
Endocrine-committed progenitor cells retain their differentiation potential in the absence of...
6
Endocrine-committed progenitor cells retain their differentiation potential in the absence of neurogenin-3 expression Krishna Prasadan, Sidhartha Tulachan, Ping Guo, Chiyo Shiota, Sohail Shah, George Gittes * Division of Pediatric General and Thoracic Surgery, Children’s Hospital of University of Pittsburgh Medical Center, One Children’s Hospital Drive, 4401 Penn Ave., Rangos Research Center, Pittsburgh, PA 15224, United States article info Article history: Received 4 May 2010 Available online 13 May 2010 Keywords: Pancreas development Progenitor cells Neurogenin-3 SOX9 Duct cells abstract Neurogenin-3 (ngn-3) expression is critical for endocrine development in the developing pancreas. We found that when ngn-3 was inhibited in an E11.5 pancreas, using either morpholino antisense or siRNA, it led to a significant decrease in endocrine differentiation after seven days in culture. Endocrine differ- entiation was rescued when ngn-3 inhibition was withdrawn after three days of culture, suggesting that the embryonic pancreas retains progenitor cells with the ability to differentiate into endocrine cell types when ngn-3 expression recurs. To determine whether the rescue phenomenon observed after withdraw- ing ngn-3 antisense treatment was the result of the original endocrine-committed cells reinitiating endo- crine differentiation, or was instead due to new recruitment of later progenitor cells, we blocked ngn-3 expression for only the last four days of a seven-day culture. Here, insulin-positive differentiation was slightly reduced, but there was a normal number of glucagon-positive cells. In addition, there was an increase in SOX9-positive cells in ngn-3 inhibited, as well as in ngn-3 rescued pancreata, with a signifi- cant proportion of these SOX9-positive cells co-localized with DBA, an early ductal marker. This increased number of cells with co-localization of SOX9 and DBA could indicate an increased number of endocrine progenitor cells. Ó 2010 Elsevier Inc. All rights reserved. 1. Introduction The organogenesis of the pancreas is spatio-temporally con- trolled by the sequential activation of a specific cascade of tran- scription factors. The homeodomain protein pdx-1 expression in the pre-pancreatic primitive foregut at the 10–12 somite stage (E8.5) [1,2] is critical for proper pancreatic development [3]. Both mice and humans lacking pdx-1 are born without a pancreas [1,2]. Pdx-1 expression is followed by the expression of several lineage-regulating molecules including notch-1, neurogenin-3 (ngn-3) and pax-6. Notch-1 proteins regulate developmental lineage decisions. The activated notch-1 intracellular domain activates transcription of basic helix-loop-helix (bHLH) Hes genes [4], and in the pancreas, Hes-1 suppresses the expression of the pro-endocrine transcription factor ngn-3 [5]. Ngn-3 marks pancreatic cells destined for an endocrine fate [6,7]. This expression of notch-1 in the early pan- creas inhibits expression of neurogenins and prevents differentia- tion of early pancreatic progenitor cells to their endocrine fate [8]. Without notch-1 signaling, cells may prematurely choose a de- fault lineage pathway. Inhibiting the notch-1 pathway during very early pancreas development leads to premature development of endocrine tissue, with no branching and a complete lack of exo- crine tissue [8]. Transgenic over-expression of neurogenin-3 in mice, or ectopic expression of neurogenin-3 in chick embryo endo- dermal cells led to premature differentiation of only glucagon and somatostatin-producing cells [9]. Ngn-3 null mutant mice lack endocrine cells, and die postnatally between day 1 and 3 due to diabetes [8]. Thus, either inhibiting the notch-1 pathway or trans- genic over-expression of ngn-3 in the early pancreas both appear to result in premature commitment of pancreatic progenitor cells to an endocrine fate, depleting residual progenitor cells. These re- sults suggest that the spatio-temporal regulation of ngn-3 is a crit- ical determinant of pancreatic endocrine progenitor cell commitment [10]. Here, we show that blocking ngn-3 expression in the E11.5 embryonic pancreas results in reduced endocrine dif- ferentiation, no change in the number of amylase-positive acinar- cells, and an apparent augmentation in the pool of undifferentiated epithelial-ductal cells. We demonstrate that endocrine differentia- tion can be later rescued in this epithelial-ductal population by restoring ngn-3 expression, thus suggesting that endocrine-com- mitted progenitor cells in the developing pancreas may retain the ability to differentiate into endocrine cells. 0006-291X/$ - see front matter Ó 2010 Elsevier Inc. All rights reserved. doi:10.1016/j.bbrc.2010.05.058 * Corresponding author. E-mail addresses: [email protected] (K. Prasadan), George.gittes@ch- p.edu (G. Gittes). Biochemical and Biophysical Research Communications 396 (2010) 1036–1041 Contents lists available at ScienceDirect Biochemical and Biophysical Research Communications journal homepage: www.elsevier.com/locate/ybbrc
Transcript of Endocrine-committed progenitor cells retain their differentiation potential in the absence of...

