andreverse signalling on the attachment spreading and migration ofMSC in vitro using an established spreading assay [20] Culturedhuman MCS exposed to EphB2-Fc or EphB4-Fc (5 μgmL) weresignificantly rounder (pb0001 ANOVA Dunnett test n=3 marrowdonors) and smaller (pb0001 ANOVA Dunnett test n=3 marrowdonors) as demonstrated with F-actin staining compared to controlhuman-Fc (Figs 3AndashC) Multiple signalling pathways were activat-ed to promote the contraction and rounding up ofMSC Incubation ofMSC with signalling pathway-specific inhibitors prior to performingthe spreading assay revealed that both the Src kinase pathway (PP2)and the PI3Kinase pathway (LY294002)were activated upon ephrin-B reverse signalling in response to EphB2-Fc while the PI3Kinaseand JNK pathways were predominantly activated in response toEphB4-Fc (Fig 3D) No difference inmorphologywas observedwhenMSC were cultured in the presence of ephrin-B1-Fc or ephrin-B2-Fc
R data were expressed as the gene transcript relative to β-actin indicates that EphB2t the meanplusmnSEM from 3 independent experiments for each donor MSC-1 MSC-2 andphrin-B proteins expressed by cultured MSC and the corresponding β-actin controls
Fig 2 Expression of B-subclass Eph receptors and ephrin ligands in human bone marrow trephines Paraffin-embedded sections of human trephine were stained with antibodiesreactive to human EphB receptors (brown) EphB1 EphB2 and EphB4 and ephrin-B ligands (brown) ephrin-B1 ephrin-B2 ephrin-B3 and counterstained with haematoxylin (blue)EphB receptors and ephrin-B ligands localised to osteoblasts (arrowheads) osteocytes (asterisks) multinucleated cells and perivascular cells (CD146 staining arrow) asdemonstrated with serial sections stained with perivascular marker CD146 (brown) Scale bar=50 μm
537A Arthur et al Bone 48 (2011) 533ndash542
(Figs 3AndashC) Supplementary Fig 1 represents a supportive studydemonstrating that MSC spreading and attachment are mediated byreverse signalling In these experiments shRNA ephrin-B2 knock-down MSC failed to undergo rounding in the presence of theephrinB2 high affinity receptor partner EphB4-Fc when comparedto the shRNA scramble nonsilencing control MSC However round-ing was still observed with both the ephrin-B2 knockdown andscramble MSC lines in the presence the ephrinB1 high affinitybinding partner EphB2-Fc demonstrating the specificity of EphB4for ephrin-B2
Migration assays using transwells were performed to examineMSC migration in vitro as previously described [20] Clustered solubleEphB2 EphB4 ephrin-B1 or ephrin-B2-Fc were added to the bottomchambers and the MSC were seeded into the top chambers of thetranswells Those cells that migrated through the transwell poreswithin 24 h were fixed and stained with DAPI before imaging Theresults indicated that human MSC migration was unaffected byephrin-B reverse signalling in the presence of EphB2-Fc or EphB4-Fc
(5 μgmL) when compared to the human-Fc (Fig 4A) However MSCmigration was significantly increased (pb001 ANOVA Dunnett testn=3 marrow donors) in the presence of ephrin-B1-Fc (5 μgmL)(Fig 4A) which appeared to be partially but significantly reversed(pb0001 ANOVA Dunnett test n=3 marrow donors) in thepresence of either the Src kinase inhibitor PP2 or the Abl kinaseinhibitor Imatinib (Fig 4B) Furthermore EphB2 forward signallingwas found to mediate migration where a shRNA EphB2 knockdownMSC line exhibited a defective migratory response to ephrin-B1-Fc incontrast to the shRNA scramble nonsilencing control MSC line(Fig 4C)
EphBephrin-B interactions influence MSC osteogenic and chondrogenicdifferentiations
To decipher the role of EphBephrin-B molecules in osteogenesishuman MSC were cultured in osteogenic inductive media in thepresence of EphB-Fc or ephrin-B-Fc molecules (Figs 5A and C)
Fig 3 Reverse ephrin-B signalling inhibits human MSC attachment and spreading (A) Representative images of human MSC cultured with 5 μgmL EphB2-Fc EphB4-Fc ephrin-B1-Fc or ephrin-B2-Fc attached to the surface of the well (BndashC) MSC were significantly rounder (pb0001) (B) and smaller (pb0001) (C) when cultured with EphB2-Fc or EphB4-Fcrespectively but not with ephrin-B1-Fc or ephrin-B2-Fc compared to the control human-Fc Data are meanplusmnSEM from 3 independent marrow donors per treatment (D) DMSOcontrol EphB2-Fc or EphB4-Fc mediated inhibition of MSC attachment was reversed in the presence of Src kinase inhibitor (25 μM PP2) PI3K inhibitor (10 μM LY294002) or JNKinhibitor (30 μM SP600125) (E) Graphs for inhibitor spreading analysis Scale bar=50 μm All data present the meanplusmnSEM from 3 independent bone marrow donors analysed byone-way ANOVA Dunnett test
538 A Arthur et al Bone 48 (2011) 533ndash542
Following 4 weeks of culture MSC produced significantly (pb001ANOVA Dunnett test n=4 bone marrow donors) more mineral inthe presence of EphB2-Fc when compared to the control human-Fcas demonstrated by Alizarin Red staining and calcium release fromthe extracellular matrix (Figs 5A and C) Furthermore MSC culturedin osteogenic media were significantly inhibited (pb001 ANOVADunnett test n=4 marrow donors) from forming mineral whencultured with blocking peptides for EphB1 (EWLS) and EphB4(TNYL) when compared to the control peptide (RTVA) (Figs 5Band D)
In parallel studies the role of Eph and ephrin molecules in theearly stages of chondrogenesis based on glycosaminoglycan (GAG)synthesis was also examined Consistent with the osteogenicdifferentiation significantly (pb005 ANOVA Dunnett test n=3marrow donors)more GAGwas produced in the presence of EphB2-Fcwhen compared to human-Fc (5 μgmL) following high densityculture in the presence of TGFβ-1 (Fig 5E) Interestingly whilereverse signalling enhanced GAG production blocking the EphB2receptor significantly inhibited (pb001 ANOVA Dunnett test n=3marrow donors) GAG synthesis (Fig 5F)
Fig 4 Forward EphB signalling promotes MSC migration (A) Transwell migration assays demonstrated that human MSC significantly increased (+pb001 ANOVA Dunnett test)their migration in the presence of 5 μgmL ephrin-B1-Fc compared to control human-Fc (B) ephrin-B1-Fc mediated MSC migration (DMSO control) was significantly reduced(plusmnpb0001) in the presence of PP2 (10 μM) and (^pb001 ANOVA Dunnett test) the presence of Imatinib (3 μM) in contrast to MEKERK inhibitor (U0126) p38 MAPK inhibitorSB203580 (SB) and LY294002 (LY) (C) MSCmigration mediated by ephrin-B1 activation was significantly inhibited in a shRNA EphB2 knockdownMSC line in either the presence ofeither 5 μgmL human-Fc (p=00014 paired Student t-test) or ephrin-B2-Fc (p=000037 Student t-test) when compared to the shRNA scramble nonsilencing control MSC Alldata represent the meanplusmnSEM from 3 independent bone marrow donors Images represent nuclei stained with DAPI Image J software was used to count nuclei in five replicatefields per chamber Scale bar=50 μm
539A Arthur et al Bone 48 (2011) 533ndash542
Discussion
In the present study we demonstrated the functional significanceof Ephephrin-B molecules expressed by MSC in relation to cellularattachment migration and osteochondral differentiation Cell motilityrelies on the convergence of a number of molecular processesincluding the establishment of focal adhesions followed by theattachment of F-actin among other associated molecules This allowsthe extension of actin processes in the form of filopodia andlamellopodia which serve to lengthen the cell [4344] These eventscan be activated by a number of ECM chemotaxis and guidancemolecules The Eph family is well known for inhibiting cell motilityparticularly during the process of boundary formation of neural crestcells restricting veins and arteries and assisting in the maintenance ofneural intestinal and dental pulp stem cell niches [172145ndash49] Inthis study we showed that activated ephrin-B reverse signalling actsto inhibit human MSC attachment and spreading by potentially
restricting focal adhesion maturation as previously shown for dentalpulp stem cells [20] The EphBephrin-B interactions trigger a numberof signalling pathways to mediate functions such as cell adhesionmigration proliferation or differentiation We demonstrated thatephrin-B induced inhibition of attachment and spreading wasmediated in part by signalling through the Src PI3Kinase and theJNK pathways This was somewhat surprising since ephrin-B reversesignalling is thought to be mediated predominantly through the Srcmolecule Grb4 or the PDZ domain However it has been demon-strated that ephrin-B1 can also signal through the JNK pathwayindependently of tyrosine kinase phosphorylation [5051] Converse-ly EphB2 forward signalling stimulated via interactions with ephrin-B1 promoted MSC migration mediated through the Src and Abl-Crksignalling pathways Therefore EphBephrin-B interactions may playa role in the recruitment of MSC from their perivascular niche towardsareas of tissue damage via the activation of either the EphB receptorsor their respective ephrin-B ligands [43742]
Fig 5 Reverse signalling promotes osteochondral differentiation (AndashD) Human MSC were cultured in osteogenic conditions for four weeks in the presence of 5 μgmL EphB2-FcEphB4-Fc ephrin-B1-Fc ephrin-B2-Fc or control human-Fc (A C) MSC cultured in the presence of EphB2-Fc produced significantly more mineral (^pb001) as demonstrated withAlizarin Red staining and (C) calcium concentration when compared to human-Fc (B D) Human MSC mineral production as demonstrated by (B) Alizarin Red staining and(D) calcium concentration was significantly inhibited in the presence of either EWLS (p=001) or TNYL (^p=001) EphB1 and EphB4 blocking peptides respectively compared tocontrol peptide RTVA (E F) HumanMSC cultured in the presence of chondrogenic media and EphB and ephrin-B-Fc molecules demonstrated significantly more GAG synthesis whencultured in the presence of (E) EphB2-Fc (pb005) when compared to the human-Fc control and significantly less GAG production when cultured with EphB2 blocking peptideSNEW (pb001) compared to peptide control RTVA All data represent the meanplusmnSEM for 3ndash4 independent marrow donors analysed by one-way ANOVA Dunnett test
540 A Arthur et al Bone 48 (2011) 533ndash542
Previously published mouse models have reported that EphBforward signalling was important for mouse osteoblastic differenti-ation [910] Similarly we observed that mineralisation of humanMSCwas decreased in the presence of the blocking peptides for EphB1 andEphB4 However our observations also suggest that reverse signallingstimulated by EphB2-Fc appears to significantly increase mineralproduction of MSC cultured in osteo-inductive media While ephrin-B-Fc molecules failed to stimulate EphB forward induced osteogen-esis addition of blocking peptides to EphB receptors inhibitedmineralproduction indicating that these blocking peptides may function notonly to block EphB forward signalling [5253] but may also inhibitreverse signalling Importantly the crystallographic studies havedemonstrated that the GndashH loop of the ephrin-B ligand physicallybinds to the ligand-binding cavity of the EphB receptor whichcoincides with the DndashE GndashH and JndashK loops resulting in eitherstabilised activation of EphB receptor or the ephrin-B ligand [54ndash57]Therefore it is plausible that by specifically blocking the ephrin-Bbinding cavity of EphB receptors these endogenous receptors may nolonger activate their cognate ephrin-B ligands While we cannotspecifically state which ligands are being blocked it could bepostulated based on binding affinity studies [58] that ephrin-B2may be predominantly required for osteoblastic differentiationHowever we cannot rule out the importance of ephrin-B1 based onthe transwells migration assays which showed that ephrin-B1activated EphB4 forward signalling Furthermore the findings ofephrin-B1 transgenic studies confirmed that reverse signallingmodulates the osteogenic response of bone marrow stromal cellsspecifically mediated through ephrin-B1 activation following stimu-lation with EphB2-Fc [33] The signalling mechanism required for theincrease in osteogenesis was mediated via PDZ signalling It appearsthat the formation of a PDZ complex containing NHERF1 PTPN13 andTAZ with ephrin-B1 results in the dephosphorylation of TAZ which
allows its translocation into the nucleus and subsequent activation ofosteogenic specific transcription factor Osterix [33]
Of note Zhao and colleagues also demonstrated high geneexpression of ephrin-B1 and ephrin-B2 by osteoblasts which werealso confirmed by Allan et al [910] However the functionalrelevance of this expression was not investigated in detail Further-more while Zhao and colleagues did use EphB4-Fc in their osteogenicassays these experiments did not demonstrate a significant differencein mineral formation [9] This finding is consistent with ourobservations as MSC produced more mineral only in response toEphB2-Fc As ephrin-B1 binds with highest affinity to EphB2 [58] itmay be assumed that MSC osteogenesis may be mediated by theEphB2ephrin-B1 receptor-ligand pairing however our blockingpeptide studies suggested that perhaps it may be mediated byephrin-B2 Our data show that EphB1 and EphB4 but not EphB2blocking peptides significantly reduce mineral formation in humanMSC where ephrin-B2 binds with highest affinity to EphB1 and EphB4[58] This observation is in line with the findings of Allan et al whoshowed that mouse Kusa 4b10 cells reduced mineral formation in thepresence of EphB4 blocking peptide TNYL at consistent concentra-tions used in the present study [10]
Alternatively the variation in EphBephrin-B signalling activatingMSC osteogenesis in this study may be attributed to the origin of theMSC used for each study The studies performed by Zhao et al andAllan et al utilized osteogenic cells derived from calvarial bone whichis formed by intramembranous ossification while the long bonesundergo predominantly endochondral ossification [5960] To accountfor these discrepancies we performed comparative studies usingbone marrow derived mouse MSC isolated from C57BL6 mousefemurs and found that these cells showed similar responses to theirhuman counterparts (unpublished observations) Furthermore theephrin-B1 transgenic studies also utilised stromal cells from the
541A Arthur et al Bone 48 (2011) 533ndash542
marrow rather than the calvarial bone identified similar findings tothe present study [33]
Studies were also performed to investigate the contribution of theEphephrin-B family in the early stages of chondrogenesis Similar tothe osteo-inductive assays we demonstrated that ephrin-B reversesignalling mediated by EphB2-Fc stimulated an increase in GAGproduction which could be inhibited by blocking the function ofendogenous EphB2 Again this observation would suggest that byblocking endogenous EphB2 ligand-binding and function may also beinhibited However unlike the mineralisation assay these findingssuggest that perhaps ephrin-B1 the highest binding affinity cognateligand for EphB2 [58] may be important for endochondral formationThis is a novel observation that the B-subclass Ephephrins may beinvolved in endochondral ossification following fracture and byhuman MSC To date the only studies to our knowledge investigatingthe B-subclass demonstrate that osteoarthritic subchondral bonemetabolism involves EphB4ephrin-B2 signalling The activation ofEphB4 by ephrin-B2 inhibits the expression of factors such as matrixmetalloproteinases interleukins and RANKL which result in reducedresorption activity leading to osteoarthritic subchondral bone [6162]Our study establishes the potential role of the EphBephrin-Bsignalling in human MSC attachmentspreading cell migration andchondrogenic and osteogenic differentiations
Supplementarymaterials related to this article can be found onlineat doi101016jbone201010180
Acknowledgments
This study was supported by a Robinson Institute Research Grantin Aid and NHMRC Project Grant 565176
References
[1] Sacchetti B Funari A Michienzi S Di Cesare S Piersanti S Saggio I et al Self-renewing osteoprogenitors in bone marrow sinusoids can organize a hematopoi-etic microenvironment Cell 2007131324ndash36
[2] Shi S Gronthos S Perivascular niche of postnatal mesenchymal stem cells inhuman bone marrow and dental pulp J Bone Miner Res 200318696ndash704
[3] Mankani MH Kuznetsov SA Robey PG Formation of hematopoietic territories andbone by transplanted human bone marrow stromal cells requires a critical celldensity Exp Hematol 200735995ndash1004
[4] Arthur A Zannettino A Gronthos S The therapeutic applications of multipotentialmesenchymalstromal stem cells in skeletal tissue repair J Cell Physiol 2008218237ndash45
[5] Komano Y Nanki T Hayashida K Taniguchi K Miyasaka N Identification of ahuman peripheral blood monocyte subset that differentiates into osteoclastsArthritis Res Ther 20068R152
[6] Karsenty G Role of Cbfa1 in osteoblast differentiation and function Semin Cell DevBiol 200011343ndash6
[7] Karsenty G Ducy P Starbuck M Priemel M Shen J Geoffroy V et al Cbfa1 as aregulator of osteoblast differentiation and function Bone 199925107ndash8
[8] Olsen BR Reginato AM Wang W Bone development Annu Rev Cell Dev Biol200016191ndash220
[9] Zhao C Irie N Takada Y Shimoda K Miyamoto T Nishiwaki T et al BidirectionalephrinB2-EphB4 signaling controls bone homeostasis Cell Metab 20064111ndash21
[10] Allan EH Hausler KD Wei T Gooi JH Quinn JM Crimeen-Irwin B et al EphrinB2regulation by PTH and PTHrP revealed by molecular profiling in differentiatingosteoblasts J Bone Miner Res 2008231170ndash81
[11] Irie N Takada Y Watanabe Y Matsuzaki Y Naruse C Asano M et al Bidirectionalsignaling through ephrinA2-EphA2 enhances osteoclastogenesis and suppressesosteoblastogenesis J Biol Chem 200928414637ndash44
[12] Cooke JE Moens CB Boundary formation in the hindbrain Eph only it weresimple Trends Neurosci 200225260ndash7
[13] Mellitzer G Xu Q Wilkinson DG Eph receptors and ephrins restrict cellintermingling and communication Nature 199940077ndash81
[14] Jayasena CS Flood WD Koblar SA High EphA3 expressing ophthalmic trigeminalsensory axons are sensitive to ephrin-A5-Fc implications for lobe specific axonguidance Neuroscience 200513597ndash109
[15] Koblar SA Krull CE Pasquale EB McLennan R Peale FD Cerretti DP et al Spinalmotor axons and neural crest cells use different molecular guides for segmentalmigration through the rostral half-somite J Neurobiol 200042437ndash47
[16] Chumley MJ Catchpole T Silvany RE Kernie SG Henkemeyer M EphB receptorsregulate stemprogenitor cell proliferation migration and polarity duringhippocampal neurogenesis J Neurosci 20072713481ndash90
[17] Merrill AE Bochukova EG Brugger SM Ishii M Pilz DTWall SA et al Cell mixing ata neural crest-mesoderm boundary and deficient ephrin-Eph signaling in thepathogenesis of craniosynostosis Hum Mol Genet 2006151319ndash28
[18] Sakai D Wakamatsu Y Regulatory mechanisms for neural crest formation CellsTissues Organs 200517924ndash35
[19] Okubo T Yanai N Obinata M Stromal cells modulate ephrinB2 expression andtransmigration of hematopoietic cells Exp Hematol 200634330ndash8
[20] Stokowski A Shi S Sun T Bartold PM Koblar SA Gronthos S EphBephrin-Binteraction mediates adult stem cell attachment spreading and migrationimplications for dental tissue repair Stem Cells 200725156ndash64
[21] Arthur A Koblar S Shi S Gronthos S EphephrinB mediate dental pulp stem cellmobilization and function J Dent Res 200988829ndash34
[22] Merlos-Suarez A Batlle E Eph-ephrin signalling in adult tissues and cancer CurrOpin Cell Biol 200820194ndash200
[23] Moore R Larue L Cell surface molecules and truncal neural crest ontogeny aperspective Birth Defects Res C Embryo Today 200472140ndash50
[24] Picco V Hudson C Yasuo H EphrinndashEph signalling drives the asymmetric division ofnotochordneural precursors in Ciona embryos Development 20071341491ndash7
[25] Sancho E Batlle E Clevers H Live and let die in the intestinal epithelium Curr OpinCell Biol 200315763ndash70
[26] Wang Z Cohen K Shao Y Mole P Dombkowski D Scadden DT Ephrin receptorEphB4 regulates ES cell differentiation of primitive mammalian hemangioblastsblood cardiomyocytes and blood vessels Blood 2004103100ndash9
[27] Compagni A Logan M Klein R Adams RH Control of skeletal patterning byephrinB1-EphB interactions Dev Cell 20035217ndash30
[28] Davy A Aubin J Soriano P Ephrin-B1 forward and reverse signaling are requiredduring mouse development Genes Dev 200418572ndash83
[29] Wieland I Jakubiczka S Muschke P CohenM Thiele H Gerlach KL et al Mutationsof the ephrin-B1 gene cause craniofrontonasal syndrome Am J Hum Genet2004741209ndash15
[30] Ting MC Wu NL Roybal PG Sun J Liu L Yen Y et al EphA4 as an effector of Twist1in the guidance of osteogenic precursor cells during calvarial bone growth and incraniosynostosis Development 2009136855ndash64
[31] Varelias A Koblar SA Cowled PA Carter CD Clayer M Human osteosarcomaexpresses specific ephrin profiles implications for tumorigenicity and prognosisCancer 200295862ndash9
[32] Xing W Baylink D Kesavan C Hu Y Kapoor S Chadwick RB et al Global geneexpression analysis in the bones reveals involvement of several novel genes andpathways in mediating an anabolic response of mechanical loading in mice J CellBiochem 2005961049ndash60
[33] Xing W Kim J Wergedal J Chen ST Mohan S Ephrin B1 regulates bone marrowstromal cell differentiation and bone formation by influencing TAZ transactivationvia complex formation with NHERF1 Mol Cell Biol 201030(3)711ndash21
[34] Twigg SR Kan R Babbs C Bochukova EG Robertson SP Wall SA et al Mutations ofephrin-B1 (EFNB1) a marker of tissue boundary formation cause craniofronto-nasal syndrome Proc Natl Acad Sci USA 20041018652ndash7
[35] Gronthos S Simmons PJ The growth factor requirements of STRO-1-positivehuman bone marrow stromal precursors under serum-deprived conditions invitro Blood 199585929ndash40
[36] Pittenger MF Mackay AM Beck SC Jaiswal RK Douglas R Mosca JD et alMultilineage potential of adult human mesenchymal stem cells Science1999284143ndash7
[37] Gronthos S Zannettino AC Hay SJ Shi S Graves SE Kortesidis A et al Molecularand cellular characterisation of highly purified stromal stem cells derived fromhuman bone marrow J Cell Sci 20031161827ndash35
[38] Salvucci O Yao L Villalba S Sajewicz A Pittaluga S Tosato G Regulation ofendothelial cell branching morphogenesis by endogenous chemokine stromal-derived factor-1 Blood 2002992703ndash11
[39] Fitter S Dewar AL Kostakis P To LB Hughes TP Roberts MM et al Long-termimatinib therapy promotes bone formation in CML patients Blood 20081112538ndash47
[40] Isenmann S Arthur A Zannettino AC Turner JL Shi S Glackin CA et al TWISTfamily of basic helix-loop-helix transcription factors mediate human mesenchy-mal stem cell growth and commitment Stem Cells 2009272457ndash68
[41] Diamond P Labrinidis A Martin SK Farrugia AN Gronthos S To LB et al Targeteddisruption of the CXCL12CXCR4 axis inhibits osteolysis in a murine model ofmyeloma-associated bone loss J Bone Miner Res 2009241150ndash61
[42] Zannettino AC Paton S Arthur A Khor F Itescu S Gimble JM et al Multipotentialhuman adipose-derived stromal stem cells exhibit a perivascular phenotype invitro and in vivo J Cell Physiol 2008214413ndash21
[43] DeMali KA Burridge K Coupling membrane protrusion and cell adhesion J Cell Sci20031162389ndash97
[44] de Hoog CL Foster LJ Mann M RNA and RNA binding proteins participate in earlystages of cell spreading through spreading initiation centers Cell 2004117649ndash62
[45] Foo SS Turner CJ Adams S Compagni A Aubyn D Kogata N et al Ephrin-B2controls cell motility and adhesion during blood-vessel-wall assembly Cell2006124161ndash73
[46] Zhang J Hughes S Role of the ephrin and Eph receptor tyrosine kinase families inangiogenesis and development of the cardiovascular system J Pathol 2006208453ndash61
[47] Arthur A Rychkov G Shi S Koblar SA Gronthos S Adult human dental pulp stemcells differentiate toward functionally active neurons under appropriate environ-mental cues Stem Cells 2008261787ndash95
[48] Holmberg J Genander M Halford MM Anneren C Sondell M Chumley MJ et alEphB receptors coordinate migration and proliferation in the intestinal stem cellniche Cell 20061251151ndash63
542 A Arthur et al Bone 48 (2011) 533ndash542
[49] Holmberg J Armulik A Senti KA Edoff K Spalding K Momma S et al Ephrin-A2reverse signaling negatively regulates neural progenitor proliferation andneurogenesis Genes Dev 200519462ndash71
[50] Xu Z Lai KO Zhou HM Lin SC Ip NY Ephrin-B1 reverse signaling activates JNKthrough a novel mechanism that is independent of tyrosine phosphorylation J BiolChem 200327824767ndash75
[51] Huynh-Do U Vindis C Liu H Cerretti DP McGrew JT Enriquez M et al Ephrin-B1transduces signals to activate integrin-mediated migration attachment andangiogenesis J Cell Sci 20021153073ndash81
[52] Koolpe M Burgess R Dail M Pasquale EB EphB receptor-binding peptidesidentified by phage display enable design of an antagonist with ephrin-likeaffinity J Biol Chem 200528017301ndash11
[53] Chrencik JE Brooun A Recht MI Nicola G Davis LK Abagyan R et al Three-dimensional structure of the EphB2 receptor in complex with an antagonis-tic peptide reveals a novel mode of inhibition J Biol Chem 200728236505ndash13
[54] Noren NK Lu M Freeman AL Koolpe M Pasquale EB Interplay between EphB4 ontumor cells and vascular ephrin-B2 regulates tumor growth Proc Natl Acad SciUSA 20041015583ndash8
[55] Himanen JP Chumley MJ LackmannM Li C BartonWA Jeffrey PD et al Repellingclass discrimination ephrin-A5 binds to and activates EphB2 receptor signalingNat Neurosci 20047501ndash9
[56] Himanen JP Rajashankar KR Lackmann M Cowan CA Henkemeyer M NikolovDB Crystal structure of an Eph receptor-ephrin complex Nature 2001414933ndash8
[57] Chrencik JE BroounA RechtMI KrausML KoolpeM Kolatkar AR et al Structure andthermodynamic characterization of the EphB4Ephrin-B2 antagonist peptidecomplex reveals thedeterminants for receptor specificity Structure 200614321ndash30
[58] Pasquale EB The Eph family of receptors Curr Opin Cell Biol 19979608ndash15[59] Opperman LA Cranial sutures as intramembranous bone growth sites Dev Dyn
2000219472ndash85[60] Kronenberg HM Developmental regulation of the growth plate Nature 2003423
332ndash6[61] Kwan Tat S Pelletier JP Amiable N Boileau C Lajeunesse D Duval N et al
Activation of the receptor EphB4 by its specific ligand ephrin B2 in humanosteoarthritic subchondral bone osteoblasts Arthritis Rheum 2008583820ndash30
[62] Kwan Tat S Pelletier JP Amiable N Boileau C Lavigne M Martel-Pelletier JTreatment with ephrin B2 positively impacts the abnormal metabolism of humanosteoarthritic chondrocytes Arthritis Res Ther 200911R119

534 A Arthur et al Bone 48 (2011) 533ndash542
Recent studies suggest that Ephephrin molecules of bothsubclasses regulate bone homeostasis [9ndash11] These membranebound receptor tyrosine kinases predominantly function by inhibitoryor repulsive cellular responses via cellndashcell contact where theresponses can be mediated through the receptor (forward signalling)andor the ligand (reverse signalling) [1213] The involvement of theEph family in cell motility and cell fate decisions is well defined duringaxon guidance [1415] neural progenitor migration [16] and neuralcrest boundary formation [1718] EphBephrin-B signalling alsomodulates hematopoietic cell migration where EphB4-expressinghematopoietic cell migration is repressed by ephrin-B2 expressingstromal cells [19]
We have previously demonstrated that the bi-directional interac-tion between the receptor and ligand restricts human dental pulpstem cells (DPSC) to their perivascular niche under normal physio-logical conditions wheremobilisation of DPSC occurs following injurythrough the down regulation of ephrin-B1 [2021] The Eph family ofreceptor tyrosine kinases have also been implicated in cell fatedecisions [22] of neural crest cells [23] neuronal precursors [24]intestinal epithelial cells [25] hematopoietic lineages [26] skeletaldevelopment [27ndash30] and now in bone remodelling [9ndash1131]
The importance of Ephephrin molecules during bone formation[3233] and bone homeostasis [9] has recently been highlighted by thestudies showing that the osteogenic potential of osteoblasts wasenhanced by forward signalling via EphB4 while the differentiationand function of osteoclasts was inhibited by reverse signalling throughephrin-B2 [9] EphB2 and ephrin-B1were also implicated in this processhowever to a lesser extent Moreover Allan et al [10] have reported anadditional layer of complexity in this process whereby osteoblastexpressionof ephrin-B2was regulatedbyparathyroidhormone receptor1 in a dose dependent manner which in turn may regulate boneformationor osteogenesis in aparacrineor autocrinemanner [10]Whilestudies examining the role of Ephephrinmolecules in bone homeostasishave focused on EphB4 and ephrin-B2 ephrin-B1 expression within thetumour tissue and the surrounding blood vessels has been correlated topoor clinical prognosis for osteosarcoma [31] Moreover completeablation of ephrin-B1 results in perinatal lethality associated with arange of phenotypes including defects in neural crest cell-derivedtissues incomplete body wall closure and abnormal skeletal patterning[28] More specifically ephrin-B1 is important for the correct formationof cranial sutures digit and rib formation and sternebrae fusion [2734]Twigg et al [34] have also demonstrated that unrelated females withcraniofrontonasal syndrome (CFNS) displayed heterozygous ephrin-B1loss of function which was correlated to disrupted boundary formationin developing coronal sutures [34] Furthermore a recent report by Xinget al [33] suggests that ephrin-B1 reverse signalling is important forbone formation [33] with in vivo and in vitro experiments showing thatephrin-B1 osteogenesis is mediated by the activation of the ephrin-B1PDZ domain Dephosphorylation of PDZ bindingmotif TAZ results in itsnuclear translocation and subsequent activation of Osterix an osteo-blast-specific transcription factor [33]