Biochemical and Biophysical Research Communications 396 (2010) 1036–1041
Contents lists available at ScienceDirect
Biochemical and Biophysical Research Communications
journal homepage: www.elsevier .com/locate /ybbrc
Endocrine-committed progenitor cells retain their differentiation potentialin the absence of neurogenin-3 expression
Krishna Prasadan, Sidhartha Tulachan, Ping Guo, Chiyo Shiota, Sohail Shah, George Gittes *
Division of Pediatric General and Thoracic Surgery, Children’s Hospital of University of Pittsburgh Medical Center, One Children’s Hospital Drive, 4401 Penn Ave.,Rangos Research Center, Pittsburgh, PA 15224, United States
a r t i c l e i n f o
Article history:Received 4 May 2010Available online 13 May 2010
Keywords:Pancreas developmentProgenitor cellsNeurogenin-3SOX9Duct cells
0006-291X/$ - see front matter � 2010 Elsevier Inc. Adoi:10.1016/j.bbrc.2010.05.058
* Corresponding author.E-mail addresses: [email protected] (K.
p.edu (G. Gittes).
a b s t r a c t
Neurogenin-3 (ngn-3) expression is critical for endocrine development in the developing pancreas. Wefound that when ngn-3 was inhibited in an E11.5 pancreas, using either morpholino antisense or siRNA,it led to a significant decrease in endocrine differentiation after seven days in culture. Endocrine differ-entiation was rescued when ngn-3 inhibition was withdrawn after three days of culture, suggesting thatthe embryonic pancreas retains progenitor cells with the ability to differentiate into endocrine cell typeswhen ngn-3 expression recurs. To determine whether the rescue phenomenon observed after withdraw-ing ngn-3 antisense treatment was the result of the original endocrine-committed cells reinitiating endo-crine differentiation, or was instead due to new recruitment of later progenitor cells, we blocked ngn-3expression for only the last four days of a seven-day culture. Here, insulin-positive differentiation wasslightly reduced, but there was a normal number of glucagon-positive cells. In addition, there was anincrease in SOX9-positive cells in ngn-3 inhibited, as well as in ngn-3 rescued pancreata, with a signifi-cant proportion of these SOX9-positive cells co-localized with DBA, an early ductal marker. This increasednumber of cells with co-localization of SOX9 and DBA could indicate an increased number of endocrineprogenitor cells.
� 2010 Elsevier Inc. All rights reserved.
1. Introduction
The organogenesis of the pancreas is spatio-temporally con-trolled by the sequential activation of a specific cascade of tran-scription factors. The homeodomain protein pdx-1 expression inthe pre-pancreatic primitive foregut at the 10–12 somite stage(E8.5) [1,2] is critical for proper pancreatic development [3]. Bothmice and humans lacking pdx-1 are born without a pancreas[1,2]. Pdx-1 expression is followed by the expression of severallineage-regulating molecules including notch-1, neurogenin-3(ngn-3) and pax-6.
Notch-1 proteins regulate developmental lineage decisions. Theactivated notch-1 intracellular domain activates transcription ofbasic helix-loop-helix (bHLH) Hes genes [4], and in the pancreas,Hes-1 suppresses the expression of the pro-endocrine transcriptionfactor ngn-3 [5]. Ngn-3 marks pancreatic cells destined for anendocrine fate [6,7]. This expression of notch-1 in the early pan-creas inhibits expression of neurogenins and prevents differentia-tion of early pancreatic progenitor cells to their endocrine fate
ll rights reserved.
Prasadan), George.gittes@ch-
[8]. Without notch-1 signaling, cells may prematurely choose a de-fault lineage pathway. Inhibiting the notch-1 pathway during veryearly pancreas development leads to premature development ofendocrine tissue, with no branching and a complete lack of exo-crine tissue [8]. Transgenic over-expression of neurogenin-3 inmice, or ectopic expression of neurogenin-3 in chick embryo endo-dermal cells led to premature differentiation of only glucagon andsomatostatin-producing cells [9]. Ngn-3 null mutant mice lackendocrine cells, and die postnatally between day 1 and 3 due todiabetes [8]. Thus, either inhibiting the notch-1 pathway or trans-genic over-expression of ngn-3 in the early pancreas both appearto result in premature commitment of pancreatic progenitor cellsto an endocrine fate, depleting residual progenitor cells. These re-sults suggest that the spatio-temporal regulation of ngn-3 is a crit-ical determinant of pancreatic endocrine progenitor cellcommitment [10]. Here, we show that blocking ngn-3 expressionin the E11.5 embryonic pancreas results in reduced endocrine dif-ferentiation, no change in the number of amylase-positive acinar-cells, and an apparent augmentation in the pool of undifferentiatedepithelial-ductal cells. We demonstrate that endocrine differentia-tion can be later rescued in this epithelial-ductal population byrestoring ngn-3 expression, thus suggesting that endocrine-com-mitted progenitor cells in the developing pancreas may retainthe ability to differentiate into endocrine cells.
K. Prasadan et al. / Biochemical and Biophysical Research Communications 396 (2010) 1036–1041 1037
2. Materials and methods
We blocked ngn-3 expression in cultured pancreas using anantisense protocol described earlier [11]. The details of the re-agents and the methods are provided as Supplementary material.
3. Results
3.1. Endocrine differentiation in early embryonic pancreas requiresneurogenin-3 expression
Ngn-3 is a critical regulator of early endocrine commitmentduring pancreas development [12]. Ngn-3 expression is first seenin the developing pancreas around day 11.5 [13]. To identifywhether ngn-3 expression is necessary for further endocrinerecruitment after this initial commitment, we blocked ngn-3expression in vitro in freshly harvested day 11.5 embryonic pancre-
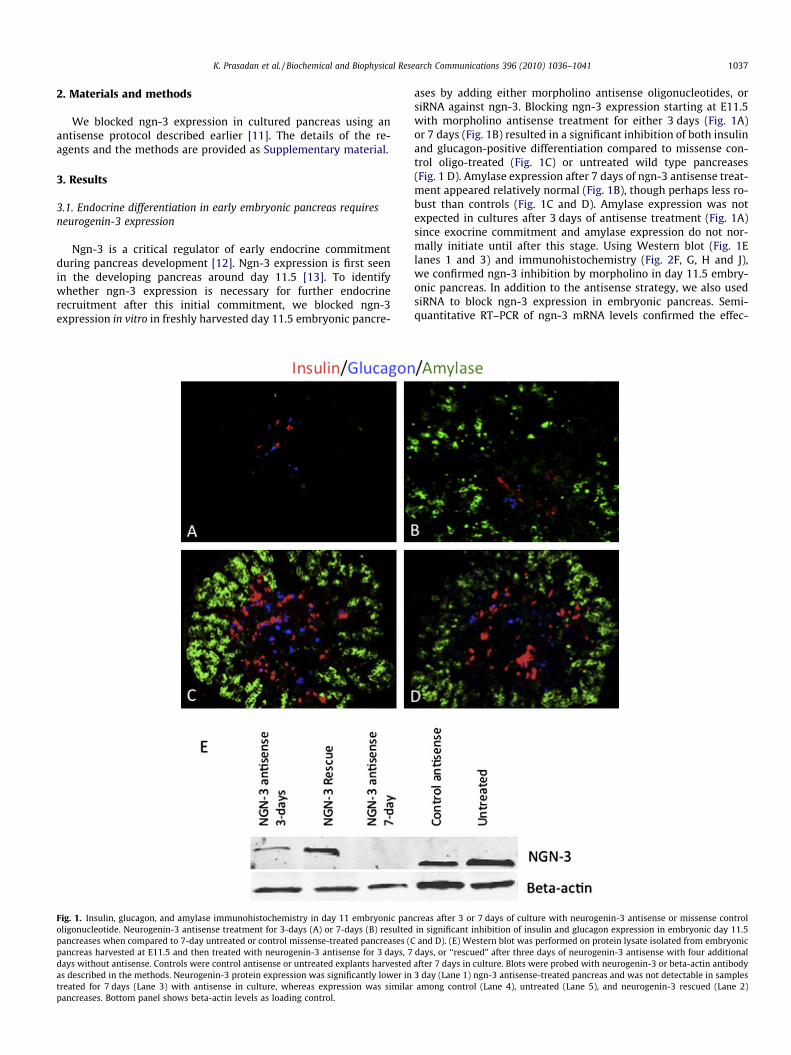
Fig. 1. Insulin, glucagon, and amylase immunohistochemistry in day 11 embryonic panoligonucleotide. Neurogenin-3 antisense treatment for 3-days (A) or 7-days (B) resultedpancreases when compared to 7-day untreated or control missense-treated pancreases (Cpancreas harvested at E11.5 and then treated with neurogenin-3 antisense for 3 days, 7days without antisense. Controls were control antisense or untreated explants harvestedas described in the methods. Neurogenin-3 protein expression was significantly lower intreated for 7 days (Lane 3) with antisense in culture, whereas expression was similarpancreases. Bottom panel shows beta-actin levels as loading control.
ases by adding either morpholino antisense oligonucleotides, orsiRNA against ngn-3. Blocking ngn-3 expression starting at E11.5with morpholino antisense treatment for either 3 days (Fig. 1A)or 7 days (Fig. 1B) resulted in a significant inhibition of both insulinand glucagon-positive differentiation compared to missense con-trol oligo-treated (Fig. 1C) or untreated wild type pancreases(Fig. 1 D). Amylase expression after 7 days of ngn-3 antisense treat-ment appeared relatively normal (Fig. 1B), though perhaps less ro-bust than controls (Fig. 1C and D). Amylase expression was notexpected in cultures after 3 days of antisense treatment (Fig. 1A)since exocrine commitment and amylase expression do not nor-mally initiate until after this stage. Using Western blot (Fig. 1Elanes 1 and 3) and immunohistochemistry (Fig. 2F, G, H and J),we confirmed ngn-3 inhibition by morpholino in day 11.5 embry-onic pancreas. In addition to the antisense strategy, we also usedsiRNA to block ngn-3 expression in embryonic pancreas. Semi-quantitative RT–PCR of ngn-3 mRNA levels confirmed the effec-
creas after 3 or 7 days of culture with neurogenin-3 antisense or missense controlin significant inhibition of insulin and glucagon expression in embryonic day 11.5and D). (E) Western blot was performed on protein lysate isolated from embryonic
days, or ‘‘rescued” after three days of neurogenin-3 antisense with four additionalafter 7 days in culture. Blots were probed with neurogenin-3 or beta-actin antibody3 day (Lane 1) ngn-3 antisense-treated pancreas and was not detectable in samplesamong control (Lane 4), untreated (Lane 5), and neurogenin-3 rescued (Lane 2)
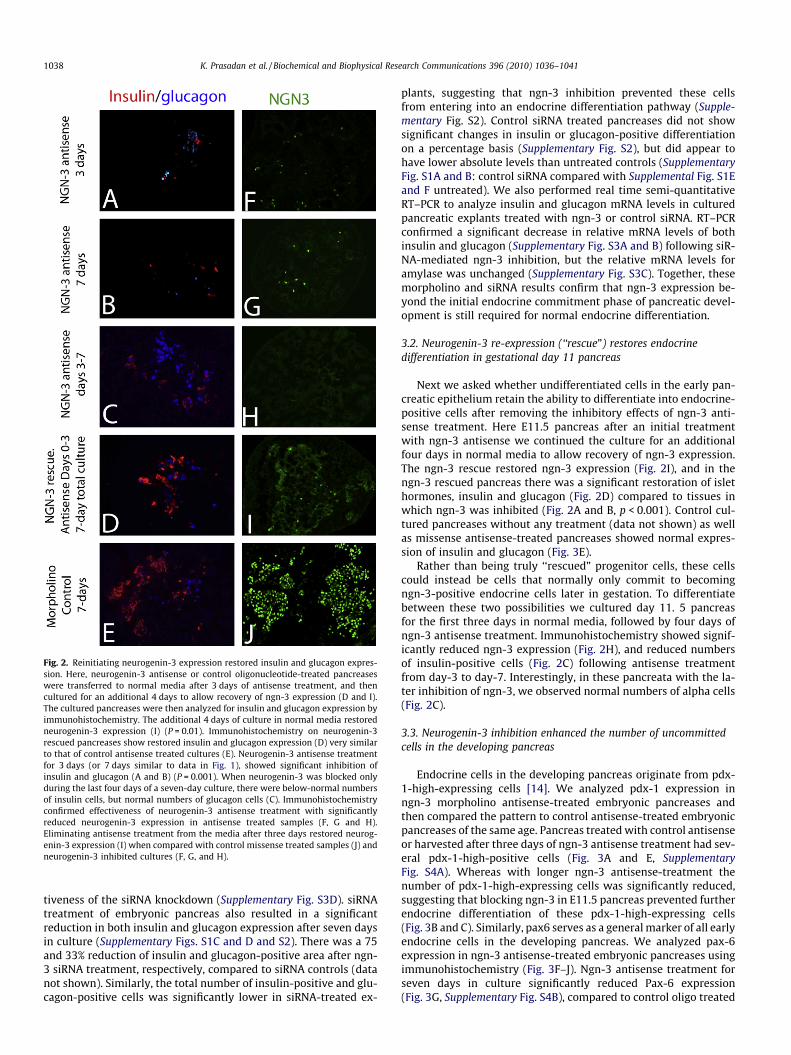
Fig. 2. Reinitiating neurogenin-3 expression restored insulin and glucagon expres-sion. Here, neurogenin-3 antisense or control oligonucleotide-treated pancreaseswere transferred to normal media after 3 days of antisense treatment, and thencultured for an additional 4 days to allow recovery of ngn-3 expression (D and I).The cultured pancreases were then analyzed for insulin and glucagon expression byimmunohistochemistry. The additional 4 days of culture in normal media restoredneurogenin-3 expression (I) (P = 0.01). Immunohistochemistry on neurogenin-3rescued pancreases show restored insulin and glucagon expression (D) very similarto that of control antisense treated cultures (E). Neurogenin-3 antisense treatmentfor 3 days (or 7 days similar to data in Fig. 1), showed significant inhibition ofinsulin and glucagon (A and B) (P = 0.001). When neurogenin-3 was blocked onlyduring the last four days of a seven-day culture, there were below-normal numbersof insulin cells, but normal numbers of glucagon cells (C). Immunohistochemistryconfirmed effectiveness of neurogenin-3 antisense treatment with significantlyreduced neurogenin-3 expression in antisense treated samples (F, G and H).Eliminating antisense treatment from the media after three days restored neurog-enin-3 expression (I) when compared with control missense treated samples (J) andneurogenin-3 inhibited cultures (F, G, and H).