Table 1RT-PCR primers generated for human specific sequences
Gene Accession Forwar
β-actin NM_001101 GATCAEphB1 AF037331 GTGGCEphB2 AF025304 ATGAAEphB3 NM_004443 CACTGEphB4 NM_004444 GCCGCEphB6 NM_004445 TGGGAephrin-B1 NM_004429 AGCTCephrin-B2 NM_004093 CCTCTCephrin-B3 NM_001406 AGGTG
Collectively these studies implicate a role of Ephephrinmoleculesin postnatal haematopoietic and skeletal tissue homeostasis Howev-er the role of Ephephrin family members has not been addressed inhuman MSC based systems of migration and osteogenic differentia-tion In this study the expression of EphBephrin-B molecules wasexamined in cultured human MSC Functional studies were alsoperformed to determine the potential role of EphBephrin-B interac-tions in human MSC attachment migration and osteochondraldifferentiation
Materials and methods
Cell culture
STRO-1 immunoselected MSC [35] were prepared from bonemarrow mononuclear cells isolated from normal human BM aspiratesas previously described [35ndash37] Primary cultures were established byculturing 5times104cm2 STROminus1+MNC in alpha Modification of EaglesMedium(αMEM) supplementedwith 20 (vv) fetal calf serum 2 mML-glutamine 1 mM sodium pyruvate 100 μM L-ascorbate-2-phosphateand penicillin (50 iumL)streptomycin sulphate (50 μgmL) at 37 degChumidified atmosphere in the presence of 5 CO2 Upon achieving 90confluence adherent cells were liberated by trypsinEDTA treatmentcounted and then re-plated at 8times103cm2 in αMEM growth mediumsupplemented with 10 FCS
Real-time polymerase chain reaction (RT-PCR)
Total RNA was extracted from human culture expanded MSC togenerate cDNA as previously described [20] Two micrograms of RNAwas reverse transcribed with superscript III (Invitrogen MulgraveVictoria Australia) Real-time PCR reactions were performed usingSyberGreen (SABiosciences Fredrick MD USA) and Rotorgene 3000series (Corbett Life Sciences Sydney NSW Australia) Primer sets(Table 1) were used at a final concentration of 100 ngμL and reactionsfor each sample and were performed in triplicate
Immunocytochemistry
Human bone marrow trephine samples were fixed in 10 bufferedformalin for 48 h at room temperature then decalcified in 10 EDTApH 80 and 005 PFA at 4 degC prior to paraffin embeddingDeparaffinised 5 μm sections were blocked for 30 min at roomtemperature with 3 normal horse serum and either 3 normalrabbit or goat serum depending on the secondary antibody Sectionswere stained with either 1ndash2 μgL goat IgG1 anti-EphB1 EphB2ephrin-B3 or rabbit IgG anti-EphB4 ephrin-B1 ephrin-B2 and thecorresponding isotype negative controls rabbit IgG or Goat IgG incorresponding blocking buffer overnight at 4 degC Samples werewashed and incubated with either rabbit anti-goat biotin (VectorLaboratory) or goat anti-rabbit biotin (Vector Laboratory) in
d 5prime-3prime Reverse 5prime-3prime
TTGCTCCTCCTGAGC GTCATAGTCCGCCTAGAAGCTACGATGAAAACCTGAAC CTGGTTGGGCTCGAAGACATCACGATCCGCACGTA TTGGTCCGTAGCCAGTTGTTCTAGTTCATGGAAAACTGC GTAGGATCGGAGGGGTCATCAGCTTTGGAAGAG CATCCAGGCCGCTCAGTTCGAGGTGAGTGTTCTG CCACATGACATGCCTCAAAGGCCTCAACCCCAAGTT GGCAGATGATGTCCAGCTTGTCTCAACTGTGCCAAA CCCAGAGGTTAGGGCTGAATTCTTCTCCGAGTGG GGAGGTTGCATTGCTGGT
535A Arthur et al Bone 48 (2011) 533ndash542
corresponding blocking buffer for two hours at room temperatureSections were washed with PBS and incubated with Streptavidin-HRP(Pierce Thermo Scientific) for one hour at room temperature in thecorresponding blocking buffer Samples were washed again andvisualised with DAB substrate (DAKO Glostrup Denmark) Sectionswere counterstained with haematoxylin and mounted with DePix(BDH MERCK Whitehouse Station NJ USA)
Western blot
Cell lysates were prepared as previously described [38] andequivalent amounts of protein (40 μg) separated on 8 or 10 SDSPAGE gels and transferred to PVDF membranes Immunoblotting wasperformed as previously described [38] with the following antibodiesgoat anti-EphB1 (Santa Cruz) EphB2 EphB3 EphB4 EphB6 ephrin-B1ephrin-B2 and ephrin-B3 (RampD Systems Minneapolis MN USA)Following incubationwith either anti-goat Ig (Millipore) or anti-mouseIgG (Chemicon Melbourne Victoria Australia) alkaline phosphataseconjugated secondary antibodies (Millipore Billerica USA) mem-branes were developed with enhanced chemifluorescence substrate(GE Healthcare Little Chalfont USA) Membranes were washed andthen incubated in stripping buffer (Western Blot Recycling Kit AlphaDiagnostic Intl Inc) for 15 min atRT beforebeingused to re-probewithmouse anti-human β-actin (Sigma) Quantitative analysis was per-formed using ImageQuant software (GE Healthcare)
Spreading assay
The spreading assay was conducted as previously described [20]Briefly Fc-fusion proteins (Human-IgG-Fc 1 mgmL EphB-Fc andephrin-B-Fc 200 μgmL RampD Systems) were pre-clustered with a 10-fold concentration of Human anti-Goat IgG (24 mgmL) in PBS for 1 hat room temperature Ninety-six well flat-bottom plates were coatedwith Poly-L-lysine (001 solution Sigma) for 5 min at roomtemperature Following the pre-clustering of the Fc the sampleswere incubated in coated wells for 2 h at 37 degC in a 5 CO2 incubatorthen washed with PBS Cells were liberated using PUCKS-EDTA (5 mMKCl 130 mMNaCl 3 mMNaHCO3 5 mM D-glucose 10 mMHEPES (pH73) and 1 mMEDTA) and resuspended in growthmedia 8times103cellswell was incubated for 3 h at 37 degC in 5 CO2 For the inhibitor assaycells were incubated for 30 min at room temperature with signallinginhibitor PP2 (CalBiochem San Diago CA USA) LY294002 (Sigma)SP600125 (Sigma) or 01 DMSO prior to being added to the well Thecultures were then fixed and stained with phalloidin-TRITC Forconsistency images of the cultures were taken in the centre of eachwell
Migration assay
The migration assay was conducted as previously described [20]Briefly pre-clustered Fc-fusion proteins (5 μgmL) were added to thebottom chambers of transwells Cultured MSC were liberated usingPUCKS-EDTA washed and resuspended in growthmedia then seededat 3times104cellswell into the top chamber of each transwell (CorningCorning NY USA) for 24 h at 37 degC in 5 CO2 For the inhibitor assaycells were incubated for 30 min at room temperature with signallinginhibitor U0126 (Cell Signalling Technology Danvers MA USA) PP2SB203580 (CalBiochem) LY Imatinib (Kindly provided by NovartisBasel Switzerland) 01 DMSO or pre-incubated with 100 μMblocking peptides specific for EphB1 (EWLSPNLAPSVR) EphB2(SNEWILPRLPQH) EphB4 (TNYLFSPNGPIARAW) or control peptide(RTVAHHGGLYHTNAEVK) (Mimotopes Clayton Victoria Australia)for one hour on ice prior to being added to the transwell The cultureswere then fixed and the inside of the transwell was cleared beforestaining with DAPI The assay was repeated in duplicate wells andimages were taken at 3 random positions within the well for each
well The number of nuclei were counted in each image andrepresented as the average cell countfield of view
Mineralisation assay
Single cell suspensions were seeded at 8000 cellscm2 in osteo-genic-induction media [37] in the presence of either pre-clusteredEphB2-Fc EphB4-Fc human-Fc (5 μgmL) or EphB blocking peptides(50 μM) After 4 weeks the mineralisedmatrix was dissolved in 06 MHCl and calcium levels quantitated by the Cresolphthalein Complex-one assay (Thermo Electron Corporation Melbourne Victoria USA)Calcium levels were normalised to cell number by quantitation of DNAcontent per well using Hoescht as described previously [39] Inaddition mineralised matrix formation was visualised by Alizarin RedStaining as previously described [39]
Glycoasminoglycan synthesis assay
Cultured MSC were liberated using PUCKS-EDTA and seeded as asingle cell suspension in duplicate 96-well plate at 5times104cellswell inchondrogenic media [40] containing pre-clustered humanEphBephrin-B-Fc (5 μgmL) or EphB blocking peptides (50 μM) and10 ngmL TGF-β1 at 37 degC 5 CO2 After 48 h the media was replacedwith a final concentration of 1 μCi100 μLwell Sulfur-35 radionuclide(35SO4
2minus Perkin Elmer Boston MA USA) The cells were thenincubated 37 degC 5 CO2 for a further 16 h Plates were treated withPapain digestion solution (Papain (20 UmL) NaAc (01 M) EDTA(24 mM) and N-Acetyl-L-Cysteine (20 mM) in water) at 65 degC for 3 hfollowed by Cetyl pyridinyl chloride (CPC) solution (25 CPC 40Ethanol in water) and Chondroitin sulphate A (CSA) solution (CSASigma 1 mgmL final) for a further 5 min Digested samples wereharvested onto glass filter membranes (Packard CT USA) using a cellharvester (Filtermate Packard) and then analysed using thescintillation counter (Top Count NXT Perkin Elmer Boston MAUSA) Top Count NTX program (v 253) GAG synthesis was quantifiedrelative to DNA concentration using replicate plates
shRNA knockdown
RNA duplexes targeting human EphB2 and ephrin-B2 were clonedinto the pFIV-H1-copGFP lentiviral vector as per manufacturersinstructions (System Biosciences Karrinyup WA Australia) TheshRNA oligonucleotide sequences used in this study were EphB2shRNA_1 (gatccggtgtgcaacgtgtttgagttcaagagactcaaacacgttgca-caccttttttgctagcg) EphB2 shRNA_2 (gatccgctgtgatttccagtgtcaatttcaa-gagaattgacactggaaatcacagcttttttgctagcg) ephrin-B2 shRNA_1 and(gatccccaggaataaagatccaacaattcaagagattgttggatctttattcctggttttttgc-tagcg) shRNA scramble nonsilencing control (gatccagttaatatcaa-gacgctgttcaagagacagcgtcttgatattaactttttttgctagcg) Viral supernatantcollected from transfected HEK-293T cells was used to infect primaryMSC from the top 10 of GFP-expressing cells using FACS aspreviously described [41] Cell lines with greater than 60 reductionin EphB2 and ephrin-B2 gene expressions compared to correspondingshRNA scramble MSC lines were used for the functional studies
Statistics
Treated groups were compared with their corresponding controlusing the paired Students t-tests Differences were considered to bestatistically significant when the P value was less than 005 Multipletreated groups were compared with the control group using one-wayANOVA where significance (P valueb005) was determined post hocusing the Dunnett test
536 A Arthur et al Bone 48 (2011) 533ndash542
Results
Characterisation of EphBephrin-B molecules expressed by human MSC
Gene expression analyses using real time PCR (Fig 1A) confirmedthat the B-subclass Ephephrin molecules were expressed by cultureexpanded human MSC At the protein level human MSC demonstrat-ed similar levels of ephrin-B1 (003plusmn0001 SD n=2 MSC donors)ephrin-B2 (003plusmn0002 SD n=2 MSC donors) and ephrin-B3(003plusmn0004 SD n=2 MSC donors) when compared to thecorresponding β-actin controls using ImageQuant software HigherEphB4 protein expression (393plusmn0075 SD n=2 MSC donors)correlated to high gene expression in contrast to EphB1 (048plusmn0216 SD n=2 MSC donors) EphB2 (012plusmn0055 SD n=2 MSCdonors) EphB3 (026plusmn0141 SD n=2 MSC donors) and EphB6(010plusmn0063 SD n=2 MSC donors) when compared to thecorresponding β-actin controls Immunohistochemical staining ofnormal human trephine biopsies obtained from MGUS patients (nopresence of lytic disease and less than 10 plasma cells in the bonemarrow) confirmed the expression of EphB and ephrin-B (Fig 2)molecules in vivo Consistent with the in vitro characterization EphB2EphB4 and ephrin-B1 expressions appeared most pronounced withinthe human bone localising to bone lining cells osteoblasts and cellswithin the marrow where ephrin-B1 was also found to be expressedby osteocytes The protein expression of ephrin-B2 was less intenselocalising to bone lining cells and osteoclasts (Fig 2) These findings
Fig 1 Expression of B-subclass Eph receptors and ephrin ligands by MSC (A) Real-time PCEphB4 and ephrin-B2 are most highly expressed by cultured MSC The histograms represenMSC-3 (B) The data depict representative Western blot analyses of different EphB and efollowing stripping and re-probing of each membrane
were inconsistent with the in vitro observations where ephrin-B3 wasexpressed by a large number of bone marrow cells (Fig 2)Furthermore these experiments demonstrated that EphB1 B2 B4and ephrin-B1 -B2 and -B3 molecules were expressed at varyingintensities around blood vessels similar to that observed for theperivascular and MSC associated marker CD146 (Fig 2) [43742]
EphBephrin-B interaction inhibit spreading via reverse signalling whilepromoting migration by forward signalling
Studies were conducted to determine the effect of forward andreverse signalling on the attachment spreading and migration ofMSC in vitro using an established spreading assay [20] Culturedhuman MCS exposed to EphB2-Fc or EphB4-Fc (5 μgmL) weresignificantly rounder (pb0001 ANOVA Dunnett test n=3 marrowdonors) and smaller (pb0001 ANOVA Dunnett test n=3 marrowdonors) as demonstrated with F-actin staining compared to controlhuman-Fc (Figs 3AndashC) Multiple signalling pathways were activat-ed to promote the contraction and rounding up ofMSC Incubation ofMSC with signalling pathway-specific inhibitors prior to performingthe spreading assay revealed that both the Src kinase pathway (PP2)and the PI3Kinase pathway (LY294002)were activated upon ephrin-B reverse signalling in response to EphB2-Fc while the PI3Kinaseand JNK pathways were predominantly activated in response toEphB4-Fc (Fig 3D) No difference inmorphologywas observedwhenMSC were cultured in the presence of ephrin-B1-Fc or ephrin-B2-Fc
R data were expressed as the gene transcript relative to β-actin indicates that EphB2t the meanplusmnSEM from 3 independent experiments for each donor MSC-1 MSC-2 andphrin-B proteins expressed by cultured MSC and the corresponding β-actin controls
Fig 2 Expression of B-subclass Eph receptors and ephrin ligands in human bone marrow trephines Paraffin-embedded sections of human trephine were stained with antibodiesreactive to human EphB receptors (brown) EphB1 EphB2 and EphB4 and ephrin-B ligands (brown) ephrin-B1 ephrin-B2 ephrin-B3 and counterstained with haematoxylin (blue)EphB receptors and ephrin-B ligands localised to osteoblasts (arrowheads) osteocytes (asterisks) multinucleated cells and perivascular cells (CD146 staining arrow) asdemonstrated with serial sections stained with perivascular marker CD146 (brown) Scale bar=50 μm
537A Arthur et al Bone 48 (2011) 533ndash542
(Figs 3AndashC) Supplementary Fig 1 represents a supportive studydemonstrating that MSC spreading and attachment are mediated byreverse signalling In these experiments shRNA ephrin-B2 knock-down MSC failed to undergo rounding in the presence of theephrinB2 high affinity receptor partner EphB4-Fc when comparedto the shRNA scramble nonsilencing control MSC However round-ing was still observed with both the ephrin-B2 knockdown andscramble MSC lines in the presence the ephrinB1 high affinitybinding partner EphB2-Fc demonstrating the specificity of EphB4for ephrin-B2
Migration assays using transwells were performed to examineMSC migration in vitro as previously described [20] Clustered solubleEphB2 EphB4 ephrin-B1 or ephrin-B2-Fc were added to the bottomchambers and the MSC were seeded into the top chambers of thetranswells Those cells that migrated through the transwell poreswithin 24 h were fixed and stained with DAPI before imaging Theresults indicated that human MSC migration was unaffected byephrin-B reverse signalling in the presence of EphB2-Fc or EphB4-Fc
(5 μgmL) when compared to the human-Fc (Fig 4A) However MSCmigration was significantly increased (pb001 ANOVA Dunnett testn=3 marrow donors) in the presence of ephrin-B1-Fc (5 μgmL)(Fig 4A) which appeared to be partially but significantly reversed(pb0001 ANOVA Dunnett test n=3 marrow donors) in thepresence of either the Src kinase inhibitor PP2 or the Abl kinaseinhibitor Imatinib (Fig 4B) Furthermore EphB2 forward signallingwas found to mediate migration where a shRNA EphB2 knockdownMSC line exhibited a defective migratory response to ephrin-B1-Fc incontrast to the shRNA scramble nonsilencing control MSC line(Fig 4C)
EphBephrin-B interactions influence MSC osteogenic and chondrogenicdifferentiations
To decipher the role of EphBephrin-B molecules in osteogenesishuman MSC were cultured in osteogenic inductive media in thepresence of EphB-Fc or ephrin-B-Fc molecules (Figs 5A and C)
Fig 3 Reverse ephrin-B signalling inhibits human MSC attachment and spreading (A) Representative images of human MSC cultured with 5 μgmL EphB2-Fc EphB4-Fc ephrin-B1-Fc or ephrin-B2-Fc attached to the surface of the well (BndashC) MSC were significantly rounder (pb0001) (B) and smaller (pb0001) (C) when cultured with EphB2-Fc or EphB4-Fcrespectively but not with ephrin-B1-Fc or ephrin-B2-Fc compared to the control human-Fc Data are meanplusmnSEM from 3 independent marrow donors per treatment (D) DMSOcontrol EphB2-Fc or EphB4-Fc mediated inhibition of MSC attachment was reversed in the presence of Src kinase inhibitor (25 μM PP2) PI3K inhibitor (10 μM LY294002) or JNKinhibitor (30 μM SP600125) (E) Graphs for inhibitor spreading analysis Scale bar=50 μm All data present the meanplusmnSEM from 3 independent bone marrow donors analysed byone-way ANOVA Dunnett test
538 A Arthur et al Bone 48 (2011) 533ndash542
Following 4 weeks of culture MSC produced significantly (pb001ANOVA Dunnett test n=4 bone marrow donors) more mineral inthe presence of EphB2-Fc when compared to the control human-Fcas demonstrated by Alizarin Red staining and calcium release fromthe extracellular matrix (Figs 5A and C) Furthermore MSC culturedin osteogenic media were significantly inhibited (pb001 ANOVADunnett test n=4 marrow donors) from forming mineral whencultured with blocking peptides for EphB1 (EWLS) and EphB4(TNYL) when compared to the control peptide (RTVA) (Figs 5Band D)
In parallel studies the role of Eph and ephrin molecules in theearly stages of chondrogenesis based on glycosaminoglycan (GAG)synthesis was also examined Consistent with the osteogenicdifferentiation significantly (pb005 ANOVA Dunnett test n=3marrow donors)more GAGwas produced in the presence of EphB2-Fcwhen compared to human-Fc (5 μgmL) following high densityculture in the presence of TGFβ-1 (Fig 5E) Interestingly whilereverse signalling enhanced GAG production blocking the EphB2receptor significantly inhibited (pb001 ANOVA Dunnett test n=3marrow donors) GAG synthesis (Fig 5F)
Fig 4 Forward EphB signalling promotes MSC migration (A) Transwell migration assays demonstrated that human MSC significantly increased (+pb001 ANOVA Dunnett test)their migration in the presence of 5 μgmL ephrin-B1-Fc compared to control human-Fc (B) ephrin-B1-Fc mediated MSC migration (DMSO control) was significantly reduced(plusmnpb0001) in the presence of PP2 (10 μM) and (^pb001 ANOVA Dunnett test) the presence of Imatinib (3 μM) in contrast to MEKERK inhibitor (U0126) p38 MAPK inhibitorSB203580 (SB) and LY294002 (LY) (C) MSCmigration mediated by ephrin-B1 activation was significantly inhibited in a shRNA EphB2 knockdownMSC line in either the presence ofeither 5 μgmL human-Fc (p=00014 paired Student t-test) or ephrin-B2-Fc (p=000037 Student t-test) when compared to the shRNA scramble nonsilencing control MSC Alldata represent the meanplusmnSEM from 3 independent bone marrow donors Images represent nuclei stained with DAPI Image J software was used to count nuclei in five replicatefields per chamber Scale bar=50 μm
539A Arthur et al Bone 48 (2011) 533ndash542
Discussion
In the present study we demonstrated the functional significanceof Ephephrin-B molecules expressed by MSC in relation to cellularattachment migration and osteochondral differentiation Cell motilityrelies on the convergence of a number of molecular processesincluding the establishment of focal adhesions followed by theattachment of F-actin among other associated molecules This allowsthe extension of actin processes in the form of filopodia andlamellopodia which serve to lengthen the cell [4344] These eventscan be activated by a number of ECM chemotaxis and guidancemolecules The Eph family is well known for inhibiting cell motilityparticularly during the process of boundary formation of neural crestcells restricting veins and arteries and assisting in the maintenance ofneural intestinal and dental pulp stem cell niches [172145ndash49] Inthis study we showed that activated ephrin-B reverse signalling actsto inhibit human MSC attachment and spreading by potentially
restricting focal adhesion maturation as previously shown for dentalpulp stem cells [20] The EphBephrin-B interactions trigger a numberof signalling pathways to mediate functions such as cell adhesionmigration proliferation or differentiation We demonstrated thatephrin-B induced inhibition of attachment and spreading wasmediated in part by signalling through the Src PI3Kinase and theJNK pathways This was somewhat surprising since ephrin-B reversesignalling is thought to be mediated predominantly through the Srcmolecule Grb4 or the PDZ domain However it has been demon-strated that ephrin-B1 can also signal through the JNK pathwayindependently of tyrosine kinase phosphorylation [5051] Converse-ly EphB2 forward signalling stimulated via interactions with ephrin-B1 promoted MSC migration mediated through the Src and Abl-Crksignalling pathways Therefore EphBephrin-B interactions may playa role in the recruitment of MSC from their perivascular niche towardsareas of tissue damage via the activation of either the EphB receptorsor their respective ephrin-B ligands [43742]
Fig 5 Reverse signalling promotes osteochondral differentiation (AndashD) Human MSC were cultured in osteogenic conditions for four weeks in the presence of 5 μgmL EphB2-FcEphB4-Fc ephrin-B1-Fc ephrin-B2-Fc or control human-Fc (A C) MSC cultured in the presence of EphB2-Fc produced significantly more mineral (^pb001) as demonstrated withAlizarin Red staining and (C) calcium concentration when compared to human-Fc (B D) Human MSC mineral production as demonstrated by (B) Alizarin Red staining and(D) calcium concentration was significantly inhibited in the presence of either EWLS (p=001) or TNYL (^p=001) EphB1 and EphB4 blocking peptides respectively compared tocontrol peptide RTVA (E F) HumanMSC cultured in the presence of chondrogenic media and EphB and ephrin-B-Fc molecules demonstrated significantly more GAG synthesis whencultured in the presence of (E) EphB2-Fc (pb005) when compared to the human-Fc control and significantly less GAG production when cultured with EphB2 blocking peptideSNEW (pb001) compared to peptide control RTVA All data represent the meanplusmnSEM for 3ndash4 independent marrow donors analysed by one-way ANOVA Dunnett test
540 A Arthur et al Bone 48 (2011) 533ndash542
Previously published mouse models have reported that EphBforward signalling was important for mouse osteoblastic differenti-ation [910] Similarly we observed that mineralisation of humanMSCwas decreased in the presence of the blocking peptides for EphB1 andEphB4 However our observations also suggest that reverse signallingstimulated by EphB2-Fc appears to significantly increase mineralproduction of MSC cultured in osteo-inductive media While ephrin-B-Fc molecules failed to stimulate EphB forward induced osteogen-esis addition of blocking peptides to EphB receptors inhibitedmineralproduction indicating that these blocking peptides may function notonly to block EphB forward signalling [5253] but may also inhibitreverse signalling Importantly the crystallographic studies havedemonstrated that the GndashH loop of the ephrin-B ligand physicallybinds to the ligand-binding cavity of the EphB receptor whichcoincides with the DndashE GndashH and JndashK loops resulting in eitherstabilised activation of EphB receptor or the ephrin-B ligand [54ndash57]Therefore it is plausible that by specifically blocking the ephrin-Bbinding cavity of EphB receptors these endogenous receptors may nolonger activate their cognate ephrin-B ligands While we cannotspecifically state which ligands are being blocked it could bepostulated based on binding affinity studies [58] that ephrin-B2may be predominantly required for osteoblastic differentiationHowever we cannot rule out the importance of ephrin-B1 based onthe transwells migration assays which showed that ephrin-B1activated EphB4 forward signalling Furthermore the findings ofephrin-B1 transgenic studies confirmed that reverse signallingmodulates the osteogenic response of bone marrow stromal cellsspecifically mediated through ephrin-B1 activation following stimu-lation with EphB2-Fc [33] The signalling mechanism required for theincrease in osteogenesis was mediated via PDZ signalling It appearsthat the formation of a PDZ complex containing NHERF1 PTPN13 andTAZ with ephrin-B1 results in the dephosphorylation of TAZ which
allows its translocation into the nucleus and subsequent activation ofosteogenic specific transcription factor Osterix [33]
Of note Zhao and colleagues also demonstrated high geneexpression of ephrin-B1 and ephrin-B2 by osteoblasts which werealso confirmed by Allan et al [910] However the functionalrelevance of this expression was not investigated in detail Further-more while Zhao and colleagues did use EphB4-Fc in their osteogenicassays these experiments did not demonstrate a significant differencein mineral formation [9] This finding is consistent with ourobservations as MSC produced more mineral only in response toEphB2-Fc As ephrin-B1 binds with highest affinity to EphB2 [58] itmay be assumed that MSC osteogenesis may be mediated by theEphB2ephrin-B1 receptor-ligand pairing however our blockingpeptide studies suggested that perhaps it may be mediated byephrin-B2 Our data show that EphB1 and EphB4 but not EphB2blocking peptides significantly reduce mineral formation in humanMSC where ephrin-B2 binds with highest affinity to EphB1 and EphB4[58] This observation is in line with the findings of Allan et al whoshowed that mouse Kusa 4b10 cells reduced mineral formation in thepresence of EphB4 blocking peptide TNYL at consistent concentra-tions used in the present study [10]
Alternatively the variation in EphBephrin-B signalling activatingMSC osteogenesis in this study may be attributed to the origin of theMSC used for each study The studies performed by Zhao et al andAllan et al utilized osteogenic cells derived from calvarial bone whichis formed by intramembranous ossification while the long bonesundergo predominantly endochondral ossification [5960] To accountfor these discrepancies we performed comparative studies usingbone marrow derived mouse MSC isolated from C57BL6 mousefemurs and found that these cells showed similar responses to theirhuman counterparts (unpublished observations) Furthermore theephrin-B1 transgenic studies also utilised stromal cells from the
541A Arthur et al Bone 48 (2011) 533ndash542
marrow rather than the calvarial bone identified similar findings tothe present study [33]
Studies were also performed to investigate the contribution of theEphephrin-B family in the early stages of chondrogenesis Similar tothe osteo-inductive assays we demonstrated that ephrin-B reversesignalling mediated by EphB2-Fc stimulated an increase in GAGproduction which could be inhibited by blocking the function ofendogenous EphB2 Again this observation would suggest that byblocking endogenous EphB2 ligand-binding and function may also beinhibited However unlike the mineralisation assay these findingssuggest that perhaps ephrin-B1 the highest binding affinity cognateligand for EphB2 [58] may be important for endochondral formationThis is a novel observation that the B-subclass Ephephrins may beinvolved in endochondral ossification following fracture and byhuman MSC To date the only studies to our knowledge investigatingthe B-subclass demonstrate that osteoarthritic subchondral bonemetabolism involves EphB4ephrin-B2 signalling The activation ofEphB4 by ephrin-B2 inhibits the expression of factors such as matrixmetalloproteinases interleukins and RANKL which result in reducedresorption activity leading to osteoarthritic subchondral bone [6162]Our study establishes the potential role of the EphBephrin-Bsignalling in human MSC attachmentspreading cell migration andchondrogenic and osteogenic differentiations
Supplementarymaterials related to this article can be found onlineat doi101016jbone201010180
Acknowledgments
This study was supported by a Robinson Institute Research Grantin Aid and NHMRC Project Grant 565176
References
[1] Sacchetti B Funari A Michienzi S Di Cesare S Piersanti S Saggio I et al Self-renewing osteoprogenitors in bone marrow sinusoids can organize a hematopoi-etic microenvironment Cell 2007131324ndash36
[2] Shi S Gronthos S Perivascular niche of postnatal mesenchymal stem cells inhuman bone marrow and dental pulp J Bone Miner Res 200318696ndash704
[3] Mankani MH Kuznetsov SA Robey PG Formation of hematopoietic territories andbone by transplanted human bone marrow stromal cells requires a critical celldensity Exp Hematol 200735995ndash1004
[4] Arthur A Zannettino A Gronthos S The therapeutic applications of multipotentialmesenchymalstromal stem cells in skeletal tissue repair J Cell Physiol 2008218237ndash45