1038 K. Prasadan et al. / Biochemical and Biophysical Research Communications 396 (2010) 1036–1041
tiveness of the siRNA knockdown (Supplementary Fig. S3D). siRNAtreatment of embryonic pancreas also resulted in a significantreduction in both insulin and glucagon expression after seven daysin culture (Supplementary Figs. S1C and D and S2). There was a 75and 33% reduction of insulin and glucagon-positive area after ngn-3 siRNA treatment, respectively, compared to siRNA controls (datanot shown). Similarly, the total number of insulin-positive and glu-cagon-positive cells was significantly lower in siRNA-treated ex-
plants, suggesting that ngn-3 inhibition prevented these cellsfrom entering into an endocrine differentiation pathway (Supple-mentary Fig. S2). Control siRNA treated pancreases did not showsignificant changes in insulin or glucagon-positive differentiationon a percentage basis (Supplementary Fig. S2), but did appear tohave lower absolute levels than untreated controls (SupplementaryFig. S1A and B: control siRNA compared with Supplemental Fig. S1Eand F untreated). We also performed real time semi-quantitativeRT–PCR to analyze insulin and glucagon mRNA levels in culturedpancreatic explants treated with ngn-3 or control siRNA. RT–PCRconfirmed a significant decrease in relative mRNA levels of bothinsulin and glucagon (Supplementary Fig. S3A and B) following siR-NA-mediated ngn-3 inhibition, but the relative mRNA levels foramylase was unchanged (Supplementary Fig. S3C). Together, thesemorpholino and siRNA results confirm that ngn-3 expression be-yond the initial endocrine commitment phase of pancreatic devel-opment is still required for normal endocrine differentiation.
3.2. Neurogenin-3 re-expression (‘‘rescue”) restores endocrinedifferentiation in gestational day 11 pancreas
Next we asked whether undifferentiated cells in the early pan-creatic epithelium retain the ability to differentiate into endocrine-positive cells after removing the inhibitory effects of ngn-3 anti-sense treatment. Here E11.5 pancreas after an initial treatmentwith ngn-3 antisense we continued the culture for an additionalfour days in normal media to allow recovery of ngn-3 expression.The ngn-3 rescue restored ngn-3 expression (Fig. 2I), and in thengn-3 rescued pancreas there was a significant restoration of islethormones, insulin and glucagon (Fig. 2D) compared to tissues inwhich ngn-3 was inhibited (Fig. 2A and B, p < 0.001). Control cul-tured pancreases without any treatment (data not shown) as wellas missense antisense-treated pancreases showed normal expres-sion of insulin and glucagon (Fig. 3E).
Rather than being truly ‘‘rescued” progenitor cells, these cellscould instead be cells that normally only commit to becomingngn-3-positive endocrine cells later in gestation. To differentiatebetween these two possibilities we cultured day 11. 5 pancreasfor the first three days in normal media, followed by four days ofngn-3 antisense treatment. Immunohistochemistry showed signif-icantly reduced ngn-3 expression (Fig. 2H), and reduced numbersof insulin-positive cells (Fig. 2C) following antisense treatmentfrom day-3 to day-7. Interestingly, in these pancreata with the la-ter inhibition of ngn-3, we observed normal numbers of alpha cells(Fig. 2C).
3.3. Neurogenin-3 inhibition enhanced the number of uncommittedcells in the developing pancreas
Endocrine cells in the developing pancreas originate from pdx-1-high-expressing cells [14]. We analyzed pdx-1 expression inngn-3 morpholino antisense-treated embryonic pancreases andthen compared the pattern to control antisense-treated embryonicpancreases of the same age. Pancreas treated with control antisenseor harvested after three days of ngn-3 antisense treatment had sev-eral pdx-1-high-positive cells (Fig. 3A and E, SupplementaryFig. S4A). Whereas with longer ngn-3 antisense-treatment thenumber of pdx-1-high-expressing cells was significantly reduced,suggesting that blocking ngn-3 in E11.5 pancreas prevented furtherendocrine differentiation of these pdx-1-high-expressing cells(Fig. 3B and C). Similarly, pax6 serves as a general marker of all earlyendocrine cells in the developing pancreas. We analyzed pax-6expression in ngn-3 antisense-treated embryonic pancreases usingimmunohistochemistry (Fig. 3F–J). Ngn-3 antisense treatment forseven days in culture significantly reduced Pax-6 expression(Fig. 3G, Supplementary Fig. S4B), compared to control oligo treated
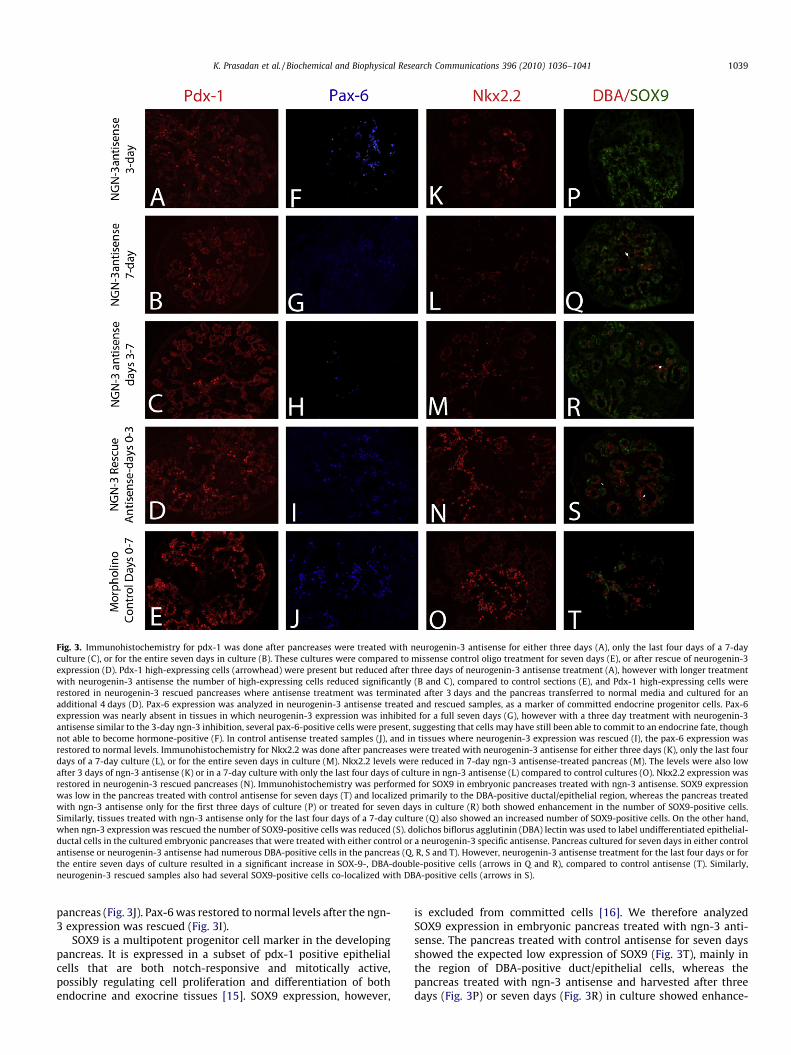
Fig. 3. Immunohistochemistry for pdx-1 was done after pancreases were treated with neurogenin-3 antisense for either three days (A), only the last four days of a 7-dayculture (C), or for the entire seven days in culture (B). These cultures were compared to missense control oligo treatment for seven days (E), or after rescue of neurogenin-3expression (D). Pdx-1 high-expressing cells (arrowhead) were present but reduced after three days of neurogenin-3 antisense treatment (A), however with longer treatmentwith neurogenin-3 antisense the number of high-expressing cells reduced significantly (B and C), compared to control sections (E), and Pdx-1 high-expressing cells wererestored in neurogenin-3 rescued pancreases where antisense treatment was terminated after 3 days and the pancreas transferred to normal media and cultured for anadditional 4 days (D). Pax-6 expression was analyzed in neurogenin-3 antisense treated and rescued samples, as a marker of committed endocrine progenitor cells. Pax-6expression was nearly absent in tissues in which neurogenin-3 expression was inhibited for a full seven days (G), however with a three day treatment with neurogenin-3antisense similar to the 3-day ngn-3 inhibition, several pax-6-positive cells were present, suggesting that cells may have still been able to commit to an endocrine fate, thoughnot able to become hormone-positive (F). In control antisense treated samples (J), and in tissues where neurogenin-3 expression was rescued (I), the pax-6 expression wasrestored to normal levels. Immunohistochemistry for Nkx2.2 was done after pancreases were treated with neurogenin-3 antisense for either three days (K), only the last fourdays of a 7-day culture (L), or for the entire seven days in culture (M). Nkx2.2 levels were reduced in 7-day ngn-3 antisense-treated pancreas (M). The levels were also lowafter 3 days of ngn-3 antisense (K) or in a 7-day culture with only the last four days of culture in ngn-3 antisense (L) compared to control cultures (O). Nkx2.2 expression wasrestored in neurogenin-3 rescued pancreases (N). Immunohistochemistry was performed for SOX9 in embryonic pancreases treated with ngn-3 antisense. SOX9 expressionwas low in the pancreas treated with control antisense for seven days (T) and localized primarily to the DBA-positive ductal/epithelial region, whereas the pancreas treatedwith ngn-3 antisense only for the first three days of culture (P) or treated for seven days in culture (R) both showed enhancement in the number of SOX9-positive cells.Similarly, tissues treated with ngn-3 antisense only for the last four days of a 7-day culture (Q) also showed an increased number of SOX9-positive cells. On the other hand,when ngn-3 expression was rescued the number of SOX9-positive cells was reduced (S). dolichos biflorus agglutinin (DBA) lectin was used to label undifferentiated epithelial-ductal cells in the cultured embryonic pancreases that were treated with either control or a neurogenin-3 specific antisense. Pancreas cultured for seven days in either controlantisense or neurogenin-3 antisense had numerous DBA-positive cells in the pancreas (Q, R, S and T). However, neurogenin-3 antisense treatment for the last four days or forthe entire seven days of culture resulted in a significant increase in SOX-9-, DBA-double-positive cells (arrows in Q and R), compared to control antisense (T). Similarly,neurogenin-3 rescued samples also had several SOX9-positive cells co-localized with DBA-positive cells (arrows in S).