[5] Komano Y Nanki T Hayashida K Taniguchi K Miyasaka N Identification of ahuman peripheral blood monocyte subset that differentiates into osteoclastsArthritis Res Ther 20068R152
[6] Karsenty G Role of Cbfa1 in osteoblast differentiation and function Semin Cell DevBiol 200011343ndash6
[7] Karsenty G Ducy P Starbuck M Priemel M Shen J Geoffroy V et al Cbfa1 as aregulator of osteoblast differentiation and function Bone 199925107ndash8
[8] Olsen BR Reginato AM Wang W Bone development Annu Rev Cell Dev Biol200016191ndash220
[9] Zhao C Irie N Takada Y Shimoda K Miyamoto T Nishiwaki T et al BidirectionalephrinB2-EphB4 signaling controls bone homeostasis Cell Metab 20064111ndash21
[10] Allan EH Hausler KD Wei T Gooi JH Quinn JM Crimeen-Irwin B et al EphrinB2regulation by PTH and PTHrP revealed by molecular profiling in differentiatingosteoblasts J Bone Miner Res 2008231170ndash81
[11] Irie N Takada Y Watanabe Y Matsuzaki Y Naruse C Asano M et al Bidirectionalsignaling through ephrinA2-EphA2 enhances osteoclastogenesis and suppressesosteoblastogenesis J Biol Chem 200928414637ndash44
[12] Cooke JE Moens CB Boundary formation in the hindbrain Eph only it weresimple Trends Neurosci 200225260ndash7
[13] Mellitzer G Xu Q Wilkinson DG Eph receptors and ephrins restrict cellintermingling and communication Nature 199940077ndash81
[14] Jayasena CS Flood WD Koblar SA High EphA3 expressing ophthalmic trigeminalsensory axons are sensitive to ephrin-A5-Fc implications for lobe specific axonguidance Neuroscience 200513597ndash109
[15] Koblar SA Krull CE Pasquale EB McLennan R Peale FD Cerretti DP et al Spinalmotor axons and neural crest cells use different molecular guides for segmentalmigration through the rostral half-somite J Neurobiol 200042437ndash47
[16] Chumley MJ Catchpole T Silvany RE Kernie SG Henkemeyer M EphB receptorsregulate stemprogenitor cell proliferation migration and polarity duringhippocampal neurogenesis J Neurosci 20072713481ndash90
[17] Merrill AE Bochukova EG Brugger SM Ishii M Pilz DTWall SA et al Cell mixing ata neural crest-mesoderm boundary and deficient ephrin-Eph signaling in thepathogenesis of craniosynostosis Hum Mol Genet 2006151319ndash28
[18] Sakai D Wakamatsu Y Regulatory mechanisms for neural crest formation CellsTissues Organs 200517924ndash35
[19] Okubo T Yanai N Obinata M Stromal cells modulate ephrinB2 expression andtransmigration of hematopoietic cells Exp Hematol 200634330ndash8
[20] Stokowski A Shi S Sun T Bartold PM Koblar SA Gronthos S EphBephrin-Binteraction mediates adult stem cell attachment spreading and migrationimplications for dental tissue repair Stem Cells 200725156ndash64
[21] Arthur A Koblar S Shi S Gronthos S EphephrinB mediate dental pulp stem cellmobilization and function J Dent Res 200988829ndash34
[22] Merlos-Suarez A Batlle E Eph-ephrin signalling in adult tissues and cancer CurrOpin Cell Biol 200820194ndash200
[23] Moore R Larue L Cell surface molecules and truncal neural crest ontogeny aperspective Birth Defects Res C Embryo Today 200472140ndash50
[24] Picco V Hudson C Yasuo H EphrinndashEph signalling drives the asymmetric division ofnotochordneural precursors in Ciona embryos Development 20071341491ndash7
[25] Sancho E Batlle E Clevers H Live and let die in the intestinal epithelium Curr OpinCell Biol 200315763ndash70
[26] Wang Z Cohen K Shao Y Mole P Dombkowski D Scadden DT Ephrin receptorEphB4 regulates ES cell differentiation of primitive mammalian hemangioblastsblood cardiomyocytes and blood vessels Blood 2004103100ndash9
[27] Compagni A Logan M Klein R Adams RH Control of skeletal patterning byephrinB1-EphB interactions Dev Cell 20035217ndash30
[28] Davy A Aubin J Soriano P Ephrin-B1 forward and reverse signaling are requiredduring mouse development Genes Dev 200418572ndash83
[29] Wieland I Jakubiczka S Muschke P CohenM Thiele H Gerlach KL et al Mutationsof the ephrin-B1 gene cause craniofrontonasal syndrome Am J Hum Genet2004741209ndash15
[30] Ting MC Wu NL Roybal PG Sun J Liu L Yen Y et al EphA4 as an effector of Twist1in the guidance of osteogenic precursor cells during calvarial bone growth and incraniosynostosis Development 2009136855ndash64
[31] Varelias A Koblar SA Cowled PA Carter CD Clayer M Human osteosarcomaexpresses specific ephrin profiles implications for tumorigenicity and prognosisCancer 200295862ndash9
[32] Xing W Baylink D Kesavan C Hu Y Kapoor S Chadwick RB et al Global geneexpression analysis in the bones reveals involvement of several novel genes andpathways in mediating an anabolic response of mechanical loading in mice J CellBiochem 2005961049ndash60
[33] Xing W Kim J Wergedal J Chen ST Mohan S Ephrin B1 regulates bone marrowstromal cell differentiation and bone formation by influencing TAZ transactivationvia complex formation with NHERF1 Mol Cell Biol 201030(3)711ndash21
[34] Twigg SR Kan R Babbs C Bochukova EG Robertson SP Wall SA et al Mutations ofephrin-B1 (EFNB1) a marker of tissue boundary formation cause craniofronto-nasal syndrome Proc Natl Acad Sci USA 20041018652ndash7
[35] Gronthos S Simmons PJ The growth factor requirements of STRO-1-positivehuman bone marrow stromal precursors under serum-deprived conditions invitro Blood 199585929ndash40
[36] Pittenger MF Mackay AM Beck SC Jaiswal RK Douglas R Mosca JD et alMultilineage potential of adult human mesenchymal stem cells Science1999284143ndash7
[37] Gronthos S Zannettino AC Hay SJ Shi S Graves SE Kortesidis A et al Molecularand cellular characterisation of highly purified stromal stem cells derived fromhuman bone marrow J Cell Sci 20031161827ndash35
[38] Salvucci O Yao L Villalba S Sajewicz A Pittaluga S Tosato G Regulation ofendothelial cell branching morphogenesis by endogenous chemokine stromal-derived factor-1 Blood 2002992703ndash11
[39] Fitter S Dewar AL Kostakis P To LB Hughes TP Roberts MM et al Long-termimatinib therapy promotes bone formation in CML patients Blood 20081112538ndash47
[40] Isenmann S Arthur A Zannettino AC Turner JL Shi S Glackin CA et al TWISTfamily of basic helix-loop-helix transcription factors mediate human mesenchy-mal stem cell growth and commitment Stem Cells 2009272457ndash68
[41] Diamond P Labrinidis A Martin SK Farrugia AN Gronthos S To LB et al Targeteddisruption of the CXCL12CXCR4 axis inhibits osteolysis in a murine model ofmyeloma-associated bone loss J Bone Miner Res 2009241150ndash61
[42] Zannettino AC Paton S Arthur A Khor F Itescu S Gimble JM et al Multipotentialhuman adipose-derived stromal stem cells exhibit a perivascular phenotype invitro and in vivo J Cell Physiol 2008214413ndash21
[43] DeMali KA Burridge K Coupling membrane protrusion and cell adhesion J Cell Sci20031162389ndash97
[44] de Hoog CL Foster LJ Mann M RNA and RNA binding proteins participate in earlystages of cell spreading through spreading initiation centers Cell 2004117649ndash62
[45] Foo SS Turner CJ Adams S Compagni A Aubyn D Kogata N et al Ephrin-B2controls cell motility and adhesion during blood-vessel-wall assembly Cell2006124161ndash73
[46] Zhang J Hughes S Role of the ephrin and Eph receptor tyrosine kinase families inangiogenesis and development of the cardiovascular system J Pathol 2006208453ndash61
[47] Arthur A Rychkov G Shi S Koblar SA Gronthos S Adult human dental pulp stemcells differentiate toward functionally active neurons under appropriate environ-mental cues Stem Cells 2008261787ndash95
[48] Holmberg J Genander M Halford MM Anneren C Sondell M Chumley MJ et alEphB receptors coordinate migration and proliferation in the intestinal stem cellniche Cell 20061251151ndash63
542 A Arthur et al Bone 48 (2011) 533ndash542
[49] Holmberg J Armulik A Senti KA Edoff K Spalding K Momma S et al Ephrin-A2reverse signaling negatively regulates neural progenitor proliferation andneurogenesis Genes Dev 200519462ndash71
[50] Xu Z Lai KO Zhou HM Lin SC Ip NY Ephrin-B1 reverse signaling activates JNKthrough a novel mechanism that is independent of tyrosine phosphorylation J BiolChem 200327824767ndash75
[51] Huynh-Do U Vindis C Liu H Cerretti DP McGrew JT Enriquez M et al Ephrin-B1transduces signals to activate integrin-mediated migration attachment andangiogenesis J Cell Sci 20021153073ndash81
[52] Koolpe M Burgess R Dail M Pasquale EB EphB receptor-binding peptidesidentified by phage display enable design of an antagonist with ephrin-likeaffinity J Biol Chem 200528017301ndash11
[53] Chrencik JE Brooun A Recht MI Nicola G Davis LK Abagyan R et al Three-dimensional structure of the EphB2 receptor in complex with an antagonis-tic peptide reveals a novel mode of inhibition J Biol Chem 200728236505ndash13
[54] Noren NK Lu M Freeman AL Koolpe M Pasquale EB Interplay between EphB4 ontumor cells and vascular ephrin-B2 regulates tumor growth Proc Natl Acad SciUSA 20041015583ndash8
[55] Himanen JP Chumley MJ LackmannM Li C BartonWA Jeffrey PD et al Repellingclass discrimination ephrin-A5 binds to and activates EphB2 receptor signalingNat Neurosci 20047501ndash9
[56] Himanen JP Rajashankar KR Lackmann M Cowan CA Henkemeyer M NikolovDB Crystal structure of an Eph receptor-ephrin complex Nature 2001414933ndash8
[57] Chrencik JE BroounA RechtMI KrausML KoolpeM Kolatkar AR et al Structure andthermodynamic characterization of the EphB4Ephrin-B2 antagonist peptidecomplex reveals thedeterminants for receptor specificity Structure 200614321ndash30
[58] Pasquale EB The Eph family of receptors Curr Opin Cell Biol 19979608ndash15[59] Opperman LA Cranial sutures as intramembranous bone growth sites Dev Dyn
2000219472ndash85[60] Kronenberg HM Developmental regulation of the growth plate Nature 2003423
332ndash6[61] Kwan Tat S Pelletier JP Amiable N Boileau C Lajeunesse D Duval N et al
Activation of the receptor EphB4 by its specific ligand ephrin B2 in humanosteoarthritic subchondral bone osteoblasts Arthritis Rheum 2008583820ndash30
[62] Kwan Tat S Pelletier JP Amiable N Boileau C Lavigne M Martel-Pelletier JTreatment with ephrin B2 positively impacts the abnormal metabolism of humanosteoarthritic chondrocytes Arthritis Res Ther 200911R119

535A Arthur et al Bone 48 (2011) 533ndash542
corresponding blocking buffer for two hours at room temperatureSections were washed with PBS and incubated with Streptavidin-HRP(Pierce Thermo Scientific) for one hour at room temperature in thecorresponding blocking buffer Samples were washed again andvisualised with DAB substrate (DAKO Glostrup Denmark) Sectionswere counterstained with haematoxylin and mounted with DePix(BDH MERCK Whitehouse Station NJ USA)
Western blot
Cell lysates were prepared as previously described [38] andequivalent amounts of protein (40 μg) separated on 8 or 10 SDSPAGE gels and transferred to PVDF membranes Immunoblotting wasperformed as previously described [38] with the following antibodiesgoat anti-EphB1 (Santa Cruz) EphB2 EphB3 EphB4 EphB6 ephrin-B1ephrin-B2 and ephrin-B3 (RampD Systems Minneapolis MN USA)Following incubationwith either anti-goat Ig (Millipore) or anti-mouseIgG (Chemicon Melbourne Victoria Australia) alkaline phosphataseconjugated secondary antibodies (Millipore Billerica USA) mem-branes were developed with enhanced chemifluorescence substrate(GE Healthcare Little Chalfont USA) Membranes were washed andthen incubated in stripping buffer (Western Blot Recycling Kit AlphaDiagnostic Intl Inc) for 15 min atRT beforebeingused to re-probewithmouse anti-human β-actin (Sigma) Quantitative analysis was per-formed using ImageQuant software (GE Healthcare)
Spreading assay
The spreading assay was conducted as previously described [20]Briefly Fc-fusion proteins (Human-IgG-Fc 1 mgmL EphB-Fc andephrin-B-Fc 200 μgmL RampD Systems) were pre-clustered with a 10-fold concentration of Human anti-Goat IgG (24 mgmL) in PBS for 1 hat room temperature Ninety-six well flat-bottom plates were coatedwith Poly-L-lysine (001 solution Sigma) for 5 min at roomtemperature Following the pre-clustering of the Fc the sampleswere incubated in coated wells for 2 h at 37 degC in a 5 CO2 incubatorthen washed with PBS Cells were liberated using PUCKS-EDTA (5 mMKCl 130 mMNaCl 3 mMNaHCO3 5 mM D-glucose 10 mMHEPES (pH73) and 1 mMEDTA) and resuspended in growthmedia 8times103cellswell was incubated for 3 h at 37 degC in 5 CO2 For the inhibitor assaycells were incubated for 30 min at room temperature with signallinginhibitor PP2 (CalBiochem San Diago CA USA) LY294002 (Sigma)SP600125 (Sigma) or 01 DMSO prior to being added to the well Thecultures were then fixed and stained with phalloidin-TRITC Forconsistency images of the cultures were taken in the centre of eachwell
Migration assay
The migration assay was conducted as previously described [20]Briefly pre-clustered Fc-fusion proteins (5 μgmL) were added to thebottom chambers of transwells Cultured MSC were liberated usingPUCKS-EDTA washed and resuspended in growthmedia then seededat 3times104cellswell into the top chamber of each transwell (CorningCorning NY USA) for 24 h at 37 degC in 5 CO2 For the inhibitor assaycells were incubated for 30 min at room temperature with signallinginhibitor U0126 (Cell Signalling Technology Danvers MA USA) PP2SB203580 (CalBiochem) LY Imatinib (Kindly provided by NovartisBasel Switzerland) 01 DMSO or pre-incubated with 100 μMblocking peptides specific for EphB1 (EWLSPNLAPSVR) EphB2(SNEWILPRLPQH) EphB4 (TNYLFSPNGPIARAW) or control peptide(RTVAHHGGLYHTNAEVK) (Mimotopes Clayton Victoria Australia)for one hour on ice prior to being added to the transwell The cultureswere then fixed and the inside of the transwell was cleared beforestaining with DAPI The assay was repeated in duplicate wells andimages were taken at 3 random positions within the well for each
well The number of nuclei were counted in each image andrepresented as the average cell countfield of view
Mineralisation assay
Single cell suspensions were seeded at 8000 cellscm2 in osteo-genic-induction media [37] in the presence of either pre-clusteredEphB2-Fc EphB4-Fc human-Fc (5 μgmL) or EphB blocking peptides(50 μM) After 4 weeks the mineralisedmatrix was dissolved in 06 MHCl and calcium levels quantitated by the Cresolphthalein Complex-one assay (Thermo Electron Corporation Melbourne Victoria USA)Calcium levels were normalised to cell number by quantitation of DNAcontent per well using Hoescht as described previously [39] Inaddition mineralised matrix formation was visualised by Alizarin RedStaining as previously described [39]
Glycoasminoglycan synthesis assay
Cultured MSC were liberated using PUCKS-EDTA and seeded as asingle cell suspension in duplicate 96-well plate at 5times104cellswell inchondrogenic media [40] containing pre-clustered humanEphBephrin-B-Fc (5 μgmL) or EphB blocking peptides (50 μM) and10 ngmL TGF-β1 at 37 degC 5 CO2 After 48 h the media was replacedwith a final concentration of 1 μCi100 μLwell Sulfur-35 radionuclide(35SO4
2minus Perkin Elmer Boston MA USA) The cells were thenincubated 37 degC 5 CO2 for a further 16 h Plates were treated withPapain digestion solution (Papain (20 UmL) NaAc (01 M) EDTA(24 mM) and N-Acetyl-L-Cysteine (20 mM) in water) at 65 degC for 3 hfollowed by Cetyl pyridinyl chloride (CPC) solution (25 CPC 40Ethanol in water) and Chondroitin sulphate A (CSA) solution (CSASigma 1 mgmL final) for a further 5 min Digested samples wereharvested onto glass filter membranes (Packard CT USA) using a cellharvester (Filtermate Packard) and then analysed using thescintillation counter (Top Count NXT Perkin Elmer Boston MAUSA) Top Count NTX program (v 253) GAG synthesis was quantifiedrelative to DNA concentration using replicate plates
shRNA knockdown
RNA duplexes targeting human EphB2 and ephrin-B2 were clonedinto the pFIV-H1-copGFP lentiviral vector as per manufacturersinstructions (System Biosciences Karrinyup WA Australia) TheshRNA oligonucleotide sequences used in this study were EphB2shRNA_1 (gatccggtgtgcaacgtgtttgagttcaagagactcaaacacgttgca-caccttttttgctagcg) EphB2 shRNA_2 (gatccgctgtgatttccagtgtcaatttcaa-gagaattgacactggaaatcacagcttttttgctagcg) ephrin-B2 shRNA_1 and(gatccccaggaataaagatccaacaattcaagagattgttggatctttattcctggttttttgc-tagcg) shRNA scramble nonsilencing control (gatccagttaatatcaa-gacgctgttcaagagacagcgtcttgatattaactttttttgctagcg) Viral supernatantcollected from transfected HEK-293T cells was used to infect primaryMSC from the top 10 of GFP-expressing cells using FACS aspreviously described [41] Cell lines with greater than 60 reductionin EphB2 and ephrin-B2 gene expressions compared to correspondingshRNA scramble MSC lines were used for the functional studies
Statistics
Treated groups were compared with their corresponding controlusing the paired Students t-tests Differences were considered to bestatistically significant when the P value was less than 005 Multipletreated groups were compared with the control group using one-wayANOVA where significance (P valueb005) was determined post hocusing the Dunnett test
536 A Arthur et al Bone 48 (2011) 533ndash542
Results
Characterisation of EphBephrin-B molecules expressed by human MSC
Gene expression analyses using real time PCR (Fig 1A) confirmedthat the B-subclass Ephephrin molecules were expressed by cultureexpanded human MSC At the protein level human MSC demonstrat-ed similar levels of ephrin-B1 (003plusmn0001 SD n=2 MSC donors)ephrin-B2 (003plusmn0002 SD n=2 MSC donors) and ephrin-B3(003plusmn0004 SD n=2 MSC donors) when compared to thecorresponding β-actin controls using ImageQuant software HigherEphB4 protein expression (393plusmn0075 SD n=2 MSC donors)correlated to high gene expression in contrast to EphB1 (048plusmn0216 SD n=2 MSC donors) EphB2 (012plusmn0055 SD n=2 MSCdonors) EphB3 (026plusmn0141 SD n=2 MSC donors) and EphB6(010plusmn0063 SD n=2 MSC donors) when compared to thecorresponding β-actin controls Immunohistochemical staining ofnormal human trephine biopsies obtained from MGUS patients (nopresence of lytic disease and less than 10 plasma cells in the bonemarrow) confirmed the expression of EphB and ephrin-B (Fig 2)molecules in vivo Consistent with the in vitro characterization EphB2EphB4 and ephrin-B1 expressions appeared most pronounced withinthe human bone localising to bone lining cells osteoblasts and cellswithin the marrow where ephrin-B1 was also found to be expressedby osteocytes The protein expression of ephrin-B2 was less intenselocalising to bone lining cells and osteoclasts (Fig 2) These findings
Fig 1 Expression of B-subclass Eph receptors and ephrin ligands by MSC (A) Real-time PCEphB4 and ephrin-B2 are most highly expressed by cultured MSC The histograms represenMSC-3 (B) The data depict representative Western blot analyses of different EphB and efollowing stripping and re-probing of each membrane
were inconsistent with the in vitro observations where ephrin-B3 wasexpressed by a large number of bone marrow cells (Fig 2)Furthermore these experiments demonstrated that EphB1 B2 B4and ephrin-B1 -B2 and -B3 molecules were expressed at varyingintensities around blood vessels similar to that observed for theperivascular and MSC associated marker CD146 (Fig 2) [43742]
EphBephrin-B interaction inhibit spreading via reverse signalling whilepromoting migration by forward signalling
Studies were conducted to determine the effect of forward andreverse signalling on the attachment spreading and migration ofMSC in vitro using an established spreading assay [20] Culturedhuman MCS exposed to EphB2-Fc or EphB4-Fc (5 μgmL) weresignificantly rounder (pb0001 ANOVA Dunnett test n=3 marrowdonors) and smaller (pb0001 ANOVA Dunnett test n=3 marrowdonors) as demonstrated with F-actin staining compared to controlhuman-Fc (Figs 3AndashC) Multiple signalling pathways were activat-ed to promote the contraction and rounding up ofMSC Incubation ofMSC with signalling pathway-specific inhibitors prior to performingthe spreading assay revealed that both the Src kinase pathway (PP2)and the PI3Kinase pathway (LY294002)were activated upon ephrin-B reverse signalling in response to EphB2-Fc while the PI3Kinaseand JNK pathways were predominantly activated in response toEphB4-Fc (Fig 3D) No difference inmorphologywas observedwhenMSC were cultured in the presence of ephrin-B1-Fc or ephrin-B2-Fc
R data were expressed as the gene transcript relative to β-actin indicates that EphB2t the meanplusmnSEM from 3 independent experiments for each donor MSC-1 MSC-2 andphrin-B proteins expressed by cultured MSC and the corresponding β-actin controls
Fig 2 Expression of B-subclass Eph receptors and ephrin ligands in human bone marrow trephines Paraffin-embedded sections of human trephine were stained with antibodiesreactive to human EphB receptors (brown) EphB1 EphB2 and EphB4 and ephrin-B ligands (brown) ephrin-B1 ephrin-B2 ephrin-B3 and counterstained with haematoxylin (blue)EphB receptors and ephrin-B ligands localised to osteoblasts (arrowheads) osteocytes (asterisks) multinucleated cells and perivascular cells (CD146 staining arrow) asdemonstrated with serial sections stained with perivascular marker CD146 (brown) Scale bar=50 μm
537A Arthur et al Bone 48 (2011) 533ndash542
(Figs 3AndashC) Supplementary Fig 1 represents a supportive studydemonstrating that MSC spreading and attachment are mediated byreverse signalling In these experiments shRNA ephrin-B2 knock-down MSC failed to undergo rounding in the presence of theephrinB2 high affinity receptor partner EphB4-Fc when comparedto the shRNA scramble nonsilencing control MSC However round-ing was still observed with both the ephrin-B2 knockdown andscramble MSC lines in the presence the ephrinB1 high affinitybinding partner EphB2-Fc demonstrating the specificity of EphB4for ephrin-B2
Migration assays using transwells were performed to examineMSC migration in vitro as previously described [20] Clustered solubleEphB2 EphB4 ephrin-B1 or ephrin-B2-Fc were added to the bottomchambers and the MSC were seeded into the top chambers of thetranswells Those cells that migrated through the transwell poreswithin 24 h were fixed and stained with DAPI before imaging Theresults indicated that human MSC migration was unaffected byephrin-B reverse signalling in the presence of EphB2-Fc or EphB4-Fc
(5 μgmL) when compared to the human-Fc (Fig 4A) However MSCmigration was significantly increased (pb001 ANOVA Dunnett testn=3 marrow donors) in the presence of ephrin-B1-Fc (5 μgmL)(Fig 4A) which appeared to be partially but significantly reversed(pb0001 ANOVA Dunnett test n=3 marrow donors) in thepresence of either the Src kinase inhibitor PP2 or the Abl kinaseinhibitor Imatinib (Fig 4B) Furthermore EphB2 forward signallingwas found to mediate migration where a shRNA EphB2 knockdownMSC line exhibited a defective migratory response to ephrin-B1-Fc incontrast to the shRNA scramble nonsilencing control MSC line(Fig 4C)
EphBephrin-B interactions influence MSC osteogenic and chondrogenicdifferentiations
To decipher the role of EphBephrin-B molecules in osteogenesishuman MSC were cultured in osteogenic inductive media in thepresence of EphB-Fc or ephrin-B-Fc molecules (Figs 5A and C)
Fig 3 Reverse ephrin-B signalling inhibits human MSC attachment and spreading (A) Representative images of human MSC cultured with 5 μgmL EphB2-Fc EphB4-Fc ephrin-B1-Fc or ephrin-B2-Fc attached to the surface of the well (BndashC) MSC were significantly rounder (pb0001) (B) and smaller (pb0001) (C) when cultured with EphB2-Fc or EphB4-Fcrespectively but not with ephrin-B1-Fc or ephrin-B2-Fc compared to the control human-Fc Data are meanplusmnSEM from 3 independent marrow donors per treatment (D) DMSOcontrol EphB2-Fc or EphB4-Fc mediated inhibition of MSC attachment was reversed in the presence of Src kinase inhibitor (25 μM PP2) PI3K inhibitor (10 μM LY294002) or JNKinhibitor (30 μM SP600125) (E) Graphs for inhibitor spreading analysis Scale bar=50 μm All data present the meanplusmnSEM from 3 independent bone marrow donors analysed byone-way ANOVA Dunnett test
538 A Arthur et al Bone 48 (2011) 533ndash542
Following 4 weeks of culture MSC produced significantly (pb001ANOVA Dunnett test n=4 bone marrow donors) more mineral inthe presence of EphB2-Fc when compared to the control human-Fcas demonstrated by Alizarin Red staining and calcium release fromthe extracellular matrix (Figs 5A and C) Furthermore MSC culturedin osteogenic media were significantly inhibited (pb001 ANOVADunnett test n=4 marrow donors) from forming mineral whencultured with blocking peptides for EphB1 (EWLS) and EphB4(TNYL) when compared to the control peptide (RTVA) (Figs 5Band D)
In parallel studies the role of Eph and ephrin molecules in theearly stages of chondrogenesis based on glycosaminoglycan (GAG)synthesis was also examined Consistent with the osteogenicdifferentiation significantly (pb005 ANOVA Dunnett test n=3marrow donors)more GAGwas produced in the presence of EphB2-Fcwhen compared to human-Fc (5 μgmL) following high densityculture in the presence of TGFβ-1 (Fig 5E) Interestingly whilereverse signalling enhanced GAG production blocking the EphB2receptor significantly inhibited (pb001 ANOVA Dunnett test n=3marrow donors) GAG synthesis (Fig 5F)
Fig 4 Forward EphB signalling promotes MSC migration (A) Transwell migration assays demonstrated that human MSC significantly increased (+pb001 ANOVA Dunnett test)their migration in the presence of 5 μgmL ephrin-B1-Fc compared to control human-Fc (B) ephrin-B1-Fc mediated MSC migration (DMSO control) was significantly reduced(plusmnpb0001) in the presence of PP2 (10 μM) and (^pb001 ANOVA Dunnett test) the presence of Imatinib (3 μM) in contrast to MEKERK inhibitor (U0126) p38 MAPK inhibitorSB203580 (SB) and LY294002 (LY) (C) MSCmigration mediated by ephrin-B1 activation was significantly inhibited in a shRNA EphB2 knockdownMSC line in either the presence ofeither 5 μgmL human-Fc (p=00014 paired Student t-test) or ephrin-B2-Fc (p=000037 Student t-test) when compared to the shRNA scramble nonsilencing control MSC Alldata represent the meanplusmnSEM from 3 independent bone marrow donors Images represent nuclei stained with DAPI Image J software was used to count nuclei in five replicatefields per chamber Scale bar=50 μm
539A Arthur et al Bone 48 (2011) 533ndash542
Discussion
In the present study we demonstrated the functional significanceof Ephephrin-B molecules expressed by MSC in relation to cellularattachment migration and osteochondral differentiation Cell motilityrelies on the convergence of a number of molecular processesincluding the establishment of focal adhesions followed by theattachment of F-actin among other associated molecules This allowsthe extension of actin processes in the form of filopodia andlamellopodia which serve to lengthen the cell [4344] These eventscan be activated by a number of ECM chemotaxis and guidancemolecules The Eph family is well known for inhibiting cell motilityparticularly during the process of boundary formation of neural crestcells restricting veins and arteries and assisting in the maintenance ofneural intestinal and dental pulp stem cell niches [172145ndash49] Inthis study we showed that activated ephrin-B reverse signalling actsto inhibit human MSC attachment and spreading by potentially
restricting focal adhesion maturation as previously shown for dentalpulp stem cells [20] The EphBephrin-B interactions trigger a numberof signalling pathways to mediate functions such as cell adhesionmigration proliferation or differentiation We demonstrated thatephrin-B induced inhibition of attachment and spreading wasmediated in part by signalling through the Src PI3Kinase and theJNK pathways This was somewhat surprising since ephrin-B reversesignalling is thought to be mediated predominantly through the Srcmolecule Grb4 or the PDZ domain However it has been demon-strated that ephrin-B1 can also signal through the JNK pathwayindependently of tyrosine kinase phosphorylation [5051] Converse-ly EphB2 forward signalling stimulated via interactions with ephrin-B1 promoted MSC migration mediated through the Src and Abl-Crksignalling pathways Therefore EphBephrin-B interactions may playa role in the recruitment of MSC from their perivascular niche towardsareas of tissue damage via the activation of either the EphB receptorsor their respective ephrin-B ligands [43742]
Fig 5 Reverse signalling promotes osteochondral differentiation (AndashD) Human MSC were cultured in osteogenic conditions for four weeks in the presence of 5 μgmL EphB2-FcEphB4-Fc ephrin-B1-Fc ephrin-B2-Fc or control human-Fc (A C) MSC cultured in the presence of EphB2-Fc produced significantly more mineral (^pb001) as demonstrated withAlizarin Red staining and (C) calcium concentration when compared to human-Fc (B D) Human MSC mineral production as demonstrated by (B) Alizarin Red staining and(D) calcium concentration was significantly inhibited in the presence of either EWLS (p=001) or TNYL (^p=001) EphB1 and EphB4 blocking peptides respectively compared tocontrol peptide RTVA (E F) HumanMSC cultured in the presence of chondrogenic media and EphB and ephrin-B-Fc molecules demonstrated significantly more GAG synthesis whencultured in the presence of (E) EphB2-Fc (pb005) when compared to the human-Fc control and significantly less GAG production when cultured with EphB2 blocking peptideSNEW (pb001) compared to peptide control RTVA All data represent the meanplusmnSEM for 3ndash4 independent marrow donors analysed by one-way ANOVA Dunnett test
540 A Arthur et al Bone 48 (2011) 533ndash542
Previously published mouse models have reported that EphBforward signalling was important for mouse osteoblastic differenti-ation [910] Similarly we observed that mineralisation of humanMSCwas decreased in the presence of the blocking peptides for EphB1 andEphB4 However our observations also suggest that reverse signallingstimulated by EphB2-Fc appears to significantly increase mineralproduction of MSC cultured in osteo-inductive media While ephrin-B-Fc molecules failed to stimulate EphB forward induced osteogen-esis addition of blocking peptides to EphB receptors inhibitedmineralproduction indicating that these blocking peptides may function notonly to block EphB forward signalling [5253] but may also inhibitreverse signalling Importantly the crystallographic studies havedemonstrated that the GndashH loop of the ephrin-B ligand physicallybinds to the ligand-binding cavity of the EphB receptor whichcoincides with the DndashE GndashH and JndashK loops resulting in eitherstabilised activation of EphB receptor or the ephrin-B ligand [54ndash57]Therefore it is plausible that by specifically blocking the ephrin-Bbinding cavity of EphB receptors these endogenous receptors may nolonger activate their cognate ephrin-B ligands While we cannotspecifically state which ligands are being blocked it could bepostulated based on binding affinity studies [58] that ephrin-B2may be predominantly required for osteoblastic differentiationHowever we cannot rule out the importance of ephrin-B1 based onthe transwells migration assays which showed that ephrin-B1activated EphB4 forward signalling Furthermore the findings ofephrin-B1 transgenic studies confirmed that reverse signallingmodulates the osteogenic response of bone marrow stromal cellsspecifically mediated through ephrin-B1 activation following stimu-lation with EphB2-Fc [33] The signalling mechanism required for theincrease in osteogenesis was mediated via PDZ signalling It appearsthat the formation of a PDZ complex containing NHERF1 PTPN13 andTAZ with ephrin-B1 results in the dephosphorylation of TAZ which
allows its translocation into the nucleus and subsequent activation ofosteogenic specific transcription factor Osterix [33]
Of note Zhao and colleagues also demonstrated high geneexpression of ephrin-B1 and ephrin-B2 by osteoblasts which werealso confirmed by Allan et al [910] However the functionalrelevance of this expression was not investigated in detail Further-more while Zhao and colleagues did use EphB4-Fc in their osteogenicassays these experiments did not demonstrate a significant differencein mineral formation [9] This finding is consistent with ourobservations as MSC produced more mineral only in response toEphB2-Fc As ephrin-B1 binds with highest affinity to EphB2 [58] itmay be assumed that MSC osteogenesis may be mediated by theEphB2ephrin-B1 receptor-ligand pairing however our blockingpeptide studies suggested that perhaps it may be mediated byephrin-B2 Our data show that EphB1 and EphB4 but not EphB2blocking peptides significantly reduce mineral formation in humanMSC where ephrin-B2 binds with highest affinity to EphB1 and EphB4[58] This observation is in line with the findings of Allan et al whoshowed that mouse Kusa 4b10 cells reduced mineral formation in thepresence of EphB4 blocking peptide TNYL at consistent concentra-tions used in the present study [10]
Alternatively the variation in EphBephrin-B signalling activatingMSC osteogenesis in this study may be attributed to the origin of theMSC used for each study The studies performed by Zhao et al andAllan et al utilized osteogenic cells derived from calvarial bone whichis formed by intramembranous ossification while the long bonesundergo predominantly endochondral ossification [5960] To accountfor these discrepancies we performed comparative studies usingbone marrow derived mouse MSC isolated from C57BL6 mousefemurs and found that these cells showed similar responses to theirhuman counterparts (unpublished observations) Furthermore theephrin-B1 transgenic studies also utilised stromal cells from the
541A Arthur et al Bone 48 (2011) 533ndash542
marrow rather than the calvarial bone identified similar findings tothe present study [33]
Studies were also performed to investigate the contribution of theEphephrin-B family in the early stages of chondrogenesis Similar tothe osteo-inductive assays we demonstrated that ephrin-B reversesignalling mediated by EphB2-Fc stimulated an increase in GAGproduction which could be inhibited by blocking the function ofendogenous EphB2 Again this observation would suggest that byblocking endogenous EphB2 ligand-binding and function may also beinhibited However unlike the mineralisation assay these findingssuggest that perhaps ephrin-B1 the highest binding affinity cognateligand for EphB2 [58] may be important for endochondral formationThis is a novel observation that the B-subclass Ephephrins may beinvolved in endochondral ossification following fracture and byhuman MSC To date the only studies to our knowledge investigatingthe B-subclass demonstrate that osteoarthritic subchondral bonemetabolism involves EphB4ephrin-B2 signalling The activation ofEphB4 by ephrin-B2 inhibits the expression of factors such as matrixmetalloproteinases interleukins and RANKL which result in reducedresorption activity leading to osteoarthritic subchondral bone [6162]Our study establishes the potential role of the EphBephrin-Bsignalling in human MSC attachmentspreading cell migration andchondrogenic and osteogenic differentiations
Supplementarymaterials related to this article can be found onlineat doi101016jbone201010180
Acknowledgments
This study was supported by a Robinson Institute Research Grantin Aid and NHMRC Project Grant 565176
References
[1] Sacchetti B Funari A Michienzi S Di Cesare S Piersanti S Saggio I et al Self-renewing osteoprogenitors in bone marrow sinusoids can organize a hematopoi-etic microenvironment Cell 2007131324ndash36
[2] Shi S Gronthos S Perivascular niche of postnatal mesenchymal stem cells inhuman bone marrow and dental pulp J Bone Miner Res 200318696ndash704
[3] Mankani MH Kuznetsov SA Robey PG Formation of hematopoietic territories andbone by transplanted human bone marrow stromal cells requires a critical celldensity Exp Hematol 200735995ndash1004
[4] Arthur A Zannettino A Gronthos S The therapeutic applications of multipotentialmesenchymalstromal stem cells in skeletal tissue repair J Cell Physiol 2008218237ndash45
[5] Komano Y Nanki T Hayashida K Taniguchi K Miyasaka N Identification of ahuman peripheral blood monocyte subset that differentiates into osteoclastsArthritis Res Ther 20068R152
[6] Karsenty G Role of Cbfa1 in osteoblast differentiation and function Semin Cell DevBiol 200011343ndash6
[7] Karsenty G Ducy P Starbuck M Priemel M Shen J Geoffroy V et al Cbfa1 as aregulator of osteoblast differentiation and function Bone 199925107ndash8
[8] Olsen BR Reginato AM Wang W Bone development Annu Rev Cell Dev Biol200016191ndash220
[9] Zhao C Irie N Takada Y Shimoda K Miyamoto T Nishiwaki T et al BidirectionalephrinB2-EphB4 signaling controls bone homeostasis Cell Metab 20064111ndash21
[10] Allan EH Hausler KD Wei T Gooi JH Quinn JM Crimeen-Irwin B et al EphrinB2regulation by PTH and PTHrP revealed by molecular profiling in differentiatingosteoblasts J Bone Miner Res 2008231170ndash81
[11] Irie N Takada Y Watanabe Y Matsuzaki Y Naruse C Asano M et al Bidirectionalsignaling through ephrinA2-EphA2 enhances osteoclastogenesis and suppressesosteoblastogenesis J Biol Chem 200928414637ndash44
[12] Cooke JE Moens CB Boundary formation in the hindbrain Eph only it weresimple Trends Neurosci 200225260ndash7
[13] Mellitzer G Xu Q Wilkinson DG Eph receptors and ephrins restrict cellintermingling and communication Nature 199940077ndash81
[14] Jayasena CS Flood WD Koblar SA High EphA3 expressing ophthalmic trigeminalsensory axons are sensitive to ephrin-A5-Fc implications for lobe specific axonguidance Neuroscience 200513597ndash109
[15] Koblar SA Krull CE Pasquale EB McLennan R Peale FD Cerretti DP et al Spinalmotor axons and neural crest cells use different molecular guides for segmentalmigration through the rostral half-somite J Neurobiol 200042437ndash47
[16] Chumley MJ Catchpole T Silvany RE Kernie SG Henkemeyer M EphB receptorsregulate stemprogenitor cell proliferation migration and polarity duringhippocampal neurogenesis J Neurosci 20072713481ndash90
[17] Merrill AE Bochukova EG Brugger SM Ishii M Pilz DTWall SA et al Cell mixing ata neural crest-mesoderm boundary and deficient ephrin-Eph signaling in thepathogenesis of craniosynostosis Hum Mol Genet 2006151319ndash28
[18] Sakai D Wakamatsu Y Regulatory mechanisms for neural crest formation CellsTissues Organs 200517924ndash35
[19] Okubo T Yanai N Obinata M Stromal cells modulate ephrinB2 expression andtransmigration of hematopoietic cells Exp Hematol 200634330ndash8
[20] Stokowski A Shi S Sun T Bartold PM Koblar SA Gronthos S EphBephrin-Binteraction mediates adult stem cell attachment spreading and migrationimplications for dental tissue repair Stem Cells 200725156ndash64
[21] Arthur A Koblar S Shi S Gronthos S EphephrinB mediate dental pulp stem cellmobilization and function J Dent Res 200988829ndash34
[22] Merlos-Suarez A Batlle E Eph-ephrin signalling in adult tissues and cancer CurrOpin Cell Biol 200820194ndash200
[23] Moore R Larue L Cell surface molecules and truncal neural crest ontogeny aperspective Birth Defects Res C Embryo Today 200472140ndash50
[24] Picco V Hudson C Yasuo H EphrinndashEph signalling drives the asymmetric division ofnotochordneural precursors in Ciona embryos Development 20071341491ndash7
[25] Sancho E Batlle E Clevers H Live and let die in the intestinal epithelium Curr OpinCell Biol 200315763ndash70
[26] Wang Z Cohen K Shao Y Mole P Dombkowski D Scadden DT Ephrin receptorEphB4 regulates ES cell differentiation of primitive mammalian hemangioblastsblood cardiomyocytes and blood vessels Blood 2004103100ndash9
[27] Compagni A Logan M Klein R Adams RH Control of skeletal patterning byephrinB1-EphB interactions Dev Cell 20035217ndash30
[28] Davy A Aubin J Soriano P Ephrin-B1 forward and reverse signaling are requiredduring mouse development Genes Dev 200418572ndash83
[29] Wieland I Jakubiczka S Muschke P CohenM Thiele H Gerlach KL et al Mutationsof the ephrin-B1 gene cause craniofrontonasal syndrome Am J Hum Genet2004741209ndash15
[30] Ting MC Wu NL Roybal PG Sun J Liu L Yen Y et al EphA4 as an effector of Twist1in the guidance of osteogenic precursor cells during calvarial bone growth and incraniosynostosis Development 2009136855ndash64
[31] Varelias A Koblar SA Cowled PA Carter CD Clayer M Human osteosarcomaexpresses specific ephrin profiles implications for tumorigenicity and prognosisCancer 200295862ndash9
[32] Xing W Baylink D Kesavan C Hu Y Kapoor S Chadwick RB et al Global geneexpression analysis in the bones reveals involvement of several novel genes andpathways in mediating an anabolic response of mechanical loading in mice J CellBiochem 2005961049ndash60
[33] Xing W Kim J Wergedal J Chen ST Mohan S Ephrin B1 regulates bone marrowstromal cell differentiation and bone formation by influencing TAZ transactivationvia complex formation with NHERF1 Mol Cell Biol 201030(3)711ndash21
[34] Twigg SR Kan R Babbs C Bochukova EG Robertson SP Wall SA et al Mutations ofephrin-B1 (EFNB1) a marker of tissue boundary formation cause craniofronto-nasal syndrome Proc Natl Acad Sci USA 20041018652ndash7
[35] Gronthos S Simmons PJ The growth factor requirements of STRO-1-positivehuman bone marrow stromal precursors under serum-deprived conditions invitro Blood 199585929ndash40
[36] Pittenger MF Mackay AM Beck SC Jaiswal RK Douglas R Mosca JD et alMultilineage potential of adult human mesenchymal stem cells Science1999284143ndash7
[37] Gronthos S Zannettino AC Hay SJ Shi S Graves SE Kortesidis A et al Molecularand cellular characterisation of highly purified stromal stem cells derived fromhuman bone marrow J Cell Sci 20031161827ndash35
[38] Salvucci O Yao L Villalba S Sajewicz A Pittaluga S Tosato G Regulation ofendothelial cell branching morphogenesis by endogenous chemokine stromal-derived factor-1 Blood 2002992703ndash11
[39] Fitter S Dewar AL Kostakis P To LB Hughes TP Roberts MM et al Long-termimatinib therapy promotes bone formation in CML patients Blood 20081112538ndash47
[40] Isenmann S Arthur A Zannettino AC Turner JL Shi S Glackin CA et al TWISTfamily of basic helix-loop-helix transcription factors mediate human mesenchy-mal stem cell growth and commitment Stem Cells 2009272457ndash68
[41] Diamond P Labrinidis A Martin SK Farrugia AN Gronthos S To LB et al Targeteddisruption of the CXCL12CXCR4 axis inhibits osteolysis in a murine model ofmyeloma-associated bone loss J Bone Miner Res 2009241150ndash61
[42] Zannettino AC Paton S Arthur A Khor F Itescu S Gimble JM et al Multipotentialhuman adipose-derived stromal stem cells exhibit a perivascular phenotype invitro and in vivo J Cell Physiol 2008214413ndash21
[43] DeMali KA Burridge K Coupling membrane protrusion and cell adhesion J Cell Sci20031162389ndash97
[44] de Hoog CL Foster LJ Mann M RNA and RNA binding proteins participate in earlystages of cell spreading through spreading initiation centers Cell 2004117649ndash62
[45] Foo SS Turner CJ Adams S Compagni A Aubyn D Kogata N et al Ephrin-B2controls cell motility and adhesion during blood-vessel-wall assembly Cell2006124161ndash73
[46] Zhang J Hughes S Role of the ephrin and Eph receptor tyrosine kinase families inangiogenesis and development of the cardiovascular system J Pathol 2006208453ndash61
[47] Arthur A Rychkov G Shi S Koblar SA Gronthos S Adult human dental pulp stemcells differentiate toward functionally active neurons under appropriate environ-mental cues Stem Cells 2008261787ndash95
[48] Holmberg J Genander M Halford MM Anneren C Sondell M Chumley MJ et alEphB receptors coordinate migration and proliferation in the intestinal stem cellniche Cell 20061251151ndash63
542 A Arthur et al Bone 48 (2011) 533ndash542
[49] Holmberg J Armulik A Senti KA Edoff K Spalding K Momma S et al Ephrin-A2reverse signaling negatively regulates neural progenitor proliferation andneurogenesis Genes Dev 200519462ndash71
[50] Xu Z Lai KO Zhou HM Lin SC Ip NY Ephrin-B1 reverse signaling activates JNKthrough a novel mechanism that is independent of tyrosine phosphorylation J BiolChem 200327824767ndash75
[51] Huynh-Do U Vindis C Liu H Cerretti DP McGrew JT Enriquez M et al Ephrin-B1transduces signals to activate integrin-mediated migration attachment andangiogenesis J Cell Sci 20021153073ndash81
[52] Koolpe M Burgess R Dail M Pasquale EB EphB receptor-binding peptidesidentified by phage display enable design of an antagonist with ephrin-likeaffinity J Biol Chem 200528017301ndash11
[53] Chrencik JE Brooun A Recht MI Nicola G Davis LK Abagyan R et al Three-dimensional structure of the EphB2 receptor in complex with an antagonis-tic peptide reveals a novel mode of inhibition J Biol Chem 200728236505ndash13
[54] Noren NK Lu M Freeman AL Koolpe M Pasquale EB Interplay between EphB4 ontumor cells and vascular ephrin-B2 regulates tumor growth Proc Natl Acad SciUSA 20041015583ndash8
[55] Himanen JP Chumley MJ LackmannM Li C BartonWA Jeffrey PD et al Repellingclass discrimination ephrin-A5 binds to and activates EphB2 receptor signalingNat Neurosci 20047501ndash9
[56] Himanen JP Rajashankar KR Lackmann M Cowan CA Henkemeyer M NikolovDB Crystal structure of an Eph receptor-ephrin complex Nature 2001414933ndash8
[57] Chrencik JE BroounA RechtMI KrausML KoolpeM Kolatkar AR et al Structure andthermodynamic characterization of the EphB4Ephrin-B2 antagonist peptidecomplex reveals thedeterminants for receptor specificity Structure 200614321ndash30
[58] Pasquale EB The Eph family of receptors Curr Opin Cell Biol 19979608ndash15[59] Opperman LA Cranial sutures as intramembranous bone growth sites Dev Dyn
2000219472ndash85[60] Kronenberg HM Developmental regulation of the growth plate Nature 2003423
332ndash6[61] Kwan Tat S Pelletier JP Amiable N Boileau C Lajeunesse D Duval N et al
Activation of the receptor EphB4 by its specific ligand ephrin B2 in humanosteoarthritic subchondral bone osteoblasts Arthritis Rheum 2008583820ndash30
[62] Kwan Tat S Pelletier JP Amiable N Boileau C Lavigne M Martel-Pelletier JTreatment with ephrin B2 positively impacts the abnormal metabolism of humanosteoarthritic chondrocytes Arthritis Res Ther 200911R119

536 A Arthur et al Bone 48 (2011) 533ndash542
Results
Characterisation of EphBephrin-B molecules expressed by human MSC
Gene expression analyses using real time PCR (Fig 1A) confirmedthat the B-subclass Ephephrin molecules were expressed by cultureexpanded human MSC At the protein level human MSC demonstrat-ed similar levels of ephrin-B1 (003plusmn0001 SD n=2 MSC donors)ephrin-B2 (003plusmn0002 SD n=2 MSC donors) and ephrin-B3(003plusmn0004 SD n=2 MSC donors) when compared to thecorresponding β-actin controls using ImageQuant software HigherEphB4 protein expression (393plusmn0075 SD n=2 MSC donors)correlated to high gene expression in contrast to EphB1 (048plusmn0216 SD n=2 MSC donors) EphB2 (012plusmn0055 SD n=2 MSCdonors) EphB3 (026plusmn0141 SD n=2 MSC donors) and EphB6(010plusmn0063 SD n=2 MSC donors) when compared to thecorresponding β-actin controls Immunohistochemical staining ofnormal human trephine biopsies obtained from MGUS patients (nopresence of lytic disease and less than 10 plasma cells in the bonemarrow) confirmed the expression of EphB and ephrin-B (Fig 2)molecules in vivo Consistent with the in vitro characterization EphB2EphB4 and ephrin-B1 expressions appeared most pronounced withinthe human bone localising to bone lining cells osteoblasts and cellswithin the marrow where ephrin-B1 was also found to be expressedby osteocytes The protein expression of ephrin-B2 was less intenselocalising to bone lining cells and osteoclasts (Fig 2) These findings
Fig 1 Expression of B-subclass Eph receptors and ephrin ligands by MSC (A) Real-time PCEphB4 and ephrin-B2 are most highly expressed by cultured MSC The histograms represenMSC-3 (B) The data depict representative Western blot analyses of different EphB and efollowing stripping and re-probing of each membrane
were inconsistent with the in vitro observations where ephrin-B3 wasexpressed by a large number of bone marrow cells (Fig 2)Furthermore these experiments demonstrated that EphB1 B2 B4and ephrin-B1 -B2 and -B3 molecules were expressed at varyingintensities around blood vessels similar to that observed for theperivascular and MSC associated marker CD146 (Fig 2) [43742]
EphBephrin-B interaction inhibit spreading via reverse signalling whilepromoting migration by forward signalling
Studies were conducted to determine the effect of forward andreverse signalling on the attachment spreading and migration ofMSC in vitro using an established spreading assay [20] Culturedhuman MCS exposed to EphB2-Fc or EphB4-Fc (5 μgmL) weresignificantly rounder (pb0001 ANOVA Dunnett test n=3 marrowdonors) and smaller (pb0001 ANOVA Dunnett test n=3 marrowdonors) as demonstrated with F-actin staining compared to controlhuman-Fc (Figs 3AndashC) Multiple signalling pathways were activat-ed to promote the contraction and rounding up ofMSC Incubation ofMSC with signalling pathway-specific inhibitors prior to performingthe spreading assay revealed that both the Src kinase pathway (PP2)and the PI3Kinase pathway (LY294002)were activated upon ephrin-B reverse signalling in response to EphB2-Fc while the PI3Kinaseand JNK pathways were predominantly activated in response toEphB4-Fc (Fig 3D) No difference inmorphologywas observedwhenMSC were cultured in the presence of ephrin-B1-Fc or ephrin-B2-Fc
R data were expressed as the gene transcript relative to β-actin indicates that EphB2t the meanplusmnSEM from 3 independent experiments for each donor MSC-1 MSC-2 andphrin-B proteins expressed by cultured MSC and the corresponding β-actin controls
Fig 2 Expression of B-subclass Eph receptors and ephrin ligands in human bone marrow trephines Paraffin-embedded sections of human trephine were stained with antibodiesreactive to human EphB receptors (brown) EphB1 EphB2 and EphB4 and ephrin-B ligands (brown) ephrin-B1 ephrin-B2 ephrin-B3 and counterstained with haematoxylin (blue)EphB receptors and ephrin-B ligands localised to osteoblasts (arrowheads) osteocytes (asterisks) multinucleated cells and perivascular cells (CD146 staining arrow) asdemonstrated with serial sections stained with perivascular marker CD146 (brown) Scale bar=50 μm
537A Arthur et al Bone 48 (2011) 533ndash542
(Figs 3AndashC) Supplementary Fig 1 represents a supportive studydemonstrating that MSC spreading and attachment are mediated byreverse signalling In these experiments shRNA ephrin-B2 knock-down MSC failed to undergo rounding in the presence of theephrinB2 high affinity receptor partner EphB4-Fc when comparedto the shRNA scramble nonsilencing control MSC However round-ing was still observed with both the ephrin-B2 knockdown andscramble MSC lines in the presence the ephrinB1 high affinitybinding partner EphB2-Fc demonstrating the specificity of EphB4for ephrin-B2
Migration assays using transwells were performed to examineMSC migration in vitro as previously described [20] Clustered solubleEphB2 EphB4 ephrin-B1 or ephrin-B2-Fc were added to the bottomchambers and the MSC were seeded into the top chambers of thetranswells Those cells that migrated through the transwell poreswithin 24 h were fixed and stained with DAPI before imaging Theresults indicated that human MSC migration was unaffected byephrin-B reverse signalling in the presence of EphB2-Fc or EphB4-Fc
(5 μgmL) when compared to the human-Fc (Fig 4A) However MSCmigration was significantly increased (pb001 ANOVA Dunnett testn=3 marrow donors) in the presence of ephrin-B1-Fc (5 μgmL)(Fig 4A) which appeared to be partially but significantly reversed(pb0001 ANOVA Dunnett test n=3 marrow donors) in thepresence of either the Src kinase inhibitor PP2 or the Abl kinaseinhibitor Imatinib (Fig 4B) Furthermore EphB2 forward signallingwas found to mediate migration where a shRNA EphB2 knockdownMSC line exhibited a defective migratory response to ephrin-B1-Fc incontrast to the shRNA scramble nonsilencing control MSC line(Fig 4C)
EphBephrin-B interactions influence MSC osteogenic and chondrogenicdifferentiations
To decipher the role of EphBephrin-B molecules in osteogenesishuman MSC were cultured in osteogenic inductive media in thepresence of EphB-Fc or ephrin-B-Fc molecules (Figs 5A and C)
Fig 3 Reverse ephrin-B signalling inhibits human MSC attachment and spreading (A) Representative images of human MSC cultured with 5 μgmL EphB2-Fc EphB4-Fc ephrin-B1-Fc or ephrin-B2-Fc attached to the surface of the well (BndashC) MSC were significantly rounder (pb0001) (B) and smaller (pb0001) (C) when cultured with EphB2-Fc or EphB4-Fcrespectively but not with ephrin-B1-Fc or ephrin-B2-Fc compared to the control human-Fc Data are meanplusmnSEM from 3 independent marrow donors per treatment (D) DMSOcontrol EphB2-Fc or EphB4-Fc mediated inhibition of MSC attachment was reversed in the presence of Src kinase inhibitor (25 μM PP2) PI3K inhibitor (10 μM LY294002) or JNKinhibitor (30 μM SP600125) (E) Graphs for inhibitor spreading analysis Scale bar=50 μm All data present the meanplusmnSEM from 3 independent bone marrow donors analysed byone-way ANOVA Dunnett test
538 A Arthur et al Bone 48 (2011) 533ndash542
Following 4 weeks of culture MSC produced significantly (pb001ANOVA Dunnett test n=4 bone marrow donors) more mineral inthe presence of EphB2-Fc when compared to the control human-Fcas demonstrated by Alizarin Red staining and calcium release fromthe extracellular matrix (Figs 5A and C) Furthermore MSC culturedin osteogenic media were significantly inhibited (pb001 ANOVADunnett test n=4 marrow donors) from forming mineral whencultured with blocking peptides for EphB1 (EWLS) and EphB4(TNYL) when compared to the control peptide (RTVA) (Figs 5Band D)
In parallel studies the role of Eph and ephrin molecules in theearly stages of chondrogenesis based on glycosaminoglycan (GAG)synthesis was also examined Consistent with the osteogenicdifferentiation significantly (pb005 ANOVA Dunnett test n=3marrow donors)more GAGwas produced in the presence of EphB2-Fcwhen compared to human-Fc (5 μgmL) following high densityculture in the presence of TGFβ-1 (Fig 5E) Interestingly whilereverse signalling enhanced GAG production blocking the EphB2receptor significantly inhibited (pb001 ANOVA Dunnett test n=3marrow donors) GAG synthesis (Fig 5F)
Fig 4 Forward EphB signalling promotes MSC migration (A) Transwell migration assays demonstrated that human MSC significantly increased (+pb001 ANOVA Dunnett test)their migration in the presence of 5 μgmL ephrin-B1-Fc compared to control human-Fc (B) ephrin-B1-Fc mediated MSC migration (DMSO control) was significantly reduced(plusmnpb0001) in the presence of PP2 (10 μM) and (^pb001 ANOVA Dunnett test) the presence of Imatinib (3 μM) in contrast to MEKERK inhibitor (U0126) p38 MAPK inhibitorSB203580 (SB) and LY294002 (LY) (C) MSCmigration mediated by ephrin-B1 activation was significantly inhibited in a shRNA EphB2 knockdownMSC line in either the presence ofeither 5 μgmL human-Fc (p=00014 paired Student t-test) or ephrin-B2-Fc (p=000037 Student t-test) when compared to the shRNA scramble nonsilencing control MSC Alldata represent the meanplusmnSEM from 3 independent bone marrow donors Images represent nuclei stained with DAPI Image J software was used to count nuclei in five replicatefields per chamber Scale bar=50 μm
539A Arthur et al Bone 48 (2011) 533ndash542
Discussion
In the present study we demonstrated the functional significanceof Ephephrin-B molecules expressed by MSC in relation to cellularattachment migration and osteochondral differentiation Cell motilityrelies on the convergence of a number of molecular processesincluding the establishment of focal adhesions followed by theattachment of F-actin among other associated molecules This allowsthe extension of actin processes in the form of filopodia andlamellopodia which serve to lengthen the cell [4344] These eventscan be activated by a number of ECM chemotaxis and guidancemolecules The Eph family is well known for inhibiting cell motilityparticularly during the process of boundary formation of neural crestcells restricting veins and arteries and assisting in the maintenance ofneural intestinal and dental pulp stem cell niches [172145ndash49] Inthis study we showed that activated ephrin-B reverse signalling actsto inhibit human MSC attachment and spreading by potentially
restricting focal adhesion maturation as previously shown for dentalpulp stem cells [20] The EphBephrin-B interactions trigger a numberof signalling pathways to mediate functions such as cell adhesionmigration proliferation or differentiation We demonstrated thatephrin-B induced inhibition of attachment and spreading wasmediated in part by signalling through the Src PI3Kinase and theJNK pathways This was somewhat surprising since ephrin-B reversesignalling is thought to be mediated predominantly through the Srcmolecule Grb4 or the PDZ domain However it has been demon-strated that ephrin-B1 can also signal through the JNK pathwayindependently of tyrosine kinase phosphorylation [5051] Converse-ly EphB2 forward signalling stimulated via interactions with ephrin-B1 promoted MSC migration mediated through the Src and Abl-Crksignalling pathways Therefore EphBephrin-B interactions may playa role in the recruitment of MSC from their perivascular niche towardsareas of tissue damage via the activation of either the EphB receptorsor their respective ephrin-B ligands [43742]
Fig 5 Reverse signalling promotes osteochondral differentiation (AndashD) Human MSC were cultured in osteogenic conditions for four weeks in the presence of 5 μgmL EphB2-FcEphB4-Fc ephrin-B1-Fc ephrin-B2-Fc or control human-Fc (A C) MSC cultured in the presence of EphB2-Fc produced significantly more mineral (^pb001) as demonstrated withAlizarin Red staining and (C) calcium concentration when compared to human-Fc (B D) Human MSC mineral production as demonstrated by (B) Alizarin Red staining and(D) calcium concentration was significantly inhibited in the presence of either EWLS (p=001) or TNYL (^p=001) EphB1 and EphB4 blocking peptides respectively compared tocontrol peptide RTVA (E F) HumanMSC cultured in the presence of chondrogenic media and EphB and ephrin-B-Fc molecules demonstrated significantly more GAG synthesis whencultured in the presence of (E) EphB2-Fc (pb005) when compared to the human-Fc control and significantly less GAG production when cultured with EphB2 blocking peptideSNEW (pb001) compared to peptide control RTVA All data represent the meanplusmnSEM for 3ndash4 independent marrow donors analysed by one-way ANOVA Dunnett test
540 A Arthur et al Bone 48 (2011) 533ndash542