K. Prasadan et al. / Biochemical and Biophysical Research Communications 396 (2010) 1036–1041 1039
pancreas (Fig. 3J). Pax-6 was restored to normal levels after the ngn-3 expression was rescued (Fig. 3I).
SOX9 is a multipotent progenitor cell marker in the developingpancreas. It is expressed in a subset of pdx-1 positive epithelialcells that are both notch-responsive and mitotically active,possibly regulating cell proliferation and differentiation of bothendocrine and exocrine tissues [15]. SOX9 expression, however,
is excluded from committed cells [16]. We therefore analyzedSOX9 expression in embryonic pancreas treated with ngn-3 anti-sense. The pancreas treated with control antisense for seven daysshowed the expected low expression of SOX9 (Fig. 3T), mainly inthe region of DBA-positive duct/epithelial cells, whereas thepancreas treated with ngn-3 antisense and harvested after threedays (Fig. 3P) or seven days (Fig. 3R) in culture showed enhance-

1040 K. Prasadan et al. / Biochemical and Biophysical Research Communications 396 (2010) 1036–1041
ment in the number of SOX9-positive cells (Fig. 3R, SupplementaryFig. S5B). Similarly, tissues treated with ngn-3 antisense only forthe last four days of culture (Fig. 3Q; Supplementary Fig. S5B) alsoshowed increased number of SOX9-positive cells. Tissues in whichthe ngn-3 expression was rescued had a reduced number of SOX9-positive cells (Fig. 3S; Supplementary Fig. S5B). In addition, ngn-3antisense treatment for three days followed by rescue of ngn-3expression resulted in a significant increase in Dolichos BiflorusAgglutinin-positive cells (DBA) (Fig. 3S and T), many of which werealso positive for SOX9 expression. A similar co-localization of DBA-positive cells and SOX9 were also found in pancreas treated withngn-3 antisense for the last four days of a seven-day culture(Fig. 3Q), as well as in 7-day antisense-treated pancreas (Fig. 3R).Thus, this increase in DBA-, and SOX9-positive cells in ngn-3 anti-sense treated and rescued tissues suggests an increase in anuncommitted epithelial-ductal cell population in the mid-gesta-tional pancreas in the absence of ngn-3 expression.
Nkx2.2 expression is required for specification alpha and betacells during pancreatic islet [17,18] and in its absence the progen-itor cells are converted to ghrelin-positive cells [19]. Nkx2.2expression decreased significantly in the pancreas treated withngn-3 antisense (Fig. 3K, L M and O; Supplementary Fig. S5A). Theeffect was most significant when treated for the entire durationof the seven-day culture (Fig. 3M, Supplemental Fig. S5A). However,a significant decrease in Nkx2.2 expression was still seen whentreated with the antisense for the last four days in culture(Fig. 3L; Supplementary Fig. S5A), as well as in explants culturedwith ngn-3 antisense for only three days (Fig. 3K; SupplementaryFig. S5A). When the ngn-3 expression was rescued we foundNkx2.2 expression at similar levels as control tissues (Fig. 3N, Sup-plementary Fig. S5A).
4. Discussion
The basic HLH transcription factor ngn-3 plays an importantrole during endocrine pancreas development [12,13,21]. Ngn-3has a bi-phasic expression during pancreas development: the firstphase is during E10–E10.5 and the second phase begins at E12.5[22]. In our study, to determine the role of ngn-3 during pancreasdevelopment beyond early time points, we analyzed the expres-sion of some key transcriptional regulators of pancreatic develop-ment, as well as the expression of islet hormones in thedeveloping pancreas after blocking ngn-3 expression well after ini-tiation of pancreatic organogenesis, at day 11.5.