Previously published mouse models have reported that EphBforward signalling was important for mouse osteoblastic differenti-ation [910] Similarly we observed that mineralisation of humanMSCwas decreased in the presence of the blocking peptides for EphB1 andEphB4 However our observations also suggest that reverse signallingstimulated by EphB2-Fc appears to significantly increase mineralproduction of MSC cultured in osteo-inductive media While ephrin-B-Fc molecules failed to stimulate EphB forward induced osteogen-esis addition of blocking peptides to EphB receptors inhibitedmineralproduction indicating that these blocking peptides may function notonly to block EphB forward signalling [5253] but may also inhibitreverse signalling Importantly the crystallographic studies havedemonstrated that the GndashH loop of the ephrin-B ligand physicallybinds to the ligand-binding cavity of the EphB receptor whichcoincides with the DndashE GndashH and JndashK loops resulting in eitherstabilised activation of EphB receptor or the ephrin-B ligand [54ndash57]Therefore it is plausible that by specifically blocking the ephrin-Bbinding cavity of EphB receptors these endogenous receptors may nolonger activate their cognate ephrin-B ligands While we cannotspecifically state which ligands are being blocked it could bepostulated based on binding affinity studies [58] that ephrin-B2may be predominantly required for osteoblastic differentiationHowever we cannot rule out the importance of ephrin-B1 based onthe transwells migration assays which showed that ephrin-B1activated EphB4 forward signalling Furthermore the findings ofephrin-B1 transgenic studies confirmed that reverse signallingmodulates the osteogenic response of bone marrow stromal cellsspecifically mediated through ephrin-B1 activation following stimu-lation with EphB2-Fc [33] The signalling mechanism required for theincrease in osteogenesis was mediated via PDZ signalling It appearsthat the formation of a PDZ complex containing NHERF1 PTPN13 andTAZ with ephrin-B1 results in the dephosphorylation of TAZ which
allows its translocation into the nucleus and subsequent activation ofosteogenic specific transcription factor Osterix [33]
Of note Zhao and colleagues also demonstrated high geneexpression of ephrin-B1 and ephrin-B2 by osteoblasts which werealso confirmed by Allan et al [910] However the functionalrelevance of this expression was not investigated in detail Further-more while Zhao and colleagues did use EphB4-Fc in their osteogenicassays these experiments did not demonstrate a significant differencein mineral formation [9] This finding is consistent with ourobservations as MSC produced more mineral only in response toEphB2-Fc As ephrin-B1 binds with highest affinity to EphB2 [58] itmay be assumed that MSC osteogenesis may be mediated by theEphB2ephrin-B1 receptor-ligand pairing however our blockingpeptide studies suggested that perhaps it may be mediated byephrin-B2 Our data show that EphB1 and EphB4 but not EphB2blocking peptides significantly reduce mineral formation in humanMSC where ephrin-B2 binds with highest affinity to EphB1 and EphB4[58] This observation is in line with the findings of Allan et al whoshowed that mouse Kusa 4b10 cells reduced mineral formation in thepresence of EphB4 blocking peptide TNYL at consistent concentra-tions used in the present study [10]
Alternatively the variation in EphBephrin-B signalling activatingMSC osteogenesis in this study may be attributed to the origin of theMSC used for each study The studies performed by Zhao et al andAllan et al utilized osteogenic cells derived from calvarial bone whichis formed by intramembranous ossification while the long bonesundergo predominantly endochondral ossification [5960] To accountfor these discrepancies we performed comparative studies usingbone marrow derived mouse MSC isolated from C57BL6 mousefemurs and found that these cells showed similar responses to theirhuman counterparts (unpublished observations) Furthermore theephrin-B1 transgenic studies also utilised stromal cells from the
541A Arthur et al Bone 48 (2011) 533ndash542
marrow rather than the calvarial bone identified similar findings tothe present study [33]
Studies were also performed to investigate the contribution of theEphephrin-B family in the early stages of chondrogenesis Similar tothe osteo-inductive assays we demonstrated that ephrin-B reversesignalling mediated by EphB2-Fc stimulated an increase in GAGproduction which could be inhibited by blocking the function ofendogenous EphB2 Again this observation would suggest that byblocking endogenous EphB2 ligand-binding and function may also beinhibited However unlike the mineralisation assay these findingssuggest that perhaps ephrin-B1 the highest binding affinity cognateligand for EphB2 [58] may be important for endochondral formationThis is a novel observation that the B-subclass Ephephrins may beinvolved in endochondral ossification following fracture and byhuman MSC To date the only studies to our knowledge investigatingthe B-subclass demonstrate that osteoarthritic subchondral bonemetabolism involves EphB4ephrin-B2 signalling The activation ofEphB4 by ephrin-B2 inhibits the expression of factors such as matrixmetalloproteinases interleukins and RANKL which result in reducedresorption activity leading to osteoarthritic subchondral bone [6162]Our study establishes the potential role of the EphBephrin-Bsignalling in human MSC attachmentspreading cell migration andchondrogenic and osteogenic differentiations
Supplementarymaterials related to this article can be found onlineat doi101016jbone201010180
Acknowledgments
This study was supported by a Robinson Institute Research Grantin Aid and NHMRC Project Grant 565176
References
[1] Sacchetti B Funari A Michienzi S Di Cesare S Piersanti S Saggio I et al Self-renewing osteoprogenitors in bone marrow sinusoids can organize a hematopoi-etic microenvironment Cell 2007131324ndash36
[2] Shi S Gronthos S Perivascular niche of postnatal mesenchymal stem cells inhuman bone marrow and dental pulp J Bone Miner Res 200318696ndash704
[3] Mankani MH Kuznetsov SA Robey PG Formation of hematopoietic territories andbone by transplanted human bone marrow stromal cells requires a critical celldensity Exp Hematol 200735995ndash1004
[4] Arthur A Zannettino A Gronthos S The therapeutic applications of multipotentialmesenchymalstromal stem cells in skeletal tissue repair J Cell Physiol 2008218237ndash45
[5] Komano Y Nanki T Hayashida K Taniguchi K Miyasaka N Identification of ahuman peripheral blood monocyte subset that differentiates into osteoclastsArthritis Res Ther 20068R152
[6] Karsenty G Role of Cbfa1 in osteoblast differentiation and function Semin Cell DevBiol 200011343ndash6
[7] Karsenty G Ducy P Starbuck M Priemel M Shen J Geoffroy V et al Cbfa1 as aregulator of osteoblast differentiation and function Bone 199925107ndash8
[8] Olsen BR Reginato AM Wang W Bone development Annu Rev Cell Dev Biol200016191ndash220
[9] Zhao C Irie N Takada Y Shimoda K Miyamoto T Nishiwaki T et al BidirectionalephrinB2-EphB4 signaling controls bone homeostasis Cell Metab 20064111ndash21
[10] Allan EH Hausler KD Wei T Gooi JH Quinn JM Crimeen-Irwin B et al EphrinB2regulation by PTH and PTHrP revealed by molecular profiling in differentiatingosteoblasts J Bone Miner Res 2008231170ndash81
[11] Irie N Takada Y Watanabe Y Matsuzaki Y Naruse C Asano M et al Bidirectionalsignaling through ephrinA2-EphA2 enhances osteoclastogenesis and suppressesosteoblastogenesis J Biol Chem 200928414637ndash44
[12] Cooke JE Moens CB Boundary formation in the hindbrain Eph only it weresimple Trends Neurosci 200225260ndash7
[13] Mellitzer G Xu Q Wilkinson DG Eph receptors and ephrins restrict cellintermingling and communication Nature 199940077ndash81
[14] Jayasena CS Flood WD Koblar SA High EphA3 expressing ophthalmic trigeminalsensory axons are sensitive to ephrin-A5-Fc implications for lobe specific axonguidance Neuroscience 200513597ndash109
[15] Koblar SA Krull CE Pasquale EB McLennan R Peale FD Cerretti DP et al Spinalmotor axons and neural crest cells use different molecular guides for segmentalmigration through the rostral half-somite J Neurobiol 200042437ndash47
[16] Chumley MJ Catchpole T Silvany RE Kernie SG Henkemeyer M EphB receptorsregulate stemprogenitor cell proliferation migration and polarity duringhippocampal neurogenesis J Neurosci 20072713481ndash90
[17] Merrill AE Bochukova EG Brugger SM Ishii M Pilz DTWall SA et al Cell mixing ata neural crest-mesoderm boundary and deficient ephrin-Eph signaling in thepathogenesis of craniosynostosis Hum Mol Genet 2006151319ndash28
[18] Sakai D Wakamatsu Y Regulatory mechanisms for neural crest formation CellsTissues Organs 200517924ndash35
[19] Okubo T Yanai N Obinata M Stromal cells modulate ephrinB2 expression andtransmigration of hematopoietic cells Exp Hematol 200634330ndash8
[20] Stokowski A Shi S Sun T Bartold PM Koblar SA Gronthos S EphBephrin-Binteraction mediates adult stem cell attachment spreading and migrationimplications for dental tissue repair Stem Cells 200725156ndash64
[21] Arthur A Koblar S Shi S Gronthos S EphephrinB mediate dental pulp stem cellmobilization and function J Dent Res 200988829ndash34
[22] Merlos-Suarez A Batlle E Eph-ephrin signalling in adult tissues and cancer CurrOpin Cell Biol 200820194ndash200
[23] Moore R Larue L Cell surface molecules and truncal neural crest ontogeny aperspective Birth Defects Res C Embryo Today 200472140ndash50
[24] Picco V Hudson C Yasuo H EphrinndashEph signalling drives the asymmetric division ofnotochordneural precursors in Ciona embryos Development 20071341491ndash7
[25] Sancho E Batlle E Clevers H Live and let die in the intestinal epithelium Curr OpinCell Biol 200315763ndash70
[26] Wang Z Cohen K Shao Y Mole P Dombkowski D Scadden DT Ephrin receptorEphB4 regulates ES cell differentiation of primitive mammalian hemangioblastsblood cardiomyocytes and blood vessels Blood 2004103100ndash9
[27] Compagni A Logan M Klein R Adams RH Control of skeletal patterning byephrinB1-EphB interactions Dev Cell 20035217ndash30
[28] Davy A Aubin J Soriano P Ephrin-B1 forward and reverse signaling are requiredduring mouse development Genes Dev 200418572ndash83
[29] Wieland I Jakubiczka S Muschke P CohenM Thiele H Gerlach KL et al Mutationsof the ephrin-B1 gene cause craniofrontonasal syndrome Am J Hum Genet2004741209ndash15
[30] Ting MC Wu NL Roybal PG Sun J Liu L Yen Y et al EphA4 as an effector of Twist1in the guidance of osteogenic precursor cells during calvarial bone growth and incraniosynostosis Development 2009136855ndash64
[31] Varelias A Koblar SA Cowled PA Carter CD Clayer M Human osteosarcomaexpresses specific ephrin profiles implications for tumorigenicity and prognosisCancer 200295862ndash9
[32] Xing W Baylink D Kesavan C Hu Y Kapoor S Chadwick RB et al Global geneexpression analysis in the bones reveals involvement of several novel genes andpathways in mediating an anabolic response of mechanical loading in mice J CellBiochem 2005961049ndash60
[33] Xing W Kim J Wergedal J Chen ST Mohan S Ephrin B1 regulates bone marrowstromal cell differentiation and bone formation by influencing TAZ transactivationvia complex formation with NHERF1 Mol Cell Biol 201030(3)711ndash21
[34] Twigg SR Kan R Babbs C Bochukova EG Robertson SP Wall SA et al Mutations ofephrin-B1 (EFNB1) a marker of tissue boundary formation cause craniofronto-nasal syndrome Proc Natl Acad Sci USA 20041018652ndash7
[35] Gronthos S Simmons PJ The growth factor requirements of STRO-1-positivehuman bone marrow stromal precursors under serum-deprived conditions invitro Blood 199585929ndash40
[36] Pittenger MF Mackay AM Beck SC Jaiswal RK Douglas R Mosca JD et alMultilineage potential of adult human mesenchymal stem cells Science1999284143ndash7
[37] Gronthos S Zannettino AC Hay SJ Shi S Graves SE Kortesidis A et al Molecularand cellular characterisation of highly purified stromal stem cells derived fromhuman bone marrow J Cell Sci 20031161827ndash35
[38] Salvucci O Yao L Villalba S Sajewicz A Pittaluga S Tosato G Regulation ofendothelial cell branching morphogenesis by endogenous chemokine stromal-derived factor-1 Blood 2002992703ndash11
[39] Fitter S Dewar AL Kostakis P To LB Hughes TP Roberts MM et al Long-termimatinib therapy promotes bone formation in CML patients Blood 20081112538ndash47
[40] Isenmann S Arthur A Zannettino AC Turner JL Shi S Glackin CA et al TWISTfamily of basic helix-loop-helix transcription factors mediate human mesenchy-mal stem cell growth and commitment Stem Cells 2009272457ndash68
[41] Diamond P Labrinidis A Martin SK Farrugia AN Gronthos S To LB et al Targeteddisruption of the CXCL12CXCR4 axis inhibits osteolysis in a murine model ofmyeloma-associated bone loss J Bone Miner Res 2009241150ndash61
[42] Zannettino AC Paton S Arthur A Khor F Itescu S Gimble JM et al Multipotentialhuman adipose-derived stromal stem cells exhibit a perivascular phenotype invitro and in vivo J Cell Physiol 2008214413ndash21
[43] DeMali KA Burridge K Coupling membrane protrusion and cell adhesion J Cell Sci20031162389ndash97
[44] de Hoog CL Foster LJ Mann M RNA and RNA binding proteins participate in earlystages of cell spreading through spreading initiation centers Cell 2004117649ndash62
[45] Foo SS Turner CJ Adams S Compagni A Aubyn D Kogata N et al Ephrin-B2controls cell motility and adhesion during blood-vessel-wall assembly Cell2006124161ndash73
[46] Zhang J Hughes S Role of the ephrin and Eph receptor tyrosine kinase families inangiogenesis and development of the cardiovascular system J Pathol 2006208453ndash61
[47] Arthur A Rychkov G Shi S Koblar SA Gronthos S Adult human dental pulp stemcells differentiate toward functionally active neurons under appropriate environ-mental cues Stem Cells 2008261787ndash95
[48] Holmberg J Genander M Halford MM Anneren C Sondell M Chumley MJ et alEphB receptors coordinate migration and proliferation in the intestinal stem cellniche Cell 20061251151ndash63
542 A Arthur et al Bone 48 (2011) 533ndash542
[49] Holmberg J Armulik A Senti KA Edoff K Spalding K Momma S et al Ephrin-A2reverse signaling negatively regulates neural progenitor proliferation andneurogenesis Genes Dev 200519462ndash71
[50] Xu Z Lai KO Zhou HM Lin SC Ip NY Ephrin-B1 reverse signaling activates JNKthrough a novel mechanism that is independent of tyrosine phosphorylation J BiolChem 200327824767ndash75
[51] Huynh-Do U Vindis C Liu H Cerretti DP McGrew JT Enriquez M et al Ephrin-B1transduces signals to activate integrin-mediated migration attachment andangiogenesis J Cell Sci 20021153073ndash81
[52] Koolpe M Burgess R Dail M Pasquale EB EphB receptor-binding peptidesidentified by phage display enable design of an antagonist with ephrin-likeaffinity J Biol Chem 200528017301ndash11
[53] Chrencik JE Brooun A Recht MI Nicola G Davis LK Abagyan R et al Three-dimensional structure of the EphB2 receptor in complex with an antagonis-tic peptide reveals a novel mode of inhibition J Biol Chem 200728236505ndash13
[54] Noren NK Lu M Freeman AL Koolpe M Pasquale EB Interplay between EphB4 ontumor cells and vascular ephrin-B2 regulates tumor growth Proc Natl Acad SciUSA 20041015583ndash8
[55] Himanen JP Chumley MJ LackmannM Li C BartonWA Jeffrey PD et al Repellingclass discrimination ephrin-A5 binds to and activates EphB2 receptor signalingNat Neurosci 20047501ndash9
[56] Himanen JP Rajashankar KR Lackmann M Cowan CA Henkemeyer M NikolovDB Crystal structure of an Eph receptor-ephrin complex Nature 2001414933ndash8
[57] Chrencik JE BroounA RechtMI KrausML KoolpeM Kolatkar AR et al Structure andthermodynamic characterization of the EphB4Ephrin-B2 antagonist peptidecomplex reveals thedeterminants for receptor specificity Structure 200614321ndash30
[58] Pasquale EB The Eph family of receptors Curr Opin Cell Biol 19979608ndash15[59] Opperman LA Cranial sutures as intramembranous bone growth sites Dev Dyn
2000219472ndash85[60] Kronenberg HM Developmental regulation of the growth plate Nature 2003423
332ndash6[61] Kwan Tat S Pelletier JP Amiable N Boileau C Lajeunesse D Duval N et al
Activation of the receptor EphB4 by its specific ligand ephrin B2 in humanosteoarthritic subchondral bone osteoblasts Arthritis Rheum 2008583820ndash30
[62] Kwan Tat S Pelletier JP Amiable N Boileau C Lavigne M Martel-Pelletier JTreatment with ephrin B2 positively impacts the abnormal metabolism of humanosteoarthritic chondrocytes Arthritis Res Ther 200911R119

Fig 2 Expression of B-subclass Eph receptors and ephrin ligands in human bone marrow trephines Paraffin-embedded sections of human trephine were stained with antibodiesreactive to human EphB receptors (brown) EphB1 EphB2 and EphB4 and ephrin-B ligands (brown) ephrin-B1 ephrin-B2 ephrin-B3 and counterstained with haematoxylin (blue)EphB receptors and ephrin-B ligands localised to osteoblasts (arrowheads) osteocytes (asterisks) multinucleated cells and perivascular cells (CD146 staining arrow) asdemonstrated with serial sections stained with perivascular marker CD146 (brown) Scale bar=50 μm
537A Arthur et al Bone 48 (2011) 533ndash542
(Figs 3AndashC) Supplementary Fig 1 represents a supportive studydemonstrating that MSC spreading and attachment are mediated byreverse signalling In these experiments shRNA ephrin-B2 knock-down MSC failed to undergo rounding in the presence of theephrinB2 high affinity receptor partner EphB4-Fc when comparedto the shRNA scramble nonsilencing control MSC However round-ing was still observed with both the ephrin-B2 knockdown andscramble MSC lines in the presence the ephrinB1 high affinitybinding partner EphB2-Fc demonstrating the specificity of EphB4for ephrin-B2
Migration assays using transwells were performed to examineMSC migration in vitro as previously described [20] Clustered solubleEphB2 EphB4 ephrin-B1 or ephrin-B2-Fc were added to the bottomchambers and the MSC were seeded into the top chambers of thetranswells Those cells that migrated through the transwell poreswithin 24 h were fixed and stained with DAPI before imaging Theresults indicated that human MSC migration was unaffected byephrin-B reverse signalling in the presence of EphB2-Fc or EphB4-Fc
(5 μgmL) when compared to the human-Fc (Fig 4A) However MSCmigration was significantly increased (pb001 ANOVA Dunnett testn=3 marrow donors) in the presence of ephrin-B1-Fc (5 μgmL)(Fig 4A) which appeared to be partially but significantly reversed(pb0001 ANOVA Dunnett test n=3 marrow donors) in thepresence of either the Src kinase inhibitor PP2 or the Abl kinaseinhibitor Imatinib (Fig 4B) Furthermore EphB2 forward signallingwas found to mediate migration where a shRNA EphB2 knockdownMSC line exhibited a defective migratory response to ephrin-B1-Fc incontrast to the shRNA scramble nonsilencing control MSC line(Fig 4C)
EphBephrin-B interactions influence MSC osteogenic and chondrogenicdifferentiations
To decipher the role of EphBephrin-B molecules in osteogenesishuman MSC were cultured in osteogenic inductive media in thepresence of EphB-Fc or ephrin-B-Fc molecules (Figs 5A and C)
Fig 3 Reverse ephrin-B signalling inhibits human MSC attachment and spreading (A) Representative images of human MSC cultured with 5 μgmL EphB2-Fc EphB4-Fc ephrin-B1-Fc or ephrin-B2-Fc attached to the surface of the well (BndashC) MSC were significantly rounder (pb0001) (B) and smaller (pb0001) (C) when cultured with EphB2-Fc or EphB4-Fcrespectively but not with ephrin-B1-Fc or ephrin-B2-Fc compared to the control human-Fc Data are meanplusmnSEM from 3 independent marrow donors per treatment (D) DMSOcontrol EphB2-Fc or EphB4-Fc mediated inhibition of MSC attachment was reversed in the presence of Src kinase inhibitor (25 μM PP2) PI3K inhibitor (10 μM LY294002) or JNKinhibitor (30 μM SP600125) (E) Graphs for inhibitor spreading analysis Scale bar=50 μm All data present the meanplusmnSEM from 3 independent bone marrow donors analysed byone-way ANOVA Dunnett test
538 A Arthur et al Bone 48 (2011) 533ndash542
Following 4 weeks of culture MSC produced significantly (pb001ANOVA Dunnett test n=4 bone marrow donors) more mineral inthe presence of EphB2-Fc when compared to the control human-Fcas demonstrated by Alizarin Red staining and calcium release fromthe extracellular matrix (Figs 5A and C) Furthermore MSC culturedin osteogenic media were significantly inhibited (pb001 ANOVADunnett test n=4 marrow donors) from forming mineral whencultured with blocking peptides for EphB1 (EWLS) and EphB4(TNYL) when compared to the control peptide (RTVA) (Figs 5Band D)
In parallel studies the role of Eph and ephrin molecules in theearly stages of chondrogenesis based on glycosaminoglycan (GAG)synthesis was also examined Consistent with the osteogenicdifferentiation significantly (pb005 ANOVA Dunnett test n=3marrow donors)more GAGwas produced in the presence of EphB2-Fcwhen compared to human-Fc (5 μgmL) following high densityculture in the presence of TGFβ-1 (Fig 5E) Interestingly whilereverse signalling enhanced GAG production blocking the EphB2receptor significantly inhibited (pb001 ANOVA Dunnett test n=3marrow donors) GAG synthesis (Fig 5F)
Fig 4 Forward EphB signalling promotes MSC migration (A) Transwell migration assays demonstrated that human MSC significantly increased (+pb001 ANOVA Dunnett test)their migration in the presence of 5 μgmL ephrin-B1-Fc compared to control human-Fc (B) ephrin-B1-Fc mediated MSC migration (DMSO control) was significantly reduced(plusmnpb0001) in the presence of PP2 (10 μM) and (^pb001 ANOVA Dunnett test) the presence of Imatinib (3 μM) in contrast to MEKERK inhibitor (U0126) p38 MAPK inhibitorSB203580 (SB) and LY294002 (LY) (C) MSCmigration mediated by ephrin-B1 activation was significantly inhibited in a shRNA EphB2 knockdownMSC line in either the presence ofeither 5 μgmL human-Fc (p=00014 paired Student t-test) or ephrin-B2-Fc (p=000037 Student t-test) when compared to the shRNA scramble nonsilencing control MSC Alldata represent the meanplusmnSEM from 3 independent bone marrow donors Images represent nuclei stained with DAPI Image J software was used to count nuclei in five replicatefields per chamber Scale bar=50 μm
539A Arthur et al Bone 48 (2011) 533ndash542
Discussion
In the present study we demonstrated the functional significanceof Ephephrin-B molecules expressed by MSC in relation to cellularattachment migration and osteochondral differentiation Cell motilityrelies on the convergence of a number of molecular processesincluding the establishment of focal adhesions followed by theattachment of F-actin among other associated molecules This allowsthe extension of actin processes in the form of filopodia andlamellopodia which serve to lengthen the cell [4344] These eventscan be activated by a number of ECM chemotaxis and guidancemolecules The Eph family is well known for inhibiting cell motilityparticularly during the process of boundary formation of neural crestcells restricting veins and arteries and assisting in the maintenance ofneural intestinal and dental pulp stem cell niches [172145ndash49] Inthis study we showed that activated ephrin-B reverse signalling actsto inhibit human MSC attachment and spreading by potentially
restricting focal adhesion maturation as previously shown for dentalpulp stem cells [20] The EphBephrin-B interactions trigger a numberof signalling pathways to mediate functions such as cell adhesionmigration proliferation or differentiation We demonstrated thatephrin-B induced inhibition of attachment and spreading wasmediated in part by signalling through the Src PI3Kinase and theJNK pathways This was somewhat surprising since ephrin-B reversesignalling is thought to be mediated predominantly through the Srcmolecule Grb4 or the PDZ domain However it has been demon-strated that ephrin-B1 can also signal through the JNK pathwayindependently of tyrosine kinase phosphorylation [5051] Converse-ly EphB2 forward signalling stimulated via interactions with ephrin-B1 promoted MSC migration mediated through the Src and Abl-Crksignalling pathways Therefore EphBephrin-B interactions may playa role in the recruitment of MSC from their perivascular niche towardsareas of tissue damage via the activation of either the EphB receptorsor their respective ephrin-B ligands [43742]
Fig 5 Reverse signalling promotes osteochondral differentiation (AndashD) Human MSC were cultured in osteogenic conditions for four weeks in the presence of 5 μgmL EphB2-FcEphB4-Fc ephrin-B1-Fc ephrin-B2-Fc or control human-Fc (A C) MSC cultured in the presence of EphB2-Fc produced significantly more mineral (^pb001) as demonstrated withAlizarin Red staining and (C) calcium concentration when compared to human-Fc (B D) Human MSC mineral production as demonstrated by (B) Alizarin Red staining and(D) calcium concentration was significantly inhibited in the presence of either EWLS (p=001) or TNYL (^p=001) EphB1 and EphB4 blocking peptides respectively compared tocontrol peptide RTVA (E F) HumanMSC cultured in the presence of chondrogenic media and EphB and ephrin-B-Fc molecules demonstrated significantly more GAG synthesis whencultured in the presence of (E) EphB2-Fc (pb005) when compared to the human-Fc control and significantly less GAG production when cultured with EphB2 blocking peptideSNEW (pb001) compared to peptide control RTVA All data represent the meanplusmnSEM for 3ndash4 independent marrow donors analysed by one-way ANOVA Dunnett test
540 A Arthur et al Bone 48 (2011) 533ndash542
Previously published mouse models have reported that EphBforward signalling was important for mouse osteoblastic differenti-ation [910] Similarly we observed that mineralisation of humanMSCwas decreased in the presence of the blocking peptides for EphB1 andEphB4 However our observations also suggest that reverse signallingstimulated by EphB2-Fc appears to significantly increase mineralproduction of MSC cultured in osteo-inductive media While ephrin-B-Fc molecules failed to stimulate EphB forward induced osteogen-esis addition of blocking peptides to EphB receptors inhibitedmineralproduction indicating that these blocking peptides may function notonly to block EphB forward signalling [5253] but may also inhibitreverse signalling Importantly the crystallographic studies havedemonstrated that the GndashH loop of the ephrin-B ligand physicallybinds to the ligand-binding cavity of the EphB receptor whichcoincides with the DndashE GndashH and JndashK loops resulting in eitherstabilised activation of EphB receptor or the ephrin-B ligand [54ndash57]Therefore it is plausible that by specifically blocking the ephrin-Bbinding cavity of EphB receptors these endogenous receptors may nolonger activate their cognate ephrin-B ligands While we cannotspecifically state which ligands are being blocked it could bepostulated based on binding affinity studies [58] that ephrin-B2may be predominantly required for osteoblastic differentiationHowever we cannot rule out the importance of ephrin-B1 based onthe transwells migration assays which showed that ephrin-B1activated EphB4 forward signalling Furthermore the findings ofephrin-B1 transgenic studies confirmed that reverse signallingmodulates the osteogenic response of bone marrow stromal cellsspecifically mediated through ephrin-B1 activation following stimu-lation with EphB2-Fc [33] The signalling mechanism required for theincrease in osteogenesis was mediated via PDZ signalling It appearsthat the formation of a PDZ complex containing NHERF1 PTPN13 andTAZ with ephrin-B1 results in the dephosphorylation of TAZ which
allows its translocation into the nucleus and subsequent activation ofosteogenic specific transcription factor Osterix [33]
Of note Zhao and colleagues also demonstrated high geneexpression of ephrin-B1 and ephrin-B2 by osteoblasts which werealso confirmed by Allan et al [910] However the functionalrelevance of this expression was not investigated in detail Further-more while Zhao and colleagues did use EphB4-Fc in their osteogenicassays these experiments did not demonstrate a significant differencein mineral formation [9] This finding is consistent with ourobservations as MSC produced more mineral only in response toEphB2-Fc As ephrin-B1 binds with highest affinity to EphB2 [58] itmay be assumed that MSC osteogenesis may be mediated by theEphB2ephrin-B1 receptor-ligand pairing however our blockingpeptide studies suggested that perhaps it may be mediated byephrin-B2 Our data show that EphB1 and EphB4 but not EphB2blocking peptides significantly reduce mineral formation in humanMSC where ephrin-B2 binds with highest affinity to EphB1 and EphB4[58] This observation is in line with the findings of Allan et al whoshowed that mouse Kusa 4b10 cells reduced mineral formation in thepresence of EphB4 blocking peptide TNYL at consistent concentra-tions used in the present study [10]
Alternatively the variation in EphBephrin-B signalling activatingMSC osteogenesis in this study may be attributed to the origin of theMSC used for each study The studies performed by Zhao et al andAllan et al utilized osteogenic cells derived from calvarial bone whichis formed by intramembranous ossification while the long bonesundergo predominantly endochondral ossification [5960] To accountfor these discrepancies we performed comparative studies usingbone marrow derived mouse MSC isolated from C57BL6 mousefemurs and found that these cells showed similar responses to theirhuman counterparts (unpublished observations) Furthermore theephrin-B1 transgenic studies also utilised stromal cells from the
541A Arthur et al Bone 48 (2011) 533ndash542
marrow rather than the calvarial bone identified similar findings tothe present study [33]
Studies were also performed to investigate the contribution of theEphephrin-B family in the early stages of chondrogenesis Similar tothe osteo-inductive assays we demonstrated that ephrin-B reversesignalling mediated by EphB2-Fc stimulated an increase in GAGproduction which could be inhibited by blocking the function ofendogenous EphB2 Again this observation would suggest that byblocking endogenous EphB2 ligand-binding and function may also beinhibited However unlike the mineralisation assay these findingssuggest that perhaps ephrin-B1 the highest binding affinity cognateligand for EphB2 [58] may be important for endochondral formationThis is a novel observation that the B-subclass Ephephrins may beinvolved in endochondral ossification following fracture and byhuman MSC To date the only studies to our knowledge investigatingthe B-subclass demonstrate that osteoarthritic subchondral bonemetabolism involves EphB4ephrin-B2 signalling The activation ofEphB4 by ephrin-B2 inhibits the expression of factors such as matrixmetalloproteinases interleukins and RANKL which result in reducedresorption activity leading to osteoarthritic subchondral bone [6162]Our study establishes the potential role of the EphBephrin-Bsignalling in human MSC attachmentspreading cell migration andchondrogenic and osteogenic differentiations
Supplementarymaterials related to this article can be found onlineat doi101016jbone201010180
Acknowledgments
This study was supported by a Robinson Institute Research Grantin Aid and NHMRC Project Grant 565176
References
[1] Sacchetti B Funari A Michienzi S Di Cesare S Piersanti S Saggio I et al Self-renewing osteoprogenitors in bone marrow sinusoids can organize a hematopoi-etic microenvironment Cell 2007131324ndash36
[2] Shi S Gronthos S Perivascular niche of postnatal mesenchymal stem cells inhuman bone marrow and dental pulp J Bone Miner Res 200318696ndash704
[3] Mankani MH Kuznetsov SA Robey PG Formation of hematopoietic territories andbone by transplanted human bone marrow stromal cells requires a critical celldensity Exp Hematol 200735995ndash1004
[4] Arthur A Zannettino A Gronthos S The therapeutic applications of multipotentialmesenchymalstromal stem cells in skeletal tissue repair J Cell Physiol 2008218237ndash45
[5] Komano Y Nanki T Hayashida K Taniguchi K Miyasaka N Identification of ahuman peripheral blood monocyte subset that differentiates into osteoclastsArthritis Res Ther 20068R152
[6] Karsenty G Role of Cbfa1 in osteoblast differentiation and function Semin Cell DevBiol 200011343ndash6
[7] Karsenty G Ducy P Starbuck M Priemel M Shen J Geoffroy V et al Cbfa1 as aregulator of osteoblast differentiation and function Bone 199925107ndash8
[8] Olsen BR Reginato AM Wang W Bone development Annu Rev Cell Dev Biol200016191ndash220
[9] Zhao C Irie N Takada Y Shimoda K Miyamoto T Nishiwaki T et al BidirectionalephrinB2-EphB4 signaling controls bone homeostasis Cell Metab 20064111ndash21
[10] Allan EH Hausler KD Wei T Gooi JH Quinn JM Crimeen-Irwin B et al EphrinB2regulation by PTH and PTHrP revealed by molecular profiling in differentiatingosteoblasts J Bone Miner Res 2008231170ndash81
[11] Irie N Takada Y Watanabe Y Matsuzaki Y Naruse C Asano M et al Bidirectionalsignaling through ephrinA2-EphA2 enhances osteoclastogenesis and suppressesosteoblastogenesis J Biol Chem 200928414637ndash44
[12] Cooke JE Moens CB Boundary formation in the hindbrain Eph only it weresimple Trends Neurosci 200225260ndash7
[13] Mellitzer G Xu Q Wilkinson DG Eph receptors and ephrins restrict cellintermingling and communication Nature 199940077ndash81
[14] Jayasena CS Flood WD Koblar SA High EphA3 expressing ophthalmic trigeminalsensory axons are sensitive to ephrin-A5-Fc implications for lobe specific axonguidance Neuroscience 200513597ndash109
[15] Koblar SA Krull CE Pasquale EB McLennan R Peale FD Cerretti DP et al Spinalmotor axons and neural crest cells use different molecular guides for segmentalmigration through the rostral half-somite J Neurobiol 200042437ndash47
[16] Chumley MJ Catchpole T Silvany RE Kernie SG Henkemeyer M EphB receptorsregulate stemprogenitor cell proliferation migration and polarity duringhippocampal neurogenesis J Neurosci 20072713481ndash90
[17] Merrill AE Bochukova EG Brugger SM Ishii M Pilz DTWall SA et al Cell mixing ata neural crest-mesoderm boundary and deficient ephrin-Eph signaling in thepathogenesis of craniosynostosis Hum Mol Genet 2006151319ndash28
[18] Sakai D Wakamatsu Y Regulatory mechanisms for neural crest formation CellsTissues Organs 200517924ndash35
[19] Okubo T Yanai N Obinata M Stromal cells modulate ephrinB2 expression andtransmigration of hematopoietic cells Exp Hematol 200634330ndash8
[20] Stokowski A Shi S Sun T Bartold PM Koblar SA Gronthos S EphBephrin-Binteraction mediates adult stem cell attachment spreading and migrationimplications for dental tissue repair Stem Cells 200725156ndash64
[21] Arthur A Koblar S Shi S Gronthos S EphephrinB mediate dental pulp stem cellmobilization and function J Dent Res 200988829ndash34
[22] Merlos-Suarez A Batlle E Eph-ephrin signalling in adult tissues and cancer CurrOpin Cell Biol 200820194ndash200
[23] Moore R Larue L Cell surface molecules and truncal neural crest ontogeny aperspective Birth Defects Res C Embryo Today 200472140ndash50
[24] Picco V Hudson C Yasuo H EphrinndashEph signalling drives the asymmetric division ofnotochordneural precursors in Ciona embryos Development 20071341491ndash7
[25] Sancho E Batlle E Clevers H Live and let die in the intestinal epithelium Curr OpinCell Biol 200315763ndash70
[26] Wang Z Cohen K Shao Y Mole P Dombkowski D Scadden DT Ephrin receptorEphB4 regulates ES cell differentiation of primitive mammalian hemangioblastsblood cardiomyocytes and blood vessels Blood 2004103100ndash9
[27] Compagni A Logan M Klein R Adams RH Control of skeletal patterning byephrinB1-EphB interactions Dev Cell 20035217ndash30
[28] Davy A Aubin J Soriano P Ephrin-B1 forward and reverse signaling are requiredduring mouse development Genes Dev 200418572ndash83
[29] Wieland I Jakubiczka S Muschke P CohenM Thiele H Gerlach KL et al Mutationsof the ephrin-B1 gene cause craniofrontonasal syndrome Am J Hum Genet2004741209ndash15
[30] Ting MC Wu NL Roybal PG Sun J Liu L Yen Y et al EphA4 as an effector of Twist1in the guidance of osteogenic precursor cells during calvarial bone growth and incraniosynostosis Development 2009136855ndash64
[31] Varelias A Koblar SA Cowled PA Carter CD Clayer M Human osteosarcomaexpresses specific ephrin profiles implications for tumorigenicity and prognosisCancer 200295862ndash9
[32] Xing W Baylink D Kesavan C Hu Y Kapoor S Chadwick RB et al Global geneexpression analysis in the bones reveals involvement of several novel genes andpathways in mediating an anabolic response of mechanical loading in mice J CellBiochem 2005961049ndash60
[33] Xing W Kim J Wergedal J Chen ST Mohan S Ephrin B1 regulates bone marrowstromal cell differentiation and bone formation by influencing TAZ transactivationvia complex formation with NHERF1 Mol Cell Biol 201030(3)711ndash21
[34] Twigg SR Kan R Babbs C Bochukova EG Robertson SP Wall SA et al Mutations ofephrin-B1 (EFNB1) a marker of tissue boundary formation cause craniofronto-nasal syndrome Proc Natl Acad Sci USA 20041018652ndash7
[35] Gronthos S Simmons PJ The growth factor requirements of STRO-1-positivehuman bone marrow stromal precursors under serum-deprived conditions invitro Blood 199585929ndash40
[36] Pittenger MF Mackay AM Beck SC Jaiswal RK Douglas R Mosca JD et alMultilineage potential of adult human mesenchymal stem cells Science1999284143ndash7
[37] Gronthos S Zannettino AC Hay SJ Shi S Graves SE Kortesidis A et al Molecularand cellular characterisation of highly purified stromal stem cells derived fromhuman bone marrow J Cell Sci 20031161827ndash35
[38] Salvucci O Yao L Villalba S Sajewicz A Pittaluga S Tosato G Regulation ofendothelial cell branching morphogenesis by endogenous chemokine stromal-derived factor-1 Blood 2002992703ndash11
[39] Fitter S Dewar AL Kostakis P To LB Hughes TP Roberts MM et al Long-termimatinib therapy promotes bone formation in CML patients Blood 20081112538ndash47
[40] Isenmann S Arthur A Zannettino AC Turner JL Shi S Glackin CA et al TWISTfamily of basic helix-loop-helix transcription factors mediate human mesenchy-mal stem cell growth and commitment Stem Cells 2009272457ndash68
[41] Diamond P Labrinidis A Martin SK Farrugia AN Gronthos S To LB et al Targeteddisruption of the CXCL12CXCR4 axis inhibits osteolysis in a murine model ofmyeloma-associated bone loss J Bone Miner Res 2009241150ndash61
[42] Zannettino AC Paton S Arthur A Khor F Itescu S Gimble JM et al Multipotentialhuman adipose-derived stromal stem cells exhibit a perivascular phenotype invitro and in vivo J Cell Physiol 2008214413ndash21
[43] DeMali KA Burridge K Coupling membrane protrusion and cell adhesion J Cell Sci20031162389ndash97
[44] de Hoog CL Foster LJ Mann M RNA and RNA binding proteins participate in earlystages of cell spreading through spreading initiation centers Cell 2004117649ndash62
[45] Foo SS Turner CJ Adams S Compagni A Aubyn D Kogata N et al Ephrin-B2controls cell motility and adhesion during blood-vessel-wall assembly Cell2006124161ndash73
[46] Zhang J Hughes S Role of the ephrin and Eph receptor tyrosine kinase families inangiogenesis and development of the cardiovascular system J Pathol 2006208453ndash61
[47] Arthur A Rychkov G Shi S Koblar SA Gronthos S Adult human dental pulp stemcells differentiate toward functionally active neurons under appropriate environ-mental cues Stem Cells 2008261787ndash95
[48] Holmberg J Genander M Halford MM Anneren C Sondell M Chumley MJ et alEphB receptors coordinate migration and proliferation in the intestinal stem cellniche Cell 20061251151ndash63
542 A Arthur et al Bone 48 (2011) 533ndash542
[49] Holmberg J Armulik A Senti KA Edoff K Spalding K Momma S et al Ephrin-A2reverse signaling negatively regulates neural progenitor proliferation andneurogenesis Genes Dev 200519462ndash71
[50] Xu Z Lai KO Zhou HM Lin SC Ip NY Ephrin-B1 reverse signaling activates JNKthrough a novel mechanism that is independent of tyrosine phosphorylation J BiolChem 200327824767ndash75
[51] Huynh-Do U Vindis C Liu H Cerretti DP McGrew JT Enriquez M et al Ephrin-B1transduces signals to activate integrin-mediated migration attachment andangiogenesis J Cell Sci 20021153073ndash81
[52] Koolpe M Burgess R Dail M Pasquale EB EphB receptor-binding peptidesidentified by phage display enable design of an antagonist with ephrin-likeaffinity J Biol Chem 200528017301ndash11
[53] Chrencik JE Brooun A Recht MI Nicola G Davis LK Abagyan R et al Three-dimensional structure of the EphB2 receptor in complex with an antagonis-tic peptide reveals a novel mode of inhibition J Biol Chem 200728236505ndash13
[54] Noren NK Lu M Freeman AL Koolpe M Pasquale EB Interplay between EphB4 ontumor cells and vascular ephrin-B2 regulates tumor growth Proc Natl Acad SciUSA 20041015583ndash8
[55] Himanen JP Chumley MJ LackmannM Li C BartonWA Jeffrey PD et al Repellingclass discrimination ephrin-A5 binds to and activates EphB2 receptor signalingNat Neurosci 20047501ndash9
[56] Himanen JP Rajashankar KR Lackmann M Cowan CA Henkemeyer M NikolovDB Crystal structure of an Eph receptor-ephrin complex Nature 2001414933ndash8
[57] Chrencik JE BroounA RechtMI KrausML KoolpeM Kolatkar AR et al Structure andthermodynamic characterization of the EphB4Ephrin-B2 antagonist peptidecomplex reveals thedeterminants for receptor specificity Structure 200614321ndash30
[58] Pasquale EB The Eph family of receptors Curr Opin Cell Biol 19979608ndash15[59] Opperman LA Cranial sutures as intramembranous bone growth sites Dev Dyn
2000219472ndash85[60] Kronenberg HM Developmental regulation of the growth plate Nature 2003423
332ndash6[61] Kwan Tat S Pelletier JP Amiable N Boileau C Lajeunesse D Duval N et al
Activation of the receptor EphB4 by its specific ligand ephrin B2 in humanosteoarthritic subchondral bone osteoblasts Arthritis Rheum 2008583820ndash30
[62] Kwan Tat S Pelletier JP Amiable N Boileau C Lavigne M Martel-Pelletier JTreatment with ephrin B2 positively impacts the abnormal metabolism of humanosteoarthritic chondrocytes Arthritis Res Ther 200911R119
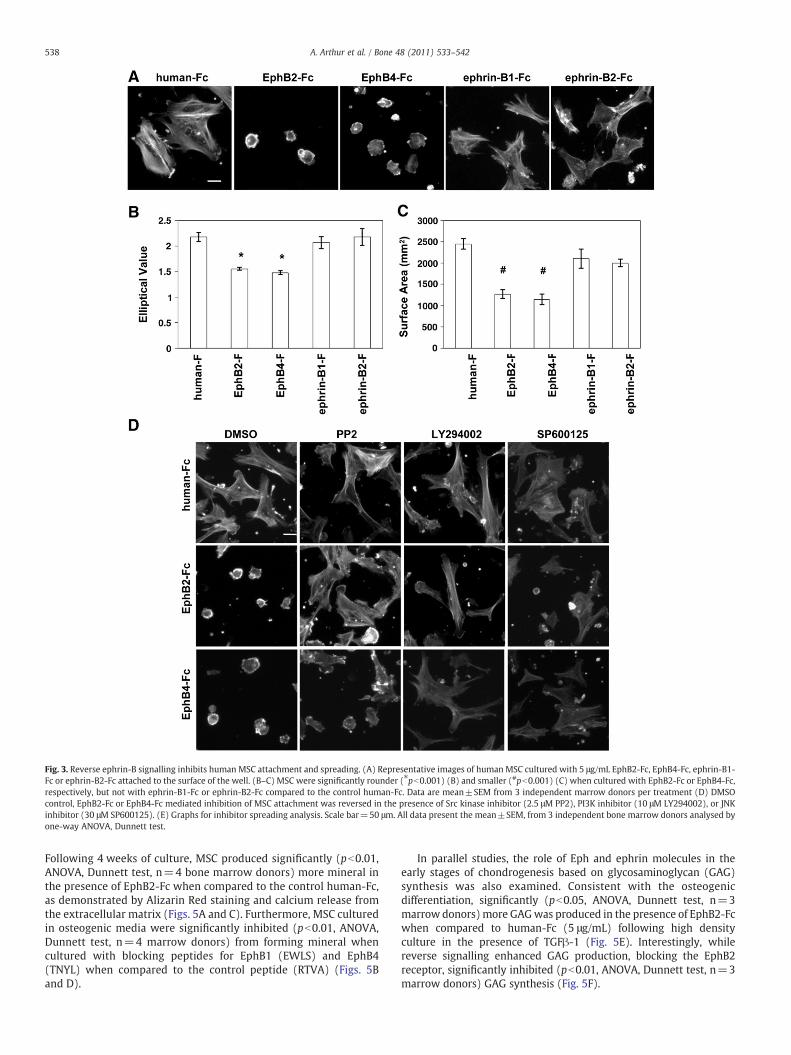
Fig 3 Reverse ephrin-B signalling inhibits human MSC attachment and spreading (A) Representative images of human MSC cultured with 5 μgmL EphB2-Fc EphB4-Fc ephrin-B1-Fc or ephrin-B2-Fc attached to the surface of the well (BndashC) MSC were significantly rounder (pb0001) (B) and smaller (pb0001) (C) when cultured with EphB2-Fc or EphB4-Fcrespectively but not with ephrin-B1-Fc or ephrin-B2-Fc compared to the control human-Fc Data are meanplusmnSEM from 3 independent marrow donors per treatment (D) DMSOcontrol EphB2-Fc or EphB4-Fc mediated inhibition of MSC attachment was reversed in the presence of Src kinase inhibitor (25 μM PP2) PI3K inhibitor (10 μM LY294002) or JNKinhibitor (30 μM SP600125) (E) Graphs for inhibitor spreading analysis Scale bar=50 μm All data present the meanplusmnSEM from 3 independent bone marrow donors analysed byone-way ANOVA Dunnett test
538 A Arthur et al Bone 48 (2011) 533ndash542
Following 4 weeks of culture MSC produced significantly (pb001ANOVA Dunnett test n=4 bone marrow donors) more mineral inthe presence of EphB2-Fc when compared to the control human-Fcas demonstrated by Alizarin Red staining and calcium release fromthe extracellular matrix (Figs 5A and C) Furthermore MSC culturedin osteogenic media were significantly inhibited (pb001 ANOVADunnett test n=4 marrow donors) from forming mineral whencultured with blocking peptides for EphB1 (EWLS) and EphB4(TNYL) when compared to the control peptide (RTVA) (Figs 5Band D)
In parallel studies the role of Eph and ephrin molecules in theearly stages of chondrogenesis based on glycosaminoglycan (GAG)synthesis was also examined Consistent with the osteogenicdifferentiation significantly (pb005 ANOVA Dunnett test n=3marrow donors)more GAGwas produced in the presence of EphB2-Fcwhen compared to human-Fc (5 μgmL) following high densityculture in the presence of TGFβ-1 (Fig 5E) Interestingly whilereverse signalling enhanced GAG production blocking the EphB2receptor significantly inhibited (pb001 ANOVA Dunnett test n=3marrow donors) GAG synthesis (Fig 5F)
Fig 4 Forward EphB signalling promotes MSC migration (A) Transwell migration assays demonstrated that human MSC significantly increased (+pb001 ANOVA Dunnett test)their migration in the presence of 5 μgmL ephrin-B1-Fc compared to control human-Fc (B) ephrin-B1-Fc mediated MSC migration (DMSO control) was significantly reduced(plusmnpb0001) in the presence of PP2 (10 μM) and (^pb001 ANOVA Dunnett test) the presence of Imatinib (3 μM) in contrast to MEKERK inhibitor (U0126) p38 MAPK inhibitorSB203580 (SB) and LY294002 (LY) (C) MSCmigration mediated by ephrin-B1 activation was significantly inhibited in a shRNA EphB2 knockdownMSC line in either the presence ofeither 5 μgmL human-Fc (p=00014 paired Student t-test) or ephrin-B2-Fc (p=000037 Student t-test) when compared to the shRNA scramble nonsilencing control MSC Alldata represent the meanplusmnSEM from 3 independent bone marrow donors Images represent nuclei stained with DAPI Image J software was used to count nuclei in five replicatefields per chamber Scale bar=50 μm
539A Arthur et al Bone 48 (2011) 533ndash542
Discussion
In the present study we demonstrated the functional significanceof Ephephrin-B molecules expressed by MSC in relation to cellularattachment migration and osteochondral differentiation Cell motilityrelies on the convergence of a number of molecular processesincluding the establishment of focal adhesions followed by theattachment of F-actin among other associated molecules This allowsthe extension of actin processes in the form of filopodia andlamellopodia which serve to lengthen the cell [4344] These eventscan be activated by a number of ECM chemotaxis and guidancemolecules The Eph family is well known for inhibiting cell motilityparticularly during the process of boundary formation of neural crestcells restricting veins and arteries and assisting in the maintenance ofneural intestinal and dental pulp stem cell niches [172145ndash49] Inthis study we showed that activated ephrin-B reverse signalling actsto inhibit human MSC attachment and spreading by potentially
restricting focal adhesion maturation as previously shown for dentalpulp stem cells [20] The EphBephrin-B interactions trigger a numberof signalling pathways to mediate functions such as cell adhesionmigration proliferation or differentiation We demonstrated thatephrin-B induced inhibition of attachment and spreading wasmediated in part by signalling through the Src PI3Kinase and theJNK pathways This was somewhat surprising since ephrin-B reversesignalling is thought to be mediated predominantly through the Srcmolecule Grb4 or the PDZ domain However it has been demon-strated that ephrin-B1 can also signal through the JNK pathwayindependently of tyrosine kinase phosphorylation [5051] Converse-ly EphB2 forward signalling stimulated via interactions with ephrin-B1 promoted MSC migration mediated through the Src and Abl-Crksignalling pathways Therefore EphBephrin-B interactions may playa role in the recruitment of MSC from their perivascular niche towardsareas of tissue damage via the activation of either the EphB receptorsor their respective ephrin-B ligands [43742]
Fig 5 Reverse signalling promotes osteochondral differentiation (AndashD) Human MSC were cultured in osteogenic conditions for four weeks in the presence of 5 μgmL EphB2-FcEphB4-Fc ephrin-B1-Fc ephrin-B2-Fc or control human-Fc (A C) MSC cultured in the presence of EphB2-Fc produced significantly more mineral (^pb001) as demonstrated withAlizarin Red staining and (C) calcium concentration when compared to human-Fc (B D) Human MSC mineral production as demonstrated by (B) Alizarin Red staining and(D) calcium concentration was significantly inhibited in the presence of either EWLS (p=001) or TNYL (^p=001) EphB1 and EphB4 blocking peptides respectively compared tocontrol peptide RTVA (E F) HumanMSC cultured in the presence of chondrogenic media and EphB and ephrin-B-Fc molecules demonstrated significantly more GAG synthesis whencultured in the presence of (E) EphB2-Fc (pb005) when compared to the human-Fc control and significantly less GAG production when cultured with EphB2 blocking peptideSNEW (pb001) compared to peptide control RTVA All data represent the meanplusmnSEM for 3ndash4 independent marrow donors analysed by one-way ANOVA Dunnett test
540 A Arthur et al Bone 48 (2011) 533ndash542
Previously published mouse models have reported that EphBforward signalling was important for mouse osteoblastic differenti-ation [910] Similarly we observed that mineralisation of humanMSCwas decreased in the presence of the blocking peptides for EphB1 andEphB4 However our observations also suggest that reverse signallingstimulated by EphB2-Fc appears to significantly increase mineralproduction of MSC cultured in osteo-inductive media While ephrin-B-Fc molecules failed to stimulate EphB forward induced osteogen-esis addition of blocking peptides to EphB receptors inhibitedmineralproduction indicating that these blocking peptides may function notonly to block EphB forward signalling [5253] but may also inhibitreverse signalling Importantly the crystallographic studies havedemonstrated that the GndashH loop of the ephrin-B ligand physicallybinds to the ligand-binding cavity of the EphB receptor whichcoincides with the DndashE GndashH and JndashK loops resulting in eitherstabilised activation of EphB receptor or the ephrin-B ligand [54ndash57]Therefore it is plausible that by specifically blocking the ephrin-Bbinding cavity of EphB receptors these endogenous receptors may nolonger activate their cognate ephrin-B ligands While we cannotspecifically state which ligands are being blocked it could bepostulated based on binding affinity studies [58] that ephrin-B2may be predominantly required for osteoblastic differentiationHowever we cannot rule out the importance of ephrin-B1 based onthe transwells migration assays which showed that ephrin-B1activated EphB4 forward signalling Furthermore the findings ofephrin-B1 transgenic studies confirmed that reverse signallingmodulates the osteogenic response of bone marrow stromal cellsspecifically mediated through ephrin-B1 activation following stimu-lation with EphB2-Fc [33] The signalling mechanism required for theincrease in osteogenesis was mediated via PDZ signalling It appearsthat the formation of a PDZ complex containing NHERF1 PTPN13 andTAZ with ephrin-B1 results in the dephosphorylation of TAZ which
allows its translocation into the nucleus and subsequent activation ofosteogenic specific transcription factor Osterix [33]
Of note Zhao and colleagues also demonstrated high geneexpression of ephrin-B1 and ephrin-B2 by osteoblasts which werealso confirmed by Allan et al [910] However the functionalrelevance of this expression was not investigated in detail Further-more while Zhao and colleagues did use EphB4-Fc in their osteogenicassays these experiments did not demonstrate a significant differencein mineral formation [9] This finding is consistent with ourobservations as MSC produced more mineral only in response toEphB2-Fc As ephrin-B1 binds with highest affinity to EphB2 [58] itmay be assumed that MSC osteogenesis may be mediated by theEphB2ephrin-B1 receptor-ligand pairing however our blockingpeptide studies suggested that perhaps it may be mediated byephrin-B2 Our data show that EphB1 and EphB4 but not EphB2blocking peptides significantly reduce mineral formation in humanMSC where ephrin-B2 binds with highest affinity to EphB1 and EphB4[58] This observation is in line with the findings of Allan et al whoshowed that mouse Kusa 4b10 cells reduced mineral formation in thepresence of EphB4 blocking peptide TNYL at consistent concentra-tions used in the present study [10]
Alternatively the variation in EphBephrin-B signalling activatingMSC osteogenesis in this study may be attributed to the origin of theMSC used for each study The studies performed by Zhao et al andAllan et al utilized osteogenic cells derived from calvarial bone whichis formed by intramembranous ossification while the long bonesundergo predominantly endochondral ossification [5960] To accountfor these discrepancies we performed comparative studies usingbone marrow derived mouse MSC isolated from C57BL6 mousefemurs and found that these cells showed similar responses to theirhuman counterparts (unpublished observations) Furthermore theephrin-B1 transgenic studies also utilised stromal cells from the
541A Arthur et al Bone 48 (2011) 533ndash542
marrow rather than the calvarial bone identified similar findings tothe present study [33]
Studies were also performed to investigate the contribution of theEphephrin-B family in the early stages of chondrogenesis Similar tothe osteo-inductive assays we demonstrated that ephrin-B reversesignalling mediated by EphB2-Fc stimulated an increase in GAGproduction which could be inhibited by blocking the function ofendogenous EphB2 Again this observation would suggest that byblocking endogenous EphB2 ligand-binding and function may also beinhibited However unlike the mineralisation assay these findingssuggest that perhaps ephrin-B1 the highest binding affinity cognateligand for EphB2 [58] may be important for endochondral formationThis is a novel observation that the B-subclass Ephephrins may beinvolved in endochondral ossification following fracture and byhuman MSC To date the only studies to our knowledge investigatingthe B-subclass demonstrate that osteoarthritic subchondral bonemetabolism involves EphB4ephrin-B2 signalling The activation ofEphB4 by ephrin-B2 inhibits the expression of factors such as matrixmetalloproteinases interleukins and RANKL which result in reducedresorption activity leading to osteoarthritic subchondral bone [6162]Our study establishes the potential role of the EphBephrin-Bsignalling in human MSC attachmentspreading cell migration andchondrogenic and osteogenic differentiations
Supplementarymaterials related to this article can be found onlineat doi101016jbone201010180
Acknowledgments
This study was supported by a Robinson Institute Research Grantin Aid and NHMRC Project Grant 565176
References
[1] Sacchetti B Funari A Michienzi S Di Cesare S Piersanti S Saggio I et al Self-renewing osteoprogenitors in bone marrow sinusoids can organize a hematopoi-etic microenvironment Cell 2007131324ndash36
[2] Shi S Gronthos S Perivascular niche of postnatal mesenchymal stem cells inhuman bone marrow and dental pulp J Bone Miner Res 200318696ndash704
[3] Mankani MH Kuznetsov SA Robey PG Formation of hematopoietic territories andbone by transplanted human bone marrow stromal cells requires a critical celldensity Exp Hematol 200735995ndash1004
[4] Arthur A Zannettino A Gronthos S The therapeutic applications of multipotentialmesenchymalstromal stem cells in skeletal tissue repair J Cell Physiol 2008218237ndash45
[5] Komano Y Nanki T Hayashida K Taniguchi K Miyasaka N Identification of ahuman peripheral blood monocyte subset that differentiates into osteoclastsArthritis Res Ther 20068R152
[6] Karsenty G Role of Cbfa1 in osteoblast differentiation and function Semin Cell DevBiol 200011343ndash6
[7] Karsenty G Ducy P Starbuck M Priemel M Shen J Geoffroy V et al Cbfa1 as aregulator of osteoblast differentiation and function Bone 199925107ndash8
[8] Olsen BR Reginato AM Wang W Bone development Annu Rev Cell Dev Biol200016191ndash220
[9] Zhao C Irie N Takada Y Shimoda K Miyamoto T Nishiwaki T et al BidirectionalephrinB2-EphB4 signaling controls bone homeostasis Cell Metab 20064111ndash21
[10] Allan EH Hausler KD Wei T Gooi JH Quinn JM Crimeen-Irwin B et al EphrinB2regulation by PTH and PTHrP revealed by molecular profiling in differentiatingosteoblasts J Bone Miner Res 2008231170ndash81
[11] Irie N Takada Y Watanabe Y Matsuzaki Y Naruse C Asano M et al Bidirectionalsignaling through ephrinA2-EphA2 enhances osteoclastogenesis and suppressesosteoblastogenesis J Biol Chem 200928414637ndash44
[12] Cooke JE Moens CB Boundary formation in the hindbrain Eph only it weresimple Trends Neurosci 200225260ndash7
[13] Mellitzer G Xu Q Wilkinson DG Eph receptors and ephrins restrict cellintermingling and communication Nature 199940077ndash81
[14] Jayasena CS Flood WD Koblar SA High EphA3 expressing ophthalmic trigeminalsensory axons are sensitive to ephrin-A5-Fc implications for lobe specific axonguidance Neuroscience 200513597ndash109
[15] Koblar SA Krull CE Pasquale EB McLennan R Peale FD Cerretti DP et al Spinalmotor axons and neural crest cells use different molecular guides for segmentalmigration through the rostral half-somite J Neurobiol 200042437ndash47
[16] Chumley MJ Catchpole T Silvany RE Kernie SG Henkemeyer M EphB receptorsregulate stemprogenitor cell proliferation migration and polarity duringhippocampal neurogenesis J Neurosci 20072713481ndash90
[17] Merrill AE Bochukova EG Brugger SM Ishii M Pilz DTWall SA et al Cell mixing ata neural crest-mesoderm boundary and deficient ephrin-Eph signaling in thepathogenesis of craniosynostosis Hum Mol Genet 2006151319ndash28
[18] Sakai D Wakamatsu Y Regulatory mechanisms for neural crest formation CellsTissues Organs 200517924ndash35
[19] Okubo T Yanai N Obinata M Stromal cells modulate ephrinB2 expression andtransmigration of hematopoietic cells Exp Hematol 200634330ndash8
[20] Stokowski A Shi S Sun T Bartold PM Koblar SA Gronthos S EphBephrin-Binteraction mediates adult stem cell attachment spreading and migrationimplications for dental tissue repair Stem Cells 200725156ndash64
[21] Arthur A Koblar S Shi S Gronthos S EphephrinB mediate dental pulp stem cellmobilization and function J Dent Res 200988829ndash34
[22] Merlos-Suarez A Batlle E Eph-ephrin signalling in adult tissues and cancer CurrOpin Cell Biol 200820194ndash200
[23] Moore R Larue L Cell surface molecules and truncal neural crest ontogeny aperspective Birth Defects Res C Embryo Today 200472140ndash50
[24] Picco V Hudson C Yasuo H EphrinndashEph signalling drives the asymmetric division ofnotochordneural precursors in Ciona embryos Development 20071341491ndash7
[25] Sancho E Batlle E Clevers H Live and let die in the intestinal epithelium Curr OpinCell Biol 200315763ndash70
[26] Wang Z Cohen K Shao Y Mole P Dombkowski D Scadden DT Ephrin receptorEphB4 regulates ES cell differentiation of primitive mammalian hemangioblastsblood cardiomyocytes and blood vessels Blood 2004103100ndash9
[27] Compagni A Logan M Klein R Adams RH Control of skeletal patterning byephrinB1-EphB interactions Dev Cell 20035217ndash30
[28] Davy A Aubin J Soriano P Ephrin-B1 forward and reverse signaling are requiredduring mouse development Genes Dev 200418572ndash83
[29] Wieland I Jakubiczka S Muschke P CohenM Thiele H Gerlach KL et al Mutationsof the ephrin-B1 gene cause craniofrontonasal syndrome Am J Hum Genet2004741209ndash15
[30] Ting MC Wu NL Roybal PG Sun J Liu L Yen Y et al EphA4 as an effector of Twist1in the guidance of osteogenic precursor cells during calvarial bone growth and incraniosynostosis Development 2009136855ndash64
[31] Varelias A Koblar SA Cowled PA Carter CD Clayer M Human osteosarcomaexpresses specific ephrin profiles implications for tumorigenicity and prognosisCancer 200295862ndash9
[32] Xing W Baylink D Kesavan C Hu Y Kapoor S Chadwick RB et al Global geneexpression analysis in the bones reveals involvement of several novel genes andpathways in mediating an anabolic response of mechanical loading in mice J CellBiochem 2005961049ndash60