Blocking ngn-3 expression in embryonic pancreases in vitro re-sulted in a significant inhibition of endocrine differentiation,however there was no effect on the number of amylase positivecells suggesting that after an initial endocrine commitment, these‘‘pre-endocrine” cells cannot be ‘‘reprogrammed” to become exo-crine cells. Lack of an effect of ngn-3 antisense on amylaseexpression, together with the demonstrated ability to rescue insu-lin and glucagon expression in pancreases previously treated withngn-3 antisense, suggest that these pancreatic progenitor cells arealready endocrine-committed. Thus, blocking ngn-3 in these cellsmay only prevent ngn-3-dependent endocrine differentiation. Weconfirmed this hypothesis (that cells retain a ngn-3-specific dif-ferentiation potential) when we rescued the insulin and gluca-gon-positive differentiation by restoring ngn-3 expressionfollowing a transient three-day inhibition of ngn-3 expression.This ability to recover endocrine differentiation suggests thatthe requirement for ngn-3 expression in endocrine differentiationpersists later into development. Specifically, endocrine progenitorcells at day 11.5 in the embryonic pancreas appear to retain theirendocrine differentiation potential for at least three days whenngn-3 expression is transiently inhibited. To determine whetherthese rescued progenitor cells may instead be arising from new
endocrine progenitor cells, not from rescued cells in which ngn-3 expression had been first inhibited, we blocked ngn-3 expres-sion only during the last four days of the culture, and then com-pared the results to cultures in which ngn-3 was inhibited onlyfor the first three days of culture. Here, instead of the completeinhibition of endocrine differentiation that we saw with inhibi-tion only for the first three days, there was instead a normalnumber of glucagon-positive cells when the ngn-3 expressionwas blocked for the final four days of culture. This presence ofa normal number of glucagon cells in a pancreas in which thengn-3 expression was blocked for the final four days of culturesuggests that the cells that initially expressed ngn-3, but wereinhibited by the antisense, had committed toward an endocrinefate, but further expression of neurogenin-3 was required for b-cell differentiation. A continued expression of ngn-3 for properb-cell differentiation, maturation, and function was also sug-gested else where [23]. In the absence of continued ngn-3 expres-sion, the committed cells may develop toward an alpha cell fate.A recent study found that pancreatic progenitor cells and alphacells adopted a beta cell fate after ectopic expression of pax-4,which in turn induced ngn-3 expression and alpha cell neogenesisfrom progenitor cells in the adult pancreas [24]. In our experi-ments, blocking ngn-3 expression early during the first three daysof culture inhibited endocrine differentiation. Inhibiting ngn-3expression later, only during the last four days of culture, pre-vented further expression of ngn-3 in committed cells, and pre-vented the formation of any new ngn-3-positive cells. Since wesaw no enhancement of alpha cells and normal insulin expressionin our rescued samples, we assume that the rescued cells are cellsin which ngn-3 expression was restored.
Furthermore ngn-3 inhibition in cultured embryonic pancreasesled to an increase in SOX9/DBA double-positive cells, markers ofundifferentiated progenitor cells and embryonic epithelial-ductalcells, respectively [20]. When ngn-3 was then rescued after the ini-tial blockage, the uncommitted progenitor cells in the ducts maycontinue to proliferate inappropriately, resulting in an increasednumber of SOX9/DBA-double-positive cells. This increase in pro-genitor cells (SOX9/DBA double-positive cells), with the subse-quent ability to recover the endocrine phenotype after recoveryof ngn-3 expression, supports the possibility that later-gestationalepithelium contains endocrine-committed progenitor cells that re-tain an endocrine differentiation potential later into development.
In the developing pancreas SOX9 maintains multipotent pro-genitors in an undifferentiated state by regulating cell proliferationand differentiation [15]. SOX9 does not co-localize with ngn-3, andis not expressed in differentiated cells, but is restricted to a subsetof pdx-1 positive cells that are both notch-responsive and mitoti-cally active [15]. In a transgenic mouse model in which pancreaticdifferentiation was blocked, enhanced expression of SOX9 wasfound throughout the tubular network of undifferentiated pdx-1-positive epithelial cells [15,25]. In our studies the increase inSOX9 expression, and the fact that several of these cells were alsoDBA-positive after ngn-3 inhibition, further suggests an increase inthe number of undifferentiated progenitor cells. Furthermore, sincewe did not see any significant change in exocrine development,and we were able to rescue the endocrine development, we believethat these SOX9/DBA double-positive cells are endocrine-commit-ted progenitor cells.
Acknowledgments
This work was supported by the NIH (GKG, RO1 DK064952, RO1DK083541-01), Pennsylvania State Tobacco Fund (GKG), and thefinancial support of Children’s Hospital of Pittsburgh. The authorswould like to thank all GKG lab members for their technical assis-tance and suggestions.

K. Prasadan et al. / Biochemical and Biophysical Research Communications 396 (2010) 1036–1041 1041
Appendix A. Supplementary material
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.bbrc.2010.05.058.
Reference
[1] U. Ahlgren, J. Jonsson, L. Jonsson, K. Simu, H. Edlund, Beta-cell-specificinactivation of the mouse Ipf1/Pdx1 gene results in loss of the beta-cellphenotype and maturity onset diabetes, Genes Dev. 12 (1998) 1763–1768.
[2] S.K. Kim, R.J. MacDonald, Signaling and transcriptional control of pancreaticorganogenesis, Curr. Opin. Genet. Dev. 12 (2002) 540–547.
[3] M.F. Offield, T.L. Jetton, P.A. Labosky, M. Ray, R.W. Stein, M.A. Magnuson, B.L.Hogan, C.V. Wright, PDX-1 is required for pancreatic outgrowth anddifferentiation of the rostral duodenum, Development 122 (1996) 983–995.
[4] S. Artavanis-Tsakonas, M.D. Rand, R.J. Lake, Notch signaling: cell fate controland signal integration in development, Science 284 (1999) 770–776.
[5] J. Jensen, E.E. Pedersen, P. Galante, J. Hald, R.S. Heller, M. Ishibashi, R.Kageyama, F. Guillemot, P. Serup, O.D. Madsen, Control of endodermalendocrine development by Hes-1, Nat. Genet. 24 (2000) 36–44.