[33] Xing W Kim J Wergedal J Chen ST Mohan S Ephrin B1 regulates bone marrowstromal cell differentiation and bone formation by influencing TAZ transactivationvia complex formation with NHERF1 Mol Cell Biol 201030(3)711ndash21
[34] Twigg SR Kan R Babbs C Bochukova EG Robertson SP Wall SA et al Mutations ofephrin-B1 (EFNB1) a marker of tissue boundary formation cause craniofronto-nasal syndrome Proc Natl Acad Sci USA 20041018652ndash7
[35] Gronthos S Simmons PJ The growth factor requirements of STRO-1-positivehuman bone marrow stromal precursors under serum-deprived conditions invitro Blood 199585929ndash40
[36] Pittenger MF Mackay AM Beck SC Jaiswal RK Douglas R Mosca JD et alMultilineage potential of adult human mesenchymal stem cells Science1999284143ndash7
[37] Gronthos S Zannettino AC Hay SJ Shi S Graves SE Kortesidis A et al Molecularand cellular characterisation of highly purified stromal stem cells derived fromhuman bone marrow J Cell Sci 20031161827ndash35
[38] Salvucci O Yao L Villalba S Sajewicz A Pittaluga S Tosato G Regulation ofendothelial cell branching morphogenesis by endogenous chemokine stromal-derived factor-1 Blood 2002992703ndash11
[39] Fitter S Dewar AL Kostakis P To LB Hughes TP Roberts MM et al Long-termimatinib therapy promotes bone formation in CML patients Blood 20081112538ndash47
[40] Isenmann S Arthur A Zannettino AC Turner JL Shi S Glackin CA et al TWISTfamily of basic helix-loop-helix transcription factors mediate human mesenchy-mal stem cell growth and commitment Stem Cells 2009272457ndash68
[41] Diamond P Labrinidis A Martin SK Farrugia AN Gronthos S To LB et al Targeteddisruption of the CXCL12CXCR4 axis inhibits osteolysis in a murine model ofmyeloma-associated bone loss J Bone Miner Res 2009241150ndash61
[42] Zannettino AC Paton S Arthur A Khor F Itescu S Gimble JM et al Multipotentialhuman adipose-derived stromal stem cells exhibit a perivascular phenotype invitro and in vivo J Cell Physiol 2008214413ndash21
[43] DeMali KA Burridge K Coupling membrane protrusion and cell adhesion J Cell Sci20031162389ndash97
[44] de Hoog CL Foster LJ Mann M RNA and RNA binding proteins participate in earlystages of cell spreading through spreading initiation centers Cell 2004117649ndash62
[45] Foo SS Turner CJ Adams S Compagni A Aubyn D Kogata N et al Ephrin-B2controls cell motility and adhesion during blood-vessel-wall assembly Cell2006124161ndash73
[46] Zhang J Hughes S Role of the ephrin and Eph receptor tyrosine kinase families inangiogenesis and development of the cardiovascular system J Pathol 2006208453ndash61
[47] Arthur A Rychkov G Shi S Koblar SA Gronthos S Adult human dental pulp stemcells differentiate toward functionally active neurons under appropriate environ-mental cues Stem Cells 2008261787ndash95
[48] Holmberg J Genander M Halford MM Anneren C Sondell M Chumley MJ et alEphB receptors coordinate migration and proliferation in the intestinal stem cellniche Cell 20061251151ndash63
542 A Arthur et al Bone 48 (2011) 533ndash542
[49] Holmberg J Armulik A Senti KA Edoff K Spalding K Momma S et al Ephrin-A2reverse signaling negatively regulates neural progenitor proliferation andneurogenesis Genes Dev 200519462ndash71
[50] Xu Z Lai KO Zhou HM Lin SC Ip NY Ephrin-B1 reverse signaling activates JNKthrough a novel mechanism that is independent of tyrosine phosphorylation J BiolChem 200327824767ndash75
[51] Huynh-Do U Vindis C Liu H Cerretti DP McGrew JT Enriquez M et al Ephrin-B1transduces signals to activate integrin-mediated migration attachment andangiogenesis J Cell Sci 20021153073ndash81
[52] Koolpe M Burgess R Dail M Pasquale EB EphB receptor-binding peptidesidentified by phage display enable design of an antagonist with ephrin-likeaffinity J Biol Chem 200528017301ndash11
[53] Chrencik JE Brooun A Recht MI Nicola G Davis LK Abagyan R et al Three-dimensional structure of the EphB2 receptor in complex with an antagonis-tic peptide reveals a novel mode of inhibition J Biol Chem 200728236505ndash13
[54] Noren NK Lu M Freeman AL Koolpe M Pasquale EB Interplay between EphB4 ontumor cells and vascular ephrin-B2 regulates tumor growth Proc Natl Acad SciUSA 20041015583ndash8
[55] Himanen JP Chumley MJ LackmannM Li C BartonWA Jeffrey PD et al Repellingclass discrimination ephrin-A5 binds to and activates EphB2 receptor signalingNat Neurosci 20047501ndash9
[56] Himanen JP Rajashankar KR Lackmann M Cowan CA Henkemeyer M NikolovDB Crystal structure of an Eph receptor-ephrin complex Nature 2001414933ndash8
[57] Chrencik JE BroounA RechtMI KrausML KoolpeM Kolatkar AR et al Structure andthermodynamic characterization of the EphB4Ephrin-B2 antagonist peptidecomplex reveals thedeterminants for receptor specificity Structure 200614321ndash30
[58] Pasquale EB The Eph family of receptors Curr Opin Cell Biol 19979608ndash15[59] Opperman LA Cranial sutures as intramembranous bone growth sites Dev Dyn
2000219472ndash85[60] Kronenberg HM Developmental regulation of the growth plate Nature 2003423
332ndash6[61] Kwan Tat S Pelletier JP Amiable N Boileau C Lajeunesse D Duval N et al
Activation of the receptor EphB4 by its specific ligand ephrin B2 in humanosteoarthritic subchondral bone osteoblasts Arthritis Rheum 2008583820ndash30
[62] Kwan Tat S Pelletier JP Amiable N Boileau C Lavigne M Martel-Pelletier JTreatment with ephrin B2 positively impacts the abnormal metabolism of humanosteoarthritic chondrocytes Arthritis Res Ther 200911R119

Fig 4 Forward EphB signalling promotes MSC migration (A) Transwell migration assays demonstrated that human MSC significantly increased (+pb001 ANOVA Dunnett test)their migration in the presence of 5 μgmL ephrin-B1-Fc compared to control human-Fc (B) ephrin-B1-Fc mediated MSC migration (DMSO control) was significantly reduced(plusmnpb0001) in the presence of PP2 (10 μM) and (^pb001 ANOVA Dunnett test) the presence of Imatinib (3 μM) in contrast to MEKERK inhibitor (U0126) p38 MAPK inhibitorSB203580 (SB) and LY294002 (LY) (C) MSCmigration mediated by ephrin-B1 activation was significantly inhibited in a shRNA EphB2 knockdownMSC line in either the presence ofeither 5 μgmL human-Fc (p=00014 paired Student t-test) or ephrin-B2-Fc (p=000037 Student t-test) when compared to the shRNA scramble nonsilencing control MSC Alldata represent the meanplusmnSEM from 3 independent bone marrow donors Images represent nuclei stained with DAPI Image J software was used to count nuclei in five replicatefields per chamber Scale bar=50 μm
539A Arthur et al Bone 48 (2011) 533ndash542
Discussion
In the present study we demonstrated the functional significanceof Ephephrin-B molecules expressed by MSC in relation to cellularattachment migration and osteochondral differentiation Cell motilityrelies on the convergence of a number of molecular processesincluding the establishment of focal adhesions followed by theattachment of F-actin among other associated molecules This allowsthe extension of actin processes in the form of filopodia andlamellopodia which serve to lengthen the cell [4344] These eventscan be activated by a number of ECM chemotaxis and guidancemolecules The Eph family is well known for inhibiting cell motilityparticularly during the process of boundary formation of neural crestcells restricting veins and arteries and assisting in the maintenance ofneural intestinal and dental pulp stem cell niches [172145ndash49] Inthis study we showed that activated ephrin-B reverse signalling actsto inhibit human MSC attachment and spreading by potentially
restricting focal adhesion maturation as previously shown for dentalpulp stem cells [20] The EphBephrin-B interactions trigger a numberof signalling pathways to mediate functions such as cell adhesionmigration proliferation or differentiation We demonstrated thatephrin-B induced inhibition of attachment and spreading wasmediated in part by signalling through the Src PI3Kinase and theJNK pathways This was somewhat surprising since ephrin-B reversesignalling is thought to be mediated predominantly through the Srcmolecule Grb4 or the PDZ domain However it has been demon-strated that ephrin-B1 can also signal through the JNK pathwayindependently of tyrosine kinase phosphorylation [5051] Converse-ly EphB2 forward signalling stimulated via interactions with ephrin-B1 promoted MSC migration mediated through the Src and Abl-Crksignalling pathways Therefore EphBephrin-B interactions may playa role in the recruitment of MSC from their perivascular niche towardsareas of tissue damage via the activation of either the EphB receptorsor their respective ephrin-B ligands [43742]
Fig 5 Reverse signalling promotes osteochondral differentiation (AndashD) Human MSC were cultured in osteogenic conditions for four weeks in the presence of 5 μgmL EphB2-FcEphB4-Fc ephrin-B1-Fc ephrin-B2-Fc or control human-Fc (A C) MSC cultured in the presence of EphB2-Fc produced significantly more mineral (^pb001) as demonstrated withAlizarin Red staining and (C) calcium concentration when compared to human-Fc (B D) Human MSC mineral production as demonstrated by (B) Alizarin Red staining and(D) calcium concentration was significantly inhibited in the presence of either EWLS (p=001) or TNYL (^p=001) EphB1 and EphB4 blocking peptides respectively compared tocontrol peptide RTVA (E F) HumanMSC cultured in the presence of chondrogenic media and EphB and ephrin-B-Fc molecules demonstrated significantly more GAG synthesis whencultured in the presence of (E) EphB2-Fc (pb005) when compared to the human-Fc control and significantly less GAG production when cultured with EphB2 blocking peptideSNEW (pb001) compared to peptide control RTVA All data represent the meanplusmnSEM for 3ndash4 independent marrow donors analysed by one-way ANOVA Dunnett test
540 A Arthur et al Bone 48 (2011) 533ndash542
Previously published mouse models have reported that EphBforward signalling was important for mouse osteoblastic differenti-ation [910] Similarly we observed that mineralisation of humanMSCwas decreased in the presence of the blocking peptides for EphB1 andEphB4 However our observations also suggest that reverse signallingstimulated by EphB2-Fc appears to significantly increase mineralproduction of MSC cultured in osteo-inductive media While ephrin-B-Fc molecules failed to stimulate EphB forward induced osteogen-esis addition of blocking peptides to EphB receptors inhibitedmineralproduction indicating that these blocking peptides may function notonly to block EphB forward signalling [5253] but may also inhibitreverse signalling Importantly the crystallographic studies havedemonstrated that the GndashH loop of the ephrin-B ligand physicallybinds to the ligand-binding cavity of the EphB receptor whichcoincides with the DndashE GndashH and JndashK loops resulting in eitherstabilised activation of EphB receptor or the ephrin-B ligand [54ndash57]Therefore it is plausible that by specifically blocking the ephrin-Bbinding cavity of EphB receptors these endogenous receptors may nolonger activate their cognate ephrin-B ligands While we cannotspecifically state which ligands are being blocked it could bepostulated based on binding affinity studies [58] that ephrin-B2may be predominantly required for osteoblastic differentiationHowever we cannot rule out the importance of ephrin-B1 based onthe transwells migration assays which showed that ephrin-B1activated EphB4 forward signalling Furthermore the findings ofephrin-B1 transgenic studies confirmed that reverse signallingmodulates the osteogenic response of bone marrow stromal cellsspecifically mediated through ephrin-B1 activation following stimu-lation with EphB2-Fc [33] The signalling mechanism required for theincrease in osteogenesis was mediated via PDZ signalling It appearsthat the formation of a PDZ complex containing NHERF1 PTPN13 andTAZ with ephrin-B1 results in the dephosphorylation of TAZ which
allows its translocation into the nucleus and subsequent activation ofosteogenic specific transcription factor Osterix [33]
Of note Zhao and colleagues also demonstrated high geneexpression of ephrin-B1 and ephrin-B2 by osteoblasts which werealso confirmed by Allan et al [910] However the functionalrelevance of this expression was not investigated in detail Further-more while Zhao and colleagues did use EphB4-Fc in their osteogenicassays these experiments did not demonstrate a significant differencein mineral formation [9] This finding is consistent with ourobservations as MSC produced more mineral only in response toEphB2-Fc As ephrin-B1 binds with highest affinity to EphB2 [58] itmay be assumed that MSC osteogenesis may be mediated by theEphB2ephrin-B1 receptor-ligand pairing however our blockingpeptide studies suggested that perhaps it may be mediated byephrin-B2 Our data show that EphB1 and EphB4 but not EphB2blocking peptides significantly reduce mineral formation in humanMSC where ephrin-B2 binds with highest affinity to EphB1 and EphB4[58] This observation is in line with the findings of Allan et al whoshowed that mouse Kusa 4b10 cells reduced mineral formation in thepresence of EphB4 blocking peptide TNYL at consistent concentra-tions used in the present study [10]
Alternatively the variation in EphBephrin-B signalling activatingMSC osteogenesis in this study may be attributed to the origin of theMSC used for each study The studies performed by Zhao et al andAllan et al utilized osteogenic cells derived from calvarial bone whichis formed by intramembranous ossification while the long bonesundergo predominantly endochondral ossification [5960] To accountfor these discrepancies we performed comparative studies usingbone marrow derived mouse MSC isolated from C57BL6 mousefemurs and found that these cells showed similar responses to theirhuman counterparts (unpublished observations) Furthermore theephrin-B1 transgenic studies also utilised stromal cells from the
541A Arthur et al Bone 48 (2011) 533ndash542
marrow rather than the calvarial bone identified similar findings tothe present study [33]
Studies were also performed to investigate the contribution of theEphephrin-B family in the early stages of chondrogenesis Similar tothe osteo-inductive assays we demonstrated that ephrin-B reversesignalling mediated by EphB2-Fc stimulated an increase in GAGproduction which could be inhibited by blocking the function ofendogenous EphB2 Again this observation would suggest that byblocking endogenous EphB2 ligand-binding and function may also beinhibited However unlike the mineralisation assay these findingssuggest that perhaps ephrin-B1 the highest binding affinity cognateligand for EphB2 [58] may be important for endochondral formationThis is a novel observation that the B-subclass Ephephrins may beinvolved in endochondral ossification following fracture and byhuman MSC To date the only studies to our knowledge investigatingthe B-subclass demonstrate that osteoarthritic subchondral bonemetabolism involves EphB4ephrin-B2 signalling The activation ofEphB4 by ephrin-B2 inhibits the expression of factors such as matrixmetalloproteinases interleukins and RANKL which result in reducedresorption activity leading to osteoarthritic subchondral bone [6162]Our study establishes the potential role of the EphBephrin-Bsignalling in human MSC attachmentspreading cell migration andchondrogenic and osteogenic differentiations
Supplementarymaterials related to this article can be found onlineat doi101016jbone201010180
Acknowledgments
This study was supported by a Robinson Institute Research Grantin Aid and NHMRC Project Grant 565176
References
[1] Sacchetti B Funari A Michienzi S Di Cesare S Piersanti S Saggio I et al Self-renewing osteoprogenitors in bone marrow sinusoids can organize a hematopoi-etic microenvironment Cell 2007131324ndash36
[2] Shi S Gronthos S Perivascular niche of postnatal mesenchymal stem cells inhuman bone marrow and dental pulp J Bone Miner Res 200318696ndash704
[3] Mankani MH Kuznetsov SA Robey PG Formation of hematopoietic territories andbone by transplanted human bone marrow stromal cells requires a critical celldensity Exp Hematol 200735995ndash1004
[4] Arthur A Zannettino A Gronthos S The therapeutic applications of multipotentialmesenchymalstromal stem cells in skeletal tissue repair J Cell Physiol 2008218237ndash45
[5] Komano Y Nanki T Hayashida K Taniguchi K Miyasaka N Identification of ahuman peripheral blood monocyte subset that differentiates into osteoclastsArthritis Res Ther 20068R152
[6] Karsenty G Role of Cbfa1 in osteoblast differentiation and function Semin Cell DevBiol 200011343ndash6
[7] Karsenty G Ducy P Starbuck M Priemel M Shen J Geoffroy V et al Cbfa1 as aregulator of osteoblast differentiation and function Bone 199925107ndash8
[8] Olsen BR Reginato AM Wang W Bone development Annu Rev Cell Dev Biol200016191ndash220
[9] Zhao C Irie N Takada Y Shimoda K Miyamoto T Nishiwaki T et al BidirectionalephrinB2-EphB4 signaling controls bone homeostasis Cell Metab 20064111ndash21
[10] Allan EH Hausler KD Wei T Gooi JH Quinn JM Crimeen-Irwin B et al EphrinB2regulation by PTH and PTHrP revealed by molecular profiling in differentiatingosteoblasts J Bone Miner Res 2008231170ndash81
[11] Irie N Takada Y Watanabe Y Matsuzaki Y Naruse C Asano M et al Bidirectionalsignaling through ephrinA2-EphA2 enhances osteoclastogenesis and suppressesosteoblastogenesis J Biol Chem 200928414637ndash44
[12] Cooke JE Moens CB Boundary formation in the hindbrain Eph only it weresimple Trends Neurosci 200225260ndash7
[13] Mellitzer G Xu Q Wilkinson DG Eph receptors and ephrins restrict cellintermingling and communication Nature 199940077ndash81
[14] Jayasena CS Flood WD Koblar SA High EphA3 expressing ophthalmic trigeminalsensory axons are sensitive to ephrin-A5-Fc implications for lobe specific axonguidance Neuroscience 200513597ndash109
[15] Koblar SA Krull CE Pasquale EB McLennan R Peale FD Cerretti DP et al Spinalmotor axons and neural crest cells use different molecular guides for segmentalmigration through the rostral half-somite J Neurobiol 200042437ndash47
[16] Chumley MJ Catchpole T Silvany RE Kernie SG Henkemeyer M EphB receptorsregulate stemprogenitor cell proliferation migration and polarity duringhippocampal neurogenesis J Neurosci 20072713481ndash90
[17] Merrill AE Bochukova EG Brugger SM Ishii M Pilz DTWall SA et al Cell mixing ata neural crest-mesoderm boundary and deficient ephrin-Eph signaling in thepathogenesis of craniosynostosis Hum Mol Genet 2006151319ndash28
[18] Sakai D Wakamatsu Y Regulatory mechanisms for neural crest formation CellsTissues Organs 200517924ndash35
[19] Okubo T Yanai N Obinata M Stromal cells modulate ephrinB2 expression andtransmigration of hematopoietic cells Exp Hematol 200634330ndash8
[20] Stokowski A Shi S Sun T Bartold PM Koblar SA Gronthos S EphBephrin-Binteraction mediates adult stem cell attachment spreading and migrationimplications for dental tissue repair Stem Cells 200725156ndash64
[21] Arthur A Koblar S Shi S Gronthos S EphephrinB mediate dental pulp stem cellmobilization and function J Dent Res 200988829ndash34
[22] Merlos-Suarez A Batlle E Eph-ephrin signalling in adult tissues and cancer CurrOpin Cell Biol 200820194ndash200
[23] Moore R Larue L Cell surface molecules and truncal neural crest ontogeny aperspective Birth Defects Res C Embryo Today 200472140ndash50
[24] Picco V Hudson C Yasuo H EphrinndashEph signalling drives the asymmetric division ofnotochordneural precursors in Ciona embryos Development 20071341491ndash7
[25] Sancho E Batlle E Clevers H Live and let die in the intestinal epithelium Curr OpinCell Biol 200315763ndash70
[26] Wang Z Cohen K Shao Y Mole P Dombkowski D Scadden DT Ephrin receptorEphB4 regulates ES cell differentiation of primitive mammalian hemangioblastsblood cardiomyocytes and blood vessels Blood 2004103100ndash9
[27] Compagni A Logan M Klein R Adams RH Control of skeletal patterning byephrinB1-EphB interactions Dev Cell 20035217ndash30
[28] Davy A Aubin J Soriano P Ephrin-B1 forward and reverse signaling are requiredduring mouse development Genes Dev 200418572ndash83
[29] Wieland I Jakubiczka S Muschke P CohenM Thiele H Gerlach KL et al Mutationsof the ephrin-B1 gene cause craniofrontonasal syndrome Am J Hum Genet2004741209ndash15
[30] Ting MC Wu NL Roybal PG Sun J Liu L Yen Y et al EphA4 as an effector of Twist1in the guidance of osteogenic precursor cells during calvarial bone growth and incraniosynostosis Development 2009136855ndash64
[31] Varelias A Koblar SA Cowled PA Carter CD Clayer M Human osteosarcomaexpresses specific ephrin profiles implications for tumorigenicity and prognosisCancer 200295862ndash9
[32] Xing W Baylink D Kesavan C Hu Y Kapoor S Chadwick RB et al Global geneexpression analysis in the bones reveals involvement of several novel genes andpathways in mediating an anabolic response of mechanical loading in mice J CellBiochem 2005961049ndash60
[33] Xing W Kim J Wergedal J Chen ST Mohan S Ephrin B1 regulates bone marrowstromal cell differentiation and bone formation by influencing TAZ transactivationvia complex formation with NHERF1 Mol Cell Biol 201030(3)711ndash21
[34] Twigg SR Kan R Babbs C Bochukova EG Robertson SP Wall SA et al Mutations ofephrin-B1 (EFNB1) a marker of tissue boundary formation cause craniofronto-nasal syndrome Proc Natl Acad Sci USA 20041018652ndash7
[35] Gronthos S Simmons PJ The growth factor requirements of STRO-1-positivehuman bone marrow stromal precursors under serum-deprived conditions invitro Blood 199585929ndash40
[36] Pittenger MF Mackay AM Beck SC Jaiswal RK Douglas R Mosca JD et alMultilineage potential of adult human mesenchymal stem cells Science1999284143ndash7
[37] Gronthos S Zannettino AC Hay SJ Shi S Graves SE Kortesidis A et al Molecularand cellular characterisation of highly purified stromal stem cells derived fromhuman bone marrow J Cell Sci 20031161827ndash35
[38] Salvucci O Yao L Villalba S Sajewicz A Pittaluga S Tosato G Regulation ofendothelial cell branching morphogenesis by endogenous chemokine stromal-derived factor-1 Blood 2002992703ndash11
[39] Fitter S Dewar AL Kostakis P To LB Hughes TP Roberts MM et al Long-termimatinib therapy promotes bone formation in CML patients Blood 20081112538ndash47
[40] Isenmann S Arthur A Zannettino AC Turner JL Shi S Glackin CA et al TWISTfamily of basic helix-loop-helix transcription factors mediate human mesenchy-mal stem cell growth and commitment Stem Cells 2009272457ndash68
[41] Diamond P Labrinidis A Martin SK Farrugia AN Gronthos S To LB et al Targeteddisruption of the CXCL12CXCR4 axis inhibits osteolysis in a murine model ofmyeloma-associated bone loss J Bone Miner Res 2009241150ndash61
[42] Zannettino AC Paton S Arthur A Khor F Itescu S Gimble JM et al Multipotentialhuman adipose-derived stromal stem cells exhibit a perivascular phenotype invitro and in vivo J Cell Physiol 2008214413ndash21
[43] DeMali KA Burridge K Coupling membrane protrusion and cell adhesion J Cell Sci20031162389ndash97
[44] de Hoog CL Foster LJ Mann M RNA and RNA binding proteins participate in earlystages of cell spreading through spreading initiation centers Cell 2004117649ndash62
[45] Foo SS Turner CJ Adams S Compagni A Aubyn D Kogata N et al Ephrin-B2controls cell motility and adhesion during blood-vessel-wall assembly Cell2006124161ndash73
[46] Zhang J Hughes S Role of the ephrin and Eph receptor tyrosine kinase families inangiogenesis and development of the cardiovascular system J Pathol 2006208453ndash61
[47] Arthur A Rychkov G Shi S Koblar SA Gronthos S Adult human dental pulp stemcells differentiate toward functionally active neurons under appropriate environ-mental cues Stem Cells 2008261787ndash95
[48] Holmberg J Genander M Halford MM Anneren C Sondell M Chumley MJ et alEphB receptors coordinate migration and proliferation in the intestinal stem cellniche Cell 20061251151ndash63
542 A Arthur et al Bone 48 (2011) 533ndash542
[49] Holmberg J Armulik A Senti KA Edoff K Spalding K Momma S et al Ephrin-A2reverse signaling negatively regulates neural progenitor proliferation andneurogenesis Genes Dev 200519462ndash71
[50] Xu Z Lai KO Zhou HM Lin SC Ip NY Ephrin-B1 reverse signaling activates JNKthrough a novel mechanism that is independent of tyrosine phosphorylation J BiolChem 200327824767ndash75
[51] Huynh-Do U Vindis C Liu H Cerretti DP McGrew JT Enriquez M et al Ephrin-B1transduces signals to activate integrin-mediated migration attachment andangiogenesis J Cell Sci 20021153073ndash81
[52] Koolpe M Burgess R Dail M Pasquale EB EphB receptor-binding peptidesidentified by phage display enable design of an antagonist with ephrin-likeaffinity J Biol Chem 200528017301ndash11
[53] Chrencik JE Brooun A Recht MI Nicola G Davis LK Abagyan R et al Three-dimensional structure of the EphB2 receptor in complex with an antagonis-tic peptide reveals a novel mode of inhibition J Biol Chem 200728236505ndash13
[54] Noren NK Lu M Freeman AL Koolpe M Pasquale EB Interplay between EphB4 ontumor cells and vascular ephrin-B2 regulates tumor growth Proc Natl Acad SciUSA 20041015583ndash8
[55] Himanen JP Chumley MJ LackmannM Li C BartonWA Jeffrey PD et al Repellingclass discrimination ephrin-A5 binds to and activates EphB2 receptor signalingNat Neurosci 20047501ndash9
[56] Himanen JP Rajashankar KR Lackmann M Cowan CA Henkemeyer M NikolovDB Crystal structure of an Eph receptor-ephrin complex Nature 2001414933ndash8
[57] Chrencik JE BroounA RechtMI KrausML KoolpeM Kolatkar AR et al Structure andthermodynamic characterization of the EphB4Ephrin-B2 antagonist peptidecomplex reveals thedeterminants for receptor specificity Structure 200614321ndash30
[58] Pasquale EB The Eph family of receptors Curr Opin Cell Biol 19979608ndash15[59] Opperman LA Cranial sutures as intramembranous bone growth sites Dev Dyn
2000219472ndash85[60] Kronenberg HM Developmental regulation of the growth plate Nature 2003423
332ndash6[61] Kwan Tat S Pelletier JP Amiable N Boileau C Lajeunesse D Duval N et al
Activation of the receptor EphB4 by its specific ligand ephrin B2 in humanosteoarthritic subchondral bone osteoblasts Arthritis Rheum 2008583820ndash30
[62] Kwan Tat S Pelletier JP Amiable N Boileau C Lavigne M Martel-Pelletier JTreatment with ephrin B2 positively impacts the abnormal metabolism of humanosteoarthritic chondrocytes Arthritis Res Ther 200911R119

Fig 5 Reverse signalling promotes osteochondral differentiation (AndashD) Human MSC were cultured in osteogenic conditions for four weeks in the presence of 5 μgmL EphB2-FcEphB4-Fc ephrin-B1-Fc ephrin-B2-Fc or control human-Fc (A C) MSC cultured in the presence of EphB2-Fc produced significantly more mineral (^pb001) as demonstrated withAlizarin Red staining and (C) calcium concentration when compared to human-Fc (B D) Human MSC mineral production as demonstrated by (B) Alizarin Red staining and(D) calcium concentration was significantly inhibited in the presence of either EWLS (p=001) or TNYL (^p=001) EphB1 and EphB4 blocking peptides respectively compared tocontrol peptide RTVA (E F) HumanMSC cultured in the presence of chondrogenic media and EphB and ephrin-B-Fc molecules demonstrated significantly more GAG synthesis whencultured in the presence of (E) EphB2-Fc (pb005) when compared to the human-Fc control and significantly less GAG production when cultured with EphB2 blocking peptideSNEW (pb001) compared to peptide control RTVA All data represent the meanplusmnSEM for 3ndash4 independent marrow donors analysed by one-way ANOVA Dunnett test
540 A Arthur et al Bone 48 (2011) 533ndash542
Previously published mouse models have reported that EphBforward signalling was important for mouse osteoblastic differenti-ation [910] Similarly we observed that mineralisation of humanMSCwas decreased in the presence of the blocking peptides for EphB1 andEphB4 However our observations also suggest that reverse signallingstimulated by EphB2-Fc appears to significantly increase mineralproduction of MSC cultured in osteo-inductive media While ephrin-B-Fc molecules failed to stimulate EphB forward induced osteogen-esis addition of blocking peptides to EphB receptors inhibitedmineralproduction indicating that these blocking peptides may function notonly to block EphB forward signalling [5253] but may also inhibitreverse signalling Importantly the crystallographic studies havedemonstrated that the GndashH loop of the ephrin-B ligand physicallybinds to the ligand-binding cavity of the EphB receptor whichcoincides with the DndashE GndashH and JndashK loops resulting in eitherstabilised activation of EphB receptor or the ephrin-B ligand [54ndash57]Therefore it is plausible that by specifically blocking the ephrin-Bbinding cavity of EphB receptors these endogenous receptors may nolonger activate their cognate ephrin-B ligands While we cannotspecifically state which ligands are being blocked it could bepostulated based on binding affinity studies [58] that ephrin-B2may be predominantly required for osteoblastic differentiationHowever we cannot rule out the importance of ephrin-B1 based onthe transwells migration assays which showed that ephrin-B1activated EphB4 forward signalling Furthermore the findings ofephrin-B1 transgenic studies confirmed that reverse signallingmodulates the osteogenic response of bone marrow stromal cellsspecifically mediated through ephrin-B1 activation following stimu-lation with EphB2-Fc [33] The signalling mechanism required for theincrease in osteogenesis was mediated via PDZ signalling It appearsthat the formation of a PDZ complex containing NHERF1 PTPN13 andTAZ with ephrin-B1 results in the dephosphorylation of TAZ which
allows its translocation into the nucleus and subsequent activation ofosteogenic specific transcription factor Osterix [33]
Of note Zhao and colleagues also demonstrated high geneexpression of ephrin-B1 and ephrin-B2 by osteoblasts which werealso confirmed by Allan et al [910] However the functionalrelevance of this expression was not investigated in detail Further-more while Zhao and colleagues did use EphB4-Fc in their osteogenicassays these experiments did not demonstrate a significant differencein mineral formation [9] This finding is consistent with ourobservations as MSC produced more mineral only in response toEphB2-Fc As ephrin-B1 binds with highest affinity to EphB2 [58] itmay be assumed that MSC osteogenesis may be mediated by theEphB2ephrin-B1 receptor-ligand pairing however our blockingpeptide studies suggested that perhaps it may be mediated byephrin-B2 Our data show that EphB1 and EphB4 but not EphB2blocking peptides significantly reduce mineral formation in humanMSC where ephrin-B2 binds with highest affinity to EphB1 and EphB4[58] This observation is in line with the findings of Allan et al whoshowed that mouse Kusa 4b10 cells reduced mineral formation in thepresence of EphB4 blocking peptide TNYL at consistent concentra-tions used in the present study [10]
Alternatively the variation in EphBephrin-B signalling activatingMSC osteogenesis in this study may be attributed to the origin of theMSC used for each study The studies performed by Zhao et al andAllan et al utilized osteogenic cells derived from calvarial bone whichis formed by intramembranous ossification while the long bonesundergo predominantly endochondral ossification [5960] To accountfor these discrepancies we performed comparative studies usingbone marrow derived mouse MSC isolated from C57BL6 mousefemurs and found that these cells showed similar responses to theirhuman counterparts (unpublished observations) Furthermore theephrin-B1 transgenic studies also utilised stromal cells from the
541A Arthur et al Bone 48 (2011) 533ndash542
marrow rather than the calvarial bone identified similar findings tothe present study [33]
Studies were also performed to investigate the contribution of theEphephrin-B family in the early stages of chondrogenesis Similar tothe osteo-inductive assays we demonstrated that ephrin-B reversesignalling mediated by EphB2-Fc stimulated an increase in GAGproduction which could be inhibited by blocking the function ofendogenous EphB2 Again this observation would suggest that byblocking endogenous EphB2 ligand-binding and function may also beinhibited However unlike the mineralisation assay these findingssuggest that perhaps ephrin-B1 the highest binding affinity cognateligand for EphB2 [58] may be important for endochondral formationThis is a novel observation that the B-subclass Ephephrins may beinvolved in endochondral ossification following fracture and byhuman MSC To date the only studies to our knowledge investigatingthe B-subclass demonstrate that osteoarthritic subchondral bonemetabolism involves EphB4ephrin-B2 signalling The activation ofEphB4 by ephrin-B2 inhibits the expression of factors such as matrixmetalloproteinases interleukins and RANKL which result in reducedresorption activity leading to osteoarthritic subchondral bone [6162]Our study establishes the potential role of the EphBephrin-Bsignalling in human MSC attachmentspreading cell migration andchondrogenic and osteogenic differentiations
Supplementarymaterials related to this article can be found onlineat doi101016jbone201010180
Acknowledgments
This study was supported by a Robinson Institute Research Grantin Aid and NHMRC Project Grant 565176
References
[1] Sacchetti B Funari A Michienzi S Di Cesare S Piersanti S Saggio I et al Self-renewing osteoprogenitors in bone marrow sinusoids can organize a hematopoi-etic microenvironment Cell 2007131324ndash36
[2] Shi S Gronthos S Perivascular niche of postnatal mesenchymal stem cells inhuman bone marrow and dental pulp J Bone Miner Res 200318696ndash704
[3] Mankani MH Kuznetsov SA Robey PG Formation of hematopoietic territories andbone by transplanted human bone marrow stromal cells requires a critical celldensity Exp Hematol 200735995ndash1004
[4] Arthur A Zannettino A Gronthos S The therapeutic applications of multipotentialmesenchymalstromal stem cells in skeletal tissue repair J Cell Physiol 2008218237ndash45
[5] Komano Y Nanki T Hayashida K Taniguchi K Miyasaka N Identification of ahuman peripheral blood monocyte subset that differentiates into osteoclastsArthritis Res Ther 20068R152
[6] Karsenty G Role of Cbfa1 in osteoblast differentiation and function Semin Cell DevBiol 200011343ndash6
[7] Karsenty G Ducy P Starbuck M Priemel M Shen J Geoffroy V et al Cbfa1 as aregulator of osteoblast differentiation and function Bone 199925107ndash8
[8] Olsen BR Reginato AM Wang W Bone development Annu Rev Cell Dev Biol200016191ndash220
[9] Zhao C Irie N Takada Y Shimoda K Miyamoto T Nishiwaki T et al BidirectionalephrinB2-EphB4 signaling controls bone homeostasis Cell Metab 20064111ndash21
[10] Allan EH Hausler KD Wei T Gooi JH Quinn JM Crimeen-Irwin B et al EphrinB2regulation by PTH and PTHrP revealed by molecular profiling in differentiatingosteoblasts J Bone Miner Res 2008231170ndash81
[11] Irie N Takada Y Watanabe Y Matsuzaki Y Naruse C Asano M et al Bidirectionalsignaling through ephrinA2-EphA2 enhances osteoclastogenesis and suppressesosteoblastogenesis J Biol Chem 200928414637ndash44
[12] Cooke JE Moens CB Boundary formation in the hindbrain Eph only it weresimple Trends Neurosci 200225260ndash7
[13] Mellitzer G Xu Q Wilkinson DG Eph receptors and ephrins restrict cellintermingling and communication Nature 199940077ndash81
[14] Jayasena CS Flood WD Koblar SA High EphA3 expressing ophthalmic trigeminalsensory axons are sensitive to ephrin-A5-Fc implications for lobe specific axonguidance Neuroscience 200513597ndash109
[15] Koblar SA Krull CE Pasquale EB McLennan R Peale FD Cerretti DP et al Spinalmotor axons and neural crest cells use different molecular guides for segmentalmigration through the rostral half-somite J Neurobiol 200042437ndash47
[16] Chumley MJ Catchpole T Silvany RE Kernie SG Henkemeyer M EphB receptorsregulate stemprogenitor cell proliferation migration and polarity duringhippocampal neurogenesis J Neurosci 20072713481ndash90
[17] Merrill AE Bochukova EG Brugger SM Ishii M Pilz DTWall SA et al Cell mixing ata neural crest-mesoderm boundary and deficient ephrin-Eph signaling in thepathogenesis of craniosynostosis Hum Mol Genet 2006151319ndash28
[18] Sakai D Wakamatsu Y Regulatory mechanisms for neural crest formation CellsTissues Organs 200517924ndash35
[19] Okubo T Yanai N Obinata M Stromal cells modulate ephrinB2 expression andtransmigration of hematopoietic cells Exp Hematol 200634330ndash8
[20] Stokowski A Shi S Sun T Bartold PM Koblar SA Gronthos S EphBephrin-Binteraction mediates adult stem cell attachment spreading and migrationimplications for dental tissue repair Stem Cells 200725156ndash64
[21] Arthur A Koblar S Shi S Gronthos S EphephrinB mediate dental pulp stem cellmobilization and function J Dent Res 200988829ndash34
[22] Merlos-Suarez A Batlle E Eph-ephrin signalling in adult tissues and cancer CurrOpin Cell Biol 200820194ndash200
[23] Moore R Larue L Cell surface molecules and truncal neural crest ontogeny aperspective Birth Defects Res C Embryo Today 200472140ndash50
[24] Picco V Hudson C Yasuo H EphrinndashEph signalling drives the asymmetric division ofnotochordneural precursors in Ciona embryos Development 20071341491ndash7
[25] Sancho E Batlle E Clevers H Live and let die in the intestinal epithelium Curr OpinCell Biol 200315763ndash70
[26] Wang Z Cohen K Shao Y Mole P Dombkowski D Scadden DT Ephrin receptorEphB4 regulates ES cell differentiation of primitive mammalian hemangioblastsblood cardiomyocytes and blood vessels Blood 2004103100ndash9
[27] Compagni A Logan M Klein R Adams RH Control of skeletal patterning byephrinB1-EphB interactions Dev Cell 20035217ndash30
[28] Davy A Aubin J Soriano P Ephrin-B1 forward and reverse signaling are requiredduring mouse development Genes Dev 200418572ndash83
[29] Wieland I Jakubiczka S Muschke P CohenM Thiele H Gerlach KL et al Mutationsof the ephrin-B1 gene cause craniofrontonasal syndrome Am J Hum Genet2004741209ndash15
[30] Ting MC Wu NL Roybal PG Sun J Liu L Yen Y et al EphA4 as an effector of Twist1in the guidance of osteogenic precursor cells during calvarial bone growth and incraniosynostosis Development 2009136855ndash64
[31] Varelias A Koblar SA Cowled PA Carter CD Clayer M Human osteosarcomaexpresses specific ephrin profiles implications for tumorigenicity and prognosisCancer 200295862ndash9
[32] Xing W Baylink D Kesavan C Hu Y Kapoor S Chadwick RB et al Global geneexpression analysis in the bones reveals involvement of several novel genes andpathways in mediating an anabolic response of mechanical loading in mice J CellBiochem 2005961049ndash60
[33] Xing W Kim J Wergedal J Chen ST Mohan S Ephrin B1 regulates bone marrowstromal cell differentiation and bone formation by influencing TAZ transactivationvia complex formation with NHERF1 Mol Cell Biol 201030(3)711ndash21
[34] Twigg SR Kan R Babbs C Bochukova EG Robertson SP Wall SA et al Mutations ofephrin-B1 (EFNB1) a marker of tissue boundary formation cause craniofronto-nasal syndrome Proc Natl Acad Sci USA 20041018652ndash7
[35] Gronthos S Simmons PJ The growth factor requirements of STRO-1-positivehuman bone marrow stromal precursors under serum-deprived conditions invitro Blood 199585929ndash40
[36] Pittenger MF Mackay AM Beck SC Jaiswal RK Douglas R Mosca JD et alMultilineage potential of adult human mesenchymal stem cells Science1999284143ndash7
[37] Gronthos S Zannettino AC Hay SJ Shi S Graves SE Kortesidis A et al Molecularand cellular characterisation of highly purified stromal stem cells derived fromhuman bone marrow J Cell Sci 20031161827ndash35
[38] Salvucci O Yao L Villalba S Sajewicz A Pittaluga S Tosato G Regulation ofendothelial cell branching morphogenesis by endogenous chemokine stromal-derived factor-1 Blood 2002992703ndash11
[39] Fitter S Dewar AL Kostakis P To LB Hughes TP Roberts MM et al Long-termimatinib therapy promotes bone formation in CML patients Blood 20081112538ndash47
[40] Isenmann S Arthur A Zannettino AC Turner JL Shi S Glackin CA et al TWISTfamily of basic helix-loop-helix transcription factors mediate human mesenchy-mal stem cell growth and commitment Stem Cells 2009272457ndash68
[41] Diamond P Labrinidis A Martin SK Farrugia AN Gronthos S To LB et al Targeteddisruption of the CXCL12CXCR4 axis inhibits osteolysis in a murine model ofmyeloma-associated bone loss J Bone Miner Res 2009241150ndash61
[42] Zannettino AC Paton S Arthur A Khor F Itescu S Gimble JM et al Multipotentialhuman adipose-derived stromal stem cells exhibit a perivascular phenotype invitro and in vivo J Cell Physiol 2008214413ndash21
[43] DeMali KA Burridge K Coupling membrane protrusion and cell adhesion J Cell Sci20031162389ndash97
[44] de Hoog CL Foster LJ Mann M RNA and RNA binding proteins participate in earlystages of cell spreading through spreading initiation centers Cell 2004117649ndash62
[45] Foo SS Turner CJ Adams S Compagni A Aubyn D Kogata N et al Ephrin-B2controls cell motility and adhesion during blood-vessel-wall assembly Cell2006124161ndash73
[46] Zhang J Hughes S Role of the ephrin and Eph receptor tyrosine kinase families inangiogenesis and development of the cardiovascular system J Pathol 2006208453ndash61
[47] Arthur A Rychkov G Shi S Koblar SA Gronthos S Adult human dental pulp stemcells differentiate toward functionally active neurons under appropriate environ-mental cues Stem Cells 2008261787ndash95
[48] Holmberg J Genander M Halford MM Anneren C Sondell M Chumley MJ et alEphB receptors coordinate migration and proliferation in the intestinal stem cellniche Cell 20061251151ndash63
542 A Arthur et al Bone 48 (2011) 533ndash542
[49] Holmberg J Armulik A Senti KA Edoff K Spalding K Momma S et al Ephrin-A2reverse signaling negatively regulates neural progenitor proliferation andneurogenesis Genes Dev 200519462ndash71
[50] Xu Z Lai KO Zhou HM Lin SC Ip NY Ephrin-B1 reverse signaling activates JNKthrough a novel mechanism that is independent of tyrosine phosphorylation J BiolChem 200327824767ndash75
[51] Huynh-Do U Vindis C Liu H Cerretti DP McGrew JT Enriquez M et al Ephrin-B1transduces signals to activate integrin-mediated migration attachment andangiogenesis J Cell Sci 20021153073ndash81
[52] Koolpe M Burgess R Dail M Pasquale EB EphB receptor-binding peptidesidentified by phage display enable design of an antagonist with ephrin-likeaffinity J Biol Chem 200528017301ndash11
[53] Chrencik JE Brooun A Recht MI Nicola G Davis LK Abagyan R et al Three-dimensional structure of the EphB2 receptor in complex with an antagonis-tic peptide reveals a novel mode of inhibition J Biol Chem 200728236505ndash13
[54] Noren NK Lu M Freeman AL Koolpe M Pasquale EB Interplay between EphB4 ontumor cells and vascular ephrin-B2 regulates tumor growth Proc Natl Acad SciUSA 20041015583ndash8
[55] Himanen JP Chumley MJ LackmannM Li C BartonWA Jeffrey PD et al Repellingclass discrimination ephrin-A5 binds to and activates EphB2 receptor signalingNat Neurosci 20047501ndash9
[56] Himanen JP Rajashankar KR Lackmann M Cowan CA Henkemeyer M NikolovDB Crystal structure of an Eph receptor-ephrin complex Nature 2001414933ndash8
[57] Chrencik JE BroounA RechtMI KrausML KoolpeM Kolatkar AR et al Structure andthermodynamic characterization of the EphB4Ephrin-B2 antagonist peptidecomplex reveals thedeterminants for receptor specificity Structure 200614321ndash30
[58] Pasquale EB The Eph family of receptors Curr Opin Cell Biol 19979608ndash15[59] Opperman LA Cranial sutures as intramembranous bone growth sites Dev Dyn
2000219472ndash85[60] Kronenberg HM Developmental regulation of the growth plate Nature 2003423
332ndash6[61] Kwan Tat S Pelletier JP Amiable N Boileau C Lajeunesse D Duval N et al
Activation of the receptor EphB4 by its specific ligand ephrin B2 in humanosteoarthritic subchondral bone osteoblasts Arthritis Rheum 2008583820ndash30
[62] Kwan Tat S Pelletier JP Amiable N Boileau C Lavigne M Martel-Pelletier JTreatment with ephrin B2 positively impacts the abnormal metabolism of humanosteoarthritic chondrocytes Arthritis Res Ther 200911R119

541A Arthur et al Bone 48 (2011) 533ndash542
marrow rather than the calvarial bone identified similar findings tothe present study [33]
Studies were also performed to investigate the contribution of theEphephrin-B family in the early stages of chondrogenesis Similar tothe osteo-inductive assays we demonstrated that ephrin-B reversesignalling mediated by EphB2-Fc stimulated an increase in GAGproduction which could be inhibited by blocking the function ofendogenous EphB2 Again this observation would suggest that byblocking endogenous EphB2 ligand-binding and function may also beinhibited However unlike the mineralisation assay these findingssuggest that perhaps ephrin-B1 the highest binding affinity cognateligand for EphB2 [58] may be important for endochondral formationThis is a novel observation that the B-subclass Ephephrins may beinvolved in endochondral ossification following fracture and byhuman MSC To date the only studies to our knowledge investigatingthe B-subclass demonstrate that osteoarthritic subchondral bonemetabolism involves EphB4ephrin-B2 signalling The activation ofEphB4 by ephrin-B2 inhibits the expression of factors such as matrixmetalloproteinases interleukins and RANKL which result in reducedresorption activity leading to osteoarthritic subchondral bone [6162]Our study establishes the potential role of the EphBephrin-Bsignalling in human MSC attachmentspreading cell migration andchondrogenic and osteogenic differentiations
Supplementarymaterials related to this article can be found onlineat doi101016jbone201010180
Acknowledgments
This study was supported by a Robinson Institute Research Grantin Aid and NHMRC Project Grant 565176
References
[1] Sacchetti B Funari A Michienzi S Di Cesare S Piersanti S Saggio I et al Self-renewing osteoprogenitors in bone marrow sinusoids can organize a hematopoi-etic microenvironment Cell 2007131324ndash36
[2] Shi S Gronthos S Perivascular niche of postnatal mesenchymal stem cells inhuman bone marrow and dental pulp J Bone Miner Res 200318696ndash704
[3] Mankani MH Kuznetsov SA Robey PG Formation of hematopoietic territories andbone by transplanted human bone marrow stromal cells requires a critical celldensity Exp Hematol 200735995ndash1004
[4] Arthur A Zannettino A Gronthos S The therapeutic applications of multipotentialmesenchymalstromal stem cells in skeletal tissue repair J Cell Physiol 2008218237ndash45
[5] Komano Y Nanki T Hayashida K Taniguchi K Miyasaka N Identification of ahuman peripheral blood monocyte subset that differentiates into osteoclastsArthritis Res Ther 20068R152
[6] Karsenty G Role of Cbfa1 in osteoblast differentiation and function Semin Cell DevBiol 200011343ndash6
[7] Karsenty G Ducy P Starbuck M Priemel M Shen J Geoffroy V et al Cbfa1 as aregulator of osteoblast differentiation and function Bone 199925107ndash8
[8] Olsen BR Reginato AM Wang W Bone development Annu Rev Cell Dev Biol200016191ndash220
[9] Zhao C Irie N Takada Y Shimoda K Miyamoto T Nishiwaki T et al BidirectionalephrinB2-EphB4 signaling controls bone homeostasis Cell Metab 20064111ndash21
[10] Allan EH Hausler KD Wei T Gooi JH Quinn JM Crimeen-Irwin B et al EphrinB2regulation by PTH and PTHrP revealed by molecular profiling in differentiatingosteoblasts J Bone Miner Res 2008231170ndash81
[11] Irie N Takada Y Watanabe Y Matsuzaki Y Naruse C Asano M et al Bidirectionalsignaling through ephrinA2-EphA2 enhances osteoclastogenesis and suppressesosteoblastogenesis J Biol Chem 200928414637ndash44
[12] Cooke JE Moens CB Boundary formation in the hindbrain Eph only it weresimple Trends Neurosci 200225260ndash7
[13] Mellitzer G Xu Q Wilkinson DG Eph receptors and ephrins restrict cellintermingling and communication Nature 199940077ndash81
[14] Jayasena CS Flood WD Koblar SA High EphA3 expressing ophthalmic trigeminalsensory axons are sensitive to ephrin-A5-Fc implications for lobe specific axonguidance Neuroscience 200513597ndash109
[15] Koblar SA Krull CE Pasquale EB McLennan R Peale FD Cerretti DP et al Spinalmotor axons and neural crest cells use different molecular guides for segmentalmigration through the rostral half-somite J Neurobiol 200042437ndash47
[16] Chumley MJ Catchpole T Silvany RE Kernie SG Henkemeyer M EphB receptorsregulate stemprogenitor cell proliferation migration and polarity duringhippocampal neurogenesis J Neurosci 20072713481ndash90
[17] Merrill AE Bochukova EG Brugger SM Ishii M Pilz DTWall SA et al Cell mixing ata neural crest-mesoderm boundary and deficient ephrin-Eph signaling in thepathogenesis of craniosynostosis Hum Mol Genet 2006151319ndash28
[18] Sakai D Wakamatsu Y Regulatory mechanisms for neural crest formation CellsTissues Organs 200517924ndash35
[19] Okubo T Yanai N Obinata M Stromal cells modulate ephrinB2 expression andtransmigration of hematopoietic cells Exp Hematol 200634330ndash8
[20] Stokowski A Shi S Sun T Bartold PM Koblar SA Gronthos S EphBephrin-Binteraction mediates adult stem cell attachment spreading and migrationimplications for dental tissue repair Stem Cells 200725156ndash64
[21] Arthur A Koblar S Shi S Gronthos S EphephrinB mediate dental pulp stem cellmobilization and function J Dent Res 200988829ndash34
[22] Merlos-Suarez A Batlle E Eph-ephrin signalling in adult tissues and cancer CurrOpin Cell Biol 200820194ndash200
[23] Moore R Larue L Cell surface molecules and truncal neural crest ontogeny aperspective Birth Defects Res C Embryo Today 200472140ndash50
[24] Picco V Hudson C Yasuo H EphrinndashEph signalling drives the asymmetric division ofnotochordneural precursors in Ciona embryos Development 20071341491ndash7
[25] Sancho E Batlle E Clevers H Live and let die in the intestinal epithelium Curr OpinCell Biol 200315763ndash70
[26] Wang Z Cohen K Shao Y Mole P Dombkowski D Scadden DT Ephrin receptorEphB4 regulates ES cell differentiation of primitive mammalian hemangioblastsblood cardiomyocytes and blood vessels Blood 2004103100ndash9
[27] Compagni A Logan M Klein R Adams RH Control of skeletal patterning byephrinB1-EphB interactions Dev Cell 20035217ndash30
[28] Davy A Aubin J Soriano P Ephrin-B1 forward and reverse signaling are requiredduring mouse development Genes Dev 200418572ndash83
[29] Wieland I Jakubiczka S Muschke P CohenM Thiele H Gerlach KL et al Mutationsof the ephrin-B1 gene cause craniofrontonasal syndrome Am J Hum Genet2004741209ndash15
[30] Ting MC Wu NL Roybal PG Sun J Liu L Yen Y et al EphA4 as an effector of Twist1in the guidance of osteogenic precursor cells during calvarial bone growth and incraniosynostosis Development 2009136855ndash64
[31] Varelias A Koblar SA Cowled PA Carter CD Clayer M Human osteosarcomaexpresses specific ephrin profiles implications for tumorigenicity and prognosisCancer 200295862ndash9
[32] Xing W Baylink D Kesavan C Hu Y Kapoor S Chadwick RB et al Global geneexpression analysis in the bones reveals involvement of several novel genes andpathways in mediating an anabolic response of mechanical loading in mice J CellBiochem 2005961049ndash60
[33] Xing W Kim J Wergedal J Chen ST Mohan S Ephrin B1 regulates bone marrowstromal cell differentiation and bone formation by influencing TAZ transactivationvia complex formation with NHERF1 Mol Cell Biol 201030(3)711ndash21
[34] Twigg SR Kan R Babbs C Bochukova EG Robertson SP Wall SA et al Mutations ofephrin-B1 (EFNB1) a marker of tissue boundary formation cause craniofronto-nasal syndrome Proc Natl Acad Sci USA 20041018652ndash7
[35] Gronthos S Simmons PJ The growth factor requirements of STRO-1-positivehuman bone marrow stromal precursors under serum-deprived conditions invitro Blood 199585929ndash40
[36] Pittenger MF Mackay AM Beck SC Jaiswal RK Douglas R Mosca JD et alMultilineage potential of adult human mesenchymal stem cells Science1999284143ndash7
[37] Gronthos S Zannettino AC Hay SJ Shi S Graves SE Kortesidis A et al Molecularand cellular characterisation of highly purified stromal stem cells derived fromhuman bone marrow J Cell Sci 20031161827ndash35
[38] Salvucci O Yao L Villalba S Sajewicz A Pittaluga S Tosato G Regulation ofendothelial cell branching morphogenesis by endogenous chemokine stromal-derived factor-1 Blood 2002992703ndash11
[39] Fitter S Dewar AL Kostakis P To LB Hughes TP Roberts MM et al Long-termimatinib therapy promotes bone formation in CML patients Blood 20081112538ndash47
[40] Isenmann S Arthur A Zannettino AC Turner JL Shi S Glackin CA et al TWISTfamily of basic helix-loop-helix transcription factors mediate human mesenchy-mal stem cell growth and commitment Stem Cells 2009272457ndash68
[41] Diamond P Labrinidis A Martin SK Farrugia AN Gronthos S To LB et al Targeteddisruption of the CXCL12CXCR4 axis inhibits osteolysis in a murine model ofmyeloma-associated bone loss J Bone Miner Res 2009241150ndash61
[42] Zannettino AC Paton S Arthur A Khor F Itescu S Gimble JM et al Multipotentialhuman adipose-derived stromal stem cells exhibit a perivascular phenotype invitro and in vivo J Cell Physiol 2008214413ndash21
[43] DeMali KA Burridge K Coupling membrane protrusion and cell adhesion J Cell Sci20031162389ndash97
[44] de Hoog CL Foster LJ Mann M RNA and RNA binding proteins participate in earlystages of cell spreading through spreading initiation centers Cell 2004117649ndash62
[45] Foo SS Turner CJ Adams S Compagni A Aubyn D Kogata N et al Ephrin-B2controls cell motility and adhesion during blood-vessel-wall assembly Cell2006124161ndash73
[46] Zhang J Hughes S Role of the ephrin and Eph receptor tyrosine kinase families inangiogenesis and development of the cardiovascular system J Pathol 2006208453ndash61
[47] Arthur A Rychkov G Shi S Koblar SA Gronthos S Adult human dental pulp stemcells differentiate toward functionally active neurons under appropriate environ-mental cues Stem Cells 2008261787ndash95
[48] Holmberg J Genander M Halford MM Anneren C Sondell M Chumley MJ et alEphB receptors coordinate migration and proliferation in the intestinal stem cellniche Cell 20061251151ndash63
542 A Arthur et al Bone 48 (2011) 533ndash542
[49] Holmberg J Armulik A Senti KA Edoff K Spalding K Momma S et al Ephrin-A2reverse signaling negatively regulates neural progenitor proliferation andneurogenesis Genes Dev 200519462ndash71
[50] Xu Z Lai KO Zhou HM Lin SC Ip NY Ephrin-B1 reverse signaling activates JNKthrough a novel mechanism that is independent of tyrosine phosphorylation J BiolChem 200327824767ndash75
[51] Huynh-Do U Vindis C Liu H Cerretti DP McGrew JT Enriquez M et al Ephrin-B1transduces signals to activate integrin-mediated migration attachment andangiogenesis J Cell Sci 20021153073ndash81
[52] Koolpe M Burgess R Dail M Pasquale EB EphB receptor-binding peptidesidentified by phage display enable design of an antagonist with ephrin-likeaffinity J Biol Chem 200528017301ndash11
[53] Chrencik JE Brooun A Recht MI Nicola G Davis LK Abagyan R et al Three-dimensional structure of the EphB2 receptor in complex with an antagonis-tic peptide reveals a novel mode of inhibition J Biol Chem 200728236505ndash13
[54] Noren NK Lu M Freeman AL Koolpe M Pasquale EB Interplay between EphB4 ontumor cells and vascular ephrin-B2 regulates tumor growth Proc Natl Acad SciUSA 20041015583ndash8
[55] Himanen JP Chumley MJ LackmannM Li C BartonWA Jeffrey PD et al Repellingclass discrimination ephrin-A5 binds to and activates EphB2 receptor signalingNat Neurosci 20047501ndash9
[56] Himanen JP Rajashankar KR Lackmann M Cowan CA Henkemeyer M NikolovDB Crystal structure of an Eph receptor-ephrin complex Nature 2001414933ndash8
[57] Chrencik JE BroounA RechtMI KrausML KoolpeM Kolatkar AR et al Structure andthermodynamic characterization of the EphB4Ephrin-B2 antagonist peptidecomplex reveals thedeterminants for receptor specificity Structure 200614321ndash30
[58] Pasquale EB The Eph family of receptors Curr Opin Cell Biol 19979608ndash15[59] Opperman LA Cranial sutures as intramembranous bone growth sites Dev Dyn
2000219472ndash85[60] Kronenberg HM Developmental regulation of the growth plate Nature 2003423
332ndash6[61] Kwan Tat S Pelletier JP Amiable N Boileau C Lajeunesse D Duval N et al
Activation of the receptor EphB4 by its specific ligand ephrin B2 in humanosteoarthritic subchondral bone osteoblasts Arthritis Rheum 2008583820ndash30
[62] Kwan Tat S Pelletier JP Amiable N Boileau C Lavigne M Martel-Pelletier JTreatment with ephrin B2 positively impacts the abnormal metabolism of humanosteoarthritic chondrocytes Arthritis Res Ther 200911R119

542 A Arthur et al Bone 48 (2011) 533ndash542
[49] Holmberg J Armulik A Senti KA Edoff K Spalding K Momma S et al Ephrin-A2reverse signaling negatively regulates neural progenitor proliferation andneurogenesis Genes Dev 200519462ndash71
[50] Xu Z Lai KO Zhou HM Lin SC Ip NY Ephrin-B1 reverse signaling activates JNKthrough a novel mechanism that is independent of tyrosine phosphorylation J BiolChem 200327824767ndash75
[51] Huynh-Do U Vindis C Liu H Cerretti DP McGrew JT Enriquez M et al Ephrin-B1transduces signals to activate integrin-mediated migration attachment andangiogenesis J Cell Sci 20021153073ndash81
[52] Koolpe M Burgess R Dail M Pasquale EB EphB receptor-binding peptidesidentified by phage display enable design of an antagonist with ephrin-likeaffinity J Biol Chem 200528017301ndash11
[53] Chrencik JE Brooun A Recht MI Nicola G Davis LK Abagyan R et al Three-dimensional structure of the EphB2 receptor in complex with an antagonis-tic peptide reveals a novel mode of inhibition J Biol Chem 200728236505ndash13
[54] Noren NK Lu M Freeman AL Koolpe M Pasquale EB Interplay between EphB4 ontumor cells and vascular ephrin-B2 regulates tumor growth Proc Natl Acad SciUSA 20041015583ndash8
[55] Himanen JP Chumley MJ LackmannM Li C BartonWA Jeffrey PD et al Repellingclass discrimination ephrin-A5 binds to and activates EphB2 receptor signalingNat Neurosci 20047501ndash9
[56] Himanen JP Rajashankar KR Lackmann M Cowan CA Henkemeyer M NikolovDB Crystal structure of an Eph receptor-ephrin complex Nature 2001414933ndash8
[57] Chrencik JE BroounA RechtMI KrausML KoolpeM Kolatkar AR et al Structure andthermodynamic characterization of the EphB4Ephrin-B2 antagonist peptidecomplex reveals thedeterminants for receptor specificity Structure 200614321ndash30
[58] Pasquale EB The Eph family of receptors Curr Opin Cell Biol 19979608ndash15[59] Opperman LA Cranial sutures as intramembranous bone growth sites Dev Dyn
2000219472ndash85[60] Kronenberg HM Developmental regulation of the growth plate Nature 2003423
332ndash6[61] Kwan Tat S Pelletier JP Amiable N Boileau C Lajeunesse D Duval N et al
Activation of the receptor EphB4 by its specific ligand ephrin B2 in humanosteoarthritic subchondral bone osteoblasts Arthritis Rheum 2008583820ndash30
[62] Kwan Tat S Pelletier JP Amiable N Boileau C Lavigne M Martel-Pelletier JTreatment with ephrin B2 positively impacts the abnormal metabolism of humanosteoarthritic chondrocytes Arthritis Res Ther 200911R119
Copyright © 2022 FDOKUMEN




