[6] S.E. Schonhoff, M. Giel-Moloney, A.B. Leiter, Minireview: development anddifferentiation of gut endocrine cells, Endocrinology 145 (2004) 2639–2644.
[7] S.B. Smith, H. Watada, M.S. German, Neurogenin3 activates the isletdifferentiation program while repressing its own expression, Mol.Endocrinol. 18 (2004) 142–149.
[8] A. Apelqvist, H. Li, L. Sommer, P. Beatus, D.J. Anderson, T. Honjo, M. Hrabe deAngelis, U. Lendahl, H. Edlund, Notch signalling controls pancreatic celldifferentiation, Nature 400 (1999) 877–881.
[9] E. Lammert, J. Brown, D.A. Melton, Notch gene expression during pancreaticorganogenesis, Mech. Dev. 94 (2000) 199–203.
[10] K.A. Johansson, U. Dursun, N. Jordan, G. Gu, F. Beermann, G. Gradwohl, A.Grapin-Botton, Temporal control of neurogenin3 activity in pancreasprogenitors reveals competence windows for the generation of differentendocrine cell types, Dev. Cell 12 (2007) 457–465.
[11] K. Prasadan, E. Daume, B. Preuett, T. Spilde, A. Bhatia, H. Kobayashi, M.Hembree, P. Manna, G.K. Gittes, Glucagon is required for early insulin-positivedifferentiation in the developing mouse pancreas, Diabetes 51 (2002) 3229–3236.
[12] G. Gradwohl, A. Dierich, M. LeMeur, F. Guillemot, Neurogenin3 is required forthe development of the four endocrine cell lineages of the pancreas, Proc. Natl.Acad. Sci. USA 97 (2000) 1607–1611.
[13] G. Gu, J. Dubauskaite, D.A. Melton, Direct evidence for the pancreatic lineage:NGN3+ cells are islet progenitors and are distinct from duct progenitors,Development 129 (2002) 2447–2457.
[14] Y. Fujitani, S. Fujitani, D.F. Boyer, M. Gannon, Y. Kawaguchi, M. Ray, M. Shiota,R.W. Stein, M.A. Magnuson, C.V. Wright, Targeted deletion of a cis-regulatoryregion reveals differential gene dosage requirements for Pdx1 in foregut organdifferentiation and pancreas formation, Genes Develop. 20 (2006) 253–266.
[15] P.A. Seymour, K.K. Freude, M.N. Tran, E.E. Mayes, J. Jensen, R. Kist, G. Scherer,M. Sander, SOX9 is required for maintenance of the pancreatic progenitor cellpool, Proc. Natl. Acad. Sci. USA 104 (2007) 1865–1870.
[16] P.A. Seymour, K.K. Freude, C.L. Dubois, H.P. Shih, N.A. Patel, M. Sander, Adosage-dependent requirement for SOX9 in pancreatic endocrine cellformation, Dev. Biol. 323 (2008) 19–30.
[17] M.J. Doyle, Z.L. Loomis, L. Sussel, Nkx2.2-repressor activity is sufficient tospecify alpha-cells and a small number of beta-cells in the pancreatic islet,Development 134 (2007) 515–523.
[18] M. Sander, L. Sussel, J. Conners, D. Scheel, J. Kalamaras, F. Dela Cruz, V.Schwitzgebel, A. Hayes-Jordan, M. German, Homeobox gene Nkx6.1 liesdownstream of Nkx2.2 in the major pathway of beta-cell formation in thepancreas, Development 127 (2000) 5533–5540.
[19] C.L. Prado, A.E. Pugh-Bernard, L. Elghazi, B. Sosa-Pineda, L. Sussel, Ghrelin cellsreplace insulin-producing beta cells in two mouse models of pancreasdevelopment, Proc. Natl. Acad. Sci. USA 101 (2004) 2924–2929.
[20] H. Kobayashi, T. Spilde, Z. Li, J. Marosky, A. Bhatia, M. Hembree, K. Prasadan, B.Preuett, G. Gittes, Lectin as a marker for staining and purification of embryonicpancreatic epithelium, Biochem. Biophys. Res. Commun. 293 (2002) 691–697.
[21] V.M. Schwitzgebel, D.W. Scheel, J.R. Conners, J. Kalamaras, J.E. Lee, D.J.Anderson, L. Sussel, J.D. Johnson, M.S. German, Expression of neurogenin3reveals an islet cell precursor population in the pancreas, Development 127(2000) 3533–3542.
[22] A. Villasenor, D.C. Chong, O. Cleaver, Biphasic Ngn3 expression in thedeveloping pancreas, Dev. Dyn. 237 (2008) 3270–3279.
[23] S. Wang, J.N. Jensen, P.A. Seymour, W. Hsu, Y. Dor, M. Sander, M.A. Magnuson,P. Serup, G. Gu, Sustained Neurog3 expression in hormone-expressing isletcells is required for endocrine maturation and function, Proc. Natl. Acad. Sci.USA 106 (2009) 9715–9720.
[24] P. Collombat, X. Xu, P. Ravassard, B. Sosa-Pineda, S. Dussaud, N. Billestrup, O.D.Madsen, P. Serup, H. Heimberg, A. Mansouri, The ectopic expression of Pax4 inthe mouse pancreas converts progenitor cells into alpha and subsequentlybeta cells, Cell 138 (2009) 449–462.
[25] G.A. Norgaard, J.N. Jensen, J. Jensen, FGF10 signaling maintains the pancreaticprogenitor cell state revealing a novel role of Notch in organ development,Dev. Biol. 264 (2003) 323–338.




















